pathogenesis of spontaneous preterm birth
DESCRIPTION
preterm birthTRANSCRIPT

599
Pathogenesis of Spontaneous Preterm BirthCATALIN S. BUHIMSCHI, MD | JANE E. NORMAN, MD
39
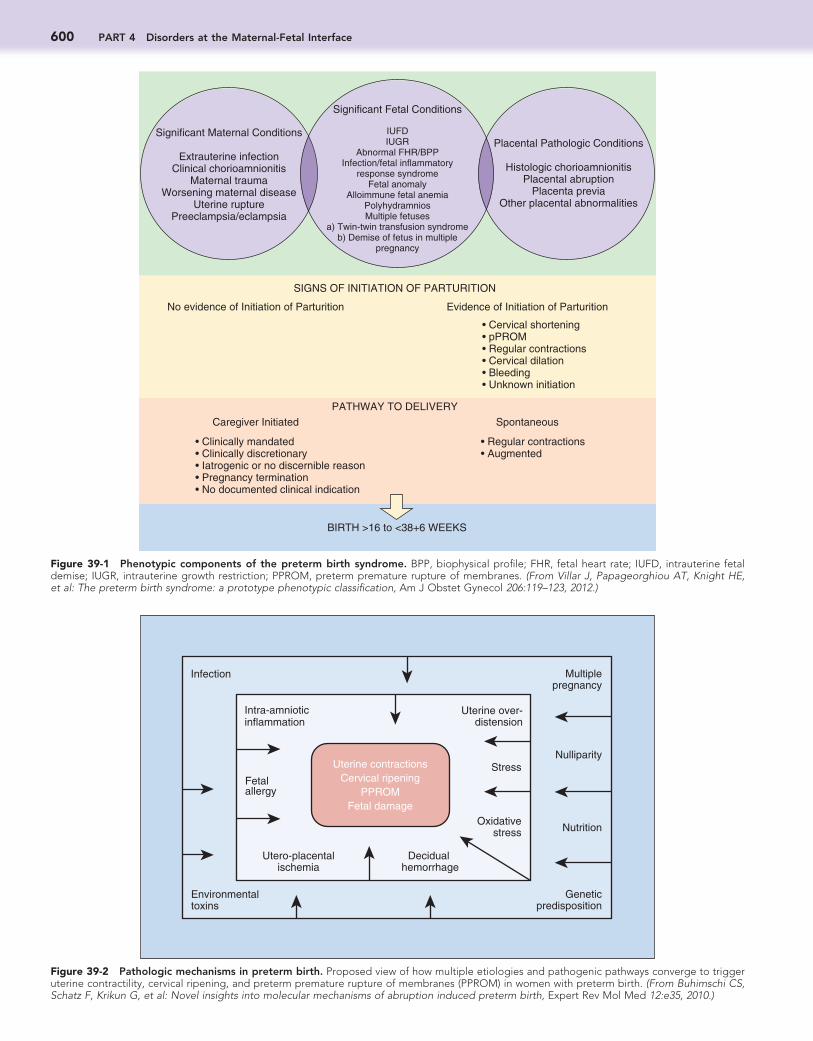
Preterm Birth Syndrome: New Phenotypic ClassificationPreterm birth (PTB) occurs between fetal viability and 37 com-pleted weeks of gestation.1 The definition of viability is contro-versial because of the increasing frequency of survival at progressively lower gestational ages. Most countries define it as a lower limit of 20 to 22 weeks, but this varies, preventing straightforward comparison of reported rates of neonatal mor-tality and morbidity.2 A recent influential report has suggested that a less arbitrary definition of PTB would include all births (including live births, stillbirths, and pregnancy terminations) occurring from 16 weeks 0 days to 38 weeks 6 days (i.e., 112 to 272 days).3 The rationale for the latter limit is that births between 37 and 39 weeks are associated with greater short- and long-term morbidity than those after 39 weeks,4 whereas the rationale for the early limit is that the pathologies inducing spontaneous abortion between 16 and 20 weeks are similar to those inducing PTB at a later gestation. Where accurate record-ing of gestational age is not possible—for example, in resource-poor countries—a birth weight of 500 g has historically been used to define the lower limit of viability. However, this approach leads to inaccuracies, because viable neonates born after 24 weeks may be affected by intrauterine growth restriction (IUGR), and some pre-viable infants may weigh more than 500 g.
Worldwide, approximately 1.1 million neonates die from prematurity-related complications.5 Rates of PTB vary around the world, with the United States having among the highest incidences.6 In 2010, the PTB rate in the United States was around 12.0%, representing a progressive decline over the past 4 years.7 Its decline from 2008 to 2010 was most noticeable among infants born at 34 to 36 weeks (late preterm). However, the percentage of infants born at less than 34 weeks also dropped, from 3.56% to 3.50%. A reduction in PTB rates was seen for most age, race, and ethnic groupings. By contrast, the birth rate for infants having low birth weight (LBW; <2500 g) was unchanged from 2008 to 2010, at 8.15%. Regretfully, the percentage of newborns delivered at very low birth weight (1500 g) declined only minimally, from 1.46% in 2008 to 1.45% in 2010. This is significant, as very-low-birth-weight premature newborns are at the highest risk for early death or disability.8
Traditional classification systems categorize PTBs as either spontaneous or indicated. Spontaneous preterm labor can occur either with intact membranes or with prelabor (prema-ture) rupture of the fetal membranes (PROM). Indicated PTBs
can result from induction of preterm labor or from preterm cesarean delivery for maternal or fetal indications (e.g., pre-eclampsia, IUGR).9 The Global Alliance to Prevent Prematurity and Stillbirth (GAPPS) has recently proposed an alternative classification system that is likely to be very influential.10 Under this paradigm, PTB is categorized according to the clinical phe-notype consisting of (1) one or more conditions of the mother, placenta, or fetus; (2) the presence or absence of signs of par-turition; and (3) the pathway to delivery (spontaneous or care-giver initiated) (Fig. 39-1). There is some overlap between the GAPPS classification system and the traditional criteria—for example, indicated preterm delivery in the previous system cor-responds to caregiver-initiated parturition in the new system, and it normally occurs in the absence of parturition. In this chapter, we address PTB under the new classification system, and we describe it according to the maternal, placental, and fetal phenotype.10 Under the new classification system, PTB rates include infants born between 37 + 0 and 38 + 6 weeks of preg-nancy, and thus these rates are about 28% higher than previ-ously. Unless otherwise indicated, it is assumed that for the conditions described in this chapter, there is evidence of partu-rition, and the pathway to delivery is spontaneous. The mecha-nisms of disease responsible for indicated preterm deliveries with caregiver-initiated parturition are discussed in other chap-ters. About 75% all PTBs have a spontaneous pathway to deliv-ery: About 45% (23.2% to 64.1%) are from preterm labor with intact membranes, and about 30% (7.1% to 51.2%) are from preterm labor after PROM.11-14
Mechanisms of Spontaneous Preterm BirthTHE COMMON PATHWAY
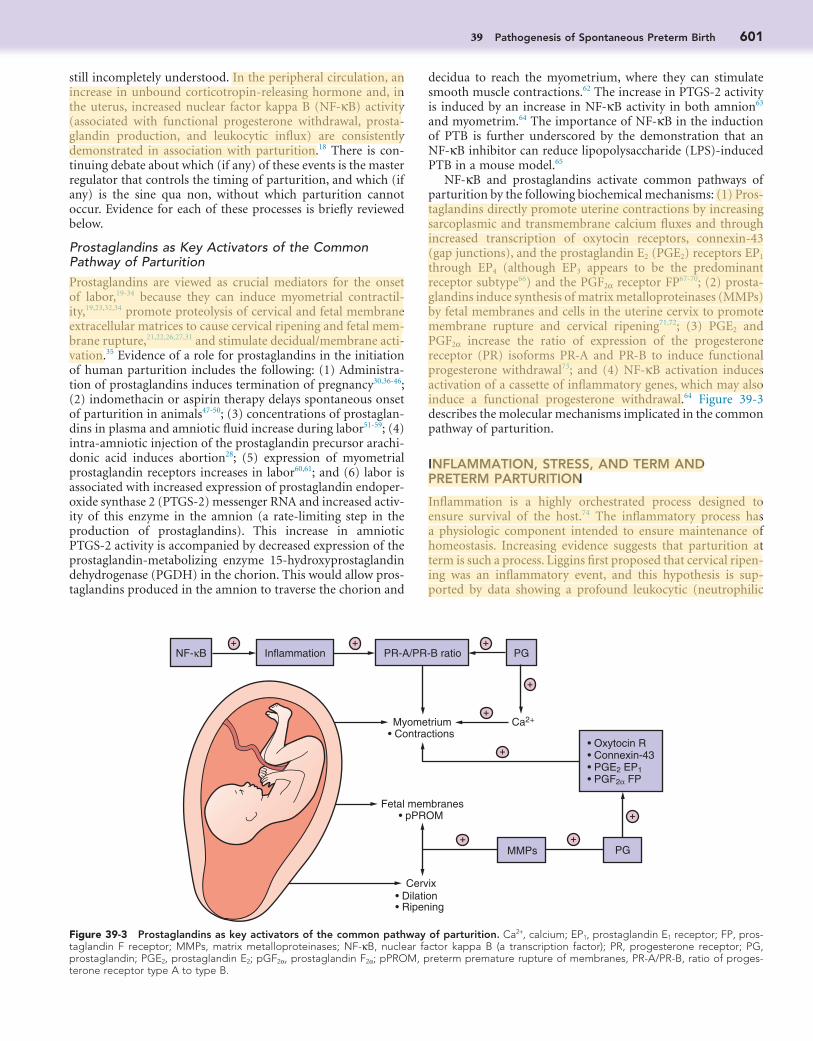
Term and preterm labor share common pathways, which include increased uterine contractility, cervical ripening, and mem-brane rupture leading to fetal prematurity and damage.15 However, whereas term birth results from physiologic activation of these common pathways, PTB results from a disease process (or pathologic activation) (Fig. 39-2) that activates one or more of the components of the common pathway via similar or alter-native mechanisms.16,17
The common pathway of parturition includes anatomic, bio-chemical, immunologic, endocrinologic, and clinical changes.16 Although the anatomic and clinical events have been studied in detail, the biochemical, immunologic, and endocrine events are
600 PART 4 Disorders at the Maternal-Fetal Interface
Figure 39-1 Phenotypic components of the preterm birth syndrome. BPP, biophysical profile; FHR, fetal heart rate; IUFD, intrauterine fetal demise; IUGR, intrauterine growth restriction; PPROM, preterm premature rupture of membranes. (From Villar J, Papageorghiou AT, Knight HE, et al: The preterm birth syndrome: a prototype phenotypic classification, Am J Obstet Gynecol 206:119–123, 2012.)
Significant Maternal Conditions
Extrauterine infectionClinical chorioamnionitis
Maternal traumaWorsening maternal disease
Uterine rupturePreeclampsia/eclampsia
Significant Fetal Conditions
IUFDIUGR
Abnormal FHR/BPPInfection/fetal inflammatory
response syndromeFetal anomaly
Alloimmune fetal anemiaPolyhydramniosMultiple fetuses
a) Twin-twin transfusion syndromeb) Demise of fetus in multiple
pregnancy
SIGNS OF INITIATION OF PARTURITION
PATHWAY TO DELIVERY
BIRTH >16 to <38+6 WEEKS
Caregiver Initiated Spontaneous
• Regular contractions• Augmented
• Clinically mandated• Clinically discretionary• Iatrogenic or no discernible reason• Pregnancy termination• No documented clinical indication
No evidence of Initiation of Parturition Evidence of Initiation of Parturition
• Cervical shortening• pPROM• Regular contractions• Cervical dilation• Bleeding• Unknown initiation
Placental Pathologic Conditions
Histologic chorioamnionitisPlacental abruption
Placenta previaOther placental abnormalities
Figure 39-2 Pathologic mechanisms in preterm birth. Proposed view of how multiple etiologies and pathogenic pathways converge to trigger uterine contractility, cervical ripening, and preterm premature rupture of membranes (PPROM) in women with preterm birth. (From Buhimschi CS, Schatz F, Krikun G, et al: Novel insights into molecular mechanisms of abruption induced preterm birth, Expert Rev Mol Med 12:e35, 2010.)
Multiplepregnancy
Geneticpredisposition
Intra-amnioticinflammation
Uterine over-distension
Decidual hemorrhage
Stress
Oxidativestress
Fetal allergy
Nutrition
Environmental toxins
Infection
Nulliparity
Utero-placental ischemia
Uterine contractions Cervical ripening
PPROM Fetal damage
39 Pathogenesis of Spontaneous Preterm Birth 601
decidua to reach the myometrium, where they can stimulate smooth muscle contractions.62 The increase in PTGS-2 activity is induced by an increase in NF-κB activity in both amnion63 and myometrim.64 The importance of NF-κB in the induction of PTB is further underscored by the demonstration that an NF-κB inhibitor can reduce lipopolysaccharide (LPS)-induced PTB in a mouse model.65
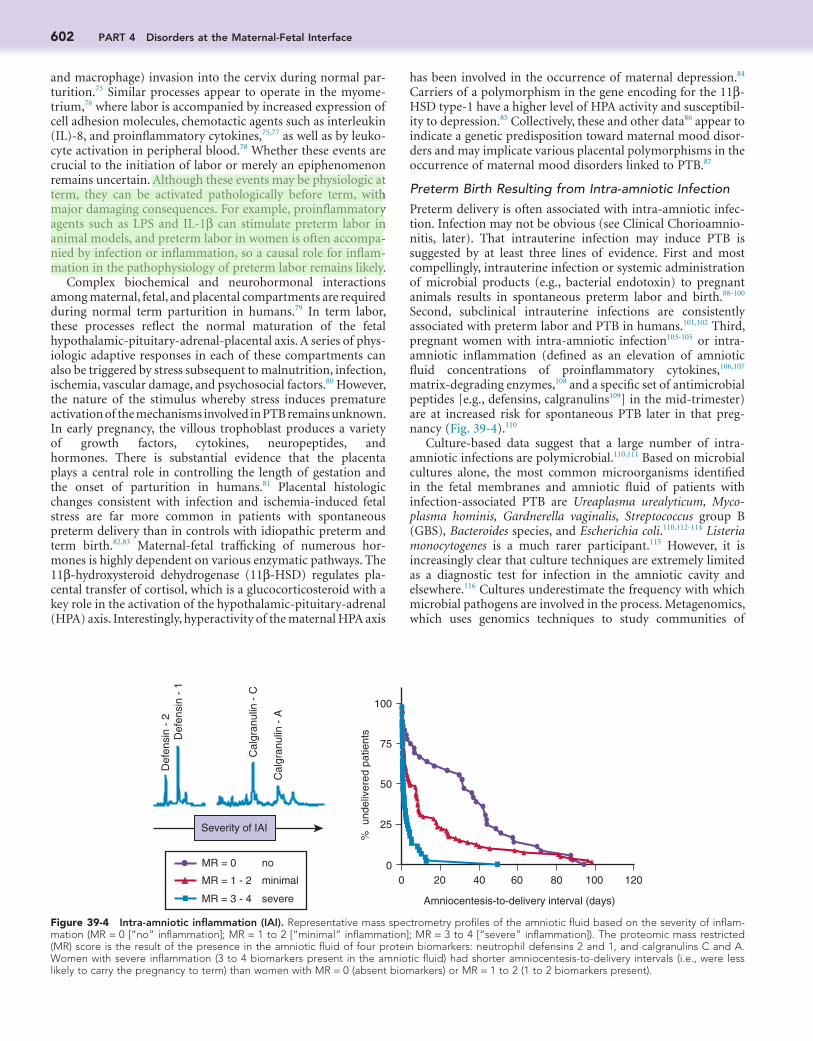
NF-κB and prostaglandins activate common pathways of parturition by the following biochemical mechanisms: (1) Pros-taglandins directly promote uterine contractions by increasing sarcoplasmic and transmembrane calcium fluxes and through increased transcription of oxytocin receptors, connexin-43 (gap junctions), and the prostaglandin E2 (PGE2) receptors EP1 through EP4 (although EP3 appears to be the predominant receptor subtype66) and the PGF2α receptor FP67-70; (2) prosta-glandins induce synthesis of matrix metalloproteinases (MMPs) by fetal membranes and cells in the uterine cervix to promote membrane rupture and cervical ripening71,72; (3) PGE2 and PGF2α increase the ratio of expression of the progesterone receptor (PR) isoforms PR-A and PR-B to induce functional progesterone withdrawal73; and (4) NF-κB activation induces activation of a cassette of inflammatory genes, which may also induce a functional progesterone withdrawal.64 Figure 39-3 describes the molecular mechanisms implicated in the common pathway of parturition.
INFLAMMATION, STRESS, AND TERM AND PRETERM PARTURITION
Inflammation is a highly orchestrated process designed to ensure survival of the host.74 The inflammatory process has a physiologic component intended to ensure maintenance of homeostasis. Increasing evidence suggests that parturition at term is such a process. Liggins first proposed that cervical ripen-ing was an inflammatory event, and this hypothesis is sup-ported by data showing a profound leukocytic (neutrophilic
still incompletely understood. In the peripheral circulation, an increase in unbound corticotropin-releasing hormone and, in the uterus, increased nuclear factor kappa B (NF-κB) activity (associated with functional progesterone withdrawal, prosta-glandin production, and leukocytic influx) are consistently demonstrated in association with parturition.18 There is con-tinuing debate about which (if any) of these events is the master regulator that controls the timing of parturition, and which (if any) is the sine qua non, without which parturition cannot occur. Evidence for each of these processes is briefly reviewed below.
Prostaglandins as Key Activators of the Common Pathway of ParturitionProstaglandins are viewed as crucial mediators for the onset of labor,19-34 because they can induce myometrial contractil-ity,19,23,32,34 promote proteolysis of cervical and fetal membrane extracellular matrices to cause cervical ripening and fetal mem-brane rupture,21,22,26,27,31 and stimulate decidual/membrane acti-vation.35 Evidence of a role for prostaglandins in the initiation of human parturition includes the following: (1) Administra-tion of prostaglandins induces termination of pregnancy30,36-46; (2) indomethacin or aspirin therapy delays spontaneous onset of parturition in animals47-50; (3) concentrations of prostaglan-dins in plasma and amniotic fluid increase during labor51-59; (4) intra-amniotic injection of the prostaglandin precursor arachi-donic acid induces abortion28; (5) expression of myometrial prostaglandin receptors increases in labor60,61; and (6) labor is associated with increased expression of prostaglandin endoper-oxide synthase 2 (PTGS-2) messenger RNA and increased activ-ity of this enzyme in the amnion (a rate-limiting step in the production of prostaglandins). This increase in amniotic PTGS-2 activity is accompanied by decreased expression of the prostaglandin-metabolizing enzyme 15-hydroxyprostaglandin dehydrogenase (PGDH) in the chorion. This would allow pros-taglandins produced in the amnion to traverse the chorion and
Figure 39-3 Prostaglandins as key activators of the common pathway of parturition. Ca2+, calcium; EP1, prostaglandin E1 receptor; FP, pros-taglandin F receptor; MMPs, matrix metalloproteinases; NF-κB, nuclear factor kappa B (a transcription factor); PR, progesterone receptor; PG, prostaglandin; PGE2, prostaglandin E2; pGF2α, prostaglandin F2α; pPROM, preterm premature rupture of membranes, PR-A/PR-B, ratio of proges-terone receptor type A to type B.
Myometrium • Contractions
Fetal membranes • pPROM
PG
Ca2+ • Oxytocin R• Connexin-43• PGE2 EP1• PGF2α FP
+
+
+
+ +
+
MMPs
+ Inflammation PR-A/PR-B ratio PGNF-κB
+ +
Cervix • Dilation • Ripening
602 PART 4 Disorders at the Maternal-Fetal Interface
has been involved in the occurrence of maternal depression.84 Carriers of a polymorphism in the gene encoding for the 11β-HSD type-1 have a higher level of HPA activity and susceptibil-ity to depression.85 Collectively, these and other data86 appear to indicate a genetic predisposition toward maternal mood disor-ders and may implicate various placental polymorphisms in the occurrence of maternal mood disorders linked to PTB.87
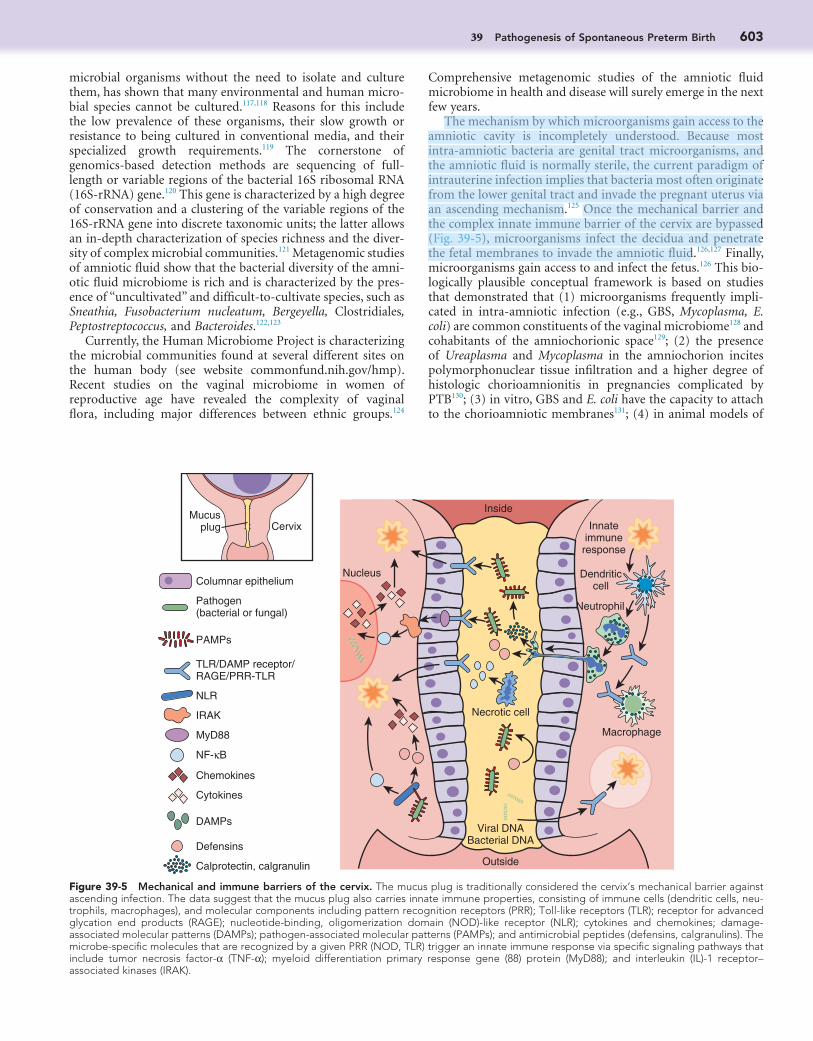
Preterm Birth Resulting from Intra-amniotic InfectionPreterm delivery is often associated with intra-amniotic infec-tion. Infection may not be obvious (see Clinical Chorioamnio-nitis, later). That intrauterine infection may induce PTB is suggested by at least three lines of evidence. First and most compellingly, intrauterine infection or systemic administration of microbial products (e.g., bacterial endotoxin) to pregnant animals results in spontaneous preterm labor and birth.88-100 Second, subclinical intrauterine infections are consistently associated with preterm labor and PTB in humans.101,102 Third, pregnant women with intra-amniotic infection103-105 or intra-amniotic inflammation (defined as an elevation of amniotic fluid concentrations of proinflammatory cytokines,106,107 matrix-degrading enzymes,108 and a specific set of antimicrobial peptides [e.g., defensins, calgranulins109] in the mid-trimester) are at increased risk for spontaneous PTB later in that preg-nancy (Fig. 39-4).110
Culture-based data suggest that a large number of intra-amniotic infections are polymicrobial.110,111 Based on microbial cultures alone, the most common microorganisms identified in the fetal membranes and amniotic fluid of patients with infection-associated PTB are Ureaplasma urealyticum, Myco-plasma hominis, Gardnerella vaginalis, Streptococcus group B (GBS), Bacteroides species, and Escherichia coli.110,112-114 Listeria monocytogenes is a much rarer participant.115 However, it is increasingly clear that culture techniques are extremely limited as a diagnostic test for infection in the amniotic cavity and elsewhere.116 Cultures underestimate the frequency with which microbial pathogens are involved in the process. Metagenomics, which uses genomics techniques to study communities of
and macrophage) invasion into the cervix during normal par-turition.75 Similar processes appear to operate in the myome-trium,76 where labor is accompanied by increased expression of cell adhesion molecules, chemotactic agents such as interleukin (IL)-8, and proinflammatory cytokines,75,77 as well as by leuko-cyte activation in peripheral blood.78 Whether these events are crucial to the initiation of labor or merely an epiphenomenon remains uncertain. Although these events may be physiologic at term, they can be activated pathologically before term, with major damaging consequences. For example, proinflammatory agents such as LPS and IL-1β can stimulate preterm labor in animal models, and preterm labor in women is often accompa-nied by infection or inflammation, so a causal role for inflam-mation in the pathophysiology of preterm labor remains likely.
Complex biochemical and neurohormonal interactions among maternal, fetal, and placental compartments are required during normal term parturition in humans.79 In term labor, these processes reflect the normal maturation of the fetal hypothalamic-pituitary-adrenal-placental axis. A series of phys-iologic adaptive responses in each of these compartments can also be triggered by stress subsequent to malnutrition, infection, ischemia, vascular damage, and psychosocial factors.80 However, the nature of the stimulus whereby stress induces premature activation of the mechanisms involved in PTB remains unknown. In early pregnancy, the villous trophoblast produces a variety of growth factors, cytokines, neuropeptides, and hormones. There is substantial evidence that the placenta plays a central role in controlling the length of gestation and the onset of parturition in humans.81 Placental histologic changes consistent with infection and ischemia-induced fetal stress are far more common in patients with spontaneous preterm delivery than in controls with idiopathic preterm and term birth.82,83 Maternal-fetal trafficking of numerous hor-mones is highly dependent on various enzymatic pathways. The 11β-hydroxysteroid dehydrogenase (11β-HSD) regulates pla-cental transfer of cortisol, which is a glucocorticosteroid with a key role in the activation of the hypothalamic-pituitary-adrenal (HPA) axis. Interestingly, hyperactivity of the maternal HPA axis
Figure 39-4 Intra-amniotic inflammation (IAI). Representative mass spectrometry profiles of the amniotic fluid based on the severity of inflam-mation (MR = 0 [“no” inflammation]; MR = 1 to 2 [“minimal” inflammation]; MR = 3 to 4 [“severe” inflammation]). The proteomic mass restricted (MR) score is the result of the presence in the amniotic fluid of four protein biomarkers: neutrophil defensins 2 and 1, and calgranulins C and A. Women with severe inflammation (3 to 4 biomarkers present in the amniotic fluid) had shorter amniocentesis-to-delivery intervals (i.e., were less likely to carry the pregnancy to term) than women with MR = 0 (absent biomarkers) or MR = 1 to 2 (1 to 2 biomarkers present).
0 20 40 60 80 100 1200
25
50
75
100
Amniocentesis-to-delivery interval (days)
Severity of IAI
MR = 0
MR = 1 - 2
MR = 3 - 4
no
minimal
severe
Def
ensi
n -
2 D
efen
sin
- 1
Cal
gran
ulin
- C
Cal
gran
ulin
- A
% u
ndel
iver
ed p
atie
nts
39 Pathogenesis of Spontaneous Preterm Birth 603
Comprehensive metagenomic studies of the amniotic fluid microbiome in health and disease will surely emerge in the next few years.
The mechanism by which microorganisms gain access to the amniotic cavity is incompletely understood. Because most intra-amniotic bacteria are genital tract microorganisms, and the amniotic fluid is normally sterile, the current paradigm of intrauterine infection implies that bacteria most often originate from the lower genital tract and invade the pregnant uterus via an ascending mechanism.125 Once the mechanical barrier and the complex innate immune barrier of the cervix are bypassed (Fig. 39-5), microorganisms infect the decidua and penetrate the fetal membranes to invade the amniotic fluid.126,127 Finally, microorganisms gain access to and infect the fetus.126 This bio-logically plausible conceptual framework is based on studies that demonstrated that (1) microorganisms frequently impli-cated in intra-amniotic infection (e.g., GBS, Mycoplasma, E. coli) are common constituents of the vaginal microbiome128 and cohabitants of the amniochorionic space129; (2) the presence of Ureaplasma and Mycoplasma in the amniochorion incites polymorphonuclear tissue infiltration and a higher degree of histologic chorioamnionitis in pregnancies complicated by PTB130; (3) in vitro, GBS and E. coli have the capacity to attach to the chorioamniotic membranes131; (4) in animal models of
microbial organisms without the need to isolate and culture them, has shown that many environmental and human micro-bial species cannot be cultured.117,118 Reasons for this include the low prevalence of these organisms, their slow growth or resistance to being cultured in conventional media, and their specialized growth requirements.119 The cornerstone of genomics-based detection methods are sequencing of full-length or variable regions of the bacterial 16S ribosomal RNA (16S-rRNA) gene.120 This gene is characterized by a high degree of conservation and a clustering of the variable regions of the 16S-rRNA gene into discrete taxonomic units; the latter allows an in-depth characterization of species richness and the diver-sity of complex microbial communities.121 Metagenomic studies of amniotic fluid show that the bacterial diversity of the amni-otic fluid microbiome is rich and is characterized by the pres-ence of “uncultivated” and difficult-to-cultivate species, such as Sneathia, Fusobacterium nucleatum, Bergeyella, Clostridiales, Peptostreptococcus, and Bacteroides.122,123
Currently, the Human Microbiome Project is characterizing the microbial communities found at several different sites on the human body (see website commonfund.nih.gov/hmp). Recent studies on the vaginal microbiome in women of reproductive age have revealed the complexity of vaginal flora, including major differences between ethnic groups.124
Figure 39-5 Mechanical and immune barriers of the cervix. The mucus plug is traditionally considered the cervix’s mechanical barrier against ascending infection. The data suggest that the mucus plug also carries innate immune properties, consisting of immune cells (dendritic cells, neu-trophils, macrophages), and molecular components including pattern recognition receptors (PRR); Toll-like receptors (TLR); receptor for advanced glycation end products (RAGE); nucleotide-binding, oligomerization domain (NOD)-like receptor (NLR); cytokines and chemokines; damage-associated molecular patterns (DAMPs); pathogen-associated molecular patterns (PAMPs); and antimicrobial peptides (defensins, calgranulins). The microbe-specific molecules that are recognized by a given PRR (NOD, TLR) trigger an innate immune response via specific signaling pathways that include tumor necrosis factor-α (TNF-α); myeloid differentiation primary response gene (88) protein (MyD88); and interleukin (IL)-1 receptor–associated kinases (IRAK).
Columnar epitheliumNucleus
CervixMucus
plug
Inside
Innateimmune
response
Dendriticcell
NeutrophilPathogen(bacterial or fungal)
PAMPs
TLR/DAMP receptor/RAGE/PRR-TLR
NLR
IRAK
MyD88
Necrotic cell
Macrophage
Viral DNABacterial DNA
Outside
NF-κB
Chemokines
Cytokines
Defensins
Calprotectin, calgranulin
DAMPs

604 PART 4 Disorders at the Maternal-Fetal Interface
that the balance among proinflammatory and anti-inflammatory cytokine responses can dictate the intensity and possible resolu-tion of an infectious process.
The biologic activity of the TLRs depends not only on the presence of bacterial pathogen–associated molecular patterns but on a palette of intracellular signaling adaptors (e.g., MyD88) and co-receptor molecules (e.g., CD14) that associate with TLRs in complex supramolecular arrangements.146 Equally important is that TLR signaling can be elicited by endogenous damage-associated molecular patterns (DAMPs).147 Like cyto-kines, DAMPs (i.e., high-mobility group box-1 [HMGB1, or amphoterin], S100β proteins) are endogenous proinflamma-tory and pro-oxidative stress molecules. Acting through TLR2, TLR4, and the receptor for advanced glycation end products (RAGE), DAMPs recruit inflammatory cells, which in turn amplify innate immune responses and enhance levels of cyto-kine activation.147 It was reported that the RAGE-DAMP system is present in women with PTB and intra-amniotic infection.148 Activation of the RAGE-DAMP system correlates with the degree of inflammation and oxidative stress damage in amnion epithelial, decidual, and extravillous trophoblast cells (Fig. 39-6).149 PAMPs and DAMPs may continue to keep active the processes that lead to fetal cellular damage.
Last, the roles of soluble receptor modulators (soluble TLR2, soluble TNF receptor-1, soluble IL-6 receptor, soluble glycopro-tein (gp)130, and soluble RAGE) in fine-tuning human TLR-mediated signaling have just begun to be elucidated.150-153 Downstream of the TLR receptor, other molecules such as pro-kineticin amplify the inflammatory response,154 so that lentivi-ral knockout of the prokineticin receptor inhibits the ability of the myometrium to produce proinflammatory cytokines in response to LPS.155
Role of Proinflammatory Agents in Preterm BirthInflammation and its mediators (e.g., chemokines such as IL-8; proinflammatory cytokines such as IL-1β and TNF-α; and others, such as platelet activating factor and prostaglandins) are central to infection-induced PTB. IL-1β was the first cytokine implicated in the onset of infection-associated PTB.156 Evidence of the role of IL-1β in the pathogenesis of PTB includes the following: (1) It is synthesized by human decidua in response to bacterial products157; (2) it stimulates prostaglandin produc-tion by human amnion and decidua158; (3) IL-1α and IL-1β concentrations and IL-1-like bioactivity are increased in the amniotic fluid of women with preterm labor and infection159; (4) intravenous IL-1β stimulates uterine contractions160; and (5) administration of IL-1 to pregnant animals induces preterm labor and birth,161 and this effect can be blocked by the admin-istration of its natural antagonist, the IL-1 receptor antagonist (IL-1ra).162
Evidence supporting the role of TNF-α in the mechanisms of infection-associated PTB include the following: (1) TNF-α stimulates prostaglandin production by amnion, decidua, and myometrium95; (2) human decidua can produce TNF-α in response to bacterial products163,164; (3) amniotic fluid TNF-α bioactivity and immunoreactive concentrations are elevated in women with preterm labor and intra-amniotic infection165; (4) in women with preterm PROM and intra-amniotic infection, TNF-α concentrations are higher in the presence of labor165; (5) TNF-α can stimulate the production of MMPs,166,167 which have been implicated in membrane rupture168-170; (6) application of TNF-α to the cervix induces changes that resemble cervical
infection-induced PTB, transcervical and choriodecidual inoc-ulation of GBS is followed by transmigration of bacteria from the choriodecidual space to the amniotic fluid cavity, a graded amniotic-fluid leukocyte infiltration response, and levels of proinflammatory cytokines (tumor necrosis factor-α [TNF-α], IL-6, IL-1β), prostaglandins (PGE2, PGF2α), and uterine activity.132,133
A secondary route of intra-amniotic infection is probably hematogenous transplacental seeding of the fetus, with the infectious organisms, in particular Haemophilus influenzae or F. nucleatum, originating from other parts of the body including the mouth.134,135 Iatrogenic infections during invasive proce-dures such as chorionic villous sampling, amniocentesis, and cordocentesis are also possible.136 Retrograde microbial seeding of the amniotic fluid through the fallopian tubes or coloniza-tion of the uterine endometrium before implantation has also been proposed.126 Compelling evidence in support of these pathways remains to be provided.
Emerging evidence suggests that microorganisms are “sensed” by the innate components of the immune system,137 leading to a cascade of events that culminate in PTB. These sensing components include soluble pattern-recognition recep-tors (PRRs), lectin, and C-reactive protein. The transmembrane PRRs include scavenger receptors, C-type lectins, and Toll-like receptors (TLRs). Intracellular PRRs include NOD1 and NOD2, retinoic acid–induced gene type 1, and melanoma differentiation-associated protein 5, which mediate recognition of intracellular pathogens (e.g., viruses).138 The best-studied PRRs are the TLRs.137 Because of their strategic positioning at the maternal-fetal interface (the decidua),139 fetal membranes, and myome-trium,140 TLR4 and TLR2 are considered major mediators by which the maternal and fetal reproductive tissues can respond to infection. TLR4 is recognized as the membrane-bound recep-tor that triggers LPS signaling of gram-negative microbes.141 A strain of mice bearing a spontaneous disabling mutation for TLR4 is less likely than wild-type mice to have preterm delivery after intrauterine inoculation of heat-killed bacteria or admin-istration of LPS.98,142 TLR2 has been shown to be involved in recognition of lipoproteins, peptidoglycan, and glycolipids of gram-positive bacteria and Mycoplasmataceae.143 How TLRs distinguish between commensal and pathogenic microorgan-isms in vaginal or other sites remains unknown.
Although the full spectrum of TLR-mediated responses remains to be elucidated, it is known that, once engaged by pathogen-associated molecular patterns (PAMPs), these bacte-rial sensors trigger a downstream molecular chain of events that lead to synthesis and release of proinflammatory cytokines such as TNF-α; interferon-γ; cytokines IL-12, IL-6, IL-1β; and many others via an NF-κB–mediated mechanism.144 Key chemokines secreted after TLR activation include IL-8; monocyte chemoat-tractant proteins 1, 2, 3, and 4; macrophage inflammatory pro-teins 1α and 1β; and RANTES (regulated on activation, normal T cell expressed and secreted). Traditionally, it is believed that activation of TLRs induces a T helper cell 1 (TH1) cytokine-type response (i.e., IL-2, interferon-γ, lymphotoxins). However, using genetically engineered animal models and a variety of in vitro cell culture systems, Pulendran and colleagues showed that TLR4 engagement can also trigger a TH2 cytokine reaction con-sistent with the release of IL-4, IL-5, IL-6, and the anti-inflammatory cytokine IL-10, depending on bacterial type.145 The significance of this observation during human pregnancy remains to be clarified, but these results underline the concept

39 Pathogenesis of Spontaneous Preterm Birth 605
Thus, neutrophil secretion of cytokines and chemokines prob-ably follows their recognition of a large repertoire of bacterial PAMPs and cellular DAMPs. Taken together, these observations highlight the maternal and fetal involvement in the process of intra-amniotic inflammation and the role of mother and fetus in amplification of the inflammatory status of the amniotic fluid and tissue damage in a forward loop fashion.200
Increasing evidence points to a role for complement in inflammation-induced PTB. Increased cervical deposition of the split complement product C3 was noted in mouse models of preterm labor induced both by LPS and by progesterone withdrawal.201 Although work in this area is in its infancy, there is evidence that complement activation is restricted to preterm labor and is absent from physiologic parturition at term.202,203 Whether these inflammatory agents truly operate indepen-dently from intrauterine infection (i.e., sterile inflammation) or whether intrauterine infection mediates all of these effects is unclear, and reexamination using modern techniques to iden-tify intrauterine infection is required (see Preterm Birth Result-ing from Intra-amniotic Infection, earlier).
In addition to the proinflammatory events just described, a wide variety of anti-inflammatory mediators are now known to operate in the pregnant uterus. The most widely known of these is the anti-inflammatory cytokine IL-10, which is thought to be important for the maintenance of pregnancy.204-206 Its concen-trations are increased in intra-amniotic inflammation,207 sug-gesting that IL-10 may play a role in damping the inflammatory response208-213 and may have therapeutic value.214-219 IL-10 knockout mice are more sensitive to LPS-induced preterm labor than wild-type mice—a defect that is ameliorated by external IL-10 administration.220 In wild-type animals, exogenous IL-10 also attenuates the preterm labor phenotype. For example, in a nonhuman primate model of intrauterine infection, pregnant
ripening171; (7) TNF-α can induce preterm parturition when administered systemically to pregnant animals172,173; and (8) TNF-α and IL-1β enhance IL-8 expression by decidual cells, and this chemokine is strongly expressed by term decidual cells in the presence of chorioamnionitis.174
Other cytokines and chemokines (IL-6,175-180 IL-10,160,181,182 IL-16,183 IL-18,184 colony-stimulating factors,185-187 macrophage migration inhibitory factor,188 IL-8,187,189-193 monocyte chemo-tactic protein-1,194 epithelial cell–derived neutrophil-activating peptide-78,195 and RANTES196) have also been implicated in infection-induced PTB. The redundancy of the cytokine network implicated in parturition is such that blockade of a single cytokine is unlikely to be sufficient to prevent PTB in the setting of infection. For example, preterm labor after exposure to infection can occur in knockout mice for the IL-1 type I receptor, suggesting that IL-1 is sufficient, but not necessary, for the onset of parturition in the context of intra-amniotic infec-tion or inflammation.197 However, blockade of multiple signal-ing pathways (e.g., IL-1β and TNF-α) in a double-knockout mouse model decreased the rate of PTBs after the administra-tion of microorganisms.173
In the setting of intra-amniotic infection, a large array of cytokines (e.g., IL-6, IL-8, IL-1β, granulocyte-macrophage colony-stimulating factor) are found in the amniotic fluid. The sources of amniotic fluid cytokines probably include decidua, fetal membranes, and the fetus. However, independent of the source, amniotic fluid IL-6 and many other cytokines induce recruitment of fetal neutrophils.198 Cytokines also induce degranulation of neutrophilic granulocytes with release of MMP-1 (collagenase). With the exception of TLR3, human leu-kocytes express the mRNAs of TLR1 through TLR10.199 Expres-sion of TLR2 is higher in circulating leukocytes obtained from women in labor than from pregnant women not in labor.78
Figure 39-6 Working model for the potential role of the RAGE-DAMP system leading to inflammation, oxidative stress, and fetal damage. DAMPs, damage-associated molecular patterns; HMGB1, high-mobility group box-1; PAMPs, pathogen-associated molecular patterns; RAGE, receptor for advanced glycation end products; ROS, reac-tive oxygen species; S100A8, calgranulin A; S100A12, cal-granulin C.
Systemic orascending infection
(PAMPs)
InflammationCytokines, chemokines
Oxidative stressFree radicals, ROS
FETAL CELLULAR DAMAGE
RAGE activation
Trauma, abruption or intra-amniotic bleeding
(DAMPs)
Release of damage-associatedmolecular patterns (DAMPs)
HMGB1, S100 proteins (S100A12, S100A8) etc.

606 PART 4 Disorders at the Maternal-Fetal Interface
intra-amniotic inflammatory response triggered by such bacte-ria, separately or as a group, remains to be determined.
STRETCH AND PARTURITION
Myometrial Stretch and Term and Preterm BirthDuring human pregnancy, significant physical and biochemical adaptive transformations of the myometrium are required to aid the development and growth of the fetus. These transforma-tions facilitate conversion of the uterus into a thin-walled muscular organ and maintain myometrial quiescence.230 Math-ematical models derived from studies aimed to understand myocardial contractility indicate that wall stress (applied force per unit of cross-sectional area) is directly proportional to intracavitary pressure and radius of the curve, but inversely proportional to the thickness of the muscular wall.231 The rel-evance of this model for the pregnant uterus is that the thick-ness of the myometrium and intra-amniotic pressure both influence uterine wall stress.232
Intra-amniotic pressure remains low through human gesta-tion.233 A low pressure is achieved through various electrophysi-ologic (e.g., by decreased number of gap junctions)234 and biomolecular (e.g., by hormonal signals that stimulate macro-phage migration, by release of cytokines, by activation of inflammatory transcription factors) processes that maintain a state of uterine quiescence in the setting of progressive myome-trial stretch.235 The mechanisms that signal conversion of the myometrium from a quiescent to a highly contractile state are unknown. However, it is reasonable to propose that several of these mechanisms are mechanically activated.235
A large body of clinical data implicates excessive myometrial stretch in the genesis of PTB. For example, a high amniotic fluid index (AFI ≥ 25 cm) is associated with a significantly increased incidence of PTB.236 Polyhydramnios and multiple gestations are the most relevant examples.236,237 There has been increased interest in identifying the molecular mechanisms responsible for the onset of uterine contractility.238 Progesterone receptor transcriptional activity has been proposed as critical for the preservation of myometrial relaxation.238 This inhibitory effect seems to be mediated by repressing the expression of genes that encode contraction-associated proteins.238,239 Two such genes are connexin-43 and oxytocin receptor. The connexin-43 gene encodes a protein with critical roles in synchronizing myome-trial contractile activity,234 and oxytocin receptor gene controls responsiveness of myometrial cells to oxytocin.240 Mechanical stretch, however, upregulates expression of connexin-43, an effect that is inhibited by progesterone.241 In vitro data demon-strated that the upregulation of the expression of oxytocin-receptor mRNA that occurs as a result of myometrial stretching is controlled via DNA binding to various transcription factors, including activator protein-1 (AP-1) and CCAAT/enhancer binding protein (C/EBP)-β.242 Interestingly, the transcription factor NF-κB did not increase the promoter activity of the oxytocin receptor gene.242 That mechanical stimulation of the uterine wall promotes expression of oxytocin receptor mRNA, and that this effect is favored by progesterone withdrawal, was confirmed in vivo.243
Myometrial elongation stimulates the expression of a variety of cytokines and chemokines (e.g., CCL2, CXCL8, CXCL1, CCL2) with a characteristic proinflammatory profile for preterm labor tissues.244,245 In various experimental models, the primary mediator of myometrial stretch-induced inflammation
rhesus monkeys were allocated to one of three interventional groups: (1) intra-amniotic IL-1β infusion with maternal dexa-methasone intravenously; (2) intra-amniotic IL-1β and IL-10; or (3) intra-amniotic IL-1β administered alone. Dexametha-sone and IL-10 treatment significantly reduced IL-1β-induced uterine contractility. The amniotic fluid concentrations of TNF-α and leukocyte counts were also decreased by IL-10 treat-ment.160 In addition to these beneficial effects on inhibition of contractility and inflammation, administration of IL-10 in animal models of infection has also been associated with improved pregnancy outcome.214,221
Another major group of anti-inflammatory molecules, the lipoxins,222 are also expressed in the reproductive tract.223 Lipox-ins are part of a group of pro-resolution molecules that appear to actively terminate the inflammatory process, promoting neu-trophil engulfment and inhibiting proinflammatory cytokine expression. Although their role in infection-induced PTB has not been elucidated, they circulate in increasing concentrations as pregnancy advances, their receptor is present in the myome-trium of pregnant women, and they attenuate the myometri-um’s proinflammatory cytokine response to LPS.223 Thus, they also show promise as therapeutic agents for infection-induced preterm labor.
In summary, there is increasing interest in the use of anti-inflammatory strategies—either for upregulating endogenously produced molecules or for external application of anti-inflammatory agents to treat preterm labor.224 These issues will be discussed further in the chapter on treatment of preterm labor.
Bacterial Species and the Intensity of Intra-amniotic Inflammatory ResponsesOnce present in the amniotic fluid, microorganisms can stimu-late the production of proinflammatory cytokines through acti-vation of fetal membrane TLR receptors.225 Several microorganisms (e.g., Ureaplasma, Mycoplasma) are tradition-ally considered to have low virulence.113,226 Studies describing the presence of Ureaplasma parvum and M. hominis in the amniotic fluid of second-trimester asymptomatic women are in support of this concept.103 Menon and coworkers demonstrated in vitro that in comparison with gram-positive and other gram-negative bacteria, Ureaplasma has a lower proinflammatory effect on fetal membranes.225 However, isolation of Ureaplasma and Mycoplasma in the amniotic fluid has been consistently associated with a wide range of adverse outcomes, such as early abortion, stillbirth, prematurity, and neonatal morbidity and mortality.227 Although an intense intra-amniotic inflammatory response is often encountered at the time of clinical onset of PTB, these studies prove association, not causation.228 Evidence that these so called silent microorganisms are capable of trig-gering an inflammatory response in vivo that can induce PTB was recently provided by Novy and coworkers.229 They inocu-lated U. urealyticum and M. hominis into the amniotic fluid of rhesus monkeys, which resulted in an increase in a myometrial contractile activity that was preceded by an intense proinflam-matory cytokine response and prostaglandin synthesis.229
Invasion of the amniotic fluid with gram-positive anaerobes, E. coli, and GBS results in intra-amniotic inflammation and fetal sepsis.114 However, intra-amniotic inflammation can also occur in the absence of positive amniotic-fluid culture results.110 As previously mentioned, “uncultivated” or difficult-to- cultivate bacteria may play an important role. The extent of

39 Pathogenesis of Spontaneous Preterm Birth 607
and chorion, but not in decidua.259 Studies using an in vitro cell culture model for fetal membrane distention demon-strated upregulation of proinflammatory genes, including those for IL-8 and pre-B-cell colony-enhancing factor (visfa-tin).260 Stretching of the fetal membrane in vitro results in overexpression of various genes, specifically those for IL-8, interleukin-enhancer binding factor 2, huntingtin-interacting protein 2, and interferon-stimulated gene encoding a 54-kDa transcript.261
These molecular mechanisms, schematically represented in Figure 39-7, highlight that myometrial and excessive mechani-cal stretch of the fetal membranes could lead to PTB through integration of multiple cellular and extracellular signaling path-ways. Each of these mechanisms can be linked to various phe-notypic components of the PTB syndrome: polyhydramnios, multiple fetuses, and twin-twin transfusion syndrome.
ACTIVATION OF THE MATERNAL-FETAL HPA AXIS IN TERM AND PRETERM BIRTH
Considerable and well-deserved attention has been paid to glu-cocorticoid physiology during human pregnancy. Substantial research data indicate the existence of a positive feedback loop involving glucocorticoids, proinflammatory cytokines, prosta-glandins, surfactant protein-A, and 11β-HSD type-1 in human fetal membranes in women who are going to have a preterm delivery.262 There is evidence to support the hypothesis that this mechanism is active in human preterm parturition in the setting of infection-induced histologic chorioamnionitis.263 Additionally, in the model of preterm parturition, stress may be involved in the production of abundant biologically active glu-cocorticoids and prostaglandins, which might promote acceler-ated fetal maturation and initiation of parturition.262
A schematic representation of the physiology of the fetal hypothalamic-pituitary-adrenal-placental axis in pregnancy is presented in Figure 39-8. Corticotropin-releasing hormone (CRH; a 41-amino-acid peptide) appears to be the mediator of stress-associated preterm deliveries.264 The glucocorticosteroid
appears to be NF-κB.244 These experiments provide support for the concept that uterine distention carries an inflammatory component. Other mechanisms that can lead to activation of myometrial contractility subsequent to excessive mechanical stretch are (1) increases in the transcription factor AP-1; (2) activation of mitogen-activated protein kinase (MAPK)-depen-dent and cyclooxygenase (COX)-2-mediated prostaglandin synthesis246,247; (3) downregulation in the expression of stretch-dependent K+ (SDK) channels248; (4) changes in the expression of transient receptor potential canonical (TRPC) proteins with a role in store-operated calcium entry in human myometrial smooth muscle cells249; and (5) upregulation in the expression pattern of gastrin-releasing peptide, a molecule with agonistic contractile properties.250
Fetal Membrane Stretch and Term and Preterm BirthFetal membranes carry important protective and biochemical functions. In physiologic pregnancy, the fetal membranes undergo progressive mechanical stretch, allowing their accom-modation to the growing uterus.251 The human fetal mem-branes are complex tissues composed of highly specialized epithelial and mesenchymal cells embedded in an extracellular matrix composed primarily of collagen and proteoglycans.252 Interstitial types I and III collagens predominate and represent the main structural components that maintain the mechanical integrity of the membranes.252 Collagen can be stretched but it is not elastic, whereas other structural components (e.g., elastin) of the extracellular matrix are both stretchable and elastic.
By comparing the surface area of the fetal membranes with that of the uterine cavity, Parry-Jones and Priya demonstrated that after 28 weeks of gestation, the intact fetal membranes are under tension and in a state of active stretch.253 A significant decrease in the elasticity of membranes that ruptured before labor was observed independent of gestational age. In a study conducted by Millar and coworkers, the investigators confirmed marked differences in elasticity between individual membranes and a diminished ability of the membranes that rupture before term to stretch.254
Traditionally, rupture of the fetal membranes was viewed as a mechanical event.255 However, although this might hold true for specific clinical cases, maintenance of the tensile strength of fetal membranes appears to involve a highly coordinated balance between synthesis and degradation of various components of the extracellular matrix.256 It has been proposed that changes in the membranes, including decreased collagen content, altered collagen structure, and increased collagenolytic and proinflam-matory activity, are associated with preterm PROM and PTB.253 In vitro, stretching of the fetal membranes induces collagenase activity.257 Thus, it is possible that, in vivo, a loaded state of the fetal membranes might facilitate their susceptibility to enzy-matic degradation. Mechanical stimulation of the amnion cells results in increased expression of COX-2 and PGE2 production, suggesting a feed-forward loop under which stimulation of uterine contractions magnifies the degree of stretching to the point of rupture and preterm delivery.258 Because this phenom-enon is mediated through members of the AP-1 family of tran-scription factors (i.e., Fos and Jun) and NF-κB, involvement of various inflammatory pathways has also been also proposed. Synthesis and expression of the proinflammatory cytokines IL-8 and IL-1β are increased after exposure of the whole mem-branes and amnion cells to mechanical stretch.257-259 Interest-ingly, expression of IL-8 was upregulated in both amnion
Figure 39-7 Proposed mechanisms by which stretch can induce preterm labor. ECM, extracellular matrix; IL-8, interleukin-8; MAPK, mitogen-activated protein kinase; MMPs, matrix metalloproteinases; PG, prostaglandins; PPROM, preterm premature rupture of membranes; PTL, preterm labor.
Rapid increases in myometrial stretch due to polyhydramnios,multifetal gestations or uterine anatomic abnormalities
Integrin-MAPKsignaling
PG, oxytocinreceptors, IL-8
MMPs PGIL-8 IL-8 PG
Cervix
PTL !/" PPROM
Myometrium Amniochorion
Contractions ECM degradation

608 PART 4 Disorders at the Maternal-Fetal Interface
expression of the mRNAs of CRH and its receptor CRH-R1 was higher in pregnancies complicated by preterm PROM and chorioamnionitis.274 In their experimental setup, endotoxin increased trophoblast CRH, urocortin-2, and CRH-R1 mRNA expression in a dosage-dependent manner. Moreover, prosta-glandins increase cervical expression of IL-8, which recruits and activates neutrophils, releasing additional MMPs and collage-nases, which can promote cervical extracellular matrix disorga-nization and weakening of the fetal membranes.71 The secretion of IL-8 and MMP-1 was significantly higher, and MMP-3 secre-tion was lower, in preterm cervical fibroblasts. In summary, cervical ripening seems to have an inflammatory component, with CRH possibly contributing to its initiation. However, preterm and term cervical fibroblasts might have different phe-notypes based on different secretion patterns of IL-8, MMP-1, and MMP-3.71
Progesterone is a hormone with key roles in human parturi-tion. Data published by several groups suggest that CRH directly modulates the endocrine function of placental trophoblasts, including production of progesterone275 and estrogens.276 Keeping in mind the common pathway to parturition, it is plausible that CRH-induced PGE2, and PGF2α increase the expression of the PR-A isoform and decrease that of the PR-B isoform in myometrium, cervix, and decidua.73,277,278 Because PR-A antagonizes many of the classic PR-mediated genomic effects of PR-B, prostaglandins appear to induce a functional progesterone withdrawal. Decidual cells, and not amnion and chorion cells, seem to be the direct target of progesterone during human pregnancy.279 This assertion is supported by Merlino and colleagues, who reported that in contrast to the intense nuclear PR mRNA and protein expression observed in decidual cells, PR expression is barely detectable in amnion and chorion.279
Experimental data also suggest that a functional increase in myometrial CRH signaling may lead to activation of myome-trial contractility and labor. A direct CRH signaling effect is possible based on the observation that both CRH-R1 and CRH-R2 are expressed in pregnant upper- and lower-segment human myometrium.280 Placing these observations in the context of labor is difficult because the protein level of CRH-R1 in the upper contractile segment was significantly downregu-lated in pregnancy, with a further decrease at the onset of labor. No significant changes in CRH-R2 expression were observed in either upper- or lower-segment myometrium. There is evidence for a myometrial relaxing effect of CRH, favoring uterine qui-escence.281 Therefore, the role of CRH in controlling activation of myometrial contractility, both term and preterm, continues to be an enigma.
Fetal Control of the Onset of ParturitionUsing matched maternal and fetal pairs samples, Lockwood and coworkers evaluated activation of the maternal-fetal HPA axis in patients undergoing cordocentesis during the second half of gestation.282 The authors noted that in physiologic pregnancy, placenta-derived maternal serum CRH values correlated better with fetal (r = 0.40) but only modestly with maternal (r = 0.28) cortisol levels. Based on these findings, it is possible that placental-derived CRH stimulates the release of fetal pituitary adrenocorticotropin to enhance fetal adrenal cortisol produc-tion, which further stimulates placental CRH release. At term, cortisol released into the amniotic fluid can directly stimulate fetal membrane prostaglandin production by increasing
hormone cortisol displays an inhibitory effect on the hypotha-lamic CRH production.264 On the other hand, cortisol stimu-lates the placental production of CRH.264 A positive, feed-forward system of CRH is a unique biologic feature of the placenta, causing progressive increases in placental CRH production as pregnancy advances to term. The effect of CRH seems to be broad, based on its expression by various placental, chorionic, amniotic, and decidual cells.265 In uncomplicated pregnancies, maternal plasma free CRH levels rise exponentially during the second half of pregnancy and peak during labor.81 The expo-nential rise in maternal plasma CRH concentration is associated with a concomitant fall in levels of CRH binding protein, leading to a rapid increase in maternal circulating levels of bioavailable CRH. This suggests that CRH may act directly as a trigger for parturition in humans. The CRH concentration across the gestation curve in women with subsequent PTB runs parallel but to the left of the CRH curve for term pregnancy.266 Despite these findings, it is unclear whether precocious eleva-tion of maternal plasma CRH levels is an epiphenomenon or a trigger for preterm delivery mechanisms.267 Because CRH maternal plasma concentrations are elevated in both term and preterm parturition, it appears that CRH is part of a common pathway of labor.
Several effector mechanisms have been proposed as being involved in activation of the common pathway of labor by CRH. First, the output of PGF2α and PGE2 production and synthesis is stimulated by CRH in amniotic, chorionic, decidual, and placental cells.268,269 Cortisol synthesized in response to CRH can increase amnion COX-2 expression while inhibiting chori-onic PGDH expression.270-272 The net result would be an increase in the bioavailability of prostaglandins. Prenatal stress increases not only prostaglandin levels but also that of maternal circulat-ing inflammatory markers (e.g., IL-6, IL-8, TNF-α) that are associated with prematurity.273 The link between stress hor-mones and various inflammatory signaling pathways in preg-nancies complicated by infection and histologic chorioamnionitis has been demonstrated.274 Torricelli and associates showed that
Figure 39-8 The fetal hypothalamic-pituitary-adrenal-placental axis in pregnancy. ACTH, adrenocorticotropic hormone; CRH, corticotropin-releasing hormone.
!
!
Stress
Hypothalamus
Pituitary
CRH
ACTH
Cortisol
Placenta,decidua, andamniochorion
Adrenal gland
(")
(")

39 Pathogenesis of Spontaneous Preterm Birth 609
the relative proportions of the nuclear PR-A and PR-B, through which progesterone is thought to exert the bulk of its actions. PR-A is thought to act as an endogenous repressor of PR-B, with an increase in the ratio of myometrial PR-A to PR-B, which decreases the response of the uterus to the relaxant effect of progesterone. The role of other progesterone receptors such as PR-C and PR cofactors and repressors is still debated,303 and work continues into downstream mediators such as the miR-200 family and its targets, zinc finger E-box binding homeobox proteins ZEB1 and ZEB2.239 Regardless, progesterone and NF-κB (see later) seem to exert a mutually repressive effect on each other’s actions, generating a feed-forward loop when one starts to predominate.63,304
GENE-ENVIRONMENT INTERACTION
A gene-environment interaction is said to be present when the risk for a disease (occurrence or severity) among individuals exposed to both the genotype and environmental triggers is either more severe or less severe than that predicted from the presence of either the genotype or the environmental exposure alone.305,306 There is evidence for a gene-environment interac-tion in infection-related PTB.307 In a case-control study, patients who had a spontaneous preterm delivery (>37 weeks) were compared with controls delivered after 37 weeks. The environ-mental exposure was bacterial vaginosis diagnosed by symp-tomatic vaginal discharge, a positive whiff test, and clue cells on a wet preparation. The genotype of interest was TNF-α allele 2.308 The authors found that patients with both bacterial vagi-nosis and the TNF-α allele 2 had an odds ratio (OR) of 6.1 (95% confidence interval [CI], 1.9 to 21) for spontaneous PTB, and that this OR was higher than for patients with either bacte-rial vaginosis or carriage of the TNF-α allele alone, suggesting that a gene-environment interaction predisposes to PTB.307,309
A schematic representation of the principal molecular and biochemical mechanisms responsible for the main pathways of preterm parturition is presented in Figure 39-9.
amniotic COX-2 expression and inhibiting the chorionic prostaglandin-metabolizing enzyme PGDH.271,283 This suggests that a local amniotic-fluid positive feedback loop exists to tie fetal HPA axis maturation to parturition.
Compelling data indicate that the fetus actively participates in controlling the timing of labor via production of adrenal hormonal precursors.284 This argument is supported by the evi-dence that at term, before the onset of labor, the weight and volume of the fetal adrenal gland equals that of the adult.285 An important role of the fetus in stress-induced PTB has been proposed.286-288 By using volume analysis—virtual organ computer-aided analysis (VOCAL)—of three-dimensional (3D) ultrasonographic images, Turan and colleagues demon-strated that a birth-weight-corrected fetal adrenal gland volume of greater than 422 mm3/kg was a significant predictor of PTB.289 Development of the fetal adrenal zone of the fetal adrenal gland after 28 to 30 weeks’ gestation creates the context of a stress-induced activation of the placental-fetal HPA axis and enhancement of placental estrogen production. This is because a unique adaptation evolved in primates: placental expression of CRH.290 Placental CRH stimulates the fetal adrenal zone, an adrenal structure unique to primates, to produce dehy-droepiandrosterone sulfate (DHEAS), which is converted to estrogen by the placenta. In addition, CRH can directly augment fetal adrenal DHEAS synthesis.291 Remarkably, fetuses exposed to intra-amniotic inflammation also have higher adrenal gland volumes and lower cortisol-to-DHEAS ratios.292 Placental sul-fatases facilitate conversion of fetal adrenal gland DHEAS to estradiol, estrone, and estriol. These estrogens increase myome-trial expression of contraction-associated proteins such as oxy-tocin receptor and connexin-43.293,294 Because reductions in PR-B expression lead to increased expression of the estrogen receptor-β (ER-β), placental estrogen production would act synergistically to prostaglandin-induced increases in myome-trial ER-β expression.295
PROGESTERONE WITHDRAWAL AND PARTURITION
The role of progesterone in the timing of the onset of human parturition has long puzzled reproductive biologists. Progesterone—literally, “in favor of carrying [a baby]”—is secreted initially by the corpus luteum and then by the placenta in large amounts during pregnancy. It maintains uterine quies-cence by inhibiting myometrial contractions.296 In many species, including most mammals, the onset of parturition is triggered (in part) by an increase in circulating estrogen and a decrease in circulating progesterone levels.297 Although there is no acute change in progesterone levels at the time of parturition,298 the importance of progesterone in human pregnancy is shown by the efficacy of the antiprogesterone mifepristone as an abortion-inducing agent in early pregnancy299 and its actions in inducing labor in later pregnancy300 (although its side-effect profile is such that it should be used only in the scenario of intrauterine fetal death). Additionally, progesterone administration reduces the rate of spontaneous PTB by around 50% in women at high risk because of a history of a previous PTB and in those who are at risk because of a short cervix.301
Although progesterone levels do not change as human labor starts, increasing evidence suggests that labor is associated with a functional progesterone withdrawal (see Mesiano and cowork-ers for a review302). Briefly, although circulating progesterone levels do not change, parturition is associated with changes in
Figure 39-9 Principal biochemical mechanisms responsible for the main pathways of preterm parturition. COX-2, cyclooxygenase-2; CRH, corticotropin-releasing hormone; IL, interleukin; MMPs, matrix metalloproteinases; PGDH, prostaglandin dehydrogenase; pPROM, preterm premature rupture of membranes; PR-B, progesterone recep-tor type B; PTL, preterm labor; TNF-α, tumor necrosis factor-α.
Inflammation
Thrombin
PTL or pPROM
CRHEstrogen
Stretch
Integrins
Abruption Stress
COX2PGDHPR-B
MMPsIL-6 and 8
IL-1βTNF-α

610 PART 4 Disorders at the Maternal-Fetal Interface
phenotype coincided with a higher myometrial expres-sion of antiapoptotic proteins (BCL2 and BCL2L1 [for-merly BCL-xL]).
2. Synthetic, in which the myometrial cells underwent hypertrophy, as demonstrated by a higher protein-to-DNA ratio in the second half of pregnancy. This stage coincided with a higher secretion of extracellular matrix proteins from the myocytes, in particular collagen I and collagen III, as well as a high concentration of caldesmon (a marker of synthetic phenotype).
3. Contractile, which occurred at the end of pregnancy and coincided with low myometrial expression of interstitial matrix proteins and high expression of components of the basement membrane (laminin and collagen IV).
In humans, restrictions on tissue access mean that compari-sons are largely limited to those between pregnant women delivered in labor at term and women delivered before the onset of labor. Gene microarray studies suggest that various cellular processes, including inflammation, transcriptional regulation, and intracellular signaling, are upregulated in laboring com-pared with nonlaboring myometrium, with these processes overlapping but being slightly different from those occurring in the cervix and fetal membranes.325,326 Notwithstanding the important contribution that arrays made to understand-ing of myometrial physiology, it is increasingly recognized that computerized modeling has much to contribute to the understanding of uterine contractions. A model would inte-grate state-of-the-art knowledge in cardiac electrophysiology, biochemistry/gene expression, and anatomy, and it would provide an in silico arena for testing of novel therapies. This approach, already well advanced in cardiac pathophysiology,327 is at a much earlier stage for pregnant uterine physiology.328
CERVICAL ADAPTATION AND REMODELING DURING HUMAN PREGNANCY
Traditionally, it was held that the closed cervix holds the fetus inside the uterus, and that progressively more forceful myome-trial contractions lead to cervical effacement and dilation.329 However, 2D and 3D ultrasound evaluation of the cervix estab-lished that during human gestation, cervical shortening and decreases in cervical volume often occur at an “asymptomatic” stage, before the onset of uterine contractions.330,331
As noted, it was recently proposed that classification of the preterm parturition syndrome based on phenotype, rather than on clinical signs or symptoms, may facilitate a better under-standing of the etiology of PTB.10 From this perspective, a short or a dilated cervix may be the first clinical manifestation of a parturition process triggered as a result of decidual activation332 or uterine contractility.330 The complexity of the issue is empha-sized by the observation that in the Preterm Prediction Study, a short cervix (≤2.0, ≤2.5, ≤3.0 cm), as seen by sonography, had a low sensitivity but a high specificity for prediction of PTB before 35 weeks.330,333 A cervical length of 2.5 cm or less at 22 to 24 6
7 weeks was associated with spontaneous PTB in only 18% and 27% of women prior to 35 and 37 weeks, respectively. This suggests that the majority of women with a short cervix by sonography will not deliver prematurely. On the basis of these observations, Iams and colleagues proposed that a short cervical length may represent a spontaneous preterm parturi-tion phenotype characterized by asymptomatic shortening of the cervix, but not decidual and myometrial activation.330
Spontaneous Preterm Parturition as a SyndromeIt is increasingly clear that preterm labor is not a single disease, but a syndrome with multiple causes. The classification system used in this chapter is the system proposed by a project funded by the Global Alliance to Prevent Prematurity and Stillbirth.10 Because the etiology of preterm labor is often not known, this system has deliberately avoided classification based on cause and has chosen a system based on phenotype. The phenotypes are based on the following: (1) significant maternal conditions (e.g., extrauterine infection, clinical chorioamnionitis, maternal trauma, worsening maternal disease, uterine rupture, preeclampsia/eclampsia), significant fetal conditions (e.g., fetal demise, IUGR, abnormal fetal heart rate or biophysical profile, infection or fetal inflammatory response syndrome, fetal anomaly, alloimmune fetal anemia, polyhydramnios, multiple fetuses), and pathologic placental conditions (e.g., histologic chorioamnionitis, placental abruption, placenta previa); (2) the presence or absence of signs of initiation of parturition; and (3) whether the pathway to delivery is caregiver initiated or spon-taneous. The aim is to provide a classification system to “use in both population surveillance and research, so that when specific types of PTBs are discussed, studied, or compared across popu-lations or over time, categories have consistent definitions that are widely understood and accepted.”10 Such an aim is laudable, and this is the classification system followed here, but before it is widely adopted and used, a paradigm shift will have to occur in many clinicians’ approach to and understanding of PTB (see Fig. 39-1).
MYOMETRIAL CONTRACTILITY
Myometrial contractions are a hallmark of parturition, both at term and before term. The biochemistry of myometrial con-tractility has been extensively reviewed.310,311 Contraction of individual myocytes is achieved by increasing intracellular calcium levels, which ultimately promotes phosphorylation of myosin, and hence increased actin-myosin cross-links and contraction. During labor, the individual myocytes contract together as a functional syncytium. This increased coordination is induced by gap junction formation, which increases cell-to-cell communication. Gap junctions develop in myometrium before labor and disappear after delivery.234,312-315 Expression of gap junction protein, connexin-43, in human myometrium is similar in both term and preterm labor.241,316-319 These findings suggest that the appearance of gap junctions and increased expression of connexin-43 (contraction-associated pro-teins318,320,321) are part of the underlying series of molecular and cellular events responsible for the switch from contractures to contractions before the onset of parturition. Estrogen, proges-terone, and prostaglandins have all been implicated in the regu-lation of gap junction formation, and they influence connexin-43 expression.67,322,323
Lye and colleagues324 proposed that the myometrium under-goes sequential phenotypic remodeling during pregnancy. Their studies were undertaken in rodents but have implications for humans. Three distinct stages of rat gestational myometrial development were recognized:
1. Proliferative, in which the number of myocytes increased, as demonstrated by greater levels of cell nuclear antigen labeling and protein expression in early pregnancy. This

39 Pathogenesis of Spontaneous Preterm Birth 611
The net enzymatic activity of MMPs, if there is any, is modulated by their interaction with tissue inhibitors of MMP (TIMPs) and various cytokines.354 Peptidyl lysine oxidase, copper, and vitamins C and E are also important regulators of collagen metabolism, directly involved in its synthesis and degradation.355,356
Animal studies have generated a large body of knowledge about processes involved in pregnancy-related changes to the cervix.337 From this research we have learned that before cervical ripening, the collagen is dense, organized, rigid, and not exten-sible.339 Collagen’s 3D structure, which limits access of degrad-ing collagenases and permits cross-linking between fibrils, might play a role. By the end of the first trimester, collagen becomes less tightly packed.337 As a result, the cervix becomes softer but retains its tensile strength. Cross-links between col-lagen molecules are essential for providing strength. Several investigators have focused their attention on decorin (dermatan sulfate proteoglycan), which seems to be implicated in the process of collagen reorganization and cross-linking.357,358 Animal studies revealed that an increase in the decorin-to-col-lagen ratio was associated with disorganization and rearrange-ment of collagen fibrils, followed by a marked decrease in mechanical strength.359
Orientation of the collagen fibrils is not the only element involved in regulating the preparative process of the cervix for labor. For example, a decline in the collagen type I mRNA expression was observed in human gestation, suggesting that a decreased synthesis of this collagen could be involved in the process of uterine cervical ripening.360 This finding was in agreement with light-induced autofluorescence measurements of the human cervix.361 By using this noninvasive technology, Maul and coworkers provided evidence that a gradual decrease in cervical collagen concentration occurs with advancing gestation.361
Other elements of the extracellular matrix, such as proteo-glycans, elastin, and its hydration status, reliant on negatively charged glycosaminoglycans and levels of vascular endothelial growth factor (VEGF), are also considered important determi-nants of cervical biomechanics.362,363 Glycosaminoglycans such as hyaluronic acid are distributed widely throughout connective tissues, including the uterine cervix.364 These molecules have a high affinity for water and therefore may control tissue hydra-tion, which is an essential element of cervical ripening.365 A decrease in collagen content was repeatedly proposed as one of the mechanisms responsible for cervical ripening.339 The high affinity of the glycosaminoglycans for water may artificially decrease the cervical collagen concentration. This premise is supported by studies that refute changes in collagen content of the cervix across gestation.342
Hyaluronidase is an intrinsic enzyme that catalyzes the hydrolysis of hyaluronic acid, effectively creating low-molecular-weight hyaluronic acid molecules.364 This catalyzing action lowers the viscosity of the hyaluronic acid, thus increasing tissue permeability to water. Hyaluronidase modifies the tensile visco-elastic properties of the rat cervix, but its role in human cervical remodeling remains to be determined.366 Elastin may have a role in cervical dilation and tissue compliance to stretch. This con-clusion is mostly supported by histologic data demonstrating fragmentation of the elastin fibers in women with an incompe-tent cervix.367
Relaxin is a two-chain peptide hormone that serves an important role in cervical growth and remodeling associated
Understanding the process of cervical functional adaptation to pregnancy has become critical for a better comprehension of the mechanisms responsible for initiation of human parturi-tion, cervical insufficiency, and spontaneous preterm labor. Today, it is recognized that cervical biology undergoes major enzymatic and biomechanical transformations that differ from those of the myometrium.334,335 Thus, although anatomically part of the uterus, the cervix should be viewed as a separate, complex, and heterogeneous organ.336
For most of a normal human gestation, the cervix remains closed and firm. The current working model of parturition indicates that the cervix must undergo a multistep adaptive process: (1) softening (chronic, slow, progressive); (2) ripening (precedes labor); (3) effacement and dilation (acute, occurs within hours); and (4) repair (occurs after delivery for several weeks).336,337 Each of these phases involves distinct biochemical, biomechanical, and molecular events, which could be phenotype dependent. This assertion is supported by studies conducted in animals with various genetic backgrounds (high-regenerative repair versus low-regenerative high-fibrotic repair).338
The cervix is a composite viscoelastic material consisting of elastic (collagen and elastin) and viscous macromolecular com-ponents (sulfated glycosaminoglycans and proteoglycans).339 The ratios of constitutive elements of the cervix vary by the region of the cervix they occupy.340,341 During each phase of the adaptive process, the complex interaction between connective tissue, extracellular matrix (collagen, elastin, macromolecular proteoglycans), smooth muscle, and fibroblasts dictates the mechanical behavior of the uterine cervix.342
Collagen makes up almost 90% of the cervix343 and is believed to be the most critical element responsible for maintenance of tissue structural integrity.339,344 Major cervical collagens are types I and III.345 Interestingly, interstitial collagens types I and III also predominate and maintain the mechanical integrity of the amnion.252 This observation implies that various factors (e.g., inflammation) may modify the biology of the cervical and fetal membrane tissues in parallel. Collagen is actively synthe-sized during pregnancy, and it is remodeled by the interplay of neutrophils, fibroblasts, and various enzymatic pathways.346-348 The role of MMPs in cervical ripening remains incompletely understood.337 A possible role of MMPs in the process of adap-tation and collagen remodeling was supported by data showing that in pregnant rabbits the antiprogesterone onapristone (ZK 98.299) augmented the cervical mRNA expression levels of MMP-3 (or stromelysin-1).349 In addition, studies conducted in rodents revealed that systemically administered PGE2 elevated the cervical tissue levels of MMP-2 and MMP-9.350 This effect was predominantly seen at term, not before term. However, the role of collagenases during the process of cervical ripening has been challenged.351 Incubation of the cervical tissue with MMP-1 altered neither the stiffness nor the extensibility of the rat cervix. Biomechanical experimentation revealed that the changes in physical properties of the rat cervix during physio-logic ripening are similar to those induced by PGE2 and anti-progestin, and they consist of increased extensibility, compliance, and strength. They cannot be attributed to increased collage-nase activity, which decreases tissue compliance and strength.351 Studies conducted in healthy pregnant women suggest that the functional relevance of MMPs is probably minimal.352,353 This assertion is based on the observation that cervicovaginal MMP-9 did not change with spontaneous labor or rupture of mem-branes at term and did not predict success of labor induction.352

612 PART 4 Disorders at the Maternal-Fetal Interface
the clinical data demonstrating that administration of the anti-progesterone RU486 increases the likelihood of a favorable cervix.300 However, RU486 alone is not sufficient to induce labor, implying that factors involved in controlling the activa-tion of myometrial contractility play a decisive role. Under-standing the molecular mechanism by which progesterone maintains a state of cervical competency proved to be a chal-lenge.384 It has been postulated that alterations in the expression of PR isoforms and changes in the metabolism of estrogen and progesterone are associated with cervical changes in human parturition.337,385 Data generated using mice deficient in steroid 5α-reductase type-1, an enzyme with an essential role in cervical progesterone catabolism, indicate that at least part of progesterone’s effect on cervical remodeling is controlled by this enzyme.386 It has been also proposed that in the cervix, progesterone is an important regulator of hyaluronic acid and MMP metabolism, and it affects the intensity of an inflam-matory response after activation of various inflammatory pathways.201,387,388
The laboring cervix is histologically characterized by an abundance of neutrophils and macrophages, and by an out-pouring of proinflammatory cytokines.389,390 Young and col-laborators reported that in the human cervix, IL-6, IL-8, and TNF-α were localized to leukocytes, glandular and surface epi-thelium, and stromal cells.391 Although these data might argue that the cervical biology is heavily dependent on various inflam-matory processes, especially at term, it is important to recognize that during ripening, the influx of monocytes into the cervix depends on the loss of progesterone function.392 Furthermore, the timing of inflammatory cell migration and activation in the pregnant cervix of mice deficient in 5α-reductase type-1 (Srd5a1−/−) suggests a role for the inflammatory cells and acti-vation of downstream signaling pathways of various cytokines, in postpartum remodeling rather than in the cervical ripening phase.393 As animal and human labor begins and the cervix dilates, there is increased activity of inflammatory mediators such as IL-1β and IL-8 that can activate various NF-κB–depen-dent signaling pathways.394,395 Expression of proinflammatory cytokines stimulates synthesis and activation of collagenases, elastases, MMPs, and possibly nitric oxide synthases.396 The increase in IL-6 stimulates prostaglandin and leukotriene pro-duction, potentially causing dilation of cervical vessels and pro-moting extravasation of various inflammatory cells.176 Proteases released by degranulating neutrophils encounter an already destabilized collagenous fiber network. In this context, collagen disorganization can be further augmented by collagenases, even in the absence of a significant change in their level of expression. If true, the process should be strictly time limited, because the sustained action of proteases may cause severe tissue damage. Differential expression of nitric oxide synthase in the uterus and cervix during pregnancy has been described.334 Nitric oxide production is upregulated in the cervix during labor, an effect that is opposite from that in the myometrium (i.e., anti-contractile).335 This increase is accompanied by softening of the cervix, and blockade of nitric oxide reduces cervical distensibil-ity.335 The mechanism of nitric oxide–induced cervical ripening during pregnancy may be mediated in part by increased PGF2α,397 but not by cytokine synthesis.398
What can be concluded from the large body of published research is that cervical adaptation to pregnancy cannot be the result of a single factor, and that various pathways should be involved simultaneously. Indeed, various genes, with roles in
with pregnancy.368,369 Specifically, relaxin-deficient mice display difficult parturition, an event also observed in relaxin leucine-rich repeat containing G protein–coupled receptors (LGR) knockout mice.369 In humans, relaxin may manifest its effect via an increase in collagenase activity and increased glycosamino-glycan synthesis.368 Other collagen accessory proteins such as thrombospondin-2, tenascin-C and lysil-hydroxylase, with an important role in collagen cross-linking, seem to be also involved.370,371
Significant gestational changes also occur in the cellular compartment of the cervix.341 Human studies demonstrated that apoptosis of stromal cells may be involved in cervical remodeling.372 A higher number of apoptotic nuclei were seen in laboring than in nonlaboring cervix, suggesting an increased rate of apoptosis as pregnancy progresses to term. Based on animal studies, it was proposed that relaxin, estrogens, and progesterone are important regulators of apoptosis.373 Interest-ingly, all of these three hormones promoted cell proliferation and repressed apoptosis by unknown mechanisms. Availability of co-regulatory proteins (nuclear receptor co-repressor tran-scriptional factor) at different stages of pregnancy, or the local ratios of various hormones may contribute to the process.374 Apoptosis is followed by infiltration of macrophages and neu-trophils that add to disorganization and dispersion of the col-lagen and elastin fibers.343 The increase in decorin expression and the resulting collagen disorganization promote the influx of water, enhancing the ability of the cervix to dilate during labor.344
At the end of pregnancy, changes in the mechanical behavior of the cervical tissue are the result of various biochemical trans-formations.375 What remains incompletely understood is the nature of the signals responsible for coordination of the process of cervical ripening, softening, and dilation. Chief candidate regulatory molecules are hormones (e.g., prostaglandins, pro-gesterone), cytokines, and decorin.376 Prostaglandins induce a marked increase in the decorin-to-collagen ratio,377 which in turn may provoke collagen disorganization and rearrangement of its 3D conformation. In various studies, prostaglandin-induced cervical ripening was characterized by extracellular matrix transformations similar to physiologic ripening such as diminished collagen concentration, increased synthesis of hydrophilic proteoglycans, and increased collagen solubility.378 The mechanisms are extremely complex and may involve aug-mentation of glycosaminoglycan synthesis by IL-1β, and the neutrophil chemoattractant IL-8.71 IL-8 is also thought to initi-ate cervical ripening by promoting neutrophil chemotaxis to and activation in cervical stroma.71 Interestingly, prostaglandins had no effect on hyaluronic acid synthesis, so tissue hydration most probably entails other mechanisms. Additional factors involved may include relationships among prostaglandins and the receptors for estrogen (EP4), androgens, and progesterone (PR-A, PR-B).379
The important role of progesterone for maintenance of cer-vical competency has long been postulated.380 Today, the atten-tion is even more focused on progesterone because of the results of randomized clinical trials that pointed to its potential role in prevention of premature birth.381,382 In an animal model, Stys and colleagues demonstrated that progesterone has different effects in the myometrium and in the cervix, supporting the hypothesis that it prevents term and PTB by acting on both myometrium and cervix.383 In humans, the contribution of pro-gesterone to the process of cervical remodeling is supported by

39 Pathogenesis of Spontaneous Preterm Birth 613
the cervix of inflammation-induced PTB.410 On the other hand, term labor was not associated with a differential expression of immune pathways. Genes responsible for the expression of various cytokines (TNF-α, IL-6, IL-1β) and MMPs (MMP-2, MMP-9) were upregulated in the cervix of animals with intra-uterine infection.410 Notably, expression of cytokines was not increased in term cervices but was amplified in the postpartum period. These findings argue that, in the postpartum period, cytokines may play an important cervical reparative role or may confer immune protection.
Premature cervical ripening is a feature of patients with mul-tiple gestations, and is rarely seen in diethylstilbestrol-exposed women.411,412 IL-8,413,414 MMPs,31,348 prostaglandins,71,337,415 and nitric oxide335 have been implicated in the control of cervical ripening. These mediators may be synthesized in response to amniochorion stretch and may exercise part of their biologic effects in parturition by degradation of the cervical extracellular matrix.
It has been suggested that cervical insufficiency is character-ized by a “muscular cervix,” with low collagen and high smooth muscle content.416 However, cervical insufficiency does not appear to be associated with a constitutionally low collagen content or collagen of inferior mechanical quality. Therefore, the hypothesis that a muscular cervix with an abundance of smooth muscle cells contributes to the development of cervical insufficiency is not supported by the present studies aimed to address this problem.
Collectively, the data presented here support the view that PTB is not an accelerated form of physiologic cervical ripening.
DECIDUAL ACTIVATION AND BLEEDING
Decidual activation refers to a complex set of pathophysiologic events that lead to separation of the fetal amniochorionic mem-branes from the decidua, bleeding, spontaneous rupture of the membranes, activation of myometrial contractility, and expul-sion of the placenta. The relative contributions of the decidual activation and bleeding process to PTB vary.
During gestation, the chorioamniotic membranes gradually fuse with the decidua. Before delivery, biochemical and molecu-lar occur that allow separation and expulsion of the fetal mem-branes. Fibronectins have multiple binding sites to permit cell binding and interaction with cytoskeletal organization to effect cell migration, adhesion, and decidual cell differentia-tion.417-419 Elastase-induced release and degradation of the gly-cosylated cellular fibronectin (i.e., fetal fibronectin) diminishes the binding ability of this glycoprotein for components of the extracellular matrix, thereby facilitating separation of the fetal membranes from the decidua.420 Detection of fetal fibro-nectin into cervical and vaginal secretions before preterm parturition421-423 is one of the most clinically useful biomarkers of PTB.424-426
Placental abruption can be viewed as a binary event in which molecular signals involved in decidual bleeding arise as a con-sequence of either inflammation or aberrant coagulation (Fig. 39-10). Although relatively distinct from other causes of prematurity, it is believed that many of the molecular events responsible for decidual activation and abruption are inflammatory. The acute lesions of chorioamnionitis and funisitis (e.g., hematoma, fibrin deposition, compressed villi, hemosiderin-laden histiocytes) are frequently associated with
cell adhesion, regulation of extracellular matrix, and inflamma-tion, were found to be differentially expressed in the ripened cervix.326,399 Current data reveal that cervical shortening at term was associated with downregulation of bone morphogenetic protein-7, claudin-1, β6 integrin, and endometrial progesterone-induced protein mRNA. The clinical significance of these dis-coveries remains to be determined.400
CERVICAL ADAPTATION AND REMODELING DURING PRETERM BIRTH
Cervical adaptation and remodeling is a complex process. However, a relevant question is whether cervical ripening and dilation at term and preterm are driven by the same molecular mechanisms. A study by Akins and colleagues demonstrated that collagen morphology in premature cervical remodeling is different from that in physiologic term ripening.401
Because the processes of term and preterm cervical ripening and dilation have been associated with various inflammatory events, inflammation was consistently presented as the final common pathway of parturition.326 This hypothesis was sup-ported by data that demonstrated that in the absence of infec-tion, IL-8, IL-6, and monocyte chemotactic protein-1 (MCP-1) were significantly elevated in human term and preterm cervical tissues.402 As shown, asymptomatic women with intra-amniotic infection or inflammation had higher degrees of cervical short-ening and dilation.110,403-405 Thus, it is reasonable to assume that in some clinical circumstances, a short or a dilated cervix favors microbial invasion of the gestational sac via an ascending mech-anism.125 In such cases, processes involved in premature cervical shortening and dilation might represent the first event leading toward premature birth. However, the reverse is possible—that is, intra-amniotic infection may induce an outpouring of inflammatory mediators that in turn might persuade ripening and opening of the cervix.
The clinical implication is that presence in the cervicovaginal fluid of specific biomarkers may reflect activation of signaling pathways involved in premature ripening, cervical dilation, and premature birth. Additional nuances concerning the involved molecular networks have been described with the recent advances in genomic and proteomic technology.406,407 Compre-hensive survey of the cervicovaginal fluid proteome showed that calgranulin-A, calgranulin-B, azurocidin, insulin-like growth factor–binding protein-1, and defensins were found upregulated in the cervicovaginal fluids of women at risk for PTB.406,407 Most of these biomarkers are part of the innate immunity defense mechanism of the cervix and are constitu-tively expressed in the cervical mucus.407 Their differential expression408 may mediate the process of leukocyte infiltration, shown previously to be involved in cervical remodeling.390
To understand the process of cervical ripening and dilation in the setting of intra-amniotic infection-induced PTB, several investigators have used animal models of intrauterine infection. With respect to the profile of cervical inflammatory infiltrate, Holt and coworkers suggested that there might be differences between various mechanisms involved in triggering PTB.409 The monocyte population dominated the progesterone withdrawal mechanism, whereas neutrophils governed the process of endotoxin-induced PTB. A comparison of the genes differen-tially expressed in the cervix of animals in which PTB was induced by mifepristone or endotoxin showed that genes involved in immunity and inflammation were upregulated in

614 PART 4 Disorders at the Maternal-Fetal Interface
delivery is defined by distinct transcriptional profiles.435 Studies demonstrating that inflammatory reactions (dependent or independent of infection) can activate the coagulation pathways emphasize the important role of inflammation in decidual bleeing.429 Cytokines (e.g., IL-1β, IL-6) act on decidual vascular surfaces and increase the expression of leukocyte interactive proteins such as P-selectin, E-selectin, vascular cell adhesion molecule, and intercellular adhesion molecule-1.436-438 This phe-nomenon may lead to decidual neutrophil infiltration, vascular damage, access of coagulation factors (factor VII) to the peri-vascular adventitial tissue factor, and generation of thrombin.439 Rosen and colleagues proposed that when the process of decid-ual thrombin activation overwhelms the physiologic anticoagu-lant and fibrinolytic response, the abruption process becomes systemic, as assessed by circulating maternal plasma thrombin-antithrombin complexes.440 Furthermore, this phenomenon was mechanistically linked to adverse pregnancy outcomes, such as preterm PROM.441
Decidua is a rich source of tissue factor, the primary initiator of coagulation.439 The role of decidual cell–expressed tissue factor in preservation of uterine hemostasis and its involvement in the abruption process has been summarized in several excel-lent reviews.439,442 Taylor and colleagues demonstrated in an animal (dog) model of LPS-induced sepsis that infusion of low concentrations of thrombin was protective against death.443 Thus, it is possible for low levels of thrombin generated in the early inflammatory phase of an abruption process to be benefi-cial. This may explain the high frequency of histopathologic lesions suggestive of chronic decidual bleeding in the absence
histologic abruption.427 Histologic evaluation of the vasculopa-thy attending decidual hemorrhage provides evidence that the nature of the damaging process is frequently chronic.83,428,429 The most frequent histopathologic lesions suggestive of chronic decidual bleeding are chronic deciduitis and villitis, infarct and necrosis, spiral vessels with absence of physiologic trans-formation and increased numbers of circulating nucleated erythrocytes, vascular thrombosis, and villous fibrosis and hypovascularity.428
Maintenance of appropriate hemostasis in human decidua is central to normal human implantation and placental develop-ment.430 Survival of the embryo and development of the fetus requires that extravillous trophoblasts gain access to the mater-nal circulation by penetrating the uterine spiral arteries without causing hemorrhage.431 This process is gradual and well coor-dinated. Disarray of the highly controlled and synchronized molecular mechanisms at the maternal-fetal interface increases the risk for hemorrhage, leading to abortion, abruption, and stillbirth. The key histologic finding in placental abruption is hemorrhage in the decidua basalis.432 This hemorrhage is believed to result from pathologic processes damaging the vas-cular endothelium.433
Incorporating the pathophysiology of abruption in the framework of inflammation or bleeding alone is difficult. This is because the molecular signals involved in decidual bleeding are potentially triggered by both pathologic inflammation and aberrant coagulation.15,434 Interestingly analysis of the expressed decidual transcripts and proteins suggests that each spontane-ous, infection-induced, and abruption-associated preterm
Hypoxia FR / ROS
↑ Decidual TF expression
↑ Thrombin
Abruption induced PTB
Inflammation Inflammation
↑ sFlt1
↓ Angiogenesis
↑ Decidual cellVEGF
Reduced or intermittentuteroplacental blood flow
Decidual endothelialcell dysfunction
1st t
rimes
ter
Figure 39-10 Binary theory of decidual bleeding and inflammation in pathogenesis of placental abruption–induced preterm birth (PTB). A reduced or intermittent uteroplacental blood flow causes focal decidual hypoxia and free radicals (FR) and reactive oxygen species (ROS) (through reperfusion injury). Hypoxia induces the expression of decidual vascular endothelium growth factor (VEGF). This angiogenic factor acts directly on decidual endothelial cells to enhance permeability and degrade the vascular wall through matrix metalloproteinase (MMP)-2 generation. This leads to hemorrhage and aberrant endothelial expression of tissue factor (TF), generating additional thrombin that further induces TF expression and uteroplacental thromboses, which exacerbate reduced blood flow. Free radicals and ROS induce endothelial cell injury, which allows perivascular leakage of coagulation factors, including factor VIIa, which then comes in contact with TF, activating the extrinsic coagulation pathway. The resulting thrombin further induces TF expression as well as expression of inflammatory cytokines, leading to inflammation in the absence of a microbial attack. sFlt1, soluble fms-like tyrosine kinase-1.

39 Pathogenesis of Spontaneous Preterm Birth 615
hypothesis is that progesterone withdrawal may induce an increase in the decidual expression of MMP-1 and MMP-3, which would promote extracellular matrix and fibrillar collagen degradation, preceding bleeding, premature separation of the placenta, and preterm PROM.
Taken together, the results of these studies may explain at least partially the potential role of progesterone in preventing decidual activation and bleeding, by inhibiting the general proteolytic and inflammatory activity at the maternal-fetal interface. The mechanism by which progesterone function is balanced in the setting of high maternal circulatory levels is unknown. Abruption-associated PTB is accompanied by reduced immunostaining for PR, and thrombin inhibits PR protein and mRNA expression in cultured term decidual cells.460a
Epidemiologic studies suggest that for some women, the recurrence risk for placental abruption is higher than the general background risk, and that thromboembolic events occur more commonly among female relatives of women with placental abruption.461-463 However, genetic studies involving candidate genes dispute this association.464,465 Zdoukopoulos and Zintzaras tried to identify the most frequent gene polymor-phisms associated with placental abruption.466 A positive asso-ciation was identified for Arg506Gln (OR = 3.4; 95% CI, 1.4 to 8.3) and G20210A (OR = 6.7; 95% CI, 3.2 to 13.0) polymor-phisms. The positive correlation between placental abruption and prothrombin gene mutation (G20210A) was confirmed in nulliparous women (OR = 12.1; 95% CI, 2.4 to 60.4).467 On the other hand, a study conducted by the Maternal-Fetal Medicine Units Network argues against this association, suggesting that the practice of screening women without a history of thrombo-sis or adverse pregnancy outcomes for the G20210A prothrom-bin gene mutation polymorphism is not necessary.468
PRETERM PREMATURE RUPTURE OF MEMBRANES
Preterm PROM (pPROM) is one of the leading risk factors for PTB. The amniochorion has a unique biology characterized by distinctive molecular, enzymatic, and biomechanical transfor-mations.235 Multiple pPROM risk factors have been identi-fied.11,469 Whatever the cause, the final common pathway must be weakness of the amniochorion membrane that allows rupture. Clinicians and scientists have traditionally attributed rupture of the membranes to mechanical stress, particularly that associated with uterine contractions. The molecular mech-anisms that could trigger pPROM after excessive mechanical stretch were presented earlier.
Fetal membranes are pluristratified structures whose com-position ensures their cohesion, elasticity, and mechanical strength. The strength of the fetal membranes is derived from both synthesis- and protease-induced degradation of the com-ponents of the extracellular matrix. The mechanisms respon-sible for degradation of the extracellular matrix are tightly regulated by several MMPs (MMP-1, 2, 3, and 9) and TIMP-1 and TIMP-2.11 A marked decidual infiltrate of neutrophils, which are a rich source of the extracellular matrix–degrading proteases elastase and MMPs, are associated with abruption-induced pPROM.428,429 This phenomenon occurs in the pres-ence or in the absence of infection. Predominant activities for MMP-1,470 MMP-8,471 and MMP-9,472 and a low concentration of TIMP-1 and TIMP-2,473 have been demonstrated in the amniotic fluid of women with pPROM. This implies that at least part of the MMPs’ bioavailability at the site of
of clinical manifestations of the disease.428,429 Various processes that lead to vascular disruption and the interaction of circulat-ing factor VIIa with decidual cell membrane–bound tissue factor generate thrombin, explaining the strong association between abruption and disseminated intravascular coagula-tion.444 The mechanisms responsible for the stimulation of myometrial contractions in the presence of intrauterine hemor-rhage and the specific role played by the thrombin have been defined.445-447 Elovitz and coworkers demonstrated that intra-uterine inoculation of whole blood stimulated rat myometrial contractility.445 Furthermore, fresh whole blood stimulated myometrial contractility in vitro, and this effect was partially blunted by hirudin, a thrombin inhibitor.445 In vitro, thrombin induced cytosolic calcium concentration oscillations that were similar to those produced by oxytocin.445 These studies con-firmed that membrane receptor-Gq protein and protease-activated receptor-1 (PAR1) coupling events play an important role in modulating thrombin stimulation of myometrial smooth muscle.445,447
Thrombin may exert pleiotropic effects on decidual cells. Genomic studies have provided a better understanding of the process of endometrial decidualization.448 Among the genes responsible for the normal phenotypic and morphologic remodeling processes of the human decidua, the homeobox (HOX) gene family appears to be critical.449 In vitro, thrombin decreased gene expression of HOXA9, HOXA10, and HOXA11. Furthermore, thrombin decreased HOXA10 mRNA and protein levels. IL-1β, a cytokine with important regulatory roles in pros-taglandin production,450 mimicked the effect of thrombin on HOXA10 gene expression. These observations provide proof of the concept that two recognized mediators of decidual inflam-mation and PTB, thrombin and IL-1β, may operate by reducing the expression of HOXA10 gene.
The inflammatory events that follow abruption can occur dependent on or independent of progesterone.451,452 Coincident with activation of the coagulation cascade, decidual injury causes release of cytokines.453 Cytokines act in an autocrine or paracrine manner to elicit and increase the synthesis of MMPs454 and VEGF.451 It has been proposed that TNF-α, IL-1β, IL-6, IL-8, and IL-11 are involved.453 Other cytokines, such as granulocyte-macrophage colony–stimulating factor (GM-CSF), monocyte chemoattractant protein-1 (CCL2), and colony-stimulating factor-1 (CSF1), may be implicated in the regula-tory process of decidual activation and bleeding.438,455
Progesterone appears to create both a hemostatic and an antiproteolytic milieu in the decidua. Using a primary cell culture experimental setup, Schatz and colleagues demonstrated that first-trimester decidua expresses tissue factor and plas-minogen activator inhibitor-1 (PAI-1), which is considered a fast inhibitor of the primary fibrinolytic agent tissue plasmino-gen activator (tPA).456 There is experimental evidence that pro-gestins are exercising a similar effect in cultured term decidual cells.457 The molecular mechanisms through which progestins promote decidual hemostasis via enhanced expression of tissue factor, the primary initiator of hemostasis, and PAI-1, the primary antifibrinolytic compound, involve epidermal growth factor receptor (EGFR) and induction of transcription factors Sp-1 and Sp-3.458
In addition to controlling the decidual hemostasis, proges-tins inhibit the proteolytic activity of collagenase MMP-1 and MMP-3.459,460 Because antiprogestins (e.g., RU486) reverse the progestin-inhibited expression of MMPs, an attractive

616 PART 4 Disorders at the Maternal-Fetal Interface
membranes is reduction-oxidation (redox) regulated.489 By using an in vitro amniochorion explant model, it was shown that MMP-9 activity was directly increased by superoxide anion, a byproduct of macrophages and neutrophils that are abun-dantly present in both amniochorion and decidua of pregnan-cies complicated by infection and decidual hemorrhage. N-Acetylcysteine decreased the amniochorionic MMP-9 activ-ity, suggesting that this glutathione precursor may have thera-peutic value.489
Vaginal bleeding is a well-recognized risk factor for PTB. Women who are experiencing vaginal bleeding during the first trimester have an increased risk for PTB (adjusted relative risk [RR] = 2; 95% CI, 1.6 to 2.5).490 Moreover, the risk for pPROM is increased if vaginal bleeding persists more than one trimester during gestation (OR = 7.4; 95% CI, 2.2 to 25.6).491 Much inter-est has been shown about understanding the relationship between decidual bleeding, activation of MMPs, and pPROM. It was hypothesized that in abruption thrombin generated from decidual cell–expressed tissue factor can indirectly promote pPROM via enhanced expressed MMP-1 and stromelysin-1 (MMP-3).441,492 Furthermore, in vitro, thrombin enhances the expression of MMP-9 in fetal membranes at term.493 Along with data confirming that the maternal coagulation system is acti-vated in women with ruptured membranes,441 the results of these experiments support the premise that thrombin is central to the pathophysiology of pPROM. Kumar and colleagues demonstrated that thrombin and TRAP (specific agonist for thrombin receptor PAR1) weaken the fetal membranes in a dosage-dependent manner.494 Thrombin appears to exercise this effect in a direct fashion, whereas cytokines such as TNF-α and IL-1β require the presence of choriodecidua.494 Interestingly, thrombin, but not TNF-α and IL-1β, exhibited protein MMP-9 and decreased TIMP-3 production in isolated amniochorion cells. Because thrombin-induced decreases in biomechanical strength of the amniochorion are reversed by α-lipoic acid, it is probable that PKB/Akt, NF-κB, or nuclear factor–erythroid 2–related factor 2 (Nrf2) signal transduction pathways, or more than one of these, are directly involved.494,495
Phenotypic Components of the Preterm Birth SyndromeMATERNAL CONDITIONS
Extrauterine InfectionGiven the very close link between intrauterine infection and preterm labor (reviewed later), it is not surprising that there is also a strong association between maternal extrauterine infec-tion and PTB.
Globally, maternal infections with human immunodefi-ciency virus (HIV) and malaria are important contributors to PTB. In a large population-based study, women infected with HIV were more likely than uninfected women to give birth before term.496 In this study, antiretroviral treatment of HIV reduced rates of adverse outcomes, including PTB, but the spe-cific antiretroviral agents used were not reported. Although this study did not differentiate between spontaneous and caregiver-initiated PTB, others have shown that both are increased in women infected with HIV.497 Accumulating evidence now sug-gests that the use of modern highly active antiretroviral therapy (HAART, which involves multiple drugs and clearly reduces
amniochorion injury and rupture is the result of activation and degranulation of fetal neutrophils.198,474 In addition to collagen, gelatinases cleave fibronectin and proteoglycans, facilitating detachment of fetal membranes from the lower uterine segment, disruption of the extracellular matrix, and rupture.11,475
Activation of MMPs and other proteases may occur depen-dent on or independent of inflammation and infection.472 The existence of a focal defective area, rather than generally weak-ened fetal membranes, has been proposed.476 This theory is based on the observation that an excessive matrix remodeling process occurs in the region of fetal membranes overlying the internal os of the cervix.477,478 Histologic examination of the “restricted zone of extreme altered morphology” reveals marked swelling and disruption of the fibrillar collagen network in the compact, fibroblast, and spongy layers of the fetal membranes.478 Observations by Bell and colleagues suggest that changes in the zone of altered morphology are more extensive in the setting of pPROM.479 A significant decrease in the amount of collagen type I, III, or V has been reported in the zone of altered mor-phology, and studies conducted by Lappas and associates dem-onstrate that remodeling of the extracellular matrix in the supracervical area is probably related to activation of MAPK/AP-1, and NF-κB-dependent signaling pathways.480,481 Enhanced expression of tenascin, an extracellular matrix protein expressed during tissue remodeling and wound healing, may also be involved.479,482 Identification of tenascin in the fetal membranes signifies the presence of injury and a wound healing–like response that may precede pPROM. Studies conducted by George and colleagues suggest that apoptosis is accelerated in the chorion of pPROM women, and that this phenomenon is of higher intensity in the setting of chorioamnionitis.483
Although the precise mechanism responsible for pro-grammed cell death in the fetal membranes is unknown, it is conceivable that cytokines play a role.484 Kumar and coworkers showed that TNF-α and IL-1β incubated amniochorion exhib-ited a dosage-dependent decrease in the mechanical force required to reach the breaking point of the tissue.485 Both TNF-α and IL-1β stimulated the production of immunoreac-tive MMP-9 and a decrease in TIMP-3, suggesting collagen remodeling and apoptosis in fetal membranes exposed to increased cytokine levels. The observation that α-lipoic acid inhibits cytokine-induced fetal membrane weakening suggests that this mechanism is NF-κB mediated.486 Fortunato and Menon showed that IL-1β increased caspase 2, 3, 8, and 9 activi-ties, whereas IL-6-treated membranes did not exhibit a signifi-cant change.484 However, the role of IL-6 and its alternative trans-signaling pathway remains to be determined, especially when the levels of this cytokine and that of soluble gp130 mol-ecule were found to be significantly decreased in women with pPROM and intra-amniotic infection compared with women with intra-amniotic infection and intact membranes.153 A pos-sible explanation for the low cytokine levels in the amniotic fluid of women with pPROM could be fetuin-mediated aggre-gation of amniotic fluid cytokines and proteins into calcifying nanoparticles that may play an important pathophysiologic role in rupture of the amniochorion (increasing necrosis and apoptosis).487
The observed reduced antioxidant glutathione peroxidase and superoxide dismutase enzyme activity in the supracervical area of the fetal membranes implies that local oxidative stress is an important factor predisposing to pPROM.488 Compelling evidence suggests that MMP activity in the human fetal

39 Pathogenesis of Spontaneous Preterm Birth 617
between intrauterine infection and inflammation (whether clinically apparent or not) and preterm labor under this heading of clinical chorioamnionitis (because of the consistency in pathophysiology), while acknowledging that subclinical infec-tion or inflammation may not strictly fulfill the recently pro-posed clinical phenotype of clinical chorioamnionitis.
Maternal Trauma and Uterine RuptureAlthough major maternal trauma is uncommon during preg-nancy, it is associated with an increased risk for immediate or delayed PTB. Population-based studies suggest that the rate of trauma sufficient to require hospitalization is around 2 to 3.5 per 1000 pregnancies.515,516 In some women, the trauma is so severe that immediate PTB occurs either as a result of maternal injury, or as a result of fetal or maternal death, or to facilitate maternal resuscitation, although the absolute risk for immedi-ate delivery is small. Women with major injuries who survive, and women with minor trauma, have about double the odds of either spontaneous preterm labor or placental abruption during the remainder of the pregnancy.515-517
Other forms of maternal trauma that make important con-tributions to PTB include treatment for preinvasive cervical carcinoma518 (see Cervical Disorders, later) and previous cesar-ean delivery, which leads to an increased risk for stillbirth in the subsequent pregnancy after around 34 weeks’ gestation (hazard ratio, about 2) compared with those with a previous vaginal delivery.519 Although previous cesarean delivery is a common antecedent of uterine rupture during labor, it is possible for uterine rupture to occur de novo. Regardless of the setting of the uterine rupture, PTB is commonly associated.520
Worsening Maternal Disease, Including PreeclampsiaThe final maternal phenotypic components of the PTB syn-drome include worsening maternal disease, preeclampsia, and eclampsia. These conditions are risk factors for caregiver-initiated pathways to PTB (typical OR = 3 to 5),14 although a recent population-based study demonstrated that maternal dia-betes and preeclampsia were also associated with increased odds of spontaneous preterm labor.14
Maternal Stress and AnxietyA large number of epidemiologic studies confirmed that mater-nal psychosocial stress is, worldwide, an independent factor for PTB.521-525 Although there is no universally accepted defini-tion of stress, it is generally the result of an interaction between a person and the environment in which there is a divergence between environmental demand and the individual’s psycho-logical, social, and biologic resources. From this perspective, the nature and timing of the stressful events may vary from a heavy workload to anxiety and depression.526,527 Population-based research conducted with pregnant women of different sociodemographic backgrounds and races showed that peri-conception stress and anxiety are independently associated with increased rates of spontaneous PTB.528,529 Stress during preg-nancy also has adverse consequences, with the risk for PTB apparently being higher when the stress exposure occurs during the second half of gestation (months 5 to 6) (OR = 1.24; 99% CI, 1.08 to 1.42).524 Because of the large heterogeneity of study designs, populations, and the associated behavioral risk factors, the magnitude of the effect is often difficult to determine. However, the general consensus is that the overall impact is modest.529,530
vertical transmission of HIV) is itself a risk factor for spontane-ous and caregiver-induced PTB, even after adjustment for confounders.498 The mechanisms of the increase in preterm labor associated with HIV infection and with HAART are unknown.
Malarial infection increases the risk for both stillbirth and PTB,499 probably by a combination of mechanisms including placental dysfunction, anemia, and maternal sepsis. In endemic areas, intermittent presumptive treatment for malaria is recom-mended during pregnancy to reduce perinatal mortality and low birth weight,500 although the evidence that such a strategy also prevents prematurity is weak.
More localized maternal extrauterine infections, whether symptomatic or asymptomatic, also increase the risk for preterm labor. The link between asymptomatic bacteriuria and PTB is well established.501 Women hospitalized with appendicitis are also more likely to give birth before term.502 These links also appear to hold true for emerging infections—for example, the risk for spontaneous preterm labor is some two to three times higher in women infected with the H1N1 influenza virus, with a significant increase in the risk for stillbirth and perinatal mortality compared with uninfected women.503 The pathophys-iology linking extrauterine infection to preterm labor is uncer-tain. For some infections, such as periodontitis, a mechanism by which mouth microorganisms induce bacteremia and hence reach the uterine has been postulated, 504 although evidence to support this claim is weak.
Clinical ChorioamnionitisLarsen and coworkers provided one of the first pieces of evi-dence that intrauterine infection is an important trigger for PTB.505 There is now compelling microbiologic evidence to suggest that intrauterine infection may contribute to about 25% of PTBs, with bacterial involvement as high as 80% for early gestation, declining to about 10% toward term (ascertainment of intrauterine infection notwithstanding).200,506,507
Clinically, intrauterine infection can manifest with obvious signs of maternal and or fetal infection (maternal fever, tachy-cardia, uterine tenderness, leukocytosis, and foul odor of the amniotic fluid). Multiple studies confirmed the non-overlapping nature of histologic and clinical chorioamnionitis.200,508,509 Clin-ical chorioamnionitis occurs in just 20% of pregnancies complicated by intra-amniotic inflammation and histologic chorioamnionitis. Because previous studies had associated short- and long-term follow-up characteristics to distinct pla-cental lesions,510-513 the results of histologic examination of the placenta (as performed by a perinatal pathologist) was used as an intermediate-outcome variable when evaluating perfor-mance of new diagnostic tests.514 Recognition of clinical cho-rioamnionitis is also a challenge in the setting of maternal systemic inflammatory conditions unrelated to obstetric causes (e.g., pyelonephritis, appendicitis, pneumonia).
In the newly proposed classification system, clinical chorio-amnionitis is defined as “clinically suspected intrauterine infec-tion, manifest by maternal fever and rupture of the membranes plus two features from maternal tachycardia, uterine tender-ness, purulent amniotic fluid, fetal tachycardia and maternal leukocytosis.”10 It is acknowledged that this somewhat strict definition would not include many women presenting in spon-taneous preterm labor and subsequently found, on careful testing, to have microorganisms in the uterine cavity. However, for the purposes of this chapter, we will describe all the links

618 PART 4 Disorders at the Maternal-Fetal Interface
implementation of behavioral interventions reduces the occur-rence of PTB (OR = 0.42; 95% CI, 0.19 to 0.93).543 Fourth, fetal exposure to maternal stress may have sustained programming effects on the HPA axis responsiveness and sympathetic nervous system functionality later in life, which seems to be sex (male) dependent.544
Complex biochemical and neurohormonal interactions between maternal, fetal, and placental compartments are required during normal and premature human parturition.79 A series of physiologic adaptive responses in each of these com-partments can be triggered by stress subsequent to malnutri-tion, infection, ischemia, vascular damage, and psychosocial factors.80 However, the nature of the stimulus whereby stress induces premature activation of the mechanisms involved in PTB remains unknown. The pathways by which stress can induce preterm labor are represented in Figure 39-11. There is substantial evidence that the placenta plays a central role in controlling the length of gestation and the onset of parturition in humans.81 Thus, placental histologic changes consistent with infection- and ischemia-induced fetal stress are far more common in patients with spontaneous PTB than in idiopathic preterm and term birth controls.82,83 Maternal-fetal trafficking of numerous hormones is highly dependent on various enzy-matic pathways. For example, 11β-HSD regulates placental transfer of cortisol, which is a glucocorticosteroid with a key role in activation of the HPA axis. Interestingly, hyperactivity of the maternal HPA axis has been involved in the occurrence of maternal depression.84 Carriers of a polymorphism in the gene encoding for 11β-HSD type-1 have a higher level of HPA activity and susceptibility to depression.85 Collectively, these and other data86 appear to indicate a genetic predisposition toward maternal mood disorders and may implicate various placental polymorphisms in the occurrence of maternal mood disorders linked to PTB.87
Cervical DisordersCervical insufficiency can produce a wide spectrum of diseases,545 including recurrent pregnancy loss in the mid-trimester and spon-taneous PTB later in gestation. The latter often appears with
Data derived from registry-linked births have noted slightly higher odds of PTB in women with post-traumatic stress dis-order, with an OR that varies from 2.5 (95% CI, 1.05 to 5.84)531 to 2.3 (95% CI, 0.82 to 6.38).532 Exposure to specific severe disaster events and the intensity of the disaster experience are considered predictors of poor pregnancy outcomes.532 Antena-tal depressive symptoms affect approximately 18% of pregnant women.533 What can be concluded from the large body of litera-ture is that during pregnancy, women with depression are at increased risk for PTB.534 The extent of the effect varies depend-ing on the method of ascertainment of depression, socioeco-nomic status, and race.535 For example, depression among African-American women is associated with an adjusted OR for PTB of 1.96 (95% CI, 1.04 to 3.72).536 Absence of similar find-ings in Hispanic and non-Hispanic white populations suggests ethnic disparity in the effect of stress in the United States.
The clinical relevance of the impact of various maternal stressors on the occurrence of PTB is multidimensional. First, comorbidity of depression and anxiety creates the context for the worst pregnancy outcome.537 Second, women with signifi-cant psychosocial stress factors and psychiatric conditions (depression, anxiety, bipolar disease) are frequently prescribed medications that may affect the growth and development of the fetus.538 However, data on the use of antidepressant medication during pregnancy is reassuring overall—treatment can be rec-ommended in view of the risks associated with absence of treatment.539-541 In a large study, exposure to selective serotonin reuptake inhibitors (SSRIs) during the first trimester was not associated with increased risk for congenital malformations in general (OR = 1.22; 95% CI, 0.81 to 1.84) or increased risk for PTB (OR = 1.21; 95% CI, 0.87 to 1.69). Others have shown a modest impact on PTB. Yonkers and colleagues542 found that use of an SSRI, both with (OR = 2.1; 95% CI, 1.0 to 4.6) and without (OR = 1.6; 95% CI, 1.0 to 2.5) a major depressive episode, was associated with PTB. A major depressive episode without SSRI use (OR = 1.2; 95% CI, 0.68 to 2.1) had no clear effect on PTB risk. Use of SSRIs in pregnancy was associated with increases in spontaneous but not medically indicated PTB.542 Third, early recognition of maternal stress and
Figure 39-11 Proposed pathways by which stress can induce preterm labor. ACTH, adrenocortico-tropic hormone; CAPs, contraction-associated pro-teins; CRH, corticotropin-releasing hormone; DHEA, dehydroepiandrosterone; E1-E3, estrone, estradiol, and estriol; EP1 and EP3, prostaglandin E receptors types 1 and 3; ER-α, estrogen receptor-α; FP, pros-taglandin F receptor; HPA, hypothalamic-pituitary-adrenal; PG, prostaglandins; PR, prostaglandin receptor; PROM, premature rupture of membranes.
Maternal stress
Activation of fetal HPA axis Placentalinsufficiency
Cortisol
CRH E1-E3
PG
Cervical change Preterm PROM Contractions
Myometrial (PR-A/B, and ER-!)enhances c-jun causingincrease in CAPs, FP, EP1, EP3
Placenta, membranes,and decidua
(!) CRH
ACTH
Fetal adrenalzone
DHEA/16-OH DHEA
Placentalsulfatases
!

39 Pathogenesis of Spontaneous Preterm Birth 619
A steady decline in fetal mortality at 28 weeks of gestation or more has paralleled the increase in late PTBs. It is reasonable to propose that better screening for syphilis, anemia, diabetes, IUGR, preeclampsia, and PTB, and the development of rigorous clinical protocols for evaluation of fetal heart rate, contributed to the observed reduced rate of stillbirth, at the expense of caregiver-initiated PTB. However, despite implementation of various clinical strategies, fetal deaths at 20 to 27 weeks of gesta-tion did not decline, thereby implying increased vulnerability of the fetus at this gestational age.
Among the major risk factors for stillbirth worldwide are uteroplacental vascular insufficiency, abruption, smoking, maternal medical conditions (e.g., lupus, chronic hypertension, antiphospholipid syndrome, thyroid disease, cholestasis, throm-bophilia), African-American race, maternal nutritional defi-ciencies, obesity, congenital and aneuploidy preeclampsia, infections (e.g., parvovirus, cytomegalovirus, syphilis, malaria, Listeria monocytogenes), chorioamnionitis, multiple gestations, and umbilical cord complications.557 Many of these are well-recognized risk factors for either spontaneous or caregiver-initiated PTB (with the latter resulting from nonreassuring fetal status in utero or a gestational age at which the risk for stillbirth outweighs the risk for prematurity-related complications).558
The existence of various common factors that underlie both stillbirth and PTB is supported by large epidemiologic studies. Gordon and coworkers found that the risk for stillbirth in a subsequent pregnancy was significantly increased if the initial pregnancy was complicated by delivery of a small-for-gestational-age (SGA) preterm neonate (7/1000 live births).559 These findings are consistent with the work of Surkan and col-leagues, who found a similar association between PTB and still-birth (19/1000 live births).560 These concepts are incorporated in the PTB classification system proposed by the Global Alliance on Prematurity and Stillbirth, which suggested that PTBs should be classified using a single system, regardless of whether the infant is alive or dead at birth.10
A reasonable hypothesis is that the health of the placenta is a common denominator in the two conditions.561 The following evidence supports this concept: (1) In animal models of LPS-induced PTB, cytotoxic natural killer cells infiltrate the placenta and are associated with placental cell death, a phenomenon similar to that observed in inflammation-induced fetal demise562,563; (2) medical conditions associated with stillbirth (e.g., severe thrombophilia) and spontaneous PTB share a common constellation of placental histologic features (e.g., thrombosis, infarction, perivillous fibrin deposition, inflamma-tion)428,564-566; (3) failure to modify spiral arteries resulting from defective endovascular trophoblast invasion is associated with stillbirth567; and (4) pregnancy loss rate is significantly higher in patients with confined placental mosaicism than in the cyto-genetically normal cohort.568 As whole exomic and genomic sequencing becomes readily available, a common set of genes that perturb the normal physiology of the placenta and uterus in pregnancies complicated by stillbirth and PTB may be found. Maternal stress521-524 and socioeconomic status569 have been linked to PTB as well as stillbirth. Therefore, residual confound-ing related to social factors linked to stillbirth may be important determinants of the PTB syndrome phenotype.
Intrauterine Growth RestrictionDiagnosis and management of IUGR is one of the most common and challenging problems in modern obstetrics. Confusion in
bulging membranes in the absence of significant uterine contrac-tility or rupture of the membrane, as well as with precipitous labor at term. Cervical insufficiency may be the result of a congenital disorder (e.g., hypoplastic cervix or diethylstilbestrol exposure in utero), surgical trauma (e.g., conization resulting in substantial loss of connective tissue), or traumatic damage of the structural integ-rity of the cervix (e.g., by repeated cervical dilation).546
Most cases of cervical insufficiency reflect not primary cervi-cal disease leading to premature remodeling but other patho-logic processes such as infection, which has been reported in 50% of patients presenting with acute cervical insufficiency,547 or recurrent decidual hemorrhage. The reader is referred to a detailed review of this condition and the role of cervical cerclage in the prevention of PTB.548
Increasing evidence implicates treatment for pre-invasive cervical carcinoma as a risk factor for PTB. Treatments associ-ated with an increased risk for PTB include cold knife coniza-tion (RR = 2.59; 95% CI, 1.80 to 3.72) and large loop excision (RR = 1.70; 95% CI, 1.24 to 2.35).549 There is a greater effect on delivery before 30 weeks, with cold knife conization associated with an RR of 5.33 (95% CI, 1.63 to 17.40).550 Cold knife coniza-tion is also associated with an increase in perinatal mortality.550 These data are important because the large number of women treated means that treatment for pre-invasive cervical disease could become a major contributor to rates of PTB.
The mechanisms by which treatment for pre-invasive disease increases the risk for prematurity are unclear.551 One hypothesis is simply that mechanical disruption of the cervix weakens the cervix. In support of this hypothesis, there is evidence that the depth of the excision correlates with the risk for prematurity, with an estimated 6% increase in risk for each additional 1 mm of tissue excised (OR = 1.06; 95% CI, 1.03 to 1.09).552 More recent data suggest that cervical pre-invasive disease and spontaneous preterm labor may share common risk factors but are not directly linked. These common risk factors may include human papil-loma viruses and other microbial infections in the cervix, or defects in the immune response.551,553 For example, in a study of more than 170,000 women, the increased risk for PTB applied to both women undergoing colposcopy only and women under-going a single excisional treatment, with both having an increased risk compared with women with normal Pap smears.554 A meta-analysis suggested that, when all the relevant studies were com-bined, an increased risk for PTB remains for women having excisional (but not ablative) treatment, compared with an untreated comparison group, although the relative risk is less than when an external comparison group (presumably not sub-jected to the same common risk factors) is used.518
FETAL CONDITIONS
Intrauterine Fetal Demise
A variety of definitions and gestational age cutoff levels are used for reporting stillbirth.555 The ages range from 20 to 28 weeks, and birth weights range from 350 to 1000 g.555 The World Health Organization defines stillbirth as death before expulsion or extraction from the mother of a product of conception, independent of gestational age, showing no signs of life as indi-cated by the absence of breathing, heartbeat, pulsation of the umbilical cord, or movements of voluntary muscles. The U.S. fetal mortality rate declined for the past few decades, reaching an all-time low level of approximately 6 per 1000 live births.556

620 PART 4 Disorders at the Maternal-Fetal Interface
was to assess whether appropriate clinical decisions (i.e., con-tinuation of pregnancy or caregiver-initiated PTB) can be made on the basis of these two tests to avoid neurologic morbidity and stillbirth in preterm infants delivered in the setting of IUGR or intrauterine infection. Although the original studies were encouraging,594 the overall consensus was that neither the NST nor the BPP had good sensitivity for predicting poor neurode-velopment (e.g., cerebral palsy) or infectious complications (e.g., neonatal sepsis).589,592,595 Steroids that are universally rec-ommended to avoid prematurity-related complications might add to the complexity of the clinical decision process because they can alter both fetal heart rate596 and BPP scores.597 There-fore, other tests such as umbilical artery and venous Doppler velocimetry were proposed to be incorporated in the clinical algorithms to indicate the appropriate time of delivery for fetuses at risk.598
The fetus relies on the placenta to ensure adequate oxygen and nutritional transfer from the mother.599 However, in the context of an acutely or chronically impaired oxygen transfer or placental inflammatory dysfunction, abnormal BPP scores and fetal heart rate monitoring patterns may become the first clinical manifestation of such an intrauterine process.600,601 This paradigm is supported by the histopathologic evidence that abnormal fetal heart rates and BPP scores are more frequently present in association with thrombotic vasculopathy, villous endothelial necrosis, placental infarcts, acute umbilical cord vasculitis (funisitis), and histologic chorioamnionitis.589,593,602 Thus, an abnormal fetal heart rate or BPP is not the cause but rather a symptom of an underlying etiology leading to sponta-neous PTB or caregiver-initiated prematurity.
Infection and the Fetal Inflammatory Response SyndromeInfection and the inflammatory response syndrome are defined as a fetal phenotype leading to preterm labor in the new clas-sification system.10 Although there is extensive overlap between these conditions and clinical chorioamnionitis, increasing evi-dence suggests that the fetal inflammatory response syndrome (FIRS), which is almost always associated with fetal infection, is a distinct entity.
FIRS was initially described in pregnancies complicated by preterm labor and pPROM.176,396 It was defined as a fetal plasma concentration of IL-6 greater than 11 pg/mL.396 There are close parallels with the adult systemic inflammatory response syn-drome, with similar peripheral blood leukocyte transcriptomic responses.603 Fetuses with elevated plasma IL-6 concentrations have higher rates of severe neonatal morbidity and a shorter cordocentesis-to-delivery interval than those with IL-6 concen-trations lower than 11 pg/mL.176,335,397,398 The histopathologic landmarks of FIRS are funisitis and chorionic vasculitis.399 The disorder can also be diagnosed by measurement of C-reactive protein concentrations in umbilical cord blood.326 Fetuses with FIRS have more systemic derangements, including hematologic abnormalities (neutrophilia), and a higher median nucleated red blood cell count, than those without elevated IL-6.400 In addition, they have biochemical evidence of fetal stress, as manifested by a fetal plasma ratio of cortisol to DHEAS,401 congenital fetal dermatitis,402 fetal cardiac dysfunction,110 involution of the thymus,403 and abnormalities of the fetal lung125,371,373,398,404-407 and brain.31,71,83,408-433,335 FIRS was initially described in the context of fetal infection, and it is more common in infants with demonstrable microorganisms (such as Mycoplasma and
terminology and lack of uniform diagnostic criteria play an important role. For example, the terms SGA and IUGR are fre-quently used interchangeably, but some suggest that SGA is more appropriate for the newborn570 and that IUGR should refer to the fetus.571 The American College of Obstetricians and Gynecolo-gists favors a sonographic estimated fetal weight of less than the 10th percentile for IUGR.572 There is evidence demonstrating that most of the adverse perinatal outcomes are primarily confined to infants below the fifth or third percentile at birth.573
The underlying etiology of IUGR is important and fre-quently assists with determination of the timing of delivery. Overall fetal growth and development in utero is highly depen-dent on maternal, fetal, or placental factors, and perturbations occurring in any of these compartments can lead to IUGR. A detailed list of various maternal, fetal, and placental conditions linked to IUGR is presented in Chapter 47. However, the risk factors for IUGR, stillbirth, and PTB overlap.
The relationship between IUGR and prematurity is complex. Traditionally, the etiology of IUGR is unknown in approxi-mately 60% of cases.574 In such cases, gestational age may play a considerable role because major complications of prematurity wane considerably after 34 weeks of gestation.575 Therefore, after 34 weeks of gestation, a caregiver-initiated premature delivery of the idiopathic IUGR fetus is indicated to avoid still-birth.575 In other clinical situations (e.g., preeclampsia) mater-nal status is the primary indication for premature delivery of an IUGR fetus. However, several lines of epidemiologic evi-dence suggest a link between spontaneous PTB and fetal growth restriction.559,560,576
Placental association with IUGR is unique because it can be the primary cause (e.g., in the case of mosaicism) of fetal growth restriction.577 However, placental mosaicism does not appear to be associated with an increased risk for spontaneous PTB.568 IUGR is often related to placental abnormalities, including partial abruptions, previa, infarcts, and hematomas. Salafia and colleagues described histologic changes of the placenta in preg-nancies complicated by IUGR.578 Placental lesions were infarc-tion, chronic villitis, hemorrhagic endovasculitis, and placental vascular thromboses. One or more of these lesions were present in 55% of IUGR cases. The spectrum of placental lesions was not uniform across cases. These observations suggest that in some cases of IUGR, decidual hemorrhage is the primary mech-anism of disease, whereas in others, inflammation is responsible for commencement of myometrial contractility or pPROM. In some other IUGR cases, chronic villitis and hemorrhagic endo-vasculitis tend to occur together, implying that the two path-ways can also act in parallel.578-580
Abnormal Fetal Heart Rate or Biophysical ProfileA number of methods for assessing fetal well-being, including the nonstress test (NST) and the biophysical profile (BPP), are routinely used to assess fetal behavioral response to intrauterine stress.581,582 Maturation and maintenance of the structural and functional integrity of the fetal autonomic nervous system is responsible for the changes in fetal heart and biophysical activ-ity (movements, breathing, tonus) observed in utero at various gestational ages.583 Approximately 80% of normally developed fetuses at 28 to 32 weeks demonstrate BPP test scores and fetal heart rate reactivity appreciated as normal (using the criteria of 8/8 or 8/10, and 10-beat, respectively).584,585
Many studies assessed the relationship between prematurity, NST, and BPP.586-593 The purpose of this large body of research

39 Pathogenesis of Spontaneous Preterm Birth 621
However, 50% to 60% of polyhydramnios cases are classified as idiopathic.628 In such situations, reduction of the physio-logic amniotic fluid turnover may result from aberrant expres-sion of members of the transmembrane channel family of aquaporins.629
Perturbation of the amniotic fluid flow resulting from fetal conditions can lead to volume abnormalities such as polyhy-dramnios. Irregular swallowing (from obstruction, or neuro-logic)630,631 and gastrointestinal tract anomalies (tracheoesophageal fistula, congenital diaphragmatic hernia)632,633 may yield hydram-nios in anomalous fetuses. In such clinical scenarios, it is reason-able to propose that excessive myometrial and fetal membrane stretch leads to premature activation of uterine contractility, cer-vical ripening, and dilation. However, clinical studies estimate that PTB occurs in 18.5% of cases with mild (AFI, 25 to 30 cm), 21.8% with moderate (AFI, 30.1 to 35 cm), and 14.3% with severe (AFI > 35.1 cm) polyhydramnios.620 Fetuses with congeni-tal malformations and those of diabetic mothers have a signifi-cantly higher incidence of PTB than fetuses with unexplained polyhydramnios. Thus, the underlying cause of polyhydramnios, rather than the relative excess of amniotic fluid, may determine the occurrence of PTB.620
Multiple PregnanciesPremature birth in multiple pregnancies is seven times greater than in singletons.634 The gestational age at delivery decreases with increasing numbers of fetuses. The average gestational age of delivery with twins is 35 weeks, compared with 30 weeks for quadruplets.635 Overall, 52.2% of multiple births deliver before 37 weeks and 10.7% before 32 weeks.636 These observations implicate extreme mechanical stretching in the pathophysiol-ogy of multiple pregnancy–related PTB.
Sfakianaki and coworkers explored whether the higher uterine volume of multiple gestation could lead to a thinner uterine wall and increased myometrial wall stress, and therefore perhaps explain the tendency for twin gestations to deliver earlier than singletons.637 However, there was no significant ultrasonographic change in the myometrial thickness of the uterine body across pregnancy in women who deliver twins term or preterm, even though there is a substantial increase in the uterine volume in the multiple pregnancies. However, thinning of the lower uterine segment occurred earlier in twin pregnancies destined to deliver before term. Based on these findings, it was proposed that the uterus of the women who deliver preterm twins have a natural limitation of adaptation to the increased length, volume, or weight, and that this limit may be reached at an earlier point in gestation than with twins delivering at term.
Structural and functional differences in the myometrium in twin versus singleton pregnancies might be anticipated, as the myometrium in a multiple pregnancy is exposed to a greater degree of stretch. However, studies comparing myometrium of singleton and multiple pregnancies found no difference in the expression of G-protein (Gsα, which mediates cyclic AMP syn-thesis and relaxation), gap junction proteins (connexin-43, connexin-26), and prostaglandins (EP1, EP3, and EP4).638 Turton and coworkers reported preliminary data that oxytocin aug-ments contractions to a greater extent in myometrium from twin pregnancies than myometrium from singletons in vitro.639 Further studies are required to determine whether this effect is stretch dependent.
There is much heterogeneity among patients with multiple gestations. In some, infection plays a central role.640-642 In others,
Ureaplasma) in their cord blood.510 However, FIRS can be trig-gered by other fetal stressors such as hypoxemia.604
Fetal AnomalyAbout 2% to 3% of all pregnancies are complicated by a fetal anomaly.605,606 The incidence of fetal anomalies in monozygotic twins, compared with those in singletons or dizygotic twins, is increased by 5% to 6%.607 Overall, the antenatal detection rate of fetal structural anomalies is approximately 45%, with a range of 15% to 85%.608 Recognition of fetal structural anomalies varies based on severity or whether the antenatal screening is performed on a low- or a high-risk population.608 Approxi-mately 18% of fetal structural anomalies are associated with chromosomal aneuploidy, but in about 50% of the cases, no cause is identified.609 Congenital heart defects are the most common nonchromosomal anomalies (6.5/1000 births), fol-lowed by limb defects (3.8/1000 births), anomalies of the urinary system (3.1/1000 births), and nervous system defects (2.3/1000 births).610
Most fetal anomalies carry no risk in utero. However, approximately 2% of cases of fetal anomalies, and 25% to 28% of fetuses with aneuploidy, end as stillbirth.611,612 To avoid death in utero, antenatal testing is often recommended for fetuses with anomalies or chromosomal abnormalities appreciated as nonlethal. This approach significantly increases the chance of caregiver-initiated prematurity to facilitate immediate direct care of the newborn.609
Fetal surgery consists of in utero treatment of various con-genital malformations. Implementation of various forms of in utero fetal surgical reparative techniques (e.g., meningomyelo-cele, spina bifida, congenital diaphragmatic hernia) is associated with potential benefits that must be weighed against the increased likelihood of maternal and fetal complications, such as induced PTB, pPROM, infection, and placental abruption.613-615
An increased risk for spontaneous PTB was reported in preg-nancies complicated by multiple anomalies,616 gastroschisis,617 and twin pregnancies with one anomalous fetus.618 The under-lying etiology remains unknown, but anomalies associated with polyhydramnios remain at highest risk. In such cases, excessive myometrial stretch may play a role.236,237
PolyhydramniosDuring human gestation, amniotic fluid is of great importance. In the gestational sac, it protects the fetus from trauma and favors fetal musculoskeletal and lung development. As noted in Chapter 3, the complex nature of the amniotic fluid dynamics reflects contributions from maternal and multiple fetal systems (cutaneous, renal, respiratory, digestive, placental, fetal mem-brane).619 Volume-regulatory mechanisms that are involved during the second half of pregnancy in controlling the amount of amniotic fluid are fetal (urine, lung, swallowing) and amnio-chorion absorption dependent.
Polyhydramnios, or excess amniotic fluid, complicates approximately 1% to 2% of pregnancies and has been associ-ated with a variety of adverse pregnancy outcomes, including PTB.620 Traditionally, polyhydramnios was diagnosed when the AFI of the deepest pool was greater than 8 cm, or when the sum of the AFIs of four quadrants was greater than 24 cm.621 Mater-nal, fetal, and placental conditions associated with polyhydram-nios include maternal diabetes mellitus622 and insipidus,623 rhesus iso-immunization,624 congenital and chromosomal abnormalities,625 multiple gestation,626 and placental tumors.627

622 PART 4 Disorders at the Maternal-Fetal Interface
histologic findings. First, the inflammatory lesions responsible for similar outcomes are characterized by a high degree of het-erogeneity and poor to moderate intraoperator and interopera-tor variability.658 Second, the intricacy and redundancy of biologic processes responsible for cellular and tissue injury might lead to identical pathologic footprints in the context of distinctive triggers. Third, a mild degree of histologic chorio-amnionitis may occur after normal labor at term, without pathologic consequences for the newborn.656,659
Placental AbruptionPlacental abruption represents hemorrhage into the decidua basalis, with complete or partial separation of the placenta from its implantation site.432 The incidence of placental abruption varies from 0.5% to 2%, based on the clinical definition and the criteria applied to characterize its intensity.427,660 The peak rate of abruption is at 24 to 27 weeks of gestation,661 with an occur-rence of PTB as high as 40% (RR = 3.9; 95% CI, 3.5 to 4.4). The high rate of abruption-related perinatal mortality, calculated to be approximately 119 in 1000 births, is heavily confounded by prematurity.662 In addition, the abruption-derived prematurity may have long-term consequences for the surviving infants (e.g., hypoxic ischemic encephalopathy, intraventricular hem-orrhage, bronchopulmonary dysplasia, cerebral palsy).
The classic clinical presentation of abruption manifests as vaginal bleeding, uterine contractions, or abnormal fetal heart rate, or a combination of these. A challenging situation is a concealed abruption, when bleeding is not clinically observable. In this situation, a diagnosis of placental abruption can be established on the basis of fresh macroscopic or histologic examination of the placenta.428 The converging point between abruption and intra-amniotic infection is histologic chorioam-nionitis. The acute lesions of histologic chorioamnionitis are frequently associated with evidence of abruption (hematoma, fibrin deposition, compressed villi, hemosiderin-laden histio-cytes).427 The key histologic finding in placental abruption is hemorrhage in the decidua basalis,432 which is thought to result from pathologic processes damaging the vascular endothelium at the maternal-fetal interface.433 Important to understanding the relationship between placental abruption and PTB is that decidual hemorrhage is a risk factor for preterm contractions and pPROM.663 This could be the consequence of bleeding-induced neutrophil infiltration, activation of MMPs, and pros-taglandin synthesis.664 A comprehensive description of the molecular mechanisms and pathways governing the process of placental abruption–induced PTB was presented earlier.
Placenta PreviaA diagnosis of placenta previa is established when the placenta is inserted into the lower uterine segment and partially or entirely covers the cervix.665 Placenta previa complicates about 0.3% to 0.8% of pregnancies and is one of the most frequent causes of painless bleeding during the second half of gesta-tion.665,666 Risk factors for placenta previa include a history of prior cesarean delivery, uterine surgery, termination of preg-nancy, smoking, advanced maternal age, multiparity, drug abuse, and multiple gestations.665 Antepartum bleeding is a strong predictive risk factor for PTB. In such clinical scenarios, over 50% of the women are delivered before term.667 The median gestational age of the first episode of bleeding is approx-imately 30 weeks, with a median interval of 20 days between the first bleeding episode and delivery.667
cervical shortening and dilation, coupled with acute inflamma-tory lesions of the placenta, may cause stress and decidual hemorrhage-induced PTB.643,644 Assisted reproductive technol-ogy, which is responsible for approximately 15% to 20% of multiple births,645 demonstrates increased rates of perinatal complications—preterm delivery as well as maternal complica-tions, such as preeclampsia, gestational diabetes, placenta previa, and placental abruption.646 It is not possible to separate risks related to assisted reproductive technology from those caused by underlying reproductive pathology, or from the medical condition requiring delivery. These separate mecha-nisms of disease may operate alone or in conjunction with uterine overdistention to activate the components of the common pathway.647
PLACENTAL PATHOLOGIC CONDITIONS
Histologic Chorioamnionitis
A large body of research points toward the choriodecidua as a major site of inflammatory processes linked to PTB.139,514,648,649 In the setting of silent chorioamnionitis, inflammatory cyto-kines and chemokines, released as a result of engagement of TLRs, lead to recruitment of inflammatory cells such as mac-rophages, dendritic cells, and neutrophils, with the final purpose of killing the invading pathogens and halting their spread into the amniotic fluid and to the fetus.438,648 The leukocytes invading the chorion and amnion are maternal in origin.650 The process of inflammatory cell migration into the decidual and fetal membrane tissue is tightly controlled and involves chemotactic factors (chemokines) and cell adhesion molecules (e.g., selec-tins, integrins). The resultant microenvironment is rich in inflammatory mediators that induce tissue damage and result in cytokine and chemokine translocation in the amniotic fluid. Nitric oxide, vascular endothelium growth factor, and angio-poietins seem to be involved in the process of cytokine transfer across fetal membranes.648,651
The inflammatory events occurring in the choriodecidua and fetal membranes are important because they could lead to premature activation of myometrial contractility and PTB through synthesis and release of free radicals, prostaglandins, and metalloprotease.489,652,653 Based on these observations, it is reasonable to assume that the development of a maternal inflammatory response (deciduitis) has some diagnostic poten-tial, even if the process at the time of evaluation is subclinical.
Examination of the placenta has been the first step in patho-logically classifying the wide range of clinical phenotypes linked to PTB (e.g., infection, abruption, hypoxia) and poor neurode-velopmental outcome of the neonate. A significant focus has been on antenatal inflammatory processes.654-656 The proximity of the placenta to the fetus, and their common embryologic origin have facilitated a significant number of studies that linked placental inflammatory lesions to short- and long-term neonatal outcomes such as cerebral palsy.657 The major draw-back is that pathologic examination of the placenta is possible only after birth. As a result, histologic biomarkers are irrelevant during the antenatal period, because they do not allow initia-tion of therapies meant to prevent either PTB or adverse neonatal outcomes. Their overall usefulness is for postnatal counseling and research purposes.
Pathologic examination of the placenta has a further limita-tion: the relatively large subjectivity in interpretation of

39 Pathogenesis of Spontaneous Preterm Birth 623
vascular tumors (chorangioma, teratoma), accessory lobes, and vasa previa.677-679 The vast majority of the literature describing these umbilical cord and placental abnormalities is descriptive. Many cord and placental abnormalities occur in association with complex congenital anomalies, chromosomal aneuploidy, IUGR, hydrops, histologic chorioamnionitis, or abruption, or as complications of multifetal gestation. On the basis of existing knowledge, it is difficult to determine if pathophysiologic events characteristic for each abnormality are independently respon-sible for triggering PTB, or if premature delivery is the result of several overlapping factors.
SummaryPreterm labor, pPROM, and cervical insufficiency are syn-dromes caused by various pathologic processes leading to decidual activation, increased myometrial contractility, cervical remodeling, and membrane rupture. The clinical presentation depends on the nature and timing of the insults affecting the various components of the uterine common pathway of partu-rition. The revised classification system for PTB depends largely on clinical phenotype. Although it requires a paradigm shift to think of preterm parturition as a syndrome, it should facilitate a more accurate comparison of causes of PTB in various popu-lations and with regard to trends over time. The revised clas-sification system has important implications for understanding the biology of preterm parturition, as well as its diagnosis, treat-ment, and prevention.
The complete reference list is available online at www.expertconsult.com.
The pathophysiology of bleeding in placenta previa includes tearing of the placental attachment over the lower uterine segment or the internal os of the cervix. Bleeding is augmented by the inability of the myometrial fibers of the lower uterine segment to contract and thereby close the bleeding vessels. A variety of regulatory molecules play functional roles in con-trolling the process of trophoblast invasion during implanta-tion and placentation.668-670 These include vasoactive and cell surface proteins, proteases, cytokines, chemokines, and growth factors.670 The underlying cause of the excessive myometrial penetration that characterizes placenta accreta, increta, or per-creta accompanying placenta previa remains largely unknown.665 Excessive trophoblast invasion can trigger profound vaginal or intraperitoneal bleeding. Recent data suggest that a lower sys-temic level of free VEGF and a switch of the interstitial extravil-lous trophoblasts to a metastable cell phenotype characterize placenta previa with excessive myometrial invasion and bleed-ing.671 The local hemostatic milieu of the human decidua plays an important role in generating uterine contractions and ampli-fication of bleeding through mechanisms that are similar to those involved in placental abruption.
Other Placental AbnormalitiesUmbilical cord abnormalities, such as single umbilical artery, varices and aneurysms, thrombosis of umbilical vessels, hema-toma, abnormal insertion of the blood vessels, and excessive coiling, knots, and entanglement of the cords, in monoamniotic twin pregnancies can be associated with abnormal fetal growth, development, and behavior that can contribute to premature delivery.672-676 Placental pathologic conditions previously associ-ated with PTB include gestational trophoblastic diseases,

39 Pathogenesis of Spontaneous Preterm Birth 623.e1
REFERENCES
1. Mazaki-Tovi S, Romero R, Kusanovic JP, et al: Recurrent preterm birth, Semin Perinatol 31:142–158, 2007.
2. Joseph KS, Liu S, Rouleau J, et al; for the Fetal and Infant Health Study Group of the Cana-dian Perinatal Surveillance System: Influence of definition based versus pragmatic birth reg-istration on international comparisons of peri-natal and infant mortality: population based retrospective study, BMJ 344:e746, 2012.
3. Kramer MS, Papageorghiou A, Culhane J, et al: Challenges in defining and classifying the preterm birth syndrome, Am J Obstet Gynecol 206(2):108–112, 2012.
4. MacKay DF, Smith GC, Dobbie R, et al: Gesta-tional age at delivery and special educational need: retrospective cohort study of 407,503 schoolchildren, PLoS Med 7(6):e1000289, 2010.
5. Liu L, Johnson HL, Cousens S, et al; Child Health Epidemiology Reference Group of WHO and UNICEF: Global, regional, and national causes of child mortality: an updated systematic analysis for 2010 with time trends since 2000, Lancet 379(9832):2151–2161, 2012.
6. March of Dimes Foundation: March of Dimes White Paper on preterm birth: the global and regional toll, White Plains, NY, 2009, March of Dimes Foundation.
7. Hamilton BE, Martin JA, Ventura SJ: Births: preliminary data for 2010, Natl Vital Stat Rep 60:2, 2011.
8. Msall ME: Developmental vulnerability and resilience in extremely preterm infants, JAMA 292:2399–2401, 2004.
9. Wong AE, Grobman WA: Medically indicated: iatrogenic prematurity, Clin Perinatol 38(3):423–439, 2011.
10. Villar J, Papageorghiou AT, Knight HE, et al: The preterm birth syndrome: a prototype phe-notypic classification, Am J Obstet Gynecol 206(2):119–123, 2012.
11. Parry S, Strauss JF III: Premature rupture of the fetal membranes, N Engl J Med 338:663–670, 1998.
12. Moutquin JM: Classification and heterogene-ity of preterm birth, BJOG 110(Suppl 20):30–33, 2003.
13. Hsu HW, Figueroa JP, Honnebier MB, et al: Power spectrum analysis of myometrial elec-tromyogram and intrauterine pressure changes in the pregnant rhesus monkey in late gesta-tion, Am J Obstet Gynecol 161:467–473, 1989.
14. Norman JE, Morris C, Chalmers J: The effect of changing patterns of obstetric care in Scot-land (1980–2004) on rates of preterm birth and its neonatal consequences. Perinatal Data-base Study, PLoS Med 6(9):e1000153, 2009.
15. Lockwood CJ, Kuczynski E: Risk stratification and pathological mechanisms in preterm delivery, Paediatr Perinat Epidemiol 15(Suppl 2):78–89, 2001.
16. Romero R, Gomez R, Mazor M, et al: The preterm labor syndrome. In Elder MG, Romero R, Lamont RF, editors: Preterm labor, New York, 1997, Churchill Livingstone, pp 29–49.
17. Romero R, Avila C, Sepulveda W, et al: The role of systemic and intrauterine infection in preterm labor. In Fuchs A, Fuchs F, Stubblefield P, editors: Preterm birth: causes, prevention, and management, New York, 1993, McGraw-Hill.
18. Smith R: Parturition, N Engl J Med 356(3):271–283, 2007.
19. Bennett PR, Elder MG, Myatt L: The effects of lipoxygenase metabolites of arachidonic acid on human myometrial contractility, Prosta-glandins 33:837–844, 1987.
20. Bleasdale JE, Johnston JM: Prostaglandins and human parturition: regulation of arachidonic acid mobilization, Rev Perinat Med 5:151, 1985.
21. Calder A: Pharmacological management of the unripe cervix in the human. In Naftolin F, Stubblefield P, editors: Dilatation of the uterine cervix, New York, 1980, Raven Press, pp 317.
22. Calder AA, Greer IA: Pharmacological modu-lation of cervical compliance in the first and second trimesters of pregnancy, Semin Perina-tol 15:162–172, 1991.
23. Carraher R, Hahn DW, Ritchie DM, et al: Involvement of lipoxygenase products in myo-metrial contractions, Prostaglandins 26:23–32, 1983.
24. Challis JRG: Endocrine control of parturition, Physiol Rev 59:863, 1979.
25. Challis JR, Olson D: Parturition. In Knobil E, Neill J, editors: The physiology of reproduction, New York, 1988, Raven Press, p 2177.
26. Ellwood DA, Mitchell MD, Anderson AB, et al: The in vitro production of prostanoids by the human cervix during pregnancy: preliminary observations, Br J Obstet Gynaecol 87:210–214, 1980.
27. Greer I: Cervical ripening. In Drife J, Calder A, editors: Prostaglandins and the uterus, London, 1992, Springer-Verlag, p 191.
28. MacDonald PC, Schultz FM, Duenhoelter JH, et al: Initiation of human parturition. I: Mech-anism of action of arachidonic acid, Obstet Gynecol 44:629–636, 1980.
29. Mitchell MD: The mechanism(s) of human parturition, J Dev Physiol 6:107–118, 1984.
30. Novy MJ, Liggins GC: Role of prostaglandins, prostacyclin, and thromboxanes in the physi-ologic control of the uterus and in parturition, Semin Perinatol 4:45–66, 1980.
31. Rajabi M, Solomon S, Poole AR: Hormonal regulation of interstitial collagenase in the uterine cervix of the pregnant guinea pig, Endocrinology 128:863–871, 1991.
32. Ritchie DM, Hahn DW, McGuire JL: Smooth muscle contraction as a model to study the medi-ator role of endogenous lipoxygenase products of arachidonic acid, Life Sci 34:509–513, 1984.
33. Thorburn GD, Challis JR: Endocrine control of parturition, Physiol Rev 59:863–918, 1979.
34. Wiqvist N, Lindblom B, Wikland M, et al: Prostaglandins and uterine contractility, Acta Obstet Gynecol Scand Suppl 113:23–29, 1983.
35. Romero R, Mazor M, Munoz H, et al: The preterm labor syndrome, Ann N Y Acad Sci 734:414–429, 1994.
36. Ekman G, Forman A, Marsal K, et al: Intravagi-nal versus intracervical application of prosta-glandin E2 in viscous gel for cervical priming and induction of labor at term in patients with an unfavorable cervical state, Am J Obstet Gynecol 147:657–661, 1983.
37. Embrey MP: Induction of abortion by prosta-glandins E1 and E2, Br Med J 1:258–260, 1970.
38. Gordon-Wright AP, Elder MG: Prostaglandin E2 tablets used intravaginally for the induction of labour, Br J Obstet Gynaecol 86:32–36, 1979.
39. Husslein P: Use of prostaglandins for induc-tion of labor, Semin Perinatol 15:173–181, 1991.
40. Husslein P: Prostaglandins for induction of labour. In Drife J, Calder A, editors: Prostaglan-dins and the uterus, London, 1992, Springer-Verlag.
41. Karim SM, Filshie GM: Therapeutic abortion using prostaglandin F2alpha, Lancet 1:157–159, 1970.
42. Macer J, Buchanan D, Yonekura ML: Induction of labor with prostaglandin E2 vaginal sup-positories, Obstet Gynecol 63:664–668, 1984.
43. MacKenzie IZ: Prostaglandins and midtrimes-ter abortion. In Drife J, Calder A, editors: Pros-taglandins and the uterus, London, 1992, Springer-Verlag, p 119.
44. World Health Organization Task Force: Repeated vaginal administration of 15-methyl pgf2 alpha for termination of pregnancy in the 13th to 20th week of gestation, Contraception 16:175, 1977.
45. World Health Organization Task Force: Com-parison of intra-amniotic prostaglandin f2 alpha and hypertonic saline for second trimes-ter abortion, Br Med J 1:1373, 1976.
46. World Health Organization Task Force: Termi-nation of second trimester pregnancy by intra-muscular injection of 16-phenoxy-w-17, 18, 19, 20-tetranor PGE methyl sulfanilamide, Int J Gynaecol Obstet 16:175, 1982.
47. Giri SN, Stabenfeldt GH, Moseley TA, et al: Role of eicosanoids in abortion and its preven-tion by treatment with flunixin meglumine in cows during the first trimester of pregnancy, Zentralbl Veterinarmed A 38:445–459, 1991.
48. Harper MJ, Skarnes RC: Inhibition of abortion and fetal death produced by endotoxin or prostaglandin F2alpha, Prostaglandins 2:295–309, 1972.
49. Keirse MJ: Eicosanoids in human pregnancy and parturition. In Mitchell M, editor: Eico-sanoids in reproduction, Boca Raton, FL, 1990, CRC Press, p 199.
50. Skarnes RC, Harper MJ: Relationship between endotoxin-induced abortion and the synthesis of prostaglandin F, Prostaglandins 1:191–203, 1972.
51. Keirse MJ: Endogenous prostaglandins in human parturition. In Keirse MA, Gravenhorst J, editors: Human parturition, The Hague, Netherlands, 1979, Nijhoff, p 101.
52. Romero R, Emamian M, Quintero R, et al: Amniotic fluid prostaglandin levels and intra-amniotic infections, Lancet 1:1380, 1986.
53. Romero R, Emamian M, Wan M, et al: Increased concentrations of arachidonic acid lipoxygenase metabolites in amniotic fluid during parturition, Obstet Gynecol 70:849–851, 1987.
54. Romero R, Emamian M, Wan M, et al: Prosta-glandin concentrations in amniotic fluid of women with intra-amniotic infection and preterm labor, Am J Obstet Gynecol 157:1461–1467, 1987.
55. Romero R, Wu YK, Mazor M, et al: Amniotic fluid prostaglandin E2 in preterm labor, Pros-taglandins Leukot Essent Fatty Acids 34:141–145, 1988.
56. Romero R, Wu YK, Mazor M, et al: Increased amniotic fluid leukotriene C4 concentration in term human parturition, Am J Obstet Gynecol 159:655–657, 1988.
57. Romero R, Wu YK, Sirtori M, et al: Amniotic fluid concentrations of prostaglandin F2 alpha, 13,14-dihydro-15-keto-prostaglandin F2 alpha

623.e2 PART 4 Disorders at the Maternal-Fetal Interface
(PGFM) and 11-deoxy-13,14-dihydro-15-keto-11, 16-cyclo-prostaglandin E2 (PGEM-LL) in preterm labor, Prostaglandins 37:149–161, 1989.
58. Sellers SM, Mitchell MD, Anderson AB, et al: The relation between the release of prostaglan-dins at amniotomy and the subsequent onset of labour, Br J Obstet Gynaecol 88:1211–1216, 1981.
59. Romero R, Baumann P, Gonzalez R, et al: Amniotic fluid prostanoid concentrations increase early during the course of spontane-ous labor at term, Am J Obstet Gynecol 171:1613–1620, 1994.
60. Brodt-Eppley J, Myatt L: Prostaglandin recep-tors in lower segment myometrium during gestation and labor, Obstet Gynecol 93:89–93, 1999.
61. Matsumoto T, Sagawa N, Yoshida M, et al: The prostaglandin E2 and F2 alpha receptor genes are expressed in human myometrium and are down-regulated during pregnancy, Biochem Biophys Res Commun 238:838–841, 1997.
62. Mohan AR, Loudon JA, Bennett PR: Molecular and biochemical mechanisms of preterm labour, Semin Fetal Neonatal Med 9:437–444, 2004.
63. Allport VC, Pieber D, Slater DM, et al: Human labour is associated with nuclear factor-kappaB activity which mediates cyclo-oxygenase-2 expression and is involved with the “functional progesterone withdrawal”, Mol Hum Reprod 7(6):581–586, 2001.
64. Khanjani S, Kandola MK, Lindstrom TM, et al: NF-κB regulates a cassette of immune/inflammatory genes in human pregnant myo-metrium at term, J Cell Mol Med 15(4):809–824, 2011.
65. Pirianov G, Waddington SN, Lindström TM, et al: The cyclopentenone 15-deoxy-delta 12,14-prostaglandin J(2) delays lipopolysac-charide-induced preterm delivery and reduces mortality in the newborn mouse, Endocrinol-ogy 150(2):699–706, 2009.
66. Arulkumaran S, Kandola MK, Hoffman B, et al: The roles of prostaglandin EP 1 and 3 receptors in the control of human myometrial contractility, J Clin Endocrinol Metab 97(2):489–498, 2012.
67. Cook JL, Zaragoza DB, Sung DH, et al: Expres-sion of myometrial activation and stimulation genes in a mouse model of preterm labor: myometrial activation, stimulation, and preterm labor, Endocrinology 141:1718–1728, 2000.
68. Myatt L, Lye SJ: Expression, localization and function of prostaglandin receptors in myo-metrium, Prostaglandins Leukot Essent Fatty Acids 70:137–148, 2004.
69. Olson DM: The role of prostaglandins in the initiation of parturition, Best Pract Res Clin Obstet Gynaecol 17:717–730, 2003.
70. Fetalvero KM, Zhang P, Shyu M, et al: Prosta-cyclin primes pregnant human myometrium for an enhanced contractile response in partu-rition, J Clin Invest 118(12):3966–3979, 2008.
71. Denison FC, Calder AA, Kelly RW: The action of prostaglandin E2 on the human cervix: Stimulation of interleukin 8 and inhibition of secretory leukocyte protease inhibitor, Am J Obstet Gynecol 180:614–620, 1999.
72. Yoshida M, Sagawa N, Itoh H, et al: Prostaglan-din F(2alpha), cytokines and cyclic mechanical stretch augment matrix metalloproteinase-1 secretion from cultured human uterine
cervical fibroblast cells, Mol Hum Reprod 8:681–687, 2002.
73. Madsen G, Zakar T, Ku CY, et al: Prostaglan-dins differentially modulate progesterone receptor-A and -B expression in human myo-metrial cells: evidence for prostaglandin-induced functional progesterone withdrawal, J Clin Endocrinol Metab 89:1010–1013, 2004.
74. Martinon F, Mayor A, Tschopp J: The inflam-masomes: guardians of the body, Annu Rev Immunol 27:229–265, 2009.
75. Osman I, Young A, Ledingham MA, et al: Leu-kocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term, Mol Hum Reprod 9:41–45, 2003.
76. Thomson AJ, Telfer JF, Young A, et al: Leuko-cytes infiltrate the myometrium during human parturition: further evidence that labour is an inflammatory process, Hum Reprod 14:229–236, 1999.
77. Ledingham MA, Thomson AJ, Jordan F, et al: Cell adhesion molecule expression in the cervix and myometrium during pregnancy and parturition, Obstet Gynecol 97:235–242, 2001.
78. Yuan M, Jordan F, McInnes I, et al: Leukocytes are primed in peripheral blood in preparation for activation during term and preterm labour, Mol Hum Reprod 15:713–724, 2009.
79. Petraglia F, Imperatore A, Challis JR: Neuroen-docrine mechanisms in pregnancy and partu-rition, Endocr Rev 31(6):783–816, 2010.
80. Wadhwa PD, Entringer S, Buss C, et al: The contribution of maternal stress to preterm birth: issues and considerations, Clin Perinatol 38(3):351–384, 2011.
81. McLean M, Bisits A, Davies J, et al: A placental clock controlling the length of human preg-nancy, Nat Med 1(5):460–463, 1995.
82. Germain AM, Carvajal J, Sanchez M, et al: Preterm labor: placental pathology and clinical correlation, Obstet Gynecol 94:284–289, 1999.
83. Arias F, Rodriquez L, Rayne SC, et al: Maternal placental vasculopathy and infection: two dis-tinct subgroups among patients with preterm labor and preterm ruptured membranes, Am J Obstet Gynecol 168:585–591, 1993.
84. Frodl T, O’Keane V: How does the brain deal with cumulative stress? A review with focus on developmental stress, HPA axis function and hippocampal structure in humans, Neurobiol Dis 52:24–37, 2013.
85. Dekker MJ, Tiemeier H, Luijendijk HJ, et al: The effect of common genetic variation in 11β-hydroxysteroid dehydrogenase type 1 on hypothalamic-pituitary-adrenal axis activity and incident depression, J Clin Endocrinol Metab 97(2):E233–E237, 2012.
86. Craddock N, Forty L: Genetics of affective (mood) disorders, Eur J Hum Genet 14:660–668, 2006.
87. Svensson AC, Pawitan Y, Cnattingius S, et al: Familial aggregation of small-for-gestational-age births: the importance of fetal genetic effects, Am J Obstet Gynecol 194:475–479, 2006.
88. Elovitz MA, Mrinalini C: Animal models of preterm birth, Trends Endocrinol Metab 15:479–487, 2004.
89. Fidel PL Jr, Romero R, Wolf N, et al: Systemic and local cytokine profiles in endotoxin-induced preterm parturition in mice, Am J Obstet Gynecol 170:1467–1475, 1994.
90. Gravett MG, Witkin SS, Haluska GJ, et al: An experimental model for intraamniotic
infection and preterm labor in rhesus monkeys, Am J Obstet Gynecol 171:1660–1667, 1994.
91. Hirsch E, Saotome I, Hirsh D: A model of intrauterine infection and preterm delivery in mice, Am J Obstet Gynecol 172:1598–1603, 1995.
92. Kullander S: Fever and parturition: an experi-mental study in rabbits, Acta Obstet Gynecol Scand Suppl 66:77–85, 1977.
93. McDuffie RS Jr, Sherman MP, Gibbs RS: Amni-otic fluid tumor necrosis factor-alpha and interleukin-1 in a rabbit model of bacterially induced preterm pregnancy loss, Am J Obstet Gynecol 167:1583–1588, 1992.
94. McKay DG, Wong TC: The effect of bacterial endotoxin on the placenta of the rat, Am J Pathol 42:357–377, 1963.
95. Romero R, Mazor M, Wu YK, et al: Infection in the pathogenesis of preterm labor, Semin Perinatol 12:262–279, 1988.
96. Romero R, Munoz H, Gomez R, et al: Antibi-otic therapy reduces the rate of infection-induced preterm delivery and perinatal mortality, Am J Obstet Gynecol 170:390, 1994.
97. Takeda Y, Tsuchiya I: Studies on the pathologi-cal changes caused by the injection of the Shwartzman filtrate and the endotoxin into pregnant rabbits, Jap J Exp Med 21:9–16, 1953.
98. Wang H, Hirsch E: Bacterially-induced preterm labor and regulation of prostaglandin-metabolizing enzyme expression in mice: the role of toll-like receptor 4, Biol Reprod 69:1957–1963, 2003.
99. Zahl PA, Bjerknes C: Induction of decidua-placental hemorrhage in mice by the endotox-ins of certain gram-negative bacteria, Proc Soc Exp Biol Med 54:329–332, 1943.
100. Gibbs RS, McDuffie RS Jr, Kunze M, et al: Experimental intrauterine infection with Pre-votella bivia in New Zealand White rabbits, Am J Obstet Gynecol 190:1082–1086, 2004.
101. Gomez R, Ghezzi F, Romero R, et al: Premature labor and intra-amniotic infection: clinical aspects and role of the cytokines in diagnosis and pathophysiology, Clin Perinatol 22:281–342, 1995.
102. Romero R, Espinoza J, Chaiworapongsa T, et al: Infection and prematurity and the role of preventive strategies, Semin Neonatol 7:259–274, 2002.
103. Cassell GH, Davis RO, Waites KB, et al: Isolation of Mycoplasma hominis and Ureaplasma urea-lyticum from amniotic fluid at 16–20 weeks of gestation: potential effect on outcome of preg-nancy, Sex Transm Dis 10:294–302, 1983.
104. Gray DJ, Robinson HB, Malone J, et al: Adverse outcome in pregnancy following amniotic fluid isolation of Ureaplasma urealyticum, Prenat Diagn 12:111–117, 1992.
105. Horowitz S, Mazor M, Romero R, et al: Infec-tion of the amniotic cavity with Ureaplasma urealyticum in the midtrimester of pregnancy, J Reprod Med 40:375–379, 1995.
106. Romero R, Munoz H, Gomez R, et al: Two thirds of spontaneous abortion/fetal deaths after genetic amniocentesis are the result of a pre-existing sub-clinical inflammatory process of the amniotic cavity, Am J Obstet Gynecol 172:S261, 1995.
107. Wenstrom KD, Andrews WW, Hauth JC, et al: Elevated second-trimester amniotic fluid interleukin-6 levels predict preterm delivery, Am J Obstet Gynecol 178:546–550, 1998.
108. Yoon BH, Oh SY, Romero R, et al: An elevated amniotic fluid matrix metalloproteinase-8

39 Pathogenesis of Spontaneous Preterm Birth 623.e3
level at the time of mid-trimester genetic amniocentesis is a risk factor for spontaneous preterm delivery, Am J Obstet Gynecol 185:1162–1167, 2001.
109. Buhimschi IA, Christner R, Buhimschi CS: Proteomic biomarker analysis of amniotic fluid for identification of intra-amniotic inflammation, BJOG 112:173–181, 2005.
110. Buhimschi CS, Bhandari V, Hamar BD, et al: Proteomic profiling of the amniotic fluid to detect inflammation, infection, and neonatal sepsis, PLoS Med 4:e18, 2007.
111. Klein LL, Gibbs RS: Infection and preterm birth, Obstet Gynecol Clin North Am 32:397–410, 2005.
112. Hillier SL, Martins J, Krohn M, et al: A case-control study of chorioamnionic infection and histologic chorioamnionitis in prematurity, N Engl J Med 319:972–978, 1988.
113. Andrews WW, Goldenberg RL, Hauth JC: Preterm labor: emerging role of genital tract infections, Infect Agents Dis 4:196–211, 1995.
114. Pettker CM, Buhimschi IA, Magloire LK, et al: Value of placental microbial evaluation in diagnosing intra-amniotic infection, Obstet Gynecol 109:739–749, 2007.
115. Muller G: Listerellosis and amniotic fluid infection, Geburtshilfe Frauenheilkd 16:496–507, 1956.
116. Mancini N, Carletti S, Ghidoli N, et al: The era of molecular and other non-culture-based methods in diagnosis of sepsis, Clin Microbiol Rev 23:235–251, 2010.
117. Chen K, Pachter L: Bioinformatics for whole-genome shotgun sequencing of microbial communities, PLoS Comput Biol 1:106–112, 2005.
118. Hugenholtz P, Goebel BM, Pace NR: Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity, J Bac-teriol 180:4765–4774, 1998.
119. Vartoukian SR, Palmer RM, Wade WG: Strate-gies for culture of “unculturable” bacteria, FEMS Microbiol Lett 309:1–7, 2010.
120. Lane DJ, Pace B, Olsen GJ, et al: Rapid deter-mination of 16S ribosomal RNA sequences for phylogenetic analyses, Proc Natl Acad Sci U S A 82:6955–6959, 1985.
121. Kim M, Morrison M, Yu Z: Evaluation of dif-ferent partial 16S rRNA gene sequence regions for phylogenetic analysis of microbiomes, J Microbiol Methods 84:81–87, 2011.
122. Han YW, Shen T, Chung P, et al: Uncultivated bacteria as etiologic agents of intra-amniotic inflammation leading to preterm birth, J Clin Microbiol 47:38–47, 2009.
123. DiGiulio DB, Romero R, Amogan HP, et al: Microbial prevalence, diversity and abundance in amniotic fluid during preterm labor: a molecular and culture-based investigation, PLoS One 3:e3056, 2008.
124. Ravel J, Gajer P, Abdo Z, et al: Vaginal micro-biome of reproductive-age women, Proc Natl Acad Sci U S A 108(Suppl 1):4680–4687, 2011.
125. Buhimschi CS, Baumbusch MA, Campbell KH, et al: Insight into innate immunity of the uterine cervix as a host defense mechanism against infection and preterm birth, Expert Rev Obstet Gynecol 4:9–15, 2009.
126. Goldenberg RL, Hauth JC, Andrews WW: Intrauterine infection and preterm delivery, N Engl J Med 342:1500–1507, 2000.
127. Buhimschi CS, Bhandari V, Han YW, et al: Using proteomics in perinatal and neonatal sepsis: hopes and challenges for the future, Curr Opin Infect Dis 22:235–243, 2009.
128. Martin DH: The microbiota of the vagina and its influence on women’s health and disease, Am J Med Sci 343:2–9, 2012.
129. Steel JH, Malatos S, Kennea N, et al: Bacteria and inflammatory cells in fetal membranes do not always cause preterm labor, Pediatr Res 57:404–411, 2005.
130. Kundsin RB, Driscoll SG, Monson RR, et al: Association of Ureaplasma urealyticum in the placenta with perinatal morbidity and mortal-ity, N Engl J Med 310:941–945, 1984.
131. Galask RP, Varner MW, Petzold CR, et al: Bac-terial attachment to the chorioamniotic mem-branes, Am J Obstet Gynecol 148:915–928, 1984.
132. Dombroski RA, Woodard DS, Harper MJ, et al: A rabbit model for bacteria-induced preterm pregnancy loss, Am J Obstet Gynecol 163:1938–1943, 1990.
133. Grigsby PL, Novy MJ, Waldorf KM, et al: Cho-riodecidual inflammation: a harbinger of the preterm labor syndrome, Reprod Sci 17:85–94, 2010.
134. Warren S, Tristram S, Bradbury RS: Maternal and neonatal sepsis caused by Haemophilus influenzae type d, J Med Microbiol 59:370–372, 2010.
135. Han YW, Fardini Y, Chen C, et al: Term still-birth caused by oral Fusobacterium nuclea-tum, Obstet Gynecol 115:442–445, 2010.
136. Cederholm M, Haglund B, Axelsson O: Mater-nal complications following amniocentesis and chorionic villus sampling for prenatal karyo-typing, BJOG 110:392–399, 2003.
137. Janeway C, Travers P, Walport M, et al: Innate immunity. In Janeway C, Travers P, Walport M, et al, editors: Immunobiology, New York, 2005, Garland Science, pp 37–102.
138. Hargreaves DC, Medzhitov R: Innate sensors of microbial infection, J Clin Immunol 25:503–510, 2005.
139. Krikun G, Lockwood CJ, Abrahams VM, et al: Expression of Toll-like receptors in the human decidua, Histol Histopathol 22:847–854, 2007.
140. Youssef RE, Ledingham MA, Bollapragada SS, et al: The role of toll-like receptors (TLR-2 and -4) and triggering receptor expressed on myeloid cells 1 (TREM-1) in human term and preterm labor, Reprod Sci 16(9):843–856, 2009.
141. Miyake K: Endotoxin recognition molecules, Toll-like receptor 4-MD-2, Semin Immunol 16:11–16, 2004.
142. Elovitz MA, Wang Z, Chien EK, et al: A new model for inflammation-induced preterm birth: the role of platelet-activating factor and toll-like receptor-4, Am J Pathol 163:2103–2111, 2003.
143. Lien E, Sellati TJ, Yoshimura A, et al: Toll-like receptor 2 functions as a pattern recognition receptor for diverse bacterial products, J Biol Chem 274:33419–33425, 1999.
144. Dev A, Iyer S, Razani B, et al: NF-κB and innate immunity, Curr Top Microbiol Immunol 349:115–143, 2011.
145. Pulendran B, Kumar P, Cutler CW, et al: Lipo-polysaccharides from distinct pathogens induce different classes of immune responses in vivo, J Immunol 167:5067–5076, 2001.
146. Miyake K: Innate immune sensing of patho-gens and danger signals by cell surface Toll-like receptors, Semin Immunol 19:3–10, 2007.
147. Lotze MT, Zeh HJ, Rubartelli A, et al: The grateful dead: damage-associated molecular pattern molecules and reduction/oxidation regulate immunity, Immunol Rev 220:60–81, 2007.
148. Buhimschi IA, Zhao G, Pettker CM, et al: The receptor for advanced glycation end products (RAGE) system in women with intraamniotic infection and inflammation, Am J Obstet Gynecol 196:181.e1–181.e13, 2007.
149. Buhimschi IA, Buhimschi CS: Proteomics/diagnosis of chorioamnionitis and relation-ships with the fetal exposome, Semin Fetal Neo-natal Med 17;36–45, 2012.
150. Dulay AT, Buhimschi CS, Zhao G, et al: Soluble TLR2 is present in human amniotic fluid and modulates the intraamniotic inflammatory response to infection, J Immunol 182:7244–7253, 2009.
151. Menon R, Velez DR, Morgan N, et al: Genetic regulation of amniotic fluid TNF-alpha and soluble TNF receptor concentrations affected by race and preterm birth, Hum Genet 124:243–253, 2008.
152. Buhimschi CS, Baumbusch MA, Dulay AT, et al: Characterization of RAGE, HMGB1, and S100beta in inflammation-induced preterm birth and fetal tissue injury, Am J Pathol 175:958–975, 2009.
153. Lee SY, Buhimschi IA, Dulay AT, et al: IL-6 trans-signaling system in intra-amniotic inflammation, preterm birth, and preterm pre-mature rupture of the membranes, J Immunol 186(5):3226–3236, 2011.
154. Catalano RD, Lannagan TR, Gorowiec M, et al: Prokineticins: novel mediators of inflamma-tory and contractile pathways at parturition? Mol Hum Reprod 16(5):311–319, 2010.
155. Gorowiec MR, Catalano RD, Norman JE, et al: Prokineticin 1 induces inflammatory response in human myometrium: a potential role in ini-tiating term and preterm parturition, Am J Pathol 179(6):2709–2719, 2011.
156. Romero R, Durum SK, Dinarello CA, et al: Interleukin-1: a signal for the initiation of labor in chorioamnionitis. 33rd Annual Meeting for the Society for Gynecologic Inves-tigation, Toronto, Ontario, 1986.
157. Romero R, Wu YK, Brody DT, et al: Human decidua: a source of interleukin-1, Obstet Gynecol 73:31–34, 1989.
158. Romero R, Durum S, Dinarello CA, et al: Interleukin-1 stimulates prostaglandin biosyn-thesis by human amnion, Prostaglandins 37:13–22, 1989.
159. Romero R, Brody DT, Oyarzun E, et al: Infec-tion and labor: III. Interleukin-1—A signal for the onset of parturition, Am J Obstet Gynecol 160:1117–1123, 1989.
160. Sadowsky DW, Novy MJ, Witkin SS, et al: Dexamethasone or interleukin-10 blocks interleukin-1beta-induced uterine contrac-tions in pregnant rhesus monkeys, Am J Obstet Gynecol 188:252–263, 2003.
161. Romero R, Mazor M, Tartakovsky B: Systemic administration of interleukin-1 induces preterm parturition in mice, Am J Obstet Gynecol 165:969–971, 1991.
162. Romero R, Tartakovsky B: The natural interleukin-1 receptor antagonist prevents interleukin-1-induced preterm delivery in mice, Am J Obstet Gynecol 167:1041–1045, 1992.
163. Casey ML, Cox SM, Beutler B, et al: Cachectin/tumor necrosis factor-alpha formation in human decidua: potential role of cytokines in infection-induced preterm labor, J Clin Invest 83:430–436, 1989.
164. Romero R, Mazor M, Manogue K, et al: Human decidua: a source of cachectin-tumor necrosis factor, Eur J Obstet Gynecol Reprod Biol 41:123–127, 1991.

623.e4 PART 4 Disorders at the Maternal-Fetal Interface
165. Romero R, Manogue KR, Mitchell MD, et al: Infection and labor: IV. Cachectin-tumor necrosis factor in the amniotic fluid of women with intraamniotic infection and preterm labor, Am J Obstet Gynecol 161:336–341, 1989.
166. Fortunato SJ, Menon R, Lombardi SJ: Role of tumor necrosis factor-[alpha] in the premature rupture of membranes and preterm labor pathways, Am J Obstet Gynecol 187:1159–1162, 2002.
167. Watari M, Watari H, DiSanto ME, et al: Pro-inflammatory cytokines induce expression of matrix-metabolizing enzymes in human cervi-cal smooth muscle cells, Am J Pathol 54:1755–1762, 1999.
168. Athayde N, Edwin SS, Romero R, et al: A role for matrix metalloproteinase-9 in spontaneous rupture of the fetal membranes, Am J Obstet Gynecol 79:1248–1253, 1998.
169. Maymon E, Romero R, Pacora P, et al: Evidence of in vivo differential bioavailability of the active forms of matrix metalloproteinases 9 and 2 in parturition, spontaneous rupture of membranes, and intra-amniotic infection, Am J Obstet Gynecol 183:887–894, 2000.
170. Romero R, Chaiworapongsa T, Espinoza J, et al: Fetal plasma MMP-9 concentrations are elevated in preterm premature rupture of the membranes, Am J Obstet Gynecol 187:1125–1130, 2002.
171. Chwalisz K, Benson M, Scholz P, et al: Cervical ripening with the cytokines interleukin 8, interleukin 1 beta and tumour necrosis factor alpha in guinea-pigs, Hum Reprod 9:2173–2181, 1994.
172. Kajikawa S, Kaga N, Futamura Y, et al: Lipotei-choic acid induces preterm delivery in mice, J Pharmacol Toxicol Methods 39:147–154, 1998.
173. Hirsch E, Filipovich Y, Mahendroo M: Signal-ing via the type I IL-1 and TNF receptors is necessary for bacterially induced preterm labor in a murine model, Am J Obstet Gynecol 194:1334–1340, 2006.
174. Lockwood CJ, Arcuri F, Toti P, et al: Tumor necrosis factor-alpha and interleukin-1beta regulate interleukin-8 expression in third tri-mester decidual cells: implications for the genesis of chorioamnionitis, Am J Pathol 169:1294–1302, 2006.
175. Andrews WW, Hauth JC, Goldenberg RL, et al: Amniotic fluid interleukin-6: correlation with upper genital tract microbial colonization and gestational age in women delivered after spon-taneous labor versus indicated delivery, Am J Obstet Gynecol 173:606–612, 1995.
176. Cox SM, King MR, Casey ML, et al: Interleukin-1 beta, -1 alpha, and -6 and pros-taglandins in vaginal/cervical fluids of preg-nant women before and during labor, J Clin Endocrinol Metab 77:805–815, 1993.
177. Gomez R, Romero R, Galasso M, et al: The value of amniotic fluid interleukin-6, white blood cell count, and gram stain in the diagno-sis of microbial invasion of the amniotic cavity in patients at term, Am J Reprod Immunol 32:200–210, 1994.
178. Hillier SL, Witkin SS, Krohn MA, et al: NB, Eschenbach DA. The relationship of amniotic fluid cytokines and preterm delivery, amniotic fluid infection, histologic chorioamnionitis, and chorioamnion infection, Obstet Gynecol 81:941–948, 1993.
179. Messer J, Eyer D, Donato L, et al: Evaluation of interleukin-6 and soluble receptors of tumor necrosis factor for early diagnosis of neonatal infection, J Pediatr 129:574–580, 1996.
180. Romero R, Avila C, Santhanam U, et al: Amni-otic fluid interleukin 6 in preterm labor: asso-ciation with infection, J Clin Invest 85:1392–1400, 1990.
181. Hanna N, Hanna I, Hleb M, et al: Gestational age-dependent expression of IL-10 and its receptor in human placental tissues and iso-lated cytotrophoblasts, J Immunol 164:5721–5728, 2000.
182. Hanna N, Bonifacio L, Weinberger B, et al: Evi-dence for interleukin-10-mediated inhibition of cyclo-oxygenase-2 expression and prosta-glandin production in preterm human pla-centa, Am J Reprod Immunol 55:19–27, 2006.
183. Athayde N, Romero R, Maymon E, et al: Inter-leukin 16 in pregnancy, parturition, rupture of fetal membranes, and microbial invasion of the amniotic cavity, Am J Obstet Gynecol 182:135–141, 2000.
184. Pacora P, Romero R, Maymon E, et al: Partici-pation of the novel cytokine interleukin 18 in the host response to intra-amniotic infection, Am J Obstet Gynecol 183:1138–1143, 2000.
185. Goldenberg RL, Andrews WW, Mercer BM, et al: The preterm prediction study: granulo-cyte colony-stimulating factor and spontane-ous preterm birth. National Institute of Child Health and Human Development Maternal-Fetal Medicine Units Network, Am J Obstet Gynecol 182:625–630, 2000.
186. Saito S, Kato Y, Ishihara Y, et al: Amniotic fluid granulocyte colony-stimulating factor in preterm and term labor, Clin Chim Acta 208:105–109, 1992.
187. Saito S, Kasahara T, Kato Y, et al: Elevation of amniotic fluid interleukin 6 (IL-6), IL-8 and granulocyte colony stimulating factor (G-CSF) in term and preterm parturition, Cytokine 5:81–88, 1993.
188. Chaiworapongsa T, Romero R, Espinoza J, et al: Macrophage migration inhibitory factor in patients with preterm parturition and microbial invasion of the amniotic cavity, J Matern Fetal Neonatal Med 18:405–416, 2005.
189. Ghezzi F, Gomez R, Romero R, et al: Elevated interleukin-8 concentrations in amniotic fluid of mothers whose neonates subsequently develop bronchopulmonary dysplasia, Eur J Obstet Gynecol Reprod Biol 78:5–10, 1998.
190. Romero R, Ceska M, Avila C, et al: Neutrophil attractant/activating peptide-1/interleukin-8 in term and preterm parturition, Am J Obstet Gynecol 165:813–820, 1991.
191. Yoon BH, Romero R, Jun JK, et al: Amniotic fluid cytokines (interleukin-6, tumor necrosis factor-alpha, interleukin-1 beta, and interleukin-8) and the risk for the develop-ment of bronchopulmonary dysplasia, Am J Obstet Gynecol 177:825–830, 1997.
192. Cherouny PH, Pankuch GA, Romero R, et al: Neutrophil attractant/activating peptide-1/interleukin-8: association with histologic cho-rioamnionitis, preterm delivery, and bioactive amniotic fluid leukoattractants, Am J Obstet Gynecol 169:1299–1303, 1993.
193. Gonzalez BE, Ferrer I, Valls C, et al: The value of interleukin-8, interleukin-6 and interleukin-1beta in vaginal wash as predictors of preterm delivery, Gynecol Obstet Invest 59:175–178, 2005.
194. Esplin MS, Romero R, Chaiworapongsa T, et al: Monocyte chemotactic protein-1 is increased in the amniotic fluid of women who deliver preterm in the presence or absence of intra-amniotic infection, J Matern Fetal Neonatal Med 17:365–373, 2005.
195. Keelan JA, Yang J, Romero RJ, et al: Epithelial cell-derived neutrophil-activating peptide-78 is present in fetal membranes and amniotic fluid at increased concentrations with intra-amniotic infection and preterm delivery, Biol Reprod 70:253–259, 2004.
196. Athayde N, Romero R, Maymon E, et al: A role for the novel cytokine RANTES in pregnancy and parturition, Am J Obstet Gynecol 181:989–994, 1999.
197. Hirsch E, Muhle RA, Mussalli GM, et al: Bac-terially induced preterm labor in the mouse does not require maternal interleukin-1 signal-ing, Am J Obstet Gynecol 186:523–530, 2002.
198. Sampson JE, Theve RP, Blatman RN, et al: Fetal origin of amniotic fluid polymorphonuclear leukocytes, Am J Obstet Gynecol 176:77–81, 1997.
199. Prince LR, Whyte MK, Sabroe I, et al: The role of TLRs in neutrophil activation, Curr Opin Pharmacol 11:397–403, 2011.
200. Buhimschi CS, Dulay AT, Abdel-Razeq S, et al: Fetal inflammatory response in women with proteomic biomarkers characteristic of intra-amniotic inflammation and preterm birth, BJOG 116:257–267, 2009.
201. Gonzalez JM, Franzke CW, Yang F, et al: Com-plement activation triggers metalloproteinases release inducing cervical remodeling and preterm birth in mice, Am J Pathol 179(2):838–849, 2011.
202. Gonzalez JM, Dong Z, Romero R, Girardi G: Cervical remodeling/ripening at term and preterm delivery: the same mechanism initi-ated by different mediators and different effec-tor cells, PLoS One 6(11):e26877, 2011.
203. Vaisbuch E, Romero R, Erez O, et al: Activation of the alternative pathway of complement is a feature of pre-term parturition but not of spontaneous labor at term, Am J Reprod Immunol 63(4):318–330, 2010.
204. Krasnow JS, Tollerud DJ, Naus G, et al: Endo-metrial Th2 cytokine expression throughout the menstrual cycle and early pregnancy, Hum Reprod 11:1747–1754, 1996.
205. Lidstrom C, Matthiesen L, Berg G, et al: Cyto-kine secretion patterns of NK cells and macro-phages in early human pregnancy decidua and blood: implications for suppressor macro-phages in decidua, Am J Reprod Immunol 50:444–452, 2003.
206. Ekerfelt C, Lidstrom C, Matthiesen L, et al: Spontaneous secretion of interleukin-4, interleukin-10 and interferon-gamma by first trimester decidual mononuclear cells, Am J Reprod Immunol 47:159–166, 2002.
207. Greig PC, Herbert WN, Robinette BL, et al: Amniotic fluid interleukin-10 concentrations increase through pregnancy and are elevated in patients with preterm labor associated with intrauterine infection, Am J Obstet Gynecol 173:1223–1227, 1995.
208. Moore KW, de Waal MR, Coffman RL, et al: Interleukin-10 and the interleukin-10 receptor, Annu Rev Immunol 19:683–765, 2001.
209. Murray PJ: Understanding and exploiting the endogenous interleukin-10/STAT3-mediated anti-inflammatory response, Curr Opin Phar-macol 6:379–386, 2006.
210. Trinchieri G: Interleukin-10 production by effector T cells: Th1 cells show self control, J Exp Med 204:239–243, 2007.
211. Berg DJ, Kuhn R, Rajewsky K, et al: Interleukin-10 is a central regulator of the response to LPS in murine models of endo-toxic shock and the Shwartzman reaction but

39 Pathogenesis of Spontaneous Preterm Birth 623.e5
not endotoxin tolerance, J Clin Invest 96:2339–2347, 1995.
212. Howard M, Muchamuel T, Andrade S, et al: Interleukin 10 protects mice from lethal endo-toxemia, J Exp Med 177:1205–1208, 1993.
213. Lang R, Rutschman RL, Greaves DR, et al: Autocrine deactivation of macrophages in transgenic mice constitutively overexpressing IL-10 under control of the human CD68 pro-moter, J Immunol 168:3402–3411, 2002.
214. Rodts-Palenik S, Wyatt-Ashmead J, Pang Y, et al: Maternal infection-induced white matter injury is reduced by treatment with interleukin-10, Am J Obstet Gynecol 191:1387–1392, 2004.
215. Chernoff AE, Granowitz EV, Shapiro L, et al: A randomized, controlled trial of IL-10 in humans: inhibition of inflammatory cytokine production and immune responses, J Immunol 154:5492–5499, 1995.
216. Huhn RD, Radwanski E, Gallo J, et al: Pharma-codynamics of subcutaneous recombinant human interleukin-10 in healthy volunteers, Clin Pharmacol Ther 62:171–180, 1997.
217. Pajkrt D, Camoglio L, Tiel-van Buul MC, et al: Attenuation of proinflammatory response by recombinant human IL-10 in human endotox-emia: effect of timing of recombinant human IL-10 administration, J Immunol 158:3971–3977, 1997.
218. Chakraborty A, Blum RA, Mis SM, et al: Phar-macokinetic and adrenal interactions of IL-10 and prednisone in healthy volunteers, J Clin Pharmacol 39:624–635, 1999.
219. Wolfberg AJ, Dammann O, Gressens P: Anti-inflammatory and immunomodulatory strate-gies to protect the perinatal brain, Semin Fetal Neonatal Med 12:296–302, 2007.
220. Robertson SA, Skinner RJ, Care AS: Essential role for IL-10 in resistance to lipopolysaccharide-induced preterm labor in mice, J Immunol 177(7):4888–4896, 2006.
221. Terrone DA, Rinehart BK, Granger JP, et al: Interleukin-10 administration and bacterial endotoxin-induced preterm birth in a rat model, Obstet Gynecol 98:476–480, 2001.
222. Serhan CN, Savill J Resolution of inflamma-tion: the beginning programs the end, Nat Immunol 6(12):1191–1197, 2005.
223. Maldonado-Pérez D, Golightly E, Denison FC, et al: A role for lipoxin A4 as anti-inflammatory and proresolution mediator in human parturi-tion, FASEB J 25(2):569–575, 2011.
224. Rinaldi SF, Hutchinson JL, Rossi AG, et al: Anti-inflammatory mediators as physiological and pharmacological regulators of parturition. Expert Rev Clin Immunol 7(5):675–696, 2011.
225. Menon R, Peltier MR, Eckardt J, et al: Diversity in cytokine response to bacteria associated with preterm birth by fetal membranes, Am J Obstet Gynecol 201:306.e1–306.e6, 2009.
226. Markenson GR, Adams LA, Hoffman DE, et al: Prevalence of Mycoplasma bacteria in amniotic fluid at the time of genetic amniocentesis using the polymerase chain reaction, J Reprod Med 48:775–779, 2003.
227. Berg TG, Philpot KL, Welsh MS, et al: Ureaplasma/Mycoplasma-infected amniotic fluid: pregnancy outcome in treated and non-treated patients, J Perinatol 19:275–277, 1999.
228. Oh KJ, Lee KA, Sohn YK, et al: Intraamniotic infection with genital mycoplasmas exhibits a more intense inflammatory response than intraamniotic infection with other microor-ganisms in patients with preterm premature
rupture of membranes, Am J Obstet Gynecol 203:211.e1–211.e8, 2010.
229. Novy MJ, Duffy L, Axthelm MK, et al: Urea-plasma parvum or Mycoplasma hominis as sole pathogens cause chorioamnionitis, preterm delivery, and fetal pneumonia in rhesus macaques, Reprod Sci 16:56–70, 2009.
230. Buhimschi CS, Buhimschi IA, Malinow AM, et al: Myometrial thickness during human labor and immediately post partum, Am J Obstet Gynecol 188:553–559, 2003.
231. Veille JC, Hosenpud JD, Morton MJ, et al: Cardiac size and function in pregnancy induced hypertension, Am J Obstet Gynecol 150:443–449, 1984.
232. Buhimschi CS, Buhimschi IA, Norwitz ER, et al: Sonographic myometrial thickness pre-dicts the latency interval of women with preterm premature rupture of the membranes and oligohydramnios, Am J Obstet Gynecol 193:762–770, 2005.
233. Fisk NM, Ronderos-Dumit D, Tannirandorn Y, et al: Normal amniotic pressure throughout gestation, Br J Obstet Gynaecol 99(1):18–22, 1992.
234. Garfield RE, Sims S, Daniel EE: Gap junctions: their presence and necessity in myometrium during parturition, Science 198(4320):958–960, 1977.
235. Mendelson CR: Minireview: fetal-maternal hormonal signaling in pregnancy and labor, Mol Endocrinol 23(7):947–954, 2009.
236. Pri-Paz S, Khalek N, Fuchs KM, et al: Maximal amniotic fluid index as a prognostic factor in pregnancies complicated by polyhydramnios, Ultrasound Obstet Gynecol 39(6):648–653, 2012.
237. Moise KJ Jr: Polyhydramnios, Clin Obstet Gynecol 40(2):266–279, 1997.
238. Zakar T, Mesiano S: How does progesterone relax the uterus in pregnancy? N Engl J Med 364(10):972–973, 2011.
239. Renthal NE, Chen CC, Williams KC, et al: miR-200 family and targets, ZEB1 and ZEB2, modu-late uterine quiescence and contractility during pregnancy and labor, Proc Natl Acad Sci U S A 107:20828–20833, 2010.
240. Blanks AM, Shmygol A, Thornton S: Regula-tion of oxytocin receptors and oxytocin recep-tor signaling, Semin Reprod Med 25(1):52–59, 2007.
241. Ou CW, Orsino A, Lye SJ: Expression of connexin-43 and connexin-26 in the rat myo-metrium during pregnancy and labor is dif-ferentially regulated by mechanical and hormonal signals, Endocrinology 138(12):5398–5407, 1997.
242. Terzidou V, Sooranna SR, Kim LU, et al: Mechanical stretch up-regulates the human oxytocin receptor in primary human uterine myocytes, J Clin Endocrinol Metab 90:237–246, 2005.
243. Ou CW, Chen ZQ, Qi S, et al: Increased expres-sion of the rat myometrial oxytocin receptor messenger ribonucleic acid during labor requires both mechanical and hormonal signals, Biol Reprod 59(5):1055–1061, 1998.
244. Hua R, Pease JE, Sooranna SR, et al: Stretch and inflammatory cytokines drive myometrial che-mokine expression via NF-κB activation, Endocrinology 153(1):481–491, 2012.
245. Shynlova O, Tsui P, Dorogin A, et al: Monocyte chemoattractant protein-1 (CCL-2) integrates mechanical and endocrine signals that mediate term and preterm labor, J Immunol 181(2):1470–1479, 2008.
246. Sooranna SR, Engineer N, Loudon JA, et al: The mitogen-activated protein kinase depen-dent expression of prostaglandin H synthase-2 and interleukin-8 messenger ribonucleic acid by myometrial cells: the differential effect of stretch and interleukin-1β, J Clin Endocrinol Metab 90(6):3517–3527, 2005.
247. Sooranna SR, Lee Y, Kim LU, et al: Mechanical stretch activates type 2 cyclooxygenase via acti-vator protein-1 transcription factor in human myometrial cells, Mol Hum Reprod 10(2):109–113, 2004.
248. Monaghan K, Baker SA, Dwyer L, et al: The stretch-dependent potassium channel TREK-1 and its function in murine myometrium, J Physiol 589(Pt 5):1221–1233, 2011.
249. Dalrymple A, Mahn K, Poston L, et al: Mechan-ical stretch regulates TRPC expression and calcium entry in human myometrial smooth muscle cells, Mol Hum Reprod 13(3):171–179, 2007.
250. Tattersall M, Cordeaux Y, Charnock-Jones S, et al: Expression of gastrin-releasing peptide is increased by prolonged stretch of human myo-metrium, and antagonists of its receptor inhibit contractility, J Physiol 590(Pt 9):2081–2093, 2012.
251. Schmidt W: The amniotic fluid compartment: the fetal habitat, Adv Anat Embryol Cell Biol 127:1–100, 1992.
252. Malak TM, Ockleford CD, Bell SC, et al: Con-focal immunofluorescence localization of col-lagen types I, III, IV, V and VI and their ultrastructural organization in term human fetal membranes, Placenta 14:385–406, 1993.
253. Parry-Jones E, Priya S: A study of the elasticity and tension of fetal membranes and of the relation of the area of the gestational sac to the area of the uterine cavity, Br J Obstet Gynaecol 83:205–212, 1976.
254. Millar LK, Stollberg J, DeBuque L, et al: Fetal membrane distention: determination of the intrauterine surface area and distention of the fetal membranes preterm and at term, Am J Obstet Gynecol 182(1 Pt 1):128–134, 2000.
255. Calvin SE, Oyen ML: Microstructure and mechanics of the chorioamnion membrane with an emphasis on fracture properties, Ann N Y Acad Sci 1101:166–185, 2007.
256. Joyce EM, Moore JJ, Sacks MS: Biomechanics of the fetal membrane prior to mechanical failure: review and implications, Eur J Obstet Gynecol Reprod Biol 144(Suppl 1):S121–S127, 2009.
257. Maradny EE, Kanayama N, Halim A, et al: Stretching of fetal membranes increases the concentration of interleukin-8 and collagenase activity, Am J Obstet Gynecol 174:843–849, 1996.
258. Mohan AR, Sooranna SR, Lindstrom TM, et al: The effect of mechanical stretch on cyclooxy-genase type 2 expression and activator protein-1 and nuclear factor-kappaB activity in human amnion cells, Endocrinology 148(4):1850–1857, 2007.
259. Maehara K, Kanayama N, Maradny EE, et al: Mechanical stretching induces interleukin-8 gene expression in fetal membranes: a possible role for the initiation of human parturition, Eur J Obstet Gynecol Reprod Biol 70(2):191–196, 1996.
260. Kendal-Wright CE, Hubbard D, Bryant-Greenwood GD: Chronic stretching of amni-otic epithelial cells increases pre-B cell colony-enhancing factor (PBEF/visfatin)

623.e6 PART 4 Disorders at the Maternal-Fetal Interface
expression and protects them from apoptosis, Placenta 29(3):255–265, 2008.
261. Nemeth E, Millar LK, Bryant-Greenwood G: Fetal membrane distention: II. Differentially expressed genes regulated by acute distention in vitro, Am J Obstet Gynecol 182(1 Pt 1):60–67, 2000.
262. Myatt L, Sun K: Role of fetal membranes in signaling of fetal maturation and parturition, Int J Dev Biol 54(2–3):545–553, 2010.
263. Han YM, Romero R, Kim YM, et al: Surfactant protein-A mRNA expression by human fetal membranes is increased in histological chorio-amnionitis but not in spontaneous labour at term, J Pathol 211(4):489–496, 2007.
264. Cheng YH, Nicholson RC, King B, et al: Gluco-corticoid stimulation of corticotropin-releasing hormone gene expression requires a cyclic adenosine 3′,5′-monophosphate regula-tory element in human primary placental cyto-trophoblast cells, J Clin Endocrinol Metab 85(5):1937–1945, 2000.
265. Challis JR, Lye SJ, Gibb W, et al: Understanding preterm labor, Ann N Y Acad Sci 943:225–234, 2001.
266. Wolfe CD, Patel SP, Linton EA, et al: Plasma corticotrophin-releasing factor (CRF) in abnormal pregnancy, Br J Obstet Gynaecol 95:1003–1006, 1988.
267. Florio P, Zatelli MC, Reis FM, et al: Corticotro-pin releasing hormone: a diagnostic marker for behavioral and reproductive disorders? Front Biosci 12:551–560, 2007.
268. Jones SA, Challis JR: Effects of corticotropin-releasing hormone and adrenocorticotropin on prostaglandin output by human placenta and fetal membranes, Gynecol Obstet Invest 29:165–168, 1990.
269. Jones SA, Challis JR: Steroid, corticotrophin-releasing hormone, ACTH and prostaglandin interactions in the amnion and placenta of early pregnancy in man, J Endocrinol 125:153–159, 1990.
270. Economopoulos P, Sun M, Purgina B, et al: Glucocorticoids stimulate prostaglandin H synthase type-2 (PGHS-2) in the fibroblast cells in human amnion cultures, Mol Cell Endocrinol 117(2):141–147, 1996.
271. Patel FA, Clifton VL, Chwalisz K, et al: Steroid regulation of prostaglandin dehydrogenase activity and expression in human term pla-centa and chorio-decidua in relation to labor, J Clin Endocrinol Metab 84:291–299, 1999.
272. Sun K, Ma R, Cui X, et al: Glucocorticoids induce cytosolic phospholipase A2 and prosta-glandin H synthase type 2 but not microsomal prostaglandin E synthase (PGES) and cytosolic PGES expression in cultured primary human amnion cells, J Clin Endocrinol Metab 88(11):5564–5571, 2003.
273. Coussons-Read ME, Lobel M, Carey JC, et al: The occurrence of preterm delivery is linked to pregnancy-specific distress and elevated inflammatory markers across gestation, Brain Behav Immun 2012.
274. Torricelli M, Novembri R, Bloise E, et al: Changes in placental CRH, urocortins, and CRH-receptor mRNA expression associated with preterm delivery and chorioamnionitis, J Clin Endocrinol Metab 96(2):534–540, 2011.
275. Yang R, You X, Tang X, et al: Corticotropin-releasing hormone inhibits progesterone production in cultured human placental tro-phoblasts, J Mol Endocrinol 37(3):533–540, 2006.
276. You X, Yang R, Tang X, et al: Corticotropin-releasing hormone stimulates estrogen biosyn-thesis in cultured human placental trophoblasts, Biol Reprod 74(6):1067–1072, 2006.
277. Stjernholm-Vladic Y, Wang H, Stygar D, et al: Differential regulation of the progesterone receptor A and B in the human uterine cervix at parturition, Gynecol Endocrinol 18:41–46, 2004.
278. Oh SY, Kim CJ, Park I, et al: Progesterone receptor isoform (A/B) ratio of human fetal membranes increases during term parturition, Am J Obstet Gynecol 193:1156–1160, 2005.
279. Merlino A, Welsh T, Erdonmez T, et al: Nuclear progesterone receptor expression in the human fetal membranes and decidua at term before and after labor, Reprod Sci 16:357–363, 2009.
280. Cong B, Zhang L, Gao L, Ni X Reduced expres-sion of CRH receptor type 1 in upper segment human myometrium during labour, Reprod Biol Endocrinol 7:43, 2009.
281. Zhang LM, Wang YK, Hui N, et al: Corticotropin-releasing hormone acts on CRH-R1 to inhibit the spontaneous contractil-ity of non-labouring human myometrium at term, Life Sci 83:620–624, 2008.
282. Lockwood CJ, Radunovic N, Nastic D, et al: Corticotropin-releasing hormone and related pituitary-adrenal axis hormones in fetal and maternal blood during the second half of preg-nancy, J Perinat Med 24:243–251, 1996.
283. Zakar T, Hirst JJ, Mijovic JE, et al: Glucocorti-coids stimulate the expression of prostaglandin endoperoxide H synthase-2 in amnion cells, Endocrinology 136:1610–1619, 1995.
284. Challis JR: CRH, a placental clock and preterm labour, Nat Med 1:416, 1995.
285. Langlois D, Li JY, Saez JM: Development and function of the human fetal adrenal cortex, J Pediatr Endocrinol 15(Suppl 5):1311–1322, 2002.
286. Economides DL, Nicolaides KH, Linton EA, et al: Plasma cortisol and adrenocorticotropin in appropriate and small for gestational age fetuses, Fetal Ther 3:158–164, 1988.
287. Amiel-Tison C, Cabrol D, Denver R, et al: Fetal adaptation to stress: Part I. Acceleration of fetal maturation and earlier birth triggered by pla-cental insufficiency in humans, Early Hum Dev 78:15–27, 2004.
288. Gravett MG, Hitti J, Hess DL, et al: Intrauterine infection and preterm delivery: evidence for activation of the fetal hypothalamic-pituitary-adrenal axis, Am J Obstet Gynecol 182(6):1404–1413, 2000.
289. Turan OM, Turan S, Funai EF, et al: Fetal adrenal gland volume: a novel method to iden-tify women at risk for impending preterm birth, Obstet Gynecol 109(4):855–862, 2007.
290. Power ML, Schulkin J: Functions of corticotropin-releasing hormone in anthro-poid primates: from brain to placenta, Am J Hum Biol 18(4):431–447, 2006.
291. Parker CR Jr, Stankovic AM, Goland RS: Corticotropin-releasing hormone stimulates steroidogenesis in cultured human adrenal cells, Mol Cell Endocrinol 155:19–25, 1999.
292. Buhimschi CS, Turan OM, Funai EF, et al: Fetal adrenal gland volume and cortisol/dehydroepiandrosterone sulfate ratio in inflammation-associated preterm birth, Obstet Gynecol 111(3):715–722, 2008.
293. Richter ON, Kubler K, Schmolling J, et al: Oxy-tocin receptor gene expression of estrogen-stimulated human myometrium in
extracorporeally perfused non-pregnant uteri, Mol Hum Reprod 10:339–346, 2004.
294. Di WL, Lachelin GC, McGarrigle HH, et al: Oestriol and oestradiol increase cell to cell communication and connexin43 protein expression in human myometrium, Mol Hum Reprod 7:671–679, 2001.
295. Mesiano S, Chan EC, Fitter JT, et al: Progester-one withdrawal and estrogen activation in human parturition are coordinated by proges-terone receptor A expression in the myome-trium, J Clin Endocrinol Metab 87:2924–2930, 2002.
296. Anderson L, Martin W, Higgins C, et al: Inhibi-tion of human myometrial contractility by progesterone does not operate via potassium channels, Reprod Sci 16:1052–1061, 2009.
297. Challis JRG, Lye SJ: Parturition. In Knobil E, Neil J, editors: The physiology of reproduction, New York, 1994, Raven Press, pp 985–1031.
298. Smith R, Smith JI, Shen X, et al: Patterns of plasma corticotropin-releasing hormone, pro-gesterone, estradiol, and estriol change and the onset of human labor, J Clin Endocrinol Metab 94(6):2066–2074, 2009.
299. Kulier R, Gülmezoglu AM, Hofmeyr GJ, et al: Medical methods for first trimester abortion, Cochrane Database Syst Rev (2):CD002855, 2004.
300. Hapangama D, Neilson JP: Mifepristone for induction of labour, Cochrane Database Syst Rev (3):CD002865, 2009.
301. Rode L, Langhoff-Roos J, Andersson C, et al: Systematic review of progesterone for the pre-vention of preterm birth in singleton pregnan-cies, Acta Obstet Gynecol Scand 88(11):1180–1189, 2009.
302. Mesiano S, Wang Y, Norwitz ER: Progesterone receptors in the human pregnancy uterus: do they hold the key to birth timing? Reprod Sci 18(1):6–19, 2011.
303. Condon JC, Hardy DB, Kovaric K, et al: Up-regulation of the progesterone receptor (PR)-C isoform in laboring myometrium by activation of nuclear factor-kappaB may con-tribute to the onset of labor through inhibition of PR function, Mol Endocrinol 20(4):764–775, 2006.
304. Tan H, Yi L, Rote NS, et al: Progesterone receptor-A and -B have opposite effects on proinflammatory gene expression in human myometrial cells: implications for progester-one actions in human pregnancy and parturi-tion, J Clin Endocrinol Metab 97(5):E719–E730, 2012.
305. Clayton D, McKeigue PM: Epidemiological methods for studying genes and environmen-tal factors in complex diseases, Lancet 358:1356–1360, 2001.
306. Tiret L: Gene-environment interaction: a central concept in multifactorial diseases, Proc Nutr Soc 61:457–463, 2002.
307. Macones GA, Parry S, Elkousy M, et al: A poly-morphism in the promoter region of TNF and bacterial vaginosis: preliminary evidence of gene-environment interaction in the etiology of spontaneous preterm birth, Am J Obstet Gynecol 190:1504–1508, 2004.
308. Roberts AK, Monzon-Bordonaba F, Van Deerlin PG, et al: Association of polymor-phism within the promoter of the tumor necrosis factor alpha gene with increased risk of preterm premature rupture of the fetal membranes, Am J Obstet Gynecol 180:1297–1302, 1999.

39 Pathogenesis of Spontaneous Preterm Birth 623.e7
309. Romero R, Chaiworapongsa T, Kuivaniemi H, et al: Bacterial vaginosis, the inflammatory response and the risk of preterm birth: a role for genetic epidemiology in the prevention of preterm birth, Am J Obstet Gynecol 190:1509–1519, 2004.
310. Young RC: Myocytes, myometrium, and uterine contractions, Ann N Y Acad Sci 1101:72–84, 2007.
311. Wray S: Uterine contraction and physiological mechanisms of modulation, Am J Physiol 264(1 Pt 1):C1–C18, 1993.
312. Cole WC, Garfield RE, Kirkaldy JS: Gap junc-tions and direct intercellular communication between rat uterine smooth muscle cells, Am J Physiol 249:C20–C31, 1985.
313. Garfield RE, Sims SM, Kannan MS, et al: Pos-sible role of gap junctions in activation of myo-metrium during parturition, Am J Physiol 235:C168–C179, 1978.
314. Garfield RE, Hayashi RH: Appearance of gap junctions in the myometrium of women during labor, Am J Obstet Gynecol 140:254–260, 1981.
315. Garfield RE, Puri CP, Csapo AI: Endocrine, structural, and functional changes in the uterus during premature labor, Am J Obstet Gynecol 142:21–27, 1982.
316. Balducci J, Risek B, Gilula NB, et al: Gap junc-tion formation in human myometrium: a key to preterm labor? Am J Obstet Gynecol 168:1609–1615, 1993.
317. Chow L, Lye SJ: Expression of the gap junction protein connexin-43 is increased in the human myometrium toward term and with the onset of labor, Am J Obstet Gynecol 170:788–795, 1994.
318. Lefebvre DL, Piersanti M, Bai XH, et al: Myo-metrial transcriptional regulation of the gap junction gene, connexin-43, Reprod Fertil Dev 7:603–611, 1995.
319. Orsino A, Taylor CV, Lye SJ: Connexin-26 and connexin-43 are differentially expressed and regulated in the rat myometrium throughout late pregnancy and with the onset of labor, Endocrinology 137:1545–1553, 1996.
320. Lye SJ: The initiation and inhibition of labour: towards a molecular understanding, Semin Reprod Endocrinol 12:284–294, 1994.
321. Lye SJ, Mitchell J, Nashman N, et al: Role of mechanical signals in the onset of term and preterm labor, Front Horm Res 27:165–178, 2001.
322. Lye SJ, Nicholson BJ, Mascarenhas M, et al: Increased expression of connexin-43 in the rat myometrium during labor is associated with an increase in the plasma estrogen: progester-one ratio, Endocrinology 132:2380–2386, 1993.
323. Petrocelli T, Lye SJ: Regulation of transcripts encoding the myometrial gap junction protein, connexin-43, by estrogen and progesterone, Endocrinology 133:284–290, 1993.
324. Lye S, Tsui P, Dorogin A, et al: Myometrial pro-gramming: a new concept underlying the maintenance of pregnancy and the initiation of labor. In VIIth International Conference on the Extracellular Matrix of the Female Reproduc-tive Tract and Simpson Symposia, Centre for Reproductive Biology, University of Edin-burgh, 2004.
325. O’Brien M, Morrison JJ, Smith TJ: Upregula-tion of PSCDBP, TLR2, TWIST1, FLJ35382, EDNRB, and RGS12 gene expression in human myometrium at labor, Reprod Sci 15(4):382–393, 2008.
326. Bollapragada S, Youssef R, Jordan F, et al: Term labor is associated with a core inflammatory response in human fetal membranes, myome-trium, and cervix, Am J Obstet Gynecol 200(1):104.e1–104.e11, 2009.
327. Aslanidi OV, Colman MA, Stott J, et al: 3D virtual human atria: a computational platform for studying clinical atrial fibrillation, Prog Biophys Mol Biol 107(1):156–168, 2011.
328. Aslanidi O, Atia J, Benson AP, et al: Towards a computational reconstruction of the electro-dynamics of premature and full term human labour, Prog Biophys Mol Biol 107(1):183–192, 2011.
329. Friedman E: The graphic analysis of labor, Am J Obstet Gynecol 68:1568–1575, 1954.
330. Iams JD, Cebrik D, Lynch C, et al: The rate of cervical change and the phenotype of sponta-neous preterm birth, Am J Obstet Gynecol 205:130.e1–130.e6, 2011.
331. Park IY, Kwon JY, Kwon JY, et al: Usefulness of cervical volume by three-dimensional ultra-sound in identifying the risk for preterm birth, Ultrasound Med Biol 37:1039–1045, 2011.
332. Weiner CP, Lee KY, Buhimschi CS, et al: Pro-teomic biomarkers that predict the clinical success of rescue cerclage, Am J Obstet Gynecol 192:710–718, 2005.
333. Iams JD, Goldenberg RL, Meis PJ, et al: The length of the cervix and the risk of spontane-ous premature delivery. National Institute of Child Health and Human Development Mater-nal Fetal Medicine Unit Network, N Engl J Med 334:567–572, 1996.
334. Garfield RE, Saade G, Buhimschi C, et al: Control and assessment of the uterus and cervix during pregnancy and labour, Hum Reprod Update 4:673–695, 1998.
335. Buhimschi I, Ali M, Jain V, et al: Differential regulation of nitric oxide in the rat uterus and cervix during pregnancy and labour, Hum Reprod 11:1755–1766, 1996.
336. Schlembach D, Mackay L, Shi L, et al: Cervical ripening and insufficiency: from biochemical and molecular studies to in vivo clinical exami-nation, Eur J Obstet Gynecol Reprod Biol 144(Suppl 1):S70–S76, 2009.
337. Word RA, Li XH, Hnat M, et al: Dynamics of cervical remodeling during pregnancy and parturition: mechanisms and current concepts, Semin Reprod Med 25:69–79, 2007.
338. Buhimschi CS, Sora N, Zhao G, et al: Genetic background affects the biomechanical behav-ior of the postpartum mouse cervix, Am J Obstet Gynecol 200:434.e1–434.e7, 2009.
339. Leppert PC: Anatomy and physiology of cervi-cal ripening, Clin Obstet Gynecol 38:267–279, 1995.
340. Yu S, Leppert PC: The collagenous tissues of the cervix during labor and delivery. In Leppert PC, Woessner JK Jr, editor: The extracellular matrix of the uterus, cervix and fetal mem-branes: synthesis, degradation and hormonal regulation, Ithaca, NY, 1991, Perinatology Press, pp 68–76.
341. Rorie DK, Newton M: Histologic and chemical studies of the smooth muscle in the human cervix and uterus, Am J Obstet Gynecol 99:466–469, 1967.
342. Myers KM, Paskaleva AP, House M, et al: Mechanical and biochemical properties of human cervical tissue, Acta Biomater 4:104–116, 2008.
343. Danforth DN: The fibrous nature of the human cervix, and its relation to the isthmic
segment in gravid and nongravid uteri, Am J Obstet Gynecol 53:541, 1947.
344. Ludmir J, Sehdev HM: Anatomy and physiol-ogy of the uterine cervix, Clinic Obstet Gynecol 43:433–439, 2000.
345. Kleissl HP, van der Rest M, Naftolin F, et al: Collagen changes in the human uterine cervix at parturition, Am J Obstet Gynecol 130:748–753, 1978.
346. Osmers R, Rath W, Adelmann-Grill BC, et al: Origin of cervical collagenase during parturi-tion, Am J Obstet Gynecol 166:1455–1460, 1992.
347. Winkler M, Rath W: Changes in the cervical extracellular matrix during pregnancy and parturition, J Perinat Med 27:45–60, 1999.
348. Malmström E, Sennström M, Holmberg A, et al: The importance of fibroblasts in remod-elling of the human uterine cervix during preg-nancy and parturition, Mol Hum Reprod 13:333–341, 2007.
349. Imada K, Sato T, Hashizume K, et al: An anti-progesterone, onapristone, enhances the gene expression of promatrix metalloproteinase 3/prostromelysin-1 in the uterine cervix of preg-nant rabbit, Biol Pharm Bull 25:1223–1227, 2002.
350. Lyons CA, Beharry KD, Nishihara KC, et al: Regulation of matrix metalloproteinases (type IV collagenases) and their inhibitors in the virgin, timed pregnant, and postpartum rat uterus and cervix by prostaglandin E(2)-cyclic adenosine monophosphate, Am J Obstet Gynecol 187:202–208, 2002.
351. Buhimschi IA, Dussably L, Buhimschi CS, et al: Physical and biomechanical characteristics of rat cervical ripening are not consistent with increased collagenase activity, Am J Obstet Gynecol 191:1695–1704, 2004.
352. Choi SJ, Jung KL, Oh SY, et al: Cervicovaginal matrix metalloproteinase-9 and cervical ripen-ing in human term parturition, Eur J Obstet Gynecol Reprod Biol 142:43–47, 2009.
353. Stygar D, Wang H, Vladic YS, et al: Increased level of matrix metalloproteinases 2 and 9 in the ripening process of the human cervix, Biol Reprod 67:889–894, 2002.
354. Becher N, Hein M, Danielsen CC, et al: Matrix metalloproteinases and their inhibitors in the cervical mucus plug at term of pregnancy, Am J Obstet Gynecol 191:1232–1239, 2004.
355. Woods JR Jr, Plessinger MA, Miller RK: Vita-mins C and E: missing links in preventing preterm premature rupture of membranes? Am J Obstet Gynecol 185: 5–10, 2001.
356. Nusgens BV, Humbert P, Rougier A, et al: Topi-cally applied vitamin C enhances the mRNA level of collagens I and III, their processing enzymes and tissue inhibitor of matrix metal-loproteinase 1 in the human dermis, J Invest Dermatol 116:853–859, 2001.
357. Leppert PC, Kokenyesi R, Klemenich CA, et al: Further evidence of a decorin-collagen interac-tion in the disruption of cervical collagen fibers during rat gestation, Am J Obstet Gynecol 182:805–811, 2000.
358. Rechberger T, Woessner JF Jr: Collagenase, its inhibitors, and decorin in the lower uterine segment in pregnant women, Am J Obstet Gynecol 168:1598–1603, 1993.
359. Kokenyesi R, Woessner JF Jr: Effects of hor-monal perturbations on the small dermatan sulfate proteoglycan and mechanical proper-ties of the uterine cervix of late pregnant rats, Connect Tissue Res 26:199–205, 1991.

623.e8 PART 4 Disorders at the Maternal-Fetal Interface
360. Iwahashi M, Muragaki Y, Ooshima A, et al: Decreased type I collagen expression in human uterine cervix during pregnancy, J Clin Endo-crinol Metab 88:2231–2235, 2003.
361. Maul H, Olson G, Fittkow CT, et al: Cervical light-induced fluorescence in humans decreases throughout gestation and before delivery: preliminary observations, Am J Obstet Gynecol 188:537–541, 2003.
362. Kokenyesi R, Woessner JF Jr: Relationship between dilatation of the rat uterine cervix and a small dermatan sulfate proteoglycan, Biol Reprod 42(1):87–97, 1990.
363. Sfakianaki AK, Buhimschi IA, Ravishankar V, et al: Relationships of maternal serum levels of vascular endothelial growth factor and tensile strength properties of the cervix in a rat model of chronic hypoxia, Am J Obstet Gynecol 198:223.e1–233.e7, 2008.
364. El Maradny E, Kanayama N, Kobayashi H, et al: The role of hyaluronic acid as a mediator and regulator of cervical ripening, Hum Reprod 12:1080–1088, 1997.
365. Uldbjerg N, Ekman G, Malmstrom A, et al: Ripening of the human uterine cervix related to changes in collagen, glycosaminoglycans, and collagenolytic activity, Am J Obstet Gynecol 147:662–666, 1983.
366. Byers BD, Bytautiene E, Costantine MM, et al: Hyaluronidase modifies the biomechanical properties of the rat cervix and shortens the duration of labor independent of myometrial contractility, Am J Obstet Gynecol 203(6):596.e1–596.e5, 2010.
367. Leppert PC, Yu SY, Keller S, et al: Decreased elastic fibers and desmosine content in incom-petent cervix, Am J Obstet Gynecol 157:1134–1139, 1987.
368. Hwang JJ, Macinga D, Rorke EA: Relaxin mod-ulates human cervical stromal cell activity, J Clin Endocrinol Metab 81:3379–3384, 1996.
369. Parry LJ, McGuane JT, Gehring HM, et al: Mechanisms of relaxin action in the reproduc-tive tract: studies in the relaxin-deficient (Rlx-/-) mouse, Ann N Y Acad Sci 1041:91–103, 2005.
370. Kokenyesi R, Armstrong LC, Agah A, et al: Thrombospondin 2 deficiency in pregnant mice results in premature softening of the uterine cervix, Biol Reprod 70:385–390, 2004.
371. Akins ML, Luby-Phelps K, Bank RA, et al: Cer-vical softening during pregnancy: regulated changes in collagen cross-linking and compo-sition of matricellular proteins in the mouse, Biol Reprod 84:1053–1062, 2011.
372. Allaire AD, D’Andrea N, Truong P, et al: Cervi-cal stroma apoptosis in pregnancy, Obstet Gynecol 97:399–403, 2001.
373. Lee HY, Sherwood OD: The effects of blocking the actions of estrogen and progesterone on the rates of proliferation and apoptosis of cer-vical epithelial and stromal cells during the second half of pregnancy in rats, Biol Reprod 73:790–797, 2005.
374. Zhang X, Jeyakumar M, Petukhov S, et al: A nuclear receptor corepressor modulates tran-scriptional activity of antagonist-occupied steroid hormone receptor, Mol Endocrinol 12:513–524, 1998.
375. Rajabi MR, Solomon S, Poole AR: Biochemical evidence of collagenase-mediated collagenoly-sis as a mechanism of cervical dilatation at parturition in the guinea pig, Biol Reprod 45:764–772, 1991.
376. De Luca A, Santra M, Baldi A, et al: Decorin-induced growth suppression is associated with
up-regulation of p21, an inhibitor of cyclin-dependent kinases, J Biol Chem 271:18961–18965, 1996.
377. Rechberger T, Abramson SR, Woessner JF Jr: Onapristone and prostaglandin E2 induction of delivery in the rat in late pregnancy: a model for the analysis of cervical softening, Am J Obstet Gynecol 175:719–723, 1996.
378. Uldbjerg N, Ekman G, Malmström A, et al: Biochemical changes in human cervical con-nective tissue after local application of prosta-glandin E2, Gynecol Obstet Invest 15:291–299, 1983.
379. Vladic-Stjernholm Y, Vladic T, Blesson CS, et al: Prostaglandin treatment is associated with a withdrawal of progesterone and andro-gen at the receptor level in the uterine cervix, Reprod Biol Endocrinol 7:116, 2009.
380. Norman J: Antiprogesterones, Br J Hosp Med 45:372–375, 1991.
381. Fonseca EB, Celik E, Parra M, et al: Progester-one and the risk of preterm birth among women with a short cervix, N Engl J Med 357:462–469, 2007.
382. DeFranco EA, O’Brien JM, Adair CD, et al: Vaginal progesterone is associated with a decrease in risk for early preterm birth and improved neonatal outcome in women with a short cervix: a secondary analysis from a ran-domized, double-blind, placebo-controlled trial, Ultrasound Obstet Gynecol 30:697–705, 2007.
383. Stys SJ, Clewell WH, Meschia G: Changes in cervical compliance at parturition indepen-dent of uterine activity, Am J Obstet Gynecol 130(4):414–418, 1978.
384. Larsen B, Hwang J: Progesterone interactions with the cervix: translational implications for term and preterm birth, Infect Dis Obstet Gynecol 2011:353297, 2011.
385. Yellon SM, Oshiro BT, Chhaya TY, et al: Remodeling of the cervix and parturition in mice lacking the progesterone receptor B isoform, Biol Reprod 85(3):498–502, 2011.
386. Mahendroo MS, Porter A, Russell DW, et al: The parturition defect in steroid 5alpha-reductase type 1 knockout mice is due to impaired cervical ripening, Mol Endocrinol 13(6):981–992, 1999.
387. Tanaka K, Nakamura T, Ikeya H, et al: Hyaluro-nate depolymerization activity induced by pro-gesterone in cultured fibroblasts derived from human uterine cervix, FEBS Lett 347:95–98, 1994.
388. Mendelson CR: Minireview: fetal-maternal hormonal signaling in pregnancy and labor, Mol Endocrinol 23(7):947–954, 2009.
389. George SS, Matthews J, Jeyaseelan L, et al: Cer-vical ripening induces labour through interleu-kin 1 beta, Aust N Z J Obstet Gynaecol 33:285–286, 1993.
390. Sakamoto Y, Moran P, Bulmer JN, et al: Mac-rophages and not granulocytes are involved in cervical ripening, J Reprod Immunol 66:161–173, 2005.
391. Young A, Thomson AJ, Ledingham M, et al: Immunolocalization of proinflammatory cyto-kines in myometrium, cervix, and fetal mem-branes during human parturition at term, Biol Reprod 66(2):445–449, 2002.
392. Timmons BC, Fairhurst AM, Mahendroo MS: Temporal changes in myeloid cells in the cervix during pregnancy and parturition, J Immunol 182(5):2700–2707, 2009.
393. Timmons BC, Mahendroo MS: Timing of neutrophil activation and expression of
proinflammatory markers do not support a role for neutrophils in cervical ripening in the mouse, Biol Reprod 74(2):236–245, 2006.
394. Ito A, Hiro D, Ojima Y, et al: Spontaneous pro-duction of interleukin-1-like factors from pregnant rabbit uterine cervix, Am J Obstet Gynecol 159:261–265, 1988.
395. Sakamoto Y, Moran P, Searle RF, et al: Interleukin-8 is involved in cervical dilatation but not in prelabour cervical ripening, Clin Exp Immunol 138(1):151–157, 2004.
396. Winkler M, Fischer DC, Ruck P, et al: Cytokine concentrations and expression of adhesion molecules in the lower uterine segment during parturition at term: relation to cervical dilata-tion and duration of labor, Z Geburtshilfe Neo-natol 202:172–175, 1998.
397. Ekerhovd E, Weijdegard B, Brannstrom M, et al: Nitric oxide induced cervical ripening in the human: involvement of cyclic guanosine monophosphate, prostaglandin F(2 alpha), and prostaglandin E(2), Am J Obstet Gynecol 186:745–750, 2002.
398. Ledingham MA, Denison FC, Kelly RW, et al: Nitric oxide donors stimulate prostaglandin F(2alpha) and inhibit thromboxane B(2) pro-duction in the human cervix during the first trimester of pregnancy, Mol Hum Reprod 5(10):973–982, 1999.
399. Hassan SS, Romero R, Tarca AL, et al: The tran-scriptome of cervical ripening in human preg-nancy before the onset of labor at term: identification of novel molecular functions involved in this process, J Matern Fetal Neona-tal Med 22(12):1183–1193, 2009.
400. Hassan SS, Romero R, Tarca AL, et al: The molecular basis for sonographic cervical short-ening at term: identification of differentially expressed genes and the epithelial-mesenchymal transition as a function of cervi-cal length, Am J Obstet Gynecol 203(5):472.e1–472.e14, 2010.
401. Akins ML, Luby-Phelps K, Mahendroo M: Second harmonic generation imaging as a potential tool for staging pregnancy and pre-dicting preterm birth, J Biomed Opt 15(2):026020, 2010.
402. Törnblom SA, Klimaviciute A, Byström B, et al: Non-infected preterm parturition is related to increased concentrations of IL-6, IL-8 and MCP-1 in human cervix, Reprod Biol Endocri-nol 3:39, 2005.
403. Rizzo G, Capponi A, Vlachopoulou A, et al: Ultrasonographic assessment of the uterine cervix and interleukin-8 concentrations in cer-vical secretions predict intrauterine infection in patients with preterm labor and intact membranes, Ultrasound Obstet Gynecol 12(2):86–92, 1998.
404. Hassan S, Romero R, Hendler I, et al: A sono-graphic short cervix as the only clinical mani-festation of intra-amniotic infection, J Perinat Med 34(1):13–19, 2006.
405. Kiefer DG, Keeler SM, Rust OA, et al: Is midtri-mester short cervix a sign of intraamniotic inflammation? Am J Obstet Gynecol 200(4):374.e1–374.e5, 2009.
406. Gravett MG, Thomas A, Schneider KA, et al: Proteomic analysis of cervical-vaginal fluid: identification of novel biomarkers for detec-tion of intra-amniotic infection, J Proteome Res 6:89–96, 2007.
407. Buhimschi IA, Buhimschi CS, Weiner CP, et al: Proteomic but not enzyme-linked immuno-sorbent assay technology detects amniotic fluid monomeric calgranulins from their

39 Pathogenesis of Spontaneous Preterm Birth 623.e9
complexed calprotectin form, Clin Diagn Lab Immunol 12(7):837–844, 2005.
408. Havelock JC, Keller P, Muleba N, et al: Human myometrial gene expression before and during parturition, Biol Reprod 72(3):707–719, 2005.
409. Holt R, Timmons BC, Akgul Y, et al: The molecular mechanisms of cervical ripening differ between term and preterm birth, Endo-crinology 152(3):1036–1046, 2011.
410. Gonzalez JM, Xu H, Chai J, et al: Preterm and term cervical ripening in CD1 mice (Mus mus-culus): similar or divergent molecular mecha-nisms? Biol Reprod 81(6):1226–1232, 2009.
411. Fox NS, Rebarber A, Klauser CK, et al: Predic-tion of spontaneous preterm birth in asymp-tomatic twin pregnancies using the change in cervical length over time, Am J Obstet Gynecol 202(2):155.e1–155.e4, 2010.
412. Levine RU, Berkowitz KM: Conservative management and pregnancy outcome in diethylstilbestrol-exposed women with and without gross genital tract abnormalities, Am J Obstet Gynecol 169(5):1125–1129, 1993.
413. Barclay CG, Brennand JE, Kelly RW, et al: Interleukin-8 production by the human cervix, Am J Obstet Gynecol 169:625–632, 1993.
414. Dubicke A, Akerud A, Sennstrom M, et al: Dif-ferent secretion patterns of matrix metallopro-teinases and IL-8 and effect of corticotropin-releasing hormone in preterm and term cervical fibroblasts, Mol Hum Reprod 14(11):641–647, 2008.
415. Calder AA: Prostaglandins and biological control of cervical function, Aust N Z J Obstet Gynaecol 34:347–351, 1994.
416. Oxlund BS, Ørtoft G, Brüel A, et al: Cervical collagen and biomechanical strength in non-pregnant women with a history of cervical insufficiency, Reprod Biol Endocrinol 8:92, 2010.
417. Yamada KM: Cell surface interactions with extracellular materials, Annu Rev Biochem 52:761–799, 1983.
418. Feinberg RF, Kliman HJ, Lockwood CJ: Is oncofetal fibronectin a trophoblast glue for human implantation? Am J Pathol 138(3):537–543, 1991.
419. Kayisli UA, Korgun ET, Akkoyunlu G, et al: Expression of integrin alpha5 and integrin beta4 and their extracellular ligands fibronec-tin and laminin in human decidua during early pregnancy and its sex steroid-mediated regula-tion, Acta Histochem 107(3):173–185, 2005.
420. Zhu BC, Laine RA: Polylactosamine glycosyl-ation on human fetal placental fibronectin weakens the binding affinity of fibronectin to gelatin, J Biol Chem 260(7):4041–4045, 1985.
421. Sibille Y, Lwebuga-Mukasa JS, Polomski L, et al: An in vitro model for polymorphonuclear-leukocyte-induced injury to an extracellular matrix: relative contribution of oxidants and elastase to fibronectin release from amnionic membranes, Am Rev Respir Dis 134(1):134–140, 1986.
422. Iams JD, Casal D, McGregor JA, et al: Fetal fibronectin improves the accuracy of diagnosis of preterm labor, Am J Obstet Gynecol 173:141–145, 1995.
423. Nageotte MP, Casal D, Senyei AE: Fetal fibro-nectin in patients at increased risk for prema-ture birth, Am J Obstet Gynecol 170:20–25, 1994.
424. Lockwood CJ, Senyei AE, Dische MR, et al: Fetal fibronectin in cervical and vaginal secre-tions as a predictor of preterm delivery, N Engl J Med 325:669–674, 1991.
425. Norman JE: Preterm labour. Cervical function and prematurity, Best Pract Res Clin Obstet Gynaecol 21:791–806, 2007.
426. Chandiramani M, Di Renzo GC, Gottschalk E, et al: Fetal fibronectin as a predictor of spon-taneous preterm birth: a European perspective, J Matern Fetal Neonatal Med 24(2):330–336, 2011.
427. Elsasser DA, Ananth CV, Prasad V, et al: New Jersey-Placental Abruption Study Investiga-tors. Diagnosis of placental abruption: rela-tionship between clinical and histopathological findings, Eur J Obstet Gynecol Reprod Biol 148(2):125–130, 2010.
428. Salafia CM, López-Zeno JA, Sherer DM, et al: Histologic evidence of old intrauterine bleed-ing is more frequent in prematurity, Am J Obstet Gynecol 173(4):1065–1070, 1995.
429. Nath CA, Ananth CV, Smulian JC, et al: Histo-logic evidence of inflammation and risk of placental abruption, Am J Obstet Gynecol 197(3):319.e1–319.e6, 2007.
430. Lockwood CJ: Pregnancy-associated changes in the hemostatic system, Clin Obstet Gynecol 49:836–843, 2006.
431. Benirschke K: Anatomical relationship between fetus and mother, Ann N Y Acad Sci 731:9–20, 1994.
432. Hall DR: Abruptio placentae and disseminated intravascular coagulopathy, Semin Perinatol 33:189–195, 2009.
433. Dommisse J, Tiltman AJ: Placental bed biop-sies in placental abruption, Br J Obstet Gynae-col 99:651–654, 2009.
434. Buhimschi CS, Rosenberg VA, Dulay AT, et al: Multidimensional system biology: genetic markers and proteomic biomarkers of adverse pregnancy outcome in preterm birth, Am J Perinatol 25(3):175–187, 2008.
435. Shankar R, Johnson MP, Williamson NA, et al: Molecular markers of preterm labor in the choriodecidua, Reprod Sci 17(3):297–310, 2010.
436. Levi M, van der Poll T. Inflammation and coagulation, Crit Care Med 38(2 Suppl):S26–S34, 2010.
437. Lockwood CJ, Murk W, Kayisli UA, et al: Pro-gestin and thrombin regulate tissue factor expression in human term decidual cells, J Clin Endocrinol Metab 94(6):2164–2170, 2009.
438. Arcuri F, Toti P, Buchwalder L, Casciaro A, et al: Mechanisms of leukocyte accumulation and activation in chorioamnionitis: interleukin 1 beta and tumor necrosis factor alpha enhance colony stimulating factor 2 expression in term decidua, Reprod Sci 16(5):453–461, 2009.
439. Lockwood CJ, Paidas M, Murk WK, et al: Involvement of human decidual cell-expressed tissue factor in uterine hemostasis and abrup-tion, Thromb Res 124(5):516–520, 2009.
440. Rosen T, Kuczynski E, O’Neill LM, et al: Plasma levels of thrombin-antithrombin complexes predict preterm premature rupture of the fetal membranes, J Matern Fetal Med 10:297–300, 2001.
441. Rosen T, Schatz F, Kuczynski E, et al: Thrombin-enhanced matrix metalloproteinase-1 expres-sion: a mechanism linking placental abruption with premature rupture of the membranes, J Matern Fetal Neonatal Med 11(1):11–17, 2002.
442. Krikun G, Lockwood CJ, Paidas MJ: Tissue factor and the endometrium: from physiology to pathology, Thromb Res 124:393–396, 2009.
443. Taylor FB Jr, Chang A, Hinshaw LB, et al: A model for thrombin protection against endo-toxin, Thromb Res 36(2):177–185, 1984.
444. Lockwood CJ, Krikun G, Papp C, et al: The role of progestationally regulated stromal cell tissue factor and type-1 plasminogen activator inhib-itor (PAI-1) in endometrial hemostasis and menstruation, Ann N Y Acad Sci 734:57–79, 1994.
445. Elovitz MA, Saunders T, Ascher-Landsberg J, et al: Effects of thrombin on myometrial con-tractions in vitro and in vivo, Am J Obstet Gynecol 183:799–804, 2000.
446. Elovitz MA, Ascher-Landsberg J, Saunders T, et al: The mechanisms underlying the stimula-tory effects of thrombin on myometrial smooth muscle, Am J Obstet Gynecol 183:674–681, 2000.
447. O’Sullivan CJ, Allen NM, O’Loughlin AJ, et al: Thrombin and PAR1-activating peptide: effects on human uterine contractility in vitro, Am J Obstet Gynecol 190:1098–1105, 2004.
448. Giudice LC: Genes associated with embryonic attachment and implantation and the role of progesterone, J Reprod Med 44:165–171, 1999.
449. Critchley HO, Saunders PT: Hormone receptor dynamics in a receptive human endometrium, Reprod Sci 16:191–199, 2009.
450. Rauk PN, Chiao JP: Interleukin-1 stimulates human uterine prostaglandin production through induction of cyclooxygenase-2 expres-sion, Am J Reprod Immunol 43:152–159, 2000.
451. Snegovskikh VV, Schatz F, Arcuri F, et al: Intra-amniotic infection upregulates decidual cell vascular endothelial growth factor (VEGF) and neuropilin-1 and -2 expression: implications for infection-related preterm birth, Reprod Sci 16:767–780, 2009.
452. Weiss A, Goldman S, Shalev E: The matrix metalloproteinases (MMPS) in the decidua and fetal membranes, Front Biosci 12:649–659, 2007.
453. Christiaens I, Zaragoza DB, Guilbert L, et al: Inflammatory processes in preterm and term parturition, J Reprod Immunol 79(1):50–57, 2008.
454. Oner C, Schatz F, Kizilay G, et al: Progestin-inflammatory cytokine interactions affect matrix metalloproteinase-1 and -3 expression in term decidual cells: implications for treat-ment of chorioamnionitis-induced preterm delivery, J Clin Endocrinol Metab 93(1):252–259, 2008.
455. Lockwood CJ, Matta P, Krikun G, et al: Regula-tion of monocyte chemoattractant protein-1 expression by tumor necrosis factor-alpha and interleukin-1beta in first trimester human decidual cells: implications for preeclampsia, Am J Pathol 168:445–452, 2006.
456. Schatz F, Lockwood CJ: Progestin regulation of plasminogen activator inhibitor type 1 in primary cultures of endometrial stromal and decidual cells, J Clin Endocrinol Metab 77:621–625, 1993.
457. Norwitz ER, Snegovskikh V, Schatz F, et al: Pro-gestin inhibits and thrombin stimulates the plasminogen activator/inhibitor system in term decidual stromal cells: implications for parturition, Am J Obstet Gynecol 196:382.e1–382.e8, 2007.
458. Krikun G, Schatz F, Mackman N, et al: Regula-tion of tissue factor gene expression in human endometrium by transcription factors Sp1 and Sp3, Mol Endocrinol 14:393–400, 2000.
459. Lockwood CJ, Krikun G, Hausknecht VA, et al: Matrix metalloproteinase and matrix metallo-proteinase inhibitor expression in endometrial stromal cells during progestin-initiated decid-ualization and menstruation-related progestin

623.e10 PART 4 Disorders at the Maternal-Fetal Interface
withdrawal, Endocrinology 139:4607–4613, 1998.
460. Schatz F, Krikun G, Runic R, et al: Implications of decidualization-associated protease expres-sion in implantation and menstruation, Semin Reprod Endocrinol 17:3–12, 1999.
460a. Lockwood CJ, Kayisli UA, Stocco C, et al: Abruption-induced preterm delivery is associ-ated with thrombin-mediated functional pro-gesterone withdrawal in decidual cells, Am J Pathol 181(6):2138–2148, 2012.
461. Rasmussen S, Irgens LM, Dalaker K: The effect on the likelihood of further pregnancy of pla-cental abruption and the rate of its recurrence, Br J Obstet Gynaecol 104:1292–1295, 1997.
462. Peltier MR: Thromboembolic diseases in fami-lies of women with placental abruption, Epide-miology 20:733–737, 2009.
463. Rasmussen S, Irgens LM: Occurrence of pla-cental abruption in relatives, BJOG 116:693–699, 2009.
464. Jaaskelainen E, Keski-Nisula L, Toivonen S, et al: Polymorphism of the interleukin 1 recep-tor antagonist (IL1Ra) gene and placental abruption, J Reprod Immunol 79:58–62, 2008.
465. Ananth CV, Peltier MR, De Marco C, et al: Associations between 2 polymorphisms in the methylenetetrahydrofolate reductase gene and placental abruption, Am J Obstet Gynecol 197:385.e1–385.e7, 2007.
466. Zdoukopoulos N, Zintzaras E: Genetic risk factors for placental abruption: a HuGE review and meta-analysis, Epidemiology 19:309–323, 2008.
467. Said JM, Higgins JR, Moses EK, et al: Inherited thrombophilia polymorphisms and pregnancy outcomes in nulliparous women, Obstet Gynecol 115:5–13, 2010.
468. Silver RM, Zhao Y, Spong CY, et al: Prothrom-bin gene G20210A mutation and obstetric complications, Obstet Gynecol 115:14–20, 2010.
469. ACOG Committee on Practice Bulletins—Obstetrics: ACOG practice bulletin no. 80: premature rupture of membranes—clinical management guidelines for obstetrician-gynecologists. (Reaffirmed 2010), Obstet Gynecol 109:1007–1019, 2007.
470. Maymon E, Romero R, Pacora P, et al: Evidence for the participation of interstitial collagenase (matrix metalloproteinase 1) in preterm pre-mature rupture of membranes, Am J Obstet Gynecol 183:914–920, 2000.
471. Maymon E, Romero R, Pacora P, et al: Human neutrophil collagenase (matrix metallopro-teinase 8) in parturition, premature rupture of the membranes, and intrauterine infection, Am J Obstet Gynecol 183(1):94–99, 2000.
472. Vadillo-Ortega F, Hernandez A, Gonzalez-Avila G, et al: Increased matrix metalloprotein-ase activity and reduced tissue inhibitor of metalloproteinases-1 levels in amniotic fluids from pregnancies complicated by premature rupture of membranes, Am J Obstet Gynecol 174:1371–1376, 1996.
473. Maymon E, Romero R, Pacora P, et al: A role for the 72 kDa gelatinase (MMP-2) and its inhibitor (TIMP-2) in human parturition, pre-mature rupture of membranes and intraamni-otic infection, J Perinat Med 29:308–316, 2001.
474. Murphy G, Reynolds JJ, Bretz U, et al: Partial purification of collagenase and gelatinase from human polymorphonuclear leucocytes: analy-sis of their actions on soluble and insoluble collagens, Biochem J 203(1):209–221, 1982.
475. Steffensen B, Chen Z, Pal S, et al: Fragmenta-tion of fibronectin by inherent autolytic and matrix metalloproteinase activities, Matrix Biol 30(1):34–42, 2011.
476. McLaren J, Malak TM, Bell SC: Structural characteristics of term human fetal mem-branes prior to labour: identification of an area of altered morphology overlying the cervix, Hum Reprod 14(1):237–241, 1999.
477. Malak TM, Bell SC: Structural characteristics of term human fetal membranes: a novel zone of extreme morphological alteration within the rupture site, Br J Obstet Gynaecol 101(5):375–386, 1994.
478. Malak TM, Mulholland G, Bell SC: Morpho-metric characteristics of the decidua, cytotro-phoblast, and connective tissue of the prelabor ruptured fetal membranes, Ann N Y Acad Sci 734:430–432, 1994.
479. Bell SC, Pringle JH, Taylor DJ, et al: Alterna-tively spliced tenascin-C mRNA isoforms in human fetal membranes, Mol Hum Reprod 5(11):1066–1076, 1999.
480. Lappas M, Riley C, Lim R, et al: MAPK and AP-1 proteins are increased in term pre-labour fetal membranes overlying the cervix: regula-tion of enzymes involved in the degradation of fetal membranes, Placenta 32:1016–1025, 2011.
481. Lappas M, Odumetse TL, Riley C, et al: Pre-labour fetal membranes overlying the cervix display alterations in inflammation and NF-kappaB signalling pathways, Placenta 29:995–1002, 2008.
482. Mackie EJ, Halfter W, Liverani D: Induction of tenascin in healing wounds, J Cell Biol 107(6 Pt 2):2757–2767, 1988.
483. George RB, Kalich J, Yonish B, et al: Apoptosis in the chorion of fetal membranes in preterm premature rupture of membranes, Am J Peri-natol 25(1):29–32, 2008.
484. Fortunato SJ, Menon R: IL-1 beta is a better inducer of apoptosis in human fetal mem-branes than IL-6, Placenta 24(10):922–928, 2003.
485. Kumar D, Fung W, Moore RM, et al: Proin-flammatory cytokines found in amniotic fluid induce collagen remodeling, apoptosis, and biophysical weakening of cultured human fetal membranes, Biol Reprod 74(1):29–34, 2006.
486. Moore RM, Novak JB, Kumar D, et al: Alpha-lipoic acid inhibits tumor necrosis factor-induced remodeling and weakening of human fetal membranes, Biol Reprod 80(4):781–787, 2009.
487. Shook LL, Buhimschi CS, Dulay AT, et al: Fetuin-mediated aggregation of amniotic fluid proteins into calcifying nanoparticles and preterm premature rupture of membranes (PPROM), Am J Obstet Gynecol 206(1):S7–S8, 2012.
488. Chai M, Barker G, Menon R, et al: Increased oxidative stress in human fetal membranes overlying the cervix from term non-labouring and post labour deliveries, Placenta 33:604–610, 2012.
489. Buhimschi IA, Kramer WB, Buhimschi CS, et al: Reduction-oxidation (redox) state regula-tion of matrix metalloproteinase activity in human fetal membranes, Am J Obstet Gynecol 182:458–464, 2000.
490. Williams MA, Mittendorf R, Lieberman E, et al: Adverse infant outcomes associated with first-trimester vaginal bleeding, Obstet Gynecol 78:14–18, 1991.
491. Harger JH, Hsing AW, Tuomala RE, et al: Risk factors for preterm premature rupture of fetal
membranes: a multicenter case-control study, Am J Obstet Gynecol 163:130–137, 1990.
492. Mackenzie AP, Schatz F, Krikun G, et al: Mech-anisms of abruption-induced premature rupture of the fetal membranes: thrombin enhanced decidual matrix metalloproteinase-3 (stromelysin-1) expression, Am J Obstet Gynecol 191:1996–2001, 2004.
493. Stephenson CD, Lockwood CJ, Ma Y, et al: Thrombin-dependent regulation of matrix metalloproteinase (MMP)-9 levels in human fetal membranes, J Matern Fetal Neonatal Med 18:17–22, 2005.
494. Kumar D, Schatz F, Moore RM, et al: The effects of thrombin and cytokines upon the biomechanics and remodeling of isolated amnion membrane, in vitro, Placenta 32(3):206–213, 2011.
495. Moore RM, Schatz F, Kumar D, et al: Alpha-lipoic acid inhibits thrombin-induced fetal membrane weakening in vitro, Placenta 31(10):886–892, 2010.
496. Habib NA, Daltveit AK, Bergsjø P, et al: Mater-nal HIV status and pregnancy outcomes in northeastern Tanzania: a registry-based study, BJOG 115(5):616–624, 2008.
497. Lopez M, Figueras F, Hernandez S, et al: Asso-ciation of HIV infection with spontaneous and iatrogenic preterm delivery: effect of HAART, AIDS 26(1):37–43, 2012.
498. Townsend C, Schulte J, Thorne C, et al: Pediat-ric Spectrum of HIV Disease Consortium, the European Collaborative Study and the National Study of HIV in Pregnancy and Childhood. Antiretroviral therapy and preterm delivery-a pooled analysis of data from the United States and Europe, BJOG 117(11):1399–1410, 2010.
499. Poespoprodjo JR, Fobia W, Kenangalem E, et al: Adverse pregnancy outcomes in an area where multidrug-resistant Plasmodium vivax and Plasmodium falciparum infections are endemic, Clin Infect Dis 46(9):1374–1381, 2008.
500. Barros FC, Bhutta ZA, Batra M, et al; GAPPS Review Group: Global report on preterm birth and stillbirth (3 of 7): evidence for effective-ness of interventions, BMC Pregnancy Child-birth 10(Suppl 1):S3, 2010.
501. Romero R, Oyarzun E, Mazor M, et al: Meta-analysis of the relationship between asymp-tomatic bacteriuria and preterm delivery/low birth weight, Obstet Gynecol 73(4):576–582, 1989.
502. Wei PL, Keller JJ, Liang HH, et al: Acute appen-dicitis and adverse pregnancy outcomes: a nationwide population-based study, J Gastro-intest Surg. 16(6):1204–1211, 2012.
503. Pierce M, Kurinczuk JJ, Spark P, et al; UKOSS: Perinatal outcomes after maternal 2009/H1N1 infection: national cohort study, BMJ 342:d3214, 2011.
504. Xiong X, Buekens P, Fraser WD, et al: Peri-odontal disease and adverse pregnancy out-comes: a systematic review, BJOG 113(2):135–143, 2006.
505. Larsen JW, Goldkrand JW, Hanson TM, et al: Intrauterine infection on an obstetric service, Obstet Gynecol 43:838–843, 1974.
506. Onderdonk AB, Hecht JL, McElrath TF, et al: Colonization of second-trimester placenta parenchyma, Am J Obstet Gynecol 199:52.e1–52.e10, 2008.
507. Watts DH, Krohn MA, Hillier SL, et al: The association of occult amniotic fluid infection with gestational age and neonatal outcome

39 Pathogenesis of Spontaneous Preterm Birth 623.e11
among women in preterm labor, Obstet Gynecol 79:351–357, 1992.
508. Smulian JC, Shen-Schwarz S, Vintzileos AM, et al: Clinical chorio-amnionitis and histologic placental inflammation, Obstet Gynecol 94:1000–1005, 1999.
509. Heller DS, Rimpel LH, Skurnick JH: Does his-tologic chorioamnionitis correspond to clini-cal chorioamnionitis? J Reprod Med 53:25–28, 2008.
510. Goldenberg RL, Andrews WW, Goepfert AR, et al: Alabama Preterm Birth Study: umbilical cord blood Ureaplasma urealyticum and Myco-plasma hominis cultures in very preterm newborn infants, Am J Obstet Gynecol 198:e1–e5, 2008.
511. Bejar R, Wozniak P, Allard M, et al: Antenatal origin of neurologic damage in newborn infants: I. Preterm infants, Am J Obstet Gynecol 159:357–363, 1988.
512. De Felice C, Toti P, Laurini RN, et al: Early neonatal brain injury in histologic chorioam-nionitis, J Pediatr 138:101–104, 2001.
513. Grafe MR: The correlation of prenatal brain damage with placental pathology, Neuropathol Exp Neurol 53:407–415, 1994.
514. Buhimschi IA, Zambrano E, Pettker CM, et al: Using proteomic analysis of the human amni-otic fluid to identify histologic chorioamnion-itis, Obstet Gynecol 111:403–412, 2008.
515. Vivian-Taylor J, Roberts CL, Chen JS, et al: Motor vehicle accidents during pregnancy: a population-based study, BJOG 119(4):499–503, 2012.
516. El-Kady D, Gilbert WM, Anderson J, et al: Trauma during pregnancy: an analysis of maternal and fetal outcomes in a large popula-tion, Am J Obstet Gynecol 190(6):1661–1668, 2004.
517. Fischer PE, Zarzaur BL, Fabian TC, et al: Minor trauma is an unrecognized contributor to poor fetal outcomes: a population-based study of 78,552 pregnancies, J Trauma 71(1):90–93, 2011.
518. Bruinsma FJ, Quinn MA: The risk of preterm birth following treatment for precancerous changes in the cervix: a systematic review and meta-analysis, BJOG 118(9):1031–1041, 2011.
519. Smith GC, Pell JP, Dobbie R: Caesarean section and risk of unexplained stillbirth in subse-quent pregnancy, Lancet 362(9398):1779–1784, 2003.
520. Ronel D, Wiznitzer A, Sergienko R, et al: Trends, risk factors and pregnancy outcome in women with uterine rupture, Arch Gynecol Obstet 285(2):317–321, 2012.
521. Lee SY, Hsu HC: Stress and health-related well-being among mothers with a low birth weight infant: the role of sleep, Soc Sci Med 74:958–965, 2012.
522. Bloch JR: Using geographical information systems to explore disparities in preterm birth rates among foreign-born and U.S.-born black mothers, J Obstet Gynecol Neonatal Nurs 40(5):544–554, 2011.
523. Torche F, Kleinhaus K: Prenatal stress, gesta-tional age and secondary sex ratio: the sex-specific effects of exposure to a natural disaster in early pregnancy, Hum Reprod 27(2):558–567, 2012.
524. Class QA, Lichtenstein P, Långström N, et al: Timing of prenatal maternal exposure to severe life events and adverse pregnancy outcomes: a population study of 2.6 million pregnancies, Psychosom Med 73(3):234–241, 2011.
525. Fransson E, Ortenstrand A, Hjelmstedt A: Antenatal depressive symptoms and preterm birth: a prospective study of a Swedish national sample, Birth 38(1):10–16, 2011.
526. Mozurkewich EL, Luke B, Avni M, et al: Working conditions and adverse pregnancy outcome: a meta-analysis, Obstet Gynecol 95(4):623–635, 2000.
527. Dunkel Schetter C, Tanner L: Anxiety, depres-sion and stress in pregnancy: implications for mothers, children, research, and practice, Curr Opin Psychiatry 25(2):141–148, 2012.
528. Copper RL, Goldenberg RL, Das A, et al: The preterm prediction study: maternal stress is associated with spontaneous preterm birth at less than thirty-five weeks’ gestation. National Institute of Child Health and Human Develop-ment Maternal-Fetal Medicine Units Network, Am J Obstet Gynecol 175:1286–1292, 1996.
529. Catov JM, Abatemarco DJ, Markovic N, et al: Anxiety and optimism associated with gesta-tional age at birth and fetal growth, Matern Child Health J 14:758–764, 2010.
530. Lockwood CJ: Stress-associated preterm deliv-ery: the role of corticotropin-releasing hormone, Am J Obstet Gynecol 180(1 Pt 3):S264–S266, 1999.
531. Lipkind HS, Curry AE, Huynh M, et al: Birth outcomes among offspring of women exposed to the September 11, 2001, terrorist attacks, Obstet Gynecol 116:917–925, 2010.
532. Xiong X, Harville EW, Mattison DR, et al: Exposure to hurricane Katrina, post-traumatic stress disorder and birth outcomes, Am J Med Sci 336(2):111–115, 2008.
533. Gavin NI, Gaynes BN, Lohr KN, et al: Perinatal depression: a systematic review of prevalence and incidence, Obstet Gynecol 106:1071–1083, 2005.
534. Alder J, Fink N, Bitzer J, et al: Depression and anxiety during pregnancy: a risk factor for obstetric, fetal and neonatal outcome? A criti-cal review of the literature, J Matern Fetal Neo-natal Med 20(3):189–209, 2007.
535. Grote NK, Bridge JA, Gavin AR, et al: A meta-analysis of depression during pregnancy and the risk of preterm birth, low birth weight, and intrauterine growth restriction, Arch Gen Psy-chiatry 67:1012–1024, 2010.
536. Orr ST, James SA, Blackmore Prince C: Mater-nal prenatal depressive symptoms and sponta-neous preterm births among African-American women in Baltimore, Maryland, Am J Epide-miol 156:797–802, 2002.
537. Ibanez G, Charles MA, Forhan A, et al; the EDEN Mother-Child Cohort Study Group: Depression and anxiety in women during pregnancy and neonatal outcome: data from the EDEN mother-child cohort, Early Hum Dev 88(8):643–649, 2012.
538. El Marroun H, Jaddoe VW, Hudziak JJ, et al: Maternal use of selective serotonin reuptake inhibitors, fetal growth, and risk of adverse birth outcomes, Arch Gen Psychiatry 69(7):706–714, 2012.
539. Buhimschi CS, Weiner CP: Medications in pregnancy and lactation: part 1. Teratology, Obstet Gynecol 113(1):166–188, 2009.
540. Buhimschi CS, Weiner CP: Medications in pregnancy and lactation: part 2. Drugs with minimal or unknown human teratogenic effect, Obstet Gynecol 113:417–432, 2009.
541. Nordeng H, van Gelder MM, Spigset O, et al: Pregnancy outcome after exposure to antide-pressants and the role of maternal depression: results from the Norwegian Mother and Child
Cohort Study, J Clin Psychopharmacol 32(2):186–194, 2012.
542. Yonkers KA, Norwitz ER, Smith MV, et al: Depression and serotonin reuptake inhibitor treatment as risk factors for preterm birth, Epidemiology 23(5):677–685, 2012.
543. El-Mohandes AA, Kiely M, Gantz MG, et al: Very preterm birth is reduced in women receiving an integrated behavioral interven-tion: a randomized controlled trial, Matern Child Health J 15(1):19–28, 2011.
544. Vedhara K, Metcalfe C, Brant H, et al: Maternal mood and neuroendocrine programming: effects of time of exposure and sex, J Neuroen-docrinol 24(7):999–1011, 2012.
545. Iams JD, Johnson FF, Sonek J, et al: Cervical competence as a continuum: a study of ultra-sonographic cervical length and obstetric per-formance, Am J Obstet Gynecol 172:1097–1103, 1995.
546. Romero R, Mazor M, Gomez R: Cervix, incom-petence and premature labor, Fetus 3:1, 1993.
547. Romero R, Gonzalez R, Sepulveda W, et al: Infection and labor: VIII. Microbial invasion of the amniotic cavity in patients with suspected cervical incompetence: prevalence and clinical significance, Am J Obstet Gynecol 167:1086–1091, 1992.
548. Romero R, Espinoza J, Erez O, et al: The role of cervical cerclage in obstetric practice: can the patient who could benefit from this proce-dure be identified? Am J Obstet Gynecol 194:1–9, 2006.
549. Kyrgiou M, Koliopoulos G, Martin-Hirsch P, et al: Obstetric outcomes after conservative treatment for intraepithelial or early invasive cervical lesions: systematic review and meta-analysis, Lancet 367(9509):489–498, 2006.
550. Arbyn M, Kyrgiou M, Simoens C, et al: Perina-tal mortality and other severe adverse preg-nancy outcomes associated with treatment of cervical intraepithelial neoplasia: meta-analysis, BMJ 337:a1284, 2008.
551. Stock SJ, Norman JE: Treatments for precur-sors of cervical cancer and preterm labour, BJOG 119(6):647–649, 2012.
552. Noehr B, Jensen A, Frederiksen K, et al: Depth of cervical cone removed by loop electrosurgi-cal excision procedure and subsequent risk of spontaneous preterm delivery, Obstet Gynecol 114(6):1232–1238, 2009.
553. Shanbhag S, Clark H, Timmaraju V, et al: Preg-nancy outcome after treatment for cervical intraepithelial neoplasia, Obstet Gynecol 114(4):727–735, 2009.
554. Reilly R, Paranjothy S, Beer H, et al: Birth out-comes following treatment for precancerous changes to the cervix: a population-based record linkage study, BJOG 119(2):236–244, 2012.
555. Frøen JF, Cacciatore J, McClure EM, et al: Lancet’s Stillbirths Series steering committee. Stillbirths: why they matter, Lancet 377(9774):1353–1366, 2011.
556. MacDorman F, Kirmeyer S: Fetal and perinatal mortality, United States, 2005, Natl Vital Stat Rep 57(8):1–19, 2009.
557. Fretts R: Stillbirth epidemiology, risk factors, and opportunities for stillbirth prevention, Clin Obstet Gynecol 53(3):588–596, 2010.
558. Goldenberg RL, Culhane JF, Iams JD, et al: Epi-demiology and causes of preterm birth, Lancet 371(9606):75–84, 2008.
559. Gordon A, Raynes-Greenow C, McGeechan K, et al: Stillbirth risk in a second pregnancy, Obstet Gynecol 119(3):509–517, 2012.

623.e12 PART 4 Disorders at the Maternal-Fetal Interface
560. Surkan PJ, Stephansson O, Dickman PW, et al: Previous preterm and small-for-gestational-age births and the subsequent risk of stillbirth, N Engl J Med 350(8):777–785, 2004.
561. Redline RW: Placental pathology: a neglected link between basic disease mechanisms and untoward pregnancy outcome, Curr Opin Obstet Gynecol 7:10–15, 1995.
562. Murphy SP, Hanna NN, Fast LD, et al: Evidence for participation of uterine natural killer cells in the mechanisms responsible for spontane-ous preterm labor and delivery, Am J Obstet Gynecol 200(3):308.e1–308.e9, 2009.
563. Murphy SP, Fast LD, Hanna NN, et al: Uterine NK cells mediate inflammation-induced fetal demise in IL-10-null mice, J Immunol 175(6):4084–4090, 2005.
564. Ryan WD, Trivedi NA, Benirschke K, et al: Pla-cental histologic criteria for diagnosis of cord accident: sensitivity and specificity, Pediatr Dev Pathol 15(4):275–280, 2012.
565. Goldenberg RL, Andrews WW, Faye-Petersen O, et al: The Alabama Preterm Birth Project: placental histology in recurrent spontaneous and indicated preterm birth, Am J Obstet Gynecol 195(3):792–796, 2006.
566. Levy RA, Avvad E, Oliveira J, et al: Placental pathology in antiphospholipid syndrome, Lupus 7:S81–S85, 1998.
567. Avagliano L, Bulfamante GP, Morabito A, et al: Abnormal spiral artery remodelling in the decidual segment during pregnancy: from his-tology to clinical correlation, J Clin Pathol 64(12):1064–1068, 2011.
568. Wapner RJ, Simpson JL, Golbus MS, et al: Cho-rionic mosaicism: association with fetal loss but not with adverse perinatal outcome, Prenat Diagn 12(5):347–355, 1992.
569. Mortensen LH, Helweg-Larsen K, Andersen AM: Socioeconomic differences in perinatal health and disease, Scand J Public Health 39(7 Suppl):110–114, 2011.
570. Battaglia FC, Lubchenco LO: A practical clas-sification of newborn infants by weight and gestational age, J Pediatr 71(2):159–163, 1967.
571. Figueras F, Gardosi J: Intrauterine growth restriction: new concepts in antenatal surveil-lance, diagnosis, and management, Am J Obstet Gynecol 204(4):288–300, 2011.
572. ACOG Practice Bulletin: intrauterine growth restriction. No. 12, January 2000. American Congress of Obstetrics and Gynecology. (Reaf-firmed 2010).
573. McIntire DD, Bloom SL, Casey BM, et al: Birth weight in relation to morbidity and mortality among newborn infants, N Engl J Med 340(16):1234–1238, 1999.
574. Hoffman HJ, Bakketeig LS: Heterogeneity of intrauterine growth retardation and recur-rence risks, Semin Perinatol 8(1):15–24, 1984.
575. Galan HL: Timing delivery of the growth-restricted fetus, Semin Perinatol 35(5):262–269, 2011.
576. Carreno CA, Costantine MM, Holland MG, et al: Approximately one-third of medically indicated late preterm births are complicated by fetal growth restriction, Am J Obstet Gynecol 204(3):263.e1–263.e4, 2011.
577. Eckmann-Scholz C, Mallek J, von Kaisenberg CS, et al: Chromosomal mosaicisms in prenatal diagnosis: correlation with first trimester screening and clinical outcome, J Perinat Med 40(3):215–223, 2012.
578. Salafia CM, Vintzileos AM, Silberman L, et al: Placental pathology of idiopathic intrauterine
growth retardation at term, Am J Perinatol 9(3):179–184, 1992.
579. Greer LG, Ziadie MS, Casey BM, et al: An immunologic basis for placental insufficiency in fetal growth restriction, Am J Perinatol 29(7):533–538, 2012.
580. Sander CM, Gilliland D, Akers C, et al: Live-births with placental hemorrhagic endovascu-litis: interlesional relationships and perinatal outcomes, Arch Pathol Lab Med 126(2):157–164, 2002.
581. Devoe LD: The nonstress test, Obstet Gynecol Clin North Am 17(1):111–128, 1990.
582. Oyelese Y, Vintzileos AM: The uses and limita-tions of the fetal biophysical profile, Clin Peri-natol 38(1):47–64, 2011.
583. Renou P, Newman W, Wood C: Autonomic control of FHR, Am J Obstet Gynecol 105:949–1053, 1969.
584. Manning FA: Fetal biophysical profile, Obstet Gynecol Clin North Am 26(4):557–577, 1999.
585. Macones GA, Hankins GD, Spong CY, et al: The 2008 National Institute of Child Health and Human Development workshop report on electronic fetal monitoring, Obstet Gynecol 112:661–666, 2008.
586. Shalev E, Dan U, Yanai N, et al: Sonography-guided fetal blood sampling for pH and blood gases in premature fetuses with abnormal fetal heart rate traces, Acta Obstet Gynecol Scand 70(7–8):539–542, 1991.
587. Jacobsson B, Hagberg G, Hagberg B, et al: Cere-bral palsy in preterm infants: a population-based case-control study of antenatal and intrapartal risk factors, Acta Paediatr 91(8):946–951, 2002.
588. Malcus P, Svenningsen N, Westgren M: Reac-tivity of non-stress tests and its relationship to outcome in infants born prior to the 33rd week of gestation, Acta Obstet Gynecol Scand 65(8):835–838, 1986.
589. Buhimschi CS, Abdel-Razeq S, Cackovic M, et al: Fetal heart rate monitoring patterns in women with amniotic fluid proteomic profiles indicative of inflammation, Am J Perinatol 25(6):359–372, 2008.
590. Andreani M, Locatelli A, Assi F, et al: Predictors of umbilical artery acidosis in preterm delivery, Am J Obstet Gynecol 197(3):303.e1–303.e5, 2007.
591. Baschat AA, Harman CR: Antenatal assessment of the growth restricted fetus, Curr Opin Obstet Gynecol 13(2):161–168, 2001.
592. Lewis DF, Adair CD, Weeks JW, et al: A ran-domized clinical trial of daily nonstress testing versus biophysical profile in the management of preterm premature rupture of membranes, Am J Obstet Gynecol 181(6):1495–1499, 1999.
593. Sherer DM, Spong CY, Salafia CM: Fetal breathing movements within 24 hours of deliv-ery in prematurity are related to histologic and clinical evidence of amnionitis, Am J Perinatol 14(6):337–340, 1997.
594. Manning FA, Bondaji N, Harman CR, et al: Fetal assessment based on fetal biophysical profile scoring: VIII. The incidence of cerebral palsy in tested and untested perinates, Am J Obstet Gynecol 178(4):696–706, 1998.
595. Baschat AA, Viscardi RM, Hussey-Gardner B, et al: Infant neurodevelopment following fetal growth restriction: relationship with antepar-tum surveillance parameters, Ultrasound Obstet Gynecol 33(1):44–50, 2009.
596. Rotmensch S, Lev S, Kovo M, et al: Effect of betamethasone administration on fetal heart
rate tracing: a blinded longitudinal study, Fetal Diagn Ther 20(5):371–376, 2005.
597. Kelly MK, Schneider EP, Petrikovsky BM, et al: Effect of antenatal steroid administration on the fetal biophysical profile, J Clin Ultrasound 28(5):224–226, 2000.
598. Gruslin A, Lemyre B: Pre-eclampsia: fetal assessment and neonatal outcomes, Best Pract Res Clin Obstet Gynaecol 25(4):491–507, 2011.
599. Battaglia FC, Regnault TR: Placental transport and metabolism of amino acids, Placenta 22:145–161, 2001.
600. Fleming AD, Salafia CM, Vintzileos AM, et al: The relationships among umbilical artery velo-cimetry, fetal biophysical profile, and placental inflammation in preterm premature rupture of the membranes, Am J Obstet Gynecol 164:38–41, 1991.
601. Salafia CM, Mangam HE, Weigl CA, et al: Abnormal FHR patterns and placental inflam-mation, Am J Obstet Gynecol 160:140–147, 1989.
602. Saleemuddin A, Tantbirojn P, Sirois K, et al: Obstetric and perinatal complications in pla-centas with fetal thrombotic vasculopathy, Pediatr Dev Pathol 13(6):459–464, 2010.
603. Madsen-Bouterse SA, Romero R, Tarca AL, et al: The transcriptome of the fetal inflamma-tory response syndrome, Am J Reprod Immunol 63(1):73–92, 2010.
604. Yafeng Dong, Weijian Hou, Jiaxue Wei, et al: Chronic hypoxemia absent bacterial infection is one cause of the fetal inflammatory response syndrome (FIRS), Reprod Sci 16(7):650–656, 2009.
605. Nelson K, Holmes LB: Malformations due to presumed spontaneous mutations in newborn infants, N Engl J Med 320(1):19–23, 1989.
606. Crane JP, LeFevre ML, Winborn RC, et al: A randomized trial of prenatal ultrasonographic screening: impact on the detection, manage-ment, and outcome of anomalous fetuses. The RADIUS Study Group, Am J Obstet Gynecol 171(2):392–399, 1994.
607. Schinzel AA, Smith DW, Miller JR: Monozy-gotic twinning and structural defects, J Pediatr 95(6):921–930, 1979.
608. Whitworth M, Bricker L, Neilson JP, et al: Ultra-sound for fetal assessment in early pregnancy, Cochrane Database Syst Rev (4):CD007058, 2010.
609. Craigo SD: Indicated preterm birth for fetal anomalies, Semin Perinatol 35(5):270–276, 2011.
610. Dolk H, Loane M, Garne E: The prevalence of congenital anomalies in Europe, Adv Exp Med Biol 686:349–364, 2010.
611. Wapner RJ: Genetics of stillbirth, Clin Obstet Gynecol 53(3):628–634, 2010.
612. ACOG Practice Bulletin No. 102: management of stillbirth, Obstet Gynecol 113(3):748–761, 2009.
613. Verbeek RJ, Heep A, Maurits NM, et al: Fetal endoscopic myelomeningocele closure pre-serves segmental neurological function, Dev Med Child Neurol 54(1):15–22, 2012.
614. Adzick NS, Thom EA, Spong CY, et al; MOMS Investigators. A randomized trial of prenatal versus postnatal repair of myelomeningocele, N Engl J Med 364(11):993–1004, 2011.
615. Deprest J, Jani J, Lewi L, et al: Fetoscopic surgery: encouraged by clinical experience and boosted by instrument innovation, Semin Fetal Neonatal Med 11(6):398–412, 2006.
616. Morken NH, Magnus P, Jacobsson B: Sub-groups of preterm delivery in the Norwegian

39 Pathogenesis of Spontaneous Preterm Birth 623.e13
Mother and Child Cohort Study, Acta Obstet Gynecol Scand 87(12):1374–1377, 2008.
617. Tam Tam KB, Briery C, Penman AD, et al: Fetal gastroschisis: epidemiological characteristics and pregnancy outcomes in Mississippi, Am J Perinatol 28(9):689–694, 2011.
618. Gul A, Cebeci A, Aslan H, et al: Perinatal out-comes of twin pregnancies discordant for major fetal anomalies, Fetal Diagn Ther 20(4):244–248, 2005.
619. Ross MG, Brace RA; National Institute of Child Health and Development Workshop Partici-pants: National Institute of Child Health and Development Conference summary: amniotic fluid biology—basic and clinical aspects, J Matern Fetal Med 10(1):2–19, 2001.
620. Many A, Hill LM, Lazebnik N, et al: The asso-ciation between polyhydramnios and preterm delivery, Obstet Gynecol 86(3):389–391, 1995.
621. Carlson DE, Platt LD, Medearis AL, et al: Quantifiable polyhydramnios: diagnosis and management, Obstet Gynecol 75(6):989–993, 1990.
622. Vink JY, Poggi SH, Ghidini A, et al: Amniotic fluid index and birth weight: is there a relation-ship in diabetics with poor glycemic control? Am J Obstet Gynecol 195(3):848–850, 2006.
623. Weinberg LE, Dinsmoor MJ, Silver RK: Severe hydramnios and preterm delivery in associa-tion with transient maternal diabetes insipi-dus, Obstet Gynecol 116 Suppl 2:547–549, 2010.
624. Chitkara U, Wilkins I, Lynch L, et al: The role of sonography in assessing severity of fetal anemia in Rh- and Kell-isoimmunized pregnancies, Obstet Gynecol 71(3 Pt 1):393–398, 1988.
625. Dashe JS, McIntire DD, Ramus RM, et al: Hydramnios: anomaly prevalence and sono-graphic detection, Obstet Gynecol 100(1):134–139, 2002.
626. Penava D, Natale R: An association of chorion-icity with preterm twin birth, J Obstet Gynaecol Can 26(6):571–574, 2004.
627. Zanardini C, Papageorghiou A, Bhide A, et al: Giant placental chorioangioma: natural history and pregnancy outcome, Ultrasound Obstet Gynecol 35(3):332–336, 2010.
628. Magann EF, Chauhan SP, Doherty DA, et al: A review of idiopathic hydramnios and preg-nancy outcomes, Obstet Gynecol Surv 62(12):795–802, 2007.
629. Prat C, Blanchon L, Borel V, et al: Ontogeny of aquaporins in human fetal membranes, Biol Reprod 86(2):48, 2012.
630. Lazar DA, Cassady CI, Olutoye OO, et al: Tra-cheoesophageal displacement index and pre-dictors of airway obstruction for fetuses with neck masses, J Pediatr Surg 47(1):46–50, 2012.
631. Obeidi N, Russell N, Higgins JR, et al: The natural history of anencephaly, Prenat Diagn 30(4):357–360, 2010.
632. Hedrick HL, Crombleholme TM, Flake AW, et al: Right congenital diaphragmatic hernia: prenatal assessment and outcome, J Pediatr Surg 39(3):319–323, 2004.
633. Houben CH, Curry JI: Current status of pre-natal diagnosis, operative management and outcome of esophageal atresia/tracheo-esoph-ageal fistula, Prenat Diagn 28(7):667–675, 2008.
634. Kurdi AM, Mesleh RA, Al-Hakeem MM, et al: Multiple pregnancy and preterm labor, Saudi Med J 25(5):632–637, 2004.
635. Martin JA, Hamilton BE, Sutton PD, et al: Births: final data for 2002, Natl Vital Stat Rep 52(10):1–113, 2003.
636. Stock S, Norman J: Preterm and term labour in multiple pregnancies, Semin Fetal Neonatal Med 15(6):336–341, 2010.
637. Sfakianaki AK, Buhimschi IA, Pettker CM, et al: Ultrasonographic evaluation of myometrial thickness in twin pregnancies, Am J Obstet Gynecol 198:530.e1–530.e10, 2008.
638. Lyall F, Lye S, Teoh T, et al: Expression of Gsalpha, connexin-43, connexin-26, and EP1, 3, and 4 receptors in myometrium of prelabor singleton versus multiple gestations and the effects of mechanical stretch and steroids on Gsalpha, J Soc Gynecol Investig 9(5):299–307, 2002.
639. Turton P, Wray S, Neilson JP, et al: Contractile properties of myometrium in twin pregnan-cies, Arch Dis Child Fetal Neonatal Ed 15:93, 2008.
640. Mazor M, Hershkovitz R, Ghezzi F, et al: Intraamniotic infection in patients with preterm labor and twin pregnancies, Acta Obstet Gynecol Scand 75:624–627, 1996.
641. Romero R, Shamma F, Avila C, et al: Infection and labor: VI. Prevalence, microbiology, and clinical significance of intraamniotic infection in twin gestations with preterm labor, Am J Obstet Gynecol 163:757–761, 1990.
642. Buhimschi IA, Buhimschi CS, Christner R, et al: Proteomics technology for the accurate diagnosis of inflammation in twin pregnan-cies, BJOG 112(2):250–255, 2005.
643. Peláez LM, Chasen ST, Baergen RN: The rela-tionship between placental histology and cer-vical length in twin gestations, J Perinat Med 38(5):485–489, 2010.
644. Salihu HM, Bekan B, Aliyu MH, et al: Perinatal mortality associated with abruptio placenta in singletons and multiples, Am J Obstet Gynecol 193(1):198–203, 2005.
645. Sunderam S, Chang J, Flowers L, et al; Centers for Disease Control and Prevention: Assisted reproductive technology surveillance—United States, 2006, MMWR Surveill Summ 58(5):1–25, 2009.
646. Reddy UM, Wapner RJ, Rebar RW, et al: Infertility, assisted reproductive technology, and adverse pregnancy outcomes: executive summary of a National Institute of Child Health and Human Development workshop, Obstet Gynecol 109(4):967–977, 2007.
647. Lockwood CJ: Testing for risk of preterm deliv-ery, Clin Lab Med 23(2):345–360, 2003.
648. Challis JR, Lockwood CJ, Myatt L, et al: Inflam-mation and pregnancy, Reprod Sci 16:206–215, 2009.
649. Buhimschi CS, Schatz F, Krikun G, et al: Novel insights into molecular mechanisms of abruption-induced preterm birth, Expert Rev Mol Med 12:e35, 2010.
650. Steel JH, O’donoghue K, Kennea NL, et al: Maternal origin of inflammatory leukocytes in preterm fetal membranes, shown by fluores-cence in situ hybridisation, Placenta 26:672–677, 2005.
651. Buhimschi CS, Bhandari V, Dulay AT, et al: Amniotic fluid angiopoietin-1, angiopoietin-2, and soluble receptor tunica interna endothelial cell kinase-2 levels and regulation in normal pregnancy and intraamniotic inflammation-induced preterm birth, J Clin Endocrinol Metab 95:3428–3436, 2010.
652. Lundin-Schiller S, Mitchell MD: Prostaglandin production by human chorion laeve cells in response to inflammatory mediators, Placenta 12(4):353–363, 1991.
653. Sato TA, Keelan JA, Mitchell MD: Critical para-crine interactions between TNF-alpha and IL-10 regulate lipopolysaccharide-stimulated human choriodecidual cytokine and prosta-glandin E2 production, J Immunol 170(1):158–166, 2003.
654. Benirschke K: Examination of the placenta, Obstet Gynecol 18:309–333, 1961.
655. Naeye RL: Disorders of the placenta and decidua. In: Disorders of the placenta, fetus and neonate: diagnosis and clinical significance, St. Louis, 1992, Mosby, pp 118–247.
656. Salafia CM, Weigl C, Silberman L: The preva-lence and distribution of acute placental inflammation in uncomplicated term preg-nancies, Obstet Gynecol 73:383–389, 1989.
657. Redline RW: Placental pathology and cerebral palsy, Clin Perinatol 33:503–516, 2006.
658. Redline RW, Faye-Petersen O, Heller D, et al: Amniotic infection syndrome: nosology and reproducibility of placental reaction patterns, Pediatr Dev Pathol 6:435–448, 2003.
659. van Hoeven KH, Anyaegbunam A, Hochster H, et al: Clinical significance of increasing histo-logic severity of acute inflammation in the fetal membranes and umbilical cord, Pediatr Pathol Lab Med 16:731–744, 1996.
660. Ananth CV, Oyelese Y, Yeo L, et al: Placental abruption in the United States, 1979 through 2001: temporal trends and potential determi-nants, Am J Obstet Gynecol 192:191–198, ••.
661. Oyelese Y, Ananth CV: Placental abruption, Obstet Gynecol 108:1005–1016, 2006.
662. Ananth CV, Wilcox AJ: Placental abruption and perinatal mortality in the United States, Am J Epidemiol 153:332–337, 2001.
663. Lockwood CJ, Toti P, Arcuri F, et al: Mecha-nisms of abruption-induced premature rupture of the fetal membranes: thrombin-enhanced interleukin-8 expression in term decidua, Am J Pathol 167(5):1443–1449, 2005.
664. Lockwood CJ, Krikun G, Rahman M, et al: The role of decidualization in regulating endome-trial hemostasis during the menstrual cycle, gestation, and in pathological states, Semin Thromb Hemost 33(1):111–117, 2007.
665. Oyelese Y, Smulian JC: Placenta previa, pla-centa accreta, and vasa previa, Obstet Gynecol 107:927–941, 2006.
666. Iyasu S, Saftlas AK, Rowley DL, et al: The epi-demiology of placenta previa in the United States, 1979 through 1987, Am J Obstet Gynecol 168:1424–1429, 1993.
667. Fishman SG, Chasen ST, Maheshwari B: Risk factors for preterm delivery with placenta previa, J Perinat Med 40(1):39–42, 2011.
668. Chaouat G, Dubanchet S, Ledée N: Cytokines: important for implantation? J Assist Reprod Genet 24:491–505, 2007.
669. Mardon H, Grewal S, Mills K: Experimental models for investigating implantation of the human embryo, Semin Reprod Med 25:410–417, 2007.
670. Dimitriadis E, Nie G, Hannan NJ, et al: Local regulation of implantation at the human fetal-maternal interface, Int J Dev Biol 54:313–322, 2010.
671. Wehrum MJ, Buhimschi IA, Salafia C, et al: Accreta complicating complete placenta previa is characterized by reduced systemic levels of vascular endothelial growth factor and by epithelial-to-mesenchymal transition of the invasive trophoblast, Am J Obstet Gynecol 204(5):411.e1–411.e11, 2011.

623.e14 PART 4 Disorders at the Maternal-Fetal Interface
672. Soregaroli M, Bonera R, Danti L, et al: Prog-nostic role of umbilical artery Doppler velo-cimetry in growth-restricted fetuses, J Matern Fetal Neonatal Med 11(3):199–203, 2002.
673. Rahemtullah A, Lieberman E, Benson C, et al: Outcome of pregnancy after prenatal diagnosis of umbilical vein varix, J Ultrasound Med 20(2):135–139, 2001.
674. de Laat MW, van Alderen ED, Franx A, et al: The umbilical coiling index in complicated
pregnancy, Eur J Obstet Gynecol Reprod Biol 130(1):66–72, 2007.
675. Heifetz SA: Thrombosis of the umbilical cord: analysis of 52 cases and literature review, Pediatr Pathol 8:37–54, 1988.
676. Dickinson JE: Monoamniotic twin pregnancy: a review of contemporary practice, Aust N Z J Obstet Gynaecol 45(6):474–478, 2005.
677. Beinder E, Voigt HJ, Jäger W, et al: [Partial hydatidiform mole in a cytogenetically normal
fetus.], Geburtshilfe Frauenheilkd 55(6):351–353, 1995.
678. Nuutinen EM, Puistola U, Herva R, et al: Two cases of large placental chorioangioma with fetal and neonatal complications, Eur J Obstet Gynecol Reprod Biol 29(4):315–320, 1988.
679. Shukunami K, Tsunezawa W, Hosokawa K, et al: Placenta previa of a succenturiate lobe: a report of two cases, Eur J Obstet Gynecol Reprod Biol 99(2):276–277, 2001.