optimal management of game and forage resources in a private fee-hunting enterprise
TRANSCRIPT

Optimal Management of Game and Forage Resources in a Private Fee-Hunting Enterprise
Ray G. Huffaker
The paper formulates a bioeconomic model of a private fee-hunting enterprise to examine the optimal management of this agroecological system under a wide range of bioeconomic circumstances. An empirical application demonstrates that the economic viability of the enterp¡ (concerning managers) and the sustainability of the underlying biological resources (concerning public regulators) depend critically on the initial endowment of resources and on the existence of publicly-mandated or self-imposed rest¡ on maximum allowable hunting rates. Perhaps surp¡ is the implication that in some circumstances regulators may be able to promote resource sustainability by relaxing hunting controls.
Key words: fee-hunting enterp¡ sustainable management.
The desire to diversify farms and ranches and disappointment over the condition of some pub- licly-managed game populations (Wenders) are motivating privately-managed "fee-hunting" en- terprises. One variation considered in Washing- ton is the Private Lands Wildlife Management Area five-year pilot program. Landowners pur- chase buck deer tags from the state, resell them to hunters willing to pay for a trophy buck hunt- ing experience, and direct the proceeds to create and maintain premium wildlife habitats on their properties. The land is opened free to state-per- mitted hunters going after does. The state re- tains ownership of the game, but the landowner manages it as if it were his own (The Spokes- man-Review).
Emergence of fee-hunting enterprises raises interesting questions regarding the economic vi- ability and biological sustainability of hunting habitat. A logical first step in confronting these questions is to provide conceptual insight into optimal enterp¡ management under the wide range of bioeconomic circumstances managers may face. The present paper applies recent lit- erature on optimal harvest of interdependent populations to formulate a normative decision
Ray F. Huffaker is associate professor in the Department of Ag- ricultural Economics, Washington State University.
This investigation was supported in part by funds provided by Washington State University.
Review coordinated by Richard Adams.
model specifying how a representative manager should control hunting to maximize net-present value of the enterp¡ Simulations generated by data collected from previous wildlife habitat studies demonstrate optimizing behavior under several bioeconomic conditions. The simula- tions are indicative of the bioeconomic circum- stances under which a manager can feasibly pur- sue a net-present-value maximizing strategy while keeping the underlying biological stocks from levels that fail to provide an adequate buffer (from a social viewpoint) against environmental shocks tfiggering population crashes.
Past Work
Traditionally, hunting has been regulated by public game managers. Thus, previous research on hunted ungulate stocks generally has focused on economic and ecological information appro- priate to the public manager's nonprofit and multiple-use decision-making context. In such a context, the general objective is to fix hunting at rates driving vegetation and ungulate popu- lations toward sustained stocks that provide a desired mix of services and an adequate buffer against environmental shocks precipitating pop- ulation crashes. Economists have studied the valuation of ungulate stocks to determine the so- cially-optimal mix of services (Keith and Lyon; Cory and Martin), and mathematical ecologists
Amer. J. Agr. Econ. 75 (August 1993): 696-710 Copyright 1993 American Agricultural Economics Association
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

Huffaker Optimal Management in a Fee-Hunting Enterprise 6 9 7
have studied the degree of stability character- izing the natural evolution of vegetation and un- gulate populations under constant publicly-set hunting rates (Metzgar and Boyd).
Standiford and Howitt extended this earlier work to the private sector by analyzing the ef- fect of hunting revenues on the decision of cattle ranchers to provide wildlife habitat on private rangelands by protecting various oak species from livestock grazing and firewood harvesting. They did not explicity model habitat/wildlife inter- actions.
Shedding conceptual light on the optimal management of non-cattle fee-hunting enter- prises requires that past work be extended to ex- plicitly model vegetation/ungulate interactions in the decision-making context of a private man- ager who exploits these biological stocks as cap- ital assets, and thus manages them according to the standard cost-benefit criterion of maximiz- ing the net present value of the enterp¡ (Clark).
Methodology
The optimal management of fee-hunting enter- prises presents the complicated and multi-fac- eted bioeconomic problem of how to control a three-trophic-level system of ungulates, their forage, and their predators (i.e., hunters). Moreover, the emphasis on trophy bucks re- quires that landowners optimally differentiate harvest by age and sex classifications. There- fore, a complete analysis of the optimal man- agement of a private fee-hunting enterprise must incorporate both facets of management: The im- pact of hunting on trophic interactions and on the sex/age structure of the ungulate popula- tion. Unfortunately, capital-theoretical frame- works conside¡ either facet independently have had to suppress the other to remain mathemat- ically tractable. (See, e.g., Derzko and Sethi; Ragozin and Brown; Wilen and Brown; Mes- terton-Gibbons). The complete optimal manage- ment problem appears to be analyzed most amenably by a piecewise approach.
This paper takes an initial step in a piecewise approach by addressing the optimal manage- ment of vegetation-ungulate-hunter (VUH) trophic interactions where the average impact of the overall ungulate population on vegetation (as opposed to the differential impact of specific sex/ age classes) is of primary importance. Modeling proceeds by embedding modified versions of the Metzgar/Boyd constant-hunting model (dis- cussed below) into an optimal control frame-
work characterized by two state variables (veg- etation and ungulates) and one control variable (hunting). The solution to the optimal control problem provides a feedback rule that adjusts hunting rates each instant in response to the sta- tus of the underlying ecological system. Inter- dependent vegetation-ungulate population dy- namics respond simultaneously to the hunting rates determined by this feedback rule, as op- posed to an exogenously determined constant- hunting rate.
Modei Formulation
The general formulation of the fee-hunting problem is
s oo
(1) max J(H) = e-8'Fo(U)toH dt
subject to
(2) 12 = V F l ( V ) - UF2(12)
(3) • = U[-F3(O) + Fa(O)] - BEs(U)
(4) 0 --- H -< H max, V(t = O) = Vo, U(t = O) = Uo
where V represents the aggregate biomass of plant species preyed upon by ungulates (state vari- able, lbs/ar, ar = unit area); U represents the density of ungulates in a single species popu- lation (state variable, hd/ar, hd = head); O = V / U (vegetation per capita) and represents prey abundance for ungulates; H represents an aver- age hunting rate over a given time period (con- trol variable, hnt/ar/t , hnt = hunters); t repre- sents a given time pe¡ and all other parameters and functions are defined below.
The assumption underlying equation (1) is that the enterprise's objective is to determine the level of either-sex hunting each period that maxi- mizes the net present value of fee hunting. The instantaneous flow of rents is
(5) Fo(U) = P U - Co
where P ($/[(hd/ar)(hnt/ar)]) and Co ($/(hnt/ ar) are unit revenue and cost parameters, re- spectively. The average fee that the enterp¡ can charge per unit density of hunting, PU ($/ (hnt/ar)), is assumed to increase linearly with the density of game. This is consistent with studies showing a direct relationship between hunters' willingness to pay per trip and the num- ber of game sighted while hunting (Loomis,
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

698 August 1993
Creel, and Cooper, pp. 4-56; Brookshire, Ran- dal, and Stoll, p. 484). The Loomis, Creel, and Cooper study also offers evidence that hunters in Montana and California are not willing to pay more if the number of competing hunters is cut in half. This offers some justification for ne- glecting congestion by not making average rev- enues an inverse function of the hunting rate, H; and thus for specifying a linear control prob- lemas required by the solution algorithm.
Equation (1) acknowledges that only the frac- tion of hunters purchasing buck tags, to, directly generates income from the marketing of big game. Bucks are assumed to be a constant frac- tion w of the total population, regardless of har- vest or stock level. The constant fraction to can be moved outside the integral in (1), andas a multiplicative constant, has no impact on the se- lection of optimal overall hunting densities. In short, once the optimal overall hunting density is determined, and the desired sex ratio reached, the enterprise can apportion a fractional share to to the sale of buck tags and the remainder 1 - to to the doe hunt. The fraction of state-permitted hunters seeking does at no additional fee has an indirect impact on income by regulating the level of intraspecific competition for vegetation among the total ungulate population.
Finally, the real per period discount rate is 6, and the landowner's selection of hunting rates to maximize (1) is subject to the control con- traint and initial populations given in (4). The maximum possible hunting rate, H max, is ah ex- ogenous parameter determined, for example, by the state that retains ownership of the ungulate population, by insurance policies covering the enterp¡ against hunter accidents, and/or by the enterprise itself.
Equations (2) and (3) comprise the ecological component of the model, and represent variations on the Metzgar/Boyd model. Metzgar and Boyd investigated vegetation-ungulate dynamics un- der constant hunting densities a s a special case of a generalized three-differential-equation eco- system model of vegetation-ungulate-predator interactions. Using information from biologists, the authors incorporated intraspecific competi- tion among predators for prey (i.e., among predators of the same species) into the birth, mortality, and feeding responses of their system by having these rates rely on the ratio of prey to predator densities (i.e., "prey abundance"), as opposed to having them rely simply on prey density as in earlier work.
Equation (2) models the net rate of change in the vegetation stock each period (lbs/ar/t) as
Amer. J. Agr. Econ.
the difference between the rates of density- dependent growth, VF~(V), and predation by un- gulates, UFz(O). Following Metzgar and Boyd, the per capita growth rate of vegetation is as- sumed to be adequately represented by the lo- gistic equation
(6) F~(V) = R(1 - V /K)
where R (1/t) represents the maximum net pro- portional growth rate, and K (lbs/ar) represents the carrying capacity of the land area for veg- etation.
A Michaelis-Menten "saturation" term is used in functions F: through F5 because it allows for the range of nonlinear functional responses that have been found appropriate for grazing inter- action systems (Metzgar and Boyd; Caughley and Lawton):
(,)x (7) G(*) =
0.05D x + (.)x
where G(*) asymptotically approaches one as the argument (*) increases; D is a 95% saturation level, i.e., G(D) = 0.95, and the parameter x determines curvature. Both D and x are related inversely to the efficiency of the process gov- erned by G(*). Metzgar and Boyd set the pa- rameter x to 1 in all of the functional responses except for Fa. This paper sets x at 1 in F4 as well, to maintain linearity of the problem in the control variable (H). The result is to lessen the impact of "energy partitioning" at low food abundance where ungulates decrease birth rates to promote adult survival.
The per capita consumption rate of vegetation by ungulates is given by
(8) F2(O) = C2G2(O)
where C2 (lbs/hd/t) represents the maximum per capita consumption rate at high prey abundance, and D2 (lbs/hd) represents the prey abundance at which ungulates are 95% satiated. As prey abundance increases from zero, per capita con- sumption increases from zero asymptotically to- ward C2.
Equation (3) stipulates that the net rate of change in the ungulate stock each period (hd/ ar/t) is the birth rate, UF4(O), less rates of mor- tality by natural causes, UF3(~2), and by hunt- ing, HFs(U). The per capita ungulate birth rate is represented by
(9) F4(~'-~) = C4G4(~'~)
where C 4 (1/ t ) is the maximum per capita birth
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

Huffaker Optimal Managemen t in a Fee-Hunt ing Enterprise 699
rate at high prey abundance, and D 4 (lbs/hd) is the prey abundance at which births are 95% maximized. As prey abundance increases from zero, per capita births increase from zero asymptotically toward C4.1
The per capita natural mortality rate is
(10) F3(O) = M1 - (Mi - M2)G3(~Q)
where M~ (1/t) is the maximum per capita death rate at low prey abundance, M2 (1/t) is the min- imum per capita death rate at high prey abun- dance, and D 3 is the prey abundance at which deaths are 95% alleviated. The per capita nat- ural mortality rate drops from M~ asymptotically toward M2 as prey increases from zero. The av- erage kill per hunter (hd/hnt) is
(1 1) F5(U) = CsGs(U)
where C5 (hd/hnt) is the game limit per hunter, and D5 (hd/ar) is the ungulate density at which the average hunter enjoys a 95% chance of suc- cess. As the ungulate density increases from zero, the average kill per hunter increases from zero asymptotically toward C5.
Parameter Scaling
The combination of analytical and numerical work needed to solve the maximization problem in (1)-(4) is facilitated by converting the prob- lem to one using dimensionless variables and parameters. Most importantly, the number of system parameters decreases from fourteen to nine, which meets the size limitations of the dy- namical-systems solution software available for this study. 2 The dimensionless model is
(12)
~oc
m a x J ( h ) = Jo e - ~ ' f ~
subject to
d v (13) - - = v[ f i (v ) - ti(0)]
dr
d u (14) - - = u[- f3(O) + fa(O) - hfs(u)]
dT
(15) m a x
O <- h <- h , v(t = O) = vo, u( t = O) = uo
wheref0(u) = cu - 1;fl(v) = r(1 - v) ; f i (O) =
1/(1 + 0);f3(0)= s(d3ml + Om2)l(d3 + 0);fa(0) = sO/(d4 + 0); fs(u) = 1/(k5 + u); 0 represents v / u ; and scaled variables and parameters are de- fined in table 1. The net present value for the dimensionless problem is given by NPV = (wCoK/O.O5D2Cs)J , but the selection of optimal hunting rates over time is not affected by the multiplicative constant.
Most of the dimensionless variables and pa- rameters have intuitive bioeconomic meaning. The scaled vegetation variable, v, measures the proportion of grazing area carrying capacity present, and is thus defined over the interval (0, 1). The dimensionless ungulate variable, u, scales the ungulate population relative to the proportion of the vegetative carrying capacity required for an ungulate to reach 95% satiated consumption each period. The scaled parameter r is a measure of the maximum grazing pressure exerted by ungulates on vegetation. The scaled discount factor, r~, compares the performance of alternative capital to that of ungulates on veg- etation. The scaled parameters di compare birth and death efficiency parameters to the feeding efficiency parameter. Finally, the dimensionless mortality parameters, mg, scale the maximum and minimum per capita ungulate mortality rates to the maximum per capita birth rate.
Solution Procedure
The solution procedure applies techniques pi- oneered by Mesterton-Gibbons (1988) to derive the optimal control rule in a linear control set- ting for the combined harvest of both predator and prey in a coupled ecosystem. Mesterton- Gibbons solved a system where predation func- tional responses are linear in the prey variable.
Table 1. Dimensionless Variables and Pa- rameters
Var iab les Pa ramete r s
1 Metzgar's and Boyd's model does not include lagged birth ef- fects. They simulated the effect of such a realistic modification in their model and concluded that "dynamics without time lags are suitably realistic" for their data set (p. 20).
2 TraX. Simulation and analysis of dynamical systems, version 1.1, dist¡ by Exeter Software, Setauket, NY (1990).
v = V / l r
u = (O.05D2/K)U h = ((O.05D2)2CffKC2)H r = (C2/O.05Dz)t
c = (P/Co)K/O.O5D~ r = O.05RDffC2 s = 0 .05C4D#C2 di = Di/D2 i = 3 ,4 mi = Mi/C4 i = 1,2 ks = O.05~D2DJK ~o = 0 .058Df fC2
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

700 August 1993 Amer. J. Agr. Econ.
His solution procedure is modified to solve the system dictated by equations (2) and (3) where only the predator species (ungulates) is har- vested, and the functions governing ungulate feeding (8), birth (9) and death (10) processes incorporate intraspecific competition.
If h~ is the control trajectory optimizing (12) subject to (13)-(15), then the Pontryagin necessary conditions require
(a) the existence of continuous adjoint func- tions Al and A2 such that
(16) dA~ OH
d'r Ov
(17) dAz OH
dz Ou
where
(18)
and
H(v, u, h, Al, A2, r) = e-~~fo(u)h + , ~ q - A(0)]
+ ,~2u[f4(0)-f3(o)- hf~(0)]
(b) H[v*(~-), u*(~-), h*(~-), Al*(~'), A*z(~'), ~'] �87 H[v*(z), u*(r), h(z), Al(z), A2(~'), r],
for all z [Tu, p. 114].
The adjoint system (16)-(17) dictates the inter- dependent optimal depreciation schedules for the vegetation and ungulate capital assets, respec- tively. The schedules require that each capital stock depreciates in value over time no faster than its marginal cont¡ to the flow of valuable output through long-term capital stock accumulation.
Following Mesterton-Gibbons, the solution procedure commences by defining the following new variables:
(19) W1 = e~'A1 v
(20) W2 = e~~)t2u
(21) r/(u, "r) = e-'P'[fo(u) - W2fs(u)]
(22) Zl(V, u) = fl(v) - f2(0)
(23) zz(0) =f4(0) - f3(0)
The transformed Hamiltonian (18) and adjoint system (16)-(17) are
(24) H = rl(u)h + e-~~[Wlzl(v, u) + W2z2 (0)]
dW1 (25) = W l ( ~ - VZlv ) - W20z20
dz
(26) dW2
-- --WlOf2e + W2(q~ + 022o + fs,,uh) - cuh dz
where Zlv, Z2o, andfsu are the first de¡ of z~, z2, and f5 with respect to v, 0, and u, re- spectively. The transformed adjoint system (25)- (26) is derived by taking the dimensionless time derivatives of (19)-(20) and then substituting in the previous adjoint system (16)-(17) and the equations of motion (13)-(14).
Necessary condition (b) above is the "opti- mality principle" that requires OH~Oh = rl(u) = 0 [by (24)]. The enterprise must adjust hunting densities each period to balance the short-term discounted marginal gains of increased hunting, e-~'fo(u), against the long-term discounted mar- ginal impacts on the value of accumulating un- gulate stocks, e -~" W2fs(u) [by (21)]. For ex- ample, above-optimal hunting densities increase short-term profits, but decrease the value of the ungulate capital stock. Ungulates that would be more valuable as reproductive stock generating enhanced future harvests, are instead liquidated with the proceeds reinvested in some lower-re- turning asset. At the margin of use, ungulates become a supe¡ asset (earning greater than a normal rate of return) that the landowner must invest in by decreasing hunting densities.
The linearity of the Hamiltonian in the control variable, h, requires a control sequence from (27) below to drive vegetation and ungulate popu- lations to the optimal singular path (along which r/(u, "r) - 0) from any given initial population level as rapidly as possible.
0 ir r/(u, ~-) < 0
(27) h ~ = h*(v, u) if ~7(u, z) = 0
h max if r/(u, r) > 0
where h*(v, u) is the optimal feedback rule that adjusts hunting rates each period to track the singular path.
The solution requires solving explicitly for h*(v, u). Taking the first and second dimen- sionless time de¡ of ~/(u, r) in (27) yields
(28) dT/ - - = e-'PT[K(u, O)W1 -- L(u, O)Wz - M(u, 0)] dz
dZr/ (29) - e-'PT[A(u, O, WI, W2)h
d'?- - B ( v , u , O, W l , W2)]
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

Huffaker
where functions K, L, M, A, and B are defined in table 2.
Because r/(u, "r) = 0 along the singular path in (27), the first and second dimensionless time derivatives in (28)-(29) also vanish. Forming the two equation system ~7 ~ 0 = d ~ / d z allows solution of the transformed adjoint va¡ along the singular path as functions of the variables and parameters of the system wl = W~(v, u) and w2 = W2(u), where Wl and w2 are given in table 2. Finally, setting d2~7/dr 2 = 0 in (29) yields the feedback control law
(30) b(v, u) uz2(O)q(v, u) + O(v, u)
h*(v,u) - - - - a(v, u) ufs(u)q(v, u)
where z2 is defined in (23); q and ~b are defined in table 2; and a(v, u) and b(v, u) result from substituting w~ and w2 into A(v, u, W~(v, u)), Wz(u)) and B(v, u, Wl(v, u), Wz(u)).
Synthesis of the Optimal Hunting Policy
The optimal control sequence of hunting rates, h ~ originating from given initial populations of vegetation and ungulates, (v, u)0, is synthesized from three phase planes, each associated with a different case in (27). The "naturally-regulated" (h = 0)and "maximal-hunting" (h = h m~x) phase planes are used as most rapid approaches to the singular path contained in the singular phase plane (h = h*(v, u)). This section considers each phase plane in turn. Due to the complexity of the un- derlying biological functions, the properties of these phase planes must be largely determined numerically. The baseline parameter values col- lected by Metzgar and Boyd are used for this purpose (table 3).
Table 2. Functions Needed to Calculate h*(v, u)
K(v, u) = f~A.O L(v, u) = z2ufs,, + Ofsz20 M(v, u) = -[~0 + cu(z2 - q~)] wl = Wl(v, u) = [foL(v, u) + f~Al]/fsK(v, u) w2 = W2(u) = fo/f5 A(v, u, W,, W2) = -uf~V~.W, + ufs[L. + fsL(v, u)]Wz +
L(v, u)cu + M.uf~ B(v, u, W,. W2) = [vz,~K(v, u) - OfzoL(v, u) - vzlKv -
uzzK.]W~ + {Ozz~[K(v, u) + L(v, u)] + vzlLv + uz2L.}Wz - q~M + M.uz2
q(v, u) = M. + cL(v, u)/f5 - K.wl + [L. + L(v, u)fs]w2 O(v, u) = [vz,JK(v, u) - 0f20L(v, u) - vz,Kvlw, + {Ozze[K(v,
u) + L(v, u) +vz~Lv - z2L(v, u)uf�91 - ~M - z2L(v, u)uc/f5
Optimal Management in a Fee-Hunting Enterprise 701
The Naturally-Regulated Phase Plane
The system of differential equations governing the natural evolution of vegetation and ungulate populations without hunting by man is derived by setting h = 0 in (13)-(14):
dv (31) - - = VZl(V, u)
dr
d u (32) - - = uz2(O),
dr
where zl and Z 2 are defined in (22) and (23). The vegetation zero-change isocline, vi, generated by the set of baseline biological parameters in table 3, assumes the familiar negative quadratic form shown in figures l a-b. The ungulate iso- cline, ui, is a line passing through the origin (figure la).
The naturally-regulated system is character- ized by a single nonzero equilibrium at the in- tersection of the two isoclines. The equilibrium can be shown analytically to be either a stable focus or node (if it occurs to the right of the maximum-sustained-yield stock level). The nu- merically-generated phase diagram for the set of baseline bioeconomic parameters shows it to be a stable focus (figure 2a). Therefore, in the ab- sence of hunting, ungulate and vegetation pop- ulations naturally cycle towards a steady state over time. Consider, for example, the dynamics initiating from various ungulate stocks when vegetation is at carrying capacity (eastern mar- gin of figure 2a). When the ungulate population is relatively small, it initially increases because high per capita food availability produces births at a higher rate than deaths. Vegetation imme- diately decreases because any positive level of consumption outpaces the near-zero vegetation growth rates associated with carrying capacity. Ungulates continue to increase until forage availability drops to the point that natural mor- tality begins to outpace births. Both stocks de- crease until ungulate grazing pressure drops suf- ficiently for both to recover, and then the cycle starts over at decreased amplitudes.
When initial ungulate populations are suffi- ciently large, both stocks undergo a combina- tion of precipitous declines and exploding re- coveries that can create sufficiently wide swings that both stocks may go extinct before they can recover. Such "extinction" trajectories originate at initial ungulate stocks greater than u = 0.1755 (2632 hd/10 sq mi) and terminate at the origin.
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

702 August 1993
Table 3. Baseline Bioeeonomic Parameter Values a
Amer. J. Agr. Econ.
Parameter Description Units Value
t a r R b
K ~ C2 b D2 b MI c
M: D3 c C4 d D4 d C5 e
05 ~ 6 f
P/Cg
time period yr 1
unit area sq mi 10 intrinsic growth rate of vegetation t-, .8
carrying capacity for vegetat ion lbs /a r 3000 max imum ungulate consumpt ion rate o f vegetat ion lbs/hd/t 1.2 prey abundance promoting 95% ungulate satiation lbs /hd 4 max imum per capita ungulate natural mortali ty rate t - ' 2.3 min imum per capita ungulate natural mortality rate t - t 0.05 prey abundance alleviating mortali ty by 95% lbs /hd 8 max imum per capita ungulate birth rate t-~ 0.4 prey abundance at which births are 95% maximized lb s /hd 8 ungulate limit per hunter (hnt) hd /hn t 1 ungulate density promot ing 95% hunter success h d / a r 10 real annual discount rate t - ' 0 .0484 ratio o f unit price parameter to unit cost ( h d / a r ) - ' 0 .02
(0 .0002 ,0 .002 ,0 .2 )
a General notes: The baseline levels of the biological parameters come from Metzgar's and Boyd's constant hunting model, and result from their literature review and consultations with biologists and game managers. b The values of R, K, and C2 come from the white-tailed deer model of Caughley and Caughley and Lawton. The value R=0.8/year implies a maximum annual growth factor of vegetation of 2.2, since disregarding the density dependent term ( - V / K ) in (6), the maximum growth rate becomes dV/dt = RV, with solution V = Voe Rt. The intrinsic growth rate consistent with any annual growth factor (VIVo) can then be solved for as R = ln(V/Vo)/t. The value C2 = 1.2 lbs/hd/t implies that about 2500 animals could feed to fullness on ten square miles during at least one year (2500 = K/C2). In setting D2, Metzgar and Boyd assume that satiation requires food availability to be several times greater than maximum consumption (Cz). Thus, D2 = C2(3.33) = 4 (lbs/ten sq mi), where the factor 3.33 is assumed to be "several." c The value M1 = 2.3/year implies 10% survival since, disregarding births and hunting mortality, the natural mortality rate is dU/dt = -F3U = -MIU, where the limit of F3 as 0 ~ 0 is M~ [by (10)]. Hence, by the same calculations done for parameter R above, the maximum death rate associated with any survival factor (U/Uo) is Mi = -ln(U/Uo)/year, where U/Uo = . 1 produces MI = 2.3/year. Likewise, the value M2 = .05/year can be shown to imply 95% survival, where the limit of Fa as 0 ~ oc is M2 and U/Uo = .95. Parameter Da is arbitrarily set at twice the level of D2 since grazing with high food abundance requires less energy and exposure to predators. d The value 6"4 = .4/t lies between values reported by Tanner and McCullough. This implies a maximum annual increase of the deer population (disregarding natural mortality or hunting) of 50%, i.e., .4 = ln(l.5)/t, where the limit of F4 as 0 ~ ~ is C4 [by (9)]. Metzgar and Boyd assume that reproduction becomes asymptotic at a similar level of food availability as survival, so that D4 = Da = 8. e There is a limit of one deer/hunter (Cs = 1) and the number of deer per ten square miles guaranteeing the average hunter a 95% chance of success is assumed by Metzgar and Boyd to be D5 = 10. This value presumes a reasonably long hunting season du¡ a susceptible
riod with highly efficient or persistent hunters. he real annual discount rate is calculated as the annual interest rate on AAA corporate bonds for February 1989 (.0964) less the
percentage change in p¡ over the previous year (.048) [Federal Reserve Bulletin, May 1989, tables A24 and A52]. : Unfortunately, fee-hunting enterp¡ ate sufficiently new in Washington that insufficient information exists to calculate either the revenue parameter P o r unit costs Co for a representative enterp¡ The best that can be done currently is to simulate the model over a wide interval of P/Co ratios in the scaled parameter "c" to determine a range of dynamic behavior. The baseline ratio is assumed to be .02 and the model is run also for values of .0002, .002 and .2.
The Maximal-Hunting Phase Plane
The system of differential equations governing the exploitation of the ungulate population at the maximum constant rate, h = ,.hmax, is
dv (33) - - = l,'Zl(V , O)
dr
du (34) - - = U[Z2(O ) -- hmaxfs(u)] .
dr
The vegetation zero-change isocline remains un- changed from the naturally-regulated case where h = 0. However, the ungulate isocline develops into a cone shape as the constant hunting density increases from zero (figure lb).
Constant hunting rates below c¡ level h ma~ = 0.000782 (3.52 hnt /10 sq mi/yr) , generate ungulate isoclines that intersect the vegetation isocline twice; thereby producing two steady states at lower- and middle-vegetation levels. These two steady states join a higher-vegetation steady state, where vegetation is at carrying capacity and ungulates are extinct, to fo rma tri- steady-state configuration. Consider, for ex- ample, the nume¡ phase dia- gram associated with h maX = 0.0007 (3.15 hnt / 10 sq mi/yr) (figure 2b). The middle-vegetation steady state, (Vm, Um) c, is a saddle point whose associated divergent separatrices feed into the two stable outer steady states. The lower-veg- etation steady state, (vi, ut)', is a focus, and the higher-vegetation steady state, (Vh, Uh)', char-
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

Huffaker Optimal Management in a Fee-Hunting Enterprise 703
ui
0.04
( V ~, U e, h : O )
0.03
v i
0.02
0.01 �9
I I I I ~ . 0.2 0.4 0.6 0.8 I
Figure la. Naturaily regulated (zero hunting) system with single steady state; darkened section of vi represents feasible solutions of ~(v, u) = 0
u i ui
0.04
o.o3 ~ v i f " ~ ( W , W , h " )
0.02 ( V . / , Umee h taz)
: " ' " - - ui
0.01
Figure lb.
(V i% Ul, e, h ' m ) I I l , I
0.2 0.4 0.6 0.8 V
Intermediate hunting pressure with tri-steady-state configuration
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

7 0 4 August 1993 Amer. J. Agr. Econ.
acterized by vegetation at carrying capacity and ungulates at extinction, is a node. 3 The conver- gent separat¡ associated with the saddle point are the thickened lines forming a cone shape. Initial vegetation/ungulate populations that fall within the cone continue (as in the naturally- regulated case) to gravitate cyclically toward a lower-vegetation focus. Initial populations on the outside evolve over time to the v-axis (i.e., un- gulate extinction), and then along the v-axis as vegetation gravitates toward its carrying capac- ity.
The imposition of a constant harvest rate in- troduces the possibility that ungulates may be driven to extinction before they can totally de- plete the vegetation stock. Consider, for ex- ample, the case where the initial vegetation pop- ulation is at carrying capacity and the initial ungulate population is greater than the upper limit of the cone (u > 0.1015). These relatively large
3 The only stability result that can be demonstra ted analyt ical ly is that the h igher-vegeta t ion equi l ibr ium is ei ther a stable focus or node.
initial populations continue (as in the naturally- regulated case) to generate dynamic forces that cause both populations to crash. However, (un- like the naturally-regulated case) the constant harvest density inhibits cyclical recovery by un- gulates, and after ungulates die out, vegetation recovers back to carrying capacity. Similarly, initial ungulate populations smaller than the lower limit of the cone eventually are hunted to ex- tinction, but more quickly.
Constant hunting rates above the critical level, h max --- 0.000782, cause the ungulate isocline to rise above the vegetation isocline; hence the system is left with the single steady state at veg- etation carrying capacity. The constant harvest rate is sufficiently large to force any initial un- gulate population toward extinction over time.
T h e S i n g u l a r P h a s e P l a n e
The system goveming population dynamics when the hunting rate is adjusted each period accord- ing to the feedback control law h*(v, u) in (30) is
< i
F i g u r e 2a . - u - 0 . 1 5
V
61; ~~ _ _ h ~ - 0 n a t u r a l l y - r e g u l a t i n g p h a s e p l a n e , w h e r e - 0 . 1 < v < 1 a n d - 0 . 0 2
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from
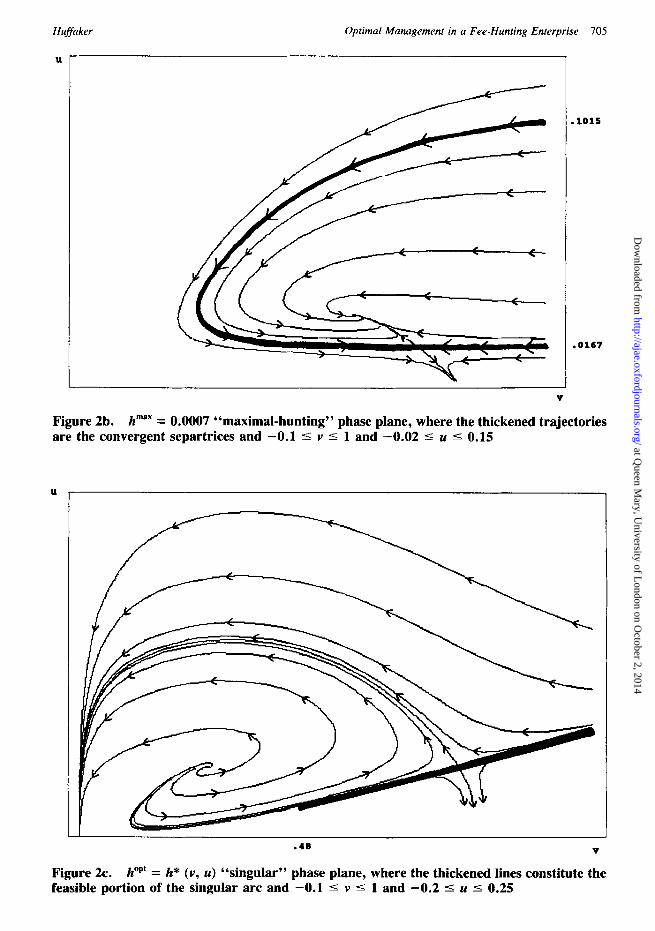
Huffaker Optimal Management in a Fee-Hunting Enterprise 705
< ( -
. Z 0 1 5
, 0167
v
Figure 2b. h max = 0.0007 "maximal -hun t ing" phase plane, where the thickened trajectories are the convergent separtrices and - 0 . 1 -< v -< 1 and - 0 . 0 2 < u <- 0.15
. 4 8 V
Figure 2c. h ~ = h* (v, u) " s ingu la r " phase plane, where the thickened lines constitute the feasible portion of the singular arc and - 0 . 1 -< v <- 1 and - 0 . 2 -< u -< 0.25
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

706 August 1993 Amer. J. Agr. Econ.
dv (35) - - = vzl(v, u)
d7
du (36) - - = u[z2(O) - h*(v, u)fs(u)].
dr
Setting du/dT = 0 in (36) and solving for h yields an ungulate zero-change isocline: h*(v, u) = z2/ fs. Equating this isocline with the control rule h*(v, u) shows that steady states in the singular plane must satisfy ~ v , u) = 0 in order for the control rule to be consistent with the ungulate isocline, where t0(v, u) is defined in table 3. Moreover, a full equilibrium requires that the solutions of ~(v, u) = 0 lie along the vegetation zero-change isocline. Finally, the control con- straint (15) requires that optimal hunting den- sities be nonnegative. Thus, only combinations of vegetation and ungulate populations lying on the vegetation isocline below the ungulate iso- cline associated with h = 0 are feasible solu- tions of to(v, u) = 0 (i.e., the thickened portion of the vegetation isocline in figure 1 a). Steady- state solutions to ~ v , u) = 0 must be found nu- merically.
Figure 2c displays the numerically-generated singular phase plane for the baseline parame- ters. As stated in the notes to table 3, no infor- mation was located to calculate the unit revenue and cost parameters, P and Co, respectively. Thus, a range of simulations were run over a wide interval of P/Co ratios in scaled parameter "c" to determine a range of dynamic behavior. The baseline ratio was assumed to be 0.02 and the model was run also for values of 0.0002, 0.002 and 0.2.
The baseline singular phase plane has two steady states. The lower-vegetation steady state is an unstable focus whose trajectories emanate towards the higher-vegetation saddle point. 4 One of these trajectories supplies the separatrix con- verging toward the saddle point from the left. The other separatrix converges from higher veg- etation levels on the right. The unstable focus is an infeasible solution (since the associated hunting rate is negative) that is shown numeri- cally to be net-present-value (npv) minimizing. Alternatively, numerical results show that the saddle point is the npv-maximizing solution, i.e., (v, u)* = (Vh, U) ~, where the steady-state hunt- ing rate is h*(v*, u*) = 0.000746 (67.818 hnt / 10 sq mi/yr) . The portion of the two convergent separat¡ for which hunting densities are pos-
4 Nothing can be said analytically regarding the stability of steady states in the singular phase plane.
itive constitutes the optimal singular path de- noted as F(v, u). The hunting rates associated with the separatrix converging from the left re- main positive for population levels greater than (v, u) = {0.48 (1440 lbs/10 sq mi), 0.01989 (298 hd/10 sq mi)} for the set of baseline bio- economic parameters. The hunting rates asso- ciated with the separatrix converging from the right are positive within the relevant range. Hence, the portion of the convergent separa- trices within 0.48 -< v -< 1 comprises the sin- gular path F(v, u) (the thickened line in figure 2c). The hunting density is adjusted over time along the singular path according to the feed- back control law h*(v, u) in (30) toward the steady-state density h*(v*, u*).
Increasing the P/Co ratio in the simulations increases the average value of hunting relative to the average cost of supplying the hunting. The landowner responds by increasing the sustained annual hunting rate. Increased sustained hunting drives steady-state ungulate densities down and less-heavily grazed steady-state vegetation stocks up. The basic configuration of an npv-maximiz- ing, higher-vegetation saddle point and an npv- minimizing, lower-vegetation unstable focus re- mains throughout the simulations. The smallest ratio (0.0002) does generate other non-maxi- mizing steady states. However, the presence of these solutions does not change the optimal strategy of following a trajectory in the natu- rally-regulated or maximal-hunting phase planes as a most-rapid-approach to the singular path, F(v, u), associated with the npv-maximizing saddle point.
The control rule given in (27) can now be re- stated more explicitly as
(37) 0 if (v, u) rests below F(v, u)
h ~ = h*(v, u) if (v, u) rests on F(v, u)
hmax if (v, u) rests above F(v, u)
where it is assumed for the time being that the feedback control rule evaluated at the net-pres- ent-value-maximizing steady state, h*(v*, u*), satisfies the control constraint (15), i.e., 0 -< h*(v*, u*) <-- h max. An enterprise beginning with initial vegetation/ungulate populations resting below the singular path F(v, u) must prohibit hunting for the interval of time needed for stocks to recover along a trajectory in the h = 0 phase plane up to singular levels (figure 3). Thus, the optimal solution initially simulates the dynamics of a naturally-regulated (unhunted) system. Once
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

Huffaker Optimal Management in a Fee-Hunting Enterprise 707
h = h m (v,u)o
r(,, .)
(v,u)*
(v,u)~
Cv,u), h=0
Figure 3. Optimal hunting sequences from initial conditions above and below F(v, u), where 0 -< h* (v*, u*) -< h max
stocks reach the singular path at levels (v, u)int, hunting begins and is adjusted according to the feedback control law. The removal of ungulates from the system pulls stocks out of dampened cycling toward a natural equilibrium and into an asymptotic increase toward the net-present-value- maximizing steady state characterized by a lower ungulate stock and an increased (less heavily grazed) vegetation stock.
Altematively, the advantage of relatively large initial stocks is that the enterprise can earn in- come during the most-rapid-approach period by setting hunting rates at the maximum tate, h max, for the length of time required for stocks to be driven downward along a trajectory in the h = h max phase plane to singular levels (v, u)int (fig- ure 3). A fraction to is apportioned to the sale of buck tags, and the remainder 1 - oJ to pop- ulation control via free doe hunting. Sufficiently large maximum hunting densities suppress the wide cycles associated with naturally-regulated dynamics, and thus permit a direct approach to the singular path.
Discussion
The manager's ability to pursue the optimal control sequence dictated by (37) for a given
initial endowment of vegetation/ungulate stocks may depend critically on the extent to which the enterprise must operate within externally- and/ or self-imposed restrictions on the maximum possible hunting density exerted on average in any year, h max. The average maximum hunting density can be restricted by decreasing the length of hunting seasons or the number of hunters. Such hunting restrictions may be imposed by state regulation or by an insurance policy protecting the enterprise against liability for hunting acci- dents. Hunting season length is perhaps the most controversial point of contention between man- agers and their public regulators (Stevens, Spokesman-Review). Moreover, the manager may voluntarily restrict maximum hunting num- bers to facilitate the supervision necessary to en- sure that hunters shoot the correct numbers and types of animals (e.g., that doe-hunters paying no fee to the manager do not kill trophy bucks) (Stevens).
As shown below, the significance of hunting- rate restrictions for economic viability is that the enterprise may be unable to sufficiently regulate intraspecific competition among ungulates for vegetation at crucial times, and thereby be forced into suboptimal (restricted) control sequences that decrease the benefits of initiating hunting from relatively large (and possibly even smaller) ini-
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

708 August 1993
U
ti
Amer. J. Agr. Econ.
( v , u ) .
hm �9
hmS
11"2 h | 0 ~ " ~ | V I | U l ) |
v
Figure 4. Blocked interval where h mx < h* (v*, u*) and initial populations are outside of the cone shown in figure 2b
tial vegetation/ungulate stocks. The signifi- cance for sustainability of the underlying bio- logical resources is that some rest¡ control sequences may dictate that stocks follow cycli- cal (as opposed to more direct) most-rapid-ap- proach paths to a npv-maximizing steady state. These cycles may drive stocks perilously close to extinction during critical time pe¡ and thus provide inadequate buffers (from a regu- latory viewpoint) against environmental shocks precipitating population crashes. Perhaps sur- prisingly, public regulators intending to pre- serve ungulate stocks with a cap on hunting (either through limited hunting seasons or tags sold to the enterprise) may instead unintention- ally impede intraspecific-competitive forces that can make stocks less susceptible to ultimate de- pletion during c¡ time periods.
Consider now how restrictions on the maxi- mum hunting rate can generate suboptimal con- trol sequences. Such rest¡ tighten the con- trol constraint, 0 -< h -< h m~x. "Blocked intervals ~ arise when the control constraint is sufficiently tight that it prevents the optimal control se- quence from tracking the singular path F(v, u) from a range of initial endowments (Clark, p. 56).
The entire singular path F(v, u) can be blocked if the maximum hunting density is smaller than that sustaining the optimal steady-state solution, i.e., h max < h*(v*, u*). In this case, driving stocks to the singular path F(v, u) is pointless because h*(v*, u*) is not feasible. The best that the enterprise can do is to maintain the hunting density at h max and d¡ stocks to a steady state in the "maximal-hunting" phase plane (Clark, pp. 96-97).
Assume, for example, that the maximum hunting density is restricted to h max - - 0.0007 (3.15 hnt/10 sq mi/y0. This level generates the "maximal-hunting" phase plane in figure 2b, and is less than the optimal sustained hunting den- sity computed for the baseline parameter values, h*(v*, u*) = 0.00078 (3.51 hnt/10 sq mi/yr). If initial stocks are within the cone formed by the convergent separatrices associated with the saddle-point equilibrium, then maximal hunting drives stocks toward the stable focus (vt, ut)'.
Alternatively, if initial stocks are outside of the cone, then the optimal strategy is more com- plicated as shown in figure 4 where (v, u)0 = (1,0.17). Here, the hunting density is set ini- tially at h max, whereupon stocks crash along the associated trajectory in the h max phase plane. Af-
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

Huffaker
ter pe¡ "r~, hunting is stopped to allow the overexploited stocks to recover along a trajec- tory in the h = 0 phase plane up to the conver- gent separatrix approaching the saddle-point equilibrium, (Vm, Un)', from the left in the h max phase plane. 5 The switch to the convergent se- paratrix and a resumption of h max takes place in period r2, where r2 is directly related to z~. In other words, the length of the recovery time is directly related to the length of time that it is postponed, ceteris paribus.
There is an economic trade-off underlying the optimal choice of the initial switching time "ti. The enterprise must select the switching time that balances the marginal benefits of operating at the maximum density with the marginal costs of shutting down operations as stocks recover up to the separat¡ approaching the saddle-point equilibrium in the h max plane. The optimizing value of ~-~ can be found by expressing the net present value of the entire hunting policy a s a function of rl, applying Leibnitz' rule to derive the net marginal present value with respect to "rl, and using numerical iterative techniques to find the relevant "r~ setting net marginal present value equal to zero (Clark, p. 75).
Portions of the singular path F(v, u) may be blocked even if h max > h*(v*, u*). Blockage oc- curs when relatively large initial stocks and sus- tained application of h rnax c a u s e wide population swings that evolve around and below the sin- guiar path in driving ungulates to extinction and vegetation to carrying capacity. Hence, the en- terprise must pursue a strategy similar to figure 4b, where the singular path is reached and fol- lowed after a recovery time of z2 - ~'1 periods, and ~-~ is determined optimally as described above.
Conclusion
This paper is motivated by the recent emergence of publicly-regulated private-fee-hunting enter- prises which seek to diversify farm and ranch operations and improve game populations. Fee hunting enterp¡ buy buck deer tags from the state, resell them to hunters, use the proceeds to invest in habitat management, and allow some level of doe hunting for free. Interest in the practice of, and the potential for, fee-hunting
5 The convergent separatrix is the first h max trajectory that ap- proaches a steady state in the maximal-hunting phase plane, and thus shortens the recovery pe¡ compared to h r~x trajecto¡ above it that spiral in toward the stable focus.
Optimal Management in a Fee-Hunting Enterprise 709
enterp¡ creates the need for conceptual in- sight into the optimal management and regula- tion of these novel agroecological systems.
The paper investigates the dynamical prop- erties of optimal management of vegetation-un- gulate-hunter interactions in such enterprises. The approach taken is dynamic optimization embed- ding a modified version of a recently developed three-trophic-level ecological interaction model of ungulates, their prey (vegetation) and their predators (hunters). The framework generates an optimal hunting sequence designed to drive initial vegetation and ungulate populations as rapidly as possible to a "singular" path. Such a path would represent the optimal approach to ul- timately sustainable populations maximizing the net present value of the enterp¡
The optimal hunting sequence is shown to de- pend critically on the interplay between the ini- tial endowment of vegetation/ungulate stocks and restrictions on the maximum possible hunt- ing density exerted in any year. Results indicate that an enterp¡ can fully profit from abundant initial stocks if it can exert relatively large av- erage maximum hunting densities. These suppress the wide cycles associated with naturally-regu- lated dynamics, permitting a direct approach to the singular path. Otherwise, the enterp¡ is forced into suboptimal management strategies dictating a cyclical most-rapid-approach, in which maximal hunting eventually drives stocks around and below the singular path until a hunting mor- atorium allows them to recover to singular lev- els. Strategies of this sort decrease the economic viability of the enterprise, and may render stocks more vulnerable to environmental shocks which precipitate population crashes. Moreover, if the maximum hunting density is below the optimal density sustaining net-present-value-maximiz- ing stocks, the best the enterp¡ can do is to drive stocks to a maximal-hunting steady-state solution.
Enterp¡ forced by regulations requiring operation within limited hunting seasons into such "blocked-interval" conditions benefit the most from policies that relax the maximum hunting rate control constraint, 0 -< h -< h max, (e.g., ex- tended hunting seasons). These enterprises are aided also by other means of decreasing ungu- late consumption of vegetation during c¡ periods, such as the offer of free doe hunting. Perhaps surp¡ is the implication that under some circumstances, public regulators can pro- mote the sustainability of vegetation/ungulate stocks by relaxing hunting restrictions.
These above results are accompanied by var-
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from

710 August 1993 Amer. J. Agr. Econ.
ious caveats, the most important of which may be that many critical factors affecting vegeta- t ion/ungulate dynamics, for example, ungulate diseases, drought, and over-winter feeding of ungulates, are not modeled directly. However , the model can be useful for indirectly analyzing the management implications of such factors. For example, the implications of short-run epidem- ics and drought conditions can be studied by perturbing vegetation/ungulate stocks away from steady state and viewing their optimal recovery paths (Wilen and Brown). The impact of more persistent diseases and drought conditions can be studied by increasing per capita ungulate death rates and lowering per capita vegetation growth rates, respectively. Finally, the impact of over- winter feeding can be studied by lowering the max imum per capita ungulate death rate at low food availabilities.
[Received D e c e m b e r 1991; f i na l revision received Augus t 1992.]
References
Brookshire, D. S., A. Randall, and J.R. Stoll. "Valuing Increments and Decrements in Natural Resource Ser- vice Flows." Amer. J. Agr. Econ. 62(August 1980): 478-88.
Clark, C. W. Mathematical Bioeconomics: The Optimal Management ofRenewable Resources, 2 ed. New York: John Wiley & Sons, 1990.
Caughley, G. "Wildlife management and the dynamics of ungulate populations, ~ in Applied Biology, Vol. I, T.H. Croaker (ed.), Academic Press London (1976).
Caughley, G., and J. H. Lawton. "Plant-Herbivore Sys- tems," in Theoretical Ecology, 2nd Ed., R. M. May (ed.), S inauer Associates, Sunderland, MA (1981).
Cory, D. C., and W. E. Martin. "Valuing wildlife for ef- ficient multiple use: Elk versus Cattle." West. J. Agr. Econ. 10(December 1985):282-93.
Derzko, N., S. P. Sethi, and G. L. Thompson. "Distrib- uted parameter systems approach to the optimal cattle ranching problem." Optimal Control Applications & Methods 1 (January-March 1980): 3-10.
Keith, J. E., and K. S. Lyon. "Valuing Wildlife Manage- ment: a Utah deer herd." West. J. Agr. Econ. 10(December 1985):216-22.
Loomis, J., M. Creel, and J. Cooper. Economic Benefits of Deer in California: Hunting and Viewing Values. Institute of Ecology Report #32, University of Cali- fornia, Davis (1989).
McCullough, D. R. The George Reserve Deer Herd: Pop- ulation ecology of a K-selected species. Ann Arbor: University of Michigan Press, 1979.
Mesterton-Gibbons, M. "On the Optimal Policy for Com- bining Harvesting of Predator and Prey." Natural Re- source Modeling 3(Summer 1988):63-90.
Metzgar, L. H., and E. Boyd. "Stability Properties in a Model of Forage-Ungulate-Predator Interactions." Natural Resource Modeling 3(Summer 1988):3-43.
Ragozin, D. L., and G. Brown Jr. "Harvest policies and nonmarket valuation in a predator-prey system." J. Environ. Econ. and Manage. 12(1985): 155-68.
Standiford, R. B., and R. E. Howitt. "Solving Empirical Bioeconomic Models: A Rangeland Management Ap- plication." Amer. J. Agr. Econ. 74(May 1992):421- 33.
Stevens, D. L. Director of Buckrun (Wildlife habitat by landowners), personal communication (February, 1992).
The Spokesman-Review. "Marketing Big Game." Spokane, Wa., Tuesday, November 12, 1991.
Tanner, J. T. "The stability and the int¡ growth rates of prey and predator populations," Ecol. 56(Summer 1975):855-67.
Tu, P. Introductory Optimization Dynamics. Springer-Ver- lag, New York, 1984.
Wenders, J. T. "Is Idaho about to run out of quality public elk hunting?" The Daily News, Pullman, Wa., Wednesday, September 25, 1991.
Wilen, J., and G. Brown Jr. "Optimal Recovery Paths for Perturbations of Trophic Level Bioeconomic Sys- tems." J. Environ. Econ. and Manage. 13(1986):225- 34.
at Queen M
ary, University of L
ondon on October 2, 2014
http://ajae.oxfordjournals.org/D
ownloaded from