pa artificial and factitious foods for the production and ... · population enhancement of...
TRANSCRIPT

Duc Tung Nguyen
Artificial and factitious foods for the production and
population enhancement of phytoseiid predatory mites
2015D
uc Tung Nguyen
Artificial an
d factitiou
s foo
ds fo
r the p
rod
uctio
n and
po
pu
lation en
han
cemen
t of p
hyto
seiid pred
atory m
ites
ISBN 978-90-5989-764-9

To my family

Promoter: Prof. dr. ir. Patrick De Clercq
Department of Crop Protection,
Faculty of Bioscience Engineering,
Ghent University, Belgium
Chair of the examination committee: Prof. dr. ir. Geert Haesaert
Department of Applied Biosciences
Faculty of Bioscience Engineering,
Ghent University, Belgium
Members of the examination committee: Prof. dr. Gilbert Van Stappen
Department of Animal Production
Faculty of Bioscience Engineering,
Ghent University, Belgium
Prof. dr. ir. Luc Tirry
Department of Crop Protection,
Faculty of Bioscience Engineering,
Ghent University, Belgium
Prof. dr. ir. Stefaan De Smet
Department of Animal Production
Faculty of Bioscience Engineering,
Ghent University, Belgium
Prof. dr. Felix Wäckers
Lancaster Environment Centre
University of Lancaster, United Kingdom
Prof. dr. Nguyen Van Dinh
Department of Entomology
Faculty of Agronomy
Vietnam National University of Agriculture, Vietnam
Dean:
Prof. dr. ir. Guido Van Huylenbroeck
Rector:
Prof. dr. Anne De Paepe

Artificial and factitious foods for the
production and population enhancement of
phytoseiid predatory mites
by
Duc Tung Nguyen
Thesis submitted in the fulfillment of the requirements for the Degree of Doctor (PhD) in
Applied Biological Sciences

Dutch translation:
Artificiële en onnatuurlijke voedselbronnen voor de productie en de populatie-ondersteuning
van roofmijten uit de familie Phytoseiidae
Please refer to this work as follows:
Nguyen, D.T. 2015. Artificial and factitious foods for the production and population
enhancement of phytoseiid predatory mites. PhD Thesis, Ghent University, Ghent, Belgium
Frontcover: A female Amblyseius swirskii
Backcover: Top: dry decapsulated cysts of A. franciscana. Middle left: prepupae of black
soldier fly, Hermetia illucens. Middle right: Ephestia kuehniella eggs. Bottom:
pupae of Chinese oak silkworm Antheraea pernyi
ISBN-number: 978-90-5989-764-9
The research was conducted at the Laboratory of Agrozoology, Department of Crop
Protection, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, B-9000
Ghent, Belgium.
The author and promoter give the permission to use this study for consultation and to copy
parts of it for personal use only. Every other use is subject to copyright laws. Permission to
reproduce any material should be obtained from the author.

Acknowledgement
Undertaking this PhD has been a truly life-changing experience for me and it would not have
been obtained without the support and guidance that I received from many people. So many
people I would like to acknowledge, however, it might not be simple to express it enough.
First of all, I would like to thank my promoter, Prof. dr. ir. Patrick De Clercq for his
brilliant supervision. He gave me a valuable opportunity to do my PhD at the Laboratory of
Agrozoology, Ghent University. For me, he is not only an excellent advisor but also a
respected colleague. He is always very kind to discuss, encourage and listen to my ideas. I
much appreciate his time spent to read, correct and comment on my manuscripts, especially
during his busy time and even his holiday period to help finish my writing on time. I would
like to thank him again for his support and encouragement.
I sincerely want to thank the members of the examination committee: Prof. dr. ir.
Geert Haesaert, Prof. dr. Gilbert Van Stappen, Prof. dr. ir. Luc Tirry, Prof. dr. ir. Stefaan De
Smet, Prof. dr. Felix Wäckers and Prof. dr. Nguyen Van Dinh for their thorough reading of
my manuscript and their useful comments and suggestions that undoubtedly helped to
improve my thesis further.
I would like to give special thanks to Dominiek Vangansbeke who accompanied me
for the past four years. It is hard for me to find enough words to express my gratitude to him
for so much help and his contributions to this thesis. I feel so lucky to have him as a colleague
in my PhD project and I enjoyed very much our discussion about the research, culture, food
and whatever. I also thank Jochem Bonte for his guidance in statistical analysis. I sincerely
thank Lü Xin and Thomas Spranghers for sharing the ideas about the Chinese oak silkworm
and black soldier fly hemolymph and providing the materials for my experiments. I would
like to acknowledge the students Simon Craeye and Vincent Bouguet for their contributions to
this dissertation. I wish you all success in your research and in your careers.
I enjoyed very much working in the office which I shared with Joachim Moens and
Annelies Scholaert and later with Veerle Van Damme and Xie Jia Qin. I am deeply grateful
for their warm friendship and help. I would like to express my sincere thanks to Leen Dierick,
Bjorn Vandekerkhove, Rik Van Caenegem and Didier Van de Velde for their help with
administrative and other issues. I would like to extend my thanks to other colleagues and
friends in the Laboratory of Agrozoology: Thijs Machtelinckx, Sara Maes, Brecht Ingels,
Veronic De Puysseleyr, Jisheng Liu, Yu Na, and more.

I would like to thank Koppert B.V., Biobest N.V., the Artemia Reference Center
(ARC) of Ghent University and the Guangdong Entomological Institute, China for support
and for providing materials used in this research.
I gratefully acknowledge the funding received towards my PhD from the Ministry of
Education and Training (MOET, project 322). I would like to express my deep appreciation to
all of my colleagues at the Entomology Department, Vietnam National University of
Agriculture for their encouragement and support. Especially, I am very thankful to Prof.
Nguyen Van Dinh for all his help, advice, and encouragement.
I would like to thank all my Vietnamese friends in Ghent, especially to Anh Ngoc,
Minh Phuong, Thu Giang, Thanh Que for their help, support and understanding, especially
during the period of high pressure from work.
I am deeply thankful to my family, specifically, my parents, my mother-in-law, and
my sisters, who have supported and encouraged me throughout my life. This last word of
acknowledgment I have saved for my dear wife Thanh Ngoc, my beloved daughter Ngoc Anh
and son Duc Anh for their love, sacrifice and endless support. Without the encouragements
from them, I don’t think I could have finished my PhD study.
Nguyen Duc Tung, January 2015

i
Table of Contents
List of abbreviations .................................................................................................. iv
Chapter 1 Introduction, objectives and thesis outline ............................................ 1
1.1 General introduction .......................................................................................... 2
1.2 Objectives of the study ........................................................................................ 2
1.3 Thesis outline ...................................................................................................... 3
Chapter 2 Bio-ecology, biological control potential and rearing of Amblyseius
swirskii: a literature review ........................................................................................ 5
2.1 General biology of the Phytoseiidae (Acari) ...................................................... 6
2.2 Amblyseius swirskii ........................................................................................... 8
2.2.1 Classification ............................................................................................... 8
2.2.2 Distribution .................................................................................................. 8
2.2.3 Morphology and identification .................................................................... 9
2.2.3.1 Eggs ...................................................................................................... 9
2.2.3.2 Larvae and nymphs .............................................................................. 9
2.2.3.3 Adult ................................................................................................... 10
2.2.4 Bio-ecology ............................................................................................... 13
2.2.4.1 Development ...................................................................................... 13
2.2.4.2 Reproduction ...................................................................................... 17
2.2.4.3 Prey spectrum and feeding behaviour ................................................ 22
2.2.5 Practical application of phytoseiid mites in biological control ................. 25
2.2.5.1 Target pests and crops ........................................................................ 25
2.2.5.2 Commercial use and release strategies ............................................... 28
2.3 Rearing of natural enemies .............................................................................. 29
2.3.1 Introduction ............................................................................................... 29
2.3.2 Factitious foods ......................................................................................... 30
2.3.3 Artificial diets ............................................................................................ 33
2.3.3.1 Types of artificial diets ....................................................................... 33
2.3.3.2 Function of diet components .............................................................. 34
2.3.3.3 Role of insect components in artificial diets ...................................... 36
2.3.3.4 Artificial diets for phytoseiid mites .................................................... 37

ii
2.3.4 Pollen ......................................................................................................... 38
Chapter 3 Development and reproduction of Amblyseius swirskii on an artificial
diet enriched with pupal hemolymph of Antheraea pernyi ................................... 41
3.1 Introduction ...................................................................................................... 42
3.2 Materials and methods ..................................................................................... 43
3.2.1 Stock colony of Amblyseius swirskii ......................................................... 43
3.2.2 Stock colony of Carpoglyphus lactis ........................................................ 44
3.2.3 Pollen ......................................................................................................... 45
3.2.4 Preparation of artificial diet ....................................................................... 45
3.2.5 Rearing microcosms .................................................................................. 45
3.2.6 Experimental setup .................................................................................... 46
3.2.7 Life table parameters calculation .............................................................. 47
3.2.8 Statistical analysis ..................................................................................... 47
3.3 Results .............................................................................................................. 48
3.4 Discussion ........................................................................................................ 51
Chapter 4 Beneficial effect of supplementing an artificial diet for Amblyseius
swirskii with Hermetia illucens hemolymph ........................................................... 55
4.1 Introduction ...................................................................................................... 56
4.2 Materials and methods ..................................................................................... 57
4.2.1 Stock colony of Amblyseius swirskii ......................................................... 57
4.2.2 Black soldier fly rearing and hemolymph collecting ................................ 57
4.2.3 Preparation of artificial diets ..................................................................... 58
4.2.4 Development and reproduction on the different artificial diets ................ 58
4.2.5 Diet switching experiment......................................................................... 58
4.2.6 Life table parameters calculation .............................................................. 59
4.2.7 Statistical analysis ..................................................................................... 59
4.3 Results .............................................................................................................. 59
4.4 Discussion ........................................................................................................ 64
Chapter 5 Different factitious and artificial foods support the continuous
rearing of Amblyseius swirskii .................................................................................. 67
5.1 Introduction ...................................................................................................... 68
5.2 Materials and methods ..................................................................................... 69
5.2.1 Stock colony of Amblyseius swirskii ......................................................... 69
5.2.2 Preparation of artificial diet ....................................................................... 69

iii
5.2.3 Experimental setup .................................................................................... 70
5.2.3.1 Multigeneration experiment ............................................................... 70
5.2.3.2 Predation experiment .......................................................................... 71
5.2.4 Life table parameters calculation .............................................................. 71
5.2.5 Statistical analysis ..................................................................................... 72
5.3 Results .............................................................................................................. 72
5.4 Discussion ........................................................................................................ 79
Chapter 6 Solid artificial diets for Amblyseius swirskii ......................................... 83
6.1 Introduction ...................................................................................................... 84
6.2 Materials and methods ..................................................................................... 85
6.2.1 Stock colony of Amblyseius swirskii ......................................................... 85
6.2.2 Preparation of artificial diet ....................................................................... 85
6.2.3 Development and reproduction on the different artificial diets ................ 86
6.2.4 Pre-establishment greenhouse experiment ................................................ 86
6.2.5 Life table parameters calculation .............................................................. 87
6.2.6 Statistical analysis ..................................................................................... 87
6.3 Results .............................................................................................................. 88
6.4 Discussion ........................................................................................................ 92
Chapter 7 Performance of four species of phytoseiid mites on artificial and
natural diets .............................................................................................................. 97
7.1 Introduction ...................................................................................................... 98
7.2 Materials and methods ................................................................................... 100
7.2.1 Stock colonies of predatory mites ........................................................... 100
7.2.2 Preparation of artificial diet ..................................................................... 100
7.2.3 Experimental setup .................................................................................. 101
7.2.4 Life table parameters calculation ............................................................ 101
7.2.5 Statistical analysis ................................................................................... 101
7.3 Results ............................................................................................................ 102
7.4 Discussion ...................................................................................................... 107
Chapter 8 General discussion, conclusions and future perspectives ................. 111
Summary ................................................................................................................. 123
Samenvatting .......................................................................................................... 127
References ............................................................................................................... 133
Curriculum vitae .................................................................................................... 155

iv
LIST OF ABBREVIATIONS
ANOVA: analysis of variance
ARC: Artemia Reference Center, Ghent University
df: degrees of freedom
L:D: light-dark cycle expressed in hours (e.g. 16:8 indicates 16h photophase and 8h
scotophase)
n: number of sampled individuals
P: significance of a statistical test
R0: net reproductive rate
RH: relative humidity
rm: intrinsic rate of increase
SE: standard error
SPSS: Statistical Product and Service Solution (formerly Statistical Package for the
Social Sciences, distributed by SPSS Inc., Chicago, Illinois, USA)
T: generation time

1
Chapter 1
INTRODUCTION, OBJECTIVES AND THESIS OUTLINE

Chapter 1
2
1.1 General introduction
Predatory mites of the Phytoseiidae family are important biological control agents of
tetranychid mites and small, soft-bodied insects like thrips and whiteflies (Chant, 1985). The
subjects of this study, the phytoseiids Amblyseius swirskii Athias-Henriot, Neoseiulus
californicus (McGregor), Neoseiulus cucumeris (Oudemans), Amblyseius andersoni Chant,
and Amblydromalus limonicus Garman & McGregor (Acarina: Phytoseiidae), are all being
used on a commercial scale as biological control agents of thrips, whiteflies and spider mites
in protected vegetable and ornamental crops.
Biological control is the use of an organism to reduce the population density of
another organism. It is the most environmentally safe pest management method and can also
be economically profitable. The potential of augmentation biological control to suppress
insect pests has been recognized for many years (Doutt and Hagen, 1949; DeBach, 1964;
Parella et al., 1992). In augmentation biological control, large numbers of beneficial
arthropods (predators, parasitoids) are mass reared and released in the field (Stinner, 1977;
Collier and Van Steenwyk, 2004). Hence, a cost-effective method for their mass production is
an essential prerequisite (van Lenteren, 2003). Rearing phytoseiid mites on plant materials
infested with natural prey has several disadvantages, such as large space requirements,
inconsistent yields of predators, harvesting difficulties and variable results with different
species (McMurtry and Scriven, 1965). Rearing procedures based on factitious prey like
storage mites (Zhang, 2003; Bolckmans and van Houten, 2006) also involve space and labor
to maintain large parallel cultures of the factitious prey. Further, there may be health risks for
workers in production facilities or greenhouses caused by allergens associated with the
factitious mite prey (Bolckmans and van Houten, 2006; Fernandez-Caldas et al., 2007). The
availability of adequate artificial or factitious foods could eliminate many of the above-
mentioned problems associated with the mass production of predatory mites (Kennett and
Hamai, 1980). In addition, such foods may be useful as food supplements to support predator
populations after release in the crop (Wade et al., 2008).
1.2 Objectives of the study
The overall objective of this study was to develop alternative food sources for phytoseiid
predatory mites (with focus on the economically important species A. swirskii) in support of
their mass production and use as food supplements to sustain their populations after release in
the crop.

Introduction, objectives and thesis outline
3
The main research questions are:
1. What is the impact of different artificial and factitious diets on the development and
reproduction of A. swirskii?
2. What is the effect of these artificial and factitious diets on the developmental,
reproductive and predation performances of A. swirskii after several generations of
rearing?
3. What is the value of artificial diets for the rearing of other commercially available
phytoseiid mites?
4. What is the most adequate formulation of artificial diets for easy and effective
application in the crop as a food supplement for predatory mites?
1.3 Thesis outline
The purpose of the literature survey in Chapter 2 is to provide an overview of the
information available on phytoseiid predatory mites, with focus on A. swirskii, treating their
morphology, bio-ecology and practical application in biological control. This chapter also
includes an introduction to some aspects of the rearing of natural enemies, with particular
attention to alternative foods. In the next three research chapters, several alternative foods are
evaluated for rearing A. swirskii. Chapter 3 investigates the effects of a meridic artificial diet
supplemented with hemolymph of oak silkworm pupae (Antheraea pernyi Guérin-Méneville
(Lepidoptera: Saturniidae)) on the development and reproduction of the predatory mite. In
Chapter 4 the potential of prepupal hemolymph of black soldier fly, Hermetia illucens (L.)
(Diptera: Stratiomyidae), to improve the basic meridic artificial diet is evaluated. The chapter
focuses on the effect of the hemolymph concentration in the artificial diet on the
developmental and reproductive performance of A. swirskii. Chapter 5 examines the use of
Ephestia kuehniella Zeller eggs (Lepidoptera: Pyralidae) and decapsulated cysts of Artemia
franciscana Kellogg (Anostraca: Artemiidae) as factitious food sources and the impact of
supplementing artificial diets for A. swirskii with these foods. The developmental,
reproductive and predatory performance of the phytoseiid on these various diets after a single
generation is compared with that after 6 generations of continuous rearing. Chapter 6 focuses
on the effect of formulation on the suitability of artificial diets for A. swirskii. Whereas in
above chapters, all diets were offered in liquid form, in this chapter different solid diets are
tested. The developmental and reproductive parameters of A. swirskii are assessed when fed
on cattail pollen (Typha latifolia L.), on lyophilized forms of liquid artificial diets
supplemented with a watery extract of decapsulated cysts of A. franciscana or with pupal

Chapter 1
4
hemolymph of A. pernyi, and on solid artificial diets supplemented with powdered dry A.
franciscana cysts or with lyophilized pupal hemolymph of A. pernyi. In Chapter 7 the
impact of a meridic artificial diet supplemented with a watery extract of A. franciscana on the
biological performance of four species of phytoseiid mites which are commercially available
in Europe (N. californicus, N. cucumeris, A. andersoni, and A. limonicus) is investigated. The
final chapter (Chapter 8) presents a general discussion of the findings in this study and
provides some further research perspectives.

5
Chapter 2
BIO-ECOLOGY, BIOLOGICAL CONTROL POTENTIAL AND REARING OF
AMBLYSEIUS SWIRSKII: A LITERATURE REVIEW

Chapter 2
6
2.1 General biology of the Phytoseiidae (Acari)
The family Phytoseiidae is the most important family of acarine predators of plant pest mites
in agriculture (Huffaker et al., 1969; Helle and Sabelis, 1985; McMurtry and Croft, 1997;
Gerson et al., 2003). Phytoseiid mites are small, free-living and are the only Mesostigmata to
have extensively exploited the foliage habitat of higher plants.
Figure 2.1 Dorsal (left) and ventral (right) aspect of a typical female adult phytoseiid mite,
including setal nomenclature (Helle and Sabelis, 1985).
The adult dorsal shield of phytoseiid species is 200-600 µm in length. The adult female
idiosoma, dorsal shield or caudoventral area have less than or equal to 38, 23 or 10 pairs of
setae, respectively; paravertical setae (z1) are absent; there are one pair of setae in the R-
series or 2 pairs of setae in the r-series and no setae in the UR-series. The palpal claw has 2
tines; corniculae are slender, proximal, parallel blade-like. The tritostemum is well developed,
with 2 lacinae. Chelicera has both digits well developed, dentition variable; anterior margin of
tectum is smooth or minutely denticulate, without processes. The peritremal shield on most
species is fused anteriorly with the dorsal shield, free or fused posteriorly with the exopodal
plate beside coxa IV. The genital setae are always inserted on the genital shield of the females.
The males have sternogenital shield and ventriana shield separate. The female genital shield
truncates posteriorly, with a straight posterior margin. Most adult females have a ventrianal

A literature review
7
shield, a few species with separate ventral and anal shields or with a simple anal shield (Chant
and McMurtry, 2007).
The life cycle of phytoseiid mites typically comprises five developmental stages: egg,
larva, protonymph, deutonymph and adult; the larval stage has only six legs whereas the
nymphal and adult stages have eight legs (Woolley, 1988).
Based on habitat and food spectrum, phytoseiid mites are classified as type I
(specialized predators of Tetranychus species), type II (selective predators of tetranychid
mites), type III (generalist predators) and type IV (specialized pollen feeders/generalist
predators) (McMurtry et al., 2013). Specialist species such as Phytoseiulus persimilis Athias-
Henriot, Neoseiulus fallacis Garman, Neoseiulus longispinosus Evans, Phytoseiulus
macropilis Banks, are known as oligophagous predators of spider mites of the family
Tetranychidae (Gerson et al., 2003; Moraes et al., 2004). Generalist phytoseiids can feed on
mites of different families: eriophyids, tarsonemids, and tydeids, and on small insect species,
such as thrips or whiteflies (Gerson and Weintraub, 2012). As a special feature, generalist
phytoseiids can also feed on plant materials, including exudates (Kreiter et al., 2002;
Nomikou et al., 2003a), pollen (Nomikou et al., 2001; Kutuk and Yigit, 2011; Goleva and
Zebitz, 2013), and nectar (van Rijn and Tanigoshi, 1999a). However, generalist species may
differ in their prey preferences and in their ability to utilize certain prey as a food source,
resulting in their species-specific suitability to control particular target pests (Schausberger
and Walzer 2001).
This study is mainly concerned with the phytoseiid mite Amblyseius swirskii which is
classified by McMurtry et al. (2013) as having a type III life style. This generalist predator is
known to utilise various prey species such as tetranychid and eriophyid mites and different
insect species including whiteflies, mealybugs, and scale crawlers. Like other type III
phytoseiid mites, this species can reproduce on a diet of pollen alone. Other species used in
this study are the commercially distributed phytoseiids Neoseiulus californicus (McGregor), N.
cucumeris (Oudemans), Amblyseius andersoni Chant, and Amblydromalus limonicus Garman
& McGregor. Based on their level of food specialization, A. andersoni, N. cucumeris, and A.
limonicus are classified by McMurtry et al. (2013) as generalist predators (type III), whereas
N. californicus is a more selective predator of tetranychid mites (type II). Although N.
californicus prefers tetranychid mites as food, it can also feed on other mite species, small
insects, such as thrips, and even pollen when the primary prey is unavailable.

Chapter 2
8
2.2 Amblyseius swirskii
2.2.1 Classification
Amblyseius swirskii was described in 1962 from Beit Dagan, Central District, Israel on
Prunus amygdalus by C. Athias-Henriot (Athias-Henriot, 1962).
Synonyms:
Suspected senior synonym: Amblyseius enab El-Badry (Faraji et al., 2011)
Senior synonym: Amblyseius rykei Pritchard & Baker (Zannou et al., 2007; Zannou and
Hanna, 2011)
Other Names: Typhlodromips swirskii (Moraes et al., 1986) and Amblyseius swerski [sic]
(Abo-Taka, 1996)
The taxonomic classification of A. swirskii is as follows:
KINGDOM Animalia
SUBKINGDOM Eumetazoa
PHYLUM Arthropoda
SUBPHYLUM Chelicerata
CLASS Arachnida
SUBCLASS Micrura
INFRACLASS Acari
SUPERORDER Anactinotrichida
ORDER Mesostigmata
SUBORDER Dermanyssina
SUPERFAMILY Ascoidea
FAMILY Phytoseiidae
GENUS Amblyseius Berlese, 1904
SPECIES swirskii Athias-Henriot 1962
2.2.2 Distribution
Amblyseius swirskii is native to the Eastern Mediterranean region. It naturally occurs in Israel,
Italy, Cyprus, Greece and Egypt, and can be found on various crops like apples, apricot, citrus,
vegetables and cotton (EPPO, 2014). In North America it was first released in 1983 for

A literature review
9
control of citrus pests in California. Since 2005, A. swirskii has been released as a biological
control agent in many European countries (Austria, Belarus, Belgium, Denmark, Finland,
France, Germany, Greece, Guernsey, Hungary, Italy, Jersey, Morocco, the Netherlands,
Norway, Poland, Spain, Turkey, UK)(EPPO, 2014), North America (Arthurs et al., 2009),
South America (Argentina, Brazil) (Cédola and Polack, 2011), North Africa (Kade et al.,
2011), Saudi Arabia (Fouly et al., 2011), China (Xia et al., 2011) and Japan (Sato and
Mochizuki, 2011). Zannou et al. (2007) and Zannou and Hanna (2011) synonymized A.
swirskii with Amblyseius rykei (Pritchard & Baker). Amblyseius rykei was reported from the
Democratic Republic of Congo, Benin, Burundi, Ghana, Kenya, Malawi, Nigeria, Tanzania
and Zimbabwe (Moraes et al., 2004; Zannou et al., 2007).
2.2.3 Morphology and identification
2.2.3.1 Eggs
Eggs are oval-shaped, pale-whitish and approximately 0.20 mm in length and 0.15 mm in
width (Figure 2.2). A. swirskii lays eggs on the underside of plant leaves, mainly at the inter-
section of main and lateral ribs. Females prefer to lay eggs on leaf hairs (trichomes) near plant
domatia (small hairy tufts or pockets found on the lower surface of some leaves) which may
be an adaptation to avoid egg predators.
Figure 2.2 Egg (right) and empty egg shell (left) of Amblyseius swirskii. Scale in μm (Photo:
Author).
2.2.3.2 Larvae and nymphs
Larvae are pale white to nearly transparent in color and approximately 0.22 mm in length and
0.16 mm in width. Larva only has three pairs of legs (Figure 2.3).

Chapter 2
10
Figure 2.3 Larva of Amblyseius swirskii with three pairs of legs. Scale in μm (Photo: Author).
The protonymph (2nd
stage) is about 0.26 mm in length and 0.16 mm in width. The
deutonymph (3rd
stage) is approximately 0.28-0.34 mm in length and 0.16-0.19 mm in width
and the difference in sizes of males and females is visible. Both nymph stages have four pairs
of legs and are darker than the larvae (Figure 2.4).
Figure 2.4 Protonymph (left) and deutonymph (right) of Amblyseius swirskii. Scale in μm
(Photo: Author).
2.2.3.3 Adult
Adult females are about 0.5 mm in length and 0.3 mm in width, shiny, pear shaped, and have
an unsegmented body and four pairs of legs. Males are smaller than females with about 0.3
mm long and 0.17 mm wide, and are usually oval (Figure 2.5). They are often more active
than females or nymphs, and can run fast. Males also may be seen guarding quiescent female
deutonymphs and will mate with the females immediately after they molt. The color of mites
may vary from deep red to pale yellow depending on the prey items eaten. Mites feeding on
thrips and whitefly are generally pale yellow to pale tan, while adults fed on eggs of the

A literature review
11
aphidophagous gall midge Aphidoletes aphidimyza (Rond.) (Diptera: Cecidomyiidae) have
been observed to be yellowish-brown to red (Messelink et al., 2011).
Figure 2.5 Female adult (left) and mating adult pair (right) of Amblyseius swirskii. Scales in
μm (Photo: Author).
Like other phytoseiid species, A. swirskii is characterized by its long legs, with the
front pair pointing forward, and by the relatively few hairs (< 20 pairs) on its back. The
predator cannot be readily distinguished from some other phytoseiid mites with the naked eye,
and positive identification requires examination under a microscope. Zannou and Hanna
(2011) and Cédola and Polack (2011) described A. swirskii specimens and provided detailed
morphometrics. The general habitus and details of diagnostic setae and spermatheca are
illustrated in Figure 2.6; the chelicerae are shown in Figure 2.7.

Chapter 2
12
Figure 2.6 Amblyseius swirskii Athias-Henriot: a, dorsal view; b, ventral view; c, detail of
serrate setae Z4 and Z5; d, spermatheca. Scales in μm (Cédola and Polack, 2011).
Figure 2.7 Scanning electron microscope image of chelicera, adaxial profile, of Amblyseius
swirskii female taken at x2,700, white scale bar is 10 μm (Adar et al., 2012).

A literature review
13
2.2.4 Bio-ecology
2.2.4.1 Development
Amblyseius swirskii passes through four immature stages (egg, larva, protonymph and
deutonymph) before reaching the adult stage. The rate of development of A. swirskii is mainly
influenced by the type of food and environmental conditions. Reported immature
developmental times for this species are quite variable, ranging from 4.8 days when the
predator is fed on Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) at 26oC (Fouly et al.,
2011) to 24.1 days on pollen of Zea mays L. at 15oC (Allen, 2010) (Table 2.1). When A.
swirskii is fed on live prey it generally develops faster than when fed on pollen. For example,
the immature developmental time of the mite reared on Aculops lycopersici (Massee) (Acari:
Eriophyidae) or Tetranychus urticae Koch (Acarina: Tetranychidae) was shorter than that on
pollen of Typha latifolia L. or Ricinus communis (L.)(Abou-Awad and Elsawi, 1992; Park et
al., 2011). Temperature is another major factors affecting the development of A. swirskii. No
egg hatching was observed to occur at 13oC and 60% relative humidity (RH) (Lee and
Gillespie, 2011). The developmental times of all stages were reported by Ali and Zaher (2007)
and Xia et al. (2011) to decrease with increasing temperature from 15 to 35oC; however Allen
(2010) and Lee and Gillespie (2011) reported that when the temperature was higher than 30oC
at 70-75% RH or than 32oC at 60% RH the development of the predator slowed down. Zaher
et al. (2007) also noted that the relative humidity influenced the developmental success of A.
swirskii, with 70 and 85% RH being the most suitable for the predator at 25oC.

Chapter 2
14
Table 2.1 Duration of total developmental time and total mortality from egg to adult of
Amblyseius swirskii at different temperatures and on different prey species
Prey species
Temp-
rature
(°C)
RH (%)
Total
develop-
ment
(days)
Total
survi-
val
(%)
Reference
Acari: Eriophyidae
Aculops lycopersici (Massee) 25 70 5.1 100 Park et al. (2011)
Aculops lycopersici (Massee) 28 70 7.0 100 Momen and Abdel-Khalek (2008)
Aculus fockeui (Nalepa and
Trouessart)
28 70 6.6 100 Momen (2009)
Cisaberoptus kenyae Keifer
(mobile stages)
25 70 20.0 - Ali and Zaher (2007)
Acari: Suidasiidae
Suidasia medanensis (Oudemans) 25 70 5.0 90 Midthassel et al. (2013)
Acari: Tarsonemidae
Polyphagotarsonemus latus
(Banks)
25 80 6.2 95 Onzo et al. (2012)
Acari: Tetranychidae
Eutetranychus orientalis (Klein) 15 70 18.5 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 20 70 14.3 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 25 70 10.8 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 28 70 9.0 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 30 70 7.7 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 32 70 6.8 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 35 70 6.1 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein)
(adults)
25 70 11.3 - Ali and Zaher (2007)
Eutetranychus orientalis (Klein)
(immatures)
25 70 10.8 - Ali and Zaher (2007)
Panonychus ulmi (Koch) (adults) 25 70 13.8 - Ali and Zaher (2007)
Tetranychus truncatus (Ehara)
(eggs & larvae)
15 85-90 20.5 - Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
18 85-90 15.9 - Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
20 85-90 11.3 - Xia et al. (2011)

A literature review
15
Table 2.1 Duration of total developmental time and total mortality from egg to adult of
Amblyseius swirskii at different temperatures and on different prey species (continued)
Prey species
Temp-
rature
(°C)
RH (%)
Total
develop-
ment
(days)
Total
survi-
val
(%)
Reference
Acari: Tetranychidae
Tetranychus truncatus (Ehara)
(eggs & larvae)
25 85-90 6.6 - Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
30 85-90 5.1 - Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
35 85-90 4.0 - Xia et al. (2011)
Tetranychus urticae Koch 26 70 5.5 - El-Laithy and Fouly (1992)
Tetranychus urticae Koch 27 70-80 11.6 100 Abou-Awad and Elsawi (1992)
Tetranychus urticae Koch (eggs) 25 70 14.7 - Ali and Zaher (2007)
Tetranychus urticae Koch
(immatures)
25 70 14.2 - Ali and Zaher (2007)
Acari: Tydeidae
Tydeus californicus (Banks)
(mobile stages)
25 70 21.4 - Ali and Zaher (2007)
Acari:Tenuipalpidae
Cenopalpus pulcher (C. & F.)
(adults)
25 70 13.2 - Ali and Zaher (2007)
Hemiptera: Aleyrodidae
Bemisia tabaci (Gennadius) 26 70 4.8 - Fouly et al. (2011)
Bemisia tabaci (Gennadius) 27 - 100 Nomikou et al. (2001)
Bemisia tabaci (Gennadius) 27 - 93 Nomikou et al. (2001)
Bemisia tabaci (Gennadius) (2nd
instar nymphs)
25 70 15.3 - Ali and Zaher (2007)
Bemisia tabaci (Gennadius) (1st
instar nymphs)
25 70 15.8 - Ali and Zaher (2007)
Bemisia tabaci (Gennadius) (eggs) 25 70 16.3 - Ali and Zaher (2007)
Hemiptera: Aphididae
Aphis durantae Theobald (adults) 25 70 19.4 - Ali and Zaher (2007)
Hemiptera: Coccidae
Chrysomphalus ficus Rilly (eggs) 25 70 18.0 - Ali and Zaher (2007)
Chrysomphalus ficus Rilly
(nymphs)
25 70 17.0 - Ali and Zaher (2007)

Chapter 2
16
Table 2.1 Duration of total developmental time and total mortality from egg to adult of
Amblyseius swirskii at different temperatures and on different prey species (continued)
Prey species
Temp-
rature
(°C)
RH (%)
Total
develop-
ment
(days)
Total
survi-
val
(%)
Reference
Hemiptera: Coccidae
Coccus hesperidium (L.) (eggs) 25 70 19.0 - Ali and Zaher (2007)
Coccus hesperidium (L.) (nymphs) 25 70 18.5 - Ali and Zaher (2007)
Thysanoptera: Thripidae
Frankliniella occidentalis
(Pergande)
25 60 7.8 67 Wimmer et al. (2008)
Thrips tabaci Lind. 25 60 7.8 78 Wimmer et al. (2008)
Pollen
Phoenix dactylifera L. 25 70 12.3 - Ali and Zaher (2007)
Ricinus communis L. 27 70-80 15.0 100 Abou-Awad and Elsawi (1992)
Typha latifolia L. 25 70 6.2 100 Park et al. (2011)
Typha latifolia L. 13 60 - 0 Lee and Gillespie (2011)
Typha latifolia L. 15 60 22.1 94-100 Lee and Gillespie (2011)
Typha latifolia L. 18 60 17.5 94-100 Lee and Gillespie (2011)
Typha latifolia L. 20 60 10.9 94-100 Lee and Gillespie (2011)
Typha latifolia L. 25 60 7.0 94-100 Lee and Gillespie (2011)
Typha latifolia L. 30 60 6.0 94-100 Lee and Gillespie (2011)
Typha latifolia L. 32 60 5.4 94-100 Lee and Gillespie (2011)
Typha latifolia L. 34 60 5.6 94-100 Lee and Gillespie (2011)
Typha latifolia L. 36 60 6.9 94-100 Lee and Gillespie (2011)
Zea mays L. 25 80 7.5 99 Onzo et al. (2012)
Zea mays L. 15 70-75 24.1 - Allen (2010)
Zea mays L. 18 70-75 20.4 - Allen (2010)
Zea mays L. 20 70-75 12.8 - Allen (2010)
Zea mays L. 23 70-75 10.7 - Allen (2010)
Zea mays L. 25 70-75 8.3 - Allen (2010)
Zea mays L. 27 70-75 6.1 - Allen (2010)
Zea mays L. 30 70-75 8.8 - Allen (2010)
Zea mays L. 25-27 60-85 6.9 Zannou and Hanna (2011)
Artificial diet
Pollen artificial diet * 27 70-80 10.9 100 Abou-Awad and Elsawi (1992)
- no data presented; * Pollen artificial diet consisted of 2/3 artificial diet (yeast, milk, cystine,
proline, arginine, sucrose, and glucose) + 1/3 pollen of Ricinus communis (L.)

A literature review
17
2.2.4.2 Reproduction
Like other phytoseiid species, females of A. swirskii need mating to produce eggs
(Helle et al., 1978; Hoy, 1985). Mating frequency affects the fecundity of the females. The
reproduction of A. swirskii females kept in the company of males throughout their lifetime
was highest (47 eggs/female), followed by that of the females paired with males every 5 days
(35 eggs/female), and was lowest for the females that only had a single mating (25
eggs/female) (Zaher et al., 2007). Reported fecundities of A. swirskii in the literature vary
largely, from 1.3 eggs/female (Lee and Gillespie, 2011) to 57.6 eggs/female (Xia et al.,
2011)(Table 2.2). The reproduction of A. swirskii is affected by the food type (prey species or
plant material), food quantity and environmental conditions. The reproduction of A. swirskii
was higher on the natural prey A. lycopersici or T. urticae than on pollen of T. latifolia or R.
communis, respectively (Abou-Awad and Elsawi, 1992; Park et al., 2011). Fouly et al. (2011)
proved that food quantity offered highly affected fecundity and all life table parameters of A.
swirskii: intrinsic rate of increase averaged 0.14, 0.17 or 0.22 females/female/day when the
predatory mites were provided with 4, 8 or 12 eggs of B. tabaci/female/day, respectively.
Different temperatures also affected egg production of the predator (Ali and Zaher, 2007; Lee
and Gillespie, 2011; Xia et al., 2011). The daily oviposition of A. swirskii increased from 0.49
to 2.4 eggs/female/day when the temperature increased from 15 to 35oC (Ali and Zaher, 2007).
Zaher et al. (2007) reported that the relative humidity also influenced the fecundity of the
predator, with females laying more eggs at 70% and 80% RH (13 and 12 eggs/female/10 days,
respectively) than at 55% and 95% RH (10 and 9 eggs/female/10 days, respectively). Zaher et
al. (2007) also reported the effect of host plant leaf surface on A. swirskii oviposition; in the
latter study, the females fed on Eutetranychus orientalis (Klein) (Acari: Tetranychidae)
deposited more eggs on the smooth leathery leaves of grapefruit (15.8 eggs/10 days) than on
the coarse reticulated leaves of guava (10.8 eggs/10 days). Yousef et al. (1982) noted the
effect of photoperiod on the reproduction of A. swirskii, with increasing photoperiods
resulting in lower prey consumption and an associated drop in fecundity.

Chapter 2
18
Table 2.2 Reproduction and intrinsic rate of increase of Amblyseius swirskii at different
temperatures and on different prey species
Prey species
Tempe
-rature
(°C)
RH
(%)
Fecundity
(eggs/♀)
Oviposition
rate
(eggs/♀/day)
Intrinsic
rate of
increase
rm
Reference
Acari: Eriophyidae
Aceria ficus (Cotte) 29 70-80 - - 0.155 Abou-Awad et al. (1999)
Aculops lycopersici (Massee) 25 70 38.1 1.90 0.201 Park et al. (2011)
Aculops lycopersici (Massee) 28 70 35.4 1.70 0.235 Momen and Abdel-
Khalek (2008)
Aculus fockeui (Nalepa &
Trouessart)
28 70 43.0 1.90 0.244 Momen (2009)
Cisaberoptus kenyae Keifer
(mobile stages)
25 70 8.8 0.83 0.027 Ali and Zaher (2007)
Rhyncaphytoptus ficifoliae
Keifer
29 70-80 - - 0.122 Abou-Awad et al. (1999)
Acari: Suidasiidae
Suidasia medanensis
(Oudemans)
25 70 22.52 1.71 0.222 Midthassel et al. (2013)
Acari: Tarsonemidae
Polyphagotarsonemus latus
(Banks)
25 80 - - 0.130 Onzo et al. (2012)
Polyphagotarsonemus latus
(Banks)
19 - - 1.69 0.139 Abou-Awad et al. (2014)
Polyphagotarsonemus latus
(Banks)
28 - - 2.36 0.170 Abou-Awad et al. (2014)
Acari: Tetranychidae
Eutetranychus orientalis (Klein) 15 70 18.3 0.49 0.069 Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 20 70 22.0 0.64 0.102 Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 25 70 38.0 1.30 0.161 Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 28 70 41.8 1.60 0.202 Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 30 70 45.7 1.90 0.227 Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 32 70 47.8 2.20 0.269 Ali and Zaher (2007)
Eutetranychus orientalis (Klein) 35 70 48.7 2.40 0.289 Ali and Zaher (2007)

A literature review
19
Table 2.2 Reproduction and intrinsic rate of increase of Amblyseius swirskii at different
temperatures and on different prey species (continued)
Prey species
Tempe
-rature
(°C)
RH
(%)
Fecundity
(eggs/♀)
Oviposition
rate
(eggs/♀/day)
Intrinsic
rate of
increase
rm
Reference
Acari: Tetranychidae
Eutetranychus orientalis (Klein)
(adults)
25 70 34.2 1.12 0.155 Ali and Zaher (2007)
Eutetranychus orientalis (Klein)
(immatures)
25 70 38.0 1.30 0.161 Ali and Zaher (2007)
Panonychus ulmi (Koch) (adult) 25 70 25.2 0.70 0.098 Ali and Zaher (2007)
Tetranychus truncatus (Ehara)
(eggs & larvae)
18 85-90 23.8 0.89 0.071 Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
20 85-90 38.3 1.43 0.098 Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
25 85-90 57.6 1.93 0.148 Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
30 85-90 40.6 2.09 0.184 Xia et al. (2011)
Tetranychus truncatus (Ehara)
(eggs & larvae)
35 85-90 35.3 2.31 0.198 Xia et al. (2011)
Tetranychus urticae Koch 26 70 - 1.24* 0.167 El-Laithy and Fouly
(1992)
Tetranychus urticae Koch 27 70-80 - 1.41 - Abou-Awad and
Elsawi (1992)
Tetranychus urticae Koch
(eggs)
25 70 21.0 0.53 0.075 Ali and Zaher (2007)
Tetranychus urticae Koch
(immatures)
25 70 22.8 0.61 0.077 Ali and Zaher (2007)
Acari:Tenuipalpidae
Cenopalpus pulcher (C. & F.)
(adults)
25 70 28.2 0.82 0.106 Ali and Zaher (2007)

Chapter 2
20
Table 2.2 Reproduction and intrinsic rate of increase of Amblyseius swirskii at different
temperatures and on different prey species (continued)
Prey species
Tempe
-rature
(°C)
RH
(%)
Fecundity
(eggs/♀)
Oviposition
rate
(eggs/♀/day)
Intrinsic
rate of
increase
rm
Reference
Hemiptera: Aleyrodidae
Bemisia tabaci (Gennadius) 26 70 - - 0.220 Fouly et al. (2011)
Bemisia tabaci (Gennadius) 27 - - 1.6** 0.213 Nomikou et al.
(2001)
Bemisia tabaci (Gennadius) 27 - - 1.7** 0.208 Nomikou et al.
(2001)
Bemisia tabaci (Gennadius) (2nd
instar nymphs)
25 70 18.5 0.46 0.063 Ali and Zaher
(2007)
Bemisia tabaci (Gennadius) (1st
instar nymphs)
25 70 18.0 0.43 0.061 Ali and Zaher
(2007)
Bemisia tabaci (Gennadius)
(eggs)
25 70 16.0 0.37 0.056 Ali and Zaher
(2007)
Hemiptera: Aphididae
Aphis durantae Theobald
(adults)
25 70 9.4 0.20 0.027 Ali and Zaher
(2007)
Hemiptera: Coccidae
Chrysomphalus ficus Rilly
(eggs)
25 70 12.0 0.26 0.042 Ali and Zaher
(2007)
Chrysomphalus ficus Rilly
(nymphs)
25 70 14.0 0.31 0.047 Ali and Zaher
(2007)
Coccus hesperidium (L.) (eggs) 25 70 10.5 0.22 0.034 Ali and Zaher
(2007)
Coccus hesperidium (L.)
(nymphs)
25 70 11.0 0.23 0.036 Ali and Zaher
(2007)
Thysanoptera: Thripidae
Frankliniella occidentalis
(Pergande)
25 60 - 0.92 0.056 Wimmer et al.
(2008)
Thrips tabaci Lind. 25 60 - 0.99 0.024 Wimmer et al.
(2008)

A literature review
21
Table 2.2 Reproduction and intrinsic rate of increase of Amblyseius swirskii at different
temperatures and on different prey species (continued)
Prey species
Tempe
-rature
(°C)
RH
(%)
Fecundity
(eggs/♀)
Oviposition
rate
(eggs/♀/day)
Intrinsic
rate of
increase
rm
Reference
Pollen
Phoenix dactylifera L. 25 70 30.5 0.94 0.129 Ali and Zaher
(2007)
Ricinus communis (L.) 27 70-80 - 0.84 - Abou-Awad and
Elsawi (1992)
Typha latifolia L. 25 70 26.8 1.60 0.185 Park et al. (2011)
Typha latifolia L. 15 60 1.3 - -0.002 Lee and Gillespie
(2011)
Typha latifolia L. 18 60 2.6 - 0.016 Lee and Gillespie
(2011)
Typha latifolia L. 20 60 14.9 - 0.076 Lee and Gillespie
(2011)
Typha latifolia L. 25 60 16.1 - 0.135 Lee and Gillespie
(2011)
Typha latifolia L. 30 60 14.5 - 0.145 Lee and Gillespie
(2011)
Typha latifolia L. 32 60 10.1 - 0.160 Lee and Gillespie
(2011)
Typha latifolia L. 34 60 9.5 - 0.134 Lee and Gillespie
(2011)
Typha latifolia L. 36 60 3.3 - 0.067 Lee and Gillespie
(2011)
Zea mays L. 25 80 - - 0.160 Onzo et al. (2012)
Zea mays L. 25-27 60-85 - 1.60 0.200 Zannou and Hanna
(2011)
Artificial diet
Pollen artificial diet *** 27 70-80 - 0.90 - Abou-Awad and
Elsawi (1992)
Artificial diet **** 27 70-80 - 0.30 - Abou-Awad et al.
(1992)
- no data presented; * the value was calculated by the average total egg laying divided by the
average oviposition period; **peak oviposition rates were calculated from the average
oviposition rates of the 2nd
to 5th
day after reaching adulthood; *** pollen artificial diet
consisted of 2/3 artificial diet (yeast, milk, cystine, proline, arginine, sucrose, and glucose) +
1/3 pollen of Ricinus communis (L.); ****artificial diet consisted of yeast, milk, cystine,
proline, arginine, sucrose, and glucose

Chapter 2
22
2.2.4.3 Prey spectrum and feeding behaviour
Like other type III generalist predatory mites, A. swirskii feeds on a broad spectrum of food
sources. The predator is an effective predator of thrips and whiteflies. Further, the predatory
mite also feeds on various phytophagous mites, including tetranychid, tarsonemid and
eriophyid mites, other hemipteran insects and eggs of several lepidopterans (Table 2.3).
Table 2.3 Natural prey of Amblyseius swirskii
Prey Classification Reference
Aceria ficus (Cotte) Acari: Eriophyidae Abou-Awad et al. (1999)
Aculops lycopersici (Nalepa
& Trouessart)
Acari: Eriophyidae Momen and Abdel-Khalek
(2008); Park et al. (2010); Park et
al. (2011)
Aculus fockeui (Massee) Acari: Eriophyidae Momen (2009)
Cisaberoptus kenyae Keifer Acari: Eriophyidae Ali and Zaher (2007)
Phyllocoptruta oleivora
(Ashmead)
Acari: Eriophyidae Maoz et al. (2014)
Rhyncaphytoptus ficifoliae
Keifer
Acari: Eriophyidae Abou-Awad et al. (1999)
Polyphagotarsonemus latus
(Banks)
Acari: Tarsonemidae van Maanen et al. (2010); Onzo et
al. (2012); Abou-Awad et al.
(2014)
Cenopalpus pulcher (C. &
F.)
Acari: Tenuipalpidae Ali and Zaher (2007)
Eutetranychus orientalis
(Klein)
Acari: Tetranychidae Ali and Zaher (2007)
Panonychus ulmi (Koch) Acari: Tetranychidae Ali and Zaher (2007)
Tetranychus truncatus
(Ehara)
Acari: Tetranychidae Xia et al. (2011)
Tetranychus urticae Koch Acari: Tetranychidae Abou-Awad et al. (2000);
Messelink et al. (2010); Xu and
Enkegaard (2010); Xiao et al.
(2013)
Tydeus californicus (Banks) Acari: Tydeidae Ali and Zaher (2007)

A literature review
23
Table 2.3 Natural prey of Amblyseius swirskii (continued)
Prey Classification Reference
Bemisia tabaci (Gennadius) Hemiptera:
Aleyrodidae
Nomikou et al. (2001); Berndt et
al. (2007); Calvo et al. (2008);
Messelink et al. (2008); Chow et
al. (2010); Fouly et al. (2011);
Park et al. (2011)
Trialeurodes vaporariorum
Westwood
Hemiptera:
Aleyrodidae
Berndt et al. (2007); Messelink et
al. (2008); Medd and Greatrex
(2014)
Aphis durantae Theobald Hemiptera: Aphididae Ali and Zaher (2007)
Chrysomphalus ficus Ashm. Hemiptera: Coccidae Ali and Zaher (2007)
Coccus hesperidum (L.) Hemiptera: Coccidae Ali and Zaher (2007)
Diaphorina citri Kuwayama Hemiptera: Psyllidae Juan-Blasco et al. (2012)
Phthorimaea operculella
(Zeller)
Lepidoptera:
Gelechiidae
El-Sawi and Momen (2005)
Spodoptera littoralis Boisd. Lepidoptera:
Noctuidae
El-Sawi and Momen (2005)
Frankliniella occidentalis
(Pergande)
Thysanoptera:
Thripidae
Messelink et al. (2008); Wimmer
et al. (2008); Chow et al. (2010);
Xu and Enkegaard (2010); Calvo
et al. (2011)
Scirtothrips dorsalis Hood Thysanoptera:
Thripidae
Arthurs et al. (2009)
Thrips tabaci Lindeman Thysanoptera:
Thripidae
Wimmer et al. (2008)
Amblyseius swirskii can also develop and reproduce on non-prey foods like pollen and
honeydew (Momen and El-Saway, 1993). Cattail pollen (T. latifolia) was reported to be an
excellent food source for the laboratory rearing of A. swirskii, with short developmental time,
low mortality under different temperatures, and an acceptable fecundity (Lee and Gillespie,
2011; Park et al., 2011). Goleva and Zebitz (2013) investigated the developmental and
reproductive performance of A. swirskii fed on pollens of 21 plant species. The results showed

Chapter 2
24
that A. swirskii can feed on 19 and develop to adulthood when feeding on 18 out of the 21
tested pollens. The latter authors classified the suitability of the pollens for preimaginal
development into six groups: (1) highly suitable: Schlumbergera hybrid, Crocus vernus Hill >
Echinocereus sp. > Paulownia tomentosa (Thunb.) Steud., Aesculus hippocastanum Haynes,
(2) suitable: Ricinus communis L. > Betula pendula Roth, Zea mays L., Tulipa gesneriana L. >
Abutilon sp. > Calla palustris L., (3) ample suitable: Cucurbita pepo L. > Pinus sylvestris L. >
Narcissus pseudonarcissus L., (4) bad: bee pollen, (5) negligible suitability: Corylus avellana
L. > Helianthus annuus L. > Poaceae mix, and (6) not edible or toxic, resulting in 100 %
mortality: Lilium martagon L., Hippeastrum sp. and Hibiscus syriacus L..
The mass rearing procedures for A. swirskii are based on the use of storage mites, like
Carpoglyphus lactis L. (Acari: Carpoglyphidae) and Thyreophagus entomophagus
(Laboulbene) (Acari: Acaridae), as a food source (Bolckmans and van Houten, 2006; Fidgett
and Stinson, 2008). Baxter et al. (2011) found that using Suidasia medanensis Oudemans
(Acari: Astigmata) in sachets as a delivery system for A. swirskii was better than the use of C.
lactis as a food mite.
The degree of cannibalism and intraguild predation varies among phytoseiids
(Schausberger, 2003; Schausberger, 2004). Avoidance of kin cannibalism is considered as an
important trait for predators to increase their inclusive fitness (Schausberger, 2003; Saito,
2010). Momen and Abdel-Khalek (2009) reported that A. swirskii females did not eat
conspecific eggs or protonymphs, but did eat larvae at very low frequencies. Zhang et al.
(2014) found that the presence of A. swirskii mothers significantly affected offspring survival
in all immature stages compared with the absence of the mothers, indicating possible maternal
cannibalism. However, the difference in immature survivorship between mother-presence and
mother-absence was small, showing that kin cannibalism is relatively low. Like cannibalism,
intraguild predation is a common phenomenon amongst phytoseiid mites when prey are
scarce. Momen and Abdel-Khalek (2009) investigated the intraguild predation and
cannibalism on eggs and immatures by the adult females of Euseius scutalis (Athias-Henriot),
Typhlodromus athiasae Porath and Swirski and A. swirskii (Acari: Phytoseiidae) under
laboratory conditions. The authors found that A. swirskii had higher predation rates on
heterospecific than on conspecific prey. The predatory mite fed more on con- and
heterospecific larvae than on con- and heterospecific protonymphs.
Predation capacity is a crucial criterion to evaluate a biological control agent. El-Laithy
and Fouly (1992) noted that A. swirskii daily consumed 10.9 nymphs of T. urticae on
mulberry leaves under conditions of 26º C and 70% RH. This result was confirmed by Xu and

A literature review
25
Enkegaard (2010) who found that the predatory mite was capable of preying on spider mite
nymphs with a consumption of 4-6 nymphs within a 12 h period at 25º C and 70 % RH.
Momen and El-Saway (1993) observed a higher number of spider mites (15 nymphs/day)
being eaten by the predator at higher temperature (27º C) and a longer starvation period (2
days). Xu and Enkegaard (2010) demonstrated that A. swirskii significantly preferred first
instar larvae of F. occidentalis to both protonymphs and deutonymphs of T. urticae spider
mites. The predator could eat 4.33-4.71 first instar larvae of the thrips per day at 25º C and 70%
RH, a value that is similar to that reported by Bolckmans et al. (2005) (4.9 thrips larvae/day)
at the same temperature. Arthurs et al. (2009) found that A. swirskii was able to eat 2.73
larvae or 1.09 adults of chilli thrips, S. dorsalis, per day in a no choice test and 1.89
larvae+0.24 adults thrips/day in a choice test at 26º C; the predator significantly preferred
larval to adult thrips. With a smaller prey like the broad mite P. latus, a female adult of A.
swirskii can consume 20.2 or 31.6 adult mites/day depending on a prey density of 25 or 50
adults, respectively at 25º C (Onzo et al., 2012). Amblyseius swirskii exhibited a type II
functional response with a maximum daily predation of 15.1 eggs of T. urticae per day at 26º
C (Xiao et al., 2013). Predation rates of female A. swirskii on eggs and 1st instars of B. tabaci
were significantly higher than on the other stages of the whitefly (20 eggs or 15 first instars
being killed by a female predator per day compared to 1.2 second instars, 0.5 third instars or
0.1 fourth instars per day) at 25º C (Nomikou et al., 2004).
2.2.5 Practical application of phytoseiid mites in biological control
2.2.5.1 Target pests and crops
In commercial augmentative biocontrol A. swirskii is mainly used to control whitefly and
thrips outbreaks in greenhouse vegetables (especially cucumber, pepper and eggplant) and
some ornamental crops (chrysanthemum, roses, gerbera) in Europe and North America
(Messelink et al., 2006; Berndt et al., 2007; Buitenhuis et al., 2010b; Chow et al., 2010).
Several studies have been carried out since the initial discovery that A. swirskii has
potential as a biological control agent of agricultural pests. Most of this work has concentrated
on the management of the cotton whitefly, B. tabaci, that causes considerable yield loss and
economic injury in various crops worldwide. Several studies have deemed A. swirskii to be an
effective biological control agent of this pest. Nomikou et al. (2002) reported that A. swirskii
was able to suppress B. tabaci population growth on cucumber plants in a greenhouse. The
numbers of whiteflies on the plants without predators was significantly higher than on plants

Chapter 2
26
with predators in weeks 5 and 7–9 after infestation. This resulted in 21-fold higher
populations in the absence of predators in week 9. On average, the whitefly populations on
plants without the predator showed a 62-fold increase, while the populations in the treatments
increased 4-fold. Hoogerbrugge et al. (2005) also concluded that A. swirskii was a promising
control agent of B. tabaci in protected sweet pepper crops. Calvo et al. (2008) noted that at
release rates of 25 and 100 mites per m2 3 weeks after establishment of B. tabaci, the whitefly
nymphs were virtually eliminated. In further experiments, Calvo et al. (2009) demonstrated
that the joint release of A. swirskii and the parasitic wasp Eretmocerus mundus Mercet
(Hymenoptera: Aphelinidae) caused a significant suppression of the whitefly population when
compared with the introduction of the parasitoid alone. The density of the cotton whitefly in
the control treatment was ca. 60 nymphs/leaf after 8 weeks, which was ca. 50 times higher
than that observed in the treatment with A. swirskii released once at a rate of 50 mites/m2. The
effectiveness of A. swirskii for the control of the greenhouse whitefly T. vaporariorum on
cucumber plants was investigated by Medd and Greatrex (2014). The authors found that the
use of loose-material products of A. swirskii resulted in significantly higher mobile mite
numbers on plants than when using sachets, but this increase in mite populations did not
correspond to a significantly greater reduction in whitefly numbers. Further, in both release
methods, the A. swirskii treatments appeared not to yield significant differences in terms of
whitefly numbers from the controls. In contrast, Berndt et al. (2007) reported that the
predatory mite led to a satisfactory control of T. vaporariorum. After release of A. swirskii six
weeks, no whitefly larvae were visible on the gerbera plants throughout the experiment.
Amblyseius swirskii also shows to be a potential predator of various thrips pests.
Messelink et al. (2006) evaluated ten phytoseiid predators for the control of the western
flower thrips, F. occidentalis, on greenhouse cucumber. The authors found that A. swirskii
reached much higher population levels resulting in a significantly better control of thrips than
the standard species N. cucumeris. Arthurs et al. (2009) conducted experiments to evaluate
two species of phytoseiid mites, N. cucumeris and A. swirskii, as predators of the chilli thrips,
S. dorsalis, on sweet pepper. The authors found that both mite species established and reduced
thrips numbers significantly over 28 days following a single release (30 mites/plant).
However, A. swirskii proved a more effective predator than N. cucumeris, consistently
maintaining thrips below 1 individual per terminal leaf, compared with up to 36 for N.
cucumeris and 70 in the control treatment.
Several studies were conducted to investigate the effectiveness of A. swirskii to control
simultaneously both whiteflies and thrips. Messelink et al. (2008) reported that A. swirskii

A literature review
27
was able to control F. occidentalis in the presence or absence of T. vaporariorum, although
whitefly control was sufficient only when thrips were also present. They concluded that
control of whiteflies was improved by the presence of thrips, but that thrips control was not
affected by the presence of whiteflies. These results contradict somewhat those of Calvo et al.
(2011) who concluded that 75 A. swirskii per m2 could be an adequate rate for controlling
both B. tabaci and F. occidentalis pests either alone or simultaneously in cucumber
greenhouses.
To reduce the expenses related to releases of the predatory bug Orius sp. (Hemiptera:
Anthocoridae), an effective but relatively expensive thrips predator, it was suggested to
reduce its release rate by simultaneously releasing the predatory mite A. swirskii, which is a
less expensive predator. Weintraub et al. (2011) noted that there was no difference in F.
occidentalis control among Orius laevigatus Fieber (Hemiptera: Anthocoridae) alone and O.
laevigatus plus A. swirskii release strategies, suggesting that a reduced release rate of the
anthocorid can maintain effective thrips control. There was also no significant difference in
the quality or quantity of the sweet pepper yield between treatments in which either 2 or 6 O.
laevigatus/m2 or 100 A. swirskii plus 2 or 6 O. laevigatus per m
2 were released. Dogramaci et
al. (2011) demonstrated that A. swirskii and Orius insidiosus (Say) (Hemiptera: Anthocoridae)
were effective predators of the chilli thrips. The thrips populations were always maintained at
equivalent or lower levels under the predator combination treatments compared with the
single predator treatments. The latter study provided evidence that intraguild interactions
between the two predator species were not sufficient to prevent them from being used
simultaneously.
El-Laithy and Fouly (1992) conducted a comparison between A. swirskii and A.
scutalis, both of which in their native environments are found widely distributed and in high
abundances in association with the two spotted spider mite, T. urticae, suggesting some
potential for use as biological control agents. The authors found the oviposition period of A.
swirskii to be almost double that of A. scutalis (22.3 compared to 12.9 days). The longevity of
both mites was similar, but A. swirskii showed a much higher fecundity and prey consumption
than A. scutalis. They concluded that A. swirskii had considerable potential as an agent against
spider mites but is not as active as the specialist phytoseiid P. persimilis, which is a widely
used and very effective predator of spider mites in glasshouse crops.

Chapter 2
28
2.2.5.2 Commercial use and release strategies
Since the start of commercial production in 2005, several biological control companies have
produced A. swirskii. The major producers are Koppert B.V. (Berkel en Rodenrijs, The
Netherlands), Biobest N.V. (Westerlo, Belgium), and Syngenta Bioline Limited (Little
Clacton, UK).
Koppert B.V. commercializes this predator under the product names SWIRSKI-
MITE®, SWIRSKI-MITE LD
® and SWIRSKI-MITE PLUS
®. SWIRSKI-MITE
® contains
50,000 predatory mites (nymphs and adults) mixed with bran in a 500 ml bottle. SWIRSKI-
MITE LD®
is a paper sachet containing 125 predatory mites and storage mites mixed with
bran, with 500 sachets per outer. SWIRSKI-MITE PLUS® is also a paper sachet containing
250 predatory mites and storage mites mixed with bran, with 100 or 500 sachets per outer
(Koppert, 2014). Biobest N.V. distributes A. swirskii under the names Swirskii-System® and
Swirskii-Breeding-System®
. Swirskii-System® consists of 25,000 or 50,000 predatory mites
in a 500 ml plastic pot. A Swirskii-Breeding- System®
breeding sachet contains approximately
250 A. swirskii in a carrier of bran and feeder mites. Each Swirskii-Long-Life-System® sachet
contains approximately 150 A. swirskii in a carrier of bran, 2 feeder mite species and an
alternative food source for these feeder mites (Biobest, 2014). Syngenta Bioline Limited
supplies the predatory mite under the products name Swirskiline®
and Bugline swirskii.
Swirskiline® Loose is a shaker tube of 1 litre containing 25,000 mites. Swirskiline
® Bulk is a
5 litre bag containing 125,000 mites. The Swirskiline®
sachet is a Gemini and hooked sachet,
each sachet containing a breeding colony of 250 predators at the time of packing. Bugline
Swirskii® is a string of sachets that can be laid onto the crop along a plant row (Syngenta,
2014).
Amblyseius swirskii is not susceptible to diapause (Bolckmans et al., 2005) it can be
used throughout much of the season provided daytime temperatures regularly exceed 22°C.
The mites are released directly in the crops in bran or vermiculite carriers sprinkled on the
leaves or substrates, but they can also be released via sachets (see below), or may be
broadcast via air blast or other automated distribution systems (Opit et al., 2005; Buitenhuis et
al., 2010a). The recommended release rates are typically between 25 and 100 mites per m²
depending on pest species, pest density, and crop. Preventive dispersal consists of 20 mites/m²,
when thrips or whiteflies are present 50 mites/m² are recommended. When the plants are
heavily infested by thrips or whiteflies the predatory mites are released at a rate of 100

A literature review
29
mites/m² in infested areas only and always in combination with other beneficials (Koppert,
2014).
Slow-release sachets (breeding systems) that contain a substitute prey (bran mites)
have been developed, and allow gradual release of the predators through a small hole in the
sachet over several weeks (Baxter et al., 2011). These sachets are water resistant and provide
resources for the reproducing mites during the release period. In this way A. swirskii can be
kept in the crop for longer periods, offering more protection, which makes it interesting for no
pollen bearing crops such as cucumber. Sachets containing 150-250 mites can be suspended
on the plants, at a rate of 1 sachet/2 m². Introduction can be repeated every 4 weeks to
maintain a continuous presence of A. swirskii in the crop (Biobest, 2014).
For crops producing little or no pollen like certain ornamental crops, vegetable crops,
and in plant nurseries, a food supplement can be distributed in the crop to enhance
establishment and population growth of A. swirskii or to help the predator population survive
periods of low prey availability. For instance, the pollen product NutrimiteTM
(consisting of
Typha angustifolia L. pollen) has been commercialized by Biobest N.V. for this purpose. The
product can be sprayed over the crop by a Nutrigun at a dosage of 500g/ha every 2 weeks
(Biobest, 2014).
2.3 Rearing of natural enemies
2.3.1 Introduction
In augmentative biological control natural enemies are mass-reared in bio-factories for release
in large numbers to obtain an immediate control of pests (van Lenteren, 2012). However,
augmentation is applied on a commercial scale in relatively few agricultural systems (van
Lenteren, 2012). One of the main reasons of the relatively low adoption of this pest
management strategy (van Lenteren, 2012) is that augmentative releases are frequently more
expensive than chemical pesticides (Collier and Van Steenwyk, 2004). Cost-effective rearing
techniques are therefore needed to make augmentation a more competitive strategy for
managing arthropod pests. One approach to facilitating this is reducing the costs associated
with rearing natural enemies by using factitious or artificial foods instead of their natural prey
or hosts (Riddick, 2009).

Chapter 2
30
2.3.2 Factitious foods
Factitious prey (hosts) are typically live, frozen, lyophilized or irradiated insects, mites and
crustaceans that support the development and reproduction of predators (parasitoids) in lieu of
natural or target prey (hosts). These food sources can not only be used for the mass production
of natural enemies but also used as a preventative strategy to help establish or maintain a
population of certain beneficial arthropods in the crop when pest populations are low, so as to
reduce the frequency of releases (Jonsson et al., 2008). The success of rearing a predator on a
factious food highly depends on its feeding habits, i.e. whether the predator is polyphagous,
oligophagous, or monophagous. Highly polyphagous species have better chances of surviving
and reproducing on factitious prey (Riddick, 2009).
Eggs of certain lepidopterans have been found to be a nutrient-rich food for several
generalist coccinellids (Herrera, 1960; Riddick, 2009). Specty et al. (2003) noted that eggs of
the Mediterranean flour moth Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) contained a
higher percentage of amino acids and lipids than the pea aphid Acyrthosiphum pisum (Harris)
(Hemiptera: Aphididae), and found that E. kuehniella eggs could support the growth and
reproduction of the multicolored Asian lady beetle Harmonia axyridis (Pallas) (Coleoptera:
Coccinellidae) to a similar and better extent as the aphid, respectively. Attia et al. (2011) and
Maes et al. (2014) proved that Cryptolaemus montrouzieri Mulsant (Coleoptera:
Coccinellidae) larvae developed faster on E. kuehniella eggs than on eggs of the mealybug
Planococcus citri (Risso)(Hemiptera: Pseudococcidae), which is the predator's natural prey.
De Clercq et al. (2005b) reported that adults of Adalia bipunctata (L.) (Coleoptera:
Coccinellidae) reared on live pea aphids, A. pisum were only half as fecund as those offered
irradiated or frozen E. kuehniella eggs, but egg hatch was significantly better on live aphids
than on flour moth eggs. Ephestia kuehniella eggs have also shown to be a suitable factitious
food for anthocorid bugs. Alauzet et al. (1992) reared Orius majusculus (Rt.) (Hemiptera:
Anthocoridae) on eggs of E. kuehniella, larvae of Cacopsylla pyri (L.) (Hemiptera: Psyllidae)
and Rhopalosiphum padi (L.) (Hemiptera: Aphididae) at five temperatures (12.5, 15, 20, 25
and 30oC). They found that at all temperatures the nymphs fed on E. kuehniella eggs
developed faster than those given the other food sources. Vacante et al. (1997) noted that a
high percentage nymphs of both Orius albidipennis (Reuter) and O. laevigatus (Hemiptera:
Anthocoridae) successfully developed to adult on diets containing E. kuehniella eggs. Chyzik
et al. (1995) reported that the fecundity and survival Orius albidipennis (Reuter) (Hemiptera:
Anthocoridae) on the onion thrips T. tabaci and on eggs of Ephestia cautella Walker

A literature review
31
(Lepidoptera: Pyralidae) were significantly higher than on the spider mite T. urticae; female
longevity was significantly higher on Ephestia eggs than on thrips and mites. Fauvel et al.
(1987) reported that the daily fecundity of Macrolophus caliginosus Wagner (Hemiptera:
Miridae) females reared on E. kuehniella eggs was approximately 3 or 8 times higher than
that of females reared on Aphis gossypii Glover (Hemiptera: Aphididae) or Tetranychus
turkestani Ugarov & Nycolsky (Acari: Tetranychidae), respectively. Vandekerkhove et al.
(2006) reported that M. caliginosus females fed E. kuehniella eggs had superior ovarian
scores and laid more eggs than those fed an artificial diet based on egg yolk. López-Arroyo et
al. (1999) evaluated developmental performance of the lacewings Ceraeochrysa cincta
(Schneider), Ceraeochrysa cubana (Hagen), and Ceraeochrysa smithi (Navas) (Neuroptera:
Chrysopidae) when larvae were reared on eggs of the moths E. kuehniella or Sitotroga
cerealella (Olivier) (Lepidoptera: Gelechiidae), or on the peach aphid Myzus persicae (Sulzer)
(Hemiptera: Aphididae). In all chrysopid species, the predators fed on moth eggs developed
significantly faster and in some cases had higher adult weights than those reared on aphids.
Pappas et al. (2011) evaluated the effect of E. kuehniella eggs and the nymphs of the aphid A.
pisum on the development and reproduction of Dichochrysa flavifrons (Brauer) and
Dichochrysa zelleri (Schneider) (Neuroptera: Chrysopidae). Preimaginal development of both
species was significantly shorter when their larvae were fed on E. kuehniella eggs than on A.
pisum nymphs. In D. flavifrons, the intrinsic rate of increase (rm ) was higher when larvae
were fed on E. kuehniella eggs than on A. pisum nymphs, while the converse was recorded in
D. zelleri. Few studies have, however, tested lepidopteran eggs for predatory mites. The eggs
of E. kuehniella were reported to support the development of the phytoseiid mite Iphiseius
degenerans (Berlese) (Acari: Phytoseiidae): 80% of the tested larvae reached adulthood, the
developmental time of females fed on this diet was same as that of those fed on sweet pepper
pollen or T. urticae brushed off onto the rearing arena , but longer than that of females reared
on almond, apple, castor bean or plum pollen (Vantornhout et al., 2004).
Cysts of the brine shrimp Artemia sp. (Anostraca: Artemiidae), a routinely used feed
in aquaculture (Van Stappen, 1996), are a non-insect type of factitious food which has been
tested for rearing a number of insect predators. Artemia cysts may have a number of practical
advantages over E. kuehniella eggs when used for rearing predatory arthropods. The market
price of decapsulated Artemia cysts is substantially lower than that of E. kuehniella eggs and
the cysts can be stored in dry form for several years in a cool and dry place without the need
for deep freezing (Arijs and De Clercq, 2001). Arijs and De Clercq (2001) were the first to
test cysts of the brine shrimp A. franciscana as a food for a heteropteran predator. The authors

Chapter 2
32
compared the development and reproduction of the anthocorid O. laevigatus on A.
franciscana cysts versus that on E. kuehniella eggs. The predators fed on hydrated
decapsulated cysts had similar developmental and reproductive rates as those fed frozen
Ephestia eggs. A follow-up study by Bonte and De Clercq (2008) confirmed that brine shrimp
cysts are suitable food for this anthocorid. Riudavets et al. (2006) compared the reproduction
of the mirid M. caliginosus and the anthocorid Orius majusculus (Reuter) (Hemiptera:
Anthocoridae), using dry cysts of Artemia sp. or frozen E. kuehniella eggs as food sources.
Whereas numbers of offspring of M. caliginosus females fed on either diet were similar, the
survival rate and numbers of offspring of O. majusculus fed on E. kuehniella eggs were higher
than on Artemia cysts. Castañé et al. (2006) found that M. caliginosus reared on dry or
hydrated cysts of Artemia sp. had the same nymphal survivorship, nymphal developmental
time, adult weight and fecundity as that obtained with E. kuehniella eggs. However, nauplii of
the brine shrimp yielded a significant reduction in survivorship, a delay in nymphal
development and a lower reproduction. Whereas Artemia cysts are less expensive than E.
kuehniella eggs and can support the development and reproduction of several predatory bugs,
prolonged rearing on the cysts as a sole food has been associated with fitness losses, at least in
Orius bugs (De Clercq et al., 2005a). For solving this problem, currently dry Artemia cysts are
routinely mixed with E. kuehniella eggs as a food for the production of different predatory
heteropterans; this practice may help to reduce inputs of expensive lepidopteran eggs and thus
rationalize the production process of these economically important predators (De Clercq et al.,
2014).
In the mass production of various generalist phytoseiids, storage mites, such as
Carpoglyphus lactis L. (Acari: Carpoglyphidae), Thyreophagus entomophagus (Laboulbene)
(Acari: Acaridae), Lepidoglyphus destructor (Acari: Glyciphagidae) and Suidasia medanensis
(Acari: Suidasiidae) are being used as primary food sources instead of natural prey
(Bolckmans and van Houten, 2006; Fidgett and Stinson, 2008; Midthassel et al., 2013).
Schliesske (1981) first used Tyrophagus casei (Oudemans) (Acari: Acaridae) as alternative
prey for N. cucumeris and Neoseiulus barkeri (Hughes) (Acari: Phytoseiidae). The author
found that both phytoseiid mites successfully developed and reproducted on this prey.
Bolckmans and van Houten (2006) tested reproduction of A. swirskii on juveniles and adults
of C. lactis and found that the daily oviposition and total fecundity of the females fed on
juvenile stages of the prey were similar as that on adult stages of the prey (1.80
eggs/female/day and 29 eggs over 16 days versus 1.84 eggs/female/day and 33 eggs over 18
days, respectively). The authors also noted that A. cucumeris was able to reproduce on C.

A literature review
33
lactis (2.13 eggs/female/day). Fidgett and Stinson (2008) reported that population of A.
cucumeris reared on T. entomophagus increased 10 fold within 14 days. This value is more
than double that routinely achieved for A. cucumeris in the commercial production on a diet
of Tyrophagus putrescentiae Schrank (Acari: Acaridae). The latter authors also indicated that
T. entomophagus and C. lactis were suitable food sources for A. swirskii, although the
population of the predator fed on the first prey was 38% greater than that on the second prey.
2.3.3 Artificial diets
Rearing arthropod parasitoids and predators in vivo requires the simultaneous rearing of
multiple organisms. Artificial diets are intended to eliminate the need to rear host (or prey)
species, thereby reducing the complexity of the system to manageable levels (Morales-Ramos
et al., 2014). Artificial diets allow rapid build-up of arthropod parasitoids or predators without
the time lag required to build up host or prey populations. It also allows to avoid risks of
diseases which may occur in the host, prey or plant culture. King et al. (1985) considered the
development of artificial diets may contribute to increase the success of augmentative
biological control, and Cohen (1992) considered the lack of suitable artificial diets the
greatest barrier to the mass production of entomophagous insects.
2.3.3.1 Types of artificial diets
Several terms are used to describe types of artificial diets. Dougherty (1959) classified
artificial diets according to three categories: holidic, meridic and oligidic, depending on
ingredients used. Holidic diets pertain to media whose intended constituents, other than
purified inert materials, have an exactly known chemical structure before compounding.
Meridic diets pertain to media composed of a holidic base to which is added at least one
substance or preparation of unknown structure (for example, protein, regardless of “purity”)
or of uncertain purity. Oligidic diets pertain to media in which crude organic materials supply
most dietary requirements. However, the distinction between these three classifications has
not always been clear; for this reason Grenier and De Clercq (2003) suggested another
classification system separating artificial diets based on the presence or absence of insect
components (i.e., tissues, hemolymph, cells, protein, amino acids, etc.). Based on formulation,
artificial diets can also be classified as (1) diets as powder or fragments, (2) semiliquid diets

Chapter 2
34
for chewing phytophages, (3) liquid diets for sucking phytophages, or (4) liquid diets for
endoparasitoids (Parra, 2012).
2.3.3.2 Function of diet components
An artificial diet for arthropods normally contains several main ingredients, including a
nitrogen source (e.g. protein, free amino acids), carbohydrates, lipids, vitamins, minerals and
water. Sometimes, other ingredients like stabilizers, preservatives, “fillers” or bulking agents
are added to the diet which helps to preserve its structure. Most successful diets contain
special components called “token stimuli” that do not have a direct nutritional function but
stimulate feeding responses (Cohen, 2004). A diet must provide all essential nutrients to allow
an arthropod to complete its development and reproduction. Whereas most nutritional studies
were performed on insect diets, very few works in this field have been done with mites.
Proteins are the source of amino acids required for the production of tissues and
enzymes in insects. Some amino acids, such as arginine, lysine, leucine, isoleucine,
tryptophan, histidine, phenylalanine, methionine, valine and threonine, are essential for
normal growth of an arthropod. Others are essential for certain species such as glycine for
some dipterous insects, alanine for Blattella cockroaches and proline for Phormia blowflies.
A good balance between different amino acids is of particular importance (Chapman, 1998).
The protein in the diet plays an important part in egg production (Johansson, 1964). Protein is
of primary importance for yolk production and is essential for oogenesis in many insects
(Chapman, 1998). In Calliphora blowflies, for instance, the intake of protein during the early
stages of egg development activates the corpora allata to secrete a factor leading to an
increase in carbohydrate intake during the period of yolk deposition (Strangways-Dixon,
1959).
Lipids including fatty acids, phospholipids and sterols are components of cell walls as
well as having other specific functions. Insects are able to synthesize many fatty acids and
phospholipids, so they are not usually essential dietary constituents, but many insects do
require a dietary source of polyunsaturated fatty acids, and all insects require sterols
(Chapman, 1998). The addition of lipids such as cholesterol has contributed to the success of
artificial diets for various insects (Hobson, 1935; Gupta et al., 2005b; Gupta et al., 2005a).
Carbohydrates are used as fuels by a majority of insects. They may be converted to
fats, and may contribute to the production of amino acids. They are, therefore, important
components of the diets of most insects, but they are not necessarily essential because they
can be synthesized from fats or amino acids (Chapman, 1998). Most insects are able to absorb

A literature review
35
and metabolize fructose and glucose, but some monosaccharides such as arabinose, ribose,
xylose, and galactose, while readily absorbed, are not metabolized (Chippendale, 1978).
Generalist feeders like herbivores and predators are able to digest disaccharides, such as
sucrose and maltose, while some parasitoids and mites are not (Singh, 1977; Cohen, 2004).
However, it is worth noting that the requirement for carbohydrates in the diet is species
dependent. Many insects, including key biocontrol agents, are fully dependent on nectar or
honeydew feeding to obtain the carbohydrates essential for their survival (Wäckers and van
Rijn, 2005).
Vitamins are divided into two groups depending on their solubility in water or lipids.
Insects need vitamins for growth and in general they can not synthesize them (Chapman,
1998). Vitamin B, a water-soluble vitamin, includes seven compounds: thiamine, riboflavin,
nicotinic acid, pyridoxine, pantothenic acid, folic acid and biotin that are required by all
insects (Chapman, 1998). Another water-soluble vitamin, vitamin C is known to play a role in
the molting and detoxification processes. It is more important in herbivorous than in
entomophagous insects (Chapman, 1998; Cohen, 2004). Lipid-soluble vitamins include
retinol, carotenoids (A), tocoferols (E), calciferol (D), and phyloquinone (K). Only vitamins A
and E are known to be required in insects, where they play a role in the synthesis of pigments
and in reproduction, respectively (Chapman, 1998).
Minerals are nearly always present as impurities in any artificial diet so that very little
work on the precise amounts required by insects has been undertaken (McFarlane, 1991).
Sodium, potassium, calcium, magnesium, chloride and phosphate are essential components of
the diet of all insects because they play a role in the functioning of insect cells (Chapman,
1998). Iron, zinc and manganese are also essential elements in insect diets because the first
element joins in cytochromes while the last two elements play a part in hardening the cuticle
in many insects (Chapman, 1998).
Water is the most fundamental nutrient and it plays an important role in all life
processes. Most insects take up sufficient water from their food or from a drinking source;
only a small group of insects can use metabolic activities to create water themselves. The
water content of an insect diet is a crucial factor: if it is too low the diet may not provide
enough water for the insect or cause difficulty in food intake, whereas if it is too high it may
encourage microbial contamination. Cohen (2004) suggested that the normal amount of water
present in the artificial diet should be the same as that in the insect's natural food. However,
sometimes water content is confused with water activity which is a thermodynamic concept
indicating the availability of water present in a given material. Water activity is a measure of

Chapter 2
36
the potential of water to move from one region to another. Different diets with the same water
content can have different water activities, resulting in different water availability for the
insect (Cohen, 2004).
2.3.3.3 Role of insect components in artificial diets
Grenier and De Clercq (2003) noted that adding insect components (i.e. tissues, hemolymph,
cells, protein, amino acids, etc.) to artificial diets enhances their acceptability and improves
their nutritional quality for a number of entomophagous insects. Artificial diets containing
insect components may be useful when predators or parasitoids require certain nutrients,
feeding stimulants, and other chemical cues found in arthropod prey or hosts (De Clercq,
2008). For instance, many artificial media for egg parasitoids of the genus Trichogramma
contain lepidopteran pupal hemolymph and/or pupal holotissues to stimulate egg laying by the
parasitoid females or to provide adequate nutrition for development of the larvae. Strand and
Vinson (1985) obtained complete in vitro culture of Trichogramma pretiosum Riley
(Hymenoptera: Trichogrammatidae) on an artificial medium supplemented with 40%
hemolymph of Manduca sexta (L.) (Lepidoptera: Sphingidae). Trichogramma dendrolimi
(Matsumura) and Trichogramma maidis Pint. et Voeg. (Hymenoptera: Trichogrammatidae)
successfully developed on artificial diets composed of egg yolk and Helicoverpa zea (Boddie)
(Lepidoptera: Noctuidae) hemolymph (Grenier and Bonnot, 1988). Trichogramma dendrolimi
was successfully reared in vitro in a medium containing hemolymph of Antheraea pernyi
Guérin-Méneville (Lepidoptera: Saturniidae) (Guan et al., 1978). The insect components also
play a positive role in the development and reproduction of other parasitoids. Dindo (1995)
reported successful in vitro culture of Brachymeria intermedia (Nees) (Hymenoptera:
Chalcididae) on bovine and chicken based commercial foods supplemented with 10-20%
pupal homogenate of Galleria mellonella L. (Lepidoptera: Pyralidae). Liu et al. (1988) noted
that the parasitoid wasp Anastatus japonicus Ashmead (Hymenoptera: Eupelmidae) could
completely develop on artificial diet containing hemolymph from the pupae of A. pernyi or
Philosamia cynthia ricini Donovan (Lepidoptera: Saturniidae) and longevity and fecundity of
artificially reared females were greater than those of parasitoids reared on host silkworm eggs.
Approximately 60% of the egg parasitoid Tetrastichus schoenobii Ferriere (Hymenoptera:
Tetrastichidae) reared on artificial diet supplemented with hemolymph from A. pernyi
completed development to the adult stage (Ding et al., 1980). Fanti and Bratti (1991) found
that a diet supplemented with G. mellonella pupal hemolymph and 20-hydroxyecdysone could

A literature review
37
support the development of the parasitoid fly Pseudogonia rufifrons (Wiedemann) (Diptera:
Tachinidae).
In comparison with insect parasitoids, the use of insect components in artificial diets
for insect predators is less studied. Ferkovich and Shapiro (2004) found that the fecundity of
the predatory anthocorid O. insidiosus was significantly increased when the bugs were fed on
an artificial diet supplemented with a cell line derived from eggs of the Indian meal moth
Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae). Matsuka and Okada (1975) reported
that the addition of pulverized drone honeybee powder to diets (consisting of yeast, chicken
liver, banana, royal jelly, pollen, and dried milk) slightly enhanced diet efficacy on
development of the ladybird H. axyridis. Bonte et al. (2010) fed the two-spot ladybird A.
bipunctata with an artificial diet based on a beef and chicken protein supplemented with
pollen and Artemia cysts. They found that developmental time increased, and body size and
fecundity of the coccinellid decreased in comparison to a diet of pea aphids. However, larval
survival rate and adult longevity were unaffected by the artificial diet as compared with the
natural aphid prey. Matsuka et al. (1982) found that adult females of Rodolia cardinalis
Mulsant (Coleoptera: Coccinellidae) lived longer but produced significantly fewer eggs when
fed an artificial diet of powdered honeybee brood with sucrose rather than live prey, Icerya
purchasi Maskell (Hemiptera: Margarodidae). Ogura and Hosoda (1995) tried to rear
Trogossita japonica Reitter (Coleoptera: Trogossitidae) on artificial diets consisting of
silkworm pupal powder, dry yeast, sucrose, peptone, squid liver oil, agar, and distilled water.
The authors found that the diet supported the development of the larvae, but diet-reared larvae
yielded smaller adults, which mated and oviposited at normal levels when provided with
natural prey.
2.3.3.4 Artificial diets for phytoseiid mites
Whereas a good body of literature exists on artificial diets for insect predators, far fewer
attempts have been made at rearing predatory mites on artificial diets (Singh, 1977).
McMurtry and Scriven (1966) reported longer developmental times and lower oviposition
rates when four phytoseiids (Amblydromalus limonicus Garman and McGregor, Amblyseius
hibisci (Chant), Typhlodromus occidentalis Nesbitt and Typhlodromus rickeri Chant) (Acari:
Phytoseiidae) were fed on various artificial diets (sucrose, molasses, yeast + sucrose or yeast
+ molasses) compared with mite prey and pollen as food sources. Shehata and Weismann
(1972) tested three artificial diets for the specialist spider mite predator P. persimilis. Their
results indicated that the larvae could develop to adults but the females failed to produce

Chapter 2
38
viable eggs. Kennett and Hamai (1980) investigated oviposition rate and developmental
capacity of 9 predaceous mites (A. hibisci, A. limonicus, Amblyseius largoensis (Muma),
Metaseiulus pomoides Schuster & Pritchard, T. occidentalis, Typhloseiopsis arboreus (Chant),
Typhloseiopsis pyri Scheuten, P. persimilis, and I. degenerans) fed on artificial and natural
diets. The authors reported that complete development and oviposition occurred for seven out
of nine species when fed on an artificial diet consisting of bee honey, sugar, yeast flakes,
yeast hydrolysate, enzymatic casein hydrolysate and fresh egg yolk. Oviposition rates of all
species fed on the artificial diet were lower than those on a natural diet. Ochieng et al. (1987)
reported that Amblyseius teke Pritchard and Baker could complete more than 25 generations
when reared on a liquid diet composed of bee honey, milk powder, egg yolk and Wesson's salt.
Abou-Awad et al. (1992) noted that the predacious mites Amblyseius gossipi El-Badry and A.
swirskii developed and reproduced successfully on artificial diets composed of yeast, milk,
cysteine, proline, arginine, sucrose, glucose, streptomycin sulphate and sorbic acid. However,
fecundity of both species fed on the artificial diet was lower than on natural prey, although the
eggs showed no abnormalities. Shih et al. (1993) conducted experiments to investigate the
responses of Euseius ovalis (Evans) to natural food resources and two artificial diets (diet A
consisted honey 15 g, ascorbic acid 100 mg, vitamin B complex 300 mg, Bovril (a
commercial beef extract) 500 mg, choline chloride 0.1 g, and distilled water 120 mL; and diet
B consisted D (-) fructose 10 g, fresh egg yolk 20 g, yeast powder 10 g, honey 10 g, choline
chloride 0.1 g, distilled water 180 ml). Immature development was successful in the first
generation but offspring was not able to complete its life cycle when maintained on the
artificial diets. The females of E. ovalis fed on artificial diets showed a shorter oviposition
period, lower daily and total reproductive rates, and shorter longevity than those fed on
natural diets. Ogawa and Osakabe (2008) investigated the development and survival of
Neoseiulus californicus (McGregor) on an artificial diet (composed of honey, sucrose,
tryptone, yeast extract, fresh egg yolk, and distilled water). The phytoseiid successfully
developed on the artificial diet, but only few eggs were deposited. The above studies suggest
that, although phytoseiid mites can develop on different artificial diets, fecundity in most
cases was inferior to that on natural or factitious prey.
2.3.4 Pollen
Pollen is an extremely rich source of many of the nutrients essential to the development and
reproduction of entomophagous arthropods. Pollen provides important food elements:
proteins, free amino acids, carbohydrates, lipids, vitamins, flavonoids, and minerals are all

A literature review
39
common in pollen grains. In fact, the protein and oil contents of pollens are both qualitatively
and quantitatively superior to those of most vegetative tissues and even many prey items.
Given its nutritional status, it is not surprising that many insects that ordinarily do not
consume plant tissues eat pollen, and that some can even complete development upon it in the
absence of prey (Lundgren, 2009b).
Predatory mites in several families (e.g. Phytoseiidae, Erythraeidae, Cheyletidae,
Stigmaeidae) feed on pollen to varying degrees. As stated above, McMurtry et al. (2013)
classify the feeding behavior of phytoseiid predatory mites into four groups, in which only
type I specialized predators of Tetranychus species not feed on pollen, whereas type II, III and
IV predatory mites are found to be more or less feeding on pollen. In the type II group, pollen
promotes reproduction of certain species, but at a much lower rate than mite prey. Most
species feeding on pollen belong to Neoseiulus genus including N. californicus (Swirski et al.,
1970), Neoseiulus fallacis (Garman) (Afifi et al., 1988), Neoseiulus idaeus Denmark & Muma
(Tanigoshi et al., 1993), Neoseiulus longispinosus (Evans) (Mori, 1977) (Acari: Phytoseiidae)
and one from Typhlodromus genus (T. rickeri) (Overmeer, 1985). In comparison to type II,
more species in the type III group have been reported feeding on pollen. These mites can
reproduce on pollen and in some species the reproduction rate is at least as high as on animal
prey (McMurtry and Rodriguez, 1987; Marisa and Sauro, 1990; Duso and Camporese, 1991).
The group of type IV specialized pollen feeders comprises only the genus Euseius; members
of this group (e.g. Euseius tularensis (Congdon) (Acari: Phytoseiidae)) can subsist on pollen
in the absence of prey with minimal reductions in fitness (Grafton-Cardwell et al., 1999).
Population increase of these species often can be correlated with pollen fallout onto the
foliage rather than the presence of any prey species (McMurtry, 1992).
In the practice of biological control, several species of predatory mites can be
provided with pollen as a supplementary or alternative food source in order to avoid
cannibalism when prey gets scarce (van Rijn and Tanigoshi, 1999b; Nichols and Altieri,
2004). Pollen can be supplied as an additional food source either by using banker plants
(Ramakers and Voet, 1996; Huang et al., 2011), by artificial pollen sources (e.g., plastic cups
with pollen placed between the crop plants) (Nomikou et al., 2010), by dusting or spraying
pollen from such various plants as cattail, castor bean, and maize (van Rijn et al., 2002;
Skirvin et al., 2006).

Chapter 2
40

41
Chapter 3
DEVELOPMENT AND REPRODUCTION OF AMBLYSEIUS SWIRSKII ON AN
ARTIFICIAL DIET ENRICHED WITH PUPAL HEMOLYMPH OF ANTHERAEA
PERNYI
This chapter is based on:
Nguyen, D. T., Vangansbeke, D., Lü, X., & De Clercq, P. 2013. Development and reproduction of
the predatory mite Amblyseius swirskii on artificial diets. BioControl, 58: 369-377.

Chapter 3
42
3.1 Introduction
Amblyseius swirskii (Athias-Henriot) (Acari: Phytoseiidae) is being used on a commercial
scale as a biological control agent of whiteflies and thrips in several greenhouse crops
(Nomikou et al., 2001; Messelink et al., 2006; Calvo et al., 2011). Amblyseius swirskii can
feed on various types of food including arthropod prey but also plant materials like pollen and
honeydew (Momen and El-Saway, 1993). Previous studies showed that A. swirskii can
successfully develop and reproduce on a wide range of food sources such as whiteflies
(Nomikou et al., 2001; Calvo et al., 2008; Messelink et al., 2008), thrips (Arthurs et al., 2009;
Chow et al., 2010), eriophyid mites (El-Laithy, 1998; Park et al., 2010), broad mites (van
Maanen et al., 2010; Onzo et al., 2012), spider mites (Xu and Enkegaard, 2010) and plant
pollen (Kutuk and Yigit, 2011; Park et al., 2011).
Depending on the species, phytoseiid mites are reared on phytophagous mite prey,
other natural foods such as pollen, or factitious prey like storage mites. In the commercial
production of A. swirskii, storage mites, including the dried fruit mite, Carpoglyphus lactis L.
(Acari: Carpoglyphidae), and Thyreophagus entomophagus (Laboulbene) (Acari: Acaridae),
are used as the primary food source (Bolckmans and van Houten, 2006; Fidgett and Stinson,
2008). The rearing procedures based on natural or factitious foods are often time-consuming
and/or expensive. The availability of an effective artificial diet for phytoseiids could eliminate
many of the problems associated with their mass rearing (Kennett and Hamai, 1980).
Whereas a good body of literature exists on artificial diets for insect predators
(Thompson, 1999; Riddick, 2009), relatively few studies have focused on the development of
artificial diets for phytoseiid mites. McMurtry and Scriven (1966) tested several artificial diets
for four phytoseiid species: Amblyseius limonicus Garman & McGregor, Amblyseius hibisci
(Chant), Typhlodromus occidentalis Nesbitt, and Typhlodromus rickeri Chant. Their results
showed that immature development was poor and oviposition rates were low compared to
natural prey or pollen. Shehata and Weismann (1972) reported that females of Phytoseiulus
persimilis (Athias-Henriot), deposited eggs when fed on three artificial diets, and larvae
developed to adulthood; however, the female offspring failed to produce eggs, and had a
smaller size and shorter longevity than conspecifics preying on T. urticae. Kennett and Hamai
(1980) found that seven out of nine species of phytoseiid mites could develop and reproduce
when fed an artificial diet composed of honey, sugar, yeast flakes, yeast hydrolysate,
enzymatic casein hydrolysate, and fresh egg yolk. However, the oviposition rate of all species
fed on artificial diet was only about one third of that achieved on natural diets. Abou-Awad et

A. swirskii artificial diet enriched A. pernyi pupal hemolymph
43
al. (1992) showed that Amblyseius gossipi El-Badry and A. swirskii developed, survived and
reproduced on several artificial diets consisting of yeast, milk, cystine, proline, arginine,
sucrose, glucose, streptomycin sulphate, and sorbic acid. However, females fed on the best
performing artificial diet had lower oviposition rates than those on natural diet. Ogawa and
Osakabe (2008) reported that an artificial diet composed of yeast components, saccharides,
and egg yolk supported development and survival of Neoseiulus californicus (McGregor), but
again fecundity on the artificial diet was low.
The above studies suggest that, although phytoseiid mites can develop on different
artificial diets, fecundity in most cases was negatively affected. Hence, for making a diet
suitable for commercial mass rearing of phytoseiids, the factor to be improved is reproduction.
Several studies have demonstrated that the use of insect materials in artificial diets can
enhance survival and oviposition of entomophagous insects (Grenier and De Clercq, 2003).
For instance, many artificial media for egg parasitoids of the genus Trichogramma contain
lepidopteran pupal hemolymph and/or pupal holotissues to stimulate egg laying by the
parasitoid females or to provide adequate nutrition for development of the larvae (e.g., Liu et
al. (1979); Strand and Vinson (1985); Nettles (1990); Lü et al. (2013)). Also, the fecundity of
the predatory anthocorid Orius insidiosus (Say) was significantly increased when the bugs
were fed on an artificial diet supplemented with a cell line derived from eggs of the Indian
meal moth Plodia interpunctella (Hübner) (Ferkovich and Shapiro, 2004). Conceivably,
insect materials like hemolymph could also be useful to supply nutritional factors for growth
and reproduction of predatory mites such as A. swirskii.
In the present study, we determined the life table parameters of A. swirskii when fed
on cattail pollen (Typha latifolia L.), dried fruit mite (C. lactis), or a meridic artificial diet
modified from Ogawa and Osakabe (2008), and a similar diet supplemented with hemolymph
of oak silkworm pupae (Antheraea pernyi Guérin-Méneville) (Lepidoptera: Saturniidae).
3.2 Materials and methods
3.2.1 Stock colony of Amblyseius swirskii
The stock colony of A. swirskii was initiated from specimens supplied by Biobest N.V.
(Westerlo, Belgium) and was cultured in a climatic cabinet set at 25 ± 1°C, 70 ± 5% RH and a
16:8 h (L:D) photoperiod. Mites were reared on green plastic arenas (10 x 10 x 0.3 cm)
(Multicel, SEDPA, France) on top of a thick foam pad placed in water in a plastic tray (20 x
13 x 5 cm). The edges of the arenas were covered with tissue paper immersed in the water to

Chapter 3
44
provide moisture and deter the mites from escaping (Figure 3.1). A small piece of sewing
thread was placed on the arenas to serve as an oviposition substrate. Every two days the eggs
were collected and transferred to new arenas. Cattail pollen (T. latifolia) was dusted every two
days as a food source for the mites.
Figure 3.1 Amblyseius swirskii colony rearing arena
3.2.2 Stock colony of Carpoglyphus lactis
A colony of C. lactis was initiated from mites supplied by Biobest N.V. and was kept in a
climatic cabinet at 25°C. Mites were reared in insect breeding dishes (10 x 4 cm) (SPL Life
Sciences Co. Ltd., Korea) with a mesh hole (4 cm in diameter) in the lid (Figure 3.2); the
dishes were placed in a foam box (30 x 20 x 20 cm) with 2 cm of water. The diet of C. lactis
was modified from Zdarkova et al. (1999), and consisted of 1/2 wheat germ (Dr. Grandel,
Augsburg, Germany), 1/3 yeast torula (MP Biomedicals LLC, Illkirch, France), and 1/6 Pond
Food Balance Sticks (Vitakraft, Bremen, Gemany); the diet was mixed with vermiculite to
provide food and shelter for the mites.
Figure 3.2 Carpoglyphus lactis colony rearing box

A. swirskii artificial diet enriched A. pernyi pupal hemolymph
45
3.2.3 Pollen
Fresh cattail pollen (T. latifolia) was also supplied by Biobest N.V. and stored at -18°C. For
the experiments, the pollen was thawed and kept in a refrigerator at 5°C for max. 1 week.
3.2.4 Preparation of artificial diet
The artificial diet was composed of honey (Meli N.V., Veurne, Belgium), sucrose (MP
Biomedicals LLC, Illkirch, France), tryptone (Fluka Analytical, Sigma-Aldrich Co., St. Louis,
USA), yeast extract (Duchefa, Haarlem, The Netherlands), fresh hen's egg yolk, and distilled
water, and was prepared according to Ogawa and Osakabe (2008), with some modifications as
laid out below. Two artificial diets (AD) were prepared.
The first diet (AD1) consisted of 5% honey, 5% sucrose, 5% tryptone, 5% yeast
extract, 10% egg yolk, and 70% distilled water (w/w). Honey, sucrose, and tryptone were
dissolved into the distilled water, after which the yeast extract and egg yolk were added.
Unlike Ogawa and Osakabe (2008), we did not filter sterilize any of the ingredients. All
ingredients were then blended using a Virtis mixer (SP Industries Inc., Gardiner, New York,
USA).
The second diet (AD2) consisted of 80% AD1 supplemented with 20% (w/w)
hemolymph of oak silkworm pupae (A. pernyi) originating from a culture at the Guangdong
Entomological Institute, China. The pupal hemolymph was collected from live A. pernyi
pupae which were immersed in a water bath at 60°C for 10 minutes to avoid melanization of
the hemolymph. After surface sterilization of the pupae with 75% ethanol, the hemolymph
was collected by pressing the pupae under sterile conditions. The hemolymph was then
lyophilized and stored in a deep freezer at -18°C. The lyophilized hemolymph was
redissolved using distilled water before being added to the diet.
For both AD1 and AD2, fresh diet was prepared every week and kept in a refrigerator
at 5°C.
3.2.5 Rearing microcosms
To examine the development and reproduction of individual A. swirskii, modified Munger
cells were used as rearing microcosms (Ogawa and Osakabe, 2008). Each cell consisted of a
transparent acrylic board (top board; 40 x 40 mm, 2 mm thick) with a 19 mm diameter hole in
the center, a black acrylic board (middle board; 40 x 40 mm, 5 mm thick) with a 18 mm

Chapter 3
46
diameter hole in the center, and another black acrylic board (bottom board; 40 x 40 mm, 2
mm thick) with a 1 mm diameter hole in the center. Clear transparency film (3M™ Dual-
Purpose Transparency Film) was placed between the top and middle boards and was pierced 4
times with a fine needle allowing ventilation but precluding escape of the mites. The hole in
the bottom board was plugged with a rolled piece of tissue paper saturated with tap water to
serve as a water source for the mites (Figure 3.3). A paper clip was used to hold the boards
together. The microcosms were placed on a plastic support containing tap water.
Figure 3.3 Rearing microcosm
3.2.6 Experimental setup
Eight hours before the start of the experiments, new black threads were placed in the stock
colony of A. swirskii. Eggs deposited on the threads were transferred individually to the
rearing microcosms; 48, 54, 55, and 53 eggs were used for the treatments with T. latifolia
pollen, C. lactis, AD1, and AD2, respectively. All diets were offered from the larval stage of
the predator on and refreshed every 2 days. In the treatment with C. lactis, predators were
supplied with a mixture of life stages of the storage mite. Artificial diets were absorbed on a
small piece of filter paper (2 x 2 mm) which was placed on the bottom board of the cells. To
obtain data on the duration of each developmental stage of A. swirskii and on mortality and
escape rates, observations were made every 24 hours until all individuals had reached
adulthood. The developmental stage of each individual was determined based on the presence
of exuviae in the cells. After completing immature development, each female was paired with
a male that was reared on the same diet as the female. Males that died during the experiment
were replaced. Adults were observed daily to determine the preoviposition and oviposition
period, longevity and fecundity. Progeny from females of the same age were transferred to
new cells and fed on the same diet as their parents in order to determine the sex ratio of the

A. swirskii artificial diet enriched A. pernyi pupal hemolymph
47
offspring for each treatment. The experiments were done in a growth chamber at 23 ± 1°C, 65
± 5% RH and a 16:8 h (L:D) photoperiod.
3.2.7 Life table parameters calculation
The intrinsic rate of increase (rm) was calculated according to the formula of Lotka (1907) and
Birch (1948):
∑𝑙𝑥𝑚𝑥𝑒−𝑟𝑚∗𝑥 = 1
where x equals the female age (days), lx is the age specific survival of the females at age x and
mx is the number of daughters produced per female at age x. The latter parameter is obtained
by multiplying the mean number of eggs laid per female by the proportion of female offspring
produced at age x. The Jackknife procedure was used according to Meyer et al. (1986) and
Hulting et al. (1990) to calculate the standard error of rm. Other parameters calculated (Maia
et al., 2000) were the generation time T, i.e. mean time span between the birth of individuals
of a generation and that of the next generation (days),
T =∑xlxmx
∑ lxmx
and the net reproductive rate, R0, i.e. the mean number of female offspring produced per
female (females/female)
R0 =∑lxmx
3.2.8 Statistical analysis
Data were subjected to statistical analysis (IBM SPSS Statistics, Ver. 20) to analyze the effect
of diet on the duration of the immature stages, preoviposition and oviposition period, daily
and total oviposition, and adult longevity of A. swirskii. When a Kolmogorov–Smirnov test
indicated that means were normally distributed, the parameter was analyzed using a one-way
analysis of variance (ANOVA). If a Levene test indicated heteroscedasticity, a Tamhane test
was used instead of Tukey's test. When means were not normally distributed, a nonparametric
Kruskal-Wallis ANOVA was used and means were separated using a Mann-Whitney U test.
Immature survival and sex ratios of the progeny were compared by means of a logistic

Chapter 3
48
regression. This regression is a generalized linear model using a probit (log odds) link and a
binomial error function. Each test consists of a regression coefficient that is calculated and
tested for being significantly different from zero, for which P-values are presented
(McCullagh and Nelder, 1989). P-values smaller than or equal to 0.05 are considered
significant. Fertility life table parameters, including the net reproductive rate (Ro), generation
time (T), and intrinsic rate of increase (rm) were estimated using the Jackknife procedure as
described by Maia et al. (2000).
3.3 Results
The developmental times of all male immature stages and egg, larval, and protonymphal
stages of females did not differ among diets (Table 3.1). However, the developmental time of
female deutonymphs was significantly affected by diet. Female deutonymphs reared on C.
lactis or AD2 took significantly less time to develop to adulthood than those fed on the other
diets. The total developmental time of females fed on C. lactis or AD2 was significantly
shorter than that of females fed on AD1. Nearly all individuals reached adulthood on the
different diets, with immature survival rates of 100, 100, 96.36, and 98.11% when the
predators were fed on T. latifolia pollen, C. lactis, AD1, and AD2, respectively. Survival rate
was not affected by diet (χ2 = 4.52; df = 3; P = 0.210).
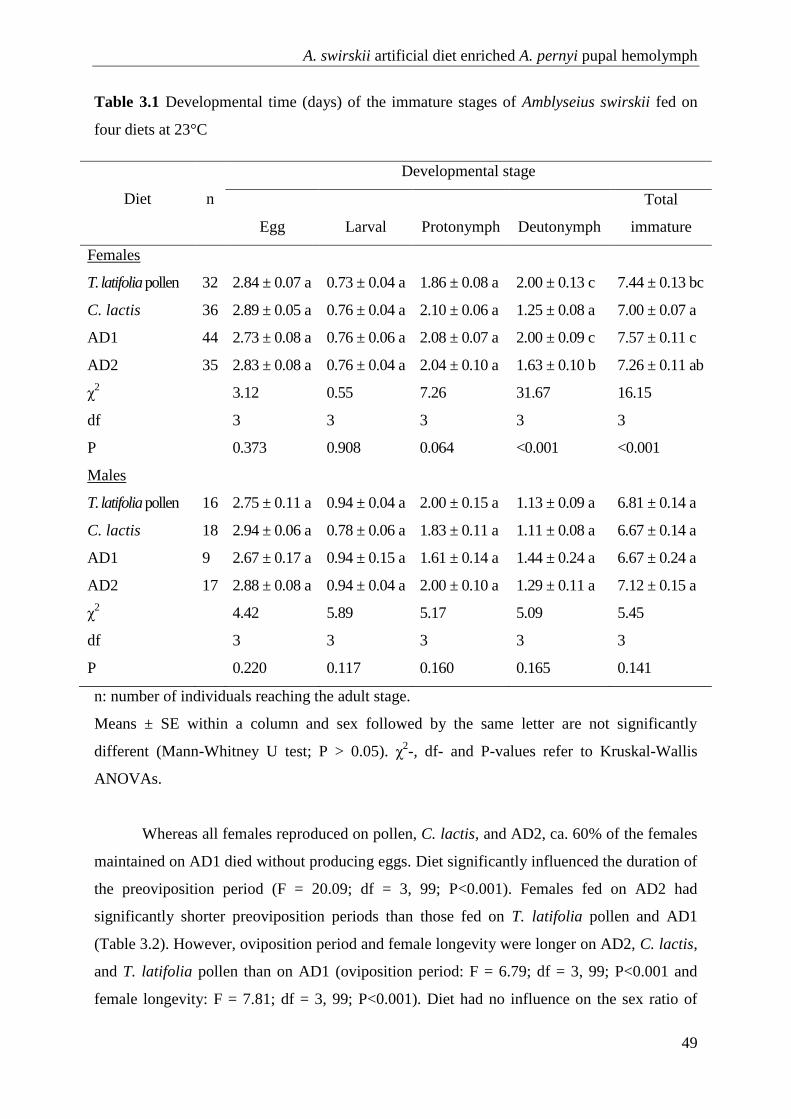
A. swirskii artificial diet enriched A. pernyi pupal hemolymph
49
Table 3.1 Developmental time (days) of the immature stages of Amblyseius swirskii fed on
four diets at 23°C
Diet n
Developmental stage
Egg Larval Protonymph Deutonymph
Total
immature
Females
T. latifolia pollen 32 2.84 ± 0.07 a 0.73 ± 0.04 a 1.86 ± 0.08 a 2.00 ± 0.13 c 7.44 ± 0.13 bc
C. lactis 36 2.89 ± 0.05 a 0.76 ± 0.04 a 2.10 ± 0.06 a 1.25 ± 0.08 a 7.00 ± 0.07 a
AD1 44 2.73 ± 0.08 a 0.76 ± 0.06 a 2.08 ± 0.07 a 2.00 ± 0.09 c 7.57 ± 0.11 c
AD2 35 2.83 ± 0.08 a 0.76 ± 0.04 a 2.04 ± 0.10 a 1.63 ± 0.10 b 7.26 ± 0.11 ab
χ2 3.12 0.55 7.26 31.67 16.15
df 3 3 3 3 3
P 0.373 0.908 0.064 <0.001 <0.001
Males
T. latifolia pollen 16 2.75 ± 0.11 a 0.94 ± 0.04 a 2.00 ± 0.15 a 1.13 ± 0.09 a 6.81 ± 0.14 a
C. lactis 18 2.94 ± 0.06 a 0.78 ± 0.06 a 1.83 ± 0.11 a 1.11 ± 0.08 a 6.67 ± 0.14 a
AD1 9 2.67 ± 0.17 a 0.94 ± 0.15 a 1.61 ± 0.14 a 1.44 ± 0.24 a 6.67 ± 0.24 a
AD2 17 2.88 ± 0.08 a 0.94 ± 0.04 a 2.00 ± 0.10 a 1.29 ± 0.11 a 7.12 ± 0.15 a
χ2 4.42 5.89 5.17 5.09 5.45
df 3 3 3 3 3
P 0.220 0.117 0.160 0.165 0.141
n: number of individuals reaching the adult stage.
Means ± SE within a column and sex followed by the same letter are not significantly
different (Mann-Whitney U test; P > 0.05). χ2-, df- and P-values refer to Kruskal-Wallis
ANOVAs.
Whereas all females reproduced on pollen, C. lactis, and AD2, ca. 60% of the females
maintained on AD1 died without producing eggs. Diet significantly influenced the duration of
the preoviposition period (F = 20.09; df = 3, 99; P<0.001). Females fed on AD2 had
significantly shorter preoviposition periods than those fed on T. latifolia pollen and AD1
(Table 3.2). However, oviposition period and female longevity were longer on AD2, C. lactis,
and T. latifolia pollen than on AD1 (oviposition period: F = 6.79; df = 3, 99; P<0.001 and
female longevity: F = 7.81; df = 3, 99; P<0.001). Diet had no influence on the sex ratio of

Chapter 3
50
offspring with the proportion of females ranging from 0.68 to 0.72 (χ2 = 2.52; df = 3, P =
0.472). The daily oviposition rate of A. swirskii reared on AD2 was significantly higher than
that of females reared on the other diets (F = 29.17; df = 3, 99; P<0.001).
Table 3.2 Reproduction and life table parameters of Amblyseius swirskii fed on four diets at
23°C
Parameter
T. latifolia
pollen
(n = 29)
C. lactis
(n = 31)
AD1
(n = 16)
AD2
(n = 27)
Preoviposition period
(days)b
3.14 ± 0.21 b 2.77 ± 0.13 ab 4.81 ± 0.44 c 2.41 ± 0.10 a
Oviposition period (days)a 25.48 ± 1.78 a 24.68 ± 1.65 a 12.81 ± 3.02 b 25.67 ± 2.05 a
Female longevity (days)a 34.45 ± 2.38 a 35.90 ± 2.05 a 22.56 ± 3.52 b 42.07 ± 2.63 a
Female proportion of the
progenyc
0.68 ± 0.007 a 0.72 ± 0.004 a 0.70 ± 0.029 a 0.71 ± 0.011 a
Oviposition rate
(eggs/female/day)b
1.18 ± 0.04 b 1.21 ± 0.03 b 0.92 ± 0.08 c 1.52 ± 0.03 a
Total number of eggs
(eggs/female)a
29.00 ± 1.72 b 29.03 ± 1.72 b 9.94 ± 2.26 d 38.26 ± 2.89 a
Net reproductive rate Ro
(females /female)b
19.71 ± 0.042 c 20.58 ± 0.04 b 6.46 ± 0.098 d 27.21 ± 0.069 a
Generation time T (days)b 19.00 ± 0.29 ab 17.33 ± 0.18 b 22.08 ± 2.50 a 18.30 ± 0.23 b
Intrinsic rate of increase rm
(females /female/day)b
0.158 ± 0.002 b 0.175 ± 0.002 a 0.104 ± 0.013 c 0.181 ± 0.002 a
n = number of reproducing females observed
Means ± SE within a row followed by the same letter are not significantly different (P>0.05;
aTukey or
bTamhane test;
cProbit (Wald Chi-square) test)
Peak oviposition was reached after 3 days on AD2, after 7 days on C. lactis and after 8
days on T. latifolia and AD1 (Figure 3.4). Also, the total number of eggs was significantly
higher for females offered AD2 versus the other diets (F = 21.78; df = 3, 99; P<0.001) (Table
3.2).

A. swirskii artificial diet enriched A. pernyi pupal hemolymph
51
Figure 3.4 Daily oviposition rate of Amblyseius swirskii fed on four diets at 23°C
Differences in developmental and reproductive characteristics were reflected in life
table statistics. Net reproductive rate (Ro) (F = 16813.98; df = 3, 99; P<0.001) of A. swirskii
fed on AD2 was significantly higher than that of predators fed on C. lactis, T. latifolia pollen,
or AD1. Mean generation time (F = 5.01; df = 3, 99; P<0.001) of predators reared on AD1
was significantly longer than that of conspecifics reared on AD2 and C. lactis. Finally, the
intrinsic rates of increase (rm) of the predators fed on AD2 and C. lactis were greater than on
the other diets (F = 45.72; df = 3, 99; P<0.001) (Table 3.2).
3.4 Discussion
The main objective of our study was to develop a suitable artificial diet for mass rearing of the
economically important phytoseiid predator A. swirskii. The artificial diets that we tested were
modified from that formulated by Ogawa and Osakabe (2008) for N. californicus and
compared with cattail pollen, a standard food in several predatory mite studies (Lee and
Gillespie, 2011; Park et al., 2011), and the dried fruit mite, C. lactis, which is routinely used
in the commercial production of A. swirskii (Bolckmans and van Houten, 2006). Our findings
indicate that A. swirskii can develop and reproduce with similar or even better success on an
artificial diet as compared to pollen or storage mites. Our results with cattail pollen and C.

Chapter 3
52
lactis are consistent with previous reports (Nomikou et al., 2003b; Bolckmans and van Houten,
2006; Fidgett and Stinson, 2008; Lee and Gillespie, 2011).
A more favorable diet will result in higher population growth of an arthropod, as
indicated by superior life table parameters (Grenier and De Clercq, 2003). The highest
intrinsic rates of increase in our study were observed when A. swirskii was fed with AD2 or C.
lactis (0.182 and 0.175 females/female/day, respectively). These figures were higher than
those reported for A. swirskii when fed on the twospotted spider mite Tetranychus urticae
Koch at 26°C (0.167) (El-Laithy and Fouly, 1992), the western flower thrips Frankliniella
occidentalis (Pergande) and onion thrips Thrips tabaci Lindeman at 25°C (0.056 and 0.024,
respectively) (Wimmer et al., 2008), and the eriophyoid fig mites Aceria ficus (Cotte) and
Rhyncaphytoptus ficifoliae Keifer at 29°C (0.155 and 0.122, respectively) (Abou-Awad et al.,
1999). On the other hand, the rm values of A. swirskii on AD2 or C. lactis calculated here were
slightly lower than the value of 0.201 reported by Park et al. (2011) when the predator was
offered the tomato russet mite Aculops lycopersici (Massee) as prey at 25°C, and when fed on
the cotton whitefly Bemisia tabaci Gennadius at 27°C (0.213) (Nomikou et al., 2001). Thus,
on both the factitious prey C. lactis and on the enriched artificial diet AD2, A. swirskii
performed well and in some cases even better than on several of its natural prey. However, it
should be noted that comparison of life table statistics among studies is complicated by
differences in experimental methods, climatic conditions and calculation of estimates, which
may explain some of the contrasting results in the literature.
The artificial diet developed by Ogawa and Osakabe (2008) supported immature
development and survival of the predatory mite N. californicus but its oviposition rate on this
diet was negligible compared with a diet of T. urticae. However, a slightly modified version
of this artificial diet used in our study (AD1, without insect hemolymph) proved suitable to
support development of A. swirskii and also allowed some reproduction. The oviposition rate
of A. swirskii on AD1 (0.92 eggs/female/day) was higher than that on an artificial diet
designed by Abou-Awad et al. (1992) (0.30 eggs/female/day); moreover, it was similar to that
of mites reared on the natural prey F. occidentalis (Wimmer et al., 2008) (0.92
eggs/female/day) and superior to that of females fed on the carmine spider mite Tetranychus
cinnabarinus (Boisduval) and brown soft scale Coccus hesperidum (L.) (0.31 and 0.02
eggs/female/day, respectively) (Ragusa and Swirski, 1977). The relatively good performance
of A. swirskii on artificial diet AD1 may be due to the generalist feeding habits of this
phytoseiid mite (Momen and El-Saway, 1993). However, this diet has still limitations in terms

A. swirskii artificial diet enriched A. pernyi pupal hemolymph
53
of fecundity, with 60% of the females failing to lay eggs and dying shortly after molting to the
adult stage.
Nettles (1990) and Grenier and De Clercq (2003) noted that adding insect components
such as hemolymph to artificial diets enhanced their acceptability and improved their
nutritional quality for a number of entomophagous insects. Hemolymph of lepidopteran larvae
or pupae has been used in artificial media for parasitoid wasps, especially for Trichogramma
species. For instance, Trichogramma pretiosum Westwood successfully completed
development in artificial medium when it was supplemented with hemolymph of Manduca
sexta (L.) (Strand and Vinson, 1985; Xie et al., 1986). Cônsoli and Parra (1997) reported that
Trichogramma galloi Zucchi and T. pretiosum could be reared in a medium consisting of 70%
Helicoverpa zea (Boddie) larval hemolymph. Pupal hemolymph of A. pernyi or Philosamia
cynthia ricini Drury was used in an artificial medium for Trichogramma dendrolimi
(Matsumura) (Liu et al., 1979) and Anastatus sp. egg parasitoids (Liu et al., 1988). Lü et al.
(2013) also indicated that insect hemolymph is a key component of artificial media for
Trichogramma spp. Our study indicates, however, that supplementing artificial diets with
insect hemolymph may also be useful to improve their nutritional value for predatory
arthropods. When 20% pupal hemolymph of A. pernyi was added to diet AD1 to create AD2
in the present study, oviposition rate and intrinsic rate of increase of A. swirskii were
substantially increased. These results suggest that the hemolymph played a positive role both
in fecundity and survival of the predator. Further research is needed to identify the
components of the pupal hemolymph which are responsible for the increased performance.
In conclusion, the artificial diet AD2 supported development and reproduction of A.
swirskii to the same extent as a factitious prey which is routinely used in the mass rearing of
the phytoseiid, indicating the potential of artificial diets to rationalize the mass production of
this economically important biological control agent. However, nutritional imbalances within
the diet is possibly expressed only in the subsequent generations (De Clercq et al., 2005a).
Therefore, developmental and reproductive performance of A. swirskii fed on AD2 will need
to be assessed over subsequent generations. Furthermore, whereas A. pernyi is currently
widely used in silk production in China, pupal hemolymph of this insect may become less
available in the future, which may result in a higher price for this diet component and thus in a
higher production cost of the artificial diet (Lü et al., 2013). Thus, future work is warranted to
reduce the percentage of pupal hemolymph in the diet or to replace it by a more easily
available nutrient.

Chapter 3
54

55
Chapter 4
BENEFICIAL EFFECT OF SUPPLEMENTING AN ARTIFICIAL DIET FOR
AMBLYSEIUS SWIRSKII WITH HERMETIA ILLUCENS HEMOLYMPH
This chapter is based on:
Nguyen, D. T., Bouguet, V., Spranghers, T., Vangansbeke, D., & De Clercq, P. 2014. Beneficial
effect of supplementing an artificial diet for Amblyseius swirskii with Hermetia illucens hemolymph.
Journal of Applied Entomology, DOI: 10.1111/jen.12188.

Chapter 4
56
4.1 Introduction
Amblyseius swirskii (Athias-Henriot) (Acari: Phytoseiidae) is a generalist predatory mite that
has been commercialized since 2005 and is presently being used on a worldwide scale as an
augmentative biological control agent against thrips and whiteflies in a range of greenhouse
crops (Nomikou et al., 2002; Messelink et al., 2006; Messelink et al., 2008). The predator can
develop and reproduce on variety of other food sources including spider mites (El-Laithy and
Fouly, 1992), eriophyid mites (Abou-Awad et al., 1999; Park et al., 2011), broad mites
(Swirski et al., 1967; van Maanen et al., 2010) and pollen (Swirski et al., 1967; Ragusa and
Swirski, 1977; Park et al., 2011).
In an augmentative biocontrol strategy, large quantities of beneficial insects and mites
are produced and released in the crop. A conservative method for mass-rearing these natural
enemies is via a so-called natural, tri-trophic system, comprising the predator (parasitoid), the
herbivorous prey (host), and the prey's host plant (De Clercq et al., 2014). In order to
rationalize the production line the number of trophic levels can be reduced, allowing to lower
costs for labour and production facilities, like greenhouses (Grenier, 2009). One way to do
this can be by using of factitious foods. In the mass production of various generalist
phytoseiids, like A. swirskii, storage mites, such as Carpoglyphus lactis L. (Acari:
Carpoglyphidae) and Thyreophagus entomophagus (Laboulbene) (Acari: Acaridae), are being
used as primary food sources instead of natural prey (Bolckmans and van Houten, 2006).
However, the rearing procedures of phytoseiid mites based on these factitious foods are often
time-consuming and allergy problems can be generated by using storage mites (Fernandez-
Caldas et al., 2007). The availability of an effective artificial diet may tackle these problems,
shorten the production line and consequently represent a step towards a more cost-effective
mass rearing (Riddick, 2009).
In Chapter 3, we found that an artificial diet enriched with pupal hemolymph of the
Chinese oak silkworm, Antheraea pernyi (Guérin-Méneville) (Lepidoptera: Saturniidae)
supported development and reproduction of A. swirskii to the same extent as the routinely
used storage mite Carpoglyphus lactis (L.). However, the silkworm hemolymph is not easily
available resulting in a higher cost and lower reliability for the continuous production of the
artificial diet (Xie et al., 1997; Heslin et al., 2005; Lü et al., 2013; Lü et al., 2014). Therefore,
if the silkworm hemolymph extract could be replaced by a more easily available and cheaper
nutrient, the cost of diet production may be further reduced.

Effect of H. illucens hemolymph in artificial diet of A. swirskii
57
The black soldier fly, Hermetia illucens (L.) (Diptera: Stratiomyidae) can feed on a
wide variety of decomposing vegetal and animal matter (James, 1935). This species is
particularly interesting because the larvae can use a wide array of waste materials, converting
them into proteins and fats that are suitable for consumption by various animals, including
poultry (Hale, 1973), swine (Newton et al., 1977), and fish (Bondari and Sheppard, 1987).
Furthermore, in organic waste management soldier fly larvae can assist in suppressing
populations of houseflies, Musca domestica L. (Diptera: Muscidae) (Furman et al., 1959), and
help to reduce infectious bacterial populations in chicken and cow manure (Erickson et al.,
2004; Liu et al., 2008). The larvae can also convert organic waste into biodiesel fuel (Li et al.,
2011a). Prepupal black soldier flies contain on average 44% dry matter, which is composed of
42% protein and 35% fat, including essential amino and fatty acids (Hale, 1973).
The hypothesis tested in this study was that the high amount of proteins and other
nutrients extracted from black soldier fly prepupae could improve the nutritional quality of an
artificial diet for the development and reproduction of the predatory mite A. swirskii. Life
table parameters of A. swirskii were determined when the predator was fed on a basic meridic
artificial diet (section 3.2.4 ) supplemented with different concentrations (0, 5, 10 and 20%) of
prepupal hemolymph of H. illucens.
4.2 Materials and methods
4.2.1 Stock colony of Amblyseius swirskii
A stock colony of A. swirskii was reared as described in section 3.2.1
4.2.2 Black soldier fly rearing and hemolymph collecting
A colony of black soldier flies, H. illucens, was initiated with specimens supplied by
Millibeter BVBA (Antwerp, Belgium). The colony was maintained in a greenhouse at the
faculty of Bioscience Engineering, Ghent University, at 25 ± 5 °C and 40 – 80 % RH. Newly
hatched larvae were placed into stainless steel containers (34 x 25 x 20 cm) with vented lids.
Larvae were reared on solid organic waste springing from a fruit and vegetable fermentation
installation. Fresh substrate was added weekly until the larvae reached the prepupal stage and
migrated out of the substrate.
Hemolymph was collected from H. illucens prepupae (ca. 5 days old), which had been
washed with tap water. Prepupae were immersed in a water bath at 60 oC for 10 min to avoid

Chapter 4
58
melanisation of the hemolymph. After surface sterilization of the prepupae with 75 % ethanol,
the hemolymph was collected by cutting the end of the prepupal abdomen and pressing the
prepupae under sterile conditions. In order to obtain 1 mL of hemolymph, about 20-25
prepupae were processed. The hemolymph was then stored in a deep freeze at -18 oC.
4.2.3 Preparation of artificial diets
The basic diet (AD0) was prepared as described in section 3.2.4 . The enriched diets AD5,
AD10 and AD20 consisted of 95%, 90% or 80% of AD0 supplemented with 5, 10 or 20%
(w/w) prepupal hemolymph of the black soldier fly, respectively. The ingredients were
blended using a Virtis mixer (SP Industries Inc., Gardiner, New York, USA). The diets were
stored in a deep freeze at -18°C and thawed before use. Thawed diets were kept in a
refrigerator at 5 °C for max. 1 week.
4.2.4 Development and reproduction on the different artificial diets
Sixty eggs (< 8 h old) of A. swirskii per treatment were transferred individually from the stock
colony to rearing microcosms that were modified from Munger cells as described in section
3.2.5 . The foods were offered ad libitum from the larval stage of the predator on, and
refreshed every 2 days. Artificial diets were absorbed on a small piece of filter paper (2 x 2
mm) which was placed on the bottom board of the cells. The mites were observed and
different parameters were recorded same as that described in section 3.2.6 . Mites that escaped
or died as a result of manipulation were excluded from data analysis; these escape and death
rates did not differ among treatments and varied between 3 and 18 % (χ2
= 5.353, df = 3, P =
0.148; Probit (Wald Chi-square) test). The experiments were done in a growth chamber at 23
± 1 °C, 65 ± 5 % RH and a 16:8 h (L:D) photoperiod.
4.2.5 Diet switching experiment
In the first experiment, most females fed on the basic diet (AD0) were not able to produce
viable eggs; therefore, a diet switching experiment was conducted to investigate the role of
prepupal hemolymph of the black soldier fly in stimulating egg laying by A. swirskii when its

Effect of H. illucens hemolymph in artificial diet of A. swirskii
59
immature stages were reared on the basic diet.Eggs (<8h old) were collected from the colony
and transferred to green plastic arenas (5 x 5 x 0.3 cm) (Multicel, SEDPA, France), placed on
a wet sponge in a vented insect breeding dish (10 x 4 cm) (SPL Life Sciences Co. Ltd., Korea)
containing water. The edges of the arenas were covered with tissue paper immersed in the
water to provide moisture and deter the mites from escaping. Every two days the juvenile
mites were offered fresh basic artificial diet AD0. When the mites reached adulthood, males
and females were paired and each pair was transferred to an individual Munger cell. Four
cohorts were presented with one of the different artificial diets (AD0, AD5, AD10 or AD20),
as described above. Adults were observed daily until the females stopped laying eggs in order
to determine the oviposition period and fecundity.
4.2.6 Life table parameters calculation
Life table parameters were calculated as explained in section 3.2.7
4.2.7 Statistical analysis
Data were subjected to statistical analysis (IBM SPSS Statistics, Ver. 21) to evaluate the
effect of diet on the developmental time, preoviposition and oviposition period, daily and total
oviposition, and adult longevity of A. swirskii. When a Kolmogorov–Smirnov test indicated
that data were normally distributed, they were analysed using a one-way analysis of variance
(ANOVA). If a Levene test indicated heteroscedasticity, a Tamhane post-hoc test was used
instead of a Tukey test. When means were not normally distributed, a nonparametric Kruskal-
Wallis ANOVA was used and means were separated using Mann-Whitney U tests. Immature
survival rates and sex ratios were compared by means of a logistic regression. This regression
is a generalized linear model using a probit (log odds) link and a binomial error function.
Each test consists of a regression coefficient that is calculated and tested for being
significantly different from zero, for which P-values are presented (McCullagh and Nelder,
1989). In all tests, P-values smaller than or equal to 0.05 were considered significant.
4.3 Results
Immature survival rates were not significantly different among diets enriched with
hemolymph of H. illucens prepupae, but these values were significantly higher than those of
predatory mites fed on the basic diet AD0 (χ2 = 12.882, df=3, P=0.005; Probit (Wald Chi-

Chapter 4
60
square) test). The survival rates were 82%, 100%, 98% and 98% when the predators were fed
on AD0, AD5, AD10 and AD20, respectively.
Table 4.1 Developmental times of immature stages of Amblyseius swirskii fed on artificial
diets enriched or not with Hermetia illucens prepupal hemolymph at 23°C
Diet n
Developmental time (days)
Larval Protonymph Deutonymph
Total immature
(Larva-adult)
Females
AD0 38 0.80 ± 0.04 a 2.14 ± 0.07 c 1.95 ± 0.07 b 4.89 ± 0.10 c
AD5 42 0.71 ± 0.04 a 1.98 ± 0.08 c 1.43 ± 0.09 a 4.12 ± 0.05 b
AD10 40 0.78 ± 0.04 a 1.68 ± 0.07 b 1.65 ± 0.08 a 4.10 ± 0.09 ab
AD20 41 0.72 ± 0.04 a 1.33 ± 0.04 a 1.85 ± 0.06 b 3.90 ± 0.05 a
χ2 3.523 57.585 22.334 69.217
df 3 3 3 3
P 0.318 <0.001 <0.001 <0.001
Males
AD0 11 0.95 ± 0.05 ab 1.95 ± 0.11 c 1.18 ± 0.18 a 4.09 ± 0.16 b
AD5 16 0.81 ± 0.06 a 1.88 ± 0.12 bc 1.00 ± 0.13 a 3.69 ± 0.12 ab
AD10 19 1.00 ± 0.00 b 1.53 ± 0.14 ab 1.32 ± 0.15 a 3.84 ± 0.16 ab
AD20 17 0.94 ± 0.04 ab 1.29 ± 0.11 a 1.24 ± 0.14 a 3.47 ± 0.12 a
χ2 10.372 15.244 2.866 8.405
df 3 3 3 3
P 0.016 0.002 0.413 0.038
n: number of individuals reaching the adult stage; AD0, AD5, AD10 and AD20: basic
artificial diet enriched with 0%, 5%, 10% and 20% of H. illucens prepupae hemolymph,
respectively. Means ± SE within a column and sex followed by the same letter are not
significantly different (Mann-Whitney U test; P > 0.05). χ2-, df- and P-values refer to Kruskal-
Wallis ANOVAs.
The developmental time of the female larvae did not differ among diets (Table 4.1).
However, diet had a significant effect on the developmental rate of the nymphal stages of the
females. Female protonymphs fed on AD20 demonstrated the fastest development, followed
by females fed on AD10 and females developed slowest in this stage on AD5 and AD0.

Effect of H. illucens hemolymph in artificial diet of A. swirskii
61
Female deutonymphs reared on AD5 or AD10 needed significantly less time to develop to
adulthood than those fed on the other diets. The total developmental time was significantly
shorter for females offered any of the enriched artificial diets versus the basic diet.
Deutonymphal times of males were not affected by diet. Whereas male larvae developed
slower on AD10 than on AD5, male protonymphs fed on AD20 and AD10 had shorter
developmental times than those fed on AD0. Male total developmental time was significantly
shorter on AD20 than on AD0.
Table 4.2 Reproduction and life table parameters of Amblyseius swirskii fed on artificial diets
enriched or not with Hermetia illucens prepupal hemolymph at 23°C
Parameter AD0
(n = 37)
AD5
(n = 36)
AD10
(n = 33)
AD20
(n = 38)
Preoviposition period (days)a 6.76 ± 0.24 c 2.44 ± 0.10 b 2.33 ± 0.11 ab 2.18 ± 0.06 a
Oviposition period (days)a 0.24 ± 0.17 b 22.33 ± 1.74 a 20.30 ± 1.30 a 22.08 ± 1.46 a
Female longevity (days)a 7.95 ± 0.90 b 33.08 ± 2.94 a 35.24 ± 3.24 a 31.03 ± 2.39 a
Female proportion of the
progenyc
- 0.68 ± 0.03 a 0.66 ± 0.03 a 0.71 ± 0.02 a
Oviposition rate
(eggs/female/day)b
0.10 ± 0.05 b 1.18 ± 0.03 a 1.19 ± 0.04 a 1.32 ± 0.05 a
Total fecundity (eggs/female)a 0.19 ± 0.12 c 25.47 ± 1.89 ab 23.70 ± 1.47 b 27.34 ± 1.52 a
Net reproductive rate R0
(females/female)b
- 14.94 ± 0.78 b 14.58 ± 0.62 b 17.60 ± 0.73 a
Generation time T (days)b - 14.84 ± 0.19 b 14.17 ± 0.20 a 13.65 ± 0.15 a
Intrinsic rate of increase rm
(females/female/day)a
- 0.182 ± 0.002 c 0.189 ± 0.002 b 0.210 ± 0.002 a
n = number of reproducing females observed
AD0, AD5, AD10 and AD20: basic artificial diet enriched with 0%, 5%, 10% and 20% of H.
illucens prepupal hemolymph, respectively.
Means ± SE within a row followed by the same letter are not significantly different (P>0.05;
aMann-Whitney U test,
bTukey test;
cProbit (Wald Chi-square) test)
The preoviposition period, oviposition period and longevity of females fed on the diets
enriched with hemolymph were significantly shortened as compared with females fed on AD0

Chapter 4
62
(Table 4.2) (preoviposition period: χ2
= 91.542; df = 3; P < 0.001; oviposition period: χ2
=
83.409; df = 3; P < 0.001 and female longevity: χ2
= 75.612; df = 3; P < 0.001). Similarly, the
oviposition rate and total number of eggs of females reared on AD0 were far lower than those
of females presented with AD5, AD10 or AD20 (oviposition rate: F = 176.613; df = 3, 140; P
< 0.001 and total number of eggs: χ2
= 75.612; df = 3; P < 0.001). Females maintained on
AD0 laid on average only 0.19 eggs during their lifetime as compared with 23.7-27.3 eggs for
those fed enriched diets; none of the eggs in the AD0 group produced viable offspring.
Consequently, life table parameters could not be calculated for the AD0 cohort. Females fed
on AD20 had significantly shorter preoviposition periods than those fed on AD5. However,
the percentage of hemolymph supplemented in the artificial diet had no influence on
oviposition period, oviposition rate and longevity of the females and on the female proportion
of their offspring (χ2 = 2.868; df = 2, P = 0.238).
Figure 4.1 Daily oviposition rate of Amblyseius swirskii fed on artificial diets enriched or not
with Hermetia illucens prepupal hemolymph at 23°C. AD0, AD5, AD10 and AD20: basic
artificial diet enriched with 0%, 5%, 10% and 20% of H. illucens prepupal hemolymph,
respectively.
Peak oviposition was reached on the 4th
day of adulthood on AD0 and AD20, on the
5th
day on AD5 and on the 8th
day on AD10. Whereas egg laying by females on AD0 was

Effect of H. illucens hemolymph in artificial diet of A. swirskii
63
almost completely terminated after 10 days of adulthood, females fed on AD5, AD10 and
AD20 continued to lay eggs until about 46 days of adulthood (Figure 4.1). The total egg
production of A. swirskii was significantly higher on AD20 than on AD10.
Life table parameters of female A. swirskii on artificial diets enriched with different
percentages of hemolymph are shown in Table 4.2. The net reproductive rate (R0) was highest
on AD20 (F = 5.339; df = 2, 104; P = 0.006), while generation times (T) were similar on
AD10 and AD20 but higher than on AD5 (F = 11.081; df = 2, 104; P < 0.001). The intrinsic
rate of increase rm was highest on AD20, followed by AD10 and lowest on AD5 (χ2
= 61.310;
df = 2; P < 0.001).
Table 4.3 Reproduction of Amblyseius swirskii fed on basic diet during the juvenile and adult
stages or switched from adulthood to artificial diets enriched with different concentrations of
Hermetia illucens prepupal hemolymph at 23°C
Diet n Oviposition
period (days)a
Oviposition rate
(eggs/female/day)b
Total fecundity
(eggs/female)b
AD0 12 5.58 ± 1.95 b 0.84 ± 0.07 b 3.50 ± 0.61 b
AD5 12 10.75 ± 1.74 a 1.21 ± 0.04 a 13.08 ± 2.14 a
AD10 9 10.89 ± 2.58 ab 1.19 ± 0.11 a 11.44 ± 2.07 a
AD20 12 11.17 ± 1.83 a 1.13 ± 0.06 a 12.00 ± 1.69 a
χ2/F 8.280 6.467 7.022
df 3 3, 41 3, 41
P 0.041 0.001 0.001
n = number of reproducing females observed. AD0, AD5, AD10 and AD20: basic artificial
diet enriched with 0%, 5%, 10% and 20% of H. illucens prepupal hemolymph, respectively.
Means ± SE within a column followed by the same letter are not significantly different
(P>0.05; aMann-Whitney U test,
bTukey test)
Diet switching influenced the reproduction of A. swirskii females (Table 4.3). Adult
females fed with AD5 and AD20 had significantly longer oviposition periods than females
maintained on AD0. Mites switched to the enriched artificial diets had significantly higher
oviposition rates and total fecundities as compared with those that were maintained on AD0,
but there was no effect of hemolymph concentration. The daily oviposition rates of females
switched to AD5, AD10 or AD20 rapidly increased in comparison to those kept on AD0 from

Chapter 4
64
the first day of the oviposition period (Figure 4.2). This trend was maintained until the
females stopped laying eggs.
Figure 4.2 Effect of diet switching from basic diet during the juvenile stages to artificial diets
enriched with different concentrations of Hermetia illucens prepupal hemolymph during the
adult stage on daily oviposition rate of Amblyseius swirskii at 23°C. AD0, AD5, AD10 and
AD20: basic artificial diet enriched with 0%, 5%, 10% and 20% of H. illucens prepupal
hemolymph, respectively.
4.4 Discussion
The development and reproduction of an arthropod natural enemy fed on an unnatural or
artificial diet are the two most important parameters to evaluate the value of the diet for
rearing purposes (Grenier and De Clercq, 2003). A more favourable diet will result in a higher
population growth, which is reflected in higher intrinsic rates of increase. In this study, A.
swirskii fed on the artificial diets enriched with prepupal hemolymph of black soldier fly, H.
illucens, had a faster development and better fecundity than when offered a basic diet without
this component. The highest intrinsic rate of increase in the current study was recorded for

Effect of H. illucens hemolymph in artificial diet of A. swirskii
65
females fed on an artificial diet supplemented with 20% of the prepupal hemolymph (AD20,
0.210 females/female/day). This growth rate is higher than the values observed on artificial
diets enriched with hemolymph of oak silkworm pupae (A. pernyi) (0.181 females/female/day
at 23 oC) (Chapter 3). The growth rate on AD20 was also superior to that reported for A.
swirskii reared on a number of natural prey species including tomato russet mite Aculops
lycopersici (Massee) (0.201 females/female/day) (Park et al., 2011), spider mite Tetranychus
urticae Koch (0.167 females/female/day) (El-Laithy and Fouly, 1992), eriophyoid fig mites
Aceria ficus (Cotte) or Rhyncaphytoptus ficifoliae Keifer (0.155 or 0.122 females/female/day,
respectively) (Abou-Awad et al., 1999) and thrips Frankliniella occidentalis (Pergande) or
Thrips tabaci Lind. (0.056 or 0.024 females/female/day, respectively) (Wimmer et al., 2008).
The highest rm-value in our study is only slightly lower than that reported by Nomikou et al.
(2001) when the predatory mite was fed on the tobacco whitefly, Bemisia tabaci (Gennadius)
(0.218 females/female/day).
Our study indicates that supplementing a nutritionally suboptimal artificial diet with
black soldier fly hemolymph significantly improved its value for supporting development and
reproduction of A. swirskii. Even at the lowest concentration of hemolymph added (5%)
juvenile survival improved, the production of viable eggs was stimulated and as a
consequence the intrinsic rate of increase substantially increased. Furthermore, in the diet
switching experiment, females reared on the basic diet in their juvenile stages and presented
with artificial diet enriched with the prepupal hemolymph in the adult stage only, showed
improved reproductive rates as compared with females maintained on the basic diet. This
confirms our earlier findings when the same basic artificial diet was enriched with 20%
hemolymph of oak silkworm pupae (A. pernyi), significantly increasing immature survival,
shortening development time and raising fecundity of A. swirskii (Chapter 3). Grenier and De
Clercq (2003) noted that adding insect components (i.e. tissues, hemolymph, cells, protein,
amino acids, etc.) to artificial diets enhances their acceptability and improves their nutritional
quality for a number of entomophagous insects. Artificial diets containing insect components
may be useful when predators or parasitoids require certain nutrients, feeding stimulants, and
other chemical cues found in arthropod prey or hosts (De Clercq, 2008). The positive role of
insect hemolymph in artificial diets is particularly well known in parasitoid wasps of the
genus Trichogramma. Strand and Vinson (1985) obtained complete in vitro culture of
Trichogramma pretiosum Riley on an artificial medium supplemented with 40% hemolymph
of Manduca sexta (L.). Trichogramma dendrolimi and T. maidis successfully developed on
artificial diets composed of egg yolk and Helicoverpa zea (Boddie) hemolymph (Grenier and

Chapter 4
66
Bonnot, 1988). Trichogramma dendrolimi was successfully reared in vitro in a medium
containing hemolymph of Antheraea pernyi (Guan et al., 1978).
The black soldier fly is considered to be a good candidate for industrial-scale
production, which is defined as a minimum reach of 1 tonne per day of fresh-weight insects
(FAO, 2012). This is related to a number of beneficial traits, including a short development
cycle, high survival of immatures and oviposition rate, good potential of biomass increase per
day (i.e. weight gain per day), high conversion rate (kg biomass gain per kg feedstock), ability
to live in high densities (kg biomass per m2); and low vulnerability to disease (Van Huis et al.,
2013). Additionally, the fly can successfully develop on a wide range of food sources
including chicken feed (Tomberlin et al., 2002; Diener et al., 2009), chicken manure
(Sheppard et al., 1994; Yu et al., 2011; Zhou et al., 2013), pig manure (Newton et al., 2005;
Zhou et al., 2013), cattle manure (Myers et al., 2008; Li et al., 2011b; Zhou et al., 2013),
human feces (Lalander et al., 2013; Banks et al., 2014), pig liver (Nguyen et al., 2013),
kitchen waste (Diener et al., 2011; Nguyen et al., 2013), fruits and vegetables (Nguyen et al.,
2013), fish offal (St-Hilaire et al., 2007; Nguyen et al., 2013), palm kernel meal (Hem et al.,
2008) and coffee pulp (Larde, 1990). These characteristics enable cost-effective production of
large amounts of insect biomass, including hemolymph. The intrinsic rate of increase of A.
swirskii fed on the basic artificial diet enriched with black soldier fly prepupal hemolymph in
this study was equally high as that of females reared on the basic diet supplemented with
hemolymph of oak silkworm pupae A. pernyi (Chapter 3). This indicates that H. illucens
prepupal hemolymph has a similar positive role in artificial diets for A. swirkii as the above-
mentioned, more expensive and/or less available arthropod materials (Lü et al., 2013; De
Clercq et al., 2014).
To our knowledge our study is the first that shows the potential of H. illucens as a
nutrient source for the rearing of beneficial arthropods. Arguably, more research is warranted
to understand the role of H. illucens hemolymph for the nutritional physiology of A. swirskii
and to explore the potential of this material to enhance the value of artificial diets for other
entomophagous mites and insects.

67
Chapter 5
DIFFERENT FACTITIOUS AND ARTIFICIAL FOODS SUPPORT THE
CONTINUOUS REARING OF AMBLYSEIUS SWIRSKII
This chapter is based on:
Nguyen, D. T., Vangansbeke, D., & De Clercq, P. (2014). Artificial and factitious foods support the
development and reproduction of the predatory mite Amblyseius swirskii. Experimental and Applied
Acarology, 62: 181-194.

Chapter 5
68
5.1 Introduction
The predatory mite Amblyseius swirskii Athias-Henriot is an economically important
biological control agent of several key pests in greenhouses, such as whiteflies, thrips,
eriophyid mites and broad mites (Nomikou et al., 2001; Messelink et al., 2008; Arthurs et al.,
2009; Park et al., 2010; van Maanen et al., 2010). Amblyseius swirskii is a type III generalist
predator, implying that it can feed on various types of food including arthropod prey, pollen,
and honeydew (McMurtry and Croft, 1997).
The potential of augmentation biological control to suppress insect pests has been
recognized for many years (Doutt and Hagen, 1949; DeBach, 1964; Parella et al., 1992).
However, augmentation is applied on a commercial scale in relatively few agricultural
systems (van Lenteren, 2012). One of the main reasons is that augmentative releases are
frequently more expensive than pesticides (Collier and Van Steenwyk, 2004). Cost-effective
rearing techniques are needed to make augmentation a more competitive strategy for
managing arthropod pests. One approach to facilitating this is reducing the costs associated
with rearing natural enemies by using factitious (i.e., unnatural) foods (Riddick, 2009).
Factitious food sources can also be used as a preventative strategy to help establish or
maintain a population of certain arthropod predators in the crop when pest populations are
low, so as to reduce the frequency of releases (Jonsson et al., 2008).
Eggs of the Mediterranean flour moth Ephestia kuehniella Zeller (Lepidoptera:
Pyralidae) have shown to be a suitable factitious food for various insect predators, including
lady beetles (De Clercq et al., 2005b), anthocorid bugs (Alauzet et al., 1992; Vacante et al.,
1997; Arijs and De Clercq, 2001), mirid bugs (Fauvel et al., 1987; Castañé et al., 2006) and
lacewings (Cohen and Smith, 1998). Cysts of the brine shrimp Artemia sp. (Anostraca:
Artemiidae) are another type of factitious food used for rearing a number of insect predators,
including Orius spp. (Hemiptera: Anthocoridae) (Arijs and De Clercq, 2001) and
Macrolophus spp. (Hemiptera: Miridae) (Castañé et al., 2006). Different species of storage
mites are routinely used as factitious prey in the mass production of phytoseiid mites
(Bolckmans and van Houten, 2006; Fidgett and Stinson, 2008).
The utilization of an artificial diet may be the next step towards a more cost-effective
rearing of predators (Riddick, 2009). Whereas several artificial diets have been developed for
predatory insects, far fewer attempts have been made at rearing predatory mites on artificial
diets. Chapter 3 reported that an artificial diet enriched with pupal hemolymph of the
Chinese oak silkworm, Antheraea pernyi (Guérin-Méneville) (Lepidoptera: Saturniidae),

Factitious and artificial foods support the continuous rearing of A. swirskii
69
allowed development and reproduction of A. swirskii to the same extent as the currently used
storage mite Carpoglyphus lactis (L.) (Acari: Carpoglyphidae). However, this hemolymph is
not easily available resulting in a higher cost and lower reliability for the diet (Xie et al., 1997;
Heslin et al., 2005). Therefore, if pupal hemolymph could be replaced by a more easily
available and cheaper nutrient, the cost of production may be reduced.
Grenier and De Clercq (2003) pointed out that, whereas measures of development and
reproduction of an arthropod natural enemy on an unnatural or artificial diet are indicative for
the value of the diet for rearing purposes, the ultimate quality parameter of an artificially
reared natural enemy is its predation or parasitization efficacy. In the present study, we
investigated the developmental, reproductive and predatory performance of A. swirskii when
fed on frozen E. kuehniella eggs, dry decapsulated cysts of A. franciscana, and on meridic
artificial diets supplemented with A. pernyi pupal hemolymph (AD1), E. kuehniella eggs
(AD2) or A. franciscana cysts (AD3). Performance of the phytoseiid after a single generation
on the various diets was compared with that after 6 generations of continuous rearing.
5.2 Materials and methods
5.2.1 Stock colony of Amblyseius swirskii
The stock colony of A. swirskii was reared as described in section 3.2.1
5.2.2 Preparation of artificial diet
Artificial diets AD1, AD2, and AD3 were prepared by supplementing a basic artificial diet
prepared according to section 3.2.4 with hemolymph from A. pernyi pupae, E. kuehniella
eggs, and decapsulated cysts of Artemia franciscana (Kellogg), respectively. Pupal
hemolymph of A. pernyi was provided in lyophilized form by the Guangdong Entomological
Institute, Guangzhou, China. Frozen eggs of E. kuehniella were supplied by Koppert B. V.
(Berkel en Rodenrijs, The Netherlands). Decapsulated A. franciscana cysts were provided by
the Artemia Reference Center (ARC) of Ghent University (Ghent, Belgium) and originated
from the Great Salt Lake (Utah, USA).

Chapter 5
70
AD1 consisted of 80% basic artificial diet supplemented with 20% (w/w) pupal
hemolymph of A. pernyi; lyophilized hemolymph was redissolved using distilled water before
being added to the diet (section 3.2.4 ). The supplements for AD2 and AD3 were prepared by
finely grinding 10 g of eggs of E. kuehniella or dry decapsulated cysts of A. franciscana,
respectively, to dust in a ceramic mortar. Next, 40 g of the basic artificial diet was added in
the mortar and the components were further ground and mixed using a pestle. The resulting
mixture was centrifuged at 12,000 rpm at 5°C for 15 min in an Eppendorf Centrifuge 5430R
(Eppendorf AG, Barkhausenweg, Hamburg, Germany), then the aqueous supernatant was
transferred to 2 ml Eppendorf tubes and subsequently stored in a freezer at -18°C.
5.2.3 Experimental setup
Eight hours before start of the experiments, new black threads were placed in the stock
colony of A. swirskii. Eggs deposited on the threads were transferred individually to the
rearing microcosms that were modified from Munger cells as described in section 3.2.5 .
Frozen eggs of E. kuehniella, dry decapsulated cysts of A. franciscana, AD1, AD2 or AD3
were supplied ad libitum. For the artificial diets, approximately 2 µl of diet was absorbed on
a small piece of filter paper (2 x 2 mm) placed on the bottom board of the rearing microcosms.
All diets were offered from the larval stage of the predator on and refreshed every 2 days. To
obtain data on the duration of each developmental stage of A. swirskii and on mortality and
escape rates, observations were made every 24 hours until all individuals had reached
adulthood. The mites were observed and the same parameters were recorded as that described
in section 3.2.6 . Mites that escaped or died due to unnatural causes were excluded from data
analysis; escape rates varied between 0 and 33%. The experiments were done in a growth
chamber at 23 ± 1°C, 65 ± 5% RH and a 16:8 h (L:D) photoperiod.
5.2.3.1 Multigeneration experiment
The offspring from first generation (G1) adults obtained in the above experiment was
maintained on the same diet up to the fifth generation. The mites were reared on green plastic
arenas (3 x 3 x 0.3 cm) (Multicel, SEDPA, France), placed on a wet sponge in a polystyrene

Factitious and artificial foods support the continuous rearing of A. swirskii
71
insect breeding dish (10 x 4 cm) (SPL Life Sciences Co. Ltd., Korea) with a mesh hole (ø 4
cm) in the lid. The dishes were kept in a growth chamber set at 23 ± 1°C, 70 ± 5% RH and a
16:8 h (L:D) photoperiod. Eggs from fifth generation females were collected to be used in an
experiment set up as described above, in order to assess the developmental and reproductive
performance in the sixth generation (G6) of rearing on each diet. However, the mites fed on
AD1 did not succeed in reaching the third generation, and could thus not be included in this
experiment.
5.2.3.2 Predation experiment
Predation capacity of female adults of A. swirskii on first instars of the Western flower thrips,
Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) was assessed in the first (G1)
and sixth (G6) generation of rearing on each of the diets, except for AD1 which was tested in
G1 only. Thrips were cultured in plastic boxes containing vermiculite and green bean pods
(Phaseolus vulgaris L.) at 23 ± 1°C, 70 ± 5% RH and a 16:8 h (L:D) photoperiod and first-
instar larvae (<24h old) were collected from the rearing containers for testing. Twenty-five
gravid females (ca. two days old) were starved for 24 h in individual glass tubes (4.5 cm in
length, 0.7 cm in diameter), stoppered with a cotton ball; starved females were not supplied
with water. Then, each female was transferred to a square bean (Phaseolus vulgaris L.) leaf
disc (25 x 25 mm) that was placed upside down on a water-saturated polyurethane sponge (10
x 50 x 50 mm) in a polystyrene insect breeding dish (100 x 40 mm) with a mesh hole (ø 40
mm) in the lid (SPL Life Sciences Co. Ltd., Korea). The four edges of the leaf disc were
covered with moist tissue paper to provide free water and preclude escaping. Fifteen first
instars of F. occidentalis) were placed on each leaf disc as prey for the mites. After 24 h, the
number of thrips corpses was recorded. Females which escaped or were found dead on the
moist tissue paper were excluded from analysis; escape rates varied between 0 and 20%.
5.2.4 Life table parameters calculation
Life table parameters were calculated as explained in section 3.2.7

Chapter 5
72
5.2.5 Statistical analysis
Two-way analysis of variance (ANOVA) (IBM, SPSS Statistics 20) was conducted to
evaluate the effects of diet and generation on the duration of the immature stages,
preoviposition and oviposition period, daily and total oviposition, adult longevity and life
table parameters of A. swirskii. When an interaction was detected between the main factors,
means were compared among diets and a pairwise multiple comparison procedure was used
(Kutner et al., 2005). When a Kolmogorov–Smirnov test indicated that means were normally
distributed, the parameter was analyzed using a one-way analysis of variance (ANOVA). If a
Levene test indicated heteroscedasticity, a Tamhane test was used instead of a Tukey test.
When means were not normally distributed, a nonparametric Kruskal-Wallis ANOVA was
used and means were separated using a Mann-Whitney U test. Immature survival rates and
sex ratios were compared by means of a logistic regression. This regression is a generalized
linear model using a probit (log odds) link and a binomial error function. Each test consists of
a regression coefficient that is calculated and tested for being significantly different from zero,
for which P-values are presented (McCullagh and Nelder, 1989). In all tests, P-values smaller
than or equal to 0.05 were considered significant.
5.3 Results
Whereas in G1 the immature mortality of A. swirskii did not differ among diets, mortality in
G6 was significantly higher than in G1, except on a diet of A. franciscana cysts (χ2=127.71,
df=8, P<0.001). In G6, mites fed on AD2 or AD3 suffered from greater mortality (65.6 and
37.1%, respectively) than those reared on E. kuehniella or A. franciscana (13.6 and 1.9%,
respectively) (Figure 5.1). Mortality of mites in G6 continuously reared on the artificial diets
occurred mainly in the egg and larval stages.

Factitious and artificial foods support the continuous rearing of A. swirskii
73
Figure 5.1 Mortality (%) of the different immature stages of Amblyseius swirskii fed in the
first (G1) and sixth (G6) generation on different factitious and artificial foods. Numbers
within or above the different sections in each bar refer to mortality percentages in the egg,
larval, protonymphal and/or deutonymphal stages.
Bars with the same letter are not significantly different (P>0.05; Probit (Wald Chi-square)
test). AD1, artificial diet 1 (with pupal hemolymph of A. pernyi); AD2, artificial diet 2 (with
E. kuehniella eggs); AD3, artificial diet 3 (with A. franciscana cysts).

Chapter 5
74
Table 5.1 Two-way ANOVA indicating the effect of diet and generation on developmental, reproductive and predation parameters of
Amblyseius swirskii
Source Duration of
immature stage
Pre-
oviposition
period
Oviposition
period
Female
longevity
Total
number
of eggs
Oviposition
rate
Net
reproductive
rate
Generation
time
Intrinsic
rate of
increase
Predation
rate Females Males
Diet F 9.948 5.79 5.658 15.271 2.075 5.341 22.730 33.975 18.674 138.47 6.405
df 3 3 3 3 3 3 3 3 3 3 3
P <0.001 0.001 0.001 <0.001 0.104 0.001 <0.001 <0.001 <0.001 <0.001 <0.001
Generation F 0.784 0.862 2.874 0.014 0.209 4.668 14.877 96.117 4.738 313.677 30.986
df 1 1 1 1 1 1 1 1 1 1 1
P 0.377 0.355 0.091 0.906 0.648 0.032 <0.001 <0.001 0.031 <0.001 <0.001
Diet x
Generation
F 4.01 0.26 2.708 2.458 0.997 0.837 2.203 11.221 7.379 81.935 0.354
df 3 3 3 3 3 3 3 3 3 3 3
P 0.008 0.854 0.046 0.064 0.395 0.475 0.089 <0.001 <0.001 <0.001 0.786
Error df 257 111 229 229 229 229 229 229 229 229 176

Factitious and artificial foods support the continuous rearing of A. swirskii
75
Two-way ANOVA (Table 5.1) showed significant interactions between diet and
generation for the developmental time of females, whereas no interaction occurred for male
developmental time. The developmental time of males was significantly affected by diet but
did not differ between generations. In G1, the males fed on E. kuehniella eggs had the longest
developmental time, but in G6 male developmental time did not differ among diets.
Developmental times of females offered E. kuehniella eggs did not differ between
generations, whereas they were shorter in G6 on A. franciscana and AD2, and longer on AD3
as compared to those in G1 (Table 5.2).
Table 5.2 Development time (days) of Amblyseius swirskii fed in the first (G1) and sixth (G6)
generation on different factitious and artificial foods
Diet/Generation n
a Developmental duration (days)
b
Females n Males n
E. kuehniella/G1 63 8.21 ± 0.25 ab 48 8.23 ± 0.46 a 13
E. kuehniella/G6 59 8.00 ± 0.25 ab 32 8.26 ± 0.57 a 19
A. franciscana/G1 55 7.67 ± 0.14 a 33 7.09 ± 0.09 bc 22
A. franciscana/G6 52 7.42 ± 0.12 bcd 33 7.33 ± 0.16 ab 18
AD1/G1 57 7.65 ± 0.18 abcd 31 6.96 ± 0.11 c 24
AD2/G1 56 7.36 ± 0.14 cd 33 7.00 ± 0.07 c 22
AD2/G6 64 6.67 ± 0.24 e 18 7.00 ± 0.41 ac 4
AD3/G1 50 7.31 ± 0.10 d 39 6.91 ± 0.09 c 11
AD3/G6 62 7.97 ± 0.14 a 29 7.50 ± 0.22 ab 10
χ2 36.75 30.99
df 8 8
P <0.001 <0.001
aInitial number of tested individuals;
bMeans ± SE; means within a column followed by the
same letter are not significantly different (P>0.05), according to a Mann-Whitney U test
(duration); χ2-, df- and P-values refer to Kruskal-Wallis ANOVAs. AD1, artificial diet 1
(with pupal hemolymph of A. pernyi); AD2, artificial diet 2 (with E. kuehniella eggs); AD3,
artificial diet 3 (with A. franciscana cysts).

Chapter 5
76
Table 5.3 Reproduction and longevity of Amblyseius swirskii in the first (G1) and sixth (G6) generation on different factitious and artificial foods
Diet/Generation n Preoviposition period (days)
a
Oviposition period (days)
a
Female longevity (days)
a
Oviposition rate (eggs/female/day)
a
Total number of eggs
(eggs/female)a
Female proportion of the
progenya
E. kuehniella/G1 42 2.41 ± 0.10 bc 21.83 ± 1.09 b 44.12 ± 3.28 a 1.48 ± 0.04 ab 31.64 ± 1.51 ab 0.71 ± 0.01 ab E. kuehniella/G6 32 3.91 ± 0.79 a 19.47 ± 1.15 b 41.06 ± 2.85 a 1.48 ± 0.05 abc 28.34 ± 1.70 b 0.66 ± 0.02 c
A. franciscana/G1 28 2.39 ± 0.13 bc 25.64 ± 1.93 ab 37.79 ± 3.04 a 1.50 ± 0.04 ab 37.71 ± 2.75 ab 0.75 ± 0.02 a A. franciscana/G6 33 2.27 ± 0.08 c 28.94 ± 1.38 a 41.79 ± 2.54 a 1.27 ± 0.04 cde 35.48 ± 1.27 a 0.71 ± 0.02 abc
AD1/G1 28 2.71 ± 0.18 ab 21.82 ± 1.31 b 40.32 ± 3.54 a 1.60 ± 0.05 a 34.50 ± 2.18 ab 0.74 ± 0.02 ab
AD2/G1 29 3.24 ± 0.29 a 33.55 ± 2.40 a 49.14 ± 2.92 a 1.13 ± 0.07 de 34.10 ± 2.07 ab 0.70 ± 0.02 bc AD2/G6 12 3.42 ± 0.42 a 28.58 ± 3.96 ab 46.00 ± 5.22 a 0.91 ± 0.09 e 26.25 ± 4.58 ab 0.72 ± 0.03 abc
AD3/G1 32 2.06 ± 0.04 d 22.50 ± 0.91 b 41.50 ± 2.42 a 1.38 ± 0.04 bcd 30.72 ± 1.22 ab 0.76 ± 0.02 a AD3/G6 29 2.21 ± 0.09 cd 25.34 ± 1.87 ab 46.76 ± 3.30 a 1.21 ± 0.04 de 29.90 ± 2.18 ab 0.75 ± 0.02 ab
χ2/F 44.19 7.54 1.22 13.43 2.82 17.90
df 8 8, 256 8, 256 8, 256 8, 256 8 P <0.001 <0.001 0.287 <0.001 0.005 0.022
aMeans ± SE; means within a column followed by the same letter are not significantly different (P > 0.05) according to a Mann-Whitney U test
(preoviposition period), Tamhane test (oviposition period, oviposition rate, total number of eggs), Tukey test (female longevity), or Probit (Wald
Chi-square) test (female proportion of progeny); χ2-, df- and P-values refer to Kruskal-Wallis ANOVAs; F-, df- and P-values refer to one-way
ANOVAs. AD1, artificial diet 1 (with pupal hemolymph of A. pernyi); AD2, artificial diet 2 (with E. kuehniella eggs); AD3, artificial diet 3
(with A. franciscana cysts).

Factitious and artificial foods support the continuous rearing of A. swirskii
77
Two-way ANOVA showed significant interactions between diet and generation for
preoviposition period but there was no interaction for the parameters oviposition period,
female longevity, oviposition rate and total number of eggs (Table 5.1). The preoviposition
periods did not differ between G1 and G6 on all diets, except E. kuehniella eggs (Table 5.3).
The shortest preoviposition period was recorded on AD3 in G1 (2.06 days). Oviposition
periods were similar in both generations and ranged from 19.5 days to 33.6 days for females
fed on E. kuehniella in G6 and AD2 in G1, respectively. Female longevity was not affected by
diet or generation. The proportion of female progeny did not differ between generations,
except on E. kuehniella. Daily oviposition rates were influenced by both diet and generation
and ranged from 0.91 to 1.60 eggs/female/day. In G1, the females reared on E. kuehniella, A.
franciscana or AD1 showed similar oviposition rates, which were in turn significantly higher
than those on AD2. Oviposition rates did not differ between generations when the mites were
fed on E. kuehniella, AD2 or AD3. No differences were observed between generations in the
total number of eggs produced when females were reared on the different diets. Total
fecundity was similar among females reared on the different diets except in G6 where the
females supplied with A. franciscana cysts produced more eggs than those given E. kuehniella
eggs.
Significant interactions between diet and generation were found for the life table
parameters net reproductive rate, generation time and intrinsic rate of increase (Table 5.1).
The net reproductive rates in G1 were significantly higher than those in G6, except on A.
franciscana (Table 5.4). In G1, different diets did not result in different net reproductive rates.
Females in G6 had longer generation times than those in G1 on all diets, except on A.
franciscana cysts, which yielded a shorter generation time in G6 than in G1. Likewise, the
intrinsic rates of increase in G1 were significantly higher than in G6, but not on a diet of A.
franciscana cysts for which similar growth rates were observed in both generations.
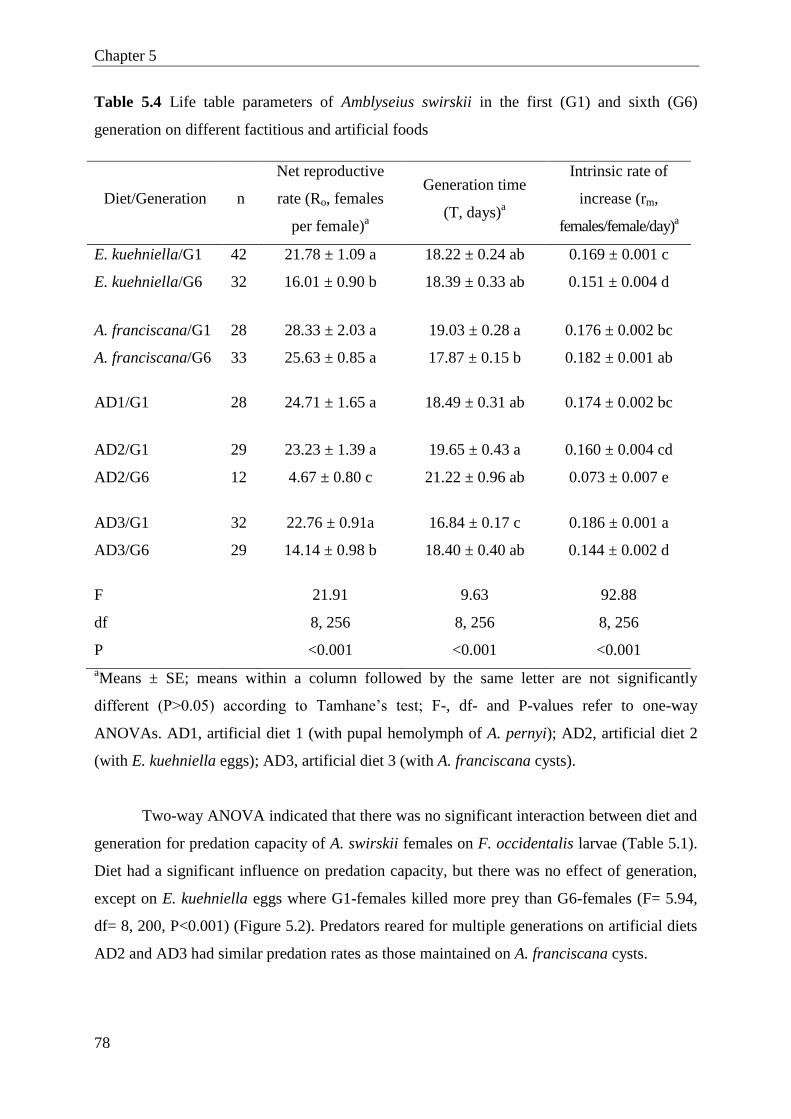
Chapter 5
78
Table 5.4 Life table parameters of Amblyseius swirskii in the first (G1) and sixth (G6)
generation on different factitious and artificial foods
Diet/Generation n
Net reproductive
rate (Ro, females
per female)a
Generation time
(T, days)a
Intrinsic rate of
increase (rm,
females/female/day)a
E. kuehniella/G1 42 21.78 ± 1.09 a 18.22 ± 0.24 ab 0.169 ± 0.001 c
E. kuehniella/G6 32 16.01 ± 0.90 b 18.39 ± 0.33 ab 0.151 ± 0.004 d
A. franciscana/G1 28 28.33 ± 2.03 a 19.03 ± 0.28 a 0.176 ± 0.002 bc
A. franciscana/G6 33 25.63 ± 0.85 a 17.87 ± 0.15 b 0.182 ± 0.001 ab
AD1/G1 28 24.71 ± 1.65 a 18.49 ± 0.31 ab 0.174 ± 0.002 bc
AD2/G1 29 23.23 ± 1.39 a 19.65 ± 0.43 a 0.160 ± 0.004 cd
AD2/G6 12 4.67 ± 0.80 c 21.22 ± 0.96 ab 0.073 ± 0.007 e
AD3/G1 32 22.76 ± 0.91a 16.84 ± 0.17 c 0.186 ± 0.001 a
AD3/G6 29 14.14 ± 0.98 b 18.40 ± 0.40 ab 0.144 ± 0.002 d
F 21.91 9.63 92.88
df 8, 256 8, 256 8, 256
P <0.001 <0.001 <0.001
aMeans ± SE; means within a column followed by the same letter are not significantly
different (P>0.05) according to Tamhane’s test; F-, df- and P-values refer to one-way
ANOVAs. AD1, artificial diet 1 (with pupal hemolymph of A. pernyi); AD2, artificial diet 2
(with E. kuehniella eggs); AD3, artificial diet 3 (with A. franciscana cysts).
Two-way ANOVA indicated that there was no significant interaction between diet and
generation for predation capacity of A. swirskii females on F. occidentalis larvae (Table 5.1).
Diet had a significant influence on predation capacity, but there was no effect of generation,
except on E. kuehniella eggs where G1-females killed more prey than G6-females (F= 5.94,
df= 8, 200, P<0.001) (Figure 5.2). Predators reared for multiple generations on artificial diets
AD2 and AD3 had similar predation rates as those maintained on A. franciscana cysts.

Factitious and artificial foods support the continuous rearing of A. swirskii
79
Figure 5.2 Mean (±SE) number of first instar Frankliniella occidentalis killed during a 24-h
period by Amblyseius swirskii females in the first (G1) and sixth (G6) generation on different
factitious and artificial foods
Bars with the same letter are not significantly different (P>0.05; Tukey test). Number of
replicates is indicated by the number in each bar. AD1, artificial diet 1 (with pupal
hemolymph of A. pernyi); AD2, artificial diet 2 (with E. kuehniella eggs); AD3, artificial diet
3 (with A. franciscana cysts).
5.4 Discussion
The main goals of this study were to (1) investigate the potential of two factitious food
sources (A. franciscana and E. kuehniella) that are routinely used for the production of a
number of insect predators, for rearing the phytoseiid predator A. swirskii, (2) evaluate the
potential of arthropod components that can improve the artificial diet developed in Chapter 3
for this predatory mite, (3) assess the effects of long-term rearing on these factitious foods and
artificial diets on the predator’s fitness.
Our results indicate that all factitious and artificial foods tested were accepted by the
different life stages of A. swirskii and sustained the development and reproduction of the
predator for at least a single generation. The highest intrinsic rates of increase in our study
were recorded in the first generation when A. swirskii was fed on decapsulated cysts of A.
franciscana and AD3, containing 20% of finely ground cysts (0.182 and 0.186
females/female/day, respectively). These values are higher than those reported on various

Chapter 5
80
natural prey, including Tetranychus urticae Koch (Acari: Tetranychidae) (0.167 at 26oC) (El-
Laithy and Fouly, 1992), F. occidentalis (0.056 at 25oC) and Thrips tabaci (Lindeman)
(Thysanoptera: Thripidae) (0.024 at 25oC) (Wimmer et al., 2008), and only slightly lower than
those reported by Park et al. (2011) on the tomato russet mite, Aculops lycopersici (Massee)
(Acari: Eriophyidae) (0.201 at 25oC) and Nomikou et al. (2001) on the cotton whitefly
Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) (0.213 at 27oC). In the first generation,
growth rates on all tested diets were also similar to or greater than those found on T. latifolia
pollen (0.158 at 23oC) and Carpoglyphus lactis (L.) (Acari: Carpoglyphidae) (0.175 at 23
oC)
(Chapter 3), which is another widely used food in the laboratory rearing and commercial
mass rearing of this phytoseiid. However, it deserves emphasis that comparison of life table
statistics among studies is complicated by differences in experimental methods, climatic
conditions and the calculation of estimates.
In a previous study, we found that enriching a basic artificial diet with pupal
hemolymph of the Chinese oak silkworm, A. pernyi, substantially improved fecundity in A.
swirskii (Chapter 3). In recent years, however, production of this silkworm in China has
decreased. As a result, pupal hemolymph is becoming less easily available and more costly
(Lü et al., 2013), rendering its use as a dietary supplement less attractive. In contrast, eggs of
E. kuehniella and cysts of A. franciscana are widely available. In the present study, growth
rates of A. swirskii on artificial diets enriched with finely ground E. kuehniella eggs (AD2) or
A. franciscana cysts (AD3) were similar to or higher than those on the diet supplemented with
pupal hemolymph of A. pernyi (AD1). Our findings suggest that certain components of E.
kuehniella eggs and A. franciscana cysts can play a critical role in stimulating development
and reproduction of A. swirskii. The positive effect of E. kuehniella on reproduction in the
present study is consistent with the result reported by Ferkovich et al. (2007) who offered the
anthocorid bug Orius insidiosus (Say) an artificial diet supplemented with protein from E.
kuehniella eggs, resulting in a higher oviposition rate compared to an unmodified artificial
diet. Further research is warranted to identify the components of the E. kuehniella eggs and A.
franciscana cysts which are responsible for the increased performance. However, A.
franciscana cysts have a number of practical advantages over E. kuehniella eggs when used
for rearing predatory arthropods. The market price of decapsulated Artemia cysts is
substantially lower than that of E. kuehniella eggs: whereas frozen E. kuehniella eggs
currently cost around 400-500 EUR/kg (F. Wäckers, 2014, pers. comm.), the market price of
decapsulated Artemia cysts is less than 10% of that (G. Van Stappen, 2013, pers. comm.).

Factitious and artificial foods support the continuous rearing of A. swirskii
81
Unlike E. kuehniella eggs, Artemia cysts can be stored in dry form for several years in a cool
and dry place without the need for deep freezing (Arijs and De Clercq, 2001).
An effective factitious or artificial food should satisfy the nutritional requirements of a
predator and ensure the continuous production of progeny of high quality (Cohen, 2004). As
nutrient imbalances in a diet may be expressed only after several generations of rearing (De
Clercq et al., 2005a), it was the objective to culture the predator for five consecutive
generations on each diet and assess its fitness in the subsequent generation. In G6, the
immature survival of predators reared on E. kuehniella, AD2 and AD3 was significantly lower
than that on the corresponding diet in G1, with stronger declines on the artificial diets than on
lepidopteran eggs. These declines were reflected in the intrinsic rates of increase on the diets.
In contrast, the predator’s performance remained at similar levels across generations on dry
decapsulated cysts of A. franciscana and allowed the production of 15 successive generations
up to present. In addition, artificial diet AD3, supplemented with A. franciscana cysts, proved
superior to the other artificial diets as it allowed to sustain A. swirskii for over 15 consecutive
generations despite lower immature survival rates, whereas the colonies on AD1 and AD3
could not be maintained beyond 3 and 9 generations, respectively.
Whereas Orius bugs can only effectively feed on fully hydrated Artemia cysts (Arijs
and De Clercq, 2001), in the present study the decapsulated cysts did not need to be hydrated
to enable the A. swirskii mites to feed upon them. Dry (i.e., containing 4% of water)
decapsulated A. franciscana cysts supplied in the microcosms, where relative humidity
exceeded 80%, appeared to take up sufficient water from the environment to allow effective
extraction of nutrients by the predatory mites. Vandekerkhove et al. (2009) reported that also
Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae) was able to feed on dry cysts
presented on leaf disks, as contact with the leaf surface allowed partial hydration (up to 20%)
of the cysts. As a practical advantage, dry cysts lead to less problems with moulds in the
rearing containers than fully hydrated cysts (Vandekerkhove et al., 2009). Likewise, all
artificial diets used in the present study were liquid, complicating their use in large-scale
cultures of the mites. Further research (Chapter 6) will focus on dry artificial diets, which are
easier to apply in the cultures and less prone to spoilage.
It is imperative that a predator retains its potential to find and kill the target prey after
long-term rearing on an unnatural prey or artificial diet (Grenier and De Clercq, 2003). Our
study indicates that A. swirskii females did not lose their ability to capture and kill live prey
after six generations of rearing on the tested diets. Only predation rates of predators
maintained on E. kuehniella eggs were slightly lower in G6 than in G1. All predation rates of

Chapter 5
82
A. swirskii females on first instars of F. occidentalis observed in the present study (5.3-8.5
thrips/day) were somewhat higher than those reported by Messelink et al. (2008).
In conclusion, the different factitious prey and artificial diets tested in the present
study supported development and reproduction of A. swirskii for a single generation, but
losses in fitness of the phytoseiid were observed after several generations of rearing on E.
kuehniella eggs and all of the artificial diets. Dry decapsulated A. franciscana cysts proved to
be the most suitable diet for the production of this economically important biological control
agent. More research will be needed to evaluate the suitability of Artemia cysts as a food in
large-scale cultures of A. swirskii or to support existing populations of the mite in the crop.
The potential of this factitious food for other predatory mites also deserves to be investigated.

83
Chapter 6
SOLID ARTIFICIAL DIETS FOR AMBLYSEIUS SWIRSKII
This chapter is based on:
Nguyen, D. T., Vangansbeke, D., & De Clercq, P. 2014. Solid artificial diets for the phytoseiid
predator Amblyseius swirskii. BioControl, 59: 719-727

Chapter 6
84
6.1 Introduction
In augmentative biological control programs, mass produced arthropod natural enemies are
often released in high numbers to obtain pest suppression (Stinner, 1977; Collier and Van
Steenwyk, 2004). Optimizing mass production methods, for instance by developing cost
effective factitious or artificial foods, may reduce the market price of biological control agents
and improve their adoption by growers (De Clercq et al., 2014). The effectiveness of
augmentative releases can also be enhanced by supplementing foods that increase survival
and reproduction of the natural enemies after release, and arrest their emigration from the
targeted area (Wade et al., 2008; Lundgren, 2009a).
The generalist predatory mite Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae)
has been shown to be an effective biological control agent of whiteflies (Nomikou et al., 2002;
Messelink et al., 2008), thrips (Messelink et al., 2006; Messelink et al., 2008) and broad mites
(van Maanen et al., 2010) in several greenhouse crops. The predatory mite also feeds on non-
prey foods like pollen and honeydew (Momen and El-Saway, 1993). The mass rearing
procedures for A. swirskii are based on the use of storage mites, like Carpoglyphus lactis L.
(Acari: Carpoglyphidae) and Thyreophagus entomophagus (Laboulbene) (Acari: Acaridae), as
a food source (Bolckmans and van Houten, 2006; Fidgett and Stinson, 2008).
In chapter 3 and 5, we found that A. swirskii performed well on liquid artificial diets
enriched with a watery extract of Artemia franciscana (Kellogg) (Anostraca: Artemiidae)
cysts or with pupal hemolymph of the Chinese oak silkworm Antheraea pernyi (Guérin-
Méneville) (Lepidoptera: Saturniidae), indicating the potential of these diets to sustain
populations of the predator in the laboratory rearing as well as in the crop after release.
However, liquid diets have certain disadvantages compared to solid diets in stickiness and in
many cases a need for encapsulation (Morales-Ramos et al., 2014).
Solid artificial diets have mostly been developed for arthropods with chewing
mouthparts (Morales-Ramos et al., 2014). However, Cohen (1998) noted that arthropods with
extra-oral digestion, which include phytoseiid mites, can also feed on solid diets. The
objectives of the present study were to determine developmental and reproductive parameters
of A. swirskii fed on cattail pollen (Typha latifolia L.), on lyophilized forms of liquid artificial
diets supplemented with a watery extract of decapsulated cysts of A. franciscana (ArF) or
with pupal hemolymph of A. pernyi (AnF), and on solid artificial diets supplemented with
powdered dry A. franciscana cysts (ArP) or with lyophilized pupal hemolymph of A. pernyi

Solid artificial diets for A. swirskii
85
(AnP). The objective of this study was to design a solid artificial diet that can be used for the
mass production of A. swirskii as well as to support its populations in the crop.
6.2 Materials and methods
6.2.1 Stock colony of Amblyseius swirskii
A stock colony of A. swirskii was reared as described in section 3.2.1
6.2.2 Preparation of artificial diet
Artificial diets ArF and AnF were prepared according section 5.2.2 . The diets were prepared
based on 80 % w/w basic liquid artificial diet (section 3.2.4 supplemented with 20 % w/w
watery extracts of decapsulated cysts of A. franciscana (ArF) or pupal hemolymph from the
Chinese oak silkworm A. pernyi (AnF), respectively. Pupal hemolymph of A. pernyi was
provided in lyophilized form by the Guangdong Entomological Institute, Guangzhou, China.
Decapsulated A. franciscana cysts were provided by the Artemia Reference Center of Ghent
University (Ghent, Belgium) and originated from the Great Salt Lake (Utah, USA).
Diets ArF and AnF were frozen at -18 °C and lyophilized at −57.2 °C and 0.034 mbar
for 6 days (VaCo 5-D freeze-dryer, Zirbus Technology Benelux B.V., Germany). Then the
lyophilized diets were finely ground (particle diameter smaller than 0.3 mm) using a Bosch
coffee grinder (Robert Bosch Hausgeräte GmbH, Munich, Germany), dispensed into 2 ml
Eppendorf tubes and stored at −18 °C.
Artificial diets ArP and AnP were powdered diets composed of 16.6 % sucrose, 16.6 %
tryptone, 16.6 % yeast extract, 6.7 % D-(+)-glucose anhydrous (MP Biomedicals LLC,
Illkirch, France), 6.7 % fructose (Sigma Aldrich Chemie GmbH, Steinheim, Germany), 16.6 %
egg yolk powder (Bouwhuis Enthoven BV, Raalte, The Netherlands), 0.13 % vitamin mix
based on the composition of bovine liver (weight percentages: 25.4 % nicotinic acid, 4.9 %
riboflavin, 0.5 % thiamine, 1.5 % vitamin B6, 12.4 % Ca-pantothenate, 1 % folic acid, 0.1 %
biotin and 54.2 % vitamin C) (Vandekerkhove et al., 2006) and 20 % w/w powdered dry
decapsulated A. franciscana cysts (ArP) or lyophilized pupal hemolymph of A. pernyi (AnP).
The ingredients of both diets were finely ground using a Bosch coffee grinder. Diet particle
diameter was measured using a Leica M80 microscope (Leica Microsystems Belgium BVBA,

Chapter 6
86
Diegem, Belgium) and AxioVision Ver. 4.8.2.0 software (Carl Zeiss MicroImaging GmbH,
Jena, Germany), indicating that the particle diameter was smaller than 0.3 mm. Next, the diets
were dispensed into 2 ml Eppendorf tubes and stored at −18 °C.
6.2.3 Development and reproduction on the different artificial diets
Eggs (less than 8 hours old) were transferred individually from the A. swirskii colony to
rearing microcosms that were modified from Munger cells as described in section 3.2.5 and
offered T. latifolia pollen or one of the artificial diets. All foods were supplied ad libitum. The
foods were offered from the larval stage of the predator on and refreshed every 2 days. After
they had been taken out of the freezer, the diets were kept in a refrigerator at 5 °C for max. 1
week. The mites were observed and different parameters were recorded same as that
described in section 3.2.6 Mites that escaped or died as a result of manipulation were
excluded from data analysis; these escape and death rates did not differ among treatments and
varied between 11 and 18 % (χ2
= 0.730, df = 4; P = 0.948; Probit (Wald Chi-square) test).
The experiments were done in a growth chamber at 23 ± 1 °C, 65 ± 5 % RH and a 16:8 h (L:D)
photoperiod.
6.2.4 Pre-establishment greenhouse experiment
The potential of powdered diets to sustain the pre-establishment of A. swirskii was tested on
chrysanthemum plants in a greenhouse at the Research Center for Ornamental Crops (PCS,
Destelbergen, Belgium). The experiments were conducted between May and August, 2014.
Four young plants (10-12 cm high with 10–20 leaves) were planted in a pot and 4 pots were
placed in a mesh cage (40 x 90 x 80 cm); 8 cages were placed on a bench for each treatment.
A single cage with 4 chrysanthemum pots was seen as one replicate. To prevent predatory
mites from migrating, each pot was placed on capillary matting saturated with water and the
plants did not touch the sides of the cages. Five adult female predatory mites, collected from
the laboratory culture with a fine brush, were released on each plant. Thereafter, three diets,
i.e. the basic (non-supplemented) powdered diet, powdered diet ArP and Nutrimite, a
commercial pollen (Biobest N.V., Westerlo, Belgium) consisting of pollen of narrow-leaved

Solid artificial diets for A. swirskii
87
cattail, Typha angustifolia L., were sprayed over the caged plants by way of a Nutrigun,
provided by Biobest N.V. In addition to the three diet supplement treatments, a control
treatment without food was included. All diets were re-applied every two weeks, according to
recommendations given by the producer (Biobest N.V.) for Nutrimite. The numbers of
predatory mites were assessed every 2 weeks in situ by counting the mobile mites on 5
random leaves per pot, resulting in a total of 20 leaves being checked in each cage.
6.2.5 Life table parameters calculation
Life table parameters were calculated as explained in section 3.2.7
6.2.6 Statistical analysis
Data were subjected to statistical analysis (IBM SPSS Statistics, Ver. 21) to evaluate the
effect of diet on the developmental time, preoviposition and oviposition period, daily and total
oviposition, and adult longevity of A. swirskii. When a Kolmogorov–Smirnov test indicated
that means were normally distributed, the parameter was analysed using a one-way analysis of
variance (ANOVA). If a Levene test indicated heteroscedasticity, a Tamhane test was used
instead of a Tukey test. When means were not normally distributed, a nonparametric Kruskal-
Wallis ANOVA was used and means were separated using Mann-Whitney U tests. Immature
survival rates and sex ratios were compared by means of a logistic regression. This regression
is a generalized linear model using a probit (log odds) link and a binomial error function.
Each test consists of a regression coefficient that is calculated and tested for being
significantly different from zero, for which P-values are presented (McCullagh and Nelder,
1989).
To test whether there was a consistent difference between densities of the predatory
mite supplemented in the greenhouse with three diets over a period of 16 weeks, the data of
nine independent monitoring times were analyzed by repeated measures ANOVA (between
subject factor: diet; within subject factor: time of sampling). If treatment differences at a
sampling date were significant, one-way analysis of variance (ANOVA) or Kruskal-Wallis
ANOVA was used to compare treatment means.
In all tests, P-values smaller than or equal to 0.05 were considered significant.

Chapter 6
88
6.3 Results
Immature survival of A. swirskii did not differ among diets ranging from 98.2 to 100 % (χ2 =
0.00, df=4, p>0.999; Probit (Wald Chi square) test). Development times of both A. swirskii
females and males were significantly affected by diet (Table 6.1). Females fed on T. latifolia
pollen developed significantly faster to adulthood than those fed on the other diets.
Development times of males offered T. latifolia pollen or both lyophilized diets (ArF and AnF)
were shorter than of those given the powdered diets (ArP and AnP).
Table 6.1 Development time (days) of Amblyseius swirskii fed on T. latifolia pollen or
different solid artificial foods
Diet Developmental duration (days)*
Females n Males n
T. latifolia 6.14 ± 0.11 a 35 5.90 ± 0.18 a 20
ArF 6.97 ± 0.16 b 35 6.25 ± 0.17 a 16
AnF 6.59 ± 0.15 b 37 6.11 ± 0.17 a 19
ArP 8.09 ± 0.08 c 34 7.92 ± 0.08 b 13
AnP 7.97 ± 0.11 c 32 7.71 ± 0.09 b 28
χ2 94.321 64.775
df 4 4
P <0.001 <0.001
n: Number of tested individuals
*Means ± SE; means within a column followed by the same letter are not significantly
different (P>0.05), according to Mann-Whitney U test; χ2-, df- and P-values refer to Kruskal-
Wallis ANOVAs. ArF or AnF: freeze-dried liquid artificial diet supplemented with A.
franciscana cysts or with pupal hemolymph of A. pernyi, respectively; ArP or AnP: basic
powdered diet supplemented with powdered A. franciscana cysts or lyophilized pupal
hemolymph of A. pernyi, respectively.

Solid artificial diets for A. swirskii
89
Table 6.2 Reproduction and longevity of Amblyseius swirskii fed on T. latifolia pollen or different solid artificial foods
Diet n Preoviposition
period (days)a
Oviposition
period (days)a
Female
longevity (days)a
Oviposition rate
(eggs/female/day)a
Total number of
eggs (eggs/female)a
Female proportion
of the progeny a
T. latifolia 31 2.35 ± 0.09 a 22.39 ± 1.21 a 36.35 ± 2.36 b 1.57 ± 0.04 a 34.29 ± 1.40 ab 0.78 ± 0.01 a
ArF 30 2.97 ± 0.16 b 24.73 ± 1.15 a 44.73 ± 2.16 ab 1.49 ± 0.03 a 36.37 ± 1.53 a 0.80 ± 0.02 a
AnF 31 2.55 ± 0.14 a 24.06 ± 1.60 a 45.90 ± 2.42 a 1.59 ± 0.05 a 36.97 ± 1.87 a 0.79 ± 0.02 a
ArP 28 3.71 ± 0.09 c 25.43 ± 1.18 a 41.07 ± 1.99 ab 1.25 ± 0.04 b 30.75 ± 0.83 b 0.76 ± 0.03 a
AnP 28 3.71 ± 0.09 c 25.89 ± 1.34 a 39.14 ± 1.82 ab 1.19 ± 0.05 b 30.79 ± 1.94 ab 0.72 ± 0.04 a
χ2/F
71.183 1.100 3.354 20.185 3.503 4.626
df
4 4, 143 4, 143 4, 143 4, 143 4
P
<0.001 0.359 0.012 <0.001 0.009 0.328
n: Number of tested females
aMeans ± SE; means within a column followed by the same letter are not significantly different (P > 0.05) according to Mann-Whitney U test
(preoviposition period), Tukey test (oviposition period, female longevity, oviposition rate), Tamhane test (total number of eggs) or Probit (Wald
Chi-square) test (female proportion of progeny); χ2-, df- and P-values refer to Kruskal-Wallis ANOVAs, and F-, df- and P-values refer to one-
way ANOVAs. ArF or AnF: freeze-dried liquid artificial diets supplemented with A. franciscana cysts or with pupal hemolymph of A. pernyi,
respectively; ArP or AnP: basic powdered diet supplemented with powdered A. franciscana cysts or lyophilized pupal hemolymph of A. pernyi,
respectively.
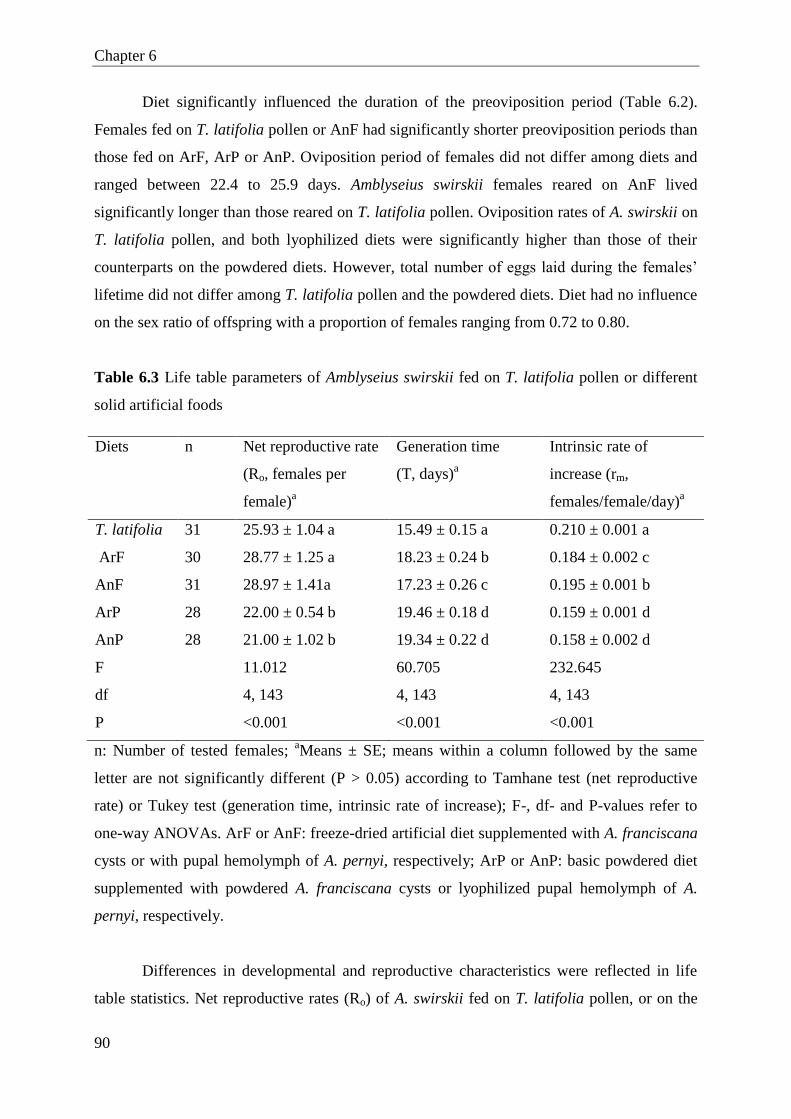
Chapter 6
90
Diet significantly influenced the duration of the preoviposition period (Table 6.2).
Females fed on T. latifolia pollen or AnF had significantly shorter preoviposition periods than
those fed on ArF, ArP or AnP. Oviposition period of females did not differ among diets and
ranged between 22.4 to 25.9 days. Amblyseius swirskii females reared on AnF lived
significantly longer than those reared on T. latifolia pollen. Oviposition rates of A. swirskii on
T. latifolia pollen, and both lyophilized diets were significantly higher than those of their
counterparts on the powdered diets. However, total number of eggs laid during the females’
lifetime did not differ among T. latifolia pollen and the powdered diets. Diet had no influence
on the sex ratio of offspring with a proportion of females ranging from 0.72 to 0.80.
Table 6.3 Life table parameters of Amblyseius swirskii fed on T. latifolia pollen or different
solid artificial foods
Diets n Net reproductive rate
(Ro, females per
female)a
Generation time
(T, days)a
Intrinsic rate of
increase (rm,
females/female/day)a
T. latifolia 31 25.93 ± 1.04 a 15.49 ± 0.15 a 0.210 ± 0.001 a
ArF 30 28.77 ± 1.25 a 18.23 ± 0.24 b 0.184 ± 0.002 c
AnF 31 28.97 ± 1.41a 17.23 ± 0.26 c 0.195 ± 0.001 b
ArP 28 22.00 ± 0.54 b 19.46 ± 0.18 d 0.159 ± 0.001 d
AnP 28 21.00 ± 1.02 b 19.34 ± 0.22 d 0.158 ± 0.002 d
F
11.012 60.705 232.645
df
4, 143 4, 143 4, 143
P
<0.001 <0.001 <0.001
n: Number of tested females; aMeans ± SE; means within a column followed by the same
letter are not significantly different (P > 0.05) according to Tamhane test (net reproductive
rate) or Tukey test (generation time, intrinsic rate of increase); F-, df- and P-values refer to
one-way ANOVAs. ArF or AnF: freeze-dried artificial diet supplemented with A. franciscana
cysts or with pupal hemolymph of A. pernyi, respectively; ArP or AnP: basic powdered diet
supplemented with powdered A. franciscana cysts or lyophilized pupal hemolymph of A.
pernyi, respectively.
Differences in developmental and reproductive characteristics were reflected in life
table statistics. Net reproductive rates (Ro) of A. swirskii fed on T. latifolia pollen, or on the

Solid artificial diets for A. swirskii
91
lyophilized diets were significantly higher than those of predators fed on both powdered diets
(Table 6.3). Generation time (T) was significantly shorter for females offered T. latifolia
pollen versus the other diets. Finally, the intrinsic rate of increase (rm) of the predator was
highest on T. latifolia pollen, followed by the lyophilized diets, and lowest on the powdered
diets.
Figure 6.1 Influence of supplemental foods on the density of Amblyseius swirskii on caged
chrysanthemum plants in a greenhouse.
Data shown are means ± SE. Means at the same monitoring date followed by the same letter
are not significantly different (P > 0.05).
The diets, sampling times, as well as their interaction significantly affected the numbers of A.
swirskii per leaf (F(3,28) = 47.557, P<0.001; F(8,21) = 75.101, P<0.001; F(24,61.508) = 7.715,
P<0.001, respectively). The population densities of the predator on all diets were significantly
higher than those in the control at all sampling times (Figure 6.1). Mite populations increased
rapidly in all diet treatments. At week 2 and 4 after release, densities of mites did not differ
among diets but were higher than those in the control (χ2=19.029, df=3, P<0.001 and
F=11.366, df=3, 28, P<0.001, for week 2 and 4, respectively). The highest densities of A.
swirskii were recorded on ArP and the basic diet 12 weeks after release (1.9 and 1.3 mites/leaf,
respectively), whereas the highest number of mites on Nutrimite (1.2 mite/leaf) was observed
eight weeks after release.

Chapter 6
92
6.4 Discussion
Artificial diets for carnivorous arthropods can have a consistency ranging from liquid to solid,
depending mostly on two factors: water content and molecular cohesion (Morales-Ramos et
al., 2014). Artificial diets have been developed for a number of phytoseiid mites, with varying
success (McMurtry and Scriven, 1966; Kennett and Hamai, 1980; Ochieng et al., 1987; Abou-
Awad et al., 1992; Ogawa and Osakabe, 2008). All of these artificial diets were liquid in form.
Liquid diets are expected to be easily accessible to predatory mites and have the practical
advantage that they can be easily sprayed over the crop when used as a supplemental food. On
the other hand, solid artificial diets have a number of advantages over liquid and semiliquid
diets, both in the laboratory rearing and when applied as supplemental foods in the crop.
These include non-stickiness and the possibility of direct presentation without the need for
encapsulation (Morales-Ramos et al., 2014). Lower water content of solid diets limits
microbial contamination and allows easier long term storage. When applied as a supplemental
food in the field, solid artificial diets are easier to distribute in the crop (e.g. by blowers
designed to distribute pollen or eggs of Ephestia kuehniella (Zeller) (Lepidoptera: Pyralidae))
and are expected to result in lower soiling of the plant surface and thus to interfere less with
plant physiology and crop quality.
Previous studies demonstrated the potential of liquid artificial diets supplemented with
pupal hemolymph of A. pernyi or with a watery extract of A. franciscana cysts to support the
development and reproduction of A. swirskii (Chapter 3 and Chapter 5). The objective of the
present study was to develop an adequate solid diet as a food for the mass rearing or for
application in the field. Our results indicate that A. swirskii performed well, both in terms of
its development and reproduction, when exclusively fed on various powdered solid artificial
diets, although its performance did not match that on T. latifolia pollen.
Among the four artificial diets tested, the highest intrinsic rate of increase of A.
swirskii (0.195 females/female/day at 23 °C) was recorded on freeze-dried artificial diet AnF,
a meridic diet enriched with pupal hemolymph of A. pernyi. Although lower than on T.
latifolia pollen in the present study, this growth rate exceeds the values reported for A.
swirskii feeding on several natural prey, including the two spotted spider mite Tetranychus
urticae Koch (Acari: Tetranychidae) (0.167 at 26 oC) (El-Laithy and Fouly, 1992), the
western flower thrips Frankliniella occidentalis (Pergande) (0.056 at 25 oC) and onion thrips
Thrips tabaci (Lindeman) (Thysanoptera: Thripidae) (0.024 at 25 oC) (Wimmer et al., 2008),

Solid artificial diets for A. swirskii
93
and the eriophyoid fig mites Aceria ficus (Cotte) (0.155 at 29 oC) and Rhyncaphytoptus
ficifoliae Keifer (0.122 at 29 oC)(Abou-Awad et al., 1999); interestingly, the growth rate on
AnF is also superior to that on the dried fruit mite C. lactis, a factitious prey which is
routinely used in the mass production of A. swirskii (0.175 at 23 oC) (Chapter 3). Arguably,
caution is warranted when comparing absolute values of intrinsic growth rates among studies
as they may be influenced by differences in climatic conditions, experimental methods, and
calculation of estimates. Interestingly, intrinsic growth rates of A. swirskii in our study are
closely correlated with oviposition rates, with about 90% of the variation in growth rates
being explained by oviposition rates. This confirms the observations by Janssen and Sabelis
(1992), who suggested that an assessment of peak oviposition may be sufficient to estimate
the reproductive performance of predatory mites, precluding the necessity to perform full life
table studies.
The fecundity of females fed on the freeze-dried liquid artificial diets ArF and AnF
was similar to that on T. latifolia pollen and higher than that on the powdered diets ArP and
AnP, which were both composed of solid basic ingredients. Parameters of development and
reproduction of females fed on lyophilized diets were similar to those on the original liquid
versions of the diets as reported in Chapter 5, indicating that the freeze-drying process did
not influence the nutritional quality of the diets and that the predator handled the solid forms
as effectively as the liquid forms. This confirms previous findings with diets for predators and
plant bugs, which have also indicated that the nutritive qualities and palatability are retained
after freeze-drying and rehydration of certain insect diets (Cohen, 2004). Freeze-dried diets
have several practical advantages over liquid diets related to storage, shipment and handling.
For instance, a freeze-dried medium may be stored at room temperature for months with no
loss of nutritional quality (Cohen, 1999). The freeze dried diets used in our study proved to be
highly hygroscopic and quickly reverted to a semi-liquid form. This liquefaction process may
have been accelerated by the high relative humidity in the Munger cells, which was always in
excess of 85 %, and may thus be slower under practical conditions. On the one hand, the
partial liquefaction of these freeze dried diets may be beneficial as it may facilitate feeding by
the mites. On the other, however, it may limit their practical applicability: for instance, when
not replaced on a regular basis like in the present study, such diets may be more prone to
microbial degradation and may thus require the addition of antimicrobial agents. Further, the
freeze-drying process also increases the cost of the artificial diets.
The availability of an adequate artificial diet would preclude the necessity to maintain
parallel cultures of prey mites in the mass production system, resulting in less labour costs

Chapter 6
94
and lower health risks for workers in the production environment (i.e. potential allergy
problems related to the use of storage mites). Even when such artificial diets appear
suboptimal as compared to storage mites in terms of predator population growth rates, they
could be used in part of the production cycle of the phytoseiids. Most ingredients of the
presented diets are relatively inexpensive and easily accessible on the market. Based on
economies of scale, prices may further be reduced if these diets are produced at a larger scale.
In order to enhance cost effectiveness as compared to the liquid diets previously designed in
Chapter 5 and their lyophilized forms tested here, the powdered diets ArP and AnP were
created by replacing the liquid ingredients of the original formulations in Chapter 5 by
powdered solid ingredients. The honey was replaced with sucrose, glucose and fructose, fresh
egg yolk was replaced with spray-dried egg yolk, and a vitamin mix was added. However, A.
swirskii fed on the powdered diets had significantly longer developmental times and lower
oviposition rates and intrinsic rates of increase than those reared on the freeze dried diets.
These differences in performance of the mite can be explained in part by differences in the
nutritional profiles of the corresponding diets. For instance, honey contains other compounds
than sugars, like vitamins, minerals and amino acids, which may have a positive impact on the
development and reproduction of A. swirskii. Guardiola et al. (1995) found that fatty acids
undergo considerable oxidation during spray-drying of egg; therefore, the nutritive value of
egg yolk powder may be lower than that of fresh egg yolk. Further, Debolt (1982) reported
differences in nutritional and sensory qualities of spray-dried egg yolk as compared to fresh
egg yolk for the mirid Lygus hesperus Knight (Hemiptera: Miridae). Nonetheless,
reproductive rates of A. swirskii on the powdered diets in the present study were comparable
to the values found in Chapter 3 on the dried fruit mite C. lactis and higher than the values
reported earlier on some of the predator’s natural prey (Ragusa and Swirski, 1977; Wimmer et
al., 2008). Additionally, the powdered diets proved to be more structurally stable than the
freeze dried diets, which may enhance their practical value in rearing systems and as
supplemental foods in the crop.
In ornamental plant production, there is often a zero tolerance towards pest presence
for aesthetic reasons (Schumacher et al., 2006). In addition, the tolerance for thrips in
chrysanthemums is very low. The availability of effective supplemental food sources should
enable to maintain a population of predatory mites in the crop at sufficiently high levels to
keep the population of thrips at a minimum level (Hoogerbrugge et al., 2008). In our pre-
establishment greenhouse experiment, the value of the powdered basic artificial diet without
or with A. franciscana cyst powder (ArP) was examined on chrysanthemum plants. The

Solid artificial diets for A. swirskii
95
results indicated that both powdered diets could support a population of A. swirskii over a
period of several weeks. The population growth of the predator on the artificial diets was
similar to that on Nutrimite, which is commercially used as a supplemental food source for
phytoseiid mites in greenhouse crops. The diets did not only succeed in maintaining the
populations of A. swirskii on the plants but they also yielded positive population growth of the
mite. Although the numbers of mites per leaf only appear to increase gradually over time, in
reality their populations exploded as the plants sprouted many more leaves over the
successive weeks. Hoogerbrugge et al. (2008) also conducted a pre-establishment experiment
with A. swirskii in chrysanthemums using seven different food sources: cysts of the brine
shrimp Artemia sp., honey bee pollen, C. lactis, C. lactis + artificial diet A (not described),
artificial diet B (not described), artificial diet B + honey bee pollen, and eggs of Ephestia
kuehniella Zeller (Lepidoptera: Pyralidae) + fresh corn pollen. The initial number of the
predatory mites introduced in their experiment was 10 females/plant, which is approximately
30 times higher than that in the present study. The authors reported the highest number of
predators to be around 8.2 mites/leaf in the treatment with C. lactis + diet A after 28 days.
This means that, although comparison of both greenhouse studies is complicated by the
different experimental conditions, the populations of A. swirskii in our study in fact increased
faster on Nutrimite and ArP than on the best diet in the study by Hoogerbrugge et al. (2008),
given the much higher initial densities in the latter study. In addition, no damage to the
chrysanthemum leaves was observed after twelve weeks in all diet treatments of our study.
However, some fungal contamination was recorded on the leaves sprayed with the powdered
artificial diets, especially where the diet grains clumped.
In conclusion, A. swirskii was capable of handling and feeding on (powdered) solid
artificial diets, allowing the mite to develop and reproduce successfully for at least a single
generation. Freeze-drying of liquid diets did not influence their acceptability and nutritional
quality for the mite. Artificial diets entirely composed of powdered/dry ingredients were
somewhat inferior to lyophilized liquid diets in terms of their nutritional value, but did
support development and reproduction of the phytoseiid and were physically more stable in
the rearing environment than the lyophilized diets. A greenhouse experiment showed that
powdered artificial diets could indeed support populations of A. swirskii in a chrysanthemum
crop. These findings indicate the potential of dry artificial diets for use in part of the
production cycle of A. swirskii or as supplemental foods to sustain its populations in the crop
after release. Arguably, more studies are needed to assess the practical applicability of
artificial diets in support of biological control programmes using phytoseiid mites. First, it is

Chapter 6
96
warranted to assess the value of the presented artificial diets for supporting the population
growth of other generalist predatory mites. The effects of long-term rearing of the mites on
artificial diets on their quality as biological control agents also need to be explored. Further,
more field research is needed to evaluate the suitability of the tested diets in the crop
environment, with regard to their practical value to support populations of A. swirskii or of
other predatory mites, and possible undesired effects of their application on populations of
arthropod pests (like omnivorous thrips who may also be able to use the diets) and on the
physiology and quality of the crop plants.

97
Chapter 7
PERFORMANCE OF FOUR SPECIES OF PHYTOSEIID MITES ON ARTIFICIAL
AND NATURAL DIETS
This chapter is based on:
Nguyen, D. T., Vangansbeke, D., & De Clercq, P. 2014. Performance of four species of phytoseiid
mites on artificial and natural diets. Biological Control, 80: 56–62, DOI:
10.1016/j.biocontrol.2014.09.016

Chapter 7
98
7.1 Introduction
Phytoseiid predatory mites are important biocontrol agents of tetranychid mites and
small, soft-bodied insects like thrips and whiteflies (Chant, 1985). In augmentation biological
control, large numbers of predaceous mites are released in the field (Stinner, 1977; Collier
and Van Steenwyk, 2004). Hence, a cost-effective method for their mass-rearing is an
essential prerequisite (van Lenteren, 2003). Rearing phytoseiid mites on plant materials
infested with natural prey has several disadvantages, such as large space requirements,
inconsistent yields of predators, harvesting difficulties and variable results with different
species (McMurtry and Scriven, 1965). Rearing procedures based on factitious prey like
storage mites (Zhang, 2003; Bolckmans and van Houten, 2006) also involve space and labor
to maintain large parallel cultures of the factitious prey. Further, there may be health risks for
workers in production facilities or releases in the crop caused by allergens associated with the
factitious mite prey (Bolckmans and van Houten, 2006; Fernandez-Caldas et al., 2007). The
availability of an adequate artificial diet could eliminate many of the above-mentioned
problems associated with the mass production of predatory mites (Kennett and Hamai, 1980).
In addition, these artificial diets may be useful as food supplements to support predator
populations after release in the crop (Wade et al., 2008).
Several artificial diets have been developed for phytoseiid mites, but the results were
mostly inferior to those on natural or factitious prey. McMurtry and Scriven (1966) reported
longer developmental times and lower oviposition rates when four phytoseiids
(Amblydromalus limonicus Garman and McGregor, Amblyseius hibisci (Chant),
Typhlodromus occidentalis Nesbitt and Typhlodromus rickeri Chant) (Acari: Phytoseiidae)
were fed on various artificial diets compared with mite prey and pollen as food sources.
Shehata and Weismann (1972) tested three artificial diets for the specialist spider mite
predator Phytoseiulus persimilis Athias-Henriot. Their results indicated that the larvae could
develop to adults but the females failed to produce viable eggs. Kennett and Hamai (1980)
investigated oviposition rate and developmental capacity of 9 predaceous mites (A. hibisci, A.
limonicus, Amblyseius largoensis (Muma), Metaseiulus pomoides Schuster & Pritchard, T.
occidentalis, Typhloseiopsis arboreus (Chant), Typhloseiopsis pyri Scheuten, P. persimilis,
and Iphiseius degenerans (Berlese)) fed on artificial and natural diets. The authors reported
that complete development and oviposition occurred for seven out of nine species when fed
on an artificial diet consisting of bee honey, sugar, yeast flakes, yeast hydrolysate, enzymatic

Performance of four phytoseiid mites on artificial diets
99
casein hydrolysate and fresh egg yolk. Oviposition rates of all species fed on the artificial diet
were lower than those on a natural diet. Ochieng et al. (1987) reported that Amblyseius teke
Pritchard and Baker could complete more than 25 generations when reared on a liquid diet
composed of bee honey, milk powder, egg yolk and Wesson's salt. Abou-Awad et al. (1992)
noted that the predacious mites Amblyseius gossipi El-Badry and Amblyseius swirskii Athias-
Henriot developed and reproduced successfully on artificial diets composed of yeast, milk,
cysteine, proline, arginine, sucrose, glucose, streptomycin sulphate and sorbic acid. However,
fecundity of both species fed on the artificial diet was lower than on natural prey, although the
eggs showed no abnormalities. Shih et al. (1993) conducted experiments to investigate the
responses of Euseius ovalis (Evans) to natural food resources and two artificial diets.
Immature development was successful in the first generation but offspring was not able to
complete its life cycle when maintained on the artificial diets. The females of E. ovalis fed on
artificial diets showed a shorter oviposition period, lower daily and total reproductive rates,
and shorter longevity than those fed on natural diets. Ogawa and Osakabe (2008) investigated
the development and survival of Neoseiulus californicus (McGregor) on an artificial diet. The
phytoseiid successfully developed on the artificial diet, but only few eggs were deposited.
In our previous studies we found that artificial diets enriched with watery extract of
dry decapsulated cysts of the brine shrimp Artemia franciscana Kellogg (Anostraca:
Artemiidae) or pupal hemolymph of the Chinese oak silkworm (Antheraea pernyi (Guérin-
Méneville)) supported development and reproduction of the generalist predatory mite A.
swirskii. The females fed on these two artificial diets displayed higher intrinsic rates of
increase than those fed on several natural prey and performed as well as those reared on the
factitious prey Carpoglyphus lactis L. (Acari: Carpoglyphidae), which is routinely used in the
mass rearing of this phytoseiid (Chapter 3 and Chapter 5). The objectives of the present
study were to assess the suitability of the artificial diet enriched with A. franciscana as an
alternative food for several other economically important predatory mites by performing full
life table studies under controlled laboratory conditions. The phytoseiids selected for testing
belong to different types based on their level of food specialization (McMurtry et al., 2013): N.
californicus is a selective predator of tetranychid mites (type II), whereas A. andersoni Chant,
N. cucumeris (Oudemans), and A. limonicus are more generalist predators (type III).

Chapter 7
100
7.2 Materials and methods
7.2.1 Stock colonies of predatory mites
Laboratory cultures of N. cucumeris and A. limonicus were initiated with specimens
supplied by Koppert B.V. (Berkel en Rodenrijs, The Netherlands) and A. andersoni was
supplied by Biobest N.V. (Westerlo, Belgium). The mites were reared on green plastic arenas
(10 x 10 x 0.3 cm) (Multicel, SEDPA, France), placed on a wet sponge in a plastic tray
containing water. The edges of the arenas were covered with tissue paper immersed in the
water to provide moisture and deter the mites from escaping. Every two days the mites were
fed with fresh cattail pollen (Typha latifolia L.), which was also supplied by Koppert B.V. and
stored at -18°C. For the experiments, pollen was thawed and kept in a refrigerator at 5°C for
max. 1 week. A small piece of sewing thread was placed on the arenas to serve as an
oviposition substrate. Every two days the eggs were collected and transferred to new arenas.
A culture of N. californicus was initiated with mites acquired from Koppert B.V. and
was reared on kidney bean leaves heavily infested with two-spotted spider mites (Tetranychus
urticae Koch). The leaves were placed upside down on a layer of water-saturated cotton in a
glass petri dish ( 133 mm), with an extra cotton layer on the leaf edges to provide free water
and prevent the mites from escaping.
Predatory mites were cultured in a growth chamber set at 25 ± 1°C, 70 ± 5% RH and a
16:8 h (L: D) photoperiod.
7.2.2 Preparation of artificial diet
Artificial diets were prepared according to section 5.2.2 : 80% basic artificial diet
supplemented with 20% (w/w) watery extract of dry decapsulated A. franciscana cysts, which
were provided by the Artemia Reference Center of Ghent University (Ghent, Belgium) and
originated from the Great Salt Lake (Utah, USA). The basic artificial diet was prepared
according to section 3.2.4 . The diet was dispensed into 2ml Eppendorf tubes and stored at
−18°C.

Performance of four phytoseiid mites on artificial diets
101
7.2.3 Experimental setup
Eight hours before start of the experiments, new pieces of sewing thread were placed
in the stock colony of N. cucumeris, A. limonicus and A. andersoni or fifty females of N.
californicus were transferred to new bean leaves infested with mixed life stages of T. urticae.
Deposited eggs (less than 8 hours old) were transferred individually to the rearing microcosms
that were modified from Munger cells as described in section 3.2.5 . Development and
reproduction of the tested phytoseiids on the artificial diet was compared with that on a
natural food source (spider mites for N. californicus or cattail pollen for N. cucumeris, A.
andersoni and A. limonicus). All food sources were offered ad libitum from the larval stage of
the predator on and were refreshed every 2 days. For the artificial diet, approximately 2 µl of
diet was absorbed on a small piece of filter paper (2 x 2 mm) placed on the bottom board of
the rearing microcosms. The mites were observed and different parameters were recorded
same as that described in section 3.2.6 . Mites that escaped or died due to unnatural causes
were excluded from data analysis. Females that died before laying eggs were excluded from
calculation of reproductive parameters. The experiments were done in a growth chamber at 23
± 1°C, 65 ± 5% RH and a 16:8 h (L: D) photoperiod.
7.2.4 Life table parameters calculation
Life table parameters were calculated as explained in section 3.2.7
7.2.5 Statistical analysis
Two-way analysis of variance (ANOVA) (IBM, SPSS Statistics 20) was conducted to
evaluate the effects of diet and species on the duration of the immature stages, preoviposition
and oviposition period, daily and total oviposition, adult longevity and life table parameters.
When an interaction was detected between the main factors, means were compared among
species and a pairwise multiple comparison procedure was used (Kutner et al., 2005). When a
Kolmogorov–Smirnov test indicated that data were normally distributed, the pairwise
comparisons among diets in each species were analyzed using Student t-tests. When data were
not normally distributed, a nonparametric Mann-Whitney U test was used. Generalized linear
model with a probit (log odds) link function and a binomial error distribution was used to
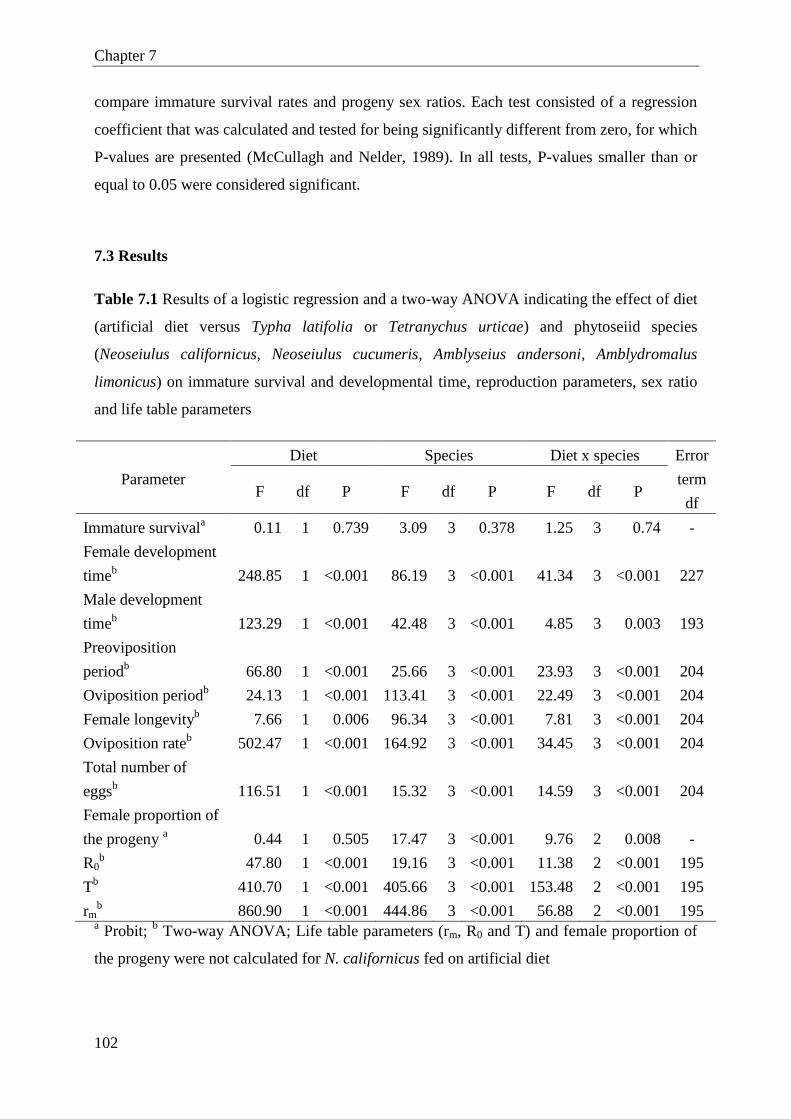
Chapter 7
102
compare immature survival rates and progeny sex ratios. Each test consisted of a regression
coefficient that was calculated and tested for being significantly different from zero, for which
P-values are presented (McCullagh and Nelder, 1989). In all tests, P-values smaller than or
equal to 0.05 were considered significant.
7.3 Results
Table 7.1 Results of a logistic regression and a two-way ANOVA indicating the effect of diet
(artificial diet versus Typha latifolia or Tetranychus urticae) and phytoseiid species
(Neoseiulus californicus, Neoseiulus cucumeris, Amblyseius andersoni, Amblydromalus
limonicus) on immature survival and developmental time, reproduction parameters, sex ratio
and life table parameters
Parameter
Diet Species Diet x species Error
term
df F df P F df P F df P
Immature survivala 0.11 1 0.739 3.09 3 0.378 1.25 3 0.74 -
Female development
timeb 248.85 1 <0.001 86.19 3 <0.001 41.34 3 <0.001 227
Male development
timeb 123.29 1 <0.001 42.48 3 <0.001 4.85 3 0.003 193
Preoviposition
periodb 66.80 1 <0.001 25.66 3 <0.001 23.93 3 <0.001 204
Oviposition periodb 24.13 1 <0.001 113.41 3 <0.001 22.49 3 <0.001 204
Female longevityb 7.66 1 0.006 96.34 3 <0.001 7.81 3 <0.001 204
Oviposition rateb 502.47 1 <0.001 164.92 3 <0.001 34.45 3 <0.001 204
Total number of
eggsb 116.51 1 <0.001 15.32 3 <0.001 14.59 3 <0.001 204
Female proportion of
the progeny a 0.44 1 0.505 17.47 3 <0.001 9.76 2 0.008 -
R0b 47.80 1 <0.001 19.16 3 <0.001 11.38 2 <0.001 195
Tb 410.70 1 <0.001 405.66 3 <0.001 153.48 2 <0.001 195
rmb 860.90 1 <0.001 444.86 3 <0.001 56.88 2 <0.001 195
a Probit;
b Two-way ANOVA; Life table parameters (rm, R0 and T) and female proportion of
the progeny were not calculated for N. californicus fed on artificial diet

Performance of four phytoseiid mites on artificial diets
103
Two-way ANOVA indicated no interaction between diet and species for immature
survival (Table 7.1), whereas interactions were found to be significant for all other parameters.
Diet had no influence on the immature survival rate of all tested species (Table 7.2). Female
developmental times were significantly shorter on natural diets (T. urticae or T. latifolia) than
on the artificial diet, except for A. limonicus. In all species, males developed faster on natural
diets than on the artificial diet.
Table 7.2 Immature survival and developmental time of different phytoseiid mites fed on
artificial diet versus natural food sources
Species Diet Proportion
surviving
Developmental duration (days)*
n Females N Males
N. californicus T. urticae 0.92 ± 0.04 a 32 5.09 ± 0.05 a 24 5.38 ± 0.18 a
Artificial 0.96 ± 0.03 a 20 7.90 ± 0.31 b 28 6.46 ± 0.15 b
χ2/ U 0.818 4.500 81.500
df/Z 1 -6.546 -4.979
P 0.366 <0.001 <0.001
N. cucumeris T. latifolia 0.98 ± 0.02 a 33 6.67 ± 0.09 a 22 6.45 ± 0.11 a
Artificial 0.98 ± 0.02 a 29 9.00 ± 0.21 b 24 8.25 ± 0.11 b
χ2/ U 0.001 0.000 5.000
df/Z 1 -6.988 -5.927
P 0.979 <0.001 <0.001
A. andersoni T. latifolia 0.96 ± 0.03 a 27 5.74 ± 0.10 a 26 5.81 ± 0.12 a
Artificial 0.98 ± 0.02 a 34 6.79 ± 0.12 b 20 6.55 ± 0.14 b
χ2/ U 0.338 118.000 113.500
df/Z 1 -5.387 -3.523
P 0.561 <0.001 <0.001
A. limonicus T. latifolia 0.98 ± 0.02 a 30 5.77 ± 0.11 a 30 5.50 ± 0.17 a
Artificial 0.97 ± 0.02 a 30 5.77 ± 0.09 a 27 6.41 ± 0.12 b
χ2/ U 0.371 443.000 150.500
df/Z 1 -0.127 -4.304
P 0.542 0.899 <0.001
n and N: Number of tested female and male individuals, respectively
*Means ± SE; means within a column and a species followed by the same letter are not
significantly different (P>0.05), according to Probit (Wald Chi-square) test (Immature
survival), or Mann-Whitney U test (Female and male developmental time). χ2-, df- and P-
values refer to Probit (Wald Chi-square) test, U-, Z-, and P- values refer to Mann-Whitney U
test.

Chapter 7
104
Preoviposition periods of A. limonicus females fed on T. latifolia and the artificial diet
were similar, whereas females of the other species fed on their respective natural diets had
significantly shorter preoviposition periods than those fed on the artificial diet (Table 7.3).
Oviposition period of N. californicus did not differ among diets, whereas in A. limonicus
females reared on artificial diet had a shorter oviposition period than those reared on cattail
pollen. In contrast, N. cucumeris and A. andersoni fed on the artificial diet had longer
oviposition periods than those reared on cattail pollen. Whereas no difference in female
longevity was observed among diets for N. californicus, females reared on artificial diet lived
shorter in A. limonicus and A. andersoni and longer in N. cucumeris, than their counterparts
fed on pollen. The total number of eggs deposited by N. californicus, N. cucumeris and A.
limonicus females reared on natural diets was significantly higher than that of females
maintained on the artificial diet but fecundity of A. andersoni was not affected by diet.
Whereas the proportions of female offspring in A. andersoni and A. limonicus were similar on
artificial and natural diets, in N. cucumeris a lower percentage of female offspring was
observed on the artificial diet than on cattail pollen. Because the offspring of N. californicus
fed on the artificial diet did not develop successfully to adulthood, the female proportion of
the progeny and life table parameters were not calculated for this treatment.

105
Table 7.3 Reproductive parameters of several phytoseiid mites fed on artificial diet versus natural food sources
Species Diets n Preoviposition
period (days)a
Oviposition
period (days)a
Female
longevity
(days)a
Oviposition rate
(eggs/female/day)a
Total number
of eggs
(eggs/female)a
Female
proportion of
the progeny a
N. californicus T. urticae 32 2.22 ± 0.10 a 19.44 ± 0.70 a 34.25 ± 2.40 a 2.76 ± 0.07 a 53.34 ± 2.05 a 0.63 ± 0.02
Artificial 10 14.30 ± 4.84 b 15.50 ± 3.40 a 38.00 ± 6.10 a 0.85 ± 0.09 b 12.90 ± 3.41 b -
U/ t/ χ2 1.000 1.135 -0.688 15.035 9.763
Z/df -5.482 9.772 40 40 40
P <0.001 0.283 0.495 <0.001 <0.001
N. cucumeris T. latifolia 33 2.52 ± 0.11 a 30.97 ± 1.43 a 52.64 ± 3.37 a 1.51 ± 0.06 a 45.45 ± 2.26 a 0.73 ± 0.02 a
Artificial 27 4.00 ± 0.16 b 50.52 ± 3.27 b 71.56 ± 4.22 b 0.52 ± 0.02 b 25.78 ± 1.68 b 0.65 ± 0.02 b
U/ t/ χ2 88.500 -5.475 -3.550 14.989 7.003 9.226
Z/df -5.531 35.747 58 38.894 56.053 1
P <0.001 <0.001 0.001 <0.001 <0.001 0.002
A. andersoni T. latifolia 27 2.26 ± 0.11 a 33.07 ± 2.15 a 55.70 ± 3.30 a 1.49 ± 0.07 a 48.56 ± 3.44 a 0.70 ± 0.01 a
Artificial 32 4.34 ± 0.18 b 50.91 ± 2.02 b 68.16 ± 2.62 b 0.92 ± 0.03 b 45.97 ± 2.10 a 0.71 ± 0.01 a
U/ t/ χ2 37.000 -6.034 -2.993 7.736 0.642 0.308
Z/df -6.189 57 57 40.001 43.942 1
P <0.001 <0.001 0.004 <0.001 0.524 0.579
A. limonicus T. latifolia 30 1.73 ± 0.08 a 16.40 ± 0.99 a 22.37 ± 1.83 a 2.57 ± 0.06 a 42.20 ± 2.81 a 0.65 ± 0.02 a
Artificial 21 1.90 ± 0.07 a 11.38 ± 0.84 b 13.10 ± 0.86 b 1.90 ± 0.08 b 22.05 ± 2.40 b 0.69 ± 0.03 a
U/ t/ χ2 261.000 3.853 4.583 6.458 5.447 0.971
Z/df -1.503 48.985 40.254 49 48.964 1
P 0.133 <0.001 <0.001 <0.001 <0.001 0.324
n: Number of tested females; aMeans ± SE; means within a column and a species followed by the same letter are not significantly different (P > 0.05)
according to Mann-Whitney U test (preoviposition period), Student’s t-test (oviposition period, female longevity, oviposition rate, total number of eggs) or
Probit (Wald Chi-square) test (female proportion of progeny). U-, Z-, and P- values refer to Mann-Whitney U test; t-, df- and P-values refer to Student’s t- test;
χ2-, df- and P-values refer to Probit (Wald Chi-square) test.

Chapter 7
106
Net reproductive rate (R0) of N. cucumeris and A. limonicus females offered pollen
was significantly higher than that of females given the artificial diet, whereas in A. andersoni
this parameter did not differ among diets. Females reared on pollen had a shorter generation
time than those on artificial diet for N. cucumeris and A. andersoni, whereas there were no
differences for A. limonicus. For all species, a higher intrinsic rate of increase (rm) was
calculated when the mites were maintained on their respective natural foods as compared to
the artificial diet.
Table 7.4 Life table parameters of different phytoseiid mites fed on artificial diet versus
natural food sources
Species Diets n
Net reproductive
rate (R0, females
per female)a
Generation
time (T,
days)a
Intrinsic rate of
increase (rm,
females/female/day)a
N. californicus T. urticae 32 29.45 ± 1.18 14.21 ± 0.17 0.238 ± 0.002
Artificial - - - -
N. cucumeris T. latifolia 33 32.97 ± 1.74 a 18.95 ± 0.27 a 0.185 ± 0.003 a
Artificial 27 17.16 ± 1.04 b 31.74 ± 0.71 b 0.090 ± 0.002 b
t 7.804 -16.850 24.357
df 51.076 33.386 58
P <0.001 <0.001 <0.001
A. andersoni T. latifolia 27 33.74 ± 2.62 a 18.03 ± 0.34 a 0.195 ± 0.002 a
Artificial 32 33.16 ± 1.82 a 24.29 ± 0.37 b 0.144 ± 0.003 b
t 0.185 -12.429 13.653
df 47.776 57 57
P 0.854 <0.001 <0.001
A. limonicus T. latifolia 30 26.96 ± 1.67 a 12.79 ± 0.19 a 0.258 ± 0.002 a
Artificial 21 13.83 ± 1.21 b 12.39 ± 0.27 a 0.212 ± 0.003 b
t 6.385 1.248 12.533
df 48.169 49 49
P <0.001 0.218 <0.001
n: Number of tested females
aMeans ± SE; means within a column and a species followed by the same letter are not
significantly different (P > 0.05) according to Student’s t- test

Performance of four phytoseiid mites on artificial diets
107
7.4 Discussion
The developmental and reproductive performances of the four predatory mites studied
were differentially affected by diet. Where differences were observed, the mites developed
faster and produced more eggs on their natural diets (spider mites for N. californicus or cattail
pollen for N. cucumeris, A. andersoni and A. limonicus) than on the artificial diet. Overall, the
percentage decrease in developmental rate, oviposition rate and intrinsic rate of increase of
the predatory mites fed on artificial diet as compared with natural diet was greatest in N.
californicus, intermediate in N. cucumeris and smallest in A. andersoni and A. limonicus. This
order reflects the level of polyphagy of the different species. N. californicus is classified as a
selective predator of tetranychid mites (McMurtry and Croft, 1997), which may explain its
relatively poor performance on the artificial diet. The more polyphagous species A. andersoni
and A. limonicus, which readily feed and reproduce also on pollens (Duso and Camporese,
1991; van Houten et al., 1995), were also able to develop and reproduce well on the artificial
diet. Our findings are consistent with those reported by McMurtry and Scriven (1966) who
tested several artificial diets for four phytoseiid mites. In the latter study, the studied type II
phytoseiids of the genus Typhlodromus (T. occidentalis and T. rickeri) did not perform well
on the tested artificial diets, whereas better results were obtained for two Amblyseius species
(A. limonicus and A. hibisci) characterized by a more generalist feeding habit.
The artificial diet in the present study supported development of N. californicus well,
with 96% of the mites reaching adulthood. Immature development of females fed on the
artificial diet was longer than that on T. urticae but was similar to that reported by Rencken
and Pringle (1998) and Kim et al. (2009) when the predatory mite was reared at 25°C with
mixed stages of T. urticae. N. californicus fed on the artificial diet at 23°C in our study had
similar developmental rates to those of the predator reared at 25°C on an artificial diet
formulated by Ogawa and Osakabe (2008). However, whereas only few eggs were produced
on the diet used by Ogawa and Osakabe (2008), N. californicus females in the present study
produced 0.85 eggs/day on a similar diet supplemented with A. franciscana. Nonetheless, egg
production on the artificial diet was substantially lower than on the natural prey, T. urticae,
and offspring could not develop successfully to adulthood.
To our knowledge, this is the first time that an artificial diet was tested for N.
cucumeris. The predatory mite developed successfully on the artificial diet, with an immature
survival of 98% and produced viable eggs, the offspring of which succeeded in reaching
adulthood (> 75% survival). A positive intrinsic rate of increase of 0.09 females/female/day-1

Chapter 7
108
was calculated on the artificial diet, showing that N. cucumeris is still able to maintain its
population when exclusively fed on the artificial diet. Thus, whereas the diet is not suitable
for the mass production of this phytoseiid, it still may have some potential to be used as a
food supplement to sustain populations of N. cucumeris in the crop (Chapter 6). Our results
also show the suitability of T. latifolia pollen as a supplemental food for N. cucumeris. The
intrinsic rate of increase of N. cucumeris fed on T. latifolia pollen (0.185 females/female/day)
is higher than the values reported for the phytoseiid when fed on pollen of apple, birch,
Christmas cactus, horse-chestnut, maize, or tulip (0.149, 0.127, 0.155, 0.180, 0.101 or 0.167
females/female/day, respectively) at 25°C (Ranabhat et al., 2014), or on castor pollen and T.
urticae (0.179 and 0.147 females/female/day, respectively) at 25 °C (van Rijn and Tanigoshi,
1999b). The rm value of N. cucumeris on T. latifolia pollen calculated in our study was
slightly lower than the value of 0.208 reported by van Rijn and Tanigoshi (1999b) when the
predator was offered pollen of broad bean (Vicia faba L.), but this difference may in part be
attributed to the higher experimental temperature (25°C) in the latter study.
Amblyseius andersoni females reared on the artificial diet produced less eggs per day
than those given T. latifolia pollen. However, the artificial diet prolonged the oviposition
period, so that an equal amount of eggs was produced on both diets. The life time fecundity of
A. andersoni females reared on T. latifolia and the artificial diet (48.56 and 45.97 eggs/female,
respectively) in this study is substantially higher than the total fecundities reported by
Lorenzon et al. (2012) for females of the species fed on T. latifolia pollen, Eotetranychus
carpini (Oudemans), Panonychus ulmi (Koch), Colomerus vitis (Pagenstecher), or the
mycelium of grape downy mildew Plasmopara viticola (Berk. & Curt.) Berl. (25.7, 24.2, 22.2,
25.6, and 9.3 eggs/female, respectively).
The artificial diet fully supported the development and reproduction of A.limonicus.
Developmental duration of females was similar on T. latifolia pollen and the artificial diet
(5.8 days). Development was shorter than that observed on dry decapsulated cysts of A.
franciscana and E. kuehniella eggs at the same temperature (6.58 and 6.85 days, respectively)
(Vangansbeke et al., 2014). The oviposition rate of A. limonicus on the present artificial diet
(1.90 eggs/female/day) was higher than that on the artificial diet designed by Kennett and
Hamai (1980) (1.22 eggs/female/day) and on the diets (sucrose, molasses, yeast + sucrose and
yeast + molasses) used by McMurtry and Scriven (1966) (0.11, 0.25, 1.13 and 1.18
egg/female/day, respectively).
The use of food supplements, consisting of alternative or artificial foods, can increase
the abundance and impact of arthropod natural enemies in crops where target prey or plant

Performance of four phytoseiid mites on artificial diets
109
foods like pollen and nectar are absent or only present at low densities (Wade et al., 2008).
While various supplemental foods appear to have potential for enhancing establishment of
insect natural enemies, only few have been reported to support beneficial Acarina (Wade et al.,
2008). For generalist predatory mites, pollen has been reported to be an adequate
supplemental food source to enhance the biological control of thrips and whiteflies on
cucumber (van Rijn et al., 2002; Nomikou et al., 2010). Typha angustifolia L. pollen
(NutrimiteTM
, Biobest NV) is commercially available to support pollen-feeding phytoseiids in
the crop. Drawbacks associated with the application of pollen are the relatively high costs
(Messelink et al., 2014) and the potential beneficial effects on pollen-feeding pests, such as
Frankliniella occidentalis Pergande (Ramakers, 1995). Certain animal foods also have
potential to support populations of phytoseiid mites, such as sterilized eggs of Ephestia
kuehniella Zeller and decapsulated cysts of A. franciscana (Vangansbeke et al., 2014).
Finding inexpensive alternative food sources is one of the major opportunities for enhancing
biological control in protected crops (Messelink et al., 2014). Several artificial diets have been
developed with the aim to reduce the production cost of phytoseiid mites or provide
supplemental foods for supporting their populations after release in the crop. For example, a
simple mixture of yeast, sugar and protein applied on chrysanthemum increased population
levels of A. swirskii (Messelink et al., 2009). In Chapter 3 and Chapter 5, artificial diets
enriched with A. franciscana or A. pernyi were found to support the development and
production of this economically important species. The findings of the present study suggest
that the same diet may also be useful to support populations of other commercially available
phytoseiids, either in the laboratory or in the field. Future studies should focus on the
optimization of artificial diets for phytoseiids in terms of their nutritional value and
formulation. In addition, when evaluating an artificial diet for field support, researchers may
not only limit their focus on promoting the predator, but also diminishing potential beneficial
effects for omnivorous pests. More work is also needed to optimize the application of
artificial diets as food sprays in the crop. Possible limitations of the use of artificial diets as
food sprays include desiccation, and chemical or microbial degradation of the foods, and crop
pollution or damage.

Chapter 7
110

111
Chapter 8
GENERAL DISCUSSION, CONCLUSIONS AND FUTURE PERSPECTIVES

Chapter 8
112
The potential for using augmentative biological control to suppress arthropod pests has been
recognized for many years (Doutt and Hagen, 1949; DeBach, 1964; Parella et al., 1992). This
method has a number of important benefits when compared with chemical control including
lower risks for the environment, lack of phytotoxic effects on the crop, and safety to
producers and consumers, allowing harvesting immediately after release of natural enemies,
unlike most chemical pesticides (van Lenteren, 2003). To date, however, augmentation
biological control is applied on a commercial scale in relatively few agricultural systems (van
Lenteren, 2012). One of the main reasons of the relatively low adoption of this pest
management strategy (van Lenteren, 2012) is that augmentative releases are usually more
expensive to growers than chemical pesticides (Collier and Van Steenwyk, 2004). One
approach to make augmentation a more competitive strategy is reducing the costs associated
with the mass production of natural enemies by using factitious or artificial foods instead of
their natural prey or hosts (Riddick, 2009).
The predatory mite Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) is an
economically important biological control agent of several key pests in greenhouses, such as
whiteflies, thrips and broad mites (Nomikou et al., 2002; Messelink et al., 2006; van Maanen
et al., 2010). The predator was first used in 2005 and is one of the 25 species that make up
more than 90% of the total market value of commercial biological control; it is currently used
in over 20 countries (Cock et al., 2010). Amblyseius swirskii is a type III generalist predator,
implying that it can feed on various types of food including various arthropod prey, pollen,
and honeydew (McMurtry et al., 2013). In the commercial mass production, A. swirskii is
being reared on storage mites, including the dried fruit mite, Carpoglyphus lactis L. (Acari:
Carpoglyphidae), and Thyreophagus entomophagus (Laboulbene) (Acari: Acaridae)
(Bolckmans and van Houten, 2006; Fidgett and Stinson, 2008).
Amblyseius swirskii can develop and reproduce well on pollens, a food source that is
rich in proteins, free amino acids, carbohydrates, lipids, vitamins, flavonoids, and minerals
(Goleva and Zebitz, 2013). The intrinsic rates of increase of the predator reared on pollens
like Typha latifolia L. or Zea mays L., as reported in Chapter 3 and by Zannou and Hanna
(2011), respectively, are higher than those observed on several of its natural prey, including
Frankliniella occidentalis (Pergande), Thrips tabaci Lind.(Wimmer et al., 2008), Bemisia
tabaci (Gennadius) (Nomikou et al., 2001), and Polyphagotarsonemus latus (Banks) (Onzo et
al., 2012). Cattail (T. latifolia) pollen has become a standard food in many laboratory studies
on phytoseiids (Nomikou et al., 2002; Park et al., 2011). However, in our study we noted that
the biological parameters of A. swirskii on T. latifolia pollen were variable depending on the

General discussion, conclusions and future perspectives
113
quality of the pollen batch used. We observed a substantial difference in rm-values (0.158 and
0.210, respectively)(Table 8.1) between the experiments in Chapter 3 and Chapter 6, using
the same environmental conditions and the same population of A. swirskii but different
batches of T. latifolia pollen. The quality of a pollen batch is influenced by harvesting time
(immature pollen grains may have a lower nutrient content), by weather conditions at the time
of pollen harvest (Coates et al., 2006), and by the duration and conditions of storage and
shipment (Bogdanov, 2004). Sufficient quantities of pollen needed for mass rearing predatory
mites may require large greenhouse areas for the production of plants, such as castor bean
(Ricinus communis L.). Alternatively, pollen can be collected in the field, like in the case of
cattail (Typha spp.). Collecting plant pollen outdoors is very labour intensive, and thus
expensive, and often only limited quantities can be collected (Bolckmans and Van Houten,
2011). Variability in quality (e.g., in terms of nutritional value or the presence of pesticide
residues or pathogenic micro-organisms), high costs and problems with continuity may
complicate the use of field collected pollens in the commercial rearing of A. swirskii or other
phytoseiids.
In the mass production of generalist phytoseiids, astigmatid mites are being used as a
primary food source (Zhang, 2003; Bolckmans and van Houten, 2006; Fidgett and Stinson,
2008; Midthassel et al., 2013). Amblyseius swirskii can be mass reared on the astigmatid mites
Suidasia medanensis (Oudemans) (Acari: Suidasidae) (Midthassel et al., 2013), Carpoglyphus
lactis L. (Acari: Carpoglyphidae) (Bolckmans and van Houten, 2006) and Thyreophagus
entomophagus (Laboulbene) (Acari: Acaridae) (Fidgett and Stinson, 2008). In Chapter 3, we
found that A. swirskii reared on C. lactis developed faster than on T. latifolia pollen whereas
the reproduction of the females fed on either food sources was not different (Table 8.1).
Midthassel et al. (2013) reported that the intrinsic rate of increase of A. swirskii fed on S.
medanensis was similar to or higher than on several of its natural arthropod prey. These
findings confirm the potential of these storage mites as a basic food for the mass production
of A. swirskii. Furthermore, the use of astigmatid mites can increase the economic efficacy of
commercial production systems as it allows the predator to be reared at high densities
(Ramakers et al., 1989; Mégevand et al., 1993). On the negative side, however, the use of
astigmatid prey mites may lead to health problems for workers in mass production facilities
caused by allergens originating from these mites (Fernandez-Caldas et al., 2007).

Chapter 8
114
Table 8.1 Intrinsic rate of increase (rm; means ± SE) of Amblyseius swirskii fed on different
artificial and factitious foods tested in the present study
Chapter Treatment Food tested Intrinsic rate
of increase
3 T. latifolia Typha latifolia L. pollen 0.158 ± 0.002
3 C. lactis Carpoglyphus lactis L. mix stages 0.175 ± 0.002
3 AD1 Liquid basic diet 0.104 ± 0.013
3 AD2 Liquid basic diet + 20% of A. pernyi pupal
hemolymph
0.181 ± 0.002
4 AD5 Liquid basic diet + 5% of H. illucens prepupal
hemolymph
0.182 ± 0.002
4 AD10 Liquid basic diet + 10% of H. illucens prepupal
hemolymph
0.189 ± 0.002
4 AD20 Liquid basic diet + 20% of H. illucens prepupal
hemolymph
0.210 ± 0.002
5 E. kuehniella/G1 Ephestia kuehniella eggs 0.169 ± 0.001
5 E. kuehniella/G6 Ephestia kuehniella eggs 0.151 ± 0.004
5 A. franciscana/G1 Decapsulated Artemia franciscana cysts 0.176 ± 0.002
5 A. franciscana/G6 Decapsulated Artemia franciscana cysts 0.182 ± 0.001
5 AD1/G1 Liquid basic diet + 20% of A. pernyi pupal hemolymph 0.174 ± 0.002
5 AD2/G1 Liquid basic diet + 20% of E. kuehniella eggs 0.160 ± 0.004
5 AD2/G6 Liquid basic diet + 20% of E. kuehniella eggs 0.073 ± 0.007
5 AD3/G1 Liquid basic diet + 20% of A. franciscana cysts 0.186 ± 0.001
5 AD3/G6 Liquid basic diet + 20% of A. franciscana cysts 0.144 ± 0.002
6 T. latifolia Typha latifolia L. pollen 0.210 ± 0.001
6 ArF Freeze-dried basic diet + 20% of A. franciscana cysts 0.184 ± 0.002
6 AnF Freeze-dried basic diet + 20% of A. pernyi pupal
hemolymph 0.195 ± 0.001
6 ArP Basic powdered diet + 20% of powdered A.
franciscana cysts 0.159 ± 0.001
6 AnP Freeze-dried basic diet + 20% of lyophilized A.
pernyi pupal hemolymph 0.158 ± 0.002
G1, G2: The first and sixth generation of rearing

General discussion, conclusions and future perspectives
115
The present study indicated that eggs of Ephestia kuehniella Zeller (Lepidoptera:
Pyralidae), a factitious food used in the mass production of various insect predators and egg
parasitoids, are also a suitable food source for A. swirskii (Chapter 5). This is consistent with
previous results of Romeih et al. (2004), who noted that A. swirskii showed a better response
to flour moth eggs than Neoseiulus californicus (McGregor), Euseius scutalis (Athias-Henriot)
(Acari: Phytoseiidae), and Agistemus exsertus Gonzalez (Acari: Stigmaeidae). The eggs also
proved to be an adequate food source for many other phytoseiid predators including
Amblydromalus limonicus Garman & McGregor (Vangansbeke et al., 2014), Iphiseius
degenerans (Berlese) (Vantornhout et al., 2004), Neoseiulus barkeri (Hughes) and Amblyseius
zaheri Yousef & El-Borolossy (Momen and El-Laithy, 2007), Euseius scutalis (Athias-
Henriot) and Agistemus exsertus Gonz. (Romeih et al., 2004)(Acari: Phytoseiidae). In most of
these studies the E. kuehniella eggs even surpassed the predators’ natural prey in terms of
developmental rate or survival. The good success obtained in several predatory mites with
these lepidopteran eggs could be related to their well-balanced nutritional composition and
easy handling by the mites. The disadvantage of E. kuehniella eggs is that they are very
expensive (with current market prices around 400-500 EUR/kg), mainly due to necessary
investments in climatisation of production facilities and health care of workers (De Clercq et
al., 2014). Furthermore, market prices for these lepidopteran eggs remain high because of a
continuously high demand for these eggs for the mass rearing of other natural enemies like
mirids, anthocorids, chrysopids, coccinellids, and trichogrammatid egg parasitoids in
commercial and research insectaries worldwide.
The value of another factitious food source, dry decapsulated Artemia franciscana
Kellogg (Anostraca: Artemiidae) cysts, was investigated in Chapter 5. The price of non-
decapsulated cysts may vary in the range 50-200 EUR/kg, depending on quality criteria such
as hatching rate, nutritional value, cyst diameter, and further according to annual fluctuations
of offer and demand. Also decapsulated cysts are offered on the market; for this purpose,
generally lower quality non-decapsulated cysts used, which have limited hatchability and thus
a lower market value for use in aquaculture (G. Van Stappen, ARC, Ghent University,
personal communication). Our experiments indicated that the high quality cysts we obtained
from the ARC of Ghent University were a highly suitable diet for the production of A. swirskii.
The mites fed on this diet developed faster than on E. kuehniella eggs and it was the only of
five tested diets not resulting in a decrease of growth rates of A. swirskii after 6 generations of
continuous rearing. Likewise, Vantornhout et al. (2004) and Vangansbeke et al. (2014) found
that dry decapsulated Artemia cysts were an acceptable food source for two other phytoseiids,

Chapter 8
116
I. degenerans and A. limonicus. In contrast, however, Hoogerbrugge et al. (2008) found only a
slightly higher population growth of A. swirskii when decapsulated Artemia sp. cysts were
supplemented in a chrysanthemum crop compared to a control treatment with no food
supplementation. The poor results with brine shrimp cysts in the latter study may be due to
lower nutritional quality of the Artemia batch used, or to an inappropriate level of hydration
or an incomplete decapsulation of the Artemia cysts. Decapsulation of brine shrimp cysts is
done by exposing the encapsulated cysts to a hypochlorite solution (Van Stappen, 1996).
Vandekerkhove et al. (2009) noted that a longer decapsulation time of A. franciscana cysts
had a positive influence on the development of the predatory bug Macrolophus pygmaeus
Rambur (Hemiptera: Miridae). Besides complete removal of the cyst shell, longer
decapsulation times may also cause damage to the hatching membrane, and as such make the
embryo more easily available for feeding predators. Given the short chelicerae of phytoseiid
mites, the decapsulation process is probably of primary importance. Vantornhout et al. (2004)
found that I. degenerans fed on encapsulated Artemia cysts failed to develop beyond the
protonymphal stage. The lack of development may be explained by the fact that I. degenerans
was not able to pierce the alveolar layer of the Artemia cysts. Further, it deserves pointing out
that hypochlorite residues from the decapsulation process may interfere with the acceptability
of decapsulated cysts for arthropod predators. The level of hydration of the cysts has also
been found to affect the immature survival and development of the predators. Vandekerkhove
et al. (2009) indicated that hydration of Artemia cysts had a significant impact on nymphal
survival of M. pygmaeus when cysts were non-decapsulated or poorly decapsulated. Arijs and
De Clercq (2001) showed that Orius laevigatus (Fieber) (Heteroptera: Anthocoridae) cannot
efficiently use dry cysts (i.e., containing less than 10% water) as food; the bugs can only
successfully develop on fully hydrated cysts. In our study, the humidity levels in the Munger
cells exceeded 80 % allowing the dry cysts to absorb some moisture; this may have facilitated
efficient extraction of the nutrients by the mites. This may have relevance not only for the use
of decapsulated Artemia cysts in a mass rearing environment, but also when used as
supplemental foods to sustain populations of predatory mites (or insects, like Macrolophus
bugs) in a greenhouse crop. The relative humidity inside greenhouses is often lower than that
observed in our Munger cells and usually fluctuates (low during the daytime and high during
nighttime). As a result, when the cysts are applied in a greenhouse crop they may experience
cycles of alternating hydration and dehydration, which may affect the feeding efficacy of
predatory mites mites on the cysts. More research is needed to optimize presentation (e.g.

General discussion, conclusions and future perspectives
117
level of decapsulation or hydration) of Artemia cysts when used in large-scale mite cultures or
as a supplemental food source for supporting their populations in the crop after release.
The availability of an effective artificial diet may allow rapid build-up of arthropod
parasitoids or predators populations, shorten the production line and consequently represent a
step towards a more cost-effective mass rearing (Riddick, 2009). Several artificial diets for
phytoseiid mites have been described by McMurtry and Scriven (1966), Shehata and
Weismann (1972), Kennett and Hamai (1980), Abou-Awad et al. (1992), and Ogawa and
Osakabe (2008). These studies suggest that, although phytoseiid mites can develop on
different artificial diets, fecundity in most cases was inferior to that on natural or factitious
prey. In Chapter 3, we noted that a slightly modified version of the artificial diet developed
by Ogawa and Osakabe (2008) was suitable to support development of A. swirskii and also
allowed some reproduction. The oviposition rate of A. swirskii on this artificial diet was
higher than that on an artificial diet designed by Abou-Awad et al. (1992). However, this diet
still has limitations in terms of fecundity, with 60% of the females failing to lay eggs and
dying shortly after molting to the adult stage. To enhance acceptability and improve
nutritional quality of artificial diets for entomophagous insects, the inclusion of arthropod
components such as hemolymph proved to be an important element (Nettles, 1990; Grenier
and De Clercq, 2003). The inclusion of insect components in artificial diets may be useful
when predators require certain nutrients, growth factors or phagostimulants found in their
natural prey (De Clercq, 2008). Our study in Chapter 3 showed that the pupal hemolymph of
the Chinese oak silkworm Antheraea pernyi Guérin-Méneville (Lepidoptera: Saturniidae)
played a positive role both in fecundity and survival of the predator. The oviposition rate and
intrinsic rate of increase of A. swirskii fed on basic diet enriched with 20% pupal hemolymph
of A. pernyi were significantly increased in comparison with the basic meridic diet. The
growth rate of A. swirskii reared on artificial diet enriched with the pupal hemolymph was
even higher than the values reported for A. swirskii when fed on natural prey like the two
spotted spider mite, western flower thrips, onion thrips, and eriophyoid fig mites (El-Laithy
and Fouly, 1992; Abou-Awad et al., 1999; Wimmer et al., 2008). Our study also showed that
the artificial diet with pupal hemolymph of A. pernyi supported development and reproduction
of A. swirskii to the same extent as the factitious prey mite C. lactis, which is routinely used in
the mass rearing of the phytoseiid, indicating the potential of an artificial diet to rationalize
the mass production of A. swirskii.
Lü et al. (2013) pointed out that the production of the oak silkworm in China has
diminished in recent years. Pupal hemolymph of this silkworm is therefore becoming less

Chapter 8
118
easily available and more costly, making its use as a dietary supplement for arthropod
parasitoids (e.g. Trichogramma spp.) and predators less attractive. In contrast, eggs of E.
kuehniella and cysts of A. franciscana are widely available. In Chapter 5, growth rates of A.
swirskii on artificial diets enriched with finely ground E. kuehniella eggs or A. franciscana
cysts were similar to or higher than those on the diet supplemented with pupal hemolymph of
A. pernyi. Our findings suggest that certain components of E. kuehniella eggs and A.
franciscana cysts can play a critical role in stimulating development and reproduction of A.
swirskii. Further research is needed to elucidate which factors are responsible for this positive
effect. Dependence on lepidopteran eggs or high quality brine shrimps cysts may, however,
complicate a reliable and cost effective artificial production of the phytoseiid. To further
reduce the cost of the artificial diet, different easily available and cheap arthropod
components were screened in the course of our study. In a preliminary experiment, we found
that the basic artificial diet supplemented with prepupal hemolymph of the black soldier fly
Hermetia illucens (L.) (Diptera: Stratiomyidae) or extract of larvae of the yellow mealworm
Tenebrio molitor L. (Coleoptera: Tenebrionidae) resulted in successful development and
reproduction of A. swirskii. However, the black soldier fly has a shorter development cycle,
higher oviposition rate and better potential of biomass increase per day as compared to the
yellow mealworm. Additionally, the fly can be cost effectively produced at a large scale on
organic waste materials. The results in Chapter 4 indicate that supplementing artificial diet
with black soldier fly hemolymph significantly improved its value for supporting
development and reproduction of A. swirskii. Even at the lowest concentration of hemolymph
added (5%), juvenile survival improved, the production of viable eggs was stimulated and as a
consequence the intrinsic rate of increase substantially increased. The intrinsic rate of increase
of A. swirskii fed on the basic artificial diet enriched with black soldier fly prepupal
hemolymph was equally high as that of females reared on the basic diet supplemented with
pupal hemolymph of A. pernyi. These results demonstrate the potential of H. illucens as a
cheap nutrient source for the rearing of A. swirskii and perhaps other beneficial arthropods.
After several consecutive generations of A. swirskii reared on the artificial diets,
nutrient imbalances became apparent to a more or less extent. In the sixth generation, the
immature survival of the predators reared on artificial diets enriched with E. kuehniella eggs
or A. franciscana cysts was significantly lower than that on the corresponding diet in the first
generation (Chapter 5). However, artificial diet supplemented with A. franciscana cysts
proved superior to the other artificial diets as it allowed to sustain A. swirskii for over 15
consecutive generations despite the lower immature survival rates, whereas the colonies kept

General discussion, conclusions and future perspectives
119
on a diet enriched with A. pernyi hemolymph or E. kuehniella egg extract could not be
maintained beyond 3 and 9 generations, respectively. In addition, A. swirskii females did not
lose their capacity to capture and kill live prey after six consecutive generations of rearing on
the tested artificial diets. Although the diet supplemented with A. franciscana cysts showed to
be the most promising diet for the production of A. swirskii, loss of quality in terms of
reduced survival after several generations of continuous rearing remains a concern. In this
context, we found that a switching to T. latifolia pollen after 6 generations of rearing on the
artificial diet could significantly increase the immature survival of the predator to more than
90%. This indicates the importance of providing different types of food (of both plant and
animal origin) to maintain the colony health of these omnivorous predators.
In Chapter 7, the diet supplemented with A. franciscana cysts was found to allow full
development and reproduction of four other phytoseiid mites species. These findings suggest
that the same diet may also be useful to support populations of other commercially available
phytoseiids, either in the laboratory or in the field. This may be of particular significance
when using the artificial diet as a supplemental food in the crop. Finding inexpensive
alternative food sources is one of the major opportunities for enhancing biological control in
protected crops (Messelink et al., 2014). Our results are indeed promising given that only few
existing supplemental foods have been reported to support beneficial Acarina (Wade et al.,
2008), and commercially available supplemental foods like Typha angustifolia L. pollen
(NutrimiteTM
, Biobest N.V.), eggs of E. kuehniella and decapsulated cysts of A. franciscana
are relatively costly.
A further objective of this study was to find the most adequate formulation of artificial
diets for easy and effective application in a laboratory culture and as a food supplement for
predatory mites in the crop. The artificial diets used in Chapters 3-5 were all liquid diets.
Solid artificial diets have a number of advantages over liquid diets including non-stickiness,
limiting microbial contamination, easy long term storage and the possibility of direct
presentation without the need for encapsulation (Morales-Ramos et al., 2014). More in
particular, solid artificial diets are easier to distribute in the crop by existing blowers designed
to distribute pollen or eggs of E. kuehniella. Solid artificial diets can also be supplied in other
ways used for pollen, like in plastic cups placed between the crop plants (Nomikou et al.,
2010) or by coating twine attached to the plants thus providing the predatory mites at the
same time with shelter and alternative food (Adar et al., 2014). The liquid artificial diets
developed in our study were solidified by freeze-drying or by replacing the liquid ingredients
of the original diet by powdered solid ingredients. In Chapter 6, we found that the fecundity

Chapter 8
120
of females fed on the freeze-dried liquid artificial diets was similar to that on T. latifolia
pollen or liquid versions of the diets and higher than that on the powdered diets. The results
indicated that the freeze-drying process did not influence the nutritional quality of the diets
and that the predator performed on the solid forms as effectively as on the liquid forms.
Freeze-dried diets have a number of practical advantages over liquid diets, related to storage,
shipment and handling. However, the freeze-dried diets used in our study proved to be highly
hygroscopic and quickly reverted to a semi-liquid form; this phenomenon may limit the
practical applicability of such diets as they may be more prone to microbial degradation.
Artificial diets entirely composed of powdered/dry ingredients were somewhat inferior to
lyophilized liquid diets in terms of their nutritional value, but did support the development
and reproduction of the phytoseiid and were physically more stable than the lyophilized diets.
In a pre-establishment experiment with A. swirskii on caged chrysanthemum plants in a
greenhouse, the powdered artificial diet with or without the addition of powdered dry A.
franciscana cysts significantly encouraged population growth of the predatory mite. Predator
densities in the treatment with powdered artificial diet were similar to those of mites supplied
with T. angustifolia pollen. Furthermore, most ingredients of the dry artificial diets tested here
are relatively inexpensive and easily accessible on the market. We estimate that the price of
our powdered artificial diet with or without A. franciscana would be approximately 170 or
100 EUR/kg. Based on economies of scale, prices may further be reduced if these diets are
produced in larger quantities. These findings indicate the potential of dry artificial diets for
use in part of the production cycle of A. swirskii or as supplemental foods to sustain its
populations in the crop after release.
In conclusion, various factitious and artificial diets tested in the present study
supported the development and reproduction of A. swirskii. The phytoseiid performed best on
decapsulated Artemia cysts and a meridic artificial diet enriched with these cysts, indicating
the potential of these foods for use in (parts of) the production cycle of the predator. The
powdered artificial diet supplemented with A. franciscana cysts also has potential for use as a
supplemental food in a conservation biological control strategy to sustain released populations
of the predatory mite in greenhouse crops. Our findings also indicate the potential of using H.
illucens as a cheap source of hemolymph in artificial diets for predatory mites and potentially
other predatory arthropods, as the fly can be cost effectively produced at a large scale on
organic waste materials. However, the effects of long-term rearing of the mites using these
artificial diets on their quality as biological control agents need to be explored. Further, field
research is needed to evaluate the suitability of the artificial diets developed in this study in

General discussion, conclusions and future perspectives
121
the crop environment, with regard to their practical value to support populations of A. swirskii
or of other predatory mites, giving attention to possible undesired effects of their application
on populations of arthropod pests (like omnivorous thrips who may also be able to use the
diets) and on the physiology and quality of the crop plants (e.g. fungal contamination).

Chapter 8
122

123
SUMMARY
The predatory mite Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) is an
economically important biological control agent of several key pests in greenhouses, such as
whiteflies, thrips, eriophyid mites and broad mites. In augmentation biological control, large
numbers of predaceous mites are released in the field. Hence, a cost-effective method for their
mass-rearing is an essential prerequisite. Rearing phytoseiid mites on plant materials infested
with natural prey has several disadvantages, such as large space requirements, inconsistent
yields of predators, harvesting difficulties and variable results with different species. Rearing
procedures based on factitious prey like storage mites also involve space and labor to
maintain large parallel cultures of the factitious prey. Further, there may be health risks for
workers in production facilities or in the crop caused by allergens associated with the
factitious mite prey. The availability of adequate artificial or factitious diets could eliminate
many of the above-mentioned problems associated with the mass production of predatory
mites. In addition, these diets may be useful as food supplements to support predator
populations after release in the crop. The overall objective of this study was to develop
alternative food sources for phytoseiid predatory mites (with focus on the economically
important species A. swirskii) in support of their mass production and use as food supplements
to sustain their populations after release in the crop.
In Chapter 3, the development, survival and reproduction of A. swirskii were assessed
when the predator was fed on cattail pollen (Typha latifolia L.), dried fruit mite
(Carpoglyphus lactis L.) (Acari: Carpoglyphidae) or on two artificial diets: a basic artificial
diet composed of honey, sucrose, tryptone, yeast extract, and egg yolk; and the basic diet
enriched with pupal hemolymph of Chinese oak silkworm (Antheraea pernyi (Guérin-
Méneville) (Lepidoptera: Saturniidae)). Mites fed on C. lactis and A. pernyi hemolymph diet
had shorter immature and preoviposition periods than those fed on the other diets. The total
number of deposited eggs was significantly higher for females fed on A. pernyi hemolymph
diet (38.25 eggs/female) than for those maintained on the other diets. The intrinsic rate of
increase (rm) of A. swirskii was highest on A. pernyi hemolymph diet (0.181) and C. lactis
(0.175), followed by T. latifolia pollen (0.158), and basic diet (0.104). The artificial diet
enriched with A. pernyi pupal hemolymph supported development and reproduction of A.
swirskii to the same extent as a factitious prey which is routinely used in the mass rearing of
the phytoseiid. These results indicate the potential of artificial diets for the mass production of
this economically important predatory mite.

Summary
124
For reducing the cost of the artificial diet, the silkworm hemolymph was replaced by
prepupal hemolymph of the black soldier fly, Hermetia illucens (L.) (Diptera: Stratiomyidae),
an easily available and cheap nutrient. In Chapter 4, the survival, development, and
reproduction of the predatory mite A. swirskii were assessed when fed on basic artificial diet
supplemented with 5, 10, or 20% of H. illucens prepupal hemolymph. Developmental time
from larva to adult was shorter for males and females offered artificial diets supplemented
with 20% hemolymph versus the basic diet. The oviposition rate and total fecundity of
females reared on the basic diet were substantially lower than those of females supplied with
the enriched diets. The intrinsic rate of increase was highest on the diet containing 20%
hemolymph, followed by those containing 10 and 5% hemolymph. In a subsequent diet
switching experiment, mites fed on the basic diet in their juvenile stages were switched upon
adulthood to diet enriched with different concentrations of H. illucens hemolymph. The
females that were fed with the enriched diets from the adult stage on had higher oviposition
rates and fecundities than those maintained on the basic diet, but their reproductive
parameters were not significantly affected by the concentration of the hemolymph in the
artificial diet. The results proved that supplementing artificial diets with black soldier fly
hemolymph significantly improved their nutritional value for A. swirskii. Findings of these
experiments indicate the potential of using H. illucens as a cheap source for hemolymph in
artificial diets, as the fly can be cost effectively produced at a large scale on organic waste
materials.
In Chapter 5, A. swirskii was reared on Ephestia kuehniella Zeller eggs (Lepidoptera:
Pyralidae), decapsulated dry cysts of the brine shrimp Artemia franciscana Kellogg
(Anostraca: Artemiidae), and on basic artificial diets supplemented with pupal hemolymph of
the Chinese oak silkworm A. pernyi (AD1), with E. kuehniella eggs (AD2) or with A.
franciscana cysts (AD3). Development, reproduction and predation capacity of the predatory
mites were assessed in the first (G1) and sixth generation (G6) of rearing on the different diets.
Immature survival rates in G1 were similar on all diets (96.8-100%). After 6 generations,
however, survival of A. swirskii was significantly reduced on all diets except on A.
franciscana cysts. Oviposition rates did not differ between generations when females were fed
on E. kuehniella, AD2 or AD3. The total number of deposited eggs was similar among diets
except in G6 where the females fed on A. franciscana cysts produced more eggs than those
maintained on E. kuehniella eggs. On most diets the intrinsic rates of increase in G1 were
superior to those in G6, except for predators supplied with A. franciscana cysts where no
differences were observed among generations. Female mites did not lose their capacity to kill
first instar Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) after six
generations on the different diets, but predation rates in G6 on E. kuehniella were lower than

Summary
125
in G1. In summary, the different factitious and artificial diets tested in this study supported the
development and reproduction of A. swirskii for a single generation but fitness losses occurred
to a varying degree after several generations on E. kuehniella eggs or the artificial diets.
Artificial diet enriched with A. franciscana cysts yielded better results than the other artificial
diets. Amblyseius swirskii performed best on decapsulated Artemia cysts indicating their
potential for use in the mass production of the predator or to sustain its populations in the crop
after release.
Solid artificial diets have a number of advantages over liquid and semiliquid diets,
both in the laboratory rearing and when applied as supplemental foods in the crop. These
include non-stickiness and the possibility of direct presentation without the need for
encapsulation. Lower water content of solid diets limits microbial contamination and allows
easier long term storage. When applied as a supplemental food in the field, solid artificial
diets are easier to distribute in the crop by existing blowers designed to distribute pollen or
eggs of E. kuehniella and are expected to result in lower soiling of the plant surface and thus
to interfere less with plant physiology and crop quality. In Chapter 6, we investigated the
survival, development and reproduction of A. swirskii fed on several dry artificial diets: the
tested diets were freeze-dried forms of previously developed liquid meridic artificial diets
supplemented with a watery extract of decapsulated cysts of A. franciscana or with pupal
hemolymph of Chinese oak silkworm A. pernyi, and newly composed powdered meridic
artificial diets (composed of sucrose, tryptone, yeast extract, glucose, fructose, egg yolk
powder, vitamin mix) supplemented with ground dry A. franciscana cysts or lyophilized
pupal hemolymph of A. pernyi. Performance of the mite on the artificial diets was compared
with that on cattail pollen (T. latifolia). Developmental time of A. swirskii females offered
lyophilized diets was significantly shorter than on powdered diets. Total fecundity was
significantly higher for females fed on the lyophilized diets than for those maintained on the
powdered diet with A. franciscana. Daily oviposition rates were similar on T. latifolia pollen
and both lyophilized diets but lower on both powdered diets. The highest intrinsic rate of
increase was observed when A. swirskii was fed on T. latifolia pollen (0.210
females/female/day), followed by the freeze dried diets enriched with A. pernyi and A.
franciscana (0.195 and 0.184 females/female/day, respectively), and the lowest growth rates
were observed on the powdered diets supplemented with A. franciscana and A. pernyi (0.159
and 0.158 females/female/day, respectively). In a greenhouse experiment, the application of
powdered artificial diets on chrysanthemum plants supported the population growth of A.
swirskii over several weeks. These results proved that the phytoseiid was able to effectively
feed on solid, powdered artificial diets. Freeze-drying of liquid diets did not influence their
value to support the development and reproduction of A. swirskii.

Summary
126
In Chapter 7 the impact of the liquid basic artificial diet supplemented with A.
franciscana watery extract on the biological performance of four species of other phytoseiid
mites which are commercially available in Europe (Neoseiulus californicus (McGregor), N.
cucumeris (Oudemans), Amblyseius andersoni Chant, and Amblydromalus limonicus Garman
& McGregor (Acari: Phytoseiidae)) is investigated. The life table parameters of these
predatory mites fed on A. franciscana artificial diet were compared to that feeding on two-
spotted spider mites Tetranychus urticae Koch (Acarina: Tetranychidae) (N. californicus) or T.
latifolia pollen (N. cucumeris, A. andersoni and A. limonicus). Diet had no influence on the
immature survival rate, ranging from 92 to 98% for all species. Female developmental times
were significantly shorter for predators offered spider mites or pollen than for those fed the
artificial diet, except in A. limonicus. The fecundity of N. californicus, N. cucumeris and A.
limonicus females given spider mites or pollen was significantly higher than that of females
presented with the artificial diet, whereas no differences among diets were observed in A.
andersoni. When N. californicus females were fed on the artificial diet, none of their offspring
succeeded in reaching adulthood. Our findings indicate the potential of this artificial diet for
use in production facilities or in the crop, especially for the more generalist predatory mites A.
andersoni and A. limonicus.
In Chapter 8, a general discussion of the findings is presented and future prospects
are discussed. It is concluded that the different factitious and artificial diets tested in the
present study supported the development and reproduction of A. swirskii. The predatory mite
performed best on decapsulated Artemia cysts and the artificial diet enriched with these cysts,
indicating the potential for use of these foods in (parts of) the production cycle of the predator.
The powdered artificial diet supplemented with A. franciscana cysts also has potential for use
as a supplemental food to sustain released populations of the predatory mite in greenhouse
crops. Our findings also indicate the potential of using H. illucens as a cheap source of
hemolymph in artificial diets for predatory mites and potentially other predatory arthropods.
However, the effects of long-term rearing of the mites on these artificial diets on their quality
as biological control agents need to be explored. Further field research is also needed to
optimize the value of these artificial diets to support the predatory mites in the crop
environment and to assess their effects on the physiology and quality of the crop plants.

127
SAMENVATTING
De roofmijt Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) is een
economisch belangrijke biologische bestrijder van verschillende plagen in kasteelten, zoals
wittevlieg, trips en verschillende soorten plaagmijten. In biologische bestrijding door middel
van vermeerdering worden grote hoeveelheden roofmijten in het veld geïntroduceerd. Een
rendabele methode om deze predators in massa te kweken is bijgevolg een belangrijke
vereiste. Het kweken van deze bladroofmijten op plantenmateriaal geïnfesteerd met hun
natuurlijke prooien heeft verschillende nadelen, zoals de noodzaak voor grote ruimtes voor de
opkweek, variabele opbrengst van de predators, moeilijkheden bij het oogsten en
verschillende resultaten bij de verschillende soorten. Ook kweekmethodes die gebaseerd zijn
op alternatieve prooien, zoals voermijten, brengen veel plaats en arbeid met zich mee. Verder
kunnen gezondheidsrisico’s optreden voor de werknemers in de productie-omgeving en in het
gewas als gevolg van allergenen geassocieerd met de voermijten. De beschikbaarheid van
geschikte kunstmatige voedselbronnen zou veel van de hierboven beschreven beperkingen bij
de massakweek van roofmijten kunnen elimineren. Bovendien kunnen deze diëten nuttig
blijken als voedingssupplementen voor populaties van de roofmijten na introductie in het
gewas. De algemene doelstelling van deze studie omvatte het ontwikkelen van alternatieve
voedselbronnen voor roofmijten van de familie Phytoseiidae (met de focus op de economisch
belangrijke soort, A. swirskii) ter ondersteuning van hun massakweek en voor gebruik als
voedingssupplementen na introductie in het gewas.
In Hoofdstuk 3 werd de ontwikkeling, overleving en reproductie van A. swirskii
nagegaan wanneer de predator werd gevoed met stuifmeel van de grote lisdodde (Typha
latifolia L.), voermijten (Carpoglyphus lactis L.) (Acari: Carpoglyphidae) of twee artificiële
diëten: een basisdieet samengesteld uit honing, sucrose, trypton, gist extract en eigeel; en het
basisdieet verrijkt met hemolymfe van poppen van de Chinese eikenzijderups (Antheraea
pernyi (Guérin-Méneville) (Lepidoptera: Saturniidae)). Roofmijten gevoed met C. lactis en
het dieet verrijkt met A. pernyi hemolymfe vertoonden een kortere ontwikkelingsduur en
preovipositieperiode dan deze gevoed op de andere diëten. Het totaal aantal afgelegde eitjes
was significant hoger voor wijfjes gevoed met het dieet verrijkt met hemolymfe van A. pernyi
(38,25 eitjes/wijfje) dan deze gevoed op de andere diëten. De intrinsieke groeisnelheid (rm)

Samenvatting
128
van A. swirskii was hoogst op het dieet verrijkt met A. pernyi hemolymfe (0,181
wijfjes/wijfje/dag) en C. lactis (0,175), gevolgd door stuifmeel van T. latifolia (0,158), en het
basisdieet (0,104). Het artificieel dieet verrijkt met hemolymfe van A. pernyi poppen
ondersteunde de ontwikkeling en reproductie van A. swirskii in dezelfde mate als de
voermijten die routinematig gebruikt wordt bij de massakweek van deze roofmijt.
Om de kost van het artificieel dieet te verminderen, werd de hemolymfe van de
zijderups vervangen door hemolymfe van prepoppen van de zwarte wapenvlieg, Hermetia
illucens (L.) (Diptera: Stratiomyidae), wat een makkelijk beschikbaar en goedkoop nutriënt is.
In Hoofdstuk 4, werd de overleving, ontwikkeling en reproductie van de roofmijt A. swirskii
nagegaan wanneer deze gevoed werd op het basisdieet verrijkt met 5, 10 of 20% hemolymfe
van prepoppen van H. illucens. De ontwikkelingsduur van larf tot adult was korter voor
mannetjes en wijfjes die artificiële diëten aangeboden kregen verrijkt met 20% hemolymfe
vergeleken met het basisdieet. De ovipositiegraad en totale fecunditeit van wijfjes gekweekt
op het basisdieet waren aanzienlijk lager dan bij deze gekweekt op de verrijkte diëten. De
intrinsieke groeisnelheid was hoogst op het dieet bestaande uit 20% hemolymfe, gevolgd door
de diëten met 10 en 5% hemolymfe. In een daaropvolgende proef werden mijten tijdens hun
onvolwassen stadia gevoed op het basisdieet, om vervolgens, na het bereiken van het adulte
stadium, het voedingsregime te wisselen naar diëten met verschillende concentraties van H.
illucens hemolymfe. Wijfjes die vanaf het adulte stadium gevoed werden op de verrijkte
diëten hadden een hogere dagelijkse en totale ei-afleg dan deze die doorgekweekt werden op
het basisdieet. Hun reproductieve parameters werden echter niet beduidend beïnvloed door de
concentratie aan hemolymfe in het artificieel dieet. De resultaten toonden aan dat het
verrijken van artificiële diëten met hemolymfe van de zwarte wapenvlieg de nutritionele
waarde voor A. swirskii significant verhoogde. De bevindingen van deze experimenten tonen
het potentieel aan van H. illucens als een goedkope bron van haemolymfe in artificiële diëten,
daar deze vlieg goedkoop en op grote schaal op organische afvalstromen kan geproduceerd
worden.
In Hoofdstuk 5 werd A. swirskii gekweekt op Ephestia kuehniella Zeller eitjes
(Lepidoptera: Pyralidae), droge gedecapsuleerde cysten van het pekelkreeftje Artemia
franciscana Kellogg (Anostraca: Artemiidae), en op een artificieel basisdieet verrijkt met
ofwel hemolymfe van poppen van de Chinese eikenzijderups A. pernyi (AD1), E. kuehniella
eggs (AD2), of A. franciscana cysten (AD3). De ontwikkeling, reproductie en
predatiecapaciteit van de roofmijten werd nagegaan in de eerste (G1) en de zesde (G6)
generatie opgekweekt op de diëten. De overlevingspercentages van de juveniele stadia in G1

Samenvatting
129
waren gelijkaardig voor alle diëten (96,8-100%). Echter, na 6 generaties verlaagde de
overleving significant op alle diëten behalve op A. franciscana cysten. De ovipositiegraden
verschilden niet tussen de generaties wanneer de wijfjes gevoed werden op E. kuehniella,
AD2 of AD3. Het totaal aantal afgelegde eitjes was gelijk voor alle diëten, behalve in G6
waar wijfjes die gevoed werden op A. franciscana cysten meer eitjes produceerden dan deze
gekweekt op E. kuehniella eitjes. Bij de meeste diëten was de intrinsieke groeisnelheid in G1
hoger dan deze in G6, behalve voor predators die A. franciscana cysten kregen aangeboden
waar geen verschillen tussen de generaties werden waargenomen. Wijfjes van de roofmijt
verloren hun capaciteit niet om eerstestadiumlarven van Frankliniella occidentalis (Pergande)
(Thysanoptera: Thripidae) te prederen na 6 generaties op de verschillende diëten, maar het
predatievermogen van de G6-roofmijten gevoed op E. kuehniella was lager dan in G1. De
verschillende alternatieve voedselbronnen en artificiële diëten in getest deze studie
ondersteunden de ontwikkeling en reproductie van A. swirskii gedurende de eerste generatie,
maar na enkele generaties werd een verlies aan fitness vastgesteld voor mijten gevoed op E.
kuehniella eitjes en de artificiële diëten. Het artificieel dieet verrijkt met A. franciscana cysten
leverde betere resultaten op dan de andere diëten. Amblyseius swirskii presteerde het best op
gedecapsuleerde Artemia cysten, wat hun potentieel aantoont voor gebruik in de
massaproductie van de predator of om populaties in het veld te ondersteunen.
Vaste diëten hebben verschillende voordelen in vergelijking met vloeibare en
halfvloeibare diëten, zowel in laboratoriumkweken of wanneer gebruikt als ondersteunende
voedingssupplementen in het gewas, zoals hun lagere kleverigheid en de mogelijkheid om ze
aan te bieden zonder de noodzaak ze eerst te encapsuleren. De lagere waterinhoud van vaste
diëten beperkt microbiële contaminatie en verhoogt de bewaartijd. Wanneer diëten als
voedingssupplement in het veld worden toegediend, zijn vaste diëten makkelijker om te
verspreiden in het gewas door gebruik te maken van blazers die ontwikkeld zijn voor
stuifmeel of E. kuehniella eitjes; bovendien zouden ze de planten minder moeten vervuilen en
op deze manier minder met de plantfysiologie en gewaskwaliteit interfereren. In Hoofdstuk 6
werd de overleving, ontwikkeling en reproductie van A. swirskii onderzocht wanneer deze
gevoed werd op verschillende droge artificiële diëten: de geteste diëten waren ofwel
gevriesdroogde formuleringen van de voorheen ontwikkelde vloeibare artificiële diëten
verrijkt met extracten van gedecapsuleerde cysten van A. franciscana of met hemolymfe van
de Chinese eikenzijderups A. pernyi, ofwel nieuw samengestelde artificiële poederdiëten
(bestaande uit sucrose, trypton, gistextract, glucose, fructose, eigeelpoeder en een
vitaminemix) verrijkt met vermaalde droge A. franciscana cysten of gelyofiliseerde

Samenvatting
130
hemolymfe van A. pernyi poppen. De prestaties van de roofmijt werden vergeleken met deze
op een dieet bestaande uit stuifmeel van de grote lisdodde (T. latifolia). De ontwikkelingsduur
van A. swirskii wijfjes die gelyofiliseerde diëten kregen aangeboden was significant korter
dan van de roofmijten gevoed op poederdiëten. De totale ei-afleg was beduidend hoger voor
wijfjes gevoed met de gelyofiliseerde diëten dan voor deze gekweekt op het poederdieet
verrijkt met A. franciscana. De dagelijkse ei-afleg was gelijkaardig op T. latifolia stuifmeel en
beide gelyofiliseerde diëten, maar was lager voor beide poederdiëten. De hoogste intrinsieke
groeisnelheid werd waargenomen wanneer A. swirskii gevoed werd op T. latifolia stuifmeel
(0,210 wijfjes/wijfje/dag), gevolgd door de gevriesdroogde diëten verrijkt met A. pernyi en A.
franciscana (0,195 en 0,184 wijfjes/wijfje/dag, respectievelijk). De laagste groeisnelheid werd
geobserveerd bij de poederdiëten verrijkt met A. franciscana en A. pernyi (0,159 en 0,158
wijfjes/wijfje/dag, respectievelijk). In een serreproef werd aangetoond dat de toediening van
de poederdiëten op chrysanten de populatiegroei van A. swirskii ondersteunde gedurende
verschillende weken. Het vriesdrogen van vloeibare diëten had geen invloed op hun
ondersteunende waarde voor de ontwikkeling en reproductie van A. swirskii.
In Hoofdstuk 7 werd de impact getest van het vloeibare basisdieet verrijkt met een
waterig extract van A. franciscana op de biologische prestaties van 4 soorten roofmijten van
de familie Phytoseiidae die commercieel beschikbaar zijn in Europa (Neoseiulus californicus
(McGregor), N. cucumeris (Oudemans), Amblyseius andersoni Chant, en Amblydromalus
limonicus Garman & McGregor (Acari: Phytoseiidae)). De levenstabelparameters van deze
roofmijten gevoed op het met A. franciscana verrijkte basisdieet werden vergeleken met deze
van roofmijten gevoed met de bonenspintmijt Tetranychus urticae Koch (Acarina:
Tetranychidae) (voor N. californicus) of T. latifolia pollen (voor N. cucumeris, A. andersoni
en A. limonicus). Het dieet had geen invloed op de overleving van de juveniele stadia, die
varieerde tussen 92 en 98% voor alle soorten. De ontwikkelingsduur van de wijfjes was
significant korter voor predators die spintmijten of pollen kregen aangeboden vergeleken met
deze gevoed op het artificieel dieet, behalve voor A. limonicus. De fecunditeit van wijfjes van
N. californicus, N. cucumeris en A. limonicus voorzien met spintmijten of pollen was
significant hoger dan deze van wijfjes die gevoed werden op het artificieel dieet; er werd
echter geen verschil tussen de diëten waargenomen voor A. andersoni. Wanneer N.
californicus wijfjes gevoed werden op het artificieel dieet, waren de nakomelingen niet in
staat het volwassen stadium te bereiken. Onze resultaten tonen het potentieel aan van dit
artificieel dieet voor verschillende Phytoseiidae roofmijten wanneer gebruikt in de

Samenvatting
131
massakweek of in het gewas, in het bijzonder voor de meer polyfage roofmijten A. andersoni
en A. limonicus.
In Hoofdstuk 8 wordt een algemene discussie van de resultaten gepresenteerd en toekomstige
perspectieven worden aangegeven. Als algemene conclusie kan worden gesteld dat de
verschillende alternatieve prooien en artificiële diëten getest in deze studie de ontwikkeling en
de reproductie van A. swirskii ondersteunden. De roofmijt presteerde het best op
gedecapsuleerde Artemia cysten en het artificieel dieet verrijkt met deze cysten, wat het
potentieel van deze voedingsbronnen aantoont in (delen van) de productiecyclus van de
predator. De poedervorm van het artificieel dieet verrijkt met A. franciscana cysten heeft ook
potentieel om gebruikt te worden als voedingssupplement om geïntroduceerde populaties van
deze roofmijt te ondersteunen in kasteelten. Onze bevindingen tonen ook de bruikbaarheid
aanvan H. illucens als een goedkope bron van hemolymfe in artificiële diëten voor roofmijten
en mogelijk ook andere predators. De effecten van het langdurig kweken van roofmijten op
deze artificiële diëten op hun kwaliteit als biologische bestrijders dient verder onderzocht te
worden. Verder praktisch onderzoek is ook aangewezen om de waarde van deze artificiële
diëten als ondersteunende voedingsbron in het gewas in te schatten en om hun effecten op de
fysiologie en kwaliteit van de gewasplanten na te gaan.

Samenvatting
132

133
REFERENCES
Abo-Taka, S.M., 1996. Studies on some ecological factors affecting phytophagous and
predacious mites in grape yards. In: Mitchell, R., Horn, D.J., Needham, G.R.,
Welbourn, W.C., Eds.), Acarology IX. Proceedings. The Ohio Biological Survey,
Columbus, USA, pp. 177-180.
Abou-Awad, A., Elsawi, S.A., 1992. Generations and reproduction in the predacious mite
Amblyseius swirskii Ath.-Hen. (Acari, Phytoseiidae) fed with artificial and natural
diets. Anzeiger für Schädlingskunde, Pflanzenschutz, Umweltschutz 65, 115-117.
Abou-Awad, B., Reda, A., Elsawi, S., 1992. Effects of artificial and natural diets on the
development and reproduction of two phytoseiid mites Amblyseius gossipi and
Amblyseius swirskii (Acari: Phytoseiidae). International Journal of Tropical Insect
Science 13, 441-445.
Abou-Awad, B.A., El-Sawaf, B.M., Abdel-Khalek, A.A., 1999. Impact of two eriophyoid fig
mites, Aceria ficus and Rhyncaphytoptus ficifoliae, as prey on postembryonic
development and oviposition rate of the predacious mite Amblyseius swirskii.
Acarologia 40, 367-371.
Abou-Awad, B.A., El-Sawaf, B.M., Reda, A.S., Abdel-Khalek, A.A., 2000. Environmental
management and biological aspects of the two eriophyoid fig mites Aceria ficus (Cotte)
and Rhyncaphytoptus ficifoliae Keifer in Egypt. Journal of Pest Science 73, 5-12.
Abou-Awad, B.A., Hafez, S.M., Farhat, B.M., 2014. Biological studies of the predacious mite
Amblyseius swirskii, a predator of the broad mite Polyphagotarsonemus latus on
pepper plants (Acari: Phytoseiidae: Tarsonemidae). Archives of Phytopathology and
Plant Protection 47, 349-354.
Adar, E., Inbar, M., Gal, S., Doron, N., Zhang, Z.Q., Palevsky, E., 2012. Plant-feeding and
non-plant feeding phytoseiids: differences in behavior and cheliceral morphology.
Experimental and Applied Acarology 58, 341-357.
Adar, E., Inbar, M., Gal, S., Gan-Mor, S., Palevsky, E., 2014. Pollen on-twine for food
provisioning and oviposition of predatory mites in protected crops. BioControl 59,
307-317.
Afifi, A., Potts, M., Patterson, C., Rodriguez, J., 1988. Pollen diet of some predator mites.
Transactions of the Kentucky Academy of Science 49, 96-100.

References
134
Alauzet, C., Dargagnon, D., Hatte, M., 1992. Production d'un Hetereroptere predateur: Orius
majusculus (Rtt.) (Heteroptera: Anthocoridae). Entomophaga 37, 249-252.
Ali, F., Zaher, M., 2007. Effect of food and temperature on the biology of Typhlodrompis
swirskii (Athias-Henriot)(Acari: Phytoseiidae). Acarines 1, 17-21.
Allen, C.M., 2010. Thermal biology and behaviour of two predatory Phytoseiid mites:
Amblyseius swirskii (Athias-Henriot)(Acari: Phytoseiidae) and Phytoseiulus longipes
(Evans)(Acari: Phytoseiidae). PhD thesis, University of Birmingham.
Arijs, Y., De Clercq, P., 2001. Rearing Orius laevigatus on cysts of the brine shrimp Artemia
franciscana. Biological Control 21, 79-83.
Arthurs, S., McKenzie, C.L., Chen, J.J., Dogramaci, M., Brennan, M., Houben, K., Osborne,
L., 2009. Evaluation of Neoseiulus cucumeris and Amblyseius swirskii (Acari:
Phytoseiidae) as biological control agents of chilli thrips, Scirtothrips dorsalis
(Thysanoptera: Thripidae) on pepper. Biological Control 49, 91-96.
Athias-Henriot, C., 1962. Amblyseius swirskii, un nouveau phytoseiide voisin d'A. andersoni
(Acariens anactinotriches). Annales de l'Ecole Nationale d'Agriculture d'Alger,
Algeria 3, 1-7.
Attia, A.R., El-Arnaouty, S., Afifi, A.I., Alla, A., 2011. Development and fecundity of the
coccinellid predator, Cryptolaemus montrouzieri Mulsant on different types of preys.
Egyptian Journal of Biological Pest Control 21, 283-289.
Banks, I.J., Gibson, W.T., Cameron, M.M., 2014. Growth rates of black soldier fly larvae fed
on fresh human faeces and their implication for improving sanitation. Tropical
medicine & international health 19, 14-22.
Baxter, I., Midthassel, A., Stepman, W., Fryer, R., Garcia, F., Lewis, J., Walker, P., Hulshof,
J., 2011. Field results of a sachet release system using the predator Amblyseius swirskii
(Athias-Henriot)(Acari: Phytoseiidae) and the factitious prey, Suidasia medanensis
Oudemans (Acari: Astigmata). IOBC/WPRS Bulletin 68, 1-4.
Berndt, O., Meyhofer, R., Richter, E., 2007. Whitefly on cut gerbera: Biological control by
parasitoids or predatory mites? Gesunde Pflanzen 59, 171-178.
Biobest, 2014. http://www.biobest.be.
Birch, L.C., 1948. The intrinsic rate of natural increase of an insect population. Journal of
Animal Ecology 17, 15-26.
Bogdanov, S., 2004. Quality and standards of pollen and beeswax. Apiacta 38, 334-341.
Bolckmans, K., van Houten, Y., Hoogerbrugge, H., 2005. Biological control of whiteflies and
western flower thrips in greenhouse sweet peppers with the phytoseiid predatory mite

References
135
Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). Second International
Symposium on Biological Control of Arthropods, pp. 555-565.
Bolckmans, K.J.F., van Houten, Y.M., 2006. Mite composition, use thereof, method for
rearing the phytoseiid predatory mite Amblyseius swirskii, rearing system for rearing
said phytoseiid mite and methods for biological pest control on a crop. WO Patent
WO/2006/057552.
Bolckmans, K.J.F., Van Houten, Y.M., 2011. Mite composition, use thereof, method for
rearing a phytoseiid predatory mite, rearing system for rearing said phytoseiid
predatory mite and methods for biological pest control on a crop. U.S. Patent No.
7,947,269.
Bondari, K., Sheppard, D., 1987. Soldier fly, Hermetia illucens L., larvae as feed for channel
catfish, Ictalurus punctatus (Rafinesque), and blue tilapia, Oreochromis aureus
(Steindachner). Aquaculture Research 18, 209-220.
Bonte, M., De Clercq, P., 2008. Developmental and reproductive fitness of Orius laevigatus
reared on factitious and artificial diets. Journal of Economic Entomology 101, 1127-
1133.
Bonte, M., Samih, M.A., De Clercq, P., 2010. Development and reproduction of Adalia
bipunctata on factitious and artificial foods. BioControl 55, 485-491.
Buitenhuis, R., Shipp, L., Scott-Dupree, C., 2010a. Dispersal of Amblyseius swirskii Athias-
Henriot (Acari: Phytoseiidae) on potted greenhouse chrysanthemum. Biological
Control 52, 110-114.
Buitenhuis, R., Shipp, L., Scott-Dupree, C., 2010b. Intra-guild vs extra-guild prey: effect on
predator fitness and preference of Amblyseius swirskii (Athias-Henriot) and
Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae). Bulletin of Entomological
Research 100, 167-173.
Calvo, F.J., Bolckmans, K., Belda, J.E., 2009. Development of a biological control-based
integrated pest management method for Bemisia tabaci for protected sweet pepper
crops. Entomologia Experimentalis et Applicata 133, 9-18.
Calvo, F.J., Bolckmans, K., Belda, J.E., 2011. Control of Bemisia tabaci and Frankliniella
occidentalis in cucumber by Amblyseius swirskii. BioControl 56, 185-192.
Calvo, J., Bolckmans, K., Belda, J.E., 2008. Controlling the tobacco whitefly Bemisia tabaci
(Genn.) (Hom.: Aleyrodidae) in horticultural crops with the predatory mite Amblyseius
swirskii (Athias-Henriot). Journal of Insect Science 8, 11-12.

References
136
Castañé, C., Quero, R., Riudavets, J., 2006. The brine shrimp Artemia sp. as alternative prey
for rearing the predatory bug Macrolophus caliginosus. Biological Control 38, 405-
412.
Cédola, C., Polack, A., 2011. Primer registro de Amblyseius swirskii (Acari: Phytoseiidae) en
Argentina. Revista de la Sociedad Entomológica Argentina 70, 375-378.
Chant, D., 1985. Systematics and morphology. In: Helle, W., Sabelis, M.W., Eds.), Spider
mites: their biology, natural enemies and control. Elsevier, Amsterdam, pp. 3-34.
Chant, D.A., McMurtry, J.A., 2007. Illustrated keys and diagnoses for the genera and
subgenera of the Phytoseiidae of the world (Acari: Mesostigmata). Indira Publishing
House.
Chapman, R.F., 1998. The insects: structure and function. Cambridge University Press, UK.
Chippendale, G., 1978. The functions of carbohydrates in insect life processes. In: Rockstein,
M., (Ed.), Biochemistry of Insects. Academic Press New York, NY, pp. 1-55.
Chow, A., Chau, A., Heinz, K.M., 2010. Compatibility of Amblyseius (Typhlodromips)
swirskii (Athias-Henriot) (Acari: Phytoseiidae) and Orius insidiosus (Hemiptera:
Anthocoridae) for biological control of Frankliniella occidentalis (Thysanoptera:
Thripidae) on roses. Biological Control 53, 188-196.
Chyzik, R., Klein, M., Ben-Dov, Y., 1995. Reproduction and survival of the predatory bug
Orius albidipennis on various arthropod prey. Entomologia Experimentalis et
Applicata 75, 27-31.
Coates, W., Ayerza, R., Palzkill, D., 2006. Supplemental pollination of jojoba—A means to
increase yields. Industrial Crops and Products 24, 41-45.
Cock, M.J., van Lenteren, J.C., Brodeur, J., Barratt, B.I., Bigler, F., Bolckmans, K., Cônsoli,
F.L., Haas, F., Mason, P.G., Parra, J.R.P., 2010. Do new access and benefit sharing
procedures under the convention on biological diversity threaten the future of
biological control? BioControl 55, 199-218.
Cohen, A.C., 1992. Using a systematic approach to develop artificial diets for predators. In:
Anderson, T.E., Leppla, N.C., Eds.), Advances in insect rearing for research and pest
management. Westview Press, Boulder, pp. 77-91.
Cohen, A.C., 1998. Solid-to-liquid feeding: the inside (s) story of extra-oral digestion in
predaceous Arthropoda. American Entomologist 44, 103-117.
Cohen, A.C., 1999. Artificial media for rearing entomophages comprising sticky, cooked
whole egg. U.S. Patent No. 5,945,271.
Cohen, A.C., 2004. Insect diets: science and technology. CRC Press, Boca Raton.

References
137
Cohen, A.C., Smith, L.K., 1998. A new concept in artificial diets for Chrysoperla rufilabris:
The efficacy of solid diets. Biological Control 13, 49-54.
Collier, T., Van Steenwyk, R., 2004. A critical evaluation of augmentative biological control.
Biological Control 31, 245-256.
Cônsoli, F.L., Parra, J.R.P., 1997. Development of an oligidic diet for in vitro rearing of
Trichogramma galloi Zucchi and Trichogramma pretiosum Riley. Biological Control
8, 172-176.
De Clercq, P., 2008. Culture of natural enemies on factitious foods and artificial diets. In:
Capinera, J.L., (Ed.), Encyclopedia of entomology. Springer, Dordrecht, pp. 1133-
1136.
De Clercq, P., Arijs, Y., Van Meir, T., Van Stappen, G., Sorgeloos, P., Dewettinck, K., Rey,
M., Grenier, S., Febvay, G., 2005a. Nutritional value of brine shrimp cysts as a
factitious food for Orius laevigatus (Heteroptera: Anthocoridae). Biocontrol Science
and Technology 15, 467-479.
De Clercq, P., Bonte, M., Van Speybroeck, K., Bolckmans, K., Deforce, K., 2005b.
Development and reproduction of Adalia bipunctata (Coleoptera: Coccinellidae) on
eggs of Ephestia kuehniella (Lepidoptera: Phycitidae) and pollen. Pest Management
Science 61, 1129-1132.
De Clercq, P., Coudron, T.A., Riddick, E.W., 2014. Production of heteropteran predators. In:
Morales-Ramos, J.A., Rojas, G., Shapiro-Ilan, D., Eds.), Mass production of beneficial
organisms: Invertebrates and entomopathogens. Elsevier, pp. 57-100.
DeBach, P., 1964. Biological control of insect pests and weeds. Reinhold Publishing Corp.,
California, USA.
Debolt, J.W., 1982. Meridic diet for rearing successive generations of Lygus hesperus. Annals
of the Entomological Society of America 75, 119-122.
Diener, S., Solano, N.M.S., Gutiérrez, F.R., Zurbrügg, C., Tockner, K., 2011. Biological
treatment of municipal organic waste using black soldier fly larvae. Waste and
Biomass Valorization 2, 357-363.
Diener, S., Zurbrugg, C., Tockner, K., 2009. Conversion of organic material by black soldier
fly larvae: establishing optimal feeding rates. Waste Management & Research 27, 603-
610.
Dindo, M., 1995. Possibilities of culturing Brachymeria intermedia (Nees)(Hymenoptera:
Chalcididae), a solitary pupal gypsy moth parasitoid, on artificial diets. IOBC/WPRS
Bulletin 18, 95-99.

References
138
Ding, D., Qiu, H., Hwang, C., 1980. In vitro rearing of an egg parasitoid, Tetrastichus
schoenobii Ferriere (Hymenoptera: Tetrastichidae). Contributions of the Shanghai
Institute of Entomology 1, 55-58.
Dogramaci, M., Arthurs, S.P., Chen, J.J., McKenzie, C., Irrizary, F., Osborne, L., 2011.
Management of chilli thrips Scirtothrips dorsalis (Thysanoptera: Thripidae) on
peppers by Amblyseius swirskii (Acari: Phytoseiidae) and Orius insidiosus (Hemiptera:
Anthocoridae). Biological Control 59, 340-347.
Dougherty, E.C., 1959. Introduction to axenic culture of invertebrate metazoan: a goal.
Annals of the New York Academy of Sciences 77, 27-54.
Doutt, R.L., Hagen, K.S., 1949. Periodic colonization of Chrysopa californica as a possible
control of mealybugs. Journal of Economic Entomology 42, 560-561.
Duso, C., Camporese, P., 1991. Developmental times and oviposition rates of predatory mites
Typhlodromus pyri and Amblyseius andersoni (Acari: Phytoseiidae) reared on
different foods. Experimental and Applied Acarology 13, 117-128.
El-Laithy, A.Y.M., 1998. Laboratory studies on growth parameters of three predatory mites
associated with eriophyid mites in olive nurseries. Journal of Plant Diseases and
Protection 105, 78-83.
El-Laithy, A.Y.M., Fouly, A.H., 1992. Life table parameters of the two phytoseiid predators
Amblyseius scutalis (Athias-Henriot) and A. swirskii A.-H.(Acari, Phytoseiidae) in
Egypt. Journal of Applied Entomology 113, 8-12.
El-Sawi, S., Momen, F., 2005. Biology of some phytoseiid predators (Acari: Phytoseiidae) on
eggs of Phthorimaea operculella and Spodoptera littoralis (Lepidoptera: Gelechiidae
and Noctuidae). Acarologia 45, 23-30.
EPPO, 2014. Commercially used biological control agents - Arachnida, Acarina. European
and Mediterranean Plant Protection Organization.
Erickson, M.C., Islam, M., Sheppard, C., Liao, J., Doyle, M.P., 2004. Reduction of
Escherichia coli O157: H7 and Salmonella enterica serovar enteritidis in chicken
manure by larvae of the black soldier fly. Journal of Food Protection 67, 685-690.
Fanti, P., Bratti, A., 1991. In vitro rearing of the larval stages of the parasitoid Pseudogonia
rufifrons Wied.(Diptera: Tachinidae): preliminary results. Insect Parasitoids 74, 449-
452.
FAO, 2012. The expert consultation meeting on assessing the potential of insects as food and
feed in assuring food security. Edible insects: future prospects for food and feed
security. FAO, Rome.

References
139
Faraji, F., Çobanoğlu, S., Çakmak, I., 2011. A checklist and a key for the Phytoseiidae species
of Turkey with two new species records (Acari: Mesostigmata). International Journal
of Acarology 37, 221-243.
Fauvel, G., Malausa, J., Kaspar, B., 1987. Etude en laboratoire des principales caracteristiques
biologiques de Macrolophus caliginosus (Heteroptera: Miridae). BioControl 32, 529-
543.
Ferkovich, S., Shapiro, J., 2004. Increased egg-laying in Orius insidiosus (Hemiptera:
Anthocoridae) fed artificial diet supplemented with an embryonic cell line. Biological
Control 31, 11-15.
Ferkovich, S.M., Venkatesan, T., Shapiro, J.P., Carpenter, J.E., 2007. Presentation of artificial
diet: effects of composition and size of prey and diet domes on egg production by
Orius insidiosus (Heteroptera: Anthocoridae). Florida Entomologist 90, 502-508.
Fernandez-Caldas, E., Iraola, V., Carnes, J., 2007. Molecular and biochemical properties of
storage mites (except Blomia species). Protein and peptide letters 14, 954-959.
Fidgett, M.J., Stinson, C.S.A., 2008. Method for rearing predatory mites. WO Patent
WO/2008/015,393.
Fouly, A., Al-Deghairi, M., Baky, N.A., 2011. Biological aspects and life tables of
Typhlodromips swirskii (Acari: Phytoseiidae) fed Bemisia tabaci (Hemiptera:
Aleyroididae). Journal of Entomology 8, 52-62.
Furman, D.P., Young, R.D., Catts, P.E., 1959. Hermetia illucens (Linnaeus) as a factor in the
natural control of Musca domestica Linnaeus. Journal of Economic Entomology 52,
917-921.
Gerson, U., Smiley, R.L., Ochoa, R., 2003. Mites (Acari) for pest control. Blackwell, Oxford.
Gerson, U., Weintraub, P.G., 2012. Mites (Acari) as a factor in greenhouse management.
Annual Review of Entomology 57, 229-247.
Goleva, I., Zebitz, C.P.W., 2013. Suitability of different pollen as alternative food for the
predatory mite Amblyseius swirskii (Acari, Phytoseiidae). Experimental and Applied
Acarology 61, 259-283.
Grafton-Cardwell, E.E., Ouyang, Y., Bugg, R.L., 1999. Leguminous cover crops to enhance
population development of Euseius tularensis (Acari: Phytoseiidae) in citrus.
Biological Control 16, 73-80.
Grenier, S., 2009. In vitro rearing of entomophagous insects-past and future trends: a
minireview. Bulletin of Insectology 62, 1-6.

References
140
Grenier, S., Bonnot, G., 1988. Development of Trichogramma dendrolimi and T. maidis
(Hymenoptera, Trichogrammatidae) in artificial media and artificial host eggs. In:
Voegele, J., (Ed.), Trichogramma and other egg parasites. Les Colloques de 1'INRA,
Paris, pp. 319-326.
Grenier, S., De Clercq, P., 2003. Comparison of artificially versus naturally reared natural
enemies and their potential for use in biological control. In: van Lenteren, J., (Ed.),
Quality control and production of biological control agents: Theory and testing
procedures. CABI Publishing, Wallingford, UK, pp. 115-131.
Guan, X., Wu, Z., Wu, T., Feng, H., 1978. Studies on rearing Trichogramma dendrolimi
Matsumura in vitro. Acta Entomologica Sinica 21, 122-126.
Guardiola, F., Codony, R., Manich, A., Rafecas, M., Boatella, J., 1995. Stability of
polyunsaturated fatty acids in egg powder processed and stored under various
conditions. Journal of Agricultural and Food Chemistry 43, 2254-2259.
Gupta, G., Rani, S., Birah, A., Raghuraman, M., 2005a. Improved artificial diet for mass
rearing of the tobacco caterpillar, Spodoptera litura (Lepidoptera: Noctuidae).
International Journal of Tropical Insect Science 25, 55-58.
Gupta, G., Rani, S., Birah, A., Raghuraman, M., 2005b. Mass rearing of the spotted bollworm,
Earias vittella (Lepidoptera: Noctuidae) on an artificial diet. International Journal of
Tropical Insect Science 25, 134-137.
Hale, O., 1973. Dried Hermetia illucens larvae (Diptera: Stratiomyidae) as a feed additive for
poultry. Journal of the Georgia Entomological Society 8, 16-20.
Helle, W., Bolland, H., Arendonk, R., Boer, R., Schulten, G., Russell, V., 1978. Genetic
evidence for biparental males in haplo-diploid predator mites (Acarina: Phytoseiidae).
Genetica 49, 165-171.
Helle, W., Sabelis, M.W., 1985. Spider mites: their biology, natural enemies and control.
Elsevier Amsterdam.
Hem, S., Toure, S., Sagbla, C., Legendre, M., 2008. Bioconversion of palm kernel meal for
aquaculture: Experiences from the forest region (Republic of Guinea). African Journal
of Biotechnology 7, 1192-1198.
Herrera, A.J.M., 1960. Investigaciones sobre la cria artificial del coccinélido Coleomegilla
maculata (De Geer). Revista Peruana de Entomologia 3, 1-6.
Heslin, L., Kopittke, R., Merritt, D., 2005. The role of insect cell lines in an artificial diet for
the parasitoid wasp, Trichogramma pretiosum. Biological Control 33, 186-193.

References
141
Hobson, R.P., 1935. On a fat-soluble growth factor required by blow-fly larvae: Identity of
the growth factor with cholesterol. Biochemical Journal 29, 2023.
Hoogerbrugge, H., Calvo, J., van Houten, Y., Bolckmans, K., 2005. Biological control of the
tobacco whitefly Bemisia tabaci with the predatory mite Amblyseius swirskii in sweet
pepper crops. IOBC/WPRS Bulletin 28, 119-122.
Hoogerbrugge, H., van Houten, Y., van Baal, E., Bolckmans, K., 2008. Alternative food
sources to enable establishment of Amblyseius swirskii (Athias-Henriot) on
chrysanthemum without pest presence. IOBC/WPRS Bulletin 32, 79.
Hoy, M.A., 1985. Recent advances in genetics and genetic improvement of the Phytoseiidae.
Annual Review of Entomology 30, 345-370.
Huang, N., Enkegaard, A., Osborne, L.S., Ramakers, P.M., Messelink, G.J., Pijnakker, J.,
Murphy, G., 2011. The banker plant method in biological control. Critical Reviews in
Plant Sciences 30, 259-278.
Huffaker, C., Vrie, M., McMurtry, J., 1969. The ecology of tetranychid mites and their natural
control. Annual Review of Entomology 14, 125-174.
Hulting, F.L., Orr, D.B., Obrycki, J.J., 1990. A computer program for calculation and
statistical comparison of intrinsic rates of increase and associated life table parameters.
Florida Entomologist 73, 601-612.
James, M., 1935. The genus Hermetia in the United States (Diptera: Stratiomyidae). Bulletin
of the Brooklyn Entomological Society 30, 165-170.
Janssen, A., Sabelis, M., 1992. Phytoseiid life-histories, local predator-prey dynamics, and
strategies for control of tetranychid mites. Experimental & Applied Acarology 14,
233-250.
Johansson, A.S., 1964. Feeding and nutrition in reproductive processes in insects. Symposia
of the Royal Entomological Society of London 2, 43-55.
Jonsson, M., Wratten, S.D., Landis, D.A., Gurr, G.M., 2008. Recent advances in conservation
biological control of arthropods by arthropods. Biological Control 45, 172-175.
Juan-Blasco, M., Qureshi, J.A., Urbaneja, A., Stansly, P.A., 2012. Predatory mite, Amblyseius
swirskii (Acari: Phytoseiidae), for biological control of Asian citrus psyllid,
Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist 95, 543-551.
Kade, N., Gueye-Ndiaye, A., Duverney, C., de Moraes, G.J., 2011. Phytoseiid mites (Acari:
Phytoseiidae) from Senegal. Acarologia 51, 133-138.

References
142
Kennett, C., Hamai, J., 1980. Oviposition and development in predaceous mites fed with
artificial and natural diets (Acari: Phytoseiidae). Entomologia Experimentalis et
Applicata 28, 116-122.
Kim, T., Ahn, J., Lee, J., 2009. Temperature-dependent developmental model of Neoseiulus
californicus (McGregor)(Acari, Phytoseiidae). Journal of Applied Entomology 133,
284-291.
King, E., Hopper, K., Powell, J., 1985. Analysis of systems for biological control of crop
arthropod pests in the US by augmentation of predators and parasites. In: Hoy, M.A.,
Herzog, D.C., Eds.), Biological Control in Agricultural IPM Systems. Academic Press,
Orlando, FL, pp. 201-227.
Koppert, 2014. http://www.koppert.com.
Kreiter, S., Tixier, M., Croft, B., Auger, P., Barret, D., 2002. Plants and leaf characteristics
influencing the predaceous mite Kampimodromus aberrans (Acari: Phytoseiidae) in
habitats surrounding vineyards. Environmental entomology 31, 648-660.
Kutner, M.H., Nachtsheim, C.J., Neter, J., Li, W., 2005. Applied linear statistical models.
McGraw-Hill, New York.
Kutuk, H., Yigit, A., 2011. Pre-establishment of Amblyseius swirskii (Athias-Henriot) (Acari:
Phytoseiidae) using Pinus brutia (Ten.) (Pinales: Pinaceae) pollen for thrips
(Thysanoptera: Thripidae) control in greenhouse peppers. International Journal of
Acarology 37, 95-101.
Lalander, C., Diener, S., Magri, M.E., Zurbrugg, C., Lindstrom, A., Vinneras, B., 2013.
Faecal sludge management with the larvae of the black soldier fly (Hermetia illucens)
- From a hygiene aspect. Science of the Total Environment 458, 312-318.
Larde, G., 1990. Recycling of coffee pulp by Hermetia illucens (Diptera, Stratiomyidae)
larvae. Biological Wastes 33, 307-310.
Lee, H.S., Gillespie, D.R., 2011. Life tables and development of Amblyseius swirskii (Acari:
Phytoseiidae) at different temperatures. Experimental and Applied Acarology 53, 17-
27.
Li, Q., Zheng, L., Cai, H., Garza, E., Yu, Z., Zhou, S., 2011a. From organic waste to biodiesel:
Black soldier fly, Hermetia illucens, makes it feasible. Fuel 90, 1545-1548.
Li, Q., Zheng, L.Y., Qiu, N., Cai, H., Tomberlin, J.K., Yu, Z.N., 2011b. Bioconversion of
dairy manure by black soldier fly (Diptera: Stratiomyidae) for biodiesel and sugar
production. Waste Management 31, 1316-1320.

References
143
Liu, Q., Tomberlin, J.K., Brady, J.A., Sanford, M.R., Yu, Z., 2008. Black soldier fly (Diptera:
Stratiomyidae) larvae reduce Escherichia coli in dairy manure. Environmental
entomology 37, 1525-1530.
Liu, W., Xie, Z., Xiao, G., Zhou, Y., Yang, D., Li, L., 1979. Rearing of the Trichogramma
dendrolimi in artificial diets. Acta Phytophyl Sin 6, 17-24.
Liu, Z.-C., Wang, Z.-Y., Sun, Y.-R., Liu, J.-F., Yang, W.-H., 1988. Studies on culturing
Anastatus sp., a parasitoid of Litchi stink bug, with artificial host eggs. In: Voegelé, J.,
Waage, J., van Lenteren, J., Eds.), Trichogramma and other egg parasites. Les
Colloques de l'INRA, pp. 353-360.
López-Arroyo, J.I., Tauber, C.A., Tauber, M.J., 1999. Effects of prey on survival,
development, and reproduction of trash-carrying chrysopids (Neuroptera:
Ceraeochrysa). Environmental entomology 28, 1183-1188.
Lorenzon, M., Pozzebon, A., Duso, C., 2012. Effects of potential food sources on biological
and demographic parameters of the predatory mites Kampimodromus aberrans,
Typhlodromus pyri and Amblyseius andersoni. Experimental and Applied Acarology
58, 259-278.
Lotka, A.J., 1907. Relation between birth rates and death rates. Science 26, 21-22.
Lü, X., Han, S., De Clercq, P., Dai, J., Li, L., 2014. Orthogonal array design for optimization
of an artificial medium for in vitro rearing of Trichogramma dendrolimi. Entomologia
Experimentalis et Applicata 152, 52-60.
Lü, X., Han, S.C., Li, L.Y., Grenier, S., De Clercq, P., 2013. The potential of trehalose to
replace insect hemolymph in artificial media for Trichogramma dendrolimi
(Hymenoptera: Trichogrammatidae). Insect Science 20, 629-636.
Lundgren, J.G., 2009a. Non-prey foods and biological control of arthropods. In: Lundgren,
J.G., (Ed.), Relationships of Natural Enemies and Non-Prey Foods. Springer, pp. 279-
307.
Lundgren, J.G., 2009b. Pollen Nutrition and Defense. In: Lundgren, J.G., (Ed.), Relationships
of Natural Enemies and Non-Prey Foods. Springer, pp. 127-139.
Maes, S., Antoons, T., Grégoire, J.-C., De Clercq, P., 2014. A semi-artificial rearing system
for the specialist predatory ladybird Cryptolaemus montrouzieri. BioControl 59, 557-
564.
Maia, A.D.N., Luiz, A.J.B., Campanhola, C., 2000. Statistical inference on associated fertility
life table parameters using jackknife technique: Computational aspects. Journal of
Economic Entomology 93, 511-518.

References
144
Maoz, Y., Gal, S., Argov, Y., Domeratzky, S., Melamed, E., Gan-Mor, S., Coll, M., Palevsky,
E., 2014. Efficacy of indigenous predatory mites (Acari: Phytoseiidae) against the
citrus rust mite Phyllocoptruta oleivora (Acari: Eriophyidae): augmentation and
conservation biological control in Israeli citrus orchards. Experimental and Applied
Acarology 63, 295-312.
Marisa, C., Sauro, S., 1990. Biological observations and life table parameters of Amblyseius
cucumeris (Oud.)(Acarina: Phytoseiidae) reared on different diets. Redia 73, 569-583.
Matsuka, M., Okada, I., 1975. Nutritional studies of an aphidophagous coccinellid, Harmonia
axyridis. I. examination of artificial diets for the larval growth with special reference
to drone honeybee powder. Bulletin Faculty of Agriculture, Tamagawa University 15,
1-9.
Matsuka, M., Watanabe, M., Niijima, K., 1982. Longevity and oviposition of vedalia beetles
on artificial diets. Environmental entomology 11, 816-819.
McCullagh, P., Nelder, J.A., 1989. Generalized linear models. Chapman and Hall/CRC,
London.
McFarlane, J., 1991. Dietary sodium, potassium and calcium requirements of the house
cricket, Acheta domesticus (l.). Comparative Biochemistry and Physiology 100A, 217-
220.
McMurtry, J., 1992. Dynamics and potential impact of ‘generalist’ phytoseiids in
agroecosystems and possibilities for establishment of exotic species. Experimental &
Applied Acarology 14, 371-382.
McMurtry, J., Croft, B., 1997. Life-styles of phytoseiid mites and their roles in biological
control. Annual Review of Entomology 42, 291-321.
McMurtry, J., Rodriguez, J., 1987. Nutritional ecology of phytoseiid mites. In: Slansky, F.,
Rodriquez, J., Eds.), Nutritional Ecology of Insects, Mites and Spiders John Wiley &
Sons Inc., New York, pp. 644.
McMurtry, J., Scriven, G., 1965. Insectary production of phytoseiid mites. Journal of
Economic Entomology 58, 282-284.
McMurtry, J., Scriven, G., 1966. Effects of artificial foods on reproduction and development
of four species of phytoseiid mites. Annals of the Entomological Society of America
59, 267-269.
McMurtry, J.A., De Moraes, G.J., Sourassou, N.F., 2013. Revision of the lifestyles of
phytoseiid mites (Acari: Phytoseiidae) and implications for biological control
strategies. Systematic and Applied Acarology 18, 297-320.

References
145
Medd, N.C., Greatrex, R.M., 2014. An evaluation of three predatory mite species for the
control of greenhouse whitefly (Trialeurodes vaporariorum). Pest Management
Science 70, 1492-1496.
Mégevand, B., Klay, A., Gnanvossou, D., Paraiso, G., 1993. Maintenance and mass rearing of
phytoseiid predators of the cassava green mite. Experimental & Applied Acarology 17,
115-128.
Messelink, G., Ramakers, P., Cortez, J., Janssen, A., 2009. How to enhance pest control by
generalist predatory mites in greenhouse crops. Proceedings of the 3rd
ISBCA,
Christchurch, New Zealand, pp. 309-318.
Messelink, G.J., Bennison, J., Alomar, O., Ingegno, B.L., Tavella, L., Shipp, L., Palevsky, E.,
Wäckers, F.L., 2014. Approaches to conserving natural enemy populations in
greenhouse crops: current methods and future prospects. BioControl 59, 377-393.
Messelink, G.J., Bloemhard, C.M.J., Cortes, J.A., Sabelis, M.W., Janssen, A., 2011.
Hyperpredation by generalist predatory mites disrupts biological control of aphids by
the aphidophagous gall midge Aphidoletes aphidimyza. Biological Control 57, 246-
252.
Messelink, G.J., Van Maanen, R., Van Holstein-Saj, R., Sabelis, M.W., Janssen, A., 2010.
Pest species diversity enhances control of spider mites and whiteflies by a generalist
phytoseiid predator. BioControl 55, 387-398.
Messelink, G.J., van Maanen, R., van Steenpaal, S.E.F., Janssen, A., 2008. Biological control
of thrips and whiteflies by a shared predator: Two pests are better than one. Biological
Control 44, 372-379.
Messelink, G.J., van Steenpaal, S.E., Ramakers, P.M., 2006. Evaluation of phytoseiid
predators for control of western flower thrips on greenhouse cucumber. BioControl 51,
753-768.
Meyer, J.S., Ingersoll, C.G., McDonald, L.L., Boyce, M.S., 1986. Estimating uncertainty in
population growth rates: jackknife vs. bootstrap techniques. Ecology 67, 1156-1166.
Midthassel, A., Leather, S.R., Baxter, I.H., 2013. Life table parameters and capture success
ratio studies of Typhlodromips swirskii (Acari: Phytoseiidae) to the factitious prey
Suidasia medanensis (Acari: Suidasidae). Experimental and Applied Acarology 61,
69-78.
Momen, F., 2009. Potential of three species of predatory phytoseiid mites as biological
control agents of the peach silver mite, Aculus fockeui (Acari: Phytoseiidae and
Eriophyidae). Acta Phytopathologica et Entomologica Hungarica 44, 151-158.

References
146
Momen, F., Abdel-Khalek, A., 2009. Cannibalism and intraguild predation in the phytoseiid
mites Typhlodromips swirskii, Euseius scutalis and Typhlodromus athiasae (Acari:
Phytoseiidae). Acarina 17, 223-229.
Momen, F., El-Saway, S., 1993. Biology and feeding behaviour of the predatory mite,
Amblyseius swirskii (Acari: Phytoseiidae). Acarologia 34, 199-204.
Momen, F.M., Abdel-Khalek, A., 2008. Effect of the tomato rust mite Aculops lycopersici
(Acari: Eriophyidae) on the development and reproduction of three predatory
phytoseiid mites. International Journal of Tropical Insect Science 28, 53-57.
Momen, F.M., El-Laithy, A.Y., 2007. Suitability of the flour moth Ephestia kuehniella
(Lepidoptera: Pyralidae) for three predatory phytoseiid mites (Acari: Phytoseiidae) in
Egypt. International Journal of Tropical Insect Science 27, 102-107.
Moraes, G.J., McMurtry, J., Denmark, H., Campos, C., 2004. A revised catalog of the mite
family Phytoseiidae (Zootaxa 434). Magnolia Press New Zealand.
Moraes, G.J., McMurtry, J.A., Denmark, H.A., 1986. A catalog of the mite family
Phytoseiidae: references to taxonomy, synonymy, distribution and habitat. Empresa
Brasileira de Pesquisa Agropecuaria, Bras'ilia, Brazil.
Morales-Ramos, J.A., Rojas, M.G., Coudron, T.A., 2014. Artificial diet development for
entomophagous arthropods. In: Morales-Ramos, J., Rojas, M.G., Shapiro-Ilan, D.,
Eds.), Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens.
Academic Press, Elsevier, pp. 203-240.
Mori, H., 1977. Notes on the biology of phytoseiid mites (Acarina: Phytoseiidae) as predators
of tetranychid mites in Japan. Review of Plant Protection Research 10, 91-101.
Myers, H.M., Tomberlin, J.K., Lambert, B.D., Kattes, D., 2008. Development of black soldier
fly (Diptera : Stratiomyidae) larvae fed dairy manure. Environmental entomology 37,
11-15.
Nettles, W.C., 1990. In vitro rearing of parasitoids: role of host factors in nutrition. Archives
of Insect Biochemistry and Physiology 13, 167-175.
Newton, G., Booram, C., Barker, R., Hale, O., 1977. Dried Hermetia illucens larvae meal as a
supplement for swine. Journal of Animal Science 44, 395-400.
Newton, G., Sheppard, D., Watson, D., Burtle, G., Dove, C., Tomberlin, J., Thelen, E., 2005.
The black soldier fly, Hermetia illucens, as a manure management/resource recovery
tool. Symposium on the state of the science of animal manure and waste management,
pp. 5-7.

References
147
Nguyen, T.T.X., Tomberlin, J.K., Vanlaerhoven, S., 2013. Influence of resources on Hermetia
illucens (Diptera: Stratiomyidae) larval development. Journal of Medical Entomology
50, 898-906.
Nichols, C., Altieri, M., 2004. The agroecological engineering: for pest management. In: Gurr,
G., Wratten, S., Altieri, M., Eds.), Ecological engineering for pest management.
CSIRO Publishing, Collingwood, pp. 33-54.
Nomikou, M., Janssen, A., Sabelis, M.W., 2003a. Phytoseiid predator of whitefly feeds on
plant tissue. Experimental and Applied Acarology 31, 27-36.
Nomikou, M., Janssen, A., Sabelis, M.W., 2003b. Phytoseiid predators of whiteflies feed and
reproduce on non-prey food sources. Experimental and Applied Acarology 31, 15-26.
Nomikou, M., Janssen, A., Schraag, R., Sabelis, M.W., 2001. Phytoseiid predators as
potential biological control agents for Bemisia tabaci. Experimental and Applied
Acarology 25, 271-291.
Nomikou, M., Janssen, A., Schraag, R., Sabelis, M.W., 2002. Phytoseiid predators suppress
populations of Bemisia tabaci on cucumber plants with alternative food. Experimental
and Applied Acarology 27, 57-68.
Nomikou, M., Janssen, A., Schraag, R., Sabelis, M.W., 2004. Vulnerability of Bemisia tabaci
immatures to phytoseiid predators: Consequences for oviposition and influence of
alternative food. Entomologia Experimentalis et Applicata 110, 95-102.
Nomikou, M., Sabelis, M.W., Janssen, A., 2010. Pollen subsidies promote whitefly control
through the numerical response of predatory mites. BioControl 55, 253-260.
Ochieng, R., Oloo, G., Amboga, E., 1987. An artificial diet for rearing the phytoseiid mite,
Amblyseius teke Pritchard and Baker. Experimental and Applied Acarology 3, 169-173.
Ogawa, Y., Osakabe, M., 2008. Development, long-term survival, and the maintenance of
fertility in Neoseiulus californicus (Acari: Phytoseiidae) reared on an artificial diet.
Experimental and Applied Acarology 45, 123-136.
Ogura, N., Hosoda, R., 1995. Rearing of a coleopterous predator, Trogossita japonica (Col.:
Trogossitidae), on artificial diets. Entomophaga 40, 371-378.
Onzo, A., Houedokoho, A.F., Hanna, R., 2012. Potential of the predatory mite, Amblyseius
swirskii to suppress the broad mite, Polyphagotarsonemus latus on the gboma
eggplant, Solanum macrocarpon. Journal of Insect Science 12, 1-11.
Opit, G.P., Nechols, J.R., Margolies, D.C., Williams, K.A., 2005. Survival, horizontal
distribution, and economics of releasing predatory mites (Acari: Phytoseiidae) using
mechanical blowers. Biological Control 33, 344-351.

References
148
Overmeer, W., 1985. Alternative prey and other food resources. In: Helle, W., Sabelis, M.W.,
Eds.), Spider mites: their biology, natural enemies and control. Elsevier, Amsterdam,
The Netherlands, pp. 131-140.
Pappas, M.L., Broufas, G.D., Tsarsitalidou, O.K., Koveos, D.S., 2011. Development and
reproduction of the lacewings Dichochrysa flavifrons and Dichochrysa zelleri
(Neuroptera: Chrysopidae) fed on two prey species. Annals of the Entomological
Society of America 104, 726-732.
Parella, M., Heinz, K., Nunney, L., 1992. Biological control through augmentative releases of
natural enemies: a strategy whose time has come. American Entomologist 38, 172-180.
Park, H.H., Shipp, L., Buitenhuis, R., 2010. Predation, development, and oviposition by the
predatory mite Amblyseius swirskii (Acari: Phytoseiidae) on tomato russet mite (Acari:
Eriophyidae). Journal of Economic Entomology 103, 563-569.
Park, H.H., Shipp, L., Buitenhuis, R., Ahn, J.J., 2011. Life history parameters of a
commercially available Amblyseius swirskii (Acari: Phytoseiidae) fed on cattail
(Typha latifolia) pollen and tomato russet mite (Aculops lycopersici). Journal of Asia-
Pacific Entomology 14, 497-501.
Parra, J.R., 2012. The evolution of artificial diets and their interactions in science and
technology. In: Panizzi, A.R., Parra, J.R., Eds.), Insect bioecology and nutrition for
integrated pest management. CRC Press, pp. 51-92.
Ragusa, S., Swirski, E., 1977. Feeding habits, post-embryonic and adult survival, mating,
virility and fecundity of the predacious mite Amblyseius swirskii (Acarina:
Phytoseiidae) on some coccids and mealybugs. Entomophaga 22, 383-392.
Ramakers, P., Dissevelt, M., Peeters, K., 1989. Large scale introductions of phytoseiid
predators to control thrips on cucumber. Mededelingen van de Faculteit
Landbouwwetenschappen, Rijksuniversiteit Gent 54, 923-929.
Ramakers, P., Voet, S., 1996. Introduction of Amblyseius degenerans for thrips control in
sweet peppers with potted castor beans as banker plants. IOBC/WPRS Bulletin 19,
127-130.
Ramakers, P.M., 1995. Biological control using oligophagous predators. In: Parker, B.L.,
Skinner, M., Lewis, T., Eds.), Thrips Biology and Management. Springer, New York,
USA, pp. 225-229.
Ranabhat, N.B., Goleva, I., Zebitz, C.P., 2014. Life tables of Neoseiulus cucumeris
exclusively fed with seven different pollens. BioControl 59, 195-203.

References
149
Rencken, I., Pringle, K., 1998. Developmental biology of Amblyseius californicus
(McGregor)(Acarina: Phytoseiidae), a predator of tetranychid mites, at three
temperatures. African Entomology 6, 41-45.
Riddick, E.W., 2009. Benefits and limitations of factitious prey and artificial diets on life
parameters of predatory beetles, bugs, and lacewings: a mini-review. BioControl 54,
325-339.
Riudavets, J., Arno, J., Castañé, C., 2006. Rearing predatory bugs with the brine shrimp
Artemia sp. as alternative prey food. IOBC/WPRS Bulletin 29, 235-240.
Romeih, A.H.M., El-Saidy, E.M.A., El-Arnaouty, S.A., 2004. Suitability of Ephestia
kuehneilla and Corycera cephalonica eggs as alternative preys for rearing predatory
mites. Egyptian Journal of Biological Pest Control 14, 101-105.
Saito, Y., 2010. Plant mites and sociality. Springer.
Sato, Y., Mochizuki, A., 2011. Risk assessment of non-target effects caused by releasing two
exotic phytoseiid mites in Japan: can an indigenous phytoseiid mite become IG prey?
Experimental and Applied Acarology 54, 319-329.
Schausberger, P., 2003. Cannibalism among phytoseiid mites: a review. Experimental &
Applied Acarology 29, 173-191.
Schausberger, P., 2004. Ontogenetic isolation favours sibling cannibalism in mites. Animal
Behaviour 67, 1031-1035.
Schliesske, J., 1981. Ueber die Technik der Massenanzucht von Raubmilben (Acari:
Phytoseiidae) unter kontrollierten bedingungen. Mededelingen van de Faculteit
Landbouwwetenschappen Rijksuniversiteit Gent 46, 511-517.
Schumacher, S.K., Marsh, T.L., Williams, K.A., 2006. Optimal pest control in greenhouse
production of ornamental crops. Agricultural Economics 34, 39-50.
Shehata, K., Weismann, L., 1972. Rearing the predacious mite Phytoseiulus persimilis Athias-
Henriot on artificial diet (Acarina: Phytoseiidae). Biologia Bratislava 27, 609-615.
Sheppard, D.C., Newton, G.L., Thompson, S.A., Savage, S., 1994. A value-added manure
management-system using the black soldier fly. Bioresource Technology 50, 275-279.
Shih, C.-I.T., Chang, H., Hsu, P., Hwang, Y., 1993. Responses of Amblyseius ovalis
(Evans)(Acarina: Phytoseiidae) to natural food resouces and two artificial diets.
Experimental and Applied Acarology 17, 503-519.
Singh, P., 1977. Artificial diets for insects, mites and spiders. Plenum Press, New York, NY.

References
150
Skirvin, D., Kravar-Garde, L., Reynolds, K., Jones, J., De Courcy Williams, M., 2006. The
influence of pollen on combining predators to control Frankliniella occidentalis in
ornamental chrysanthemum crops. Biocontrol Science and Technology 16, 99-105.
Specty, O., Febvay, G., Grenier, S., Delobel, B., Piotte, C., Pageaux, J.-F., Ferran, A.,
Guillaud, J., 2003. Nutritional plasticity of the predatory ladybeetle Harmonia axyridis
(Coleoptera: Coccinellidae): Comparison between natural and substitution prey.
Archives of Insect Biochemistry and Physiology 52, 81-91.
St-Hilaire, S., Cranfill, K., McGuire, M.A., Mosley, E.E., Tomberlin, J.K., Newton, L., Sealey,
W., Sheppard, C., Irving, S., 2007. Fish offal recycling by the black soldier fly
produces a foodstuff high in omega-3 fatty acids. Journal of the World Aquaculture
Society 38, 309-313.
Stinner, R., 1977. Efficacy of inundative releases. Annual Review of Entomology 22, 515-531.
Strand, M., Vinson, S., 1985. In vitro culture of Trichogramma pretiosum on an artificial
medium. Entomologia Experimentalis et Applicata 39, 203-209.
Strangways-Dixon, J., 1959. Hormones, selective feeding and reproduction in Calliphora.
Symposium on the Ontogenetic Development of Insects, Prague, pp. 137-139.
Swirski, E., Amitai, S., Dorzia, N., 1967. Laboratory studies on the feeding, development and
reproduction of the predaceous mites Amblyseius rubini Swirski and Amitai and
Amblyseius swirskii Athias (Acarina: Phytoseidae) on various kinds of food substances.
Israel Journal of Agricultural Research 17, 101-119.
Swirski, E., Amitai, S., Dorzia, N., 1970. Laboratory studies on the feeding habits, post-
embryonic survival and oviposition of the predaceous mites Amblyseius chilenensis
Dosse and Amblyseius hibisci Chant (Acarina: Phytoseiidae) on various kinds of food
substances. Entomophaga 15, 93-106.
Syngenta, 2014. http://www3.syngenta.com/global/Bioline/en/about-us/Pages/About-us.aspx.
Tanigoshi, L., Mégevand, B., Yaninek, J., 1993. Non-prey food for subsistence of Amblyseius
idaeus (Acari: Phytoseiidae) on cassava in Africa. Experimental & Applied Acarology
17, 91-96.
Thompson, S., 1999. Nutrition and culture of entomophagous insects. Annual Review of
Entomology 44, 561-592.
Tomberlin, J.K., Sheppard, D.C., Joyce, J.A., 2002. Selected life-history traits of black soldier
flies (Diptera : Stratiomyidae) reared on three artificial diets. Annals of the
Entomological Society of America 95, 379-386.

References
151
Vacante, V., Cocuzza, G., De Clercq, P., Van De Veire, M., Tirry, L., 1997. Development and
survival of Orius albidipennis and O. laevigatus (Het.: Anthocoridae) on various diets.
BioControl 42, 493-498.
van Houten, Y.M., Rijn, P.C., Tanigoshi, L.K., Stratum, P., Bruin, J., 1995. Preselection of
predatory mites to improve year-round biological control of western flower thrips in
greenhouse crops. Entomologia Experimentalis et Applicata 74, 225-234.
Van Huis, A., Van Itterbeeck, J., Klunder, H., Mertens, E., Halloran, A., Muir, G., Vantomme,
P., 2013. Edible insects: future prospects for food and feed security. Food and
agriculture organization of the United Nations (FAO).
van Lenteren, J., 2012. The state of commercial augmentative biological control: plenty of
natural enemies, but a frustrating lack of uptake. BioControl 57, 1-20.
van Lenteren, J.C., 2003. Quality control and production of biological control agents: theory
and testing procedures. CABI Publishing, Wallingford, UK
van Maanen, R., Vila, E., Sabelis, M.W., Janssen, A., 2010. Biological control of broad mites
(Polyphagotarsonemus latus) with the generalist predator Amblyseius swirskii.
Experimental and Applied Acarology 52, 29-34.
van Rijn, P.C., Tanigoshi, L.K., 1999a. The contribution of extrafloral nectar to survival and
reproduction of the predatory mite Iphiseius degenerans on Ricinus communis.
Experimental & Applied Acarology 23, 281-296.
van Rijn, P.C., Tanigoshi, L.K., 1999b. Pollen as food for the predatory mites Iphiseius
degenerans and Neoseiulus cucumeris (Acari: Phytoseiidae): dietary range and life
history. Experimental & Applied Acarology 23, 785-802.
van Rijn, P.C., van Houten, Y.M., Sabelis, M.W., 2002. How plants benefit from providing
food to predators even when it is also edible to herbivores. Ecology 83, 2664-2679.
Van Stappen, G., 1996. Introduction, biology and ecology of Artemia. In: Lavens, P.,
Sorgeloos, P., Eds.), Manual on the production and use of live food for aquaculture.
FAO, pp. 79-106.
Vandekerkhove, B., Baal, E.V., Bolckmans, K., De Clercq, P., 2006. Effect of diet and mating
status on ovarian development and oviposition in the polyphagous predator
Macrolophus caliginosus (Heteroptera: Miridae). Biological Control 39, 532-538.
Vandekerkhove, B., Parmentier, L., Van Stappen, G., Grenier, S., Febvay, G., Rey, M., De
Clercq, P., 2009. Artemia cysts as an alternative food for the predatory bug
Macrolophus pygmaeus. Journal of Applied Entomology 133, 133-142.

References
152
Vangansbeke, D., Nguyen, D.T., Audenaert, J., Verhoeven, R., Gobin, B., Tirry, L., De
Clercq, P., 2014. Performance of the predatory mite Amblydromalus limonicus on
factitious foods. BioControl 59, 67-77.
Vantornhout, I., Minnaert, H., Tirry, L., De Clercq, P., 2004. Effect of pollen, natural prey
and factitious prey on the development of Iphiseius degenerans. BioControl 49, 627-
644.
Wäckers, F.L., van Rijn, P.C.J., 2005. Food for protection: an introduction. In: Wäckers, F.L.,
van Rijn, P.C.J., Bruin, J., Eds.), Plant-Provided Food for Carnivorous Insects: A
Protective Mutualism and its Applications. Cambridge University Press, Cambridge.
Wade, M.R., Zalucki, M.P., Wratten, S.D., Robinson, K.A., 2008. Conservation biological
control of arthropods using artificial food sprays: current status and future challenges.
Biological Control 45, 185-199.
Weintraub, P.G., Pivonia, S., Steinberg, S., 2011. How many Orius laevigatus are needed for
effective western flower thrips, Frankliniella occidentalis, management in sweet
pepper? Crop Protection 30, 1443-1448.
Wimmer, D., Hoffmann, D., Schausberger, P., 2008. Prey suitability of western flower thrips,
Frankliniella occidentalis, and onion thrips, Thrips tabaci, for the predatory mite
Amblyseius swirskii. Biocontrol Science and Technology 18, 533-542.
Woolley, T.A., 1988. Acarology: mites and human welfare. John Wiley & Sons, Inc.
Xia, C., Zhang, Y.-X., Ji, J., Lin, J.-Z., 2011. Experimental life table for population of
Amblyseius swirskii (Athias-Henriot) fed on Tetranychus truncatus (Ehara). Fujian
Journal of Agricultural Sciences 26, 415-419.
Xiao, Y., Osborne, L.S., Chen, J., McKenzie, C.L., 2013. Functional responses and prey-stage
preferences of a predatory gall midge and two predacious mites with twospotted spider
mites, Tetranychus urticae, as host. Journal of Insect Science 13, 8.
Xie, Z.-N., Nettles, W.C., Morrison, R.K., Irie, K., Vinson, S.B., 1986. Effect of ovipositional
stimulants and diets on the growth and development of Trichogramma pretiosum in
vitro. Entomologia Experimentalis et Applicata 42, 119-124.
Xie, Z.-N., Wu, Z.-X., Nettles Jr, W.C., Saldaña, G., Nordlund, D.A., 1997. In vitro culture of
Trichogramma spp. on artificial diets containing yeast extract and ultracentrifuged
chicken egg yolk but devoid of insect. Biological Control 8, 107-110.
Xu, X.N., Enkegaard, A., 2010. Prey preference of the predatory mite, Amblyseius swirskii
between first instar western flower thrips Frankliniella occidentalis and nymphs of the
twospotted spider mite Tetranychus urticae. Journal of Insect Science 10, 149.

References
153
Yousef, A.-t., El-keifl, A.H., Metwally, A.M., 1982. Zur wirkung von temperatur und
photoperiode auf die entwicklung, ernährung und eiablage der raubmilbe Amblyseius
swirskii Ath.-Henr. (Acari, Gamasida, Phytoseiidae). Anzeiger für Schädlingskunde,
Pflanzenschutz, Umweltschutz 55, 107-109.
Yu, G.H., Cheng, P., Chen, Y.H., Li, Y.J., Yang, Z.H., Chen, Y.F., Tomberlin, J.K., 2011.
Inoculating poultry manure with companion bacteria influences growth and
development of black soldier fly (Diptera: Stratiomyidae) larvae. Environmental
entomology 40, 30-35.
Zaher, M., El-Bishlawy, S., Ali, F., 2007. Some ecological and biological studies on
Typhlodromips swirskii (Athias-Henriot)(Acari: Phytoseiidae). Acarines 1, 23-27.
Zannou, I.D., De Moraes, G.J., Ueckermann, E.A., Oliveira, A.R., Yaninek, J.S., Hanna, R.,
2007. Phytoseiid mites of the subtribe Amblyseiina (Acari: Phytoseiidae: Amblyseiini)
from sub-Saharan Africa. Zootaxa 1550, 1-47.
Zannou, I.D., Hanna, R., 2011. Clarifying the identity of Amblyseius swirskii and Amblyseius
rykei (Acari: Phytoseiidae): are they two distinct species or two populations of one
species? Experimental and Applied Acarology 53, 339-347.
Zdarkova, E., Strohalm, J., Houska, M., 1999. Effect of high pressure on Carpoglyphus lactis
L. (Acari: Carpoglyphidae). Czech Journal of Food Sciences 17, 235-237.
Zhang, Y., Ji, J., Lin, J., Chen, X., Saito, Y., 2014. Female performance towards offspring
under starved conditions in four phytoseiid species (Acari, Phytoseiidae).
Experimental and Applied Acarology DOI: 10.1007/s10493-014-9847-2.
Zhang, Z.-Q., 2003. Mites of greenhouses: identification, biology and control. CABI
Publishing, Wallingford, UK
Zhou, F., Tomberlin, J.K., Zheng, L.Y., Yu, Z.N., Zhang, J.B., 2013. Developmental and
waste reduction plasticity of three black soldier fly strains (Diptera: Stratiomyidae)
raised on different livestock manures. Journal of Medical Entomology 50, 1224-1230.

References
154

155
CURRICULUM VITAE
Personalia
Name: Nguyen Duc Tung
Date of birth: 8th
March 1979
Place of birth: Hanoi
Nationality: Vietnamese
E-mail: [email protected]
Education
2011-2014: PhD student at the Laboratory of Agrozoology, Department of Crop Protection,
Faculty of Bioscience Engineering, Ghent University, Belgium
Thesis: Artificial and factitious foods for the production and population
enhancement of phytoseiid predatory mites
Promoter: Prof. dr. ir. P. De Clercq
2007-2009: M.Sc. in Entomology
Crop Protection Cluster, College of Agriculture, University of the Philippines Los
Baños, Philippines
Thesis: Efficacy of the predatory mite Neoseiulus longispinosus (Evans) (Acari:
Phytoseiidae) to control the two-spotted spider mite Tetranychus urticae Koch
(Acari: Tetranychidae) in green bean and acceptability of the technology to
farmers in Hanoi, Vietnam.
Promoter: Dr. Celia DR. Medina

Curriculum vitae
156
1999-2003: B.Sc. in Plant Protection
College of Natural Resources and Environment, South China Agricultural
University, China
Thesis: Study on parasitic and reproductive characteristics of Chrysonotomyia
formosa (Westwood) (Hymenoptera: Eulophidae) parasitoid of vegetable leaf-
miner Liriomyza sativae Blanchard (Diptera: Agromyzidae).
Promoter: Prof. Dr. Xu Zai Fu
Professional career
2004-present: Lecturer at the Entomology Department, Faculty of Agronomy, Vietnam
National University of Agriculture, Hanoi, Vietnam.
Publications
Duc Tung Nguyen, Dominiek Vangansbeke, Xin Lu & Patrick De Clercq. 2013.
Development and reproduction of the predatory mite Amblyseius swirskii on artificial
diets. BioControl 58: 369-377.
Dominiek Vangansbeke, Lien De Schrijver, Thomas Spranghers, Joachim Audenaert, Ruth
Verhoeven, Duc Tung Nguyen, Bruno Gobin, Luc Tirry & Patrick De Clercq. 2013.
Alternating temperatures affect life table parameters of Phytoseiulus persimilis,
Neoseiulus californicus (Acari: Phytoseiidae) and their prey Tetranychus urticae (Acari:
Tetranychidae). Experimental and Applied Acarology 61: 285-298.
Dominiek Vangansbeke, Duc Tung Nguyen, Joachim Audenaert, Ruth Verhoeven, Bruno
Gobin, Luc Tirry & Patrick De Clercq. 2014. Performance of the predatory mite
Amblydromalus limonicus on factitious foods. BioControl 59: 67-77.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2014. Artificial and
factitious foods support the development and reproduction of the predatory mite
Amblyseius swirskii. Experimental and Applied Acarology 62: 181-194.

Curriculum vitae
157
Dominiek Vangansbeke, Duc Tung Nguyen, Joachim Audenaert, Ruth Verhoeven, Koen
Deforce, Bruno Gobin, Luc Tirry & Patrick De Clercq. 2014. Diet-dependent cannibalism
in the omnivorous phytoseiid mite Amblydromalus limonicus. Biological Control 74: 30-
35.
Dominiek Vangansbeke, Duc Tung Nguyen, Joachim Audenaert, Ruth Verhoeven, Bruno
Gobin, Luc Tirry & Patrick De Clercq. 2014. Food supplementation affects interactions
between a phytoseiid predator and its omnivorous prey. Biological Control 76: 95-100.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2014. Solid artificial diets
for the phytoseiid predator Amblyseius swirskii. BioControl, 59: 719-727.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2015. Performance of four
species of phytoseiid mites on artificial and natural diets. Biological Control, 80: 56-62.
Duc Tung Nguyen, Vincent Bouguet, Thomas Spranghers, Dominiek Vangansbeke & Patrick
De Clercq. 2014. Beneficial effect of supplementing an artificial diet for Amblyseius
swirskii with Hermetia illucens hemolymph. Journal of Applied Entomology, DOI:
10.1111/jen.12188.
Duc Tung Nguyen, Dominiek Vangansbeke and Patrick De Clercq. 2014. Artificial diets
support the development and reproduction of the predatory mite Amblyseius swirskii.
IOBC-WPRS Bulletin 102: 215-218.
Oral presentations
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2013. Performance of
Amblyseius swirskii Athias-Henriot and Amblydromalus limonicus Garman
(Mesostigmata: Phytoseiidae) on factitious foods and pollen. 4th
International Symposium
on Biological Control of Arthropods, March 4-8, 2013, Pucon, Chile.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2013. Artificial diets for
rearing the predatory mite Amblyseius swirskii (Acari: Phytoseiidae). 65th
International
Symposium on Crop Protection, May 21, 2013, Ghent, Belgium.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2014. Liquid and solid
artificial diets support the development and reproduction of the predatory mite

Curriculum vitae
158
Amblyseius swirskii. 14th
International Congress of Acarology, July 14-18, 2014, Kyoto,
Japan.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2014. Artificial diets
support the development and reproduction of the predatory mite Amblyseius swirskii.
IOBC Working Group on "Integrated Control in Protected Crops: Temperate Climate",
September 14-18, 2014, Gent, Belgium.
Poster presentations
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2013. Artificial and
factitious foods support the development and Reproduction of the predatory mite
Amblyseius swirskii. 4th
Meeting of the IOBC Working Group: “Integrated control of
plant-feeding mites”. September 9-12, 2013, Paphos, Cyprus.
Duc Tung Nguyen, Dominiek Vangansbeke & Patrick De Clercq. 2013. Development and
reproduction of the predatory mite Amblyseius swirskii (Athias-Henriot) (Acari:
Phytoseiidae) on artificial diets. 13th
Workshop of the IOBC Global Working Group on
Mass Rearing and Quality Assurance: “Emerging Opportunities for the Mass Production
& Quality Assurance of Invertebrates”. November 6-8, 2013, Bangalore, India.