pax9 and msx1 transcription factor genes in non-syndromic dental agenesis
TRANSCRIPT

PAX9 and MSX1 transcription factor genes in non-syndromicdental agenesis
Vanessa Rodrigues Paixao-Cortes a,1, Tatiana Braga b,1, Francisco Mauro Salzano a,Karina Mundstock b, Carlos Alberto Mundstock b, Maria Catira Bortolini a,*aDepartamento de Genetica, Instituto de Biociencias, Universidade Federal do Rio Grande do Sul,
Caixa Postal 15053, 91501-970 Porto Alegre, RS, Brazilb Faculdade de Odontologia, Universidade Federal do Rio Grande do Sul, Caixa Postal 1118, 90035-003 Porto Alegre, RS, Brazil
a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4
a r t i c l e i n f o
Article history:
Accepted 27 October 2010
Keywords:
PAX9 gene
MSX1 gene
Dental agenesis
Developmental genes
a b s t r a c t
Objective: The molecular variation of paired domain box gene 9 (PAX9) was previously
investigated by our research group and a high degree of evolutionary conservation in coding
and non-coding regions was observed except in exon 3. PAX9 is a transcription factor
important in tooth development, and we wanted to verify its role in dental agenesis in detail.
Since dental development is a complex trait we also decided to examine the influence of
another transcription factor, muscle segment homeodomain-homeobox 1 (MSX1) on it.
Design: A total of 360 consecutively ascertained patients seeking orthodontic treatment
were screened for tooth agenesis and 33% of them were found to have it. Thirty-five of those
with agenesis and 15 controls had their DNA studied for PAX9 exons 2, 3, 4 and adjacent
regions (total of 1476 base pairs, bp) as well as MSX1 exon 2 (698 bp). A trio (a proband and her
parents) was also studied.
Results: Six polymorphic sites were found, three in PAX9 exon 3 and three in MSX1 exon2.
MSX1 rs1095 derived allele occurred in individuals with agenesis only, and two other
mutations in this gene had been earlier associated with tooth agenesis. Homozygosity
for the PAX9 Ala240Pro mutation was studied in a family (proband and her parents),
suggesting recessive inheritance with variable expressivity for the dental agenesis found.
Conclusion: Common variants located out of the DNA binding domain of the two PAX9 and
MSX1 genes can also be related to tooth agenesis.
# 2010 Elsevier Ltd. All rights reserved.
avai lab le at www.sc iencedi rec t .com
journal homepage: http://www.elsevier.com/locate/aob
1. Introduction
Theroleofheterodonty for themammalian evolutionaryhistory
is well-recognized.1,2 For humans, teeth have also a prominent
relevance to socio-cultural interactions and at an individual
level can represent a bad or good life quality.3,4 Agenesis of one
or more teeth is the most common anomaly observed in the
* Corresponding author. Tel.: +55 51 3308 9844.E-mail address: [email protected] (M.C. Bortolini).
1 These two authors contributed equally to this work.0003–9969/$ – see front matter # 2010 Elsevier Ltd. All rights reservedoi:10.1016/j.archoralbio.2010.10.020
human craniofacial development.1,3,5–7 Amongst all non-syn-
dromic (familial or sporadic) agenesis conditions detected in
humans, the most common is the absence of third molar(s) – in
average about 20% of the individuals in a population do not have
at least one third molar. Upper lateral incisors and second
premolar ageneses are also common, being second in frequen-
cies (2.2% and 3.4%, respectively).8–10
d.

a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4338
Variation in these frequencies between and within conti-
nental human groups has been found. Third molar agenesis
occurrence, for example, increases in a gradient from Sub-
Saharan Africa (�2%) to Europe (�20%) and Asia (�30%).11–21
Polder et al.22, in a meta-analysis, observed that gender
differences can also be found, females being 1.4 times more
susceptible to non-syndromic dental agenesis than males.
Changes in the expression and/or structure of transcription
factors are common genetic causes of absence of one or more
teeth in non-syndromic agenesis. Mutations in the Paired Box
9 (PAX9) and in the muscle segment homeodomain-homeobox
1 (MSX1) transcription factor genes have been linked to failure
in tooth development.23–29 Up to now, 16 and 11 distinct
mutations in the PAX9 and MSX1 genes, respectively, have
been identified in humans (http://www.ncbi.nlm.nih.gov/
omim – OMIM#167416; OMIM#142893), all resulting in dental
agenesis. The majority of these mutations are located in the
PAX9 paired box domain (segment of 381 bp inside exon 2) and
in the MSX1 homeodomain (segment of 180 bp inside exon 2),
both DNA binding regions. The only mutation in the OMIM list
that is not located in a DNA binding domain, considering both
genes cited here, is a nucleotide substitution in PAX9 exon 4,
which introduces one premature stop codon.25
Pereira et al.30 demonstrated that a common polymor-
phism (Ala240Pro; rs4904210) in PAX9 exon 3 is probably
functional and could be associated with third molar agenesis
and its different distributions around the world. Their results
are in agreement with a family study that showed that the
derived allele (240Pro) has a significant role in third molar
agenesis.31,32 Pawlowska et al.29 on the other hand, suggested
that two polymorphisms in MSX1 exon 2 untranslated region
(rs8670 and rs12532) were involved with familial and sporadic
agenesis in humans. These results introduced the idea that
regions out of the DNA binding domain of these two
transcription factor genes could also be related to tooth
development.
The present report reviews the influence of genetic factors in
tooth development and describes our observations of tooth
agenesis in a family trio and a pilot study on a sample of patients
who received orthodontic treatment at an orthodontic clinic of
the Federal University of Rio Grande do Sul (UFRGS), Porto
Alegre, Brazil. Patients with tooth agenesis were screened for
molecular variation in PAX9 and MSX1 genes.
2. Subjects and methods
2.1. Participants and clinical examination
An initial group of 360 consecutively ascertained patients
who received orthodontic treatment at the UFRGS were
selected. Forty-three of them were Blacks and the remaining
(317s) were Whites. The urban complex formed by Porto
Alegre and neighbouring cities has 3,152,596 inhabitants, 7%
and 88% of whom are classified as Blacks ( pretos, in
Portuguese) and Whites (brancos), respectively (Brazilian
Institute of Geography and Statistics-IBGE, www.ibge.gov.br,
2000 census). In Brazil, skin colour rather than close or remote
ancestry is used to define an equivalent to ‘‘race’’, and in the
present study the word ‘‘Black’’ was employed to refer to
pretos or any person identified or self-identified with another
term that suggests major African ancestry, such as mulato or
pardo. ‘‘White’’ was used to define those who, based on their
physical traits and information, show no admixture with
non-Europeans. One-hundred and fifty eight of them were
males and 202 females. A total of 119 of these 360 patients
presented congenital non-syndromic dental agenesis (ab-
sence of at least one secondary tooth, including third molars).
Thirty-five of them (all White) accepted to participate in the
genetic investigation. Parents of one proband were also
studied.
Tooth agenesis was characterized by panoramic radio-
graphs and careful examination of their clinical charts.
Medical, birth defects, and family histories were gathered to
identify possible associated anomalies and to differentiate the
non-syndromic from the syndromic agenesis. No other dental
anomalies beyond agenesis were observed. We also randomly
included 15 individuals, without any dental agenesis, as a
control group (all Whites).
2.2. DNA extraction, PAX9/MSX1 PCR amplification andsequencing
Genomic DNA was extracted from saliva using the QIAamp
DNA MiniKit (Qiagen). PAX9 exon 3 (138 bp) and its 50and 30
flanking intronic segments (232 bp and 219 bp, respectively)
were amplified using primers and conditions described in
Pereira et al.30 Primers were designed to amplify PAX9 exons 2
(640 bp) and 4 (247 bp) (Table S1, Supplementary Data). With
this approach all PAX9 coding regions were covered (since
exon 1 presents just the initiation code) MSX1 exon 2 (698 bp,
involving both the homeodomain and the untranslated region)
was amplified using primers and conditions described in Xuan
et al.33. PCR products were purified using exonuclease I and
alkaline phosphatase (Amersham Biosciences). Both DNA
strands were sequenced using ABI Prism BigDye and an ABI
310 Genetic Analyzer.
2.3. Data analysis
Information about the 360 patients was collected and
organized in a database with complete dental description.
The SPSS program (version 16) was used to analyse the data
concerning dental agenesis. Nonparametric tests (distribution
free Kruskal–Wallis and Chi-square) were used to compare
agenesis by gender, age, skin colour (White or Black), tooth
types (third molars, molars, premolars, canine and incisors)
and other dental categories (left and right quadrants; upper
and lower arches).
Sequences were aligned and their quality, as well as the
precision of the resulting data, was ascertained using the
PHRED, PHRAP and CONSED program (http://www.genome.-
washington.edu). All chromatograms were visualized and
checked manually to detect possible mutations in the
sequence. Deviations from the reference sequence were
compared with available genome databases (Ensembl –
http://www.ensembl.org/index.html, UCSC Genome Browser
– http://genome.ucsc.edu) and SNP banks (dbSNP http://
www.ncbi.nlm.nih.gov/snp/, Hapmap – http://hapmap.nc-
bi.nlm.nih.gov/).

Table 1 – Number of individuals with dental agenesis.
Group Number
Total of patients 360
Non-syndromic agenesis patients 119
Dental agenesis class
Hypodontia (� 6 teeth missing) 115
Oligodontia (� 6 teeth missing) 4
Dental agenesis categories
Third molar 66
Othera 28
Third molar + othera 25
a Agenesis in almost all other teeth were found. Exceptions were
the upper central incisors (numbers 9 and 8) and lower canines (22
and 27; Fig. 1).
a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4 339
Allele frequencies were determined by direct counting.
Haplotypes for the PAX9 and MSX1 genes were estimated from
phase-unknown multi-site genotypes using multiple locus
haplotype analysis (MLOCUS).34,35
Observed allele and genotype frequencies of patients and
controls were compared by Chi-square with a 95% confidence
interval using the SPSS program (version 16).
Genes related to odontogenesis were compiled in the Gene
Ontology website database using the AmiGO browser (http://
amigo.geneontology.org/cgi-bin/amigo/browse.cgi). Associa-
tion between these genes and dental development was tested
using the STRING software (known and predicted protein–
protein interactions – http://string.embl.de/database), assum-
ing the highest confidence value (0.900).
3. Results
3.1. Dental agenesis frequencies
Table 1 shows that a total of 119 (33%) of the 360 patients
presented non-syndromic dental agenesis. Amongst them,
third molar absence was the most common, 66 (55%) of the
individuals showing it in isolation. Twenty-eight (23%)
presented agenesis of other teeth, whilst agenesis of other
Fig. 1 – Dental agenesis distribution by tooth. The universal nu
Association, was used to identify each tooth (http://www.ada.o
dental groups plus third molar was present in 25 (21%). Four
patients had oligodontia (at least six teeth missing).
Fig. 1 illustrates the dental agenesis distribution by tooth
considering the whole dentition. As indicated, third molar is
the most common missing tooth (14%), followed by premolar
(2%) and incisor (1%), whereas the occurrence of canine and
other molar ageneses is much less frequent (0.3% and 0.9%,
respectively). Comparisons between left and right quadrants,
or upper and lower arches agenesis showed no significant
statistical differences.
Females presented more tooth agenesis than males when
only upper teeth were considered (9, 10, 11, 12, 13, 14, 15, 16, 8,
7, 6, 5, 4, 3, 2, 1 teeth; Fisher’s exact test p-value = 0.037).
However, removing the upper third molars (18 and 28 teeth) of
the analysis, the significance is lost (p-value = 0.064). A
significant value between genders was also obtained when
all incisors were compared (9, 10, 8, 7, 25, 26, 24, 23 teeth;
Fisher’s exact test p-value = 0.022) (data not shown).
Whites presented more tooth agenesis than Blacks when the
upper teeth, left and right quadrant, as well as molar dental
groups were considered separately. However, the significance
of these differences is lost (with exception of the right quadrant)
when third molars are excluded from the analysis (Table S2).
Third molar agenesis frequency differences are expected, since
it is well known that third molar absence is rare in Sub-Saharan
Africans as compared to Europeans.11–21
3.2. PAX9 and MSX1 gene polymorphisms
Sequences of the DNA binding domain and of other regions of
the MSX1 and PAX9 genes were obtained for 35 patients with
distinct tooth agenesis and respective controls (Table 2). The
following PAX9 and MSX1 nucleotide sequences were submit-
ted to GenBank, IDs: HM213907–HM214140. No mutation was
found in PAX9 exons 2 and 4. Sequencing revealed, however,
six nucleotide substitutions outside the DNA binding domains
of both genes (PAX9 exon 3: rs12881240, rs4904210; 50 flanking
intronic segment of PAX9 exon 3: rs7143727; untranslated
region of MSX1 exon 2: rs8670, rs1095, rs12532), all recognized
as single-nucleotide polymorphisms in the available data-
bases (Table 2).
mbering system, which is adopted by the American Dental
rg/).
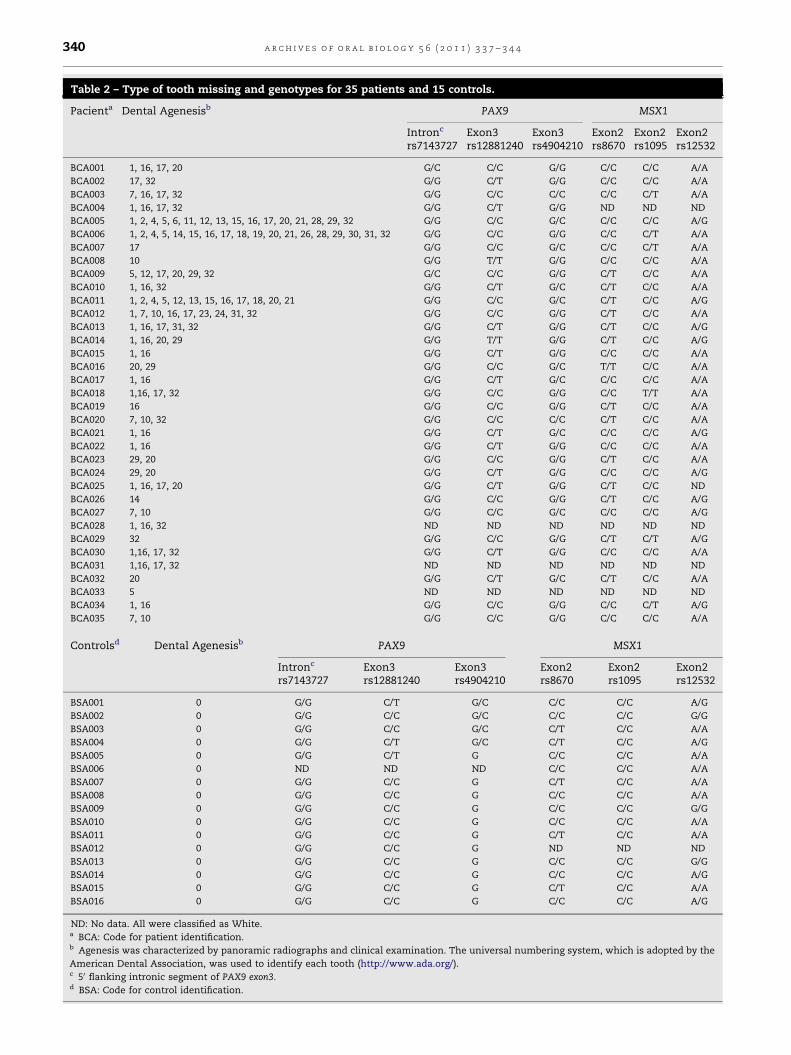
Table 2 – Type of tooth missing and genotypes for 35 patients and 15 controls.
Pacienta Dental Agenesisb PAX9 MSX1
Intronc
rs7143727Exon3rs12881240
Exon3rs4904210
Exon2rs8670
Exon2rs1095
Exon2rs12532
BCA001 1, 16, 17, 20 G/C C/C G/G C/C C/C A/A
BCA002 17, 32 G/G C/T G/G C/C C/C A/A
BCA003 7, 16, 17, 32 G/G C/C C/C C/C C/T A/A
BCA004 1, 16, 17, 32 G/G C/T G/G ND ND ND
BCA005 1, 2, 4, 5, 6, 11, 12, 13, 15, 16, 17, 20, 21, 28, 29, 32 G/G C/C G/C C/C C/C A/G
BCA006 1, 2, 4, 5, 14, 15, 16, 17, 18, 19, 20, 21, 26, 28, 29, 30, 31, 32 G/G C/C G/G C/C C/T A/A
BCA007 17 G/G C/C G/C C/C C/T A/A
BCA008 10 G/G T/T G/G C/C C/C A/A
BCA009 5, 12, 17, 20, 29, 32 G/C C/C G/G C/T C/C A/A
BCA010 1, 16, 32 G/G C/T G/C C/T C/C A/A
BCA011 1, 2, 4, 5, 12, 13, 15, 16, 17, 18, 20, 21 G/G C/C G/C C/T C/C A/G
BCA012 1, 7, 10, 16, 17, 23, 24, 31, 32 G/G C/C G/G C/T C/C A/A
BCA013 1, 16, 17, 31, 32 G/G C/T G/G C/T C/C A/G
BCA014 1, 16, 20, 29 G/G T/T G/G C/T C/C A/G
BCA015 1, 16 G/G C/T G/G C/C C/C A/A
BCA016 20, 29 G/G C/C G/C T/T C/C A/A
BCA017 1, 16 G/G C/T G/C C/C C/C A/A
BCA018 1,16, 17, 32 G/G C/C G/G C/C T/T A/A
BCA019 16 G/G C/C G/G C/T C/C A/A
BCA020 7, 10, 32 G/G C/C C/C C/T C/C A/A
BCA021 1, 16 G/G C/T G/C C/C C/C A/G
BCA022 1, 16 G/G C/T G/G C/C C/C A/A
BCA023 29, 20 G/G C/C G/G C/T C/C A/A
BCA024 29, 20 G/G C/T G/G C/C C/C A/G
BCA025 1, 16, 17, 20 G/G C/T G/G C/T C/C ND
BCA026 14 G/G C/C G/G C/T C/C A/G
BCA027 7, 10 G/G C/C G/C C/C C/C A/G
BCA028 1, 16, 32 ND ND ND ND ND ND
BCA029 32 G/G C/C G/G C/T C/T A/G
BCA030 1,16, 17, 32 G/G C/T G/G C/C C/C A/A
BCA031 1,16, 17, 32 ND ND ND ND ND ND
BCA032 20 G/G C/T G/C C/T C/C A/A
BCA033 5 ND ND ND ND ND ND
BCA034 1, 16 G/G C/C G/G C/C C/T A/G
BCA035 7, 10 G/G C/C G/G C/C C/C A/A
Controlsd Dental Agenesisb PAX9 MSX1
Intronc
rs7143727Exon3rs12881240
Exon3rs4904210
Exon2rs8670
Exon2rs1095
Exon2rs12532
BSA001 0 G/G C/T G/C C/C C/C A/G
BSA002 0 G/G C/C G/C C/C C/C G/G
BSA003 0 G/G C/C G/C C/T C/C A/A
BSA004 0 G/G C/T G/C C/T C/C A/G
BSA005 0 G/G C/T G C/C C/C A/A
BSA006 0 ND ND ND C/C C/C A/A
BSA007 0 G/G C/C G C/T C/C A/A
BSA008 0 G/G C/C G C/C C/C A/A
BSA009 0 G/G C/C G C/C C/C G/G
BSA010 0 G/G C/C G C/C C/C A/A
BSA011 0 G/G C/C G C/T C/C A/A
BSA012 0 G/G C/C G ND ND ND
BSA013 0 G/G C/C G C/C C/C G/G
BSA014 0 G/G C/C G C/C C/C A/G
BSA015 0 G/G C/C G C/T C/C A/A
BSA016 0 G/G C/C G C/C C/C A/G
ND: No data. All were classified as White.a BCA: Code for patient identification.b Agenesis was characterized by panoramic radiographs and clinical examination. The universal numbering system, which is adopted by the
American Dental Association, was used to identify each tooth (http://www.ada.org/).c 50 flanking intronic segment of PAX9 exon3.d BSA: Code for control identification.
a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4340

Table 3 – Genotype and allele frequencies (in %) for the 35 agenesis patients and 15 controls.
Variation PAX9
rs7143727 rs12881240 rs4904210 (Ala240Pro)
Patients Controls p-Valuea Patients Controls p-Valuea Samples Controls p-Valuea
Allele 0.832 0.595 0.139
G 97 100 C 75 75 G 80 73
C 3 0 T 25 25 C 20 27
Genotype
GG 94 100 CC 57 90 GG 66 73
GC 6 0 CT 37 10 GC 28 27
CC 0 0 TT 6 0 CC 6 0
Variation MSX1
rs8670 rs1095 rs12532
Patients Controls p-Valuea Patients Controls p-Valuea Patients Controls p-Valuea
Allele 0.582 0.144 0.325
C 76 83 C 89 100 C 89 67
T 24 17 T 11 0 T 11 33
Genotype
CC 55 67 CC 81 100 CC 81 53
CT 42 33 CT 16 0 CT 16 27
TT 3 0 TT 3 0 TT 3 20
a p-Values refer to the genotype comparisons.
a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4 341
There is no statistical difference between allele and
genotype distributions in patients and controls (Table 3).
Kim et al.27 and Nieminen7 suggested that MSX1 and PAX9
differ in their influence for agenesis of specific teeth. Both
genes affect third molars, but significantly higher frequencies
of agenesis for second premolars and maxillary first premolars
Fig. 2 – (a) Panoramic radiograph of II-1 (BCA003). Missing teeth a
arrow; (b) pedigree of the family considered; and (c) chromatogr
homozygous mutation for 240Pro (rs4904210) in the patient, the
profiles.
were found in association with MSX1 mutations as compared
to PAX9 mutations. Whilst, agenesis of the maxillary first and
second molars and mandibular second molars was signifi-
cantly more common in association with PAX9 nucleotide
substitutions. Usually the upper lateral or lower incisors are
also affected. Interestingly, some reports on MSX1 mutations
re marked with an T; an impacted tooth is marked with an
am showing a PAX9 exon 3 reference sequence, the
heterozygote mother and the homozygote for 240Pro father

a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4342
describe agenesis of the first permanent molars even in the
presence of second molars. We tested each agenesis dental
category for association with the MSX1 and PAX9 polymorph-
isms, and although the same general tendencies listed above
were found, the values were not significant. These results,
however, should be considered with caution since the sample
sizes used for our case–control comparisons were small.
Multiple locus haplotype analysis showed no linkage disequi-
librium between the PAX9 or MSX1 alleles.
Although the case–control results showed no association
with the PAX9 andMSX1 variation, it should be mentioned that
in two individuals, BCA003 (7, 16, 1, 17) and BCA020 (10, 7, 32;
Table S2, Supplementary Data) where the derived allele
(240Pro; PAX9 exon 3) appears in homozygosity, third molar(s),
as well as upper lateral incisor(s), are absent. For BCA003, a
woman with absence of three third molars and one upper
lateral incisor, it was possible to obtain sequences of the PAX9
exon 3 of her parents. Interestingly, her father, who presents
the four third molars missing is also homozygote for the
240Pro allele. The mother, on the other hand, is heterozygote
Fig. 3 – Fourty-two genes of the dental development network. I
Interacting Genes/P Genes/Proteins – http://string.embl.de) with
types of lines represent the kind of evidence for the association
G/C and does not present missing teeth (Fig. 2). No
homozygotes for the 240Pro allele were found in our control
sample.
4. Discussion
In the present study 33% of the subjects who received
orthodontic treatment had agenesis of one or more teeth.
Third molar is the tooth with the highest agenesis frequency,
followed by the lower premolars and upper lateral incisors.
Some differences between genders and skin colour groups
were found, but generally they disappear if third molars are
excluded of the analysis.
Sequences of the untranslated MSX1 exon 2 region, and of
the PAX9 exons 2, 3 and 4 were obtained for 35 patients with
distinct dental agenesis. Although no new or previously
described mutations located in the DNA binding domain for
both genes were identified, six substitutions located outside
this domain were found.
mage created by STRING (Search Tool for the Retrieval of
the required highest confidence score – 0.900. Different
.

a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4 343
Although the case–control results showed no significant
differences, some findings deserve a comment; for instance,
the MSX1 rs1095 derived allele appeared in agenesis affected
patients only (no mutant allele C was found in controls). This
variant was absent in a sample of Euro-descendents studied
earlier (dbSNP database – http://www.ncbi.nlm.nih.gov/snp/).
The other two MSX1 polymorphisms (rs8670 and rs12532) had
earlier been associated with dental agenesis.29 PAX9 rs7143727
derived allele also appeared in agenesis affected individuals
only (no mutant allele T was found in controls). However,
differently from the other substitutions, this variant is located
in a non-coding region (50 flanking intronic segment of PAX9
exon 3).
Anearlier familystudyshowedthat theAla240Pro (PAX9exon
3) mutation seems to produce a recessive pattern of inheritance,
since all homozygotes for it had missing third molar(s) as well as
lateral incisor(s).31,32 Additionally, Pereira et al.30 based on
population and protein modelling data, suggested that 240Pro
homozygotes might present a PAX9 protein with a slightly
reduced DNA-binding capacity, which could be specifically
associated to third molar(s) absence. Our data reinforce the role
of the Ala240Pro polymorphism in these situations, but if the
inheritance isrecessive there isvariablephenotypeexpressivity,
since the number of missing third molars is different for each
patient. Our results also indicate a possible role of this
polymorphism for lateral incisor development but in this case
other factors may be involved, since one 240Pro homozygote
studied here presents all lateral incisors (the father in Fig. 2).
Finally, it should be stressed that non-syndromic congeni-
tal missing tooth is a complex and heterogeneous trait.7 Fig. 3
shows a network involving 42 teeth development genes,
including the two studied here. Table S3 give details of each
gene of this network, their interconnections, and the wide
range of their functions. In this context, and based in our
results, MSX1 and PAX9 appear to influence different agenesis
phenotypes, with other known and unknown genes as well as
epigenetic factors having an influence in tooth development.
For instance, nine Ala240Pro G/C heterozygote patients
present third molar agenesis, whilst the trio’s mother and
other four controls with this same condition show no agenesis
(Table 2 and Fig. 2). These results illustrate the importance of
these other factors in tooth development and agenesis.
5. Conclusion
Our results support an earlier finding that the derived 240Pro
allele (PAX9 exon 3) is related with third molar agenesis and
that it may have a recessive pattern of inheritance with
variable expressivity. On the other hand, MSX1 rs1095 derived
allele appeared in agenesis affected individuals only. These
results suggest that common variants located out of the DNA
binding domain of these two transcription factor genes can
also be related to tooth agenesis.
Acknowledgement
We would like to thank the patients and controls who made
this study possible.
Funding: This research was supported by Conselho Nacional
de Desenvolvimento Cientıfico e Tecnologico and Fundacao de
Amparo a Pesquisa do Estado do Rio Grande do Sul.
Competing interest: None declared.
Ethical approval: Informed consent was obtained from all of
the participants, and the project was approved by the Research
and Ethics Committee of the Federal University of Rio Grande
do Sul. In the case of children under 15 years of age, consent
was requested from their parents or from the individual
legally in charge of the child.
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.archoral-
bio.2010.10.020.
r e f e r e n c e s
1. Vastardis H. The genetics of human tooth agenesis: newdiscoveries for understanding dental anomalies. Am J OrthodDentofacial Orthop 2000;117:650–6.
2. Yamanaka A, Yasui K, Sonomura T, Uemura M.Development of heterodont dentition in house shrew(Suncus murinus). Eur J Oral Sci 2007;115:433–40.
3. Nomura R, Shimizu T, Asada Y, Hirukawa S, Maeda T.Genetic mapping of the absence of third molars in EL miceto chromosome 3. J Dent Res 2003;82:786–90.
4. Silvestri AR, Singh I. The unresolved problem of the thirdmolar: would people be better off without it? J Am Dent Assoc2003;134:450–5.
5. Burzynski NJ, Escobar VH. Classification and geneticsof numeric anomalies of dentition. Birth Defects 1983;19:95–106.
6. Kapadia H, Mues G, D’Souza RN. Genes affectingtooth morphogenesis. Orthod Craniofacial Res 2007;10:105–13.
7. Nieminen P. Genetic basis of tooth agenesis. J Exp Zool (MolDev Evol) 2009;312B:320–42.
8. Symons AL, Stritzel F, Stamation J. Anomalies associatedwith hypodontia of the permanent lateral incisor andsecond premolar. J Clin Pediatr Dent 1993;17:109–11.
9. Arte S, Nieminen P, Apajalahti S, Haavikko K, Thesleff I,Pirinen S. Characteristics of incisor–premolar hypodontia infamilies. J Dent Res 2001;80:1445–50.
10. Matalova E, Fleischmannova J, Sharpe PT, Tucker AS. Toothagenesis: from molecular genetics to molecular dentistry. JDent Res 2008;87:617–23.
11. Bredy E, Erbring C, Hubenthal B. The incidence ofhypodontia with the presence and absence ofwisdom teeth. Dtsch Zahn Mund Kieferheilkd Zentralbl1991;79:357–63.
12. Irish JD. Biological affinities of late Pleistocene through modernAfrican aboriginal populations: the dental evidence. PhD thesis.Tempe: Arizona State University; 1993.
13. Irish JD. Characteristic high- and low-frequency dentaltraits in sub-Saharan African populations. Am J PhysAnthropol 1997;102:455–67.
14. Irish JD, Guatelli-Steinberg D. Ancient teeth and modernhuman origins: an expanded comparison of African Plio-Pleistocene and recent world dental samples. J Hum Evol2003;45:113–44.
15. Turner II CG. The first Americans: the dental evidence. NatlGeogr Res 1986;2:37–46.

a r c h i v e s o f o r a l b i o l o g y 5 6 ( 2 0 1 1 ) 3 3 7 – 3 4 4344
16. Turner II CG. Late Pleistocene and Holocene populationhistory of East Asia based on dental variation. Am J PhysAnthropol 1987;73:305–21.
17. Turner II CG, Markowitz M. Dental discontinuitybetween late Pleistocene and recent Nubians. I. Peoplingof the Eurafrican–South Asian triangle. Homo 1990;41:32–41.
18. Rozkovcova E, Markova M, Dolejsı J. Studies on agenesis ofthird molars amongst populations of different origin. Sb Lek1999;100:71–84.
19. Rozkovcova E, Markova M, Lanık J, Zvarova J. Developmentof third molar in the Czech population. Prague Med Rep2004;105:391–422.
20. Rozkovcova E, Markova M, Lanık J, Zvarova J. Agenesis ofthird molars in young Czech population. Prague Med Rep2004;105:35–52.
21. Delgado-Burbano ME. Population affinities of AfricanColombians to Sub-Saharan Africans based on dentalmorphology. Homo 2007;58:329–56.
22. Polder BJ, Vant Hof MA, Van der Linden FP, Kuijpers-Jagtman AM. A meta-analysis of the prevalence of dentalagenesis of permanent teeth. Comm Dent Oral Epidemiol2004;32:217–26.
23. Vastardis H, Karimbux N, Guthua SW, Seidman JG,Seidman CE. A human MSX1 homeodomain missensemutation causes selective tooth agenesis. Nat Genet1996;13:417–21.
24. Stockton DW, Das P, Goldenberg M, D’Souza RN, Patel PI.Mutation of PAX9 is associated with oligodontia. Nat Genet2000;24:18–9.
25. Frazier-Bowers SA, Guo DC, Cavender A, Xue L, Evans B,King T, et al. A novel mutation in human PAX9 causes molaroligodontia. J Dent Res 2002;81:129–33.
26. Klein ML, Nieminen P, Lammi L, Niebuhr E, Kreiborg S.Novel mutation of the initiation codon of PAX9 causesoligodontia. J Dent Res 2005;84:43–7.
27. Kim JW, Simmer JP, Lin BP, Hu JC. Novel MSX1 frameshiftcauses autosomal-dominant oligodontia. J Dent Res2006;85:267–71.
28. Gerits A, Nieminen P, De Muynck S, Carels C. Exclusion ofcoding region mutations in MSX1, PAX9 and AXIN2 in eightpatients with severe oligodontia phenotype. Orthod CraniofacRes 2006;9:129–36.
29. Pawlowska E, Janik-Papis K, Wisniewska-Jarosinska M,Szczepanska J, Blasiak J. Mutations in the human homeoboxMSX1 gene in the congenital lack of permanent teeth.Tohoku J Exp Med 2009;217:307–12.
30. Pereira TV, Salzano FM, Mostowska A, Ruiz-Linares WR,Chies JA, Saavedra C, et al. Natural selection andmolecular evolution in primate PAX9 gene, a majordeterminant of tooth development. Proc Natl Acad Sci USA2006;103:5676–81.
31. Trimmell JB. A mutation of PAX9 associated with congenitallymissing teeth. MSc dissertation. Kansas City: University ofMissouri; 2004.
32. Kula K, Trimmell J, Lu Y, Briscoe P, Feng JQ. Tooth agenesisin a family and homozygous PAX9 mutation in exon 3: acase report. World J Orthod 2008;9:e55–61.
33. Xuan KF, Jin Y, Liu L, Yuan L, Wen F, Yang X, et al.Identification of a novel missense mutation of MSX1 gene inChinese family with autossomal dominant oligodontia. ArchOral Biol 2008;53:773–9.
34. Weir BS. Genetic data analysis II. Sunderland, Mass: Sinauer; 1996.35. Long JC, William RC, Urbanek M. An E-M algorithm and
testing strategy for multiple-locus haplotypes. Am J HumGenet 1995;56:799–810.