original slaying dragons: limited evidence for … reptiles, the largest living lizard (the komodo...
TRANSCRIPT

ORIGINALARTICLE
Slaying dragons: limited evidence forunusual body size evolution on islands
Shai Meiri1*, Pasquale Raia2 and Albert B. Phillimore3
INTRODUCTION
Giant tortoises, enormous flightless birds and huge bears,
alongside minute deer, tiny lizards, dwarf elephants and, lately,
pygmy humans all spring to mind when the body sizes of
island vertebrates is discussed. Body size evolution on islands is
perceived to be fast (Lister, 1989; Millien, 2006) and has
produced extreme phenotypes, with the smallest or the largest
1Department of Zoology, Faculty of Life
Sciences, Tel Aviv University, 69978 Tel Aviv,
Israel, 2Dipartimento di Scienze della Terra,
Universita Federico II, 80138 Naples, Italy,3Division of Biology, Imperial College at
Silwood Park, Ascot SL5 7PY, UK
*Correspondence: Shai Meiri, Department of
Zoology, Faculty of Life Sciences, Tel Aviv
University, 69978 Tel Aviv, Israel.
E-mail: [email protected]
ABSTRACT
Aim Island taxa often attain forms outside the range achieved by mainland
relatives. Body size evolution of vertebrates on islands has therefore received
much attention, with two seemingly conflicting patterns thought to prevail: (1)
islands harbour animals of extreme size, and (2) islands promote evolution
towards medium body size (‘the island rule’). We test both hypotheses using body
size distributions of mammal, lizard and bird species.
Location World-wide.
Methods We assembled body size and insularity datasets for the world’s lizards,
birds and mammals. We compared the frequencies with which the largest or
smallest member of a group is insular with the frequencies expected if insularity is
randomly assigned within groups. We tested whether size extremes on islands
considered across mammalian phylogeny depart from a null expectation under a
Brownian motion model. We tested the island rule by comparing insular and
mainland members of (1) a taxonomic level and (2) mammalian sister species, to
determine if large insular animals tend to evolve smaller body sizes while small
ones evolve larger sizes.
Results The smallest species in a taxon (order, family or genus) are insular no
more often than would be expected by chance in all groups. The largest species
within lizard families and bird genera (but no other taxonomic levels) are insular
more often than expected. The incidence of extreme sizes in insular mammals
never departs from the null, except among extant genera, where gigantism is
marginally less common than expected under a Brownian motion null. Mammals
follow the island rule at the genus level and when comparing sister species and
clades. This appears to be driven mainly by insular dwarfing in large-bodied
lineages. A similar pattern in birds is apparent for species within orders. However,
lizards follow the converse pattern.
Main conclusions The popular misconception that islands have more than
their fair share of size extremes may stem from a greater tendency to notice
gigantism and dwarfism when they occur on islands. There is compelling evidence
for insular dwarfing in large mammals, but not in other taxa, and little evidence
for the second component of the island rule – gigantism in small-bodied taxa.
Keywords
Birds, dwarfism, evolution, gigantism, island biogeography, island rule, lizards,
mammals.
Journal of Biogeography (J. Biogeogr.) (2011) 38, 89–100
ª 2010 Blackwell Publishing Ltd www.blackwellpublishing.com/jbi 89doi:10.1111/j.1365-2699.2010.02390.x

species of many clades being insular (Hooijer, 1967; Berry,
1998; Greer, 2001; Glaw et al., 2006; Whittaker & Fernandez-
Palacios, 2007; Hedges, 2008; Losos & Ricklefs, 2009). For
example, the St Helena earwig, Labidura herculeana, the
Indonesian stick insect, Pharnacia serratipes, and the New
Zealand giant wetas (Deinacrida spp.) are probably the largest
representatives of their clades (Chown & Gaston, 2010).
Similarly, the world’s largest bat (Smith et al., 2003) is the
Philippine-endemic golden crowned flying fox, Acerodon
jubatus, the largest Quaternary bird was the Madagascan
elephant bird, Aepyornis maximus, and the largest raptor was
the New Zealand endemic Haast’s eagle, Harpagornis moorei
(Worthy et al., 2002; Murray & Vickers-Rich, 2004).
Among reptiles, the largest living lizard (the Komodo
dragon, Varanus komodoensis) and tortoises (the giant tor-
toises of Aldabra and the Galapagos, Geochelone gigantea and
Geochelone elephantopus) are insular endemics (Arnold, 1979;
Meiri, 2008). Islands also harbour the smallest members of
several clades: the smallest bird is believed to be the Cuban bee
hummingbird, Mellisuga helenaei (although Dunning, 2008b,
suggests that Thaumastura cora from mainland Peru is
smaller), and species of Caribbean Leptotyphlops and Sphaero-
dactylus are the world’s smallest snakes and lizards, respectively
(Hedges, 2008; Meiri, 2008). This perceived abundance of
insular size extremes is usually thought to be a response to the
low intensity of competition and predation both within and
across taxa: the absence of carnivorous mammals is most often
quoted as allowing the evolution of large size in birds (mainly
through the evolution of flightlessness; e.g. Bunce et al., 2005;
Murray & Vickers-Rich, 2004), reptiles (e.g. Case, 1978; Meiri,
2008) and small mammals (e.g. Angerbjorn, 1986; Adler &
Levins, 1994). Alternatively, this perceived pattern may simply
reflect an ascertainment bias, i.e. we may be more likely to
notice animals of extreme body size when they happen to live
on islands (Whittaker & Fernandez-Palacios, 2007).
By contrast, the island rule suggests that, rather than showing
size extremes on islands, insular populations should be closer to
the clade-wide median body size than their mainland counter-
parts (Lomolino, 1985, 2005). According to the island rule,
populations of small species will evolve larger size, populations
of large species will dwarf, and populations of average-sized
species will show little size evolution on islands (Lomolino, 2005;
Welch, 2009). Viewed at the clade level, the island rule predicts
stabilizing selection: on islands an individual should not be
either too large or too small. Once the optimum is reached (via
directional selection on the founding population), stabilizing
selection should maintain phenotypes around it. Interestingly,
an opposite pattern of disruptive selection and increased vari-
ance is often thought to prevail within populations on islands
(Van Valen, 1965; Scott et al., 2003; cf. Meiri et al., 2005a).
The island rule is thought to manifest the combined effects
of lower predation pressures on islands (Heaney, 1978),
character release in the absence of competitors (Dayan &
Simberloff, 1998) and the paucity of resources on islands
driving dwarfism in large-bodied forms (Lomolino, 2005).
Empirical evidence from terrestrial vertebrate studies provides
mixed support for this rule (Clegg & Owens, 2002; Boback &
Guyer, 2003; Lomolino, 2005; Meiri, 2007), with the best
support coming from data for mammals (Lomolino, 1985;
Price & Phillimore, 2007; Welch, 2009; but see Meiri et al.,
2004, 2006, 2008). Size evolution is often hypothesized to be
most drastic on small islands, with island area showing
complex interactions with body size (Heaney, 1978), but
empirical patterns are equivocal (e.g. Meiri et al., 2005b; Wu
et al., 2006; Schillaci et al., 2009).
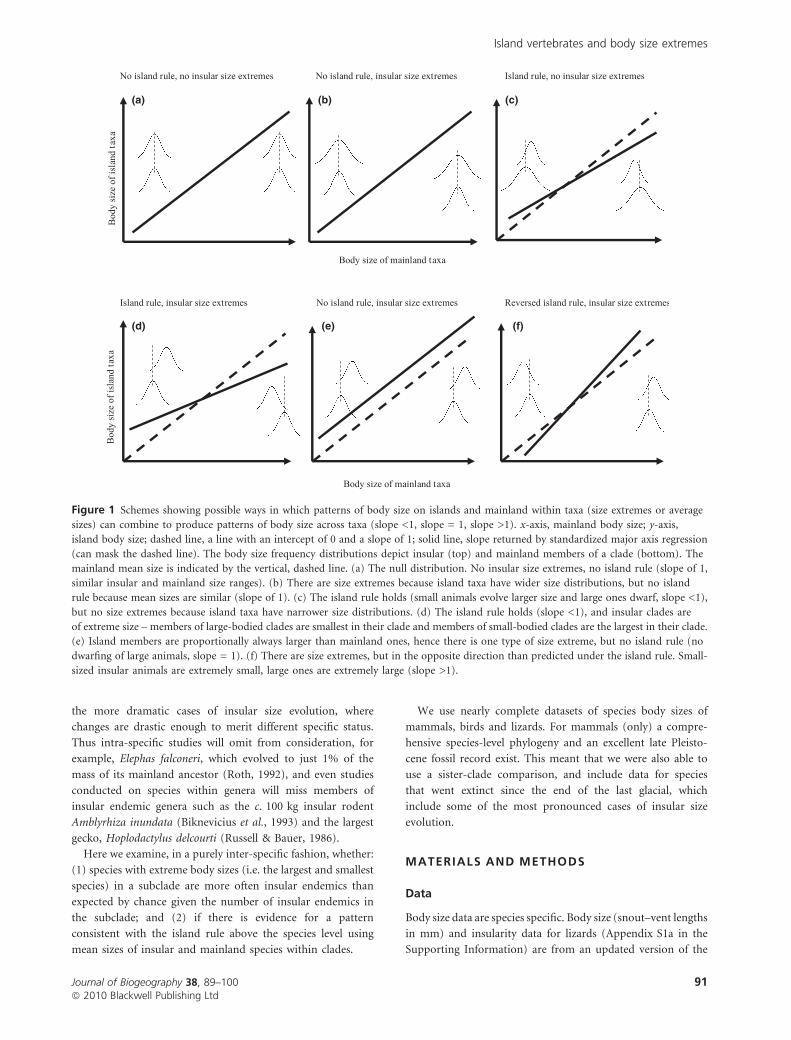
These two hypotheses, that islands should harbour extreme
sizes, and that they should harbour taxa that are closer to a clade-
wide mode, need not necessarily be contradictory (Fig. 1). If a
clade, such as mammals, has a single size attractor (e.g. Brown
et al., 1993) then members of a subclade may evolve to a size
extreme: insular members of a subclade of large-bodied animals
(e.g. elephants) will be the smallest within this subclade, but not
across the larger clade as a whole. Similarly, insular members of a
subclade of small-bodied animals (e.g. shrews) may be the largest
members of this subclade, but not the largest mammals overall.
If, however, each subclade has its own optimal size that its insular
members evolve towards (Lomolino, 2005) then islands should
harbour few size extremes (Fig. 1c).
The seemingly conflicting discussion of insular size extremes
on the one hand, and insular medium sizes on the other hand,
stems in part from the different phylogenetic and temporal scope
of studies dealing with them. Studies of size extremes are usually
conducted at the inter-specific level (Glaw et al., 2006; Hedges,
2008), and often deal with extinct taxa (Kurten, 1953; Sondaar,
1977; Steadman et al., 2002; Raia et al., 2003). The study of
evolution towards medium sizes usually involves intra-specific
studies of insular and mainland populations of extant species
(e.g. Lomolino, 1985; Boback & Guyer, 2003; Meiri, 2007).
Evolutionary processes above and below the species level
may differ through, for example, species sorting and adaptive
radiation in the former versus inter-island and island–
mainland gene flow in the latter (Jablonski, 2008). As far as
we are aware McClain et al. (2006) and Welch (2010) present
the only purely inter-specific studies of the island rule,
comparing mean sizes within genera of deep sea (= ‘insular’
in McClain et al.) and shallow sea (‘mainland’) species, rather
than using comparisons within single species. An argument for
extending tests of the island rule to the species level is that the
selection pressures thought to promote convergence on a
median body size for island populations should act similarly
on island species. The major difference between the two
scenarios is that there should be less gene flow between species
than between populations, meaning that evolution should
proceed more rapidly in the former. However, there is no
obvious reason why the type of selection and adaptive optima
should differ in these two contexts. A major advantage of
intra-specific studies of insular size evolution is that they
compare very closely related taxa (i.e. different populations
within a species), usually from areas that are in geographic
proximity. Intra-specific comparisons thus control for much of
the variation in size that is unrelated to insularity. Restricting
ourselves to intra-specific studies, however, we may be missing
S. Meiri et al.
90 Journal of Biogeography 38, 89–100ª 2010 Blackwell Publishing Ltd
the more dramatic cases of insular size evolution, where
changes are drastic enough to merit different specific status.
Thus intra-specific studies will omit from consideration, for
example, Elephas falconeri, which evolved to just 1% of the
mass of its mainland ancestor (Roth, 1992), and even studies
conducted on species within genera will miss members of
insular endemic genera such as the c. 100 kg insular rodent
Amblyrhiza inundata (Biknevicius et al., 1993) and the largest
gecko, Hoplodactylus delcourti (Russell & Bauer, 1986).
Here we examine, in a purely inter-specific fashion, whether:
(1) species with extreme body sizes (i.e. the largest and smallest
species) in a subclade are more often insular endemics than
expected by chance given the number of insular endemics in
the subclade; and (2) if there is evidence for a pattern
consistent with the island rule above the species level using
mean sizes of insular and mainland species within clades.
We use nearly complete datasets of species body sizes of
mammals, birds and lizards. For mammals (only) a compre-
hensive species-level phylogeny and an excellent late Pleisto-
cene fossil record exist. This meant that we were also able to
use a sister-clade comparison, and include data for species
that went extinct since the end of the last glacial, which
include some of the most pronounced cases of insular size
evolution.
MATERIALS AND METHODS
Data
Body size data are species specific. Body size (snout–vent lengths
in mm) and insularity data for lizards (Appendix S1a in the
Supporting Information) are from an updated version of the
No island rule, insular size extremes
Island rule, insular size extremes No island rule, insular size extremes
Island rule, no insular size extremes
Reversed island rule, insular size extremes
No island rule, no insular size extremes
Body size of mainland taxa
Body size of mainland taxa
Bod
y si
ze o
f is
land
tax
aB
ody
size
of
isla
nd t
axa
(a) (b) (c)
(d) (e) (f)
Figure 1 Schemes showing possible ways in which patterns of body size on islands and mainland within taxa (size extremes or average
sizes) can combine to produce patterns of body size across taxa (slope <1, slope = 1, slope >1). x-axis, mainland body size; y-axis,
island body size; dashed line, a line with an intercept of 0 and a slope of 1; solid line, slope returned by standardized major axis regression
(can mask the dashed line). The body size frequency distributions depict insular (top) and mainland members of a clade (bottom). The
mainland mean size is indicated by the vertical, dashed line. (a) The null distribution. No insular size extremes, no island rule (slope of 1,
similar insular and mainland size ranges). (b) There are size extremes because island taxa have wider size distributions, but no island
rule because mean sizes are similar (slope of 1). (c) The island rule holds (small animals evolve larger size and large ones dwarf, slope <1),
but no size extremes because island taxa have narrower size distributions. (d) The island rule holds (slope <1), and insular clades are
of extreme size – members of large-bodied clades are smallest in their clade and members of small-bodied clades are the largest in their clade.
(e) Island members are proportionally always larger than mainland ones, hence there is one type of size extreme, but no island rule (no
dwarfing of large animals, slope = 1). (f) There are size extremes, but in the opposite direction than predicted under the island rule. Small-
sized insular animals are extremely small, large ones are extremely large (slope >1).
Island vertebrates and body size extremes
Journal of Biogeography 38, 89–100 91ª 2010 Blackwell Publishing Ltd

dataset of Meiri (2008). Lizard taxonomy follows Uetz et al.
(2009).
Body mass data (in g) for birds are from Dunning (2008).
Where mass data were reported for multiple subspecies the mean
of these measurements was taken (intra-specific data were first
log10-transformed). For each species overall mean body mass
was estimated across means for female, male and unsexed birds.
We obtained data on avian insularity from McCall (1997), and
supplemented and verified them using Avibase (http://
avibase.bsc-eoc.org) and regional guides (Appendix S1b). Tax-
onomy follows Clements (1998), and we included species that
had been extirpated since the extinction of the dodo (17th
century). As we were interested in the effect of insularity on
birds, we excluded birds that forage at sea, which we defined as all
members of the Alcidae, Diomedeidae, Fregatidae, Hydrobat-
idae, Laridae, Pelecanidae, Pelecanoididae, Phaethontidae,
Phalacocoracidae, Procellariidae, Rynchopidae, Spheniscidae,
Stercoraridae, Sternidae and Sulidae families.
Body mass and insularity data for mammals are from the
2007 version of Smith et al. (2003) (data kindly provided by
Felisa Smith), supplemented with literature data (see Appen-
dix S1c for species for which we obtained data from sources
other than the Smith et al. database). Mammal insularity was
verified using Wilson & Reeder (2005), regional guides and
palaeontological accounts of fossil species. We use only fossil
mammals that went extinct since the end of the last glacial.
Only fossil species known to be confined to islands during the
last glacial were considered to be insular.
All analyses were conducted on log10-transformed measures
of body size, to bring the intra-class distribution of body size
closer to a normal distribution and to remove a relationship
between the mean and variance of a group (Lynch & Walsh,
1998).
Phylogeny
Complete species-level phylogenetic data exist only for mam-
mals, and we thus only use phylogenetic analysis on this group.
We used the species-level mammal supertree of Bininda-
Emonds et al. (2007). The tree was modified as follows: we
resolved taxonomic discrepancies between the tree and our
data using the taxonomy of Wilson & Reeder (2005), and
excluded species for which we had no size data and fossil
species whose island endemism was uncertain. Finally, where
the phylogenetic affinities of extinct species are well under-
stood we added them to the tree. Our modified tree includes
3961 species. Although this tree excludes some new phyloge-
netic data, it is the only comprehensive species-level tree
currently available for any vertebrate class.
Statistical analysis
Size extremes
For each of the four datasets we counted the number of genera
for which either the largest or the smallest species was insular.
Only genera having at least one insular member and one
continental member were included. We repeated this proce-
dure at the family and order levels (family level only for
lizards).
We then assessed whether the observed values departed from
the null expectation if insular status were randomly assigned
by randomly selecting n individuals per taxon (i.e. genus,
family or order), where n is the number of insular species in
that taxon. We then calculated the number of taxa for which
the largest or smallest representative had been randomly
assigned insular status. This process was repeated 10,000 times
to obtain a distribution of the null expectation for the
frequency of insular gigantism or dwarfism. We calculated the
proportion of the null distribution that was: (1) greater than or
equal to the observed value, and (2) smaller than or equal to
the observed value. The smaller of these proportions was
multiplied by 2 to give a two-tailed P-value.
Our randomization test for size extremes on islands should
be conservative for two reasons. First, we classify all species
that have at least one mainland population as ‘mainland’,
even though in some of these species the largest or smallest
populations may be insular (e.g. Kodiak brown bears, Ursus
arctos middendorffi). Second, if insular taxa are clustered
within a subclade (as is the case in mammals; Raia et al.,
2010), the variance in body size among insular members of a
subclade will be reduced, reducing the potential for the
evolution of size extremes. However, it is possible that the
ancestral members of subclades that colonize islands are
themselves biased in size with respect to the mainland
representatives of the subclade (e.g. they may be very large;
Lomolino, 1985) and this could generate an increased
incidence of insular size extremes without requiring further
evolution on islands. In the absence of adequate fossil data we
are unable to test this hypothesis.
Using the mammalian phylogeny we were able to compare
the observed incidence of size extremes with those generated
under a phylogenetically explicit null model. We conducted
1000 simulations of body size evolution across the whole
phylogeny following a Brownian motion (random walk with
constant variance) model using the evolve.phylo function in
the R library ‘ape’ (Paradis et al., 2004). A recent study on
body size evolution across the mammalian tree identified a
weak but significant signature of early burst evolution (i.e.
decelerating rates of phenotypic evolution; Cooper & Purvis,
2010). However, a constant-rate Brownian motion model did
not perform much worse, and in the context of our study
should represent an unbiased means of generating a phyloge-
netically explicit null expectation. The fossil record shows that
mammals have increased in body size through time according
to Cope’s rule (e.g., Alroy, 1998), which conflicts with a
Brownian model. Nonetheless, with respect to the hypotheses
being tested here, adding directionality (universally across
island and mainland lineages) to a random walk should not
bias our tests. For each simulation we quantified the incidence
of gigantism and dwarfism and in this manner generated a null
distribution. We then used two-tailed tests to establish the
S. Meiri et al.
92 Journal of Biogeography 38, 89–100ª 2010 Blackwell Publishing Ltd

probability of obtaining the observed incidences of gigantism
and dwarfism under the null Brownian model.
Mean size
We assessed whether the ‘island rule’ applies above the species
level. We calculated the mean log10 body size of insular and
mainland species within each genus. We tested the null
hypothesis that there are no differences between patterns of
mainland and insular size evolution across genera (Welch,
2009) by examining the slope of the island versus mainland
taxon means using standardized major axis (= reduced major
axis) regression. A slope <1 is expected under the island rule,
which predicts gigantism in small-bodied taxa and dwarfism in
large-bodied ones and a reduction of variance amongst insular
taxa as compared with their mainland counterparts (Price &
Phillimore, 2007). Different taxa within a taxonomic level will
vary in age, meaning that the expected variance of phenotypes
among them is likely to vary. This heterogeneity of expected
variance violates an assumption of standardized major axis
(SMA) regression. We therefore used a distribution-free
variant of the SMA test, as proposed by Welch (2009). The
SMA correlation coefficient (r) between x + y (where x is body
size on the mainland and y is body size on an island) and x ) y
was the test statistic. The observed correlation coefficient was
then compared with that observed when the identity of insular
means was randomized with respect to x and y (Welch, 2009).
Ten thousand randomizations were conducted and the two-
tailed P-value was the smaller of twice the proportion of
randomized correlation coefficients that were either greater
than or equal to or less than or equal to the observed
correlation. If the observed SMA correlation between x + y
and x ) y was significantly greater than expected at random
this would support the island rule.
At the family level we adjusted the protocol to control for
clade species richness, so that rather than each species
contributing equally to the family mean, each species contrib-
uted equally to the genus mean and each genus then
contributed equally to the family mean. The same procedure
was repeated at the order level with the addition of taking the
mean across families in an order. Tests with species treated
independently (i.e. simple means of all species in a family or
order) gave qualitatively the same results (not shown).
Using the modified mammal tree we located nodes that
represented a partition between a solely insular and a solely
mainland clade and estimated the mean body size for each
clade. Clades including polytomies were excluded. The sister
clades we identified are shown as Appendix S2. As the clades
involved in each island versus mainland comparison vary in
age, then under a Brownian motion model of evolution the
expected variance of the mean values of these clades is expected
to vary, violating an assumption of SMA regression (Welch,
2009). Consequently, we estimated the statistical significance
of a departure from a 1:1 relationship between the size of
island and mainland taxa by applying 10,000 randomizations
of the identity of insular and mainland clades using the
distribution-free randomization approach of Welch (2009)
described above. All statistical analyses were conducted in R (R
Development Core Team, 2008) and all tests were two-tailed.
RESULTS
Lizards
Of 5380 lizards in our dataset, 1636 are insular endemics
(Appendix S1a). The largest lizard in 9 of 19 families is an
insular endemic (see size frequency histograms in Appen-
dix S3a), while the median expected number is five (P = 0.07).
At the genus level the number of largest insular species is no
different from the null expectation. The number of insular
species that are the smallest species in their genus or family is
likewise not statistically different from that expected by chance
(Table 1).
The SMA slope between the taxon mean masses on islands
versus mainland does not depart significantly from 1 at the
genus level (Fig. 2a). At the family level the slope estimate of
1.24 [95% confidence interval (CI) = 1.02–1.50] (Fig. 2b) is
significantly steeper than 1 using both SMA (r = 0.49,
P < 0.05) and the distribution-free randomization test
(P < 0.05, similar to the pattern depicted in Fig. 1f).
Table 1 Actual and expected numbers of taxa in which insular
members are the largest or the smallest, in different taxa at dif-
ferent taxonomic levels.
Clade Level
Number
of clades Largest� Smallest�
Lizards Genus 85 30 (36) 36 (35)
Lizards Family 19 9* (5) 7 (5)
Birds Genus 223 90** (71) 73 (71)
Birds Family 86 24 (21) 21 (21)
Birds Order 19 3 (3) 6 (4)
Mammals Genus 91 29 (36) 38 (35)
Mammals Family 39 9 (11) 10 (12)
Mammals Order 12 4 (3) 2 (3)
Mammals� Genus 101 35 (40) 41 (39)
Mammals� Family 47 14 (14) 12 (14)
Mammals� Order 14 5 (3) 3 (3)
Mammals [BM] Genus 91 28* (35) 38 (35)
Mammals [BM] Family 39 9 (11) 10 (11)
Mammals [BM] Order 12 4 (3) 2 (3)
Mammals� [BM] Genus 99 33 (37) 41 (37)
Mammals� [BM] Family 46 15 (14) 12 (14)
Mammals� [BM] Order 14 6 (3) 3 (3)
*, **Denote statistical significance at the a = 0.1 and 0.01 levels,
respectively.
�The median expected values derived from our randomizations are in
parentheses.
�Including extinct species. [BM] denotes analyses conducted using
Brownian motion simulations rather than randomizations.
Note that because we only have phylogenetic data for c. 92% of
mammalian species observed numbers of size extremes can differ
between non-phylogenetic and phylogenetic analyses.
Island vertebrates and body size extremes
Journal of Biogeography 38, 89–100 93ª 2010 Blackwell Publishing Ltd

Birds
Of 8069 bird species in our dataset, 1347 are insular endemics
(we have maximum mass data for all, and for 7552, including
1268 insular endemic species, we have data on mean size;
Appendix S1b). In 90 out of 223 avian genera the largest
member of the clade is an insular endemic. This is significantly
more than expected under our randomizations (median
expected = 71; P < 0.01, Table 1). However, at the family
and order levels (see size frequency histograms in Appen-
dix S3b) the largest member of a clade is no more often an
insular endemic than expected at random. The frequency with
which the smallest member of a clade is an insular endemic
does not depart from the null expectation for any taxonomic
level. Moreover, when we repeated the analysis at the genus
level using the maximum rather than mean body size (data are
the maximum reported for a species in Dunning, 2008),
neither the frequency of gigantism nor dwarfism departed
from the null expectation (67 observed insular maxima across
219 genera, median expected = 71, P = 0.53).
There was no evidence for the island rule at the genus and
family level in birds, with slopes equal to 1.00 and 0.98,
respectively (Fig. 3a,b). The slope at the order level, however,
was significantly shallower than 1 (SMA slope = 0.76, 95%
CI = 0.66–0.88, r = )0.70, P < 0.01; Prandomization < 0.01;
Fig. 3c), even when ratites are excluded (SMA slope = 0.78,
95% CI = 0.64–0.95, r = )0.56, P < 0.05; Prandomization <
0.05).
Mammals
Of 3961 extant mammal species, 670 are insular endemics (778
of 4213 when extinct species are included). The frequency of
insular endemics that are either the largest or smallest
members of their clades does not depart from the null
expectation derived by randomization at any taxonomic level,
either including or excluding extinct taxa (Table 1, Appen-
dix S3c).
In agreement with the results from randomizations, the
frequency of gigantism and dwarfism tended not to exceed the
null expectation generated under a single-rate Brownian model
on the mammalian phylogeny. This was true for all taxonomic
levels, both including and excluding extinct species, except for
Continental mean SVL (log10 scale)
Insu
lar m
ean
SV
L (lo
g10
sca
le) (a) (b)
Figure 2 Standardized major axis regression
of mean body size [log10-transformed
snout–vent length (SVL) in mm] on islands
versus continents for species in lizard genera
(a) and families (b). The dashed line has a
slope of 1 and an intercept of 0. The solid line
represents the standardized major axis
regression slope estimate.
Continental mean mass (log10 scale)
Insu
lar m
ean
mas
s (lo
g 10
scal
e) (a) (b) (c)
Figure 3 Standardized major axis regression of mean body size (log10-transformed mass, in g) on islands versus continents for species in
bird genera (a), families (b) and orders (c). The dashed line has a slope of 1 and an intercept of 0. The solid line represents the standardized
major axis regression slope estimate.
S. Meiri et al.
94 Journal of Biogeography 38, 89–100ª 2010 Blackwell Publishing Ltd

extant mammals at the genus level, where the frequency of
gigantism was marginally lower than expected (P = 0.1).
Mammals show a general tendency to follow the island rule
at the genus level. The observed slope of mean insular clade
mass versus mean continental clade mass is significantly less
than 1 for genera both including and excluding extinct species
(extant only: SMA slope = 0.96, 95% CI = 0.93–0.99, r =
)0.24, P < 0.05; Prandomization < 0.05, Fig. 4a; extant + extinct:
slope = 0.92, 95% CI = 0.89–0.96, r = )0.41, P < 0.01;
Prandomization < 0.05, Fig. 4d; see also Fig. 1c). The slope for
families is not significantly different from 1 (slope = 1.01, 95%
CI = 0.95–1.08, and 0.95, 95% CI = 0.89–1.01, with and
without extinct species, respectively, Fig. 4b,e). The slope for
orders likewise is not significantly different from 1
(slope = 1.01, 95% CI = 0.74–1.37, and 0.87, 95% CI =
0.76–1.05, with and without extinct species, respectively;
Fig. 4c,f).
When analyses were conducted using the mammalian
phylogeny, regressing mean body sizes within insular clades
on the mean size within their mainland sister clades returned a
slope significantly shallower than 1 using SMA (n = 91,
slope = 0.931, 95% CI = 0.877–0.987, r = )0.25, P < 0.01),
but not using randomizations (P = 0.155; Fig. 5a). Restricting
the analysis to insular versus mainland sister species, we get a
slope consistent with the island rule using both tests (n = 57,
slope = 0.870, 95% CI = 0.81–0.93, r = )0.49, P < 0.01;
Prandomization < 0.05, Fig. 5b).
DISCUSSION
The largest species in bird and lizard taxa tend to be insular
more frequently than expected were insular status assigned to
species at random. Interestingly, however, this holds within
bird genera (but not when species maximum rather than mean
masses are used), whereas lizards only show gigantism among
families. Furthermore, the lizard giants are often the result of
insular radiations (e.g. Hoplodactylus, Gallotia, Cyclura) on
islands lacking mammalian carnivores (here New Zealand, the
Canaries and the Antilles, respectively). Such absence of
mammalian predation has been hypothesized to promote
lizard gigantism by allowing for more foraging, as less time is
spent hiding from predators, and by enabling lizards to evolve
the role of top predators (Case, 1982; Meiri, 2008). In birds,
the largest members of genera that are insular seem to be quite
Continental mean mass (log10 scale)
Insu
lar m
ean
mas
s (lo
g 10
scal
e)
(a) (b) (c)
(d) (e) (f)
Figure 4 Standardized major axis regression of mean body size (log10-transformed mass, in g) on islands versus continents for species in
mammalian genera (a, d), families (b, e) and orders (c, f). Plots (a)–(c) are for extant taxa only and (d)–(f) include extinct taxa (see
Materials and Methods). The dashed line has a slope of 1 and an intercept of 0. The solid line represents the standardized major axis
regression slope estimate.
Island vertebrates and body size extremes
Journal of Biogeography 38, 89–100 95ª 2010 Blackwell Publishing Ltd

evenly distributed between continental shelf, volcanic and
continental plate islands (e.g. the Philippines, New Guinea,
Tasmania, Hawaii and New Zealand). This suggests that release
from predation on islands has often promoted gigantism in
lizards and perhaps gigantism associated with flightlessness in
birds (e.g. the New Zealand moas, Sylviornis; Russell, 1877;
McNab, 1994), but perhaps a different mechanism drove
gigantism among birds that retained their power of flight (e.g.
high population density; Blondel, 2000).
Size extremes in insular mammals show no departure from
null expectations. However, mammals conform to the island
rule at the genus level, especially when fossil species are
included. This pattern also emerges when we compare sister
species, a better test of size evolution than comparing clade
members ignoring their phylogenetic affinities.
In all three taxa, deviations from a slope of 1 seem to stem
from small (mammals and birds) or large (lizards) members of
generally large-bodied forms (e.g. small elephants, artiodactyls,
ratites and ducks, large iguanas). Extinct mammals show the
most drastic cases of dwarfism. However, many of the recently
extinct insular lizards and birds are extremely large (Pregill,
1986; Blondel, 2000) and extinction in these taxa was probably
much more prevalent on islands than on mainlands, whereas
extinctions of large mammals were common in mainland
settings (Barnosky et al., 2004). Thus in early Holocene times
there were giant insular birds in orders now exhibiting an
overall tendency for small sizes on islands. Indeed very large,
recently extinct, insular birds include members of the Falcon-
iformes (e.g. Amplibuteo, Harpagornis moorei, Circus eylesi;
Worthy et al., 2002; Suarez & Olson, 2007) and Strigiformes
(e.g. Tyto riveroi, Ornimegalonyx oteroi; Alcover & McMinn,
1994), ratites (Dinornis, Aepyornis; Worthy et al., 2002),
Anseriformes (e.g. Cygnus falconeri and the ‘very large Hawaii
goose’; Milberg & Tyrberg, 1993; Paxinos et al., 1999),
Ciconiiformes (Threskiornis solitarius, perhaps a Pelecaniform;
Mourer-Chauvire et al., 1995), Gruiformes (Diaphorapteryx
hawkinsii, perhaps Aptornis; Holdaway, 1989; Raia, 2009),
Galliformes (e.g. Megavitiornis, perhaps Sylviornis; Steadman,
2006) and Columbiformes (Raphus cucullatus, Pezophaps
solitaria, Natunaornis gigoura; Steadman, 2006; Worthy,
2000). Thus, the apparent tendency for smaller mean size in
insular members of large-bodied avian orders may be a result
of human-mediated extinction (Steadman, 2006; Pavia, 2008)
rather than a feature of natural insular evolution.
The finding that bird genera harbour more insular giants
than expected by chance is surprising, given that the mean size
of species within bird genera does not seem to differ between
islands and mainlands (Fig. 2a). A thorough study of the avian
subfossil record may even reveal that when extinct taxa are
included a similar pattern will be revealed within families and
orders.
A common explanation for insular gigantism in birds and
reptiles (e.g. elephant birds, moas, Komodo dragons and the
giant skinks of Cape Verde and New Caledonia) is that they
have evolved large size on islands with no mammalian
competitors or predators (Russell, 1877; Case, 1978; McNab,
2002; Meiri, 2008). While we view this as a highly likely
explanation, we are not sure it can explain our finding that
island birds tend to be the largest members of their genera
more often than is expected by chance. Our impression from
the data is that these largest members of avian genera are
mostly found on large islands, rich in bird, reptile and often
mammal species. Being classified as congenerics of mainland
forms, insular giants seldom occupy niches vacated by
mainland mammals, and usually differ relatively little from
the size of their mainland relatives. Currently we are unable to
sufficiently explain why this pattern prevails, or why it holds
only for birds, and only within genera, and note that species
maximum sizes [probably an inferior size measure because it is
more sensitive to sample size (Meiri, 2007) but representing
about 7% more species (Dunning, 2008)] do not show the
same trend.
The evidence we find for the island rule in mammals
emerges primarily via insular dwarfism in large taxa. Curi-
ously, the tendency of large mammals to dwarf on islands (see
also Raia et al., 2010), which is corroborated by our phylo-
genetic tests, and when fossils are included, is also linked to the
absence of predators and competitors, and seems more
prevalent in herbivores than in carnivores (Raia & Meiri,
2006). McNab (2002) has claimed that gigantism in insular
Continental mean mass (log10 scale)
Insu
lar m
ean
mas
s (lo
g10
sca
le) (a) (b)
Figure 5 Standardized major axis regression
of mean body size (log10-transformed mass,
in g) on islands versus continents within (a)
mammalian sister clades and (b) a subset of
the data in (a): only sister species [only clades
in (a) where both mainland and insular
sample sizes are 1]. The dashed line has a
slope of 1 and an intercept of 0. The solid line
represents the standardized major axis
regression slope estimate.
S. Meiri et al.
96 Journal of Biogeography 38, 89–100ª 2010 Blackwell Publishing Ltd

birds is more likely in herbivorous taxa. Additionally, in lizards
insularity is often associated with large size and herbivory
(Troyer, 1983; Meiri, 2008). Gigantism may be favoured where
resources are abundant (McClain et al., 2006), and the size of
large carnivorous vertebrates may depend on the size of
available prey; thus islands lacking large herbivorous mammals
are likely also to lack large carnivores. Because mammals can
grow much larger than either birds or lizards, one might say
that even the largest avian and reptilian predators, Haast’s
eagle and the Komodo dragon, are not large predators
compared with large mammalian carnivores. Thus low preda-
tion and competition pressures on islands may tend to
produce both relatively small mammals and relatively large
lizards.
The nature of the islands that we study, in terms of their
area and isolation, climate, geology (e.g. whether they are part
of the continental shelf, part of a tectonic plate or volcanic)
and biogeographic settings (e.g. realm, ocean) may all affect
the mode of size evolution (Meiri et al., 2005b; Schillaci et al.,
2009). Moreover, these attributes may interact with the
ecological attributes of the different taxa themselves, such as
their functional group or guild, their diet and microhabitat
preferences, as well as their behaviour (Case, 1978; McNab,
1994; Raia & Meiri, 2006) in shaping the way that size evolves.
Such attributes of islands and taxa offer promising avenues for
research into size evolution on islands.
CONCLUSIONS
The evidence that insular conditions favour the evolution of
extreme sizes within clades is restricted to gigantism in lizard
families and, perhaps, bird genera, but is not found in these
groups at other taxonomic levels, and neither does it apply to
mammals. Furthermore, large insular lizards seem often to
result from radiations on oceanic islands with no mammalian
carnivores whereas giant insular bird species are scattered over
highly variable set of islands (S.M., unpublished). We thus
think it is unlikely that these two patterns have a common
explanation.
The island rule applies in a statistical sense to mammalian
species within genera, and between sister species. Biologically,
however, while dwarfism in large insular mammals seems
prevalent, we find no evidence for the second component of
the island rule – a general tendency for gigantism in small-
bodied mammals. Within small-sized lizard families insular
species are smaller than mainland ones, and within large-
bodied families insular species are larger than mainland ones,
reversing the island rule. These findings are consistent with
intra-specific studies (Lomolino, 1985; Meiri, 2007), suggesting
that similar selection pressures may operate to produce
patterns seen both within and between species. More compre-
hensive fossil data are needed to resolve the pattern of size
evolution in island birds. The different courses of size
evolution on islands taken by different taxa imply an
important role for contingency, as animals differing in their
ecology respond differently to the selective forces imposed by
agents such as resource abundance, predation and competi-
tion, which in turn differ across different islands.
ACKNOWLEDGEMENTS
We thank Liz Butcher and Barbara Sanger from the Michael
Way Library for their enormous help in obtaining literature
sources for data used in this work. Felisa Smith kindly
provided us with the latest version of the ‘Integrating
Macroecological Pattern and Processes across Scales’ (IMMPS)
working group mammalian mass database. We thank Ian
Owens for valuable discussion and Mark Lomolino, Craig
McClain, John Welch and two anonymous referees for very
helpful comments on earlier versions of this manuscript.
REFERENCES
Adler, G.H. & Levins, R. (1994) The island syndrome in rodent
populations. Quarterly Review of Biology, 69, 473–490.
Alcover, J.A. & McMinn, M. (1994) Predators of vertebrates on
islands. BioScience, 44, 12–18.
Alroy, J. (1998) Cope’s rule and the dynamics of body mass
evolution in North American fossil mammals. Science, 280,
731–734.
Angerbjorn, A. (1986) Gigantism in island populations of
wood mice (Apodemus) in Europe. Oikos, 47, 47–56.
Arnold, E.N. (1979) Indian Ocean giant tortoises: their sys-
tematics and island adaptations. Philosophical Transactions
of the Royal Society B: Biological Sciences, 286, 127–145.
Barnosky, A.D., Koch, P.L., Feranec, R.S., Wing, S.L. & Shabel,
A.B. (2004) Assessing the causes of late Pleistocene extinc-
tions on the continents. Science, 306, 70–75.
Berry, R.J. (1998) Evolution of small mammals. Evolution on
islands (ed. by P.R. Grant), pp. 35–50. Oxford University
Press, Oxford.
Biknevicius, A.R., McFarlane, D.A. & MacPhee, R.D.E. (1993)
Body size in Amblyrhiza inundata (Rodentia: Caviomorpha),
an extinct megafaunal rodent from the Anguilla Bank, West
Indies: estimates and implications. American Museum Nov-
itates, 3079, 1–25.
Bininda-Emonds, O.R.P., Cardillo, M., Jones, K.E., MacPhee,
R.D.E., Beck, R.M.D., Grenyer, R., Price, S.A., Vos, R.A.,
Gittleman, J.L. & Purvis, A. (2007) The delayed rise of
present-day mammals. Nature, 446, 507–512.
Blondel, J. (2000) Evolution and ecology of birds on islands:
trends and prospects. Vie et Milieu, 50, 205–220.
Boback, S.M. & Guyer, C. (2003) Empirical evidence for an
optimal body size in snakes. Evolution, 57, 345–351.
Brown, J.H., Marquet, P.A. & Taper, M.L. (1993) Evolution of
body size, consequences of an energetic definition of fitness.
The American Naturalist, 142, 573–584.
Bunce, M., Szulkin, M., Lerner, H.R.L., Barnes, I., Shapiro, B.,
Cooper, A. & Holdaway, R.N. (2005) Ancient DNA provides
new insights into the evolutionary history of New Zealand’s
extinct giant eagle. PLoS Biology, 3, 44–46.
Island vertebrates and body size extremes
Journal of Biogeography 38, 89–100 97ª 2010 Blackwell Publishing Ltd

Case, T.J. (1978) A general explanation for insular body size
trends in terrestrial vertebrates. Ecology, 59, 1–18.
Case, T.J. (1982) Ecology and evolution of the insular giant
chuckawallas, Sauromalus hispidus and Sauromalus varius.
Iguanas of the world: their behavior, ecology and conservation
(ed. by G.M. Burghardt and A.S. Rand), pp. 184–212. Noyes
Publications, Park Ridge, NJ.
Chown, S.L. & Gaston, K.J. (2010) Body size variation in in-
sects: a macroecological perspective. Biological Reviews, 85,
139–169.
Clegg, S.M. & Owens, I.P.F. (2002) The ‘island rule’ in birds:
medium body size and its ecological explanation. Proceed-
ings of the Royal Society B: Biological Sciences, 269, 1359–
1365.
Clements, J.F. (1998) Birds of the world: a checklist, 5th edn.
Ibis, Vista, CA.
Cooper, N. & Purvis, A. (2010) Body size evolution in mam-
mals: complexity in tempo and mode. The American Nat-
uralist, 175, 727–738.
Dayan, T. & Simberloff, D. (1998) Size patterns among com-
petitors: ecological character displacement and character
release in mammals, with special reference to island popu-
lations. Mammal Review, 28, 99–124.
Dunning, J.B. (2008) CRC handbook of avian body masses, 2nd
edn. CRC Press, London.
Glaw, F., Kosuch, J., Henkel, F.W., Sound, P. & Bohme, W.
(2006) Genetic and morphological variation of the leaf-
tailed gecko Uroplatus fimbriatus from Madagascar, with
description of a new giant species. Salamandra, 42, 129–144.
Greer, A.E. (2001) Distribution of maximum snout–vent
length among species of scincid lizards. Journal of Herpe-
tology, 35, 383–395.
Heaney, L.R. (1978) Island area and body size of insular
mammals: evidence from the tri-colored squirrel (Callosci-
urus prevosti) of Southeast Asia. Evolution, 32, 29–44.
Hedges, S.B. (2008) At the lower size limit in snakes: two new
species of threadsnakes (Squamata, Leptotyphlopidae: Lep-
totyphlops) from the Lesser Antilles. Zootaxa, 1841, 1–30.
Holdaway, R.N. (1989) New Zealand’s pre-human avifauna
and its vulnerability. New Zealand Journal of Ecology,
12(Suppl.), 11–25.
Hooijer, D.A. (1967) Indo-Australian insular elephants.
Genetica, 38, 143–162.
Jablonski, D. (2008) Species selection: theory and data. Annual
Review of Ecology, Evolution, and Systematics, 39, 501–524.
Kurten, B. (1953) On the variation and population dynamics
of fossil and recent mammal populations. Acta Zoologica
Fennica, 76, 1–122.
Lister, A.M. (1989) Rapid dwarfing of red deer on Jersey in the
last interglacial. Nature, 342, 539–542.
Lomolino, M.V. (1985) Body size of mammals on islands: the
island rule reexamined. The American Naturalist, 125, 310–
316.
Lomolino, M.V. (2005) Body size evolution in insular verte-
brates: generality of the island rule. Journal of Biogeography,
32, 1683–1699.
Losos, J.B. & Ricklefs, R.E. (2009) Adaptation and diversifi-
cation on islands. Nature, 457, 830–836.
Lynch, M. & Walsh, B. (1998) Genetics and analysis of quan-
titative traits. Sinauer Associates, Sunderland, MA.
McCall, R.A. (1997) Biological, geographical and geological
factors influencing biodiversity on islands. PhD Thesis, Uni-
versity of Oxford, Oxford.
McClain, C.R., Boyer, A.G. & Rosenberg, G. (2006) The island
rule and the evolution of body size in the deep sea. Journal of
Biogeography, 33, 1578–1584.
McNab, B.K. (1994) Energy conservation and the evolution of
flightlessness in birds. The American Naturalist, 144, 628–
642.
McNab, B.K. (2002) Minimizing energy expenditure facilitates
vertebrate persistence on oceanic islands. Ecology Letters, 5,
693–704.
Meiri, S. (2007) Size evolution in island lizards. Global Ecology
and Biogeography, 16, 702–708.
Meiri, S. (2008) Evolution and ecology of lizard body sizes.
Global Ecology and Biogeography, 17, 724–734.
Meiri, S., Dayan, T. & Simberloff, D. (2004) Body size of
insular carnivores: little support for the island rule. The
American Naturalist, 163, 469–479.
Meiri, S., Dayan, T. & Simberloff, D. (2005a) Variability and
sexual size dimorphism in carnivores: testing the niche
variation hypothesis. Ecology, 86, 1432–1440.
Meiri, S., Simberloff, D. & Dayan, T. (2005b) Insular carnivore
biogeography: island area and mammalian optimal body
size. The American Naturalist, 165, 505–514.
Meiri, S., Dayan, T. & Simberloff, D. (2006) The generality of
the island rule reexamined. Journal of Biogeography, 33,
1571–1577.
Meiri, S., Cooper, N. & Purvis, A. (2008) The island rule: made
to be broken? Proceedings of the Royal Society B: Biological
Sciences, 275, 141–148.
Milberg, P. & Tyrberg, T. (1993) Naıve birds and noble savages
– a review of man-caused prehistoric extinctions of island
birds. Ecography, 16, 229–250.
Millien, V. (2006) Morphological evolution is accelerated
among island mammals. PLoS Biology, 4, 1863–1868.
Mourer-Chauvire, C., Bour, R. & Ribes, S. (1995) Was the
solitaire of Reunion an ibis? Nature, 373, 568.
Murray, P.F. & Vickers-Rich, P. (2004) Magnificent mihirungs.
The colossal flightless birds of the Australian dreamtime.
Indiana University Press, Bloomington, IN.
Paradis, E., Claude, J. & Strimmer, K. (2004) APE: analyses of
phylogenetics and evolution in R language. Bioinformatics,
20, 289–290.
Pavia, M. (2008) The evolution dynamics of the Strigiformes in
the Mediterranean islands with the description of Aegolius
martae n. sp. (Aves, Strigidae). Quaternary International,
182, 80–89.
Paxinos, E.E., James, H.F., Olson, S.L., Sorenson, M.D., Jack-
son, J. & Fleischer, R.C. (1999) mtDNA from fossils reveals a
radiation of Hawaiian geese recently derived from the
S. Meiri et al.
98 Journal of Biogeography 38, 89–100ª 2010 Blackwell Publishing Ltd

Canada goose (Branta canadensis). Proceedings of the
National Academy of Sciences USA, 99, 1399–1404.
Pregill, G. (1986) Body size of insular lizards: a pattern of
Holocene dwarfism. Evolution, 40, 997–1008.
Price, T.D. & Phillimore, A.B. (2007) Reduced major axis
regression and the island rule. Journal of Biogeography, 34,
1998–1999.
R Development Core Team (2008) R: a language and
environment for statistical computing. R Foundation for
Statistical Computing, Vienna. Available at: http://www.
r-project.org/.
Raia, P. (2009) Gigantism. Encyclopedia of islands (ed. by R.G.
Gillespie and D.A. Clague), pp. 372–376. University of
California Press, Berkeley, CA.
Raia, P. & Meiri, S. (2006) The island rule in large mammals:
paleontology meets ecology. Evolution, 60, 1731–1742.
Raia, P., Barbera, C. & Conte, M. (2003) The fast life of a
dwarfed giant. Evolutionary Ecology, 17, 293–312.
Raia, P., Carotenuto, F. & Meiri, S. (2010) One size does not fit
all: no evidence for an optimal body size on islands. Global
Ecology and Biogeography, 19, 475–484.
Roth, V.L. (1992) Inferences from allometry and fossils:
dwarfing of elephants on islands. Oxford surveys in evolu-
tionary biology, Vol. 8 (ed. by D. Futuyma and J. Anto-
novics), pp. 259–288. Oxford University Press, Oxford.
Russell, A.P. & Bauer, A.M. (1986) Le gecko geant Hoplo-
dactylus delcourti et ses relations avec le gigantisme et
l’endemisme insulaire chez les Gekkonidae. Mesogee, 46, 25–
28.
Russell, I.C. (1877) The giant birds of New Zealand. The
American Naturalist, 11, 11–21.
Schillaci, M.A., Meijaard, E. & Clark, T. (2009) The effect of
island area on body size in a primate species from the Sunda
Shelf Islands. Journal of Biogeography, 36, 362–371.
Scott, S.N., Clegg, S.C., Blomberg, S.P., Kikkawa, J. & Owens,
I.P.F. (2003) Morphological shifts in island-dwelling passe-
rines: the roles of generalist foraging and niche expansion.
Evolution, 57, 2147–2156.
Smith, F.A., Lyons, S.K., Morgan Ernest, S.K., Jones, K.E.,
Kaufman, D.M., Dayan, T., Marquet, P.A., Brown, J.H. &
Haskell, J.P. (2003) Body mass of late Quaternary mammals.
Ecology, 84, 3403.
Sondaar, P.Y. (1977) Insularity and its effects on mammal
evolution. Major patterns of vertebrate evolution (ed. by M.K.
Hecht, P.C. Goody and B.M. Hecht), pp. 671–707. Plenum
Press, New York.
Steadman, D.W. (2006) Extinction and biogeography of tropical
Pacific birds. University of Chicago Press, Chicago.
Steadman, D.W., Pregill, G.K. & Burley, D.V. (2002) Rapid
prehistoric extinction of iguanas and birds in Polynesia.
Proceedings of the National Academy of Sciences USA, 99,
3673–3677.
Suarez, W. & Olson, S.L. (2007) The Cuban fossil eagle Aquila
borrasi Arredondo: a scaled-up version of the great black-
hawk Buteogallus urubitinga (Gmelin). Journal of Raptor
Research, 41, 288–298.
Troyer, K. (1983) The biology of iguanine lizards: present
status and future directions. Herpetologica, 39, 317–328.
Uetz, P., Goll, J. & Hallermann, J. (2009) The JCVI reptile
database. Available at: http://reptile-database.org/ (accessed
23 July 2009).
Van Valen, L.M. (1965) Morphological variation and the width
of the ecological niche. The American Naturalist, 99, 377–
390.
Welch, J.J. (2009) Testing the island rule: primates as a case
study. Proceedings of the Royal Society B: Biological Sciences,
276, 675–682.
Welch, J.J. (2010) The ‘island rule’ and deep-sea gastropods:
re-examining the evidence. PLoS ONE, 5, e8776.
doi:10.1371/journal.pone.0008776.
Whittaker, R.J. & Fernandez-Palacios, J.M. (2007) Island bio-
geography: ecology, evolution, and conservation, 2nd edn.
Oxford University Press, Oxford.
Wilson, D.E. & Reeder, D.M. (2005) (eds) Mammal species of
the world, 3rd edn. Johns Hopkins University Press, Balti-
more, MD.
Worthy, T.H. (2000) The fossil megapodes (Aves: Megapo-
diidae) of Fiji with descriptions of a new genus and two new
species. Journal of the Royal Society of New Zealand, 30, 337–
364.
Worthy, T.H., Holdaway, R. & Morris, R. (2002) The lost world
of the moa: prehistoric life of New Zealand. Indiana Univer-
sity Press, Bloomington, IN.
Wu, Z., Li, Y. & Murray, B.R. (2006) Insular shifts in body size
of rice frogs in the Zhoushan Archipelago, China. Journal of
Animal Ecology, 75, 1071–1080.
SUPPORTING INFORMATION
Additional supporting information may be found in the online
version of this article:
Appendix S1 Body size and insularity status for (a) lizards,
(b) birds and (c) mammals.
Appendix S2 Insular and mainland mammal sister clades.
Appendix S3 Size frequency histograms of species within (a)
lizard families, (b) bird orders, (c) mammal orders.
As a service to our authors and readers, this journal provides
supporting information supplied by the authors. Such mate-
rials are peer-reviewed and may be reorganized for online
delivery, but are not copy-edited or typeset. Technical support
issues arising from supporting information (other than
missing files) should be addressed to the authors.
Island vertebrates and body size extremes
Journal of Biogeography 38, 89–100 99ª 2010 Blackwell Publishing Ltd

BIOSKETCHES
Shai Meiri is a senior lecturer at the Department of Zoology, Tel Aviv University. He is interested in trait evolution, the tempo and
mode of evolution, the evolutionary implications of biogeography and vertebrate evolution.
Pasquale Raia is a post-doctoral research fellow at the Department of Earth Science, University of Naples Federico II, and a
member of the Center for Evolutionary Ecology based at Rome III University. He is interested in large mammal evolution, both at
the organismal and community levels, in response to climate change and the effect of ecological interactions.
Albert Phillimore is an Imperial College Junior Research Fellow and is interested in the influences of ecology on evolution and
speciation.
Author contributions: A.B.P., P.R. and S.M. conceived the ideas and collected the data; A.B.P. analysed the data; S.M. led the writing.
Editor: K.C. Burns
S. Meiri et al.
100 Journal of Biogeography 38, 89–100ª 2010 Blackwell Publishing Ltd