p68 rna helicase unwinds the human let-7 microrna precursor duplex and is required for...
DESCRIPTION
TRANSCRIPT
Shubert-Coleman and Henry FurneauxDavid W. Salzman, Jonathan Gene ExpressionRequired for let-7-directed Silencing oflet-7 MicroRNA Precursor Duplex and Is P68 RNA Helicase Unwinds the HumanRNAs:RNA-Mediated Regulation and Noncoding
doi: 10.1074/jbc.M705054200 originally published online August 27, 20072007, 282:32773-32779.J. Biol. Chem.
10.1074/jbc.M705054200Access the most updated version of this article at doi:
.JBC Affinity SitesFind articles, minireviews, Reflections and Classics on similar topics on the
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/282/45/32773.full.html#ref-list-1
This article cites 48 references, 20 of which can be accessed free at
at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from

P68 RNA Helicase Unwinds the Human let-7 MicroRNAPrecursor Duplex and Is Required for let-7-directedSilencing of Gene Expression*
Received for publication, June 20, 2007, and in revised form, July 31, 2007 Published, JBC Papers in Press, August 27, 2007, DOI 10.1074/jbc.M705054200
David W. Salzman, Jonathan Shubert-Coleman, and Henry Furneaux1
From the Department of Molecular, Microbial, and Structural Biology, University of Connecticut Health Center,Farmington, Connecticut 06030
MicroRNAs are short, single-stranded RNAs that arise from atransient precursor duplex. We have identified a novel activityin HeLa cell extracts that can unwind the let-7 microRNAduplex. Using partially purified material, we have shown thatmicroRNA helicase activity requires ATP and has a nativemolecularmass of�68 kDa.Affinity purification of the unwind-ing activity revealed co-purification of P68 RNA helicase.Importantly, recombinant P68 RNA helicase was sufficient tounwind the let-7 duplex.Moreover, like its native homolog, P68RNA helicase did not unwind an analogous small interferingRNA duplex. We further showed that knockdown of P68 inhib-ited let-7 microRNA function. From our data, we conclude thatP68 RNA helicase is an essential component of the let-7microRNA pathway, and in conjunction with other factors, itmay play a role in the loading of let-7microRNA into the silenc-ing complex.
It is now recognized thatmicroRNAsplay a critical role in theregulation of gene expression and that their deregulation mayunderlie many human diseases (1–5). Thus, it is important tounderstand how they are incorporated into the protein cofactorcomplexes that are essential for their suppressive activity.MicroRNAs are initially transcribed as precursor RNAs, whichfold into hairpin structures (6). The acknowledged first step ofmicroRNAmaturation is the specific endonucleolytic cleavageof the pri-microRNA near the base of the stem loop, by Drosha(7). The Drosha cleavage product is typically a 70-nucleotidestem loop RNA, containing a two-nucleotide overhang at the 3�end and a recessed 5�-phosphate (8–10). This product is a sub-strate for a second enzyme called Dicer (11). Dicer cleaves bothstrands of the hairpin precursor �21 nucleotides from the endof the stem loop creating a 19-nucleotide paired duplex withtwo-nucleotide overhangs at the 3� ends: the transientmicroRNA duplex (12–14).It is now clear that the silencing activities of microRNAs are
effected by a novel class of RNA-binding proteins called theArgonaute family (15–18). However, the affinity of these pro-
teins for duplex RNA ismuch less than that observed for single-stranded RNA (15, 19). Moreover, analysis of Argonaute/RNAcomplexes in the cell typically reveals the association of onlyone strand (the guide strand) of this transient duplex (20). Thus,there are likely to be additional factors, which unwind the tran-sient duplex and confer specificity in uptake into Argonaute.Studies on structurally different, but conceptually analogous,
siRNA2 duplexes have revealed that this specificity in uptakemay be conferred by a heterodimer of Dicer and R2D2 in whichthe stable end of the duplex is bound by R2D2 and Dicer facil-itates the loading of the loosely paired 5� end of the guide strandinto Argonaute (21–24). Although the unwinding is likely facil-itated by the cleavage of the passenger strand byArgonaute (25,26), RNA helicase A has also recently been implicated in load-ing of the guide strand of siRNA (27).Much less is known aboutthe factors that are likely required to load the guide strand of theanalogous microRNA duplex into Argonaute complexes. Incontrast to siRNA duplexes, microRNA duplexes typically con-tain unpaired bulges, which likely preclude the cleavage of thepassenger strand by Argonaute (26).An affinity-purified complex that contains Argonaute2 and
Dicer has been shown to direct the cleavage of a target mRNAdirected by a let-7 hairpin precursor (19, 28). Although thisargues that Dicer might be necessary for the unwinding of themicroRNA transient duplex, microRNA duplexes can stillsilence expression in cells that lack Dicer (29). In any event,neither the siRNA-directed nor themicroRNAduplex-directedcleavage of a mRNA has been successfully reconstituted withrecombinant proteins, and thus, it is likely that additional fac-tors are required for this step.In the present study, we have identified an activity from
human cells that promotes the ATP-dependent unwinding ofthe human let-7 microRNA precursor duplex. Further charac-terization suggested that this activity corresponded to the pre-viously described P68 RNA helicase (30–32). Indeed, we foundthat recombinant P68 RNA helicase was sufficient to unwindthe let-7 microRNA precursor duplex. Importantly, a transientknockdown of P68 abrogated let-7-directed suppression ofgene expression and indicates that P68 RNA helicase is indeedrequired to facilitate the uptake of duplex let-7 microRNA intothe silencing complex.
* This work was supported by National Institutes of Health Grants R03DA022226 and P01 HL70694. The costs of publication of this article weredefrayed in part by the payment of page charges. This article must there-fore be hereby marked “advertisement” in accordance with 18 U.S.C. Sec-tion 1734 solely to indicate this fact.
1 To whom correspondence should be addressed: University of ConnecticutHealth Center, 236 Farmington Ave., Farmington, CT 06030. Tel.: 860-679-2374; Fax: 860-679-1862; E-mail: [email protected].
2 The abbreviations used are: siRNA, small interfering RNA; GAPD, glyceralde-hyde-3-phosphate; GAPDH, glyceraldehyde-3-phosphate dehydrogen-ase; AMP-PNP, adenosine 5�-(�,�-imino)triphosphate.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 45, pp. 32773–32779, November 9, 2007© 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
NOVEMBER 9, 2007 • VOLUME 282 • NUMBER 45 JOURNAL OF BIOLOGICAL CHEMISTRY 32773 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from
EXPERIMENTAL PROCEDURES
HeLa S3 cells were obtained from the National Cell CultureCenter (Minneapolis, MN). Synthetic RNAs and siRNAs wereobtained from Dharmacon Research Inc. (Lafayette, CO).Luciferase reporter plasmids were provided by the David Bartellaboratory. The His-P68 plasmid was provided by the Zhi-RenLiu laboratory. Anti-P68 monoclonal antibody (PAB204) wasobtained fromUpstate Biochemicals, andmonoclonal antibod-ies against GAPD and Vimentin were obtained from Abcam.Preparation of HeLa Cell Extract—HeLa S3 cells (National
Cell Culture Center) were resuspended in hypotonic buffer (50mM Tris, pH 7.5, 10 mM KCl, 5 mM dithiothreitol). The swollencells were homogenized, and KCl, MgCl2, and glycerol wereadded to final concentrations of 100 mM, 2 mM, and 10%,respectively. The homogenate was centrifuged at 500� g for 10
min. The supernatant was removed,and the pellet was resuspended inbuffer A (50 mM Tris, pH 7.5, 2 mMMgCl2, 5 mM dithiothreitol, 10%glycerol). KCl was added dropwiseto a final concentration of 400 mM.The homogenate was centrifuged at10,000� g for 10min. The resultantnuclear extractwas stored at�80 °Cin aliquots.RNA Affinity Chromatography—
0.5 ml of avidin A beads (VectorLaboratories) were incubated with36 �mol of biotinylated let-7 pre-cursor hairpin RNA in 0.6 ml vol-ume with buffer A (50 mM Tris, pH7.5, 0.01% Nonidet P-40, 10% glyc-erol) at 4 °C for 8 h. Beads werewashedwith bufferA containing 1MNaCl and then equilibrated withbuffer A containing 50 mM NaCl.Nuclear extract was applied to thecolumn and washed with buffer Acontaining 50 mM NaCl. The col-umn was then eluted with buffer Acontaining a 50 mM stepwise 0.05M-0.8 M NaCl gradient.Preparation of Recombinant His-
P68 RNA Helicase—RecombinantHis-P68 was prepared as describedpreviously (31). In short, His-P68was induced in BL21 (DE3) cellswith 0.1 mM isopropyl-1-thio-�-D-galactopyranoside at 37 °C for 6 h.Bacterial pellets were resuspendedin lysis buffer (50 mM Tris, pH 8.0,0.1 M NaCl, 0.5 mM EDTA, pH 8.0)and were lysed by the addition oflysozyme (0.2 mg/ml) and TritonX-100 (1%). The resulting superna-tant was applied to a nickel-nitrilo-triacetic acid agarose column. Thecolumn was first washed with lysis
buffer containing 20mM imidazole followed by lysis buffer con-taining 20mM imidazole and 0.15 mMNaCl. Recombinant His-P68 was then eluted with buffer containing 250 mM NaCl and250 mM imidazole.Preparation of Labeled MicroRNA Precursor Duplex—The
let-7 guide strand was labeled using T4 polynucleotide kinaseand [�-32P]ATP (Amersham Biosciences). After phenol-chlo-roform extraction, it was annealed (65 °C for 5 min and then37 °C for 25 min) to a 5-fold excess of let-7 passenger strand.The duplex was then gel-purified and stored in 50 mM Tris, pH7.5, and 0.2 M potassium acetate.Precursor Duplex-unwinding Assay—Reaction mixtures
(0.02 ml) contained labeled let-7 precursor duplex (1 nM), 50mM Tris, pH 7.5, 50 mM NaCl, 2 mM MgCl2, 5 mM ATP, andnuclear extract or recombinant P68 RNA helicase as indicated.
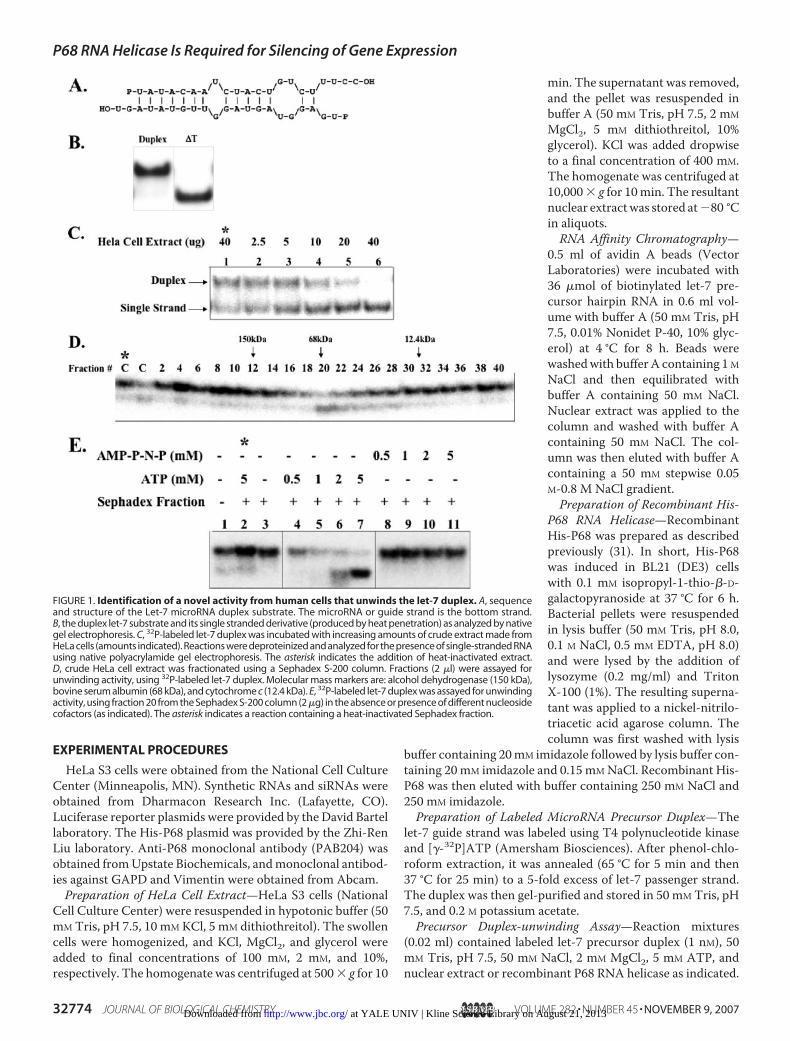
FIGURE 1. Identification of a novel activity from human cells that unwinds the let-7 duplex. A, sequenceand structure of the Let-7 microRNA duplex substrate. The microRNA or guide strand is the bottom strand.B, the duplex let-7 substrate and its single stranded derivative (produced by heat penetration) as analyzed by nativegel electrophoresis. C, 32P-labeled let-7 duplex was incubated with increasing amounts of crude extract made fromHeLa cells (amounts indicated). Reactions were deproteinized and analyzed for the presence of single-stranded RNAusing native polyacrylamide gel electrophoresis. The asterisk indicates the addition of heat-inactivated extract.D, crude HeLa cell extract was fractionated using a Sephadex S-200 column. Fractions (2 �l) were assayed forunwinding activity, using 32P-labeled let-7 duplex. Molecular mass markers are: alcohol dehydrogenase (150 kDa),bovine serum albumin (68 kDa), and cytochrome c (12.4 kDa). E, 32P-labeled let-7 duplex was assayed for unwindingactivity, using fraction 20 from the Sephadex S-200 column (2 �g) in the absence or presence of different nucleosidecofactors (as indicated). The asterisk indicates a reaction containing a heat-inactivated Sephadex fraction.
P68 RNA Helicase Is Required for Silencing of Gene Expression
32774 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 45 • NOVEMBER 9, 2007 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from

After 30 min at 37 °C, the reaction was terminated by the addi-tion of SDS-EDTAbuffer (50% glycerol, 0.1 MTris, pH 8.0, 0.5%SDS, 20 mM EDTA, 0.1% Nonidet P-40, 0.1% each bromphenolblue, and xylene cyanol) and analyzed by 15% native polyacryl-amide gel electrophoresis.Luciferase Assay of MicroRNA Function—HeLa cells were
transfected using Lipofectamine 2000 in a 12-well plate usingthe indicated amounts of firefly luciferase reporter plasmid andeither �-galactosidase or Renilla luciferase plasmid. Firefly andeither �-galactosidase or Renilla luciferase activities wereassayed 36–48 h after transfection using the Dual Luciferaseassay (Promega). Firefly activity was normalized by cotransfec-tion of either �-galactosidase or Renilla plasmids to control fortransfection efficiency.
RESULTS
The Identification of an ATP-dependent Activity That CanUnwind the let-7 MicroRNA Duplex—We elected to use thehuman let-7 duplex (Fig. 1A) as amodel since let-7 is absolutelyconserved between worms and humans and has been biochem-ically studied in many systems (12, 33–39). This choice there-fore permits the ready comparison of any identified humanpro-tein co-factors with those identified in other organisms.However, one disadvantage of the human let-7 duplex is that adiscernible level of single strand is producedmerely on incuba-tion at 37 °C. Thus, in all experiments, we included a negativecomparison control (a heat-inactivated corresponding cellular
fraction) so that we could clearlydistinguish any cellular helicaseactivity from the enzyme-indepen-dent background. Fig. 1B shows thatthe let-7 duplex was unwound onincubation with nuclear extractmade fromHeLa cells. The extent ofunwinding titrated with the amountof extract added to the reaction. Toextend this observation and toestablish the native molecularweight of the activity, we fraction-ated the extract using a SephadexS-200 column. Fractions were col-lected and assayed for helicaseactivity. There was a minor highmolecular weight species; however,themajority of the unwinding activ-ity eluted in an inclusion volumeconsistent with a native molecularmass of 68 kDa (Fig. 1C).Next, using this partially purified
material, we investigated whetherthe unwinding activity requiredATP. Little unwinding was seen inthe absence of ATP, whereas theaddition of ATP markedly stimu-lated the reaction and appeared tosaturate at 5 mM. In addition, nounwinding activity was seen in reac-tion mixtures containing AMP-
PNP, a non-hydrolyzable ATP analog (Fig. 1D). From theseobservations,weconcludedthatwehave identifiedanovelATP-dependent activity in HeLa cell extract that is capable ofunwinding the let-7microRNAduplex and that this activity hasa native molecular mass of �68 kDa.P68 RNA Helicase Co-purifies with the MicroRNA Duplex-
unwinding Activity—Our initial observations suggested thatthe activity that unwinds the let-7 duplex precursor mightcorrespond to P68 RNA helicase. P68 RNA helicase is anATP-dependent RNA-unwinding enzyme of 68 kDa that hasbeen found to be a subunit of the Drosha-processing com-plex (8, 30–32, 40). Accordingly, we elected to affinity-purifyour unwinding activity and investigate whether it co-puri-fied with P68 RNA helicase. We prepared an affinity columnby immobilizing a biotinylated hairpin sequence containingthe let-7 passenger and guide sequences (Fig. 2A) to avidinbeads.HeLa cell extract was applied, the column was washed with
low salt, and bound protein was eluted using a step gradientfrom 0.05 to 0.8 M NaCl (Fig. 2B). A significant portion of themicroRNAhelicase activitywas retained and eluted at 200–250mM NaCl (Fig. 2C). Next, we assayed for the presence of P68RNA helicase by Western blot and found that it was preciselycoincident with the unwinding activity (Fig. 2D). Thus, we con-cluded that P68 was indeed a strong candidate for themicroRNA-unwinding activity.
FIGURE 2. Purification of the let-7 microRNA helicase activity yields P68 RNA helicase as a likely candidate.A, the sequence and likely structure of the let-7 hairpin RNA used to purify the activity. Bi indicates the Biotin moiety.B, HeLa cell extract was applied to a let-7 hairpin affinity column and eluted with a salt gradient. C, fractions (2 �l)were assayed for unwinding activity, using 32P-labeled let-7 duplex. D, fractions from the let-7 hairpin affinity column(2 �l) were analyzed via Western blot for P68 RNA helicase using PAb204. Molecular size markers areindicated in kDa.
P68 RNA Helicase Is Required for Silencing of Gene Expression
NOVEMBER 9, 2007 • VOLUME 282 • NUMBER 45 JOURNAL OF BIOLOGICAL CHEMISTRY 32775 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from

Recombinant P68 RNA Helicase Is Sufficient to Unwind thelet-7 MicroRNA Duplex—Next, we investigated whetherrecombinant P68 was sufficient to unwind the let-7microRNA duplex, and if so, whether its properties resem-bled those displayed by the native activity. First, we investi-gated the structural features of the microRNA duplex thatare necessary for unwinding by the native activity. We syn-thesized two mutant derivatives of the let-7 duplex. In thefirst mutant (mutant 1), nucleotide substitutions were madeso that every nucleotide of the microRNA guide strand wasannealed to the passenger. This mutant is a “bad” siRNA inwhich the 5� end of the guide strand is annealed to the pas-
senger strand. Previous studieshave shown that the guide strandof such an siRNA will not bereadily incorporated into a silenc-ing complex (21, 22, 26, 41). Thismutant was a very poor substrate;virtually no unwinding activitywas noted even on incubation withsaturating amounts of affinity-pu-rified unwinding activity (Fig. 3, Aand B).Next, we synthesized a mutant
that lacked the internal bulges ofthe microRNA duplex yet retainsthe unpaired structure of the 5�end of the guide strand. Thismutant (mutant 2) is analogous toa “good” siRNA in which the guidestrand is readily incorporated intothe silencing complex (41). Impor-tantly, this mutant was also a verypoor substrate and indicates thatthe unwinding activity recognizedthe internal bulges and thereforecan distinguish between an siRNAduplex and a microRNA duplex.Indeed, the importance of theinternal bulges was illustrated bythe observation that a thirdmutant, in which the bulges wereretained and the 5� end of theguide was annealed to the passen-ger strand, exhibited significantunwinding Fig. 3, A and B).
Importantly, recombinant P68RNA helicase also displayed amarked preference for a microRNAduplex, and the critical role of theinternal bulges was similarly evi-dent. Thus, we concluded that P68RNAhelicase is sufficient to unwindthe let-7 duplex, and like the affini-ty-purified activity, it prefers amicroRNA duplex to an siRNAduplex (Fig. 3, A and C).P68 RNA Helicase Is Required
for let-7 MicroRNA Function in HeLa Cells—If P68 RNAhelicase was required for the unwinding of the let-7microRNA duplex, one would predict that its down-regula-tion would prevent loading of let-7 microRNA into thesilencing complex and that significant inhibition ofmicroRNA activity would result. To test this hypothesis, weutilized a reporter plasmid (pIS-Lin41(s) (42)) that containsa previously characterized let-7-response element found inlin-41 mRNA. Indeed, transfection of this plasmid into HeLacells resulted in a 22-fold repression when compared withnormalized luciferase activity in comparison with thatexpressed by the parental plasmid that lacks the response
FIGURE 3. Purified recombinant His-P68 RNA helicase is sufficient to unwind the let-7 microRNA duplex.A, 32P-labeled let-7 duplexes were analyzed for helicase activity using affinity-purified material (left panel,amounts indicated) or recombinant His-P68 (middle panel, amounts indicated). I, indicates the input into eachreaction. �T indicates heat-denatured input. The sequence and structure of each let-7 duplex are indicatednext to the companion assay. Mutations along the passenger strand are indicated in red. * and N indicate heatdenatured and native protein, respectively. B, quantification of the unwinding activity exhibited by the affinity-purified material. C, quantification of the unwinding activity exhibited by recombinant His-P68.
P68 RNA Helicase Is Required for Silencing of Gene Expression
32776 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 45 • NOVEMBER 9, 2007 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from

element (Fig. 4A). Evidence that this suppression was exertedby let-7 was provided by the observation that this suppressionwas alleviated by AntagomiRs against let-7 but not byAntagomiRs against an irrelevant microRNA (Fig. 4A).
Moreover, the suppressive activ-ity of the element was attenuatedby mutations that compromise theannealing of let-7 (Fig. 4B). In thefollowing experiment, we haveused the comparison of the lucif-erase activity between the wildtype (WT) and mutant (MUT) let-7-response elements as a measureof let-7 activity. siRNA-mediateddown-regulation of P68 RNA heli-case, but not GAPDH, attenuatedthe activity of let-7 microRNA(Fig. 4B). Importantly, GAPDHwas successfully down-regulatedas shown by Western blot analysisusing Vimentin as a loading con-trol (Fig. 4C). Thus, we concludethat P68 RNA helicase is indeedrequired for microRNA activityand is likely required for the incor-poration of the microRNA into theArgonaute2 complex.
DISCUSSION
The single-stranded small RNA-directed silencing of mRNA hasbeen reconstituted with recombi-nant Argonaute2 (18). However, theprecursor microRNA duplex-di-rected silencing of mRNA has notyet been reconstituted with the can-didate recombinant proteins. Thus,it is likely that other factors remainto be discovered. In these studies,we have identified an ATP-depend-ent unwinding activity that specifi-cally unwinds the let-7 microRNAduplex yet exhibits little activityon a derived siRNA duplex. Thisobservation reinforces the currentperception that the guide strandsof siRNA and microRNA duplexesarrive in Argonaute2 complexesthrough different pathways (20,22, 26). Indeed, it will be interest-ing to see whether RNA helicase A,which has been implicated in theunwinding of the siRNA duplex(27), is also capable of unwindingsome microRNA duplexes.Our size fractionation analysis
and affinity purification studies sug-gested that a principal component
of the unwinding activity corresponds to the P68 RNA helicase.P68 was originally identified in human cells due to its coinci-dental reactivity with a monoclonal antibody directed againstSV40 large T antigen (40).
FIGURE 4. P68 RNA helicase is required for let-7 microRNA function in HeLa cells. A, HeLa cells wereco-transfected with a �-galactosidase plasmid (100 ng) and either the parental plasmid (pIS-0) or a let-7-responsive reporter (lin41(s) WT (where WT indicates wild type)) (100 ng, as indicated), along with AntagomiRsagainst let-7 or Mir-16 (amounts indicated). Cells were incubated for 36 h and subsequently analyzed forluciferase activity. Results are normalized to cotransfected plasmid encoding �-galactosidase. B, HeLa cellswere co-transfected with a Renilla luciferase reporter (100 ng) and the pIS-0, lin-41(s) WT, or lin-41(s) MUT (whereMUT indicates mutant) (42) luciferase reporter (100 ng, as indicated) along with siRNA directed against eitherP68 RNA helicase or GAPDH (amounts indicated). Cells were incubated for 40 h and analyzed for luciferaseactivity. Results are normalized to Renilla luciferase. C, left panel, HeLa cells treated with siRNA against P68 RNAhelicase or GAPDH (amounts indicated) were analyzed for protein via Western blot. Protein levels were nor-malized to Vimentin.
P68 RNA Helicase Is Required for Silencing of Gene Expression
NOVEMBER 9, 2007 • VOLUME 282 • NUMBER 45 JOURNAL OF BIOLOGICAL CHEMISTRY 32777 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from

P68 was originally believed to be an RNA helicase due to itshomology to the DEAD box of eIF-4A, a well characterizedRNA helicase (43, 44). When P68 was assayed for unwindingactivity, it was found to be an ATP-dependent RNA helicase,which can unwind RNA duplexes in both 3� to 5� and 5� to 3�directions (30–32). The substrates unwound by P68 RNA heli-case range in size from 22 to 175 nucleotides in length andcontain overhangs of varied lengths ranging from 6 to 185nucleotides long (30–32). Thus, our observations are consist-ent with the known properties of P68 RNA helicase.P68 RNA helicase has been implicated inmany cellular func-
tions. In some cases, for example, in its perceived role as a tran-scriptional regulator, this function does not require helicaseactivity (45). In most cases, however, its touted role in mRNAsplicing, rRNA processing, and mRNA decay requires theintegrity of the helicase domain (46–48). Our studies here leadus to speculate that P68 RNA helicase might regulate theexpression of many microRNAs with a consequent pleiotropiceffect upon cellular function. It is possible that this functionmight accommodate many of the previously ascribed functionsin RNAmetabolism. In any event, the regulation of microRNAactivity would be consistent with its well described role in cel-lular proliferation. However, it remains possible that P68 RNAhelicase may only unwind particular subclasses of microRNAduplexes, and its role in a particular cell function may be pecu-liar to the miRNAs that are expressed in a given cell type.Our contention that P68 RNA helicase plays a role in the
unwinding of the let-7microRNAduplex is strengthened by theprevious observation that it is a subunit of the affinity-purifiedDrosha-processing complex (48). In addition, it has beenrecently demonstrated that mouse embryonic fibroblasts thatlack P68 RNA helicase are compromised in their expression ofmany microRNAs (48). Importantly, we show here, for the firsttime, that a recombinant RNA helicase is sufficient to unwind amicroRNA precursor duplex. From this key observation, wewould propose that P68 RNA helicase might drive the selectiveuptake of the let-7 guide strand into the silencing complex. Sofar, however, our preliminary attempts to directly demonstratethis using purified, recombinant proteins have not been suc-cessful. However, the present studies have identified an essen-tial co-factor thatmay ultimately facilitate the reconstitution ofthis critical step.
REFERENCES1. Calin, G. A., and Croce, C. M. (2006) Cancer Res. 66, 7390–73942. Ambros, V. (2001) Cell 107, 823–8263. Zamore, P. D., and Haley, B. (2005) Science 309, 1519–15244. Meister, G., and Tuschl, T. (2004) Nature 431, 343–3495. Thai, T. H., Calado, D. P., Casola, S., Ansel, K. M., Xiao, C., Xue, Y.,
Murphy, A., Frendewey, D., Valenzuela, D., Kutok, J. L., Schmidt-Sup-prian,M., Rajewsky, N., Yancopoulos, G., Rao, A., and Rajewsky, K. (2007)Science 316, 604–608
6. Lee, Y., Kim, M., Han, J., Yeom, K. H., Lee, S., Baek, S. H., and Kim, V. N.(2004) EMBO J. 23, 4051–4060
7. Lee, Y., Ahn, C., Han, J., Choi, H., Kim, J., Yim, J., Lee, J., Provost, P.,Radmark, O., Kim, S., and Kim, V. N. (2003) Nature 425, 415–419
8. Gregory, R. I., Yan, K. P., Amuthan, G., Chendrimada, T., Doratotaj, B.,Cooch, N., and Shiekhattar, R. (2004) Nature 432, 235–240
9. Han, J., Lee, Y., Yeom, K. H., Kim, Y. K., Jin, H., and Kim, V. N. (2004)Genes Dev. 18, 3016–3027
10. Han, J., Lee, Y., Yeom, K. H., Nam, J. W., Heo, I., Rhee, J. K., Sohn, S. Y.,Cho, Y., Zhang, B. T., and Kim, V. N. (2006) Cell 125, 887–901
11. Provost, P., Dishart, D., Doucet, J., Frendewey, D., Samuelsson, B., andRadmark, O. (2002) EMBO J. 21, 5864–5874
12. Hutvagner, G., McLachlan, J., Pasquinelli, A. E., Balint, E., Tuschl, T., andZamore, P. D. (2001) Science 293, 834–838
13. Macrae, I. J., Zhou, K., Li, F., Repic, A., Brooks, A.N., Cande,W.Z., Adams,P. D., and Doudna, J. A. (2006) Science 311, 195–198
14. Lee, Y. S., Nakahara, K., Pham, J.W., Kim, K., He, Z., Sontheimer, E. J., andCarthew, R. W. (2004) Cell 117, 69–81
15. Liu, J., Carmell, M. A., Rivas, F. V., Marsden, C. G., Thomson, J. M., Song,J. J., Hammond, S. M., Joshua-Tor, L., and Hannon, G. J. (2004) Science305, 1437–1441
16. Meister, G., Landthaler, M., Patkaniowska, A., Dorsett, Y., Teng, G., andTuschl, T. (2004)Mol. Cell 15, 185–197
17. Okamura, K., Ishizuka, A., Siomi, H., and Siomi, M. C. (2004) Genes Dev.18, 1655–1666
18. Rivas, F. V., Tolia, N. H., Song, J. J., Aragon, J. P., Liu, J., Hannon, G. J., andJoshua-Tor, L. (2005) Nat. Struct. Mol. Biol. 12, 340–349
19. Gregory, R. I., Chendrimada, T. P., Cooch, N., and Shiekhattar, R. (2005)Cell 123, 631–640
20. Maniataki, E., and Mourelatos, Z. (2005) Genes Dev. 19, 2979–299021. Tomari, Y., Matranga, C., Haley, B., Martinez, N., and Zamore, P. D.
(2004) Science 306, 1377–138022. Liu, X., Jiang, F., Kalidas, S., Smith,D., and Liu,Q. (2006)RNA (Cold Spring
Harbor) 12, 1514–152023. Liu, Q., Rand, T. A., Kalidas, S., Du, F., Kim, H. E., Smith, D. P., andWang,
X. (2003) Science 301, 1921–192524. Haase, A. D., Jaskiewicz, L., Zhang, H., Laine, S., Sack, R., Gatignol, A., and
Filipowicz, W. (2005) EMBO Rep. 6, 961–96725. Leuschner, P. J., Ameres, S. L., Kueng, S., and Martinez, J. (2006) EMBO
Rep. 7, 314–32026. Matranga, C., Tomari, Y., Shin, C., Bartel, D. P., and Zamore, P. D. (2005)
Cell 123, 607–62027. Robb, G. B., and Rana, T. M. (2007)Mol. Cell 26, 523–53728. Chendrimada, T. P., Gregory, R. I., Kumaraswamy, E., Norman, J., Cooch,
N., Nishikura, K., and Shiekhattar, R. (2005) Nature 436, 740–74429. Giraldez, A. J., Cinalli, R.M., Glasner,M. E., Enright, A. J., Thomson, J. M.,
Baskerville, S., Hammond, S. M., Bartel, D. P., and Schier, A. F. (2005)Science 308, 833–838
30. Iggo, R. D., and Lane, D. P. (1989) EMBO J. 8, 1827–183131. Huang, Y., and Liu, Z. R. (2002) J. Biol. Chem. 277, 12810–1281532. Hirling, H., Scheffner, M., Restle, T., and Stahl, H. (1989) Nature 339,
562–56433. Johnson, S. M., Grosshans, H., Shingara, J., Byrom, M., Jarvis, R., Cheng,
A., Labourier, E., Reinert, K. L., Brown, D., and Slack, F. J. (2005) Cell 120,635–647
34. Mayr, C., Hemann,M. T., and Bartel, D. P. (2007) Science 315, 1576–157935. Pasquinelli, A. E., Reinhart, B. J., Slack, F.,Martindale,M.Q., Kuroda,M. I.,
Maller, B., Hayward, D. C., Ball, E. E., Degnan, B., Muller, P., Spring, J.,Srinivasan, A., Fishman, M., Finnerty, J., Corbo, J., Levine, M., Leahy, P.,Davidson, E., and Ruvkun, G. (2000) Nature 408, 86–89
36. Pillai, R. S., Bhattacharyya, S. N., Artus, C. G., Zoller, T., Cougot, N.,Basyuk, E., Bertrand, E., and Filipowicz, W. (2005) Science 309,1573–1576
37. Reinhart, B. J., Slack, F. J., Basson, M., Pasquinelli, A. E., Bettinger, J. C.,Rougvie, A. E., Horvitz, H. R., and Ruvkun, G. (2000) Nature 403,901–906
38. Vella, M. C., Choi, E. Y., Lin, S. Y., Reinert, K., and Slack, F. J. (2004)GenesDev. 18, 132–137
39. Kloosterman, W. P., Wienholds, E., Ketting, R. F., and Plasterk, R. H.(2004) Nucleic Acids Res. 32, 6284–6291
40. Lane, D. P., and Hoeffler, W. K. (1980) Nature 288, 167–17041. Schwarz, D. S., Hutvagner, G., Du, T., Xu, Z., Aronin, N., and Zamore,
P. D. (2003) Cell 115, 199–20842. Lewis, B. P., Shih, I. H., Jones-Rhoades, M. W., Bartel, D. P., and Burge,
C. B. (2003) Cell 115, 787–79843. Lane, D. (1988) Nature 334, 478
P68 RNA Helicase Is Required for Silencing of Gene Expression
32778 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 45 • NOVEMBER 9, 2007 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from

44. Ray, B. K., Lawson, T. G., Kramer, J. C., Cladaras, M. H., Grifo, J. A.,Abramson, R. D., Merrick, W. C., and Thach, R. E. (1985) J. Biol. Chem.260, 7651–7658
45. Bates, G. J., Nicol, S. M., Wilson, B. J., Jacobs, A. M., Bourdon, J. C., War-drop, J., Gregory, D. J., Lane, D. P., Perkins, N. D., and Fuller-Pace, F. V.(2005) EMBO J. 24, 543–553
46. Lin, C., Yang, L., Yang, J. J., Huang, Y., and Liu, Z. R. (2005)Mol. Cell. Biol.
25, 7484–749347. Ishizuka, A., Siomi, M. C., and Siomi, H. (2002)Genes Dev. 16, 2497–250848. Fukuda, T., Yamagata, K., Fujiyama, S., Matsumoto, T., Koshida, I., Yo-
shimura, K., Mihara, M., Naitou, M., Endoh, H., Nakamura, T., Akimoto,C., Yamamoto, Y., Katagiri, T., Foulds, C., Takezawa, S., Kitagawa, H.,Takeyama, K., O’Malley, B. W., and Kato, S. (2007) Nat. Cell Biol. 9,604–611
P68 RNA Helicase Is Required for Silencing of Gene Expression
NOVEMBER 9, 2007 • VOLUME 282 • NUMBER 45 JOURNAL OF BIOLOGICAL CHEMISTRY 32779 at YALE UNIV | Kline Science Library on August 21, 2013http://www.jbc.org/Downloaded from