overcoming non-selective nodulation of lessertia by soil-borne rhizobium in the presence of...
TRANSCRIPT

REGULAR ARTICLE
Overcoming non-selective nodulation of Lessertia by soil-bornerhizobium in the presence of inoculant mesorhizobium
Macarena Gerding & Graham William O’Hara &
John Gregory Howieson & Lambert Bräu
Received: 25 June 2013 /Accepted: 28 February 2014# Springer International Publishing Switzerland 2014
AbstractBackground and aims Legumes of the South Africangenus Lessertia, along with their microsymbionts, wereintroduced into the Western Australia wheatbelt. Theyachieved poor establishment followed by weak summersurvival. This was caused in part by low levels ofnodulation with the inoculant strains, and by ineffectivenodulation with naturalized strains –an example of non-selective nodulation. The aims of this work were toassess Lessertia spp. symbiotic promiscuity, to studythe effect of increased doses of an effective inoculant
strain (WSM3565) with L. herbacea, and to study thecompetitive ability and symbiotic performance of dif-ferent Mesorhizobium strains nodulating L. diffusa.Methods A glasshouse experiment was set up to evalu-ate the ability of L. diffusa, L. capitata, L. herbacea andL. excisa to nodulate with inoculants under current usein Western Australia. To assess competitive ability twofield experiments were set up at Karridale, WesternAustralia. L. herbacea was inoculated with the strainWSM3565 at different doses and L. diffusa was inocu-lated with ten differentMesorhizobium strains. Rhizobiawere re-isolated from nodules and their identity con-firmed through PCR fingerprinting and sequencing oftheir partial dnaK.Results There were differences in promiscuity betweendifferent Lessertia spp., where L. herbacea proved to behighly promiscuous under controlled conditions.Increasing the inoculation dose of L. herbacea withWSM3565 did not improve establishment and survivalof the legume in the field. Although WSM3565 noduleoccupancy improved from 28 to 54 % with higher dosesof inoculation, none of the treatments increasedL. herbacea yield over the inoculated control. The inoc-ulation of L. diffusa with the strains WSM3598, 3636,3626 and 3565 resulted in greater biomass productionthan the uninoculated control. These strains were able tooutcompete resident rhizobia and to occupy a high(>60 %) proportion of lateral root nodules. Thenaturalised strains that achieved nodulation were identi-fied as R. leguminosarum.Conclusion The high numbers of resident rhizobia andtheir ability to rapidly nodulate Lessertia spp. are likely
Plant SoilDOI 10.1007/s11104-014-2081-1
Responsible Editor: Euan K. James.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11104-014-2081-1) contains supplementarymaterial, which is available to authorized users.
M. Gerding (*)Facultad de Agronomía, Universidad de Concepción, Av.Vicente Méndez 595, Chillán, Chilee-mail: [email protected]
G. W. O’Hara : J. G. HowiesonCentre for Rhizobium Studies, Murdoch University,South Street, Murdoch, WA 6150, Australia
G. W. O’Harae-mail: [email protected]
J. G. Howiesone-mail: [email protected]
L. BräuSchool of Life and Environmental Sciences,Deakin University, Burwood, Australiae-mail: [email protected]

to be the main reasons for the low nodule occupancyachieved by some effective inoculant strains withL. diffusa and L. herbacea. Strains WSM 3636 and3598 were very competitive on nodule occupancy andtogether with WSM 3565, WSM 3612 and WSM3626,effective on nodule formation and plant growth ofL. diffusa despite the high numbers of resident soilrhizobia. These strains and L. diffusa have potential tobe introduced as exotic legumes species and rhizobiastrains to Western Australia.
Keywords Mesorhizobium .Lessertiadiffusa .Lessertiaherbacea . Non-selective nodulation . Symbioticnitrogen fixation
Introduction
Rhizobia inoculant strains are exposed to diverse envi-ronmental and biological factors that may constrain theirability to colonise new niches and therefore reduce over-all symbiotic performance and plant productivity(Graham 2008; Zahran 1999). One well documentedconstraint is the presence of indigenous or naturalisedsoil rhizobia able to form nodules with the introducedlegume (Dowling and Broughton 1986; Thies et al.1991), which may compromise the inoculation outcome,and result in suboptimal nitrogen fixation, or in the worstcase scenario, a total displacement of the effective inoc-ulant by ineffective soil bacteria (Denton et al. 2002;Dowling and Broughton 1986; Slattery and Coventry1993, 1999; Thies et al. 1991). This kind of associationis termed non-selective nodulation, where nodule occu-pancy can be linked to numerical superiority or positionaladvantage of soil rhizobia (Yates et al. 2011).
Lessertia spp., along with their microsymbionts, wereintroduced into Western Australia in an attempt to over-come stresses such as dryland salinity and low soil pH,and to increase the diversity of perennial species inagricultural systems. The introduced species had beencollected from similar edaphic areas in South Africa;L. diffusa was collected from Southern Africa Northernand Eastern Cape which correspond to Succulent Karooand Fynbos biomes and L. herbacea from the Fynbosbiome in the Western Cape (Gerding et al. 2012). Quiteunexpectedly, Lessertia spp. showed an overall poorestablishment and survival in different sites of theWestern Australian wheat belt (Gerding et al. 2013)caused in part by poor nodulation by the inoculant strains
and by ineffective nodulation with naturalised Rhizobiumleguminosarum.
Naturalised soil rhizobia are difficult to displace(Brockwell et al. 1995; Howieson et al. 2008), especial-ly in stressful environments, where they may hold thecompetitive advantage through adaptation to those harshconditions (Dowling and Broughton 1986; Howiesonand Ballard 2004; Sessitsch et al. 2002; Thies et al.1992). Some approaches to overcome ineffective nodu-lation by soil rhizobia are to increase inocula concentra-tion (Brockwell and Bottomley 1995; Dowling andBroughton 1986; Mårtensson 1990; Roughley et al.1993), to select a host genotype capable of effectivenodulation (Brockwell et al. 1982; Demezas andBottomley 1986; Drew and Ballard 2009; Duodu et al.2005; Yates et al. 2008), to select inoculant strains with ahigher competitive ability (Aguilar et al. 2001; Amarger1981a; Hungria et al. 2003) and to choose symbiosesthat are selective (Yates et al. 2008, 2011).
Lessertia diffusa was the species that performed bestacross all the sites evaluated in the Western Australianwheatbelt but still showed a high incidence of nodulationby background bacteria (Gerding et al. 2013). This le-gume was also able to nodulate and increase dry weightwith ten different Mesorhizobium strains in glasshouseexperiments (Gerding et al. 2012), which provided anopportunity to test the symbiotic performance of differentstrains in the field. L. herbacea, on the other hand, despiteshowing good results in glasshouse experiments, did notestablish well at any of the sites. It was hypothesised thatfor L. herbacea an increase in the inoculation rate wouldenrich the number of live cells on the seed and rhizo-sphere, and therefore increasing the chances of the inoc-ulant to “outcompeting” naturalised rhizobia for infectionsites.
This work reports on the symbiotic promiscuity ofLessertia spp., and specifically on the response to in-creased doses of an effective inoculant strain with thelegume L. herbacea, and on the competitive ability andsymbiotic performance of different L. diffusa strains inthe field.
Materials and methods
Nodulation of Lessertia spp. by commercial inoculants
The ability of L. diffusa, L. capitata, L. herbacea andL. excisa to nodulate with 17 inoculants of current use in
Plant Soil

Western Australia was assessed in a glasshouse experi-ment. The inoculants used are listed in Table 1.Although the strains from Lebeckia sp. (WSM4187),Rhynchosia sp. (WSM3937) and the Microvirga spp.strains (WSM3693 and WSM3557) are not commercialinoculants, they were included in the experiment asthese plants have been introduced into the WesternAustralian wheat belt as alternative perennial legumesjust as Lessertia spp. (Ardley et al. 2012; Howiesonet al. 2013) and are likely to be sown in the sameagro-ecological areas.
The inoculant strains were obtained from theWesternAustralian Soil Microbiology (WSM) culture collection.Bacterial cultures were grown on half lupin-agar (½ LA)plates (Howieson et al. 1988) for 3 to 7 days at 28 °C.Seeds of the Lessertia spp. were scarified, sterilized thengerminated in 1.5 % (w/v) water agar plates. Followingradicle emergence, the seedlings were planted in aclosed 250 ml vial system filled with 125 ml of asteamed sand mix consisting of two parts of river sandand three parts of leached yellow sand [modified fromHowieson et al. (1995)]. Each vial contained twoLessertia species. Initially, two seedlings of each specieswere transplanted into the vial and after 3 weeks werethinned to one plant per species. After planting theseedlings, 10 ml of sterile nutrient solution devoid ofnitrogen [0.31 gL−1 MgSO4.7H2O; 0.21 gL
−1 KH2PO4;0.44 gL−1 K2SO4; 0.06 gL−1 FeEDTA; 0.05 gL−1
CaSO4; 0.116 mgL−1 H3BO3 0.116; 0.0045 mgL−1
Na2MoO4.2H2O; 0.134 mgL−1 ZnSO4.7H2O;0.01 mgL−1 MnSO4,4H2O and 0.06 mgL−1
CoSO4.7H2O (Howieson 1995)] and 20 ml of steriledeionized water were added to each vial.
Three days after planting each plant was inoculatedapproximately 109 cells ml−1 (estimated by optical den-sity (OD600nm) of 1.0) of a cell suspension obtainedfrom fresh colonies suspended in 1 % (w/v) sucrosesolution. There were three replicates for each of theinoculants, and an uninoculated control was provided.
This experiment was carried out under axenic con-ditions in a temperature controlled phytotron at 22 °Cfor 8 weeks. At harvest, plants were carefully removedfrom the vials and washed with tap water. Nodulationwas assessed and nodule characteristics were record-ed. To verify strain identity, two nodules per plantwere collected to re-isolate the bacteria and performPCR fingerprint to confirm their identity using theprimer RPO1 (5′ AATTTTCAAGCGTCGTGCCA 3′)(Richardson et al. 1995).
Field experiments
The field experiments were carried out on BoathaughEstate, Karridale, Western Australia (34°08′27.68″S;115°10′35.62″E) in August 2009. The paddock historyincluded pasture fallow in 2005 and 2006, rye grass-clover mix in 2007, with clover inoculated withWSM1325, and winter wheat and millet (summer) in2008. The soil was a grey sand duplex, with a pH of 6.1(CaCl2), 2.45 % soil organic carbon, 30 mg Kg−1 ofnitrate and 9 mg Kg−1 of ammonium. Monthly rainfallduring the experiments was: 149,8 mm in August,171,8 mm in September, 28,8 mm in October and37,8 mm in November, 2009.
Prior to sowing into moist soil the site was mowedand sprayed with glyphosate (1.5 L ha−1) to reduce thenumber of weeds. Soil samples were randomly collectedto e s t ima t e the popu l a t i on o f ind igenousR. leguminosarum bv. trifolii, R. leguminosarum bv.viciae, Sinorhizobium meliloti and Mesorhizobiumciceri bv. biserrulae through the most-probable-number (MPN) technique (Brockwell 1982) usingTrifolium subterraneum (subterranean clover), Pisumsativum (Field pea), Medicago sativa (lucerne) andBiserrula pelecinus respectively as trap plants.
Field experiment 1: effect of rate of inoculationof WSM3565 on symbiotic performancewith L. herbacea
The host species chosen for this trial was L. herbacea,which proved to be a promiscuous species in the glass-house experiment. The chosen inoculant wasWSM3565, a strain that was able to significantly in-crease L. herbacea biomass under glasshouse conditions(Gerding et al. 2012) and which had not been testedunder field conditions.
Seed inoculation
Seeds of L. herbacea were scarified and surface-sterilised for 3 min in 4 % (w/v) bleach, 1 min in 70 %(v/v) ethanol followed by six rinses in sterile de-ionisedwater. Seeds were aseptically dried in a laminar flowcabinet and stored in sterile paper bags until being slurryinoculated, lime pelletized and sown. The seed germi-nation percentage was assessed by placing three repli-cates of 100 sterilised seeds on 1.5 % (w/v) water agarplates and incubating them for 72 h at room temperature.
Plant Soil

The inoculant strain WSM3565 was grown on ½ LAagar plates at 28 °C for 5 days. One loop full of freshcells was inoculated into a 250 ml conical flask contain-ing 50 ml of ½ LA broth which was incubated on ashaker at 28 °C for 7 days until it contained approxi-mately 109 cells ml−1 (OD600nm=1.0). Eight ml of thebroth culture were inoculated into 10 g of sterile peatcontained in 30 ml sterile polycarbonate capped tubes.The inoculated peat was mixed and incubated at 28 °Cfor another 10 days until the cell number was approxi-mately 109 cells g−1. Cell number in the peat wasassessed by colony count through the Miles and Misradrop plate method (Singleton 1992) on ½ LA mediausing 10-fold dilutions.
Inoculant rates using this peat culture ranged from0.5 to 10 times the recommended peat based inoculationrate for medium size seeds (250 g of peat per 50 Kg ofseeds), equivalent to 104 rhizobial cells seed−1 (Deakeret al. 2004; Lupwayi et al. 2000). Twelve hours prior tosowing, the seeds were pelleted by mixing the amount
of peat needed to achieve each of the target doses with3 ml of 1 % methocel adhesive solution. The mixture ofmethocel and peat was applied at a rate of 200 μl for 3 gof seeds and mixed until seeds were evenly covered.Then, in a sterile 150 ml polycarbonate container theywere mixed with 1 g of dolomite lime until evenlycoated. For the maximum rate of inoculation (50 g ofpeat kg−1), the 3 g of seeds were inoculated with 300 μland were carefully mixed with dolomite to avoid seedclustering. Each of the treatments were separated intoreplicates, packed in paper envelopes and stored at 4 °Cuntil sowing (14 h later).
A sample of 100 peat-inoculated seeds per treatmentwas randomly selected to perform cell counts immedi-ately after inoculation. The peat inoculated seeds weresuspended in 50 mL solution of 0.1 % Na4P2O7.10H2Oand shaken on a rotary shaker at 200 rpm for 20 min(Pijnenborg et al. 1991). Samples were diluted in a 10 -fold dilution series in sterile saline solution (0.89 % w/vNaCl). Rhizobia cell counts were performed using the
Table 1 Legume rhizobial inoculants under current use in Western Australia and evaluated in this study
Strain Species Inoculationgroup
Target hosts
RR1128 Sinorhizobium meliloti AL Medicago sativa, M. littoralis andM. tornata
WSM1115 Sinorhizobium medicae AM Medicago truncatula, M. polymorpha, M. murex, M. sphaerocarpus,M. rugosa, M. scutellata, M. rotata
TA1 R. leguminosarum bv. trifolii B Trifolium repens, T. pratense, T. fragiferum, T. hybridum, T. alexandrinum,T. glomeratum and T. dubium
WSM1325 R. leguminosarum bv. trifolii C Trifolium subterraneum var. subterraneum, Trifolium subterraneumvar.yanninicum, Trifolium subterraneum var.brachycalycinum,T. cherleri, T. clypeatum, T. glanduliferum, T. hirtum, T. incarnatum,T. michelanium, T. purpureum, T. vesiculosum.
SU303 R. leguminosarum bv. viciae E Lathyrus cicera, Pisum sativum, Vicia narbonensis, V. sativa, V. villosa ssp.dasicarpa, V. benghalensis and V. ervilia
WSM1455 R. leguminosarum bv. viciae F Vicia faba and Lens culinaris
CC829 Mesorhizobium loti D Lotus pedunculatus
WSM1497 Mesorhizobium ciceri bv. biserrulae Biserrula pelecinus
WSM1284 Mesorhizobium ciceri bv. biserrulae Biserrula pelecinus
CC1192 Mesorhizobium ciceri N Cicer arietinum
WSM471 Bradyrhizobium sp. S Ornithopus compressus, O. pinnatus, O. sativus × compressus andO. perpusillus
WU425 Bradyrhizobium sp. G Lupinus angustifolius, L. albus, L. luteus and L. consentinii
WSM2598 Methylobacterium sp. L Lotononis bainesii
WSM3693 Microvirga zambiensis Listia angolensis
WSM3557 Microvirga lotononidis Listia angolensis
WSM3937 Burkholderia sp. Rhynchosia spp.
WSM4187 Burkholderia sp. Lebeckia spp.
Plant Soil

Miles and Misra drop plate count method. Plates wereincubated for 5 days at 28 °C.
Experimental design and sowing
The experiment was a complete randomized block de-sign with five replicates (blocks) per treatment. Thetreatments were: Uninoculated control, 1×104, 2×104,4×104 and 1×105 cells seed−1. Each plot consisted of asingle row 1.5 m long and spaced 1 m apart. The seedswere sown by hand at 1.5 cm depth at a rate of 1 g ofseed per m. Plots were hand fertilised after sowing withphosphate fertiliser (15 % available P) at a rate of 12 gper plot (equivalent to 150 kg ha−1).
Evaluations
Five plants (when present) from each plot were sampledat week 12 after sowing to assess plant dry weight andnodule occupancy. Three nodules were carefully re-moved from each plant, surface sterilised and the bacte-ria re-isolated in ½ LA media. The position of thenodules on the root was also recorded. The shoots ofthe plants sampled were dried at 60 °C for 48 h thenweighed.
All the isolates were PCR fingerprinted to confirmtheir identity using the primer RPO1 (5′ AATTTTCAAGCGTCGTGCCA 3′) (Richardson et al. 1995). Thecycling parameters were a cell lysis step of 5 min at95 °C, followed by 5 cycles at 94 °C for 30 s; 50 °C for10 s and 72 °C for 1:30 min; and then 35 cycles at 94 °Cfor 30 s, 55 °C for 25 s and 72 °C for 1:30 min and afinal extension at 72 °C for 5 min. The PCR productswere electrophoresed in 2 % (w/v) agarose gels andbands were visualized in a UV transilluminator. In caseswhere fingerprints did not match that of the inoculant,the banding patterns were further analysed to construct acladogram and to determine the number of distinctstrains. The proportion of nodule occupancy was deter-mined for each of the resident strains as well as forWSM3565.
The identity of the resident strains was estimated atthe species level by sequencing the variable 3′ endfragment of the protein encoding gene dnaK (330 bp)using the primers TSdnak2 (5′GTACATGGCCTCGCCGAGCTTCA3′) and TSdnak3 (5′AAGGAGCAGCAGATCCGCATCCA3′) (Stępkowski et al. 2003).The cycling parameters were: 7 min at 95 °C followedby 35 cycles of 94 °C for 30 s, 62 °C for 30 s and 72 °C
for 45 s and a final hold at 72 °C for 5 min. Theamplification of the gene sequence (330 bp dnaK)was verified by agarose gel electrophoresis in 1.5 %(w/v) agarose gels pre-stained with 1:10,000 SYBR ®Safe DNA gel stain (Invitrogen). The marker used was1Kb DNA ladder (Promega G5711). Electrophoresiswas carried out in tanks buffered with 1 × TAE(40 mM Tris-Acetate, 1 mM EDTA, pH 8.0) at100 V for 3 h.
Field experiment 2: competitive ability of differentstrains of Mesorhizobium for nodulation of Lessertiadiffusa
The plant chosen for this experiment was L. diffusa as itachieved the highest establishment percentages at somesites in the Wheat belt (Gerding et al. 2013). The inoc-ulant strains are indicated in Table 2. The 10 strains hadpreviously increased L. diffusa biomass in comparisonto uninoculated plants in a glasshouse experiment(Gerding et al. 2012).
Seed inoculation
Seed scarification, disinfection and coating were asdescribed for field experiment 1. For each strain, 3 g ofseeds were inoculated with 200 μl of a mixture of peatin 3 ml of 1 % methocel. Due to variations in thenumber of rhizobial cell g−1 of peat within strains,the amount of peat was adjusted to achieve the stan-dard inoculation rate for small seeded legumes of 103
cells seed−1 (Deaker et al. 2004). The inoculated seedswere then coated with 1 g of dolomite lime. Thereplicates of each treatment were packed separatelyin sterile paper bags and sown 14 h later. A sample of50 peat inoculated seeds per treatment was taken toassess rhizobia survival on the seed as described infield experiment 1.
Experimental design and sowing
The experiment was a complete randomized block de-sign with 11 treatments [ten inoculant strains (Table 2)and an uninoculated control] and 5 replicates (blocks)per treatment. Each plot consisted of a single row 1.5 mlong, with a spacing of 1 m between rows. The sowingwas by hand at a rate of 1 g of seed per m. Lines werefertilised after sowing with 12 g of phosphate fertiliser(15 % available P).
Plant Soil

Evaluations
Five individual plants were sampled from each line12 weeks after sowing for nodulation and dry mattermeasurements. Shoots were dried at 60 °C for 48 h thenweighed. Three nodules were removed from each plant,surface sterilised and the bacteria re-isolated in ½ LAmedia as described in field experiment 1. Nodules wereseparated in tap root nodules and lateral root nodules.The Mesorhizobium inoculant strains were initially dif-ferentiated from soil strains by their growth rate in ½ LAmedia, as all the inoculant strains used in this experi-ment required 5 or more days to form 1 mm colonies,whereas the commercial inoculants and the soil rhizobiaable to nodulate Lessertia in previous experiments(Gerding et al 2013) were fast growing rhizobia (2–3 days to form 1 mm colonies). A sub-sample of threeisolates from each of the plots were RPO1-PCRfingerprinted to confirm their identity. Nodule occupan-cy (%) was the criterion to assess competitiveness foreach of the strains (Brockwell et al. 1982).
Statistical analyses
To determine significant differences between treatmentsin the percentage of established plants and plant dryweight, a one-way analysis of variance was performedwith the statistics programme SPSS 13.0 for Windows(2004). When differences were determined, the treat-ment means were compared using the Fisher’s Leastsignificant difference (LSD) test (P≤0.05).
The banding patterns for the isolates were analysedusing the software BIOCapt (ALYS Technologies,
Lausanne). The genetic distance between isolates wascalculated using AFPL SURV version 1.0 (Vekemans2002) and the UPGMA cluster analyses were performedusing the NEIGHBOR application from the PHYLIPsoftware package. The cladograms were visualized inMEGA4 (Tamura et al. 2007).
The dnaK sequences of the distinct strains wereanalysed in Gene Tool Lite 1.0 (2000) software(Doubletwist, Inc., Oakland, CA, USA) and inMEGA4 (Tamura et al. 2007) as described in fieldexperiment n° 1.
Results
Nodulation of Lessertia spp. by commercial inoculants
There were clear differences in promiscuity between thedifferent Lessertia species (Table 3). Lessertia excisawas not able to nodulate with any of the commercialinoculants of current use in Western Australia. Lessertiaincana nodulated with the clover inoculants strains TA1and WSM1325. Lessertia. capitata and L. diffusa hadsimilar nodulation patterns as they were both nodulatedby the Sinorhizobium strains RR1128 and WSM1115and with Rhizobium leguminosarum bv trifolii strainsTA1 and WSM1325.
Lessertia herbacea was able to nodulate with 13 ofthe 17 inoculants tested. It was able to nodulate witheve ry S inorh i zob ium spp . and Rhi zob iumleguminosarum inoculant. It formed nodules with mostof the Mesorhizobium spp. strains except forMesorhizobium loti strain CC829 whose original host
Table 2 Mesorhizobium noculant strains for Lessertia diffusa used in field trial 2
Strain Host plant Geographic origin Source (Year isolated)
WSM2566 L. pauciflora Rooiberg pass, Western Cape SK Yates and Real (2002)
WSM2623 L. annularis Beaufort West, Western Cape NK Yates and Real (2002)
WSM3270 L. annularis Middelburg, Eastern Cape NK Law (2004)
WSM3495 L. capitata Komkans, Western Cape SK Howieson et al. (2004)
WSM3626 L. diffusa Kamieskroon, Northern Cape SK Howieson et al. (2004)
WSM3636 L. capitata Komkans, Western Cape SK Howieson et al. (2004)
WSM3565 L. diffusa Grotto Bay, Western Cape F Howieson et al. (2004)
WSM3598 L. herbacea Citrusdal, Western Cape F Howieson et al. (2004)
WSM3612 L. excisa Lamberts Bay, Western Cape F Howieson et al. (2004)
WSM3898 L. excisa Lamberts Bay, Western Cape F Howieson et al. (2004)
BIOMES: F Fynbos, NK Nama Karoo, SK Succulent Karoo
Plant Soil
is Lotus pedunculatus. None of the Bradyrhizobium sp,Methylobacterium sp and Burkholderia sp. strains wereable to nodulate L. herbacea nor any of the otherLessertia spp.
In the four Lessertia species able to nodulate with thecommercial inoculants, nodules were indeterminate,white and apparently ineffective. There were some dif-ferences in nodule morphology. The strains RR1128(S. meliloti), TA1 and WSM1325 (R. leguminosarumbv. trifolii) and strains WSM1497 and WSM1284(M. ciceri bv. biserrulae) formed elongated nodules,whereas strains WSM1115 (S. medicae) and CC1192(M. ciceri) formed fan shaped nodules.
The identity of the inoculant strains was verifiedthrough RPO1-PCR fingerprints in most of the cases.The only case were isolation was not possible was fromL. herbacea nodulating with the Listia angolensisstrains WSM3693 and WSM3557, which formedpseudo-nodules
Field experiments
The soil population of R. leguminosarum bv. trifoliinodulating T. subterraneum, estimated through theMPN technique, was 2 (0.45–4.2)×105 cells g−1 of soil.
No nodulation of V. sativa, M. sativa and B. pelecinuswas recorded at any soil dilution.
Field experiment 1: effect of rate of inoculationof WSM3565 on symbiotic performancewith L. herbacea
Viable rhizobia cells on seeds
The germination (%) of L. herbacea seed was 72.2 %.The population of rhizobia on the seed surface afterinoculation was slightly lower than the potential numberof cells per seed. For the inoculation rate of 2.5 g of peatper Kg of seed the number of rhizobia cells per seed wasof 7.2×103, which corresponded to 72 % of the expect-ed rhizobia population. For the inoculation rates of 5, 10and 50 g Kg−1, the rhizobia cell counts per seed were1.6×104 (80 % cell survival), 3.7×104 (92.5 % cellsurvival) and 8.7×104 (87 % cell survival) respectively.
Plant population and biomass production
The percentage of established plants ranged between 28and 47 %. There was no effect of the different concen-trations of the inoculant strain (WSM3565) (P>0.05) on
Table 3 Nodulation of Lessertia spp. by commercial rhizobial inoculants
Strain Species L.capitata L.diffusa L.excisa L.herbacea L.incana
RR1128 Sinorhizobium meliloti + + − + −WSM1115 Sinorhizobium medicae + + − + −TA1 R. leguminosarum bv. trifolii + + − + +
WSM1325 R. leguminosarum bv. trifolii − + − + +
SU303 R. leguminosarum bv. viciae − − − + −WSM1455 R. leguminosarum bv. viciae − − − + −CC829 Mesorhizobium loti − − − − −WSM1497 M. ciceri bv. biserrulae − − − + −WSM1284 M. ciceri bv. biserrulae − − − + −CC1192 Mesorhizobium ciceri − − − + −WSM471 Bradyrhizobium sp. − − − − −WU425 Bradyrhizobium sp. − − − − −WSM2598 Methylobacterium sp. − − − − −WSM3693 Microvirga zambiensis − − − + −WSM3557 Microvirga lotononidis − − − + −WSM3937 Burkholderia sp. − − − − −WSM4187 Burkholderia sp. − − − − −
+ All plants nodulated, − No plants nodulated
Plant Soil
the percentage of established L. herbacea plants at week12 after sowing. There were no differences (P>0.05) inshoot dry weight at different inoculation rates. The shootdry weights of the uninoculated plants were equivalentto those inoculated with increasing doses of WSM3565(Fig. 1). It is important to note that there was highvariability in plant size within plots and treatments,and that no clear signs of nitrogen deficiency (chloroticfoliage) were evident.
Nodule occupancy
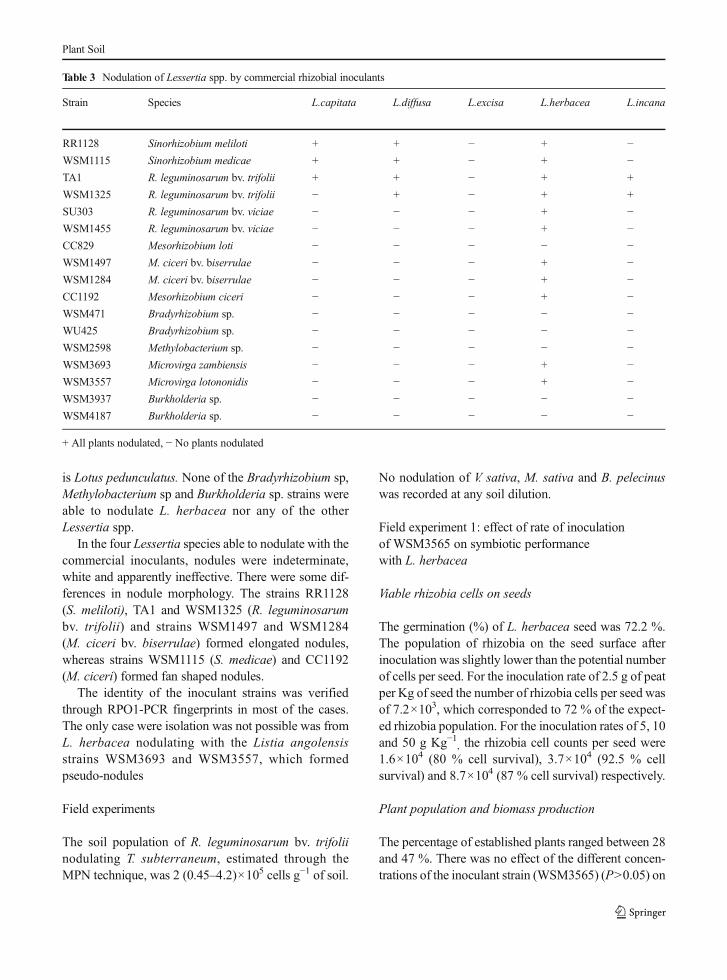
Of the 197 isolates obtained from L. herbacea nodulesacross all inoculation rates, only 78 isolates (39.5 %)matched the inoculant strain WSM3565 RPO1-PCRfingerprint. The other 119 isolates were grouped as 12distinct strains (A to L) based on their RPO1 bandingpatterns (available as supplementary material).
The nodule occupancy achieved by the inoculantstrain WSM3565 (in white, Fig. 2) ranged between 28and 31 % for the three lowest inoculation rates (7.2×103, 1.6×104 and 3.7×104 cells seed−1) and increased to54 % with the highest inoculation rate. The percentageof nodule occupancy of each of the soil naturalisedstrains (in shades of gray, Fig. 2) was different in thefive treatments, with B dominating.
Several soil strains were present in only some of thetreatments, and in a low proportion of nodules (less than3 %) and were usually represented by 1 or 2 isolates
(Strains from F to L). Strains A, B and C were thedominant resident strains and were present in every plot.
Nodule occupancy according to distribution of nodules
Overall, the percentage of nodules on the tap root occu-pied by WSM3565 was 70 % compared to only 17.7 %
Fig. 1 Shoot dry weights of L. herbacea inoculated at fourinoculation rates of strain WSM3565 (equivalent to 2.5, 5, 10and 50 g of peat kg−1 seed). Vertical lines correspond to thestandard error of means. No difference between treatments wasdetected through ANOVA (P≤0.05)
Fig. 2 Percentage nodule occupancy byMesorhizobium inoculantWSM3565 (Inwhite) and 12 soil strains (A to L; in shades of gray)in L. herbacea inoculated with: 1) 7×103 cells seed−1; 2) 1.6×104
cells seed−1; 3) 3.7×104 cells seed−1 4) 9.8×104 cells seed−1 and 5)Uninoculated control
�Fig. 3 Neighbor joining tree based on the sequencing of thepartial dnaK gene. The type strains sequences in the phylogramwere obtained from GenBank (accession number in parentheses).M.: Mesorhizobium; S.: Sinorhizobium; R.: Rhizobium and B.:Bradyrhizobium. Strains in bold correspond to naturalised soilrhizobia isolated from Lessertia herbacea at Karridale. The num-ber of isolates and their origin are indicated in brackets. Strains N1to N8 correspond to rhizobia isolated from Lessertia spp. in theWestern Australian wheatbelt (Gerding et al 2013). Bootstrapvalues are indicated on branches only when higher than 50 %
Plant Soil

Plant Soil

of the lateral roots. It is important to note that 45.7 % ofthe nodules on the tap root were desiccated due to lowrainfall periods that caused the top soil to dry, whichcorresponded to the clayey layer of the duplex soil,.Some of these dry nodules (25 %) were rehydrated inlaboratory and the rhizobia could be recovered, corre-sponding in every case to WSM3565.
Sequencing of dnaK gene
A single DNA fragment of 283 bp representing thepartial dnaK gene was amplified for the 12 soilnaturalised strains isolated from L. herbacea. Themajority of the strains clustered in the Rhizobiumgenus clade with a high bootstrap support (87 %)(Fig. 3). From these, nine of the strains (A, B, C, D,E, F, G, J and L) were closely related to the speciesR. leguminosarum, and to strains from sites from theWestern Australian wheat belt (Buntine, Katanning,Muresk and Newdegate) able to nodulate Lessertiaspp. in previous experiments (Gerding et al 2013),which are also included in this tree. Only strain I,representing two isolates, clustered apart from thisgenus and its dnaK sequence did not show similaritieswith any known species.
Field experiment 2: competitive ability of differentstrains of Mesorhizobium nodulating Lessertia diffusa
Viable rhizobia cells on seeds
Lessertia diffusa seed germination (%) on water agarmedia was 89.3 %. The number of viable rhizobia cellsper seed after inoculation was variable depending on theinoculant strain. The highest number was recorded forstrain WSM3612 (960 cells seed−1). Most of the strains(WSM2566, 3565, 3636, 3598, 3898, 2623 and 3626)were present on the seed in numbers that ranged from270 to 490 cells seed−1. The lowest numbers wereregistered for the strains WSM3270 and 3495: 191.9and 51.1 viable rhizobia cells per seed whichcorresponded to 19.2 and 5.1 % of the potential rhizo-bial population respectively.
Plant population
The percentage of established plants in the field rangedfrom 38 to 63 %. None of the inoculant treatments had a
significant effect on the numbers of L. diffusa plants(P>0.05). There was high variation between replicates.
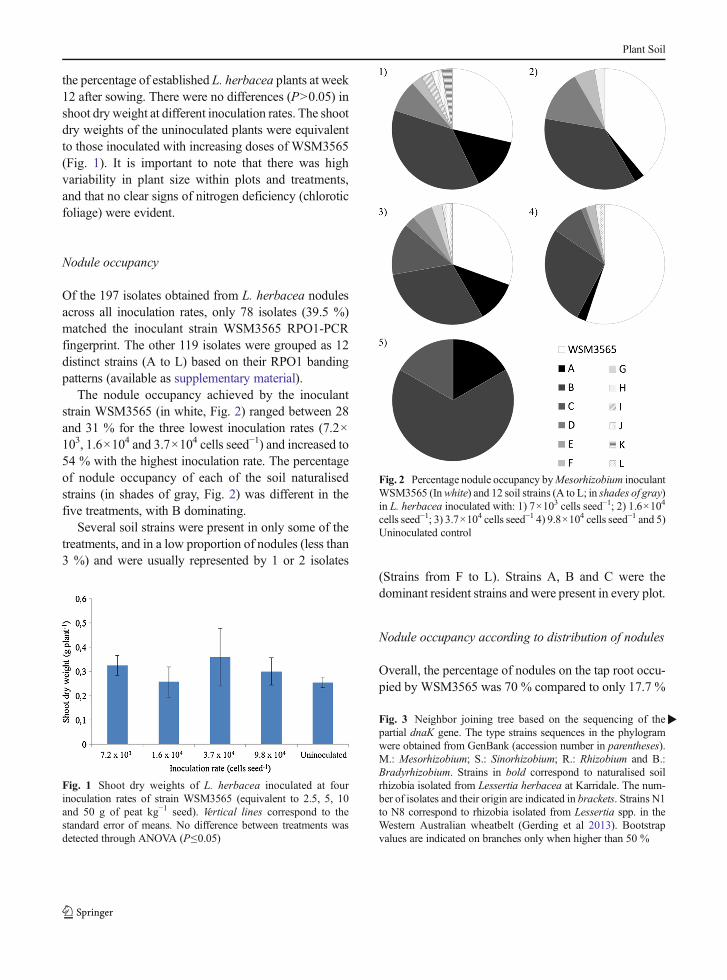
Plant biomass production and nodule occupancy
There were differences in L. diffusa dry weights for thedifferent inoculant strains (solid bars in Fig. 4). Thestrains WSM2566, 3495, 3898, 2623 and 3270 had noeffect on plant biomass in comparison to the uninocu-lated control. The highest dry weights were achievedwhen inoculating with WSM3565, 3598, 3636,3626and 3612 all of which produced higher dry weights thanthe uninoculated control (P≤0.05).
The nodule occupancy (bars with diagonal stripes inFig. 4) was variable according to the inoculant strain.The highest nodule occupancy (%) was achieved bystrains WSM3636 and WSM3598, with 81.8 and84.6 % of the nodules respectively. Three of the strainst h a t s i gn i f i c an t l y i nc r e a s ed d ry we igh t s ,WSM3565,3626 and 3612, achieved only 35, 55.5 and44.5 % of nodule occupancy respectively. Most of thestrains that did not increase dry weights (WSM2566,2623, 3898 and 3270) showed a percentage of noduleoccupancy lower than 50 % (ranging from 37.5 to47.4 %), the only exception was WSM3495 whichoccupied 52 % of the nodules.
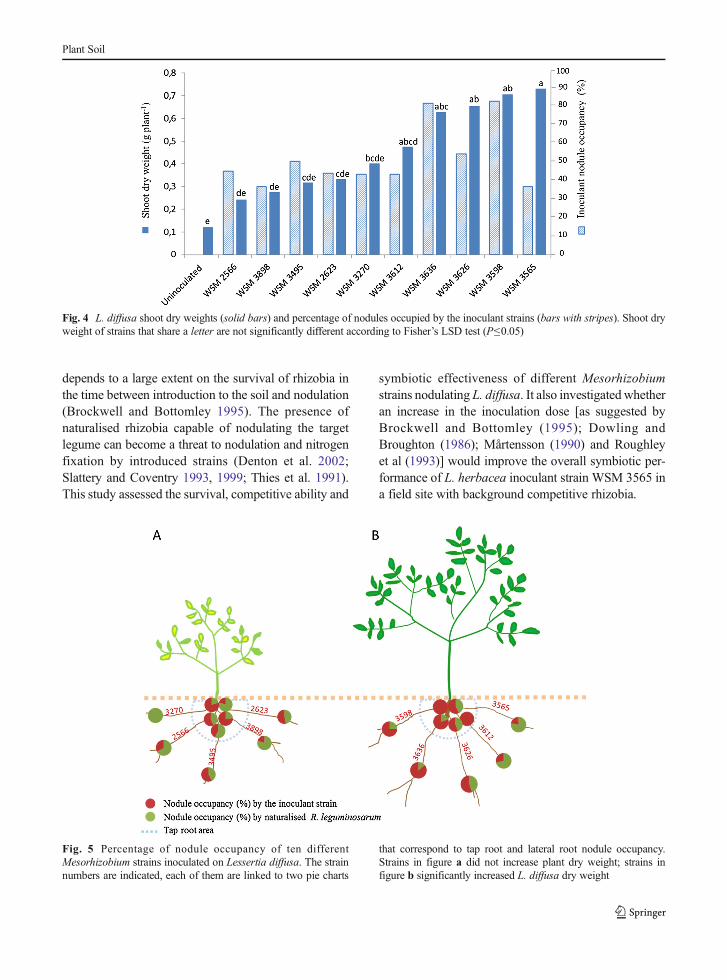
Nodule occupancy according to distribution of nodules
The percentage of the inoculant strains achieving nod-ule occupancy varied according to the position of thenodules. The amount of nodules occupied by the in-oculant strains tended to be higher when the nodulesassessed were located on the tap root, close to thecrown, than in lateral root nodules (Fig. 5). The strainsWSM3612, 3598, 3270, 3636 and 3898 achieved taproot nodule occupancies >75 %. However, onlyWSM3636 and 3598 maintained high nodule occu-pancy in lateral roots. Overall, strains that achievedover 50 % nodule occupancy on both tap and lateralroots (WSM3598, 3636 and 3626) induced a signifi-cant increase in plant biomass in comparison to theuninoculated control (Fig. 4).
Discussion
A primary aim of legume inoculation is to maximize thepotential yield from biological nitrogen fixation and this
Plant Soil
depends to a large extent on the survival of rhizobia inthe time between introduction to the soil and nodulation(Brockwell and Bottomley 1995). The presence ofnaturalised rhizobia capable of nodulating the targetlegume can become a threat to nodulation and nitrogenfixation by introduced strains (Denton et al. 2002;Slattery and Coventry 1993, 1999; Thies et al. 1991).This study assessed the survival, competitive ability and
symbiotic effectiveness of different Mesorhizobiumstrains nodulating L. diffusa. It also investigatedwhetheran increase in the inoculation dose [as suggested byBrockwell and Bottomley (1995); Dowling andBroughton (1986); Mårtensson (1990) and Roughleyet al (1993)] would improve the overall symbiotic per-formance of L. herbacea inoculant strain WSM 3565 ina field site with background competitive rhizobia.
Fig. 4 L. diffusa shoot dry weights (solid bars) and percentage of nodules occupied by the inoculant strains (bars with stripes). Shoot dryweight of strains that share a letter are not significantly different according to Fisher’s LSD test (P≤0.05)
Fig. 5 Percentage of nodule occupancy of ten differentMesorhizobium strains inoculated on Lessertia diffusa. The strainnumbers are indicated, each of them are linked to two pie charts
that correspond to tap root and lateral root nodule occupancy.Strains in figure a did not increase plant dry weight; strains infigure b significantly increased L. diffusa dry weight
Plant Soil

The isolation of R. leguminosarum from Lessertianodules in field experiments in the Western AustralianWheat belt (Gerding et al. 2013) led to the study of thesymbiotic interaction of Lessertia spp. with other inocu-lants under current use in Western Australia (Table 1), towhich these legumes might be exposed once introducedinto the field, where L. diffusa, L. capitata and L. herbaceawere able to nodulate with taxonomically and symbioti-cally different rhizobia not known for their broad hostrange (Gibson et al. 2008; Hirsch et al. 2001; López-Lara et al. 1996; Slattery et al. 2004). Nodulation withthe Burkholderia sp. strain WSM3937 was expected; asone of the L. herbacea strains (WSM3602) nodA hadmatched with the nodA corresponding to that strain in aprevious study (Gerding et al. 2012) and nodA genes areknown to correlate with host range (Laguerre et al. 2001;Perret et al. 2000; Roche et al. 1996). This might beexplained by the fact that nodA is one of the commongenes that code for the Nod factor core, which can bemodified by various chemical groups which also conferhost specificity (López-Lara et al. 1996; Perret et al. 2000).
There are various factors that contribute to host spec-ificity (Perret et al. 2000), with the amount and structureof the Nod factor molecule being attributed with thegreatest effect (Gibson et al. 2008; Hirsch et al. 2001).The rhizobia species that were able to interact withL. capitata, L. diffusa and L. herbacea produce differentNod factor structures (Oldroyd and Downie 2008; Perretet al. 2000). Although some studies report that there isno strict correlation between Nod factor and the plantsthey nodulate (Perret et al. 2000), the length and degreeof saturation of the fatty acyl group of the Nod factor canhave a function in defining specificity of interactionbetween host and bacteria and regulate nodule initiationand morphogenesis (Gibson et al. 2008; Oldroyd andDownie 2008; Sessitsch et al. 2002).
It is important to consider that a suboptimal Nodfactor can still induce nodule morphogenesis if theirconcentrations are high enough (Oldroyd and Downie2008; Walker and Downie 2000). Martínez-Romero(2003) showed that the promiscuous legumePhaseolus vulgaris is able to nodulate with many morerhizobium species under controlled axenic conditions,than when grown in the field. This could explain thevariety of strains able to nodulate L. capitata, L. diffusaand L. herbacea under glasshouse conditions, where theplants were inoculated with a high number of bacteria,and therefore not representative of the ability ofL. herbacea to nodulate with all these strains in field.
Despite the interventions of increased inoculationdoses and the improved soil and environmental condi-tions (higher average rainfall and a soil pH of 6.3),L. diffusa and L. herbacea showed overall low numbersof established plants and did not improve in comparisonto previous field trials in the Western Australian wheatbelt (Gerding et al. 2013). The seed germination for bothspecies was previously assessed in the laboratory andwas satisfactory, but in the field seeds can show areduced vigour as well as reduced viability due to det-rimental environmental factors (Bewley and Black1985; Powell 2006). Consequently, reasons for poorestablishment that could include climatic and edaphicfactors besides rhizobia need to be assessed in future.
The naturalised strains competing for nodulation inL. herbacea were identified as R. leguminosarum. Thiswas no surprise, since it had been the dominant com-peting species in previous field experiments withLessertia spp. (Gerding et al. 2013) and it is present inpopulations as high as 106 rhizobia g−1 soil in differentenvironments of Australia (Ballard et al. 2002;Brockwell et al. 2008; Denton et al. 2002; Howiesonand Ballard 2004).
When some of these competitive soil rhizobia areeffective or partially effective with the target legume,an advisable strategy is to maximise nitrogen fixation bymanipulating the rhizobial population (Aguilar et al.2001; Brockwell and Bottomley 1995; Brockwell et al.1995; Denton et al. 2003; Hungria et al. 2003;Mpepereki et al. 2000; Thies et al. 1991). This recom-mendation cannot apply for L. herbacea, as it formsineffective nodules with R. leguminosarum strains(Gerding et al. 2013). In cases like this, displacing theineffective population by the effective inoculant is achallenge, and the most viable choice seems to be toincrease inoculant dose and to select for competitivestrains to outcompete the resident soil rhizobia(Amarger 1981b; Brockwell et al. 1995; Herridge2008; Meade et al. 1985) or in the worst case scenario,to change the host (Sessitsch et al. 2002). While theapproach of increasing inoculant dose with L. herbaceadid not render the expected results, the outcomesachieved by L. diffusa inoculated with differentmicrosymbionts were more promising.
L. diffusa had been the Lessertia spp. that performedbest in former studies in the Western Australian wheatbelt (Gerding et al. 2013). In this work, although it wasaffected by ineffective nodulation with naturalisedrhizobia, the strains WSM3636, 3598, 3626, 3612 and
Plant Soil

3565 were able to increase L. diffusa dry weight, con-sistent with data obtained under controlled conditions(Gerding et al. 2012). Thies et al. (1992) suggested thathighly competitive inoculant strains are able to performwell across a range of environments. This seems to betrue for WSM3636 which was superior to other strainsin the field trial but had also been the only strain recov-ered from plants in situ and from soil (1 year afterinoculation), in experiments in the wheat belt (Gerdinget al. 2013). These results also emphasize the impor-tance of the rhizosphere colonisation by the inoculant,and lateral root infection, in the L. diffusa response toinoculation. The strains that achieved highest plant dryweight were those that were present in a high proportionin lateral root nodules that were formed later during thegrowing season. Those strains that were present mainlyin the tap root nodules but in a small proportion in newlyformed lateral nodules (WSM3898 and 3495), did nothave a significant effect on plant biomass in spite oftheir nitrogen fixing effectiveness in glasshouse experi-ments (Gerding et al. 2012).
In general, nodules present on the tap root wereoccupied in a higher proportion by the inoculant strains(Fig. 5), which is likely due to the strategic placement ofthe inoculant on the seed (Brockwell et al. 1995; Dentonet al. 2003; Hardarson et al. 1989; McDermott andGraham 1989). This is an indication that inoculationwas successful in ensuring that the inoculant strainformed the first nodules, which are the ones that startcontributing nitrogen in the earlier stages of develop-ment of the legume (Brockwell et al. 1982; Pijnenborgand Lie 1990; Spriggs and Dakora 2007). Yet, noduleformation on lateral roots is a strongly indicative of theability of the strains to move and to colonise the rhizo-sphere (McDermott and Graham 1989; Pijnenborg et al.1991; Spriggs and Dakora 2007), and become increas-ingly important at later stages in plant development(Dowling and Broughton 1986; Hardarson et al. 1989;Sessitsch et al. 2002). Moreover, lateral root nodulesmay have played an important role in these experimentsdue to the low rainfall conditions that caused the top soilto dry and therefore the earlier senescence of a greatproportion of the crown nodules. Thus, plants mighthave had to rely on nitrogen fixed by nodules formedlater, i.e. those formed on secondary roots.
In L. diffusa the proportion of nodules formed by thedifferent strains in lateral roots was variable. Most of thestrains occupied less than 55 % of lateral root nodules,and in L. herbacea the occupancy of WSM3565 did not
exceed 20 % with any of the inoculation doses. This isof concern considering that L. herbacea and L. diffusaare perennial species that should rely on the inoculantestablished with the seed in the first year during thefollowing seasons (Ballard et al. 2003; Brockwell et al.1982; Sessitsch et al. 2002) and that nodule occupancyis expected to decline over time (Brockwell et al. 1995;Denton et al. 2002; Svenning et al. 2001). Nevertheless,the strains WSM3636 and 3598 nodulating L. diffusaconsistently increased plant dry weight and were presentin 85.7 and 75.0 % of the lateral nodules respectivelyand appeared to have the capacity to colonise therhizosphere.
The comparison between L. diffusa and L. herbaceafield performances demonstrates the impact that legumehost can have in the symbiotic relationship withrhizobia. The inoculant strain WSM3565 had provedeffective with L. herbacea in glasshouse conditionsand was only partially effective with L. diffusa(Gerding et al 2012). However, in the field, WSM3565produced an increase in L. diffusa dry weight but not inL. herbacea, although it was inoculated at 10 times theinoculant dose. The exact mechanisms that allowed thisto happen are unknown. It could be due to an ability ofL. diffusa to better discriminate for effective rhizobia, ashas been reported for other legume species and cultivars(Ballard and Charman 2000; Drew and Ballard 2009;Trinick et al. 1991; Yates et al. 2005).
Inoculating legumes in soils with the presence of alarge population of competitive rhizobia is generallyregarded as challenging (Ballard and Charman 2000;Ballard et al. 2003; Brockwell et al. 1995; Dentonet al. 2002; Slattery and Coventry 1993; Thies et al.1991). Moreover, the use of legumes that are prone toform ineffective symbioses with soil rhizobia is discour-aged (Howieson and Ballard 2004). However, this studyshowed that L. diffusa strains like WSM3636 and 3898are able to outcompete local populations of rhizobia andto colonise lateral roots, and together with WSM3565,3612 and 3626 able to increase dry weight, whichmakesthem promising to develop as inoculants. There was alsoevidence of an overall higher performance of L. diffusain comparison to L. herbacea, as had been observed inprevious field experiments (Gerding et al. 2013) sug-gesting an effect of the host on strain performance in thefield. It should be noted though that L. diffusa establish-ment was still not sufficient (Li et al. 2008) and that agreat disparity between plants was observed withinplots, which may in part had had an impact in the poor
Plant Soil

plant establishment. Based on the variation observed, itcannot be discarded that within plant species there mightbe different ecotypes, which can show differences intolerance to environmental conditions (Joshi et al.2001; Vogel et al. 2005) and also in their ability to selectrhizobia (Ballard et al. 2002; Charman and Ballard2004; Drew and Ballard 2009). This suggests a needfor selection of L. diffusa for adaptation to these condi-tions and for symbiotic performance before embarkinginto new field inoculation trials.
References
Aguilar OM, López MV, Riccillo PM (2001) The diversity ofrhizobia nodulating beans in Northwest Argentina as a sourceof more efficient inoculant strains. J Biotechnol 91:181–188
Amarger N (1981a) Competition for nodule formation betweeneffective and ineffective strains of Rhizobium meliloti. SoilBiol Biochem 13:475–480
Amarger N (1981b) Selection of Rhizobium strains on their com-petitive ability for nodulation. Soil Biol Biochem 13:481–486
Ardley JK, Parker MA, DeMeyer SE, Trengove RD, O’Hara GW,Reeve WG, Yates RJ, Dilworth MJ, Willems A, HowiesonJG (2012)Microvirga lupini sp. nov.,Microvirga lotononidissp. nov. and Microvirga zambiensis sp. nov. arealphaproteobacterial root-nodule bacteria that specificallynodulate and fix nitrogen with geographically and taxonom-ically separate legume hosts. Int J Syst Evol Microbiol 62:2579–2588
Ballard RA, Charman N (2000) Nodulation and growth of pasturelegumes with naturalised soil rhizobia. 1. Annual Medicagospp. Aust J Exp Agric 40:939–948
Ballard RA, Craig AD, Charman N (2002) Nodulation and growthof pasture legumes with naturalised soil rhizobia. 2. Balansaclover (Trifolium michelanium Savi). Aust J Exp Agric 42:939–944
Ballard RA, Sheperd BR, Charman N (2003) Nodulation andgrowth of pasture legumes with naturalised soil rhizobia. 3.Lucerne (Medicago sativa L.). Aust J Exp Agric 43:135–140
Bewley JD, Black M (1985) Seeds: physiology of developmentand germination. Plenum Press, New York, p 367
Brockwell J (1982) Plant-infection counts of Rhizobia in soils. In:Vincent JM (ed) Nitrogen fixation in legumes. Academic,Australia, pp 41–58
Brockwell J, Bottomley PJ (1995) Recent advances in inoculanttechnology and prospects for the future. Soil Biol Biochem27:683–697
Brockwell J, Gault RR, Zorin M, Roberts MJ (1982) Effects ofenvironmental variables on the competition between inocu-lum strains and naturalized populations of Rhizobium trifoliifor nodulation of Trifolium subterraneum L. and on rhizobiapersistence in the soil. Aust J Agric Res 33:803–815
Brockwell J, Bottomley PJ, Thies JE (1995) Manipulation ofrhizobia microflora for improving legume productivity andsoil fertility: a critical assessment. Plant Soil 174:143–180
Brockwell J, Fettell NA, Bowman AM, Smith W, Sweeney G,Charman N, Ballard RA (2008) Symbiotic competence ofrose clover (Trifolium hirtum All.). Aust J Agric Res59:802–813
Charman N, Ballard RA (2004) Burr medic (Medicagopolymorpha L.) selections for improved N2 fixationwith naturalised soil rhizobia. Soil Biol Biochem 36:1331–1337
Deaker R, Roughley RJ, Kennedy IR (2004) Legume seed inoc-ulation technology - a review. Soil Biol Biochem 36:1275–1288
Demezas DH, Bottomley PJ (1986) Interstrain competition be-tween representatives of indigenous serotypes of Rhizobiumtrifolii. Appl Environ Microbiol 52:1020–1025
Denton MD, Coventry DR, Murphy PJ, Howieson JG, BellottiWD (2002) Competition between inoculant and naturalisedRhizobium leguminosarum bv. trifolii for nodulation of an-nual clovers in alkaline soils. Aust J Agric Res 53:1019–1026
Denton MD, Reeve WG, Howieson JG, Coventry DR (2003)Competitive abilities of common field isolates and a com-mercial strain of Rhizobium leguminosarum bv. trifolii forclover nodule occupancy. Soil Biol Biochem 35:1039–1048
Dowling DN, Broughton WJ (1986) Competition for nodulationof legumes. Annu Rev Microbiol 40:131–157
Drew EA, Ballard RA (2009) Improving N2 fixation from theplant down: compatibility of Trifolium subterraneum L. cul-tivars with soil rhizobia can influence symbiotic perfor-mance. Plant Soil 327:261–277
Duodu S, Bhuvaneswari TV, Gudmundsson J, Svenning MM(2005) Symbiotic and saprophytic survival of three un-marked Rhizobium leguminosarum biovar trifolii strains in-troduced into the field. Environ Microbiol 7:1049–1058
Gerding M, O’Hara GW, Bräu L, Nandasena KG, Howieson JG(2012) Diverse Mesorhizobium spp. with unique nodAnodulating the South African legume species of the genusLessertia. Plant Soil 359(1):385–401. doi:10.1007/s11104-012-1153-3
Gerding M, O’Hara GW, Real D, Howieson JG, Bräu L (2013)Establishment and survival of the South African legumeLessertia spp. and rhizobia inWestern Australian agriculturalsystems. Plant Soil 370(1):235–249. doi:10.1007/s11104-013-1632-1
Gibson KE, Kobayashi H, Walker G (2008) Molecular determi-nants of a symbiotic chronic infection. Annu Rev Genet 42:413–441
Graham PH (ed) (2008) Ecology of the root-nodule bacteria oflegumes. Springer, Dordrecht, pp 23–58
Hardarson G, Golbs M, Danso SKA (1989) Nitrogen fixation insoybeans (Glycine max L. Merrill) as affected by nodulationpatterns. Soil Biol Biochem 21:783–787
Herridge D (2008) Inoculation technology for legumes. In: DilworthMJ, James EK, Sprent JI, Newton WE (eds) Nitrogen-fixingleguminous symbioses. Springer, Dordrecht, pp 77–115
Hirsch AM, Lum MR, Downie JA (2001) What makes theRhizobia-Legume symbiosis so special? Update onrhizobia-Legume symbiosis. Plant Physiol 127:1484–1492
Howieson JG (1995) Characteristics of an ideotype acid tolerantpasture legume symbiosis in Mediterranean agriculture. PlantSoil 171:71–76
Howieson J, Ballard R (2004) Optimising the legume symbiosis instressful and competitive environments within southern
Plant Soil

Australia - some contemporay thoughts. Soil Biol Biochem36:1261–1273
Howieson JG, Ewing MA, D’Antuono MF (1988) Selection foracid tolerance in Rhizobium meliloti. Plant Soil 105:179–188
Howieson JG, Loi A, Carr SJ (1995) Biserrula pelecinus L. - alegume pasture species with potential for acid, duplex soilswhich is nodulated by unique root-nodule bacteria. Aust JAgric Res 46:997–1009
Howieson JG, Yates RJ, Foster KJ, Real D, Besier RB (2008)Prospects for the future use of legumes. In: Dilworth MJ,James EK, Sprent JI, Newton WE (eds) Nitrogen-fixingleguminous symbioses. Springer, Dordrecht, pp 363–393
Howieson JG, De Meyer SE, Vivas-Marfisi A, Ratnayake S,Ardley JK, Yates RJ (2013) Novel Burkholderia bacteriaisolated from Lebeckia ambigua - a perennial suffrutescentlegume of the fynbos. Soil Biol Biochem 60:55–64
Hungria M, Campo RJ, Mendes IC (2003) Benefits of inoculation ofthe common bean (Phaseolus vulgaris) crop with efficient andcompetitiveRhizobium tropici strains. Biol Fertil Soils 39:88–93
Joshi J, Schmid B, Caldeira MC, Dimitrakopoulos PG, Good J,Harris R, Hector A, Huss-Danell K, Jumpponen A, Minns A,Mulder CPH, Pereira JS, Prinz A, Scherer-Lorenzen M,Siamantziouras ASD, Terry AC, Troumbis AY, Lawton JH(2001) Local adaptation enhances performance of commonplant species. Ecol Lett 4:536–544
Laguerre G, Nour SM,Macheret V, Sanjuan J, Drouin P, Amarger N(2001) Classification of rhizobia based on nodC and nifH geneanalysis reveals a close phylogenetic relationship amongPhaseolus vulgaris symbionts. Microbiology 147:981–993
Li GD, Lodge GM, Moore GA, Craig AD, Dear BS, Boschma SP,Albertsen TO, Miller SM, Harden S, Hayes RC, Hughes SJ,Snowball R, Smith AB, Cullis BC (2008) Evaluation ofperennial pasture legumes and herbs to identify species withhigh herbage production and persistence in mixed farmingzones in southern Australia. Aust J Exp Agric 48:449–466
López-Lara IM, Blok-Tip L, Quinto C, Garcia M, Stacey G,Bloemberg GV, Lamers GEM, Lugtenberg BJJ, Thomas-Oates JE, SpainkHP (1996) NodZ ofBradyrhizobium extendsthe nodulation host range of Rhizobium by adding a fucosylresidue to nodulation signals. Mol Microbiol 21:397–408
Lupwayi NZ, Olsen PE, Sande ES, Keyser HH, Collins MM,Singleton PW, Rice WA (2000) Inoculant quality and itsevaluation. Field Crop Res 65:259–270
Mårtensson AM (1990) Competitiveness of inoculant strains ofRhizobium leguminosarum bv. trifolii in red clover usingrepeated inoculation and increased inoculum levels. Can JMicrobiol 36:136–139
Martínez-Romero E (2003) Diversity of Rhizobium-Phaseolusvulgaris symbiosis: overview and perspectives. Plant Soil252:11–23
McDermott TR, Graham PH (1989) Bradyrhizobium japonicuminoculant mobility, nodule occupancy, and acetylene reduc-tion in the soybean root system. Appl Environ Microbiol 55:2493–2498
Meade J, Higgins P, O’Gara F (1985) Studies on the inoculation andcompetitiveness of a Rhizobium leguminosarum strain in soilscontaining indigenous rhizobia. Appl Environ Microbiol 49:899–903
Mpepereki S, Javaheri F, Davis P, Giller KE (2000) Soyabeans andsustainable agriculture: promiscuous soyabeans in southernAfrica. Field Crop Res 65:137–149
Oldroyd GED, Downie JA (2008) Coordinating nodule morpho-genesis with rhizobial infection in legumes. Annu Rev PlantBiol 59:519–546
Perret X, Staehelin C, Broughton WJ (2000) Molecular basis ofsymbiotic promiscuity. Microbiol Mol Biol Rev 64:180–201
Pijnenborg JWM, Lie TA (1990) Effect of lime-pelleting on thenodulation of lucerne (Medicago sativa L.) in an acid soil: acomparative study carried out in the field, in pots and inrhizotrons. Plant Soil 121:225–234
Pijnenborg JWM, Lie TA, Zehnder AJB (1991) Nodulation oflucerne (Medicago sativa L.) in an acid soil: effects of inoc-ulum size and lime-pelleting. Plant Soil 131:1–10
Powell AA (2006) Seed vigor and its assessment. In: Basra AS(ed) Handbook of seed science and technology. HaworthPress, New York, pp 603–648
Richardson AE, Viccars LA, Watson JM, Gibson AH (1995)Differentiation of Rhizobium strains using the polymerasechain reaction with random and directed primers. Soil BiolBiochem 27:515–524
Roche P, Maillet F, Plazanet C, Debelle F, Ferro M, Truchet G,Prome J-C, Denarie J (1996) The common nodABC genes ofRhizobium meliloti are host-range determinants. Proc NatlAcad Sci U S A 93:15305–15310
Roughley RJ, Gemell LG, Thompson JA, Brockwell J (1993) Thenumber of Bradyrhizobium sp. (lupinus) applied to seed andits effect on rhizosphere colonisation, nodulation and yield oflupin. Soil Biol Biochem 25:1453–1458
Sessitsch A, Howieson JG, Perret X, Antoun H,Martínez-RomeroE (2002) Advances in Rhizobium research. Crit Rev Plant Sci21:323–378
Singleton P (1992) Introduction to bacteria for students of biology,biotechnology and medicine. Wiley, Chichester, p 221
Slattery JF, Coventry DR (1993) Variation of soil populations ofRhizobium leguminosarum bv. trifolii and the occurrence ofinoculant rhizobia in nodules of subterranean clover afterpasture renovation in North-Eastern Victoria. Soil BiolBiochem 25:1725–1730
Slattery JF, Coventry DR (1999) Persistence of introduced strainsof Rhizobium leguminosarum bv trifolii in acidic soils ofnorth-eastern Victoria. Aust J Exp Agric 39:829–837
Slattery JF, Pearce DJ, Slattery WJ (2004) Effects of residentrhizobial communities and soil type on the effective nodula-tion of pulse legumes. Soil Biol Biochem 36:1339–1346
Spriggs AC, Dakora FD (2007) Competitive ability of selectedCyclopia Vent. rhizobia under glasshouse and field condi-tions. Soil Biol Biochem 39:58–67
Stępkowski T, CzaplińskaM,MiedzinskaK,Moulin L (2003) Thevariable part of the dnaK gene as an alternative marker forphylogenetic studies of rhizobia and related alphaproteobacteria. Syst Appl Microbiol 26:483–494
Svenning MM, Gudmundsson J, Fagerli IL, Leinonen P (2001)Competition for nodule occupancy between introducedstrains of Rhizobium leguminosarum Biovar trifolii and itsinfluence on plant production. Ann Bot 88:781–787
Tamura K, Dudley J, Nei M, Kumar S (2007) MolecularEvolutionary Genetics Analysis (MEGA) software version4.0. Mol Biol Evol 24:1596–1599
Thies JE, Singleton PW, Bohlool BB (1991) Influence of the sizeof indigenous rhizobial populations on establishment andsymbiotic performance of introduced rhizobia on field-grown legumes. Appl Environ Microbiol 57:19–28
Plant Soil

Thies JE, Bohlool BB, Singleton PW (1992) Environmental ef-fects on competition for nodule occupancy between intro-duced and indigenous rhizobia and among introduced strains.Can J Microbiol 38:493–500
TrinickMJ,Miller M, Hadobas PA (1991) Formation and structureof root nodules induced on Macroptilium atropurpureuminoculated with various species of Rhizobium. Can J Bot69:1520–1532
Vekemans X (2002) AFLP-SURV version 1.0. Distributed by theauthor. Laboratoire de Genétique et Ecologie Vegétale,Université Libre de Bruxelles, Belgium
Vogel KP, Schmer MR, Mitchell RB (2005) Plant adaptationregions: ecological and climatic classification of plant mate-rials. Rangel Ecol Manag 58:315–319
Walker SA, Downie JA (2000) Entry of Rhizobium leguminosarumbv. viciae into root hairs requires minimal nod factor specific-ity, but subsequent infection thread growth requires nodO ornodE. Mol Plant-Microbe Interact 13:754–762
Yates RJ, Howieson JG, Real D, Reeve WG, Vivas-MarfisiA, O’Hara GW (2005) Evidence of selection for effec-tive nodulation in the Trifolium spp. symbiosis withRhizobium leguminosarum biovar trifolii. Aust J ExpAgric 45:189–198
Yates RJ, Howieson JG, Reeve WG, Bräu L, Speijers J,Nandasena KG, Real D, Sezmis E, O’Hara GW (2008)Host-strain mediated selection for an effective nitrogen-fixing symbiosis between Trifolium spp. and Rhizobiumleguminosarum biovar trifolii. Soil Biol Biochem 40:822–833
Yates RJ, Howieson JG, Reeve WG, O’Hara GW (2011) A re-appraisal of the biology and terminology describing rhizobialstrain success in nodule occupancy of legumes in agriculture.Plant Soil 348:255–267
Zahran HH (1999) Rhizobium-legume symbiosis and nitrogenfixation under severe conditions and in an Arid Climate.Microbiol Mol Biol Rev 63:968–989
Plant Soil