oxidative stress and its association with male infertility 6
TRANSCRIPT

57© Springer Nature Switzerland AG 2020S. J. Parekattil et al. (eds.), Male Infertility, https://doi.org/10.1007/978-3-030-32300-4_6
Oxidative Stress and Its Association with Male Infertility
Ashok Agarwal and Pallav Sengupta
6.1 Introduction
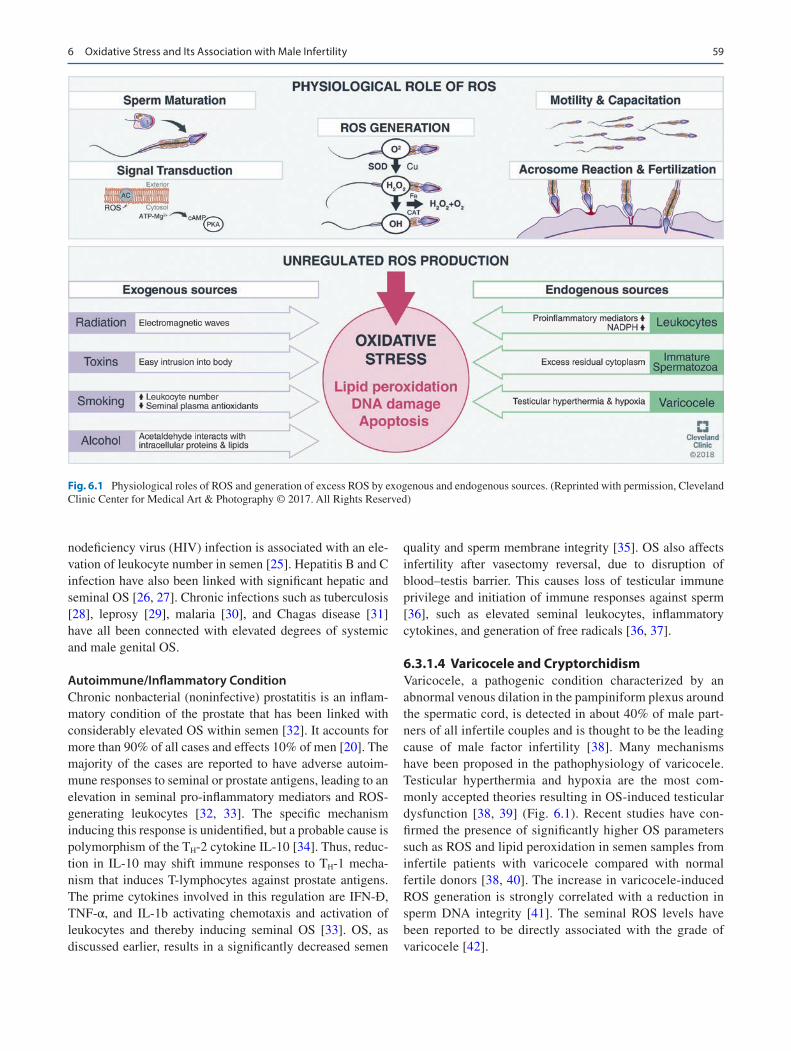
Infertility is a major global medical and social concern, and male etiology contributes to half of the overall infertility [1]. The mechanisms, by which multivariate causative factors influence male fecundity, are not yet fully understood. The advances in research in the arena of male reproductive sci-ence could potentially recognize some of the most deleteri-ous factors attributing to male factor infertility. These include physiological, environmental, as well as genetic and epigen-etic factors. At the molecular level, most of these factors interfere with normal male reproductive functioning via the imbalance in reactive oxygen species (ROS) generation and antioxidant capacity, a condition of oxidative stress (OS) [2].
ROS are the oxygen metabolism derivatives. They present as a set of toxic metabolites, while possessing some vital physiological role in normal male reproductive functions [3]. At normal physiological levels, ROS participate in essential intracellular signaling cascades to ensure proper reproduc-tive functions such as capacitation and hyperactivation of sperms, and acrosome reaction (AR) [3, 4]. On the contrary, the imbalance between ROS and antioxidant levels causes a disparity in the redox balance, which has deleterious effects over the cellular micro- and macromolecules including the carbohydrate components, lipids, proteins, and nucleic acids [5]. Spermatozoa are very vulnerable to OS as they lack proper cell repair machineries and have inadequate antioxi-dant capacity [6]. These deficiencies in spermatozoa are due to their structural peculiarity possessing diminutive cytoplas-mic content. Spermatozoa are highly susceptible to OS-induced lipid peroxidation (LPO) due to high abundance of PUFA in their cell membrane. As a result of LPO, sperma-tozoa lose their membrane permeability causing efflux of the much-needed ATPs, thereby impairing their own motility with disrupted flagellar movements [7, 8]. OS impairs the structural and functional aspects of sperm including its pro-duction, viability, and fertilizing potential [9].
This chapter provides a concise overview by discussing (a) the generation and sources of ROS in the male reproduc-tive tract, (b) physiological role of ROS in male reproductive functions, (c) pathological role of ROS in causing male infertility, and (d) assessment as well as management of OS-induced male infertility. This chapter aims to aid in bet-ter understanding of association of ROS with male infertility from physiological and clinical perspectives.
6.2 Biochemistry of Oxidative Stress
Reactive oxygen species (ROS) are generated as a result of physiological cellular metabolism. Biological energy pro-duction mostly involves oxidative phosphorylation by the enzymatically regulated reaction of oxygen with other bio-
6
A. Agarwal (*) American Center for Reproductive Medicine, Cleveland Clinic, Cleveland, OH, USAe-mail: [email protected]
P. Sengupta Department of Physiology, MAHSA University, Jenjarom, Selangor, Malaysia
Key Points• ROS mediate some essential intracellular signaling
cascades needed for normal male reproductive functions.
• Excess ROS production leads to redox imbalance and OS that cause oxidative damage to the sperm.
• ROS can affect male reproductive functions through lipid peroxidation, sperm DNA fragmentation, apoptosis of germ cells, and adversely affecting ART media.
• Laboratory assessment of ROS in male infertility includes both indirect methods via lipid peroxida-tion markers, TAC and seminal ORP, and direct method through chemiluminescence assay.

58
molecules within the mitochondria [10]. In the course of these enzymatic reductions of molecular oxygen to produce energy, free radicals are formed [11]. A free radical refers to an oxygen molecule that has a single or several unpaired electron(s) in the molecular orbitals. Superoxide anion (O2
•−) radical, the primary ROS molecule, is produced by incorpo-ration of an electron to the molecular O2. This primary prod-uct, either by direct or indirect conversions, is then transformed to secondary ROS products, namely, peroxyl radical (ROO•−), hydroxyl radical (OH•−), or hydrogen per-oxide (H2O2) [8]. Among these, H2O2 cannot be categorized as a free radical as it is devoid of any unpaired electron. Reactive species derived from nitrogen (reactive nitrogen species or RNS) also have vital physiological and pathological importance. RNS include nitrous oxide (N2O), peroxynitrite (ONOO−), peroxynitrous acid (HOONO), nitrogen dioxide (•NO2), and dinitrogen trioxides (N2O3) etc. [10, 12].
Spermatozoa generates ROS via two mechanisms: (a) in sperm plasma membrane, ROS is be produced by the nico-tinamide adenine dinucleotide phosphate (NADP) oxidase system and/or (b) in sperm mitochondria, ROS can be gener-ated via the NAD-dependent redox reaction. Spermatozoa are mitochondria-rich cells owing to their constant require-ment of energy for their motility [13]. Increased number of dysfunctional spermatozoa triggers higher ROS production in semen which in turn affects sperm mitochondrial function and motility. As discussed earlier, the leading ROS in human spermatozoa is O2
•− that reacts with itself through dismuta-tion reactions to yield H2O2. In presence of transition metals (Fe2+ and Cu2+), H2O2 and O2
•− can generate the most reactive OH•− via the Haber–Weiss reaction which can initiate a LPO cascade disrupting membrane fluidity and impairing sperm functions [14, 15].
6.3 Sources of ROS in Male Reproductive Tract
6.3.1 Endogenous Sources
6.3.1.1 LeukocytesThe significant percentage of semen specimens contain peroxidase- positive leukocytes (polymorphonuclear leuko-cytes, predominantly neutrophils, 50∼60%) and macro-phages (20∼30%) that originate from prostate gland and seminal vesicles. Leukocytospermia is also a sperm- disrupting disorder characterized by the presence of >1 × 106 WBC/mL of semen [16]. Any infection or inflammatory response triggers these cells to produce 100 times more ROS than normal production rate as a defense mechanism and also promote NADPH production through the hexose mono-phosphate (HMP) shunt [5]. ROS generation is one of the prime mechanisms of leukocytes-induced immune defense
with leukocytes being quite potent OS-inducing cells [15]. In addition, the elevation of pro-inflammatory mediators (pre-dominantly IL-6, IL-8, and TNFα) and reduction of antioxi-dants in inflammatory reactions can induce a respiratory burst resulting in OS (Fig. 6.1) [17].
6.3.1.2 Immature SpermatozoaIn normal conditions, the cytoplasm is extruded from the maturing spermatozoa to prepare itself for fertilization. Any disruption in spermiogenesis may result in retention of excess residual cytoplasm (ERC) around the mid-piece of the spermatozoon, as in immature teratozoospermic sperms (Fig. 6.1). These ERCs have considerable content of enzymes to regulate glucose metabolism, namely, the glucose-6- phosphate dehydrogenase (G6PD). They also mediate intracellular β-nicotinamide adenine dinucleotide phosphate (NADPH) production through the HMP shunt that is the major fuel for ROS production via NADPH oxi-dase located within the sperm membrane [18]. Thus, terato-zoospermic sperms produce more ROS compared with morphologically normal sperm. The calcium-dependent NADPH oxidase (NOX 5) present in the sperm differs from those present in the leukocytes, as the sperm-specific NOX5 activity is not dependent upon protein kinase-C [19]. However, the correlation of overexpression of NOX5 in spermatozoa in OS-induced infertility remains under debate.
6.3.1.3 Infections, Autoimmune/Inflammatory Conditions
Genitourinary Tract InfectionProstatitis may affect up to 50% of men worldwide, and 10% of them suffer from chronic prostatitis [20]. Bacteria that cause prostate infection are of urinary tract origin and/or transmitted sexually [21, 22]. Some important nonsexu-ally transmitted bacteria are gram-negative bacteria (E. coli, Proteus mirabilis), Streptococci (S. viridans and S. pyogenes), atypical mycoplasma strains (Ureaplasma urea-lyticum), and Mycoplasma hominis coagulase-negative Staphylococci (S. epidermidis, S. haemolyticus). These agents commonly produce acute inflammatory responses by promoting leukocyte influx into the male reproductive tract and, ultimately, triggering ROS production [13]. Viral infections by cytomegalovirus, herpes simplex virus (HSV), Epstein–Barr virus, and others can also initiate oxidative damage to spermatozoa. Herpes simplex DNA has been found in semen of 4–50% of infertile men [23], with a ten-fold increase IgM antibodies toward HSV [24].
Systemic InfectionSeveral chronic systemic infections have been correlated with increased OS in the male genital system. Human immu-
A. Agarwal and P. Sengupta
59
nodeficiency virus (HIV) infection is associated with an ele-vation of leukocyte number in semen [25]. Hepatitis B and C infection have also been linked with significant hepatic and seminal OS [26, 27]. Chronic infections such as tuberculosis [28], leprosy [29], malaria [30], and Chagas disease [31] have all been connected with elevated degrees of systemic and male genital OS.
Autoimmune/Inflammatory ConditionChronic nonbacterial (noninfective) prostatitis is an inflam-matory condition of the prostate that has been linked with considerably elevated OS within semen [32]. It accounts for more than 90% of all cases and effects 10% of men [20]. The majority of the cases are reported to have adverse autoim-mune responses to seminal or prostate antigens, leading to an elevation in seminal pro-inflammatory mediators and ROS- generating leukocytes [32, 33]. The specific mechanism inducing this response is unidentified, but a probable cause is polymorphism of the TH-2 cytokine IL-10 [34]. Thus, reduc-tion in IL-10 may shift immune responses to TH-1 mecha-nism that induces T-lymphocytes against prostate antigens. The prime cytokines involved in this regulation are IFN-Ɣ, TNF-α, and IL-1b activating chemotaxis and activation of leukocytes and thereby inducing seminal OS [33]. OS, as discussed earlier, results in a significantly decreased semen
quality and sperm membrane integrity [35]. OS also affects infertility after vasectomy reversal, due to disruption of blood–testis barrier. This causes loss of testicular immune privilege and initiation of immune responses against sperm [36], such as elevated seminal leukocytes, inflammatory cytokines, and generation of free radicals [36, 37].
6.3.1.4 Varicocele and CryptorchidismVaricocele, a pathogenic condition characterized by an abnormal venous dilation in the pampiniform plexus around the spermatic cord, is detected in about 40% of male part-ners of all infertile couples and is thought to be the leading cause of male factor infertility [38]. Many mechanisms have been proposed in the pathophysiology of varicocele. Testicular hyperthermia and hypoxia are the most com-monly accepted theories resulting in OS-induced testicular dysfunction [38, 39] (Fig. 6.1). Recent studies have con-firmed the presence of significantly higher OS parameters such as ROS and lipid peroxidation in semen samples from infertile patients with varicocele compared with normal fertile donors [38, 40]. The increase in varicocele-induced ROS generation is strongly correlated with a reduction in sperm DNA integrity [41]. The seminal ROS levels have been reported to be directly associated with the grade of varicocele [42].
Fig. 6.1 Physiological roles of ROS and generation of excess ROS by exogenous and endogenous sources. (Reprinted with permission, Cleveland Clinic Center for Medical Art & Photography © 2017. All Rights Reserved)
6 Oxidative Stress and Its Association with Male Infertility

60
Cryptorchidism is yet another cause of male factor infer-tility. It involves hypo-spermatogenesis with inability of gonocytes to mature to type-A spermatogonia [43]. Cryptorchidism-affected men may be surgically treated with orchidopexy in early stage of life still, but they will continue to possess an increased sperm ROS generation and DNA fragmentation [40].
Spermatic cord torsions are also associated with male infertility. It is suggested that OS-induced ischemia reper-fusion injury results in damage to contralateral testis and with torsions. A prolonged ischemic phase follows either surgical or spontaneous restoration of testicular blood flow. This results in sudden influx of activated leukocytes into the testis [1, 40] that accelerates the generation of free radi-cals [6].
6.3.1.5 Other Chronic DiseasesDiabetes, one of the most prevalent chronic diseases, impairs male reproductive functions influencing both spermatogen-esis and erectile processes. Diabetic men have shown higher sperm DNA fragmentation induced by OS compared with normal men [43]. Chronic inflammation and OS are also found in men with chronic kidney diseases [44]. Men with hemoglobinopathies, such as β-thalassemia, possess sys-temic OS [45] that can cause oxidative damage to sperm owing to iron overload by multiple blood transfusions [46]. Homocysteines can also lead to reproductive dysfunctions and OS by accumulated toxins [46]. Hyperhomocysteinemia generally occurs via suboptimal homocysteine re- methylation to methionine by methyl tetrahydrofolate reductase (MTHFR). This is caused by dietary folate deficiency or a single nucleotide polymorphism (SNP) in the MTHFR gene [47]. SNPs in the MTHFR gene are more commonly reported in the infertile men [48–50], rendering these men at elevated risk for homocysteine-induced OS.
6.3.2 Exogenous Sources
6.3.2.1 RadiationMobile phone radiation magnifies ROS generation in semi-nal plasma and causes deleterious effects on semen quality. It induces sperm DNA damage affecting sperm motility, count, and vitality [6, 51]. The radiofrequency electromag-netic waves interfere with the intracellular electron transfer along the membranes owing to cytosolically charged mole-cules, and thus interfering with the normal germ cell func-tions [52–54]. Radiofrequency waves affect male fertility through both thermal and nonthermal mechanisms (Fig. 6.1). Since testis depends mostly upon surface conduction for its temperature regulation, it is susceptible to damage by elec-tromagnetic energy than any other organs who mainly depend on blood flow for temperature control. Radiation
may elevate scrotal temperature, and even if it gets increased by 1 °C, normal spermatogenesis gets affected. The non-thermal effects of radiation include induction of oxidative stress or cell membrane potential alterations which disrupt germ cell proliferation and trigger apoptosis. Radiation may also cause sperm DNA fragmentation and epigenetic modi-fications. Chronic exposure to radiation severely disrupts steroidogenesis and brings about degeneration of Leydig cells [52–54].
6.3.2.2 Lifestyle FactorsSmoking disrupts the balance of ROS production with ade-quate antioxidant defense. Smoking has been reported to elevate concentrations of seminal leukocytes by 48% that accounts for the increase in seminal ROS by 107%. This leads to reduction in seminal antioxidants capacity and sub-sequent rise in 8-OHdG concentrations (a potent oxidative damage biomarker) [52]. Furthermore, smoking elevates cadmium and lead concentrations in blood and semen. This in turn increases ROS generation and damages sperm motil-ity [52]. Germ cell apoptosis and DNA damage are gener-ally identified in smokers, thus compromising male fecundity [55].
Alcohol is another inducing factor leading to uncontrolled seminal ROS generation and deterioration of antioxidant capacity. Acetaldehyde, produced as a by-product through ethanol metabolism, yields ROS via reaction with sperm cel-lular components. This significantly reduces the percentage of functional spermatozoa [56].
6.3.2.3 ToxinsIndustrialization and domestic sophistications have led to the generation of a huge mass of environmental toxins and endo-crine disruptors in an individual’s immediate environment. These intrude into the body and may potentially induce excessive testicular ROS generation (Fig. 6.1). ROS thereby impairs morphology and functions of sperms. Exposure to environmental toxins leads to ROS-induced germ cell apop-tosis [55]. Increased use of plastics have led to rise in expo-sure to phthalates which in addition to toxic metals such as lead, manganese, cadmium, chromium, and mercury afflict sperm quality, count, and spermatogenesis [52, 57].
6.4 Physiological Role of ROS
ROS, at physiological limits, is crucial for the quintessential phases of sperm development such as their maturation, capacitation, hyperactivation, and finally acrosome reaction (Fig. 6.1). The efficiency of ROS as signaling molecule attri-butes to its compatible small size, abundant generation in cells, and short half-life [58]. Redox modulation of cysteine residues is the key regulatory step by which ROS influence
A. Agarwal and P. Sengupta

61
the most of the signaling cascades. It is to be mentioned that activity of any specific enzyme is determined by the redox status of its thiol groups. ROS operates through activation of adenyl cyclase (AC), thus increasing intracellular cyclic AMP (cAMP) production. This turns on protein kinase A (PKA) that activates the downstream messengers specific to the spermatozoa maturational state [59].
6.4.1 Capacitation
Capacitation is the process of ultimate functional maturation of spermatozoa, which is essential to fertilize an ovum. Maturation of spermatozoa takes place mostly in the epidid-ymis. It involves cell membrane restructuring, surface pro-tein redistribution, and alterations in nuclear components and enzymes [3, 59]. ROS mediates the spermatozoa maturation by regulating the vital cell signaling cascades [59, 60]. Spermatozoon in human has compactly packed chromosomal DNA with smaller proteins than histone pro-teins. Chromatin stability is achieved through ROS-aided disulfide bonds between the cysteine residues of protamines. Thus, ROS helps in conferring chromatin stability and in pre-venting DNA damage. ROS also protects mitochondria from proteolytic dilapidation by participating in the development of “mitochondrial capsule” formed by disulfide bonded pro-teins [3, 61]. ROS facilitates spermatozoa capacitation by signaling cascades increasing intracellular cAMP levels. This activates protein kinase A followed by subsequent phos-phorylation of MEK (extracellular signal regulated kinase)-like proteins, sheath proteins, and threonine-glutamate-tyrosine [18, 62]. This cellular pathway ultimately leads to spermato-zoa capacitation, which is a prerequisite for acrosome reac-tions [52, 63].
6.4.2 Hyperactivation
Hyperactivation refers to a state of increased sperm motility with enhanced amplitude and asymmetric sperm flagellar movements. It confers a nonlinear movement of sperm with an increased sideward displacement of the sperm head [64]. This process just follows capacitation or can be considered as a later part of capacitation. It is required to determine proper sperm penetration into the zona pellucida for success-ful fertilization. The initiation of the process is mediated by calcium ion influx, which together with ROS activates ade-nylyl cyclase (AC), inducing cAMP production and activa-tion of PKA. This triggers NADPH oxidase to further induce ROS generation [60]. PKA triggers protein tyrosine kinases (PTK) and phosphorylates the tyrosine residues present within the axonemal fibrous sheath and flagellar cytoskele-ton. ROS induces the phosphorylation of tyrosine by influ-
encing the activation of PTK and deactivation of phosphotyrosine phosphatase (PTPase) [3, 52].
6.4.3 Acrosome Reaction
The hyperactivated spermatozoon crosses the cumulus ooph-orous, to get attached to the zona pellucida of the oocyte. This is followed by a pore formation in the extracellular matrix of zona pellucida by spermatozoa exocytotic release of proteolytic enzymes [61]. These reactions enable sperm attachment and entry into ova and together is termed as acro-some reaction. These reactions are initiated by phosphoryla-tion of tyrosine residues of membrane proteins that is aided by ROS-regulated cell signaling pathway. The primary path-way as discussed in the earlier sections involves Ca2+ influx, rise in intracellular cAMP, and PKA activation [3, 59].
6.4.4 Fertilization
ROS also assists in the process of fertilization by enhancing the membrane fluidity for sperm–oocyte fusion to occur [52]. ROS inhibits protein tyrosine phosphatase activity and thus prevents phospholipase A2 (PLA2) from getting deacti-vated. Hence, the activated PLA2 cleaves the secondary fatty acid present in the membrane to effectively enhance mem-brane fluidity [65].
6.5 Mechanisms of ROS-Mediated Male Infertility
6.5.1 Lipid Peroxidation
Sperm has substantially high lipid content in its plasma membrane. They are mostly present as PUFAs with unconju-gated double bonds in methylene groups. This weakens the methyl bond between carbon and hydrogen. Thus, hydrogen is rendered vulnerable to oxidative attack. The uncontrolled rise in intracellular ROS initiates a progressive reaction cas-cade and triggers LPO [39, 66, 67]. LPO is a self- propagating autocatalytic reaction that destroys almost 60% of the mem-brane fatty acids affecting its fluidity. It also imparts nonspe-cific permeability of the membrane and disrupts membrane receptors and enzymes that severely impairs fertilization [52, 66, 68].
Oxidative damage once initiated follows steady propaga-tion until it is terminated. “Initiation” is marked by isolation of hydrogen atoms from the carbon–carbon bonds, followed by generation of free radicals and lipid radicals. This reacts with oxygen to produce peroxyl radicals [66]. These peroxyl radicals further abstract hydrogen atom from the lipids
6 Oxidative Stress and Its Association with Male Infertility

62
(action enhanced by the presence of metals such as copper and iron). This triggers the chain of autocatalytic reaction. The “propagation” of oxidative damage refers to continua-tion of the damaging reaction with the existing radicals reacting with succeeding lipids, giving rise to toxic alde-hydes by hydroperoxide degradation (Fig. 6.2) [15, 18, 66]. Cytotoxic peroxyl and alkyl radicals are generated in a cycli-cal manner until a stable end product, malondialdehyde (MDA), is produced to terminate the reaction [66]. MDA thus bears importance as a marker to predict the level of per-oxidative damage caused to spermatozoa. Another deleteri-ous product of LPO is the hydrophilic 4-hydroxynonenal that severely impairs spermatozoa function affecting its pro-teomics and genomic constitution [52].
6.5.2 Sperm DNA Fragmentation
ROS adversely affects sperm nuclear DNA through sperm DNA fragmentation (SDF), inducing chromatin cross- linking, randomly modifying the base-pairing and chromosomal microdeletions (Fig. 6.2) [52, 66, 69]. ROS also aids adenine and pyridine nucleotides removal by initiating LPO [52, 69]. Numerous intrinsic and extrinsic factors are suggested in the etiopathogenesis of male SDF, including infection, varico-cele, advanced male age, lifestyle factors, heat stress, envi-ronmental toxins, radiations, defective protamination, and
sometimes even remain idiopathic [43, 70]. Several of the mentioned etiologies are mediated by uncontrolled ROS gen-eration leading to increased SDF [71]. Testicular SDF has been characterized by abortive germ cell apoptosis [72] and defective sperm maturation [73]. As a consequence of imbal-ance between ROS production (via testicular hypoxia, reflux of metabolites, endocrine disruption, and scrotal hyperther-mia) and protective antioxidant capacity, a state of OS is established. This induces elevated lipid peroxidation products in infertile men with pathological conditions that serve as eti-ology for SDF [74]. Moreover, treatment of such pathological state with antioxidants seems to be effective in concurrent decrease in both ROS [75] and SDF [76].
6.5.3 Apoptosis of Spermatozoa
ROS is capable of disrupting the inner and outer mitochon-drial membranes releasing cytochrome C. This cytochrome C in turn activates the apoptotic caspases [52, 62]. ROS induces apoptosis of spermatozoa by elevating cytochrome C level, which have been reported by seminal plasma analy-sis in infertile men (Fig. 6.2). The increased cytochrome C level in seminal plasma is an indicator of severe sperm mito-chondrial damage [6, 52]. Besides the defects in sperm chro-matin compaction, numerous intrinsic and extrinsic factors have been reported in the etiopathogenesis of SDF, including
Fig. 6.2 Mechanisms of ROS-induced impairments in sperm functions. (Reprinted with permission, Cleveland Clinic Center for Medical Art & Photography © 2017. All Rights Reserved)
A. Agarwal and P. Sengupta

63
varicocele, infection, advanced male age, heat stress, life-style factors, environmental toxins, ionizing and nonionizing radiations, defective protamination, and, at times, idiopathic [43, 70]. Many of the above etiologies, but not all, are medi-ated by ROS leading to high SDF [71]. Abortive apoptosis [72] and defective maturation [73] theories have been pro-posed to correlate the role of intrinsic factors in testicular SDF. Moreover, evidences showed more DNA fragmentation in epididymal and ejaculated sperm than the testicular sperm, signifying the impact of extrinsic factors in most patients [77]. The presence of a large amount of polyunsaturated fatty acids (PUFA) in the plasma membrane makes sperm suscep-tible to ROS-induced damage [78].
The close relationship between ROS and SDF is also evi-dent from the etiopathologies of all grades of clinical varico-cele. The result of imbalance of ROS (produced by testicular hypoxia, scrotal hyperthermia, reflux of metabolites, and endocrine disruption) and protective antioxidant system was demonstrated by the higher level of ROS and lipid peroxida-tion products in infertile men with varicocele than infertile men without varicocele [74]. Moreover, treatment of varico-cele is effective in decreasing both ROS [75] and SDF [76].
6.6 ROS in Assisted Reproduction
ROS has been shown to possess a significant role in fertiliza-tion. Oocyte metabolism produces ROS, but it actually lacks adequate protective antioxidant mechanisms [79], which results in OS. Moreover, preimplantation embryonic devel-opment needs a preferred modulation in cellular energetics [80]. Thus, the established OS serves as beneficial state where oxidative phosphorylation is needed as energy source for preimplantation embryo development. This oxidative phosphorylation dependency shifts to glycolysis to increase energy production [80, 81]. Oxygen may oxidatively phos-phorylate ADP to ATP, which is utilized in folliculogenesis and oocyte maturation. In case of any alterations in these reactions using oxygen, it results in excess production of ROS which adversely affects embryo development [81].
Sources of ROS differ in case of conventional IVF and for ICSI [82]. The oocytes for ICSI are devoid of cumulus cells, rendering the oocyte and the injected spermatozoa as the only potential ROS sources in the culture media environment [81, 83]. While in IVF, ROS gets generated from the several oocytes per culture dish, a large mass of cumulus cells, and also from the inseminated spermatozoa. Moreover, unlike IVF, in ICSI there is no sperm–oocyte contact, thus limiting the scope of ROS production by faulty spermatozoa [59]. This explains why lesser the concentration of spermatozoa in the in vitro culture media, higher is the possibility of proper fertilization, implantation, and pregnancy [84]. ICSI utilizes a single spermatozoon at a time and therefore a better proce-dure to minimize OS.
In either IVF or ICSI, both the culture media and exter-nal environment influence the ultimate outcomes [8]. The procedures differ mostly in the fact that for ICSI there is shorter incubation time that reduces exposure to modulat-ing external environmental factors [83, 85]. The predicted limit of the incubation time is 1–2 hours which yields bet-ter ART outcomes. In both IVF and ICSI, ROS is gener-ated from the gametes, the embryo, or may arise indirectly from external factors, such as cumulus cells, leukocytes, etc. Since fertilization followed by embryo development in vivo takes place under low oxygen tension, a state of OS in the in vitro setting definitely accounts for high failure rate in ARTs. Hence, in order to improve ART outcomes, in vivo conditions should be emulated via minimizing exposure to the factors that promote generation of excess ROS [83, 86].
6.7 Measurement of OS
ROS-induced sperm damage is a major contributing male reproductive pathology (almost 30–80% of idiopathic male infertility) [87]. Assessment of ROS levels in the infertile men thereby is very crucial for its treatment and manage-ment. However, the difficulties in ROS assessment arise from the factors such as high cost, screening inconveniences, and scarcity of accepted specific analysis method. With the advent of research and technology, more than 30 different ROS measurement assays have emerged lately that can pre-dict the level of OS in infertile men [52].
6.7.1 Assessment of Sperm OS from Routine Semen Analysis
Routine semen analysis (sperm count, morphology, and motility) provides diagnostic clues to the clinicians with asthenozoospermia being a major OS marker [88, 89]. Seminal plasma hyperviscosity is suggestive of increased MDA and reduced seminal antioxidant levels [88]. In addi-tion, Ureaplasma urealyticum infection in the semen can also increase seminal viscosity and can be associated with high seminal ROS production [88]. The presence of round sperm cells may infer leukocytospermia which is again a potent source of ROS. However, it is needed to ascertain that the round cells are actually not immature spermatozoa. For this, several other tests are required, such as peroxidase test, seminal elastase measurement, or staining of CD45 (trans-membrane glycoprotein) antibody. OS status is also pre-sumed from disruption in sperm morphology and cytoplasmic droplets featuring anomalous spermatozoa. Poor sperm membrane integrity is diagnosed by hypoos-motic swelling test (HOST), which also associates with a state of OS [52].
6 Oxidative Stress and Its Association with Male Infertility

64
6.7.2 Total Antioxidant Capacity (TAC)
Antioxidant capacity testing has been reported to potentially enhance prognostic information for infertile men. Luminol, a chemical exhibiting chemiluminescence when reacted with particular oxidizing agent, is being widely used to measure seminal total antioxidant capacity (TAC). It is quantified in contrast to a vitamin E analog “Trolox,” a water-soluble tocopherol. The results signify ROS-TAC score and aid to infer the level of antioxidants produced as a combined efforts of all seminal constituents such as vita-mins, lipids, and proteins [89].
6.7.3 Lipid Peroxidation Markers
Spermatozoa often carries accumulated lipid peroxides, pro-duced as end-products. These metabolites, hydroxynonenal, MDA, 2-propenal (acrolein), and isoprostanes, can be used as OS indicators [66]. Thiobarbituric acid (TBA) assay is used for MDA measurement. The principles include combi-nation of MDA with TBA to produce 1:2 adduct. This is a colored substance measured by fluorometry or spectropho-tometry [52, 89].
6.7.4 Seminal Oxidation-Reduction Potential (ORP)
Oxidation-reduction potential (ORP) or redox potential mea-sures the potential of electrons transfer between different chemical species [90, 91]. ORP helps to infer the association between oxidants and antioxidants and thereby suggest the level of OS. Recently, a novel method has been developed
based on galvanostatic electron measure to assess OS changes in patients in trauma [92, 93].
Seminal ORP is efficiently assessed with the MiOXSYS System (AytuBioScience, USA) which is a standardized, reproducible, and reliable method. It provides greater appli-cation for OS analysis in research and clinical settings (Fig. 6.3) [94, 95].
6.7.5 Direct Laboratory Assessments of OS
Seminal ROS can be directly measured the chemilumines-cence assays. The procedure involves a luminometer and a chemiluminescent probe such as luminal (5-amino-2,3, -dihydro-1,4-phthalazinedione). Luminol measures both extracellular and intracellular ROS. The free radicals con-tained in the semen sample produce a light signal reacting with luminal, which is converted by the luminometer to an electric signal (photon). Results are expressed as relative light units/s/106 sperm. The range of normal ROS levels in washed sperm suspensions is 0.10–1.03 units/s/106 [89].
6.8 Management of OS-Associated Male Infertility
Primary steps in OS management involve unveiling its underlying cause(s) followed by effective treatment.
6.8.1 Lifestyle Management Approach
Stress arises from both personal lifestyle adoptions and com-petitive professional load. The ill habits, such as substance
a b c
Fig. 6.3 Measurement of ORP by the MiOXSYS system. (a) MiOXSYS analyzer showing the socket and the sensor module, (b) sen-sor showing the reference cell and the sample port where the sample is
loaded, and (c) loading of semen samples on the sample port of the sensor strip. (Reprinted with permission, Cleveland Clinic Center for Medical Art & Photography © 2017. All Rights Reserved)
A. Agarwal and P. Sengupta

65
abuse, alcoholism, tobacco smoking and chewing, and an unbalanced diet are all major contributors to cause OS. Therefore, minimizing the avoidable lifestyle disorien-tations may help to prevent OS [52, 96].
Exposure to endocrine and reproductive disruptors which include pollutants, heavy metals, and other toxins greatly account for OS development. Increase in scrotal temperature via rigorous exercise, hot baths, driving for long hours, sed-entary office works, etc., induce heat stress that create OS [39, 87, 97].
6.8.2 Vitamin and Antioxidant Supplementation
Antioxidant supplementation may eradicate excess ROS or decrease its production, to halt the damaging oxidative chain reactions. On the one hand, preventive antioxidants such as the metal chelators or binding proteins, for example, trans-ferrin and lactoferrin, prevents OS by restricting ROS gen-eration [98]. On the other hand, the scavenging antioxidants (vitamins C and E) combat OS by mitigating excess ROS [99].
Antioxidants can also be classified as enzymatic and non-enzymatic. Natural antioxidants are mostly enzymatic anti-oxidants that include superoxide dismutase (SOD), catalase, and glutathione reductase (GSH). Examples of nonenzy-matic antioxidants include vitamins B, C, and E, carnitines, carotenoids, cysteines, pentoxifylline, taurine, hypotaurine, albumin, and some metals. They can also be obtained from certain foods and food supplements [52, 100, 101].
Available reports suggest antioxidant supplements are a beneficial treatment option for OS-induced male infertility that may reduce sperm DNA damage and improve semen parameters [102–107]. Specific antioxidants such as vitamin C, vitamin E, N-acetyl-cysteine, selenium, and zinc have been reported to be beneficial in treatment of OS-induced male infertility [108].
6.8.3 Surgery
Surgeries effectively correct the venous abnormality in vari-cocele patients and decrease seminal ROS, thus preventing further oxidative damage [87]. Surgical repair improves sev-eral sperm parameters and has been shown to improve preg-nancy rates as well [52, 75]. It has been reported that inflammatory and OS markers such as MDA, H2O2, and nitric oxide (NO) are reduced while antioxidant levels improved following a successful varicocelectomy [109]. This scenario suggests that varicocelectomy decreases semi-nal ROS levels and increases TAC to restore male reproduc-tive functions in varicocele patients [109, 110].
6.9 Conclusion
OS arises from disrupted balance between ROS production and their elimination. While at physiological levels, ROS contribute in maturation of sperm and normal sperm func-tions, and excessive seminal ROS induce OS that impairs sperm quality and functions. This deleterious chain of oxida-tive reactions leads to infertility. With proper assessment and management strategies, along with healthy life style prac-tices, it is possible to improve male reproductive potential in OS-induced infertile men.
6.10 Review Criteria
An extensive literature search has been performed to find the relationship between oxidative stress and male infertility using search engines such as Science Direct, OVID, Google Scholar, PubMed, and MEDLINE. The overall strategy for study identification and data extraction was based on the fol-lowing key words: “oxidative stress,” “reactive oxygen spe-cies,” “infertile men,” “infertility,” “semen parameters,” and “assisted reproduction” and the names of related oxidative- stress markers and specific ROS assessment methods. Articles published languages other than English were also considered. Data that were solely published in conference or meeting proceedings, websites, or books were not included. Websites and book-chapter citations provide conceptual con-tent only.
References
1. Agarwal A, et al. A unique view on male infertility around the globe. Reprod Biol Endocrinol. 2015;13(1):37.
2. Halliwell B, Cross CE. Oxygen-derived species: their relation to human disease and environmental stress. Environ Health Perspect. 1994;102(Suppl 10):5–12.
3. Thompson A, Agarwal A, du Plessis SS. Physiological role of reactive oxygen species in sperm function: a review. Antioxidants in male infertility: a guide for clinicians and researchers. New York, USA: Springer Science and Business Media. 2013:69–89.
4. Ochsendorf F. Infections in the male genital tract and reactive oxygen species. Hum Reprod Update. 1999;5(5):399–420.
5. Agarwal A, Saleh RA, Bedaiwy MA. Role of reactive oxygen spe-cies in the pathophysiology of human reproduction. Fertil Steril. 2003;79(4):829–43.
6. Aitken RJ, et al. Causes and consequences of oxidative stress in spermatozoa. Reprod Fertil Dev. 2016;28(1–2):1–10.
7. Alvarez JG, Storey BT. Assessment of cell damage caused by spontaneous lipid peroxidation in rabbit spermatozoa. Biol Reprod. 1984;30(2):323–31.
8. Cheeseman K, Slater T. An introduction to free radical biochemis-try. Br Med Bull. 1993;49(3):481–93.
9. Agarwal A, et al. Reactive oxygen species as an independent marker of male factor infertility. Fertil Steril. 2006;86(4):878–85.
10. Saleh RA, Agarwal A. Oxidative stress and male infertility: from research bench to clinical practice. J Androl. 2002;23(6):737–52.
6 Oxidative Stress and Its Association with Male Infertility

66
11. Valko M, et al. Free radicals and antioxidants in normal physi-ological functions and human disease. Int J Biochem Cell Biol. 2007;39(1):44–84.
12. Doshi SB, et al. Role of reactive nitrogen species in male infertil-ity. Reprod Biol Endocrinol. 2012;10:109.
13. Henkel RR. Leukocytes and oxidative stress: dilemma for sperm function and male fertility. Asian J Androl. 2011;13(1):43–52.
14. Chen SJ, et al. Influence of reactive oxygen species on human sperm functions and fertilizing capacity including therapeutical approaches. Arch Gynecol Obstet. 2013;288(1):191–9.
15. Wallach EE, Wolff H. The biologic significance of white blood cells in semen. Fertil Steril. 1995;63(6):1143–57.
16. Press W. Laboratory manual for the examination and process-ing of human semen. Geneva: World Health Organization; 2010. p. 7–113.
17. Loveland KL, et al. Cytokines in male fertility and reproductive pathologies: immunoregulation and beyond. Front Endocrinol (Lausanne). 2017;8:307.
18. Said TM, et al. Impact of sperm morphology on DNA damage caused by oxidative stress induced by β-nicotinamide adenine dinucleotide phosphate. Fertil Steril. 2005;83(1):95–103.
19. Ghanbari H, Keshtgar S, Kazeroni M. Inhibition of the CatSper channel and NOX5 enzyme activity affects the functions of the progesterone-stimulated human sperm. Iran J Med Sci. 2018;43(1):18–25.
20. Schaeffer AJ. Epidemiology and demographics of prostatitis. Andrologia. 2003;35(5):252–7.
21. Fraczek M, et al. Proinflammatory cytokines as an intermediate factor enhancing lipid sperm membrane peroxidation in in vitro conditions. J Androl. 2008;29(1):85–92.
22. Papes D, et al. Detection of sexually transmitted pathogens in patients with chronic prostatitis/chronic pelvic pain: a prospective clinical study. Int J STD AIDS. 2017;28(6):613–5.
23. Bezold G, Lange M, Peter RU. Homozygous methylenetetrahy-drofolate reductase C677T mutation and male infertility. N Engl J Med. 2001;344(15):1172–3.
24. Krause W, et al. Cellular and biochemical markers in semen indicating male accessory gland inflammation. Andrologia. 2003;35(5):279–82.
25. Chaillon A, et al. HIV trafficking between blood and semen dur-ing early untreated HIV infection. J Acquir Immune Defic Syndr. 2017;74(1):95–102.
26. Seronello S, Sheikh MY, Choi J. Redox regulation of hepati-tis C in nonalcoholic and alcoholic liver. Free Radic Biol Med. 2007;43(6):869–82.
27. Vicari E, et al. Sperm output in patients with primary infertil-ity and hepatitis B or C virus; negative influence of HBV infec-tion during concomitant varicocele. Minerva Med. 2006;97(1): 65–77.
28. Srinivasan S, et al. Oxidative stress in urogenital tuberculo-sis patients: a predisposing factor for renal stone formation—amelioration by vitamin E supplementation. Clin Chim Acta. 2004;350(1–2):57–63.
29. Vijayaraghavan R, et al. Protective role of vitamin E on the oxida-tive stress in Hansen’s disease (Leprosy) patients. Eur J Clin Nutr. 2005;59(10):1121.
30. Guha M, et al. Apoptosis in liver during malaria: role of oxida-tive stress and implication of mitochondrial pathway. FASEB J. 2006;20(8):1224–6.
31. Maçao LB, et al. Antioxidant therapy attenuates oxidative stress in chronic cardiopathy associated with Chagas’ disease. Int J Cardiol. 2007;123(1):43–9.
32. Burke AJ, et al. Inflammation and nitrosative stress effects in ovar-ian and prostate pathology and carcinogenesis. Antioxid Redox Signal. 2017;26(18):1078–90.
33. Motrich RD, et al. Reduced semen quality in chronic prostatitis patients that have cellular autoimmune response to prostate anti-gens. Hum Reprod. 2005;20(9):2567–72.
34. Shoskes DA, et al. Cytokine polymorphisms in men with chronic prostatitis/chronic pelvic pain syndrome: association with diagno-sis and treatment response. J Urol. 2002;168(1):331–5.
35. Henkel R, et al. Chronic pelvic pain syndrome/chronic prostatitis affect the acrosome reaction in human spermatozoa. World J Urol. 2006;24(1):39–44.
36. Patel AP, Smith RP. Vasectomy reversal: a clinical update. Asian J Androl. 2016;18(3):365–71.
37. Nandipati K, et al. Relationship of interleukin-6 with semen char-acteristics and oxidative stress in vasectomy reversal patients. Andrologia. 2005;37(4):131–4.
38. Agarwal A, Prabakaran S, Allamaneni SS. Relationship between oxidative stress, varicocele and infertility: a meta-analysis. Reprod Biomed Online. 2006;12(5):630–3.
39. Makker K, Agarwal A, Sharma R. Oxidative stress & male infer-tility. Indian J Med Res. 2009;129(4):357–67.
40. Cho CL, Esteves SC, Agarwal A. Novel insights into the patho-physiology of varicocele and its association with reactive oxy-gen species and sperm DNA fragmentation. Asian J Androl. 2016;18(2):186–93.
41. Majzoub A, Agarwal A, Esteves SC. Sperm DNA fragmentation testing in patients with subclinical varicocele: is there any evi-dence? Transl Androl Urol. 2017;6(Suppl 4):S459–61.
42. Will MA, et al. The great debate: varicocele treatment and impact on fertility. Fertil Steril. 2011;95(3):841–52.
43. Gunes S, Al-Sadaan M, Agarwal A. Spermatogenesis, DNA damage and DNA repair mechanisms in male infertility. Reprod Biomed Online. 2015;31(3):309–19.
44. Scholze A, et al. Oxidative stress in chronic kidney disease. Oxid Med Cell Longev. 2016;2016:8375186.
45. Hirsch RE, et al. HbE/beta-thalassemia and oxidative stress: the key to pathophysiological mechanisms and novel therapeutics. Antioxid Redox Signal. 2017;26(14):794–813.
46. Chen MJ, et al. Effect of iron overload on impaired fertility in male patients with transfusion-dependent beta-thalassemia. Pediatr Res. 2018;83(3):655–61.
47. Cornet D, et al. Association between the MTHFR-C677T iso-form and structure of sperm DNA. J Assist Reprod Genet. 2017;34(10):1283–8.
48. Bisht S, Chawla B, Dada R. Oxidative stress and polymorphism in MTHFR SNPs (677 and 1298) in paternal sperm DNA is associ-ated with an increased risk of retinoblastoma in their children: a case-control study. J Pediatr Genet. 2018;7(3):103–13.
49. Singh V, et al. SNPs in ERCC1, ERCC2, and XRCC1 genes of the DNA repair pathway and risk of male infertility in the Asian populations: association study, meta-analysis, and trial sequential analysis. J Assist Reprod Genet. 2019;36:79–90.
50. Lai H, et al. Association between genetic polymorphism in NFKB1 and NFKBIA and coronary artery disease in a Chinese Han population. Int J Clin Exp Med. 2015;8(11):21487–96.
51. Agarwal A, et al. Effect of cell phone usage on semen analysis in men attending infertility clinic: an observational study. Fertil Steril. 2008;89(1):124–8.
52. Agarwal A, et al. Effect of oxidative stress on male reproduction. World J Mens Health. 2014;32(1):1–17.
53. Houston BJ, et al. The effects of radiofrequency electromagnetic radiation on sperm function. Reproduction. 2016;152(6):R263–76.
54. Sabeti P, et al. Etiologies of sperm oxidative stress. Int J Reprod Biomed (Yazd). 2016;14(4):231–40.
55. Saleh RA, et al. Effect of cigarette smoking on levels of seminal oxidative stress in infertile men: a prospective study. Fertil Steril. 2002;78(3):491–9.
A. Agarwal and P. Sengupta

67
56. Agarwal A, Prabakaran SA. Mechanism, measurement, and prevention of oxidative stress in male reproductive physiology. Indian J Exp Biol. 2005;43(11):963–74.
57. Sengupta P, Dutta S, Krajewska-Kulak E. The disappearing sperms: analysis of reports published between 1980 and 2015. Am J Mens Health. 2017;11(4):1279–304.
58. Mittler R, et al. ROS signaling: the new wave? Trends Plant Sci. 2011;16(6):300–9.
59. Du Plessis SS, et al. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J Assist Reprod Genet. 2015;32(4):509–20.
60. Griveau JF, Le Lannou D. Reactive oxygen species and human spermatozoa: physiology and pathology. Int J Androl. 1997;20(2):61–9.
61. Fujii J, Tsunoda S. Redox regulation of fertilisation and the sper-matogenic process. Asian J Androl. 2011;13(3):420–3.
62. Fraczek M, Kurpisz M. Inflammatory mediators exert toxic effects of oxidative stress on human spermatozoa. J Androl. 2007;28(2):325–33.
63. Aitken RJ. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol Reprod Dev. 2017;84(10):1039–52.
64. Suarez SS. Control of hyperactivation in sperm. Hum Reprod Update. 2008;14(6):647–57.
65. Khosrowbeygi A, Zarghami N. Fatty acid composition of human spermatozoa and seminal plasma levels of oxidative stress bio-markers in subfertile males. Prostaglandins Leukot Essent Fatty Acids. 2007;77(2):117–21.
66. Martínez P, Proverbio F, Camejo MI. Sperm lipid peroxidation and pro-inflammatory cytokines. Asian J Androl. 2007;9(1):102–7.
67. Armstrong JS, et al. A comparison of the NADPH oxidase in human sperm and white blood cells. Int J Androl. 2002;25(4):223–9.
68. Opuwari CS, Henkel RR. An update on oxidative damage to sper-matozoa and oocytes. Biomed Res Int. 2016;2016:9540142.
69. Shen H, Ong C. Detection of oxidative DNA damage in human sperm and its association with sperm function and male infertility. Free Radic Biol Med. 2000;28(4):529–36.
70. Ahmad G, Agarwal A. Ionizing radiation and male fertility. In: Male infertility: New York, USA: Springer; 2017. p. 185–96.
71. Henkel R, et al. Effect of reactive oxygen species produced by spermatozoa and leukocytes on sperm functions in non- leukocytospermic patients. Fertil Steril. 2005;83(3):635–42.
72. Sakkas D, et al. Origin of DNA damage in ejaculated human sper-matozoa. Rev Reprod. 1999;4(1):31–7.
73. Sakkas D, et al. Relationship between the presence of endogenous nicks and sperm chromatin packaging in maturing and fertilizing mouse spermatozoa. Biol Reprod. 1995;52(5):1149–55.
74. Sakamoto Y, et al. The assessment of oxidative stress in infertile patients with varicocele. BJU Int. 2008;101(12):1547–52.
75. Hamada A, Esteves SC, Agarwal A. Insight into oxidative stress in varicocele-associated male infertility: part 2. Nat Rev Urol. 2013;10(1):26–37.
76. Wang Y-J, et al. Relationship between varicocele and sperm DNA damage and the effect of varicocele repair: a meta-analysis. Reprod Biomed Online. 2012;25(3):307–14.
77. Esteves SC, et al. Comparison of reproductive outcome in oligo-zoospermic men with high sperm DNA fragmentation undergoing intracytoplasmic sperm injection with ejaculated and testicular sperm. Fertil Steril. 2015;104(6):1398–405.
78. John Aitken R, Clarkson JS, Fishel S. Generation of reactive oxy-gen species, lipid peroxidation, and human sperm function. Biol Reprod. 1989;41(1):183–97.
79. Khazaei M, Aghaz F. Reactive oxygen species generation and use of antioxidants during in vitro maturation of oocytes. Int J Fertil Steril. 2017;11(2):63–70.
80. Ali I, et al. Reactive oxygen species-mediated unfolded pro-tein response pathways in preimplantation embryos. J Vet Sci. 2017;18(1):1–9.
81. Scott R 3rd, Zhang M, Seli E. Metabolism of the oocyte and the preimplantation embryo: implications for assisted reproduction. Curr Opin Obstet Gynecol. 2018;30(3):163–70.
82. Agarwal A, Ahmad G, Sharma R. Reference values of reactive oxygen species in seminal ejaculates using chemiluminescence assay. J Assist Reprod Genet. 2015;32(12):1721–9.
83. Agarwal A, et al. Oxidative stress in an assisted reproductive tech-niques setting. Fertil Steril. 2006;86(3):503–12.
84. Gianaroli L, et al. Prolonged sperm-oocyte exposure and high sperm concentration affect human embryo viability and pregnancy rate. Hum Reprod. 1996;11(11):2507–11.
85. Bedaiwy M, et al. Role of total antioxidant capacity in the dif-ferential growth of human embryos in vitro. Fertil Steril. 2006;86(2):304–9.
86. Sikka SC. Role of oxidative stress and antioxidants in androl-ogy and assisted reproductive technology. J Androl. 2004;25(1): 5–18.
87. Tremellen K. Oxidative stress and male infertility--a clinical per-spective. Hum Reprod Update. 2008;14(3):243–58.
88. Aydemir B, et al. The influence of oxidative damage on viscosity of seminal fluid in infertile men. J Androl. 2008;29(1):41–6.
89. Agarwal A, Majzoub A. Laboratory tests for oxidative stress. Indian J Urol: IJU. 2017;33(3):199–206.
90. McCord JM. The evolution of free radicals and oxidative stress. Am J Med. 2000;108(8):652–9.
91. Agarwal A, et al. Oxidation-reduction potential of semen: what is its role in the treatment of male infertility? Ther Adv Urol. 2016;8(5):302–18.
92. Rael LT, et al. Oxidation-reduction potential and paraoxonase- arylesterase activity in trauma patients. Biochem Biophys Res Commun. 2007;361(2):561–5.
93. Rael LT, et al. Plasma oxidation-reduction potential and pro-tein oxidation in traumatic brain injury. J Neurotrauma. 2009;26(8):1203–11.
94. Agarwal A, et al. Diagnostic application of oxidation-reduction potential assay for measurement of oxidative stress: clinical util-ity in male factor infertility. Reprod Biomed Online. 2017;34: 48–57.
95. Agarwal A, et al. MiOXSYS: a novel method of measuring oxida-tion reduction potential in semen and seminal plasma. Fertil Steril. 2016;106(3):566–73.
96. Sengupta P. Recent trends in male reproductive health problems. Asian J Pharm Clin Res. 2014;7(2):1–5.
97. Sengupta P, Dutta S. Metals. In: Skinner MK, editor. Reference module in biomedical sciences: encyclopedia of reproduction. San Diego: Elsevier; 2018.
98. Hammadeh ME, Hamad MF. Reactive oxygen species and anti-oxidant in seminal plasma and their impact on male fertility. Int J Fertil Steril. 2009;3(3):87–110.
99. McDowell LR, et al. Vitamins and minerals functioning as antiox-idants with supplementation considerations. In: Florida ruminant nutrition symposium. Gainesville: Best Western Gateway Grand; 2007.
100. Sengupta P, et al. Role of Withania somnifera (Ashwagandha) in the management of male infertility. Reprod Biomed Online. 2018;36(3):311–26.
101. Zini A, Al-Hathal N. Antioxidant therapy in male infertility: fact or fiction? Asian J Androl. 2011;13(3):374–81.
102. Agarwal A, Majzoub A. Role of antioxidants in male infertility. BJUI Knowledge. 2016;e-learning module:1–9.
103. Agarwal A, Majzoub A. Role of antioxidants in assisted reproduc-tive techniques. World J Mens Health. 2017;35(2):1–17.
6 Oxidative Stress and Its Association with Male Infertility

68
104. Magdi Y, et al. Effect of modifiable lifestyle factors and antioxi-dant treatment on semen parameters of men with severe oligoas-thenoteratozoospermia. Andrologia. 2017;49(7):e12694.
105. Majzoub A, Agarwal A. Antioxidant therapy in idiopathic oligoas-thenoteratozoospermia. Indian J Urol. 2017;33(3):207–14.
106. Majzoub A, Agarwal A, Esteves SC. Antioxidants for elevated sperm DNA fragmentation: a mini review. Transl Androl Urol. 2017;6(Suppl 4):S649–53.
107. Gharagozloo P, Aitken RJ. The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Hum Reprod. 2011;26(7):1628–40.
108. Majzoub A, Agarwal A. Systematic review of antioxidant types and doses in male infertility: benefits on semen parameters, advanced sperm function, assisted reproduction and live-birth rate. Arab J Urol. 2018;16(1):113–24.
109. Mostafa T, et al. Varicocelectomy reduces reactive oxygen species levels and increases antioxidant activity of semi-nal plasma from infertile men with varicocele. Int J Androl. 2001;24(5):261–5.
110. Chen SS, et al. Attenuation of oxidative stress after vari-cocelectomy in subfertile patients with varicocele. J Urol. 2008;179(2):639–42.
A. Agarwal and P. Sengupta