- institute of experimental medicine as cr
TRANSCRIPT

Cytotherapy, 2010; 12: 212–225
ISSN 1465-3249 print/ISSN 1477-2566 online © 2010 Informa UK Ltd. (Informa Healthcare, Taylor & Francis AS)DOI: 10.3109/14653240903440103
Co-transplantation of olfactory ensheathing glia and mesenchymal stromal cells does not have synergistic effects after spinal cord injury in the rat
TAKASHI AMEMORI1, PAVLA JENDELOVÁ1,2, KATERINA RUŽICKOVÁ1, DAVID ARBOLEDA1 & EVA SYKOVÁ1,2
1Institute of Experimental Medicine, Academy of Sciences of the Czech Republic, Prague, Czech Republic, and 2Department of Neuroscience and Center for Cell Therapy and Tissue Repair, Charles University, 2nd Medical Faculty, Prague, Czech Republic
Abstract Background aims. Olfactory ensheathing glia (OEG) and mesenchymal stromal cells (MSC) are suitable candidates for transplantation therapy of spinal cord injury (SCI). Both facilitate functional improvement after SCI by producing trophic factors and cytokines. In this study, the co-transplantation of both types of cells was studied to clarify their additive and/or synergistic effects on SCI. Methods. A balloon-induced compression lesion was used to produce SCI in rats. OEG, MSC or both OEG and MSC (3 × 105 cells of each cell type) were implanted by intraspinal injection 1 week after SCI. The effect of transplantation was assessed using behavioral, electrophysiologic and histologic methods. Results. Hindlimb function was examined with Basso, Beattie and Bresnahan (BBB) and Plantar tests. Improvement was found in all three groups of transplanted rats with different time–courses, but there was no signifi cant difference among the groups at the end of the experiment. Motor-evoked potentials after SCI decreased in amplitude from 7 mV to 10 µV. Linear regression analysis showed a modest recovery in amplitude following transplantation, but no change in the control rats. Histologic fi ndings showed that the white and gray matter were signifi cantly spared by transplantation after SCI. Conclusions. Functional improvement was achieved with transplantation of OEG and/or MSC, but the co-transplantation of OEG and MSC did not show synergistic effects. The poor migration of OEG and MSC might prevent their concerted action. Pre-treatment with a Rho antagonist and a combination of intraspinal and intravenous injection of the cells might be benefi cial for SCI therapy.
Key Words: mesenchymal stromal cells, olfactory ensheathing glia, spinal cord injury, transplantation
fi bronectin) in addition to adhesion molecules (5–7). OEG display phenotypes of both Schwann cells and astrocytes. They are genetically closer to Schwann cells than astrocytes, and astrocytes are more related to OEG than to oligodendrocytes (8). These char-acteristic features of OEG promote the ingrowth of axons from the peripheral nervous system (PNS) to the central nervous system (CNS) beyond the laminin-rich boundary established following SCI (9). Au et al. (10) considered that the production of secreted protein acidic rich in cysteine (SPARC) by OEG regulated laminin-1 production and allowed Schwann cells and astrocytes to interact, resulting in the re-entry of regenerating axons into the CNS.
MSC are multipotent progenitor cells that, in the case of severe tissue ischemia or damage, can be
Correspondence: Dr Takashi Amemori, PhD, Department of Neuroscience, Institute of Experimental Medicine, Academy of Sciences of the Czech Republic, Videnska 1083, 142 20, Prague 4, Czech Republic. E-mail: [email protected]
(Received 15 May 2009; accepted 21 October 2009)
Introduction
Cell therapy is a promising approach to restoring locomotor function after spinal cord injury (SCI). Both olfactory ensheathing glia (OEG) and mesen-chymal stromal cells (MSC) have been investigated extensively as promising candidates for cell therapy.
The implantation of OEG is known to minimize cavity and scar formation by regulating astrocytes and promoting angiogenesis and myelination (1,2). OEG decrease the levels of chondroitin sulfate pro-teoglycan (CSPG), which has inhibitory effects on growing axons following SCI (3,4). Thus OEG prob-ably create a permissive environment for spared and regenerating axons after injury by releasing growth factors (NGF, BDNF, GDNF and VEGF) and extra-cellular matrix molecules (collagen, laminin and

Co-transplantation of OEG and MSC 213
performed. A 2-french Fogarty catheter was inserted below T8, and the balloon was infl ated with 15 µL saline for 5 min at T8 (19,21). During the surgi-cal procedure, the body temperature of the animal was maintained at 37°C with a heating pad, and 3% isofl urane in air was administered at a fl ow rate of 0.3 L/min to prevent edema development as a result of low levels of anesthesia (22). Following the lesion-ing procedure, the rats were assisted in feeding and urination until they had recovered suffi ciently to perform these functions on their own. The animals received gentamicin sulfate (5 mg/kg) for 3 days to prevent post-operative infections.
OEG preparation and identifi cation
OEG were obtained from 4-week-old Wistar rats. The olfactory mucosa was scraped from both sides of the nasal septum. After incubation in Dispase II (Roche, Mannheim, Germany), the lamina pro-pria was separated mechanically from the epithelium and dissociated with collagenase type I (Sigma, St. Louis, MO, USA) (23). After a 24-h incubation in Dulbecco’s modifi ed Eagle medium (DMEM)/F12 with 10% fetal bovine serum, the cells were treated with cytosine-β-D-arabinofuranoside (5 × 10–5 M; Sigma) to eliminate rapidly dividing cells, such as fi broblasts, and expanded with bovine pituitary extract (100 µg/mL; Sigma) (24). To assess the purity of OEG cultures, cul-tured cells on glass slides were fi xed in 4% paraformal-dehyde, permeabilized in 0.1% Triton X-100, blocked in serum, and incubated with primary antibodies over-night at 4°C. Two antibodies were used to identify OEG: mouse anti-p75, low-affi nity nerve growth factor recep-tor (1:100; Chemicon, Temecula, CA, USA), and rabbit anti-S100 (1:200; Sigma). Alexa Fluor 488-conjugated goat anti-mouse IgG and Alexa Fluor 594-conjugated goat anti-rabbit IgG secondary antibodies (Sigma) were used to distinguish the two primary antibodies. Labeled cells were examined in a fl uorescent microscope (Zeiss Axioskop, Göttingen, Germany). More than 80% of cultured cells were p75/S100-positive.
MSC preparation
BM was extracted from the tibia and femur of 4-week-old Wistar rats by fl ushing with DMEM. BM cells were dissociated mechanically and plated in DMEM medium supplemented with 10% fetal bovine serum, 100 U/mL penicillin and 0.1 mg/mL streptomycin (Gibco). MSC were isolated through their adherence to plastic. After a 24-h incuba-tion, non-adherent cells, including hematopoietic stem cells, were removed by replacing the medium (19,25). MSC were used for transplantation after 6–10 passages.
attracted to the lesion site, where they can secrete cytokines (interleukins) and trophic factors that can exert an inhibitory infl uence on scarring and apopto-sis. They facilitate angiogenesis and neural regenera-tion (11). Mobilized from the bone marrow (BM), and sorted or expanded in culture, MSC can be delivered to the damaged site by direct or systemic application. Functional improvement resulting from MSC transplantation could be principally because of their neuroprotective and reconstructive infl uences at the lesion site, resulting from their secretion of various trophic factors such as cytokines and growth factors (12–14), by their down-regulation of cas-pase-3, which mediates apoptosis (15), and through their immunomodulatory and anti-apoptotic activity resulting from their production of galectin-1, angio-poietin-1, osteopontin and thrombospondin-1 and -2 (16,17), thus fostering functional recovery after SCI (18–20). Their easy accessibility and rapid expansion in culture are attractive characteristics from a thera-peutic point of view.
The co-transplantation of OEG and MSC may be benefi cial for SCI treatment through a combinatory approach: OEG can facilitate regeneration following SCI by providing guiding strands and molecules of the extracellular matrix for axonal ingrowth, while other cell types such as MSC can improve functional outcome by producing trophic factors and cytokines. In addition, OEG as well as MSC can be used in autologous transplantation protocols, and therefore no additional immunosuppression would be neces-sary. In this study, the effect of the transplantation of OEG, MSC and the co-transplantation of both OEG and MSC on behavioral, electrophysiologic and histologic outcomes after SCI was studied.
Methods
Adult male Wistar rats weighing 270–300 g were used in our experiments. The lesioned animals were divided into four groups. The fi rst group received both OEG and MSC (n � 21). The second and third groups were transplanted with only OEG (n � 22) or MSC (n � 23), respectively. The fourth group was injected with saline as a control (n � 23). All experiments were performed in accordance with the European Communities Council Directive of 24th of November 1986 (86/609/EEC) regarding the use of animals in research, and were approved by the Ethics Committee of the Institute of Experimental Medicine ASCR, Prague, Czech Republic.
SCI
Balloon compression was used to create an SCI. A small opening of the thoracic vertebra 10 (T10) was

214 T. Amemori et al.
control, n � 12). The rats were placed on a fl oor within a circular enclosure as described by Basso et al. (29). Their hindlimb joint movement, paw placement, weight support, forelimb–hindlimb co-ordination, etc., were evaluated using a 0–21-point scale.
Hindpaw withdrawal latency to a noxious thermal stimuli was assessed with a Plantar test apparatus (Ugo Basile, Comerio, Italy) (OEG/MSC, n � 10; OEG, n � 10; MSC, n � 10; control, n � 10). The animals were placed in a clear plastic chamber and acclimated for 10 min until becoming quiescent. The hindpaw received a heat stimulus through a glass plate. The withdrawal latencies were measured three times for each hindpaw at 5-min intervals. The average of three values was used for statistical evaluation.
Motor-evoked potentials
Rats were anesthetized with ketamine (100 mg/kg). One pair of needle electrodes was placed subcu-taneously over the skull for cortical stimulation and another pair in the gastrocnemius muscle for recording. The signals were fi ltered (1–5000 Hz) and recorded by a Medelec Synergy T-EP EMG/EP Monitoring System (Oxford Instruments Medical Inc., Surrey, UK). Motor-evoked potentials (MEP) were elicited once a week by transcranial stimulation (a single square-pulse electrical stimulus of 0.1 ms in duration) to measure the amplitude, latency to onset and duration of the response (OEG/MSC, n � 9; OEG, n � 10; MSC, n � 11; control, n � 7).
Histologic analysis
To count the number of transplanted GFP cells and examine the possible neural differentiation of the transplanted cells, longitudinal cryostat sections (40-µm thickness) of the spinal cord were made after 4% paraformaldehyde fi xation. Every sixth section was chosen for imaging using a fl uorescent micro-scope. The surviving cells were counted in each sec-tion, and the total number of cells was estimated according to the volume of the section in which the cells were found. The percentage of surviving trans-planted cells was calculated by dividing the estimated total number of surviving cells by the total number of transplanted cells (3 × 105 cells/rat). For immu-nohistochemistry, the sections were blocked with 1% bovine serum albumin, then incubated with anti-bodies against neural markers, anti-nestin (1:2500; Chemicon) or anti-βIII-tubulin (1:400; Sigma) overnight at 4°C, followed by incubation with the Alexa Fluor 594-conjugated secondary antibody. To assess spared descending axon tracts after SCI, the
Transplantation
The animals were transplanted 1 week after SCI. Just before transplantation, OEG were labeled with the fl uo-rescent dye 10 µM carboxy-fl uorescein diacetate succin-imidyl ester (CFDA-SE; green fl uorescence; Molecular Probes, Eugene, OR, USA), while MSC were labeled with 20 µM carboxy SNARF-1 (red fl uorescence; Molecular Probes) or CFDA-SE, to enable subsequent identifi cation of transplanted cells. Labeling was performed according to the manufacturer’s instructions, and the concentra-tions of the tracers were based on the results of Imaizumi et al. (26) and Li et al. (27). The animals were secured in a stereotaxic apparatus by holding the spinal process just anterior to the injured site of the spinal cord with a rat-specifi c vertebra holder (Cunningham spinal adaptor; Stoelting Co., Wood Dale, IL, USA). The spinal cord was exposed at T8. A total of 3 × 105 OEG and/or MSC was injected through a glass pipette at a concentration of 1 × 105 cells/µL, into the proximal, central and dis-tal parts of the lesioned spinal cord (each part received 1 µL cell suspension), at a depth of 1 mm below the dorsal surface and a rate of 1 µL/min using a Nano-Injector (Stoelting Co.); OEG/MSC-transplanted ani-mals received six injections (3 × 105 OEG and 3 × 105
MSC) instead of the three injections received by the other animals. The glass pipette was kept in place after each injection for a further minute to prevent leakage of the cell suspension. The control group received three injections of saline (1 µL/injection), also into the proxi-mal, central and distal parts of the lesioned spinal cord. Cyclosporine (10 mg/kg) and ampicillin (50–100 mg/kg) were administered intraperitoneally 1 day before transplantation and throughout the experiment (for 2 months).
OEG and MSC from green fl uorescent protein transgenic rats
To enable more easily distinguishable and stable tracking of transplanted cells, OEG and MSC expressing enhanced green fl uorescent protein (GFP) isolated from GFP+ rats were used in 2-month survival experiments. Transgenic Sprague–Dawley rats (SD-Tg(CAG-EGFP)CZ-004Osb) were kindly provided by Dr Masaru Okabe (Osaka Univer-sity, Osaka, Japan) (28), bred at the laboratory of Dr Martin Marsala (University of California, San Diego, CA, USA) and subsequently sent to our institute and bred in our animal facility.
Functional analysis
Hindlimb locomotor activity after SCI was assessed by the Basso, Beattie and Bresnahan (BBB) test (29) (OEG/MSC, n � 14; OEG, n � 15; MSC, n � 16;

Co-transplantation of OEG and MSC 215
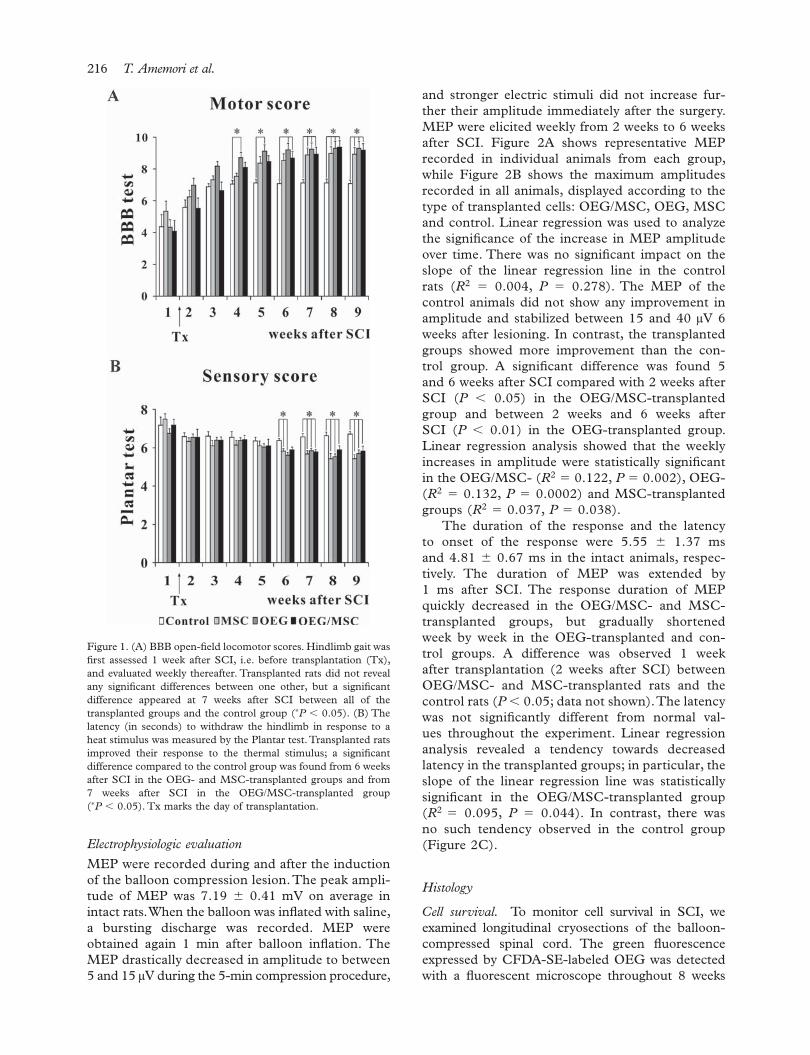
of the three joints in the hindlimb started to regain their ability to move during the fi rst post-surgical week, but the animals never placed their paws on the ground. The control animals achieved BBB scores of 7.08 � 0.24 at the end of the experiment (9 weeks after SCI, 8 weeks after transplantation) but never supported their body weight on their hind legs. During the fi rst week after transplantation, the transplanted animals already showed a tendency to improve their BBB scores. Among the transplanted groups, the fastest recovery was observed in animals transplanted with OEG. These animals showed signifi cant improvement compared with the controls as early as 4 weeks after SCI (P � 0.05) and their fi nal BBB score was 9.30 � 0.46. Animals with OEG and MSC co-grafts, even though they received six injections, showed a statistically signifi cant improve ment 6 weeks after SCI, with a fi nal BBB score of 9.18 � 0.44. One week later (i.e. 7 weeks after SCI), similar motor improvement was observed in MSC-transplanted rats (fi nal BBB score of 8.94 � 0.42) (Figure 1A). All transplanted animals recovered their gait; some of them gained the ability to support their body weight and to perform paw stepping, and two animals from the OEG- and MSC-transplanted groups showed co-ordination of the forelimbs and hindlimbs. However, in conclusion there were no signifi cant differences among any of the transplanted animal groups in terms of overall functional improve ment at the end of the experiment (P � 0.05).
Plantar test
The withdrawal latency of the hindpaws in response to a thermal stimulus was measured once a week. During the fi rst post-operative week (before trans plantation), the average latency of hindpaw withdrawal, determined from three repeated mea-surements at 5-min intervals, was 7.18 � 0.30, 6.75 � 0.24, 7.51 � 0.27 and 7.18 � 0.44 s, in OEG/MSC-, OEG- and MSC-transplanted rats and control rats, respectively. The latency of each group slightly decreased during the second week follow-ing lesion induction. The control group did not continue to improve further, but the transplanted groups shortened their withdrawal latency during each subsequent week. Signifi cant differences from the control rats (P � 0.05) started to appear in the transplanted rats during the sixth post-surgical week. The fi nal values determined 9 weeks after SCI were 5.83 � 0.25, 5.69 � 0.21 and 5.42 � 0.22 s in OEG/MSC-, OEG- and MSC-transplanted rats, respec-tively (Figure 1B); these values were signifi cantly dif-ferent (P � 0.05) from those of the control group (6.72 � 0.12 s). Hyperalgesia was never observed after OEG and/or MSC transplantation.
corticospinal tract, one of the major descending tracts in the spinal cord, was anterogradely labeled. Two weeks before termination of the experiment, 1 µL of a 10% biotinylated dextran amine (BDA; 10 000 MW; Molecular Probes) was injected bilaterally into the sensorimotor cortex at a depth of 1.5 mm from the surface with a glass micropipette. BDA-injected ani-mals were deeply anaesthetized with ketamine (100 mg/kg) and xylazine (20 mg/kg), their chests opened and transcardial perfusion performed with phosphate buffer followed by 4% paraformaldehyde solution in phosphate buffer. The spinal column was removed and left in paraformaldehyde solution overnight. The spinal cord was then carefully removed from the spi-nal cord channel and post-fi xed in the same solution for at least 1 week. An approximately 5-cm long seg-ment from the same area of each spinal cord was dissected, and 14-µm thick sagittal frozen sections were cut. To visualize the BDA-injected corticospinal tract axons, cryostat sections were incubated with avidin horseradish peroxidase and diaminobenzidine (DAB), following the instructions in the BDA neu-ronal tracer kit (Molecular Probes). Some sections were stained with hematoxylin-eosin.
Spinal compression results in the formation of cavi-ties within the spinal cord tissue and induces degen-eration of the spinal cord. The volume of the spared white and gray matter was analyzed at the end of the experiment. After perfusion with 4% paraformaldehyde, a 2-cm long segment of the spinal cord was dissected between 1-cm cranial and 1-cm caudal to the injury epicenter. Serial cross-sections (5 µm thick) were made after paraffi n embedding and stained with Luxol-fast blue and Cresyl violet to distinguish the white and gray matter. For volumetric measurements, six sections were selected at 1-mm intervals along the cranio-caudal axis, and whole images of the spinal cord were taken with an Axioskop 2 plus microscope (Zeiss) and analyzed by ImageJ software (Wayne Rasband, National Institutes of Health, Bethesda, MD, USA) (19).
Statistical analysis
The data are expressed as mean � SEM. A one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test was used to evaluate differences between groups. Regression analyzes were applied for MEP. A P-value �0.05 was considered statistically signifi cant.
Results
Functional outcome
BBB test. Hindlimb locomotion was examined weekly after SCI using an open-fi eld BBB test. The animals had a BBB score of 0 immediately after the induction of the balloon compression lesion. Some
216 T. Amemori et al.
and stronger electric stimuli did not increase fur-ther their amplitude immediately after the surgery. MEP were elicited weekly from 2 weeks to 6 weeks after SCI. Figure 2A shows representative MEP recorded in individual animals from each group, while Figure 2B shows the maximum amplitudes recorded in all animals, displayed according to the type of transplanted cells: OEG/MSC, OEG, MSC and control. Linear regression was used to analyze the signifi cance of the increase in MEP amplitude over time. There was no signifi cant impact on the slope of the linear regression line in the control rats (R2 � 0.004, P � 0.278). The MEP of the control animals did not show any improvement in amplitude and stabilized between 15 and 40 µV 6 weeks after lesioning. In contrast, the transplanted groups showed more improvement than the con-trol group. A signifi cant difference was found 5 and 6 weeks after SCI compared with 2 weeks after SCI (P � 0.05) in the OEG/MSC-transplanted group and between 2 weeks and 6 weeks after SCI (P � 0.01) in the OEG-transplanted group. Linear regression analysis showed that the weekly increases in amplitude were statistically signifi cant in the OEG/MSC- (R2 � 0.122, P � 0.002), OEG- (R2 � 0.132, P � 0.0002) and MSC-transplanted groups (R2 � 0.037, P � 0.038).
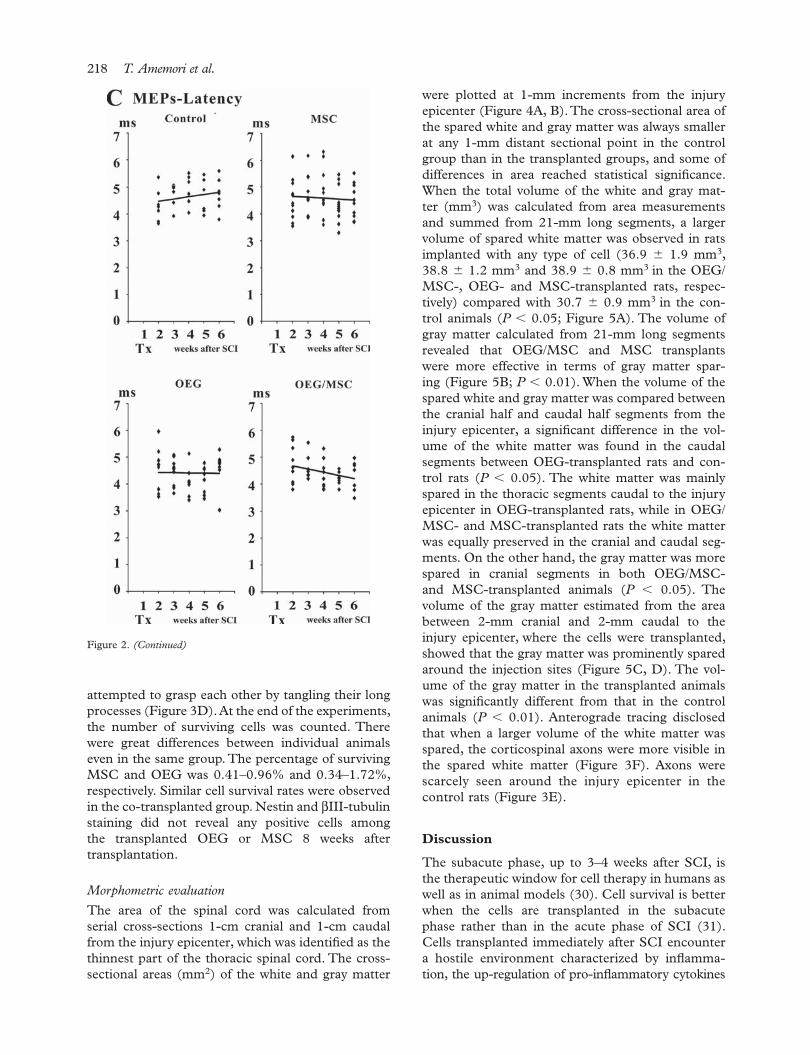
The duration of the response and the latency to onset of the response were 5.55 � 1.37 ms and 4.81 � 0.67 ms in the intact animals, respec-tively. The duration of MEP was extended by 1 ms after SCI. The response duration of MEP quickly decreased in the OEG/MSC- and MSC-transplanted groups, but gradually shortened week by week in the OEG-transplanted and con-trol groups. A difference was observed 1 week after transplantation (2 weeks after SCI) between OEG/MSC- and MSC-transplanted rats and the control rats (P � 0.05; data not shown). The latency was not signifi cantly different from normal val-ues throughout the experiment. Linear regression analysis revealed a tendency towards decreased latency in the transplanted groups; in particular, the slope of the linear regression line was statistically signifi cant in the OEG/MSC-transplanted group (R2 � 0.095, P � 0.044). In contrast, there was no such tendency observed in the control group (Figure 2C).
Histology
Cell survival. To monitor cell survival in SCI, we examined longitudinal cryosections of the balloon-compressed spinal cord. The green fl uorescence expressed by CFDA-SE-labeled OEG was detected with a fl uorescent microscope throughout 8 weeks
Electrophysiologic evaluation
MEP were recorded during and after the induction of the balloon compression lesion. The peak ampli-tude of MEP was 7.19 � 0.41 mV on average in intact rats. When the balloon was infl ated with saline, a bursting discharge was recorded. MEP were obtained again 1 min after balloon infl ation. The MEP drastically decreased in amplitude to between 5 and 15 µV during the 5-min compression procedure,
Figure 1. (A) BBB open-fi eld locomotor scores. Hindlimb gait was fi rst assessed 1 week after SCI, i.e. before transplantation (Tx), and evaluated weekly thereafter. Transplanted rats did not reveal any signifi cant differences between one other, but a signifi cant difference appeared at 7 weeks after SCI between all of the transplanted groups and the control group (∗P � 0.05). (B) The latency (in seconds) to withdraw the hindlimb in response to a heat stimulus was measured by the Plantar test. Transplanted rats improved their response to the thermal stimulus; a signifi cant difference compared to the control group was found from 6 weeks after SCI in the OEG- and MSC-transplanted groups and from 7 weeks after SCI in the OEG/MSC-transplanted group (∗P � 0.05). Tx marks the day of transplantation.

Co-transplantation of OEG and MSC 217
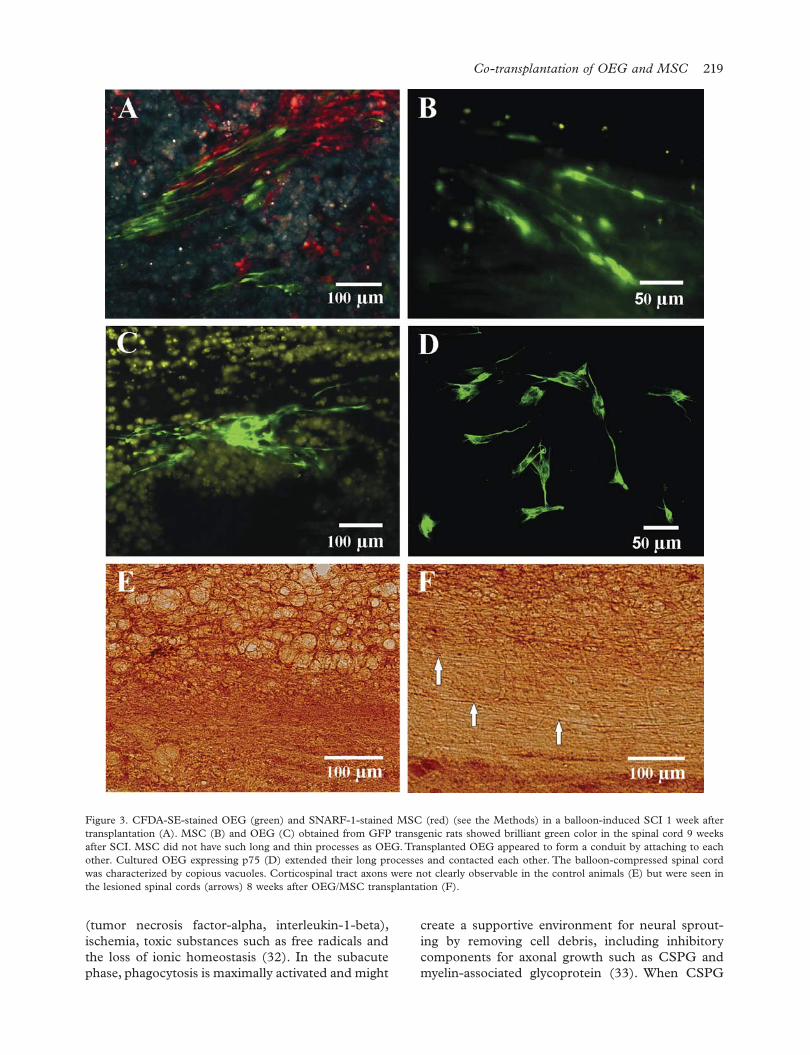
by their vivid green color in the spinal cord for 8 weeks. None of the studied cell types showed any extensive migration within the spinal cord tissue, as they were almost exclusively found around the injection sites or were slightly diffused by the injection pressure. MSC spread around the injection sites after transplantation, while OEG extended their long processes and remained in close contact with one another. It appeared as if OEG formed conduits, as they do in the lamina propria in situ to ensheath and guide the olfactory axons (Figure 3C). Such dynamic behavior of OEG was also observed in vitro: OEG
of observation after transplantation. However, SNARF-1- or CFDA-SE-labeled MSC were visible for only 4 or 5 weeks after SCI (Figure 3A); subsequently, labeled MSC were not seen. It seemed likely that the concentration of the dye was not adequate for longer observation. Therefore, to enable more reliable tracking of the cells in vivo, OEG and MSC obtained from GFP transgenic rats were used for long-term tracking. Transplants of both types of cells were injected and checked every week in the same way. Surviving MSC (Figure 3B) and OEG (Figure 3C) from GFP rats were easily recognized
Figure 2. Representative MEP recorded in transplanted and control rats 6 weeks after SCI (A). The maximum amplitudes (µV) (B) and the latency (ms) (C) obtained from individual animals are shown as a scatterplot at 1-week intervals following transplantation or saline injection (Tx). The linear regression lines are shown as solid lines.
218 T. Amemori et al.
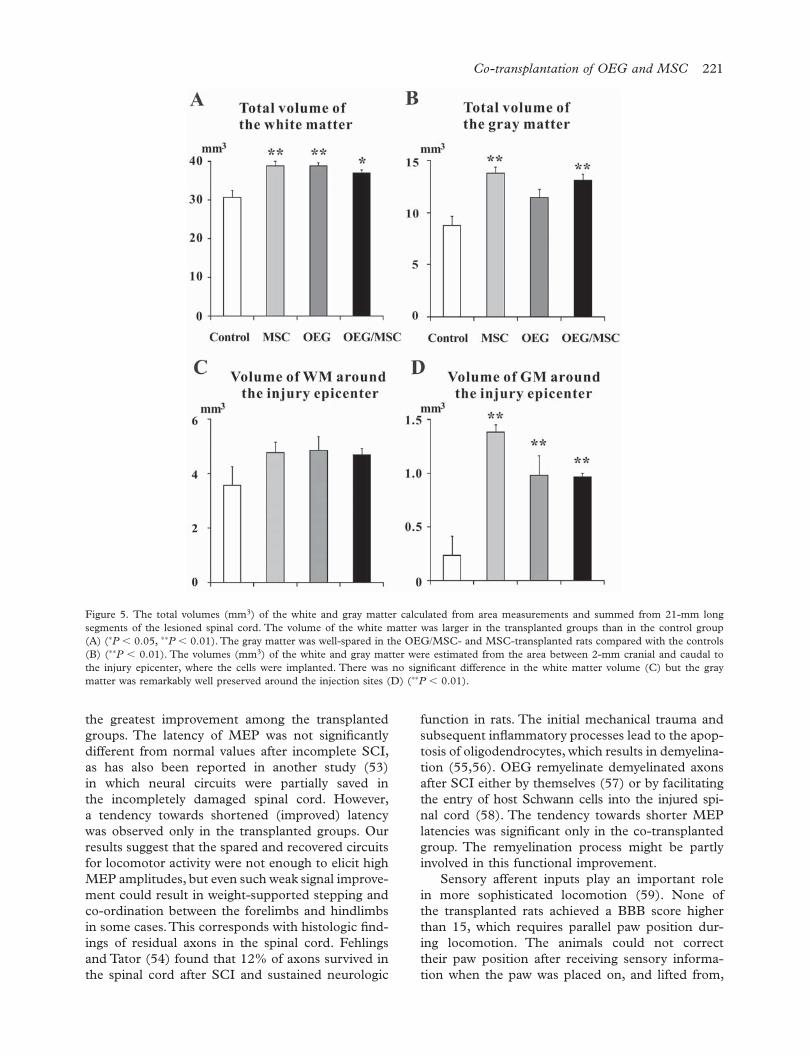
were plotted at 1-mm increments from the injury epicenter (Figure 4A, B). The cross-sectional area of the spared white and gray matter was always smaller at any 1-mm distant sectional point in the control group than in the transplanted groups, and some of differences in area reached statistical signifi cance. When the total volume of the white and gray mat-ter (mm3) was calculated from area measurements and summed from 21-mm long segments, a larger volume of spared white matter was observed in rats implanted with any type of cell (36.9 � 1.9 mm3, 38.8 � 1.2 mm3 and 38.9 � 0.8 mm3 in the OEG/MSC-, OEG- and MSC-transplanted rats, respec-tively) compared with 30.7 � 0.9 mm3 in the con-trol animals (P � 0.05; Figure 5A). The volume of gray matter calculated from 21-mm long segments revealed that OEG/MSC and MSC transplants were more effective in terms of gray matter spar-ing (Figure 5B; P � 0.01). When the volume of the spared white and gray matter was compared between the cranial half and caudal half segments from the injury epicenter, a signifi cant difference in the vol-ume of the white matter was found in the caudal segments between OEG-transplanted rats and con-trol rats (P � 0.05). The white matter was mainly spared in the thoracic segments caudal to the injury epicenter in OEG-transplanted rats, while in OEG/MSC- and MSC-transplanted rats the white matter was equally preserved in the cranial and caudal seg-ments. On the other hand, the gray matter was more spared in cranial segments in both OEG/MSC- and MSC-transplanted animals (P � 0.05). The volume of the gray matter estimated from the area between 2-mm cranial and 2-mm caudal to the injury epicenter, where the cells were transplanted, showed that the gray matter was prominently spared around the injection sites (Figure 5C, D). The vol-ume of the gray matter in the transplanted animals was signifi cantly different from that in the control animals (P � 0.01). Anterograde tracing disclosed that when a larger volume of the white matter was spared, the corticospinal axons were more visible in the spared white matter (Figure 3F). Axons were scarcely seen around the injury epicenter in the control rats (Figure 3E).
Discussion
The subacute phase, up to 3–4 weeks after SCI, is the therapeutic window for cell therapy in humans as well as in animal models (30). Cell survival is better when the cells are transplanted in the subacute phase rather than in the acute phase of SCI (31). Cells transplanted immediately after SCI encounter a hostile environment characterized by infl amma-tion, the up-regulation of pro-infl ammatory cytokines
attempted to grasp each other by tangling their long processes (Figure 3D). At the end of the experiments, the number of surviving cells was counted. There were great differences between individual animals even in the same group. The percentage of surviving MSC and OEG was 0.41–0.96% and 0.34–1.72%, respectively. Similar cell survival rates were observed in the co-transplanted group. Nestin and βIII-tubulin staining did not reveal any positive cells among the transplanted OEG or MSC 8 weeks after transplantation.
Morphometric evaluation
The area of the spinal cord was calculated from serial cross-sections 1-cm cranial and 1-cm caudal from the injury epicenter, which was identifi ed as the thinnest part of the thoracic spinal cord. The cross-sectional areas (mm2) of the white and gray matter
Figure 2. (Continued)
Co-transplantation of OEG and MSC 219
create a supportive environment for neural sprout-ing by removing cell debris, including inhibitory components for axonal growth such as CSPG and myelin-associated glycoprotein (33). When CSPG
(tumor necrosis factor-alpha, interleukin-1-beta), ischemia, toxic substances such as free radicals and the loss of ionic homeostasis (32). In the subacute phase, phagocytosis is maximally activated and might
Figure 3. CFDA-SE-stained OEG (green) and SNARF-1-stained MSC (red) (see the Methods) in a balloon-induced SCI 1 week after transplantation (A). MSC (B) and OEG (C) obtained from GFP transgenic rats showed brilliant green color in the spinal cord 9 weeks after SCI. MSC did not have such long and thin processes as OEG. Transplanted OEG appeared to form a conduit by attaching to each other. Cultured OEG expressing p75 (D) extended their long processes and contacted each other. The balloon-compressed spinal cord was characterized by copious vacuoles. Corticospinal tract axons were not clearly observable in the control animals (E) but were seen in the lesioned spinal cords (arrows) 8 weeks after OEG/MSC transplantation (F).

220 T. Amemori et al.
Rho antagonist or a Rho kinase inhibitor might be benefi cial for the co-transplantation of OEG and MSC to facilitate cell migration as well as promote axonal growth and prevent apoptosis in the injured spinal cord (40,45).
To treat SCI, cell therapy is used to ameliorate a hostile niche for neuronal sparing and regenera-tion, including the prevention of scar formation, inhibitory factors and demyelination, and to establish new neural circuits to bypass the injured sites. After SCI, the severed axons sprout and send collaterals into the gray matter (46–48) and new circuits are mediated by propriospinal neurons in the incom-pletely damaged spinal cord (49,50). Functional recovery is very much dependent on this newly established route. The MEP recorded once a week showed very variable amplitudes in each group and even in the same animal. The positions of the elec-trode for recording were not exactly the same for every measurement. Therefore, permanent implan-tation of electrodes into the muscle (51) or spinal cord (52) might be benefi cial for periodic recordings during a long-term experiment if they do not affect the animal’s locomotion and behavior. Even though the MEP had variable amplitudes and remained within microvolt-levels, the trend of their increasing amplitude, analyzed during a 1-month observation period, was statistically signifi cant in the trans-planted groups. The control group did not show any improvement in amplitude, but the transplanted groups improved their amplitude week by week; in particular, the OEG-transplanted group showed
production is blocked immediately after SCI, spon-taneous recovery is decreased. CSPG seems to exert a benefi cial effect on the injured spinal cord in the acute phase by activating microglia and macrophages (34). However, the inhibitory regulation of microglial activity is required in the later phase of SCI because neural cell damage proceeds as a result of cytokines and free radicals released from proliferating micro-glia (35). Thus OEG and MSC were transplanted at 7 days after SCI in our study.
In agreement with previous studies (19), our results show that hindlimb locomotion improved in OEG- and MSC-transplanted rats. However, their co-transplantation did not yield further additive and synergistic functional improvements. OEG and MSC appear to have a propensity to migrate towards an injury site (36–38). The GFP OEG and MSC transplants were predominantly observed around the injection sites. In our experiments, OEG and MSC were injected inside the lesions, so they would not need to migrate any considerable distance. On the other hand, their poor migration might be one of the features of OEG obtained from the lamina propria (39). Su et al. (40) suggested that post-injury up-regulation of Nogo-A on oligodendrocytes enhanced the adhesion of OEG via activation of Rho-A and limited their migration. The increased activation of Rho after SCI (41,42) also induced the formation of MSC focal adhesions (43,44). Poor cell migra-tion might interfere with the concerted actions of OEG and MSC, which were implanted by separate injections in our experiments. Pre-treatment with a
Figure 4. Cross-sectional areas (mm2) of the white (A) and gray (B) matter plotted at 1-mm increments from the injury epicenter, which is shown as 0. Numbers indicate the distance from the injury epicenter in millimeters. The spared areas are indicated according to the type of transplanted cell.
Co-transplantation of OEG and MSC 221
function in rats. The initial mechanical trauma and subsequent infl ammatory processes lead to the apop-tosis of oligodendrocytes, which results in demyelina-tion (55,56). OEG remyelinate demyelinated axons after SCI either by themselves (57) or by facilitating the entry of host Schwann cells into the injured spi-nal cord (58). The tendency towards shorter MEP latencies was signifi cant only in the co-transplanted group. The remyelination process might be partly involved in this functional improvement.
Sensory afferent inputs play an important role in more sophisticated locomotion (59). None of the transplanted rats achieved a BBB score higher than 15, which requires parallel paw position dur-ing locomotion. The animals could not correct their paw position after receiving sensory informa-tion when the paw was placed on, and lifted from,
the greatest improvement among the transplanted groups. The latency of MEP was not signifi cantly different from normal values after incomplete SCI, as has also been reported in another study (53) in which neural circuits were partially saved in the incompletely damaged spinal cord. However, a tendency towards shortened (improved) latency was observed only in the transplanted groups. Our results suggest that the spared and recovered circuits for locomotor activity were not enough to elicit high MEP amplitudes, but even such weak signal improve-ment could result in weight-supported stepping and co-ordination between the forelimbs and hindlimbs in some cases. This corresponds with histologic fi nd-ings of residual axons in the spinal cord. Fehlings and Tator (54) found that 12% of axons survived in the spinal cord after SCI and sustained neurologic
Figure 5. The total volumes (mm3) of the white and gray matter calculated from area measurements and summed from 21-mm long segments of the lesioned spinal cord. The volume of the white matter was larger in the transplanted groups than in the control group (A) (∗P � 0.05, ∗∗P � 0.01). The gray matter was well-spared in the OEG/MSC- and MSC-transplanted rats compared with the controls (B) (∗∗P � 0.01). The volumes (mm3) of the white and gray matter were estimated from the area between 2-mm cranial and caudal to the injury epicenter, where the cells were implanted. There was no signifi cant difference in the white matter volume (C) but the gray matter was remarkably well preserved around the injection sites (D) (∗∗P � 0.01).

222 T. Amemori et al.
spinal cord cranial to the injury epicenter. In other words, the gray matter sparing improves with the aid of MSC transplantation, while the white matter spar-ing resulting from OEG transplantation is likely to pro-tect spinal tract axons from degeneration during the early stages of SCI, which in turn might lead to the faster recovery of simple locomotor function. Moreover, analy zing the volume between 2-mm cranial and 2-mm caudal to the injury epicenter, where the OEG and/or MSC were mainly implanted, there was no statistical difference in the volume of the white matter between the transplanted and control groups. In contrast, a sig-nifi cant difference was found in the volume of the gray matter. OEG and MSC had the ability to spare the white and gray matter in the injured spinal cord; in par-ticular, the gray matter was well preserved around the damaged area of the spinal cord where the transplants were directly injected. The gray matter is more suscep-tible to metabolic damage than white matter. Balloon compression includes ischemic damage to the spinal cord and leads to metabolic failure in the gray matter. We showed that the transplants spared the gray matter from such damage. Without OEG and/or MSC treat-ment, the gray matter scarcely remained and the white matter was poorly spared. OEG and MSC appear to secure more descending and ascending axons by spar-ing the white matter. Therefore, they can provide more opportunities to reconstruct lost connections by sending collaterals from the spared axons to the gray matter, which was also spared by the transplants. In the gray matter, interneurons can re-establish or maintain neural connections with the PNS and CNS. If OEG and/or MSC were not implanted, the animals never showed the ability to support their body weight on their hind legs. However, we could not fi nd any unambiguous rela-tionship between the volume of the spared spinal cord tissue and behavioral improvement. Functional recov-ery must be considered from the viewpoint of synaptic reconstruction, and the white and gray matter sparing induced by transplantation increases the feasibility of this reconstruction.
Although MSC have the multipotency to differen-tiate into mesodermal and non-mesodermal lineages (64), they are prone to differentiate into the meso-dermal lineage. Their potential to differentiate into neuronal cells is limited and remains controversial (65,66). In our experiment, nestin-positive and βIII-tubulin-positive transplanted cells were not found 2 months after transplantation. Although we cannot dismiss the possibility of neural differentiation in the transplanted cells, which might help to bridge the damaged neural circuits, the trophic effects of the transplanted cells on the resident neurons are more likely to affect functional improvement positively after neural damage than the possible differentiation of the transplanted cells (67).
the fl oor. Our results did not show the same clear improvement in the Plantar test as was observed in a previous study where MSC were implanted intra-venously (19). Because intraspinal injection may cause additional damage to the dorsal columns, the results from the Plantar test may not be as reliable in the present study as in our previous MSC study. Although the transplantation of both types of cells showed similar results, their mechanisms of action are different in restoring the damaged spinal cord. It has been shown that intravenous injection of MSC 7 days after SCI improves locomotor activity and sig-nifi cantly increases spared white matter in the lesion area (19). However, no sparing of gray matter was observed. MSC are known to have immunomodula-tory effects (60). Therefore, their systemic injection may infl uence the levels of pro- and anti-infl amma-tory cytokines, and this might result in a trophic effect on functional recovery. Also, the lesion is not fully developed at 7 days after SCI, so the MSC may act as a neuroprotective agent. The slow improve-ment observed in the Plantar test might therefore be because of these different routes of cell injection: intraspinal versus intravenous.
Cell counts showed the poor survival of OEG and MSC implanted into the lesioned area. The direct injection of each cell type spared the gray matter in our study, but the injury epicenter is not a habitable environment for transplanted cells (31). Subarachnoid injection is an alternate and promising route for cell administration in order to avoid additional damage to the spinal cord and enable more effi cient cell delivery to the spinal cord (61,62), but further study is required to elucidate the environment in the cerebrospinal fl uid (CSF) for transplanted cells. MSC injected into the fourth ventricle were subsequently transported to the subarachnoid space in the spinal cord through the CSF and promoted behavioral recovery after SCI, but these MSC had disappeared by 3 weeks after injection, even using an immunosuppressant (63). Repeated suba-rachnoid administration of cells might be effective in promoting the long-term effects of the transplanted cells on the damaged host tissue.
Vanicky et al. (21) showed that balloon compression leads to a complete loss of gray matter, a partial loss of white matter and copious vacuoles. Our morphometric analysis also revealed that the cross-sectional area of the gray matter around the injury epicenter was 0–0.013 mm2 in the control animals. However, the gray mat-ter always remained spared in the treated animals. The white matter was equally spared in the lesioned spinal cord with the OEG/MSC and MSC transplants, while OEG transplants preserved the white matter in the cau-dal segments more than in the cranial segments. The gray matter was spared to a greater extent in the MSC/OEG- and MSC-transplanted rats, especially in the

Co-transplantation of OEG and MSC 223
Lakatos A, Barnett, SC, Franklin JM. Olfactory ensheathing 4. cells induce less host astrocyte response and chondroitin sul-phate proteoglycan expression than Schwann cells following transplantation into adult CNS white matter. Exp Neurol. 2003;184:237–46.Ramon-Cueto A, Avila J. Olfactory ensheathing glia: proper-5. ties and function. Brain Res Bull. 1998;46:175–87.Woodhall E, West AK, Chuah MI. Cultured olfactory 6. ensheathing cells express nerve growth factor, brain-derived neurotrophic factor, glia cell line-derived neurotrophic factor and their receptors. Mol Brain Res. 2001;88:203–13.Au E, Roskams AJ. Olfactory ensheathing cells of the lamina 7. propria in vivo and in vitro. Glia. 2003;41:224–36.Rojas-Mayorquin AE, Torres-Ruiz NM, Orituno-Sahagun D, 8. Gudino-Cabrera G. Microarray analysis of striatal embry-onic stem cells induced to differentiate by ensheathing cell conditioned media. Dev Dyn. 2008;237:979–94.Ramon-Cueto A, Nieto-Sampedro M. Regeneration into 9. the spinal cord of transected dorsal root axons is promoted by ensheathing glia transplants. Exp Neurol. 1994;127:232–44.Au E, Richter MW, Tetzlaff W, Aebersold R, Sage EH, 10. Roskams AJ. SPARC from olfactory ensheathing cells stimu-lates Schwann cells to promote neurite outgrowth and enhances spinal cord repair. J Neurosci. 2007;27:7208–21.Caplan AI. Adult mesenchymal stem cells for tissue engi-11. neering versus regenerative medicine. J Cell Physiol. 2007;213:341–7.Caplan AI, Dennis JE. Mesenchymal stem cells as trophic 12. mediators. J Cell Biochem. 2006;93:1076–84.Opydo-Canek M. Bone marrow stromal cells in traumatic 13. brain injury therapy: true perspective or false hope? Acta Neurobiol Exp. 2007;67:187–95.Chen L, Tredget EE, Wu PYG, Wu Y. Paracrine factors of 14. mesenchymal stem cells recruit macrophages and endothe-lial lineage cells and enhance wound healing. PloS One. 2008;3:e1886.Dasari VR, Spomar DG, Cady C, Gujrati M, Rao JS, 15. Dinh DH. Mesenchymal stem cells from rat bone marrow downregulate caspase-3-mediated apoptotic pathway after spinal cord injury in rats. Neurochem Res. 2007;32:2080–93.Ucelli A, Moretta L, Pistoia V. Immunoregulatory function of 16. mesenchymal stem cells. Eur J Immunol. 2006;36:2566–73.Pedemonte E, Benvenuto F, Casazza S, Mancardi G, 17. Oksenberg JR, Uccelli A, et al. The molecular signature of therapeutic mesenchymal stem cells exposes the architec-ture of the hematopoietic stem cell niche synapse. BMC Genomics. 2007;8:65.Sykova E, Jendelova P. In vivo tracking of stem cells in brain 18. and spinal cord injury. Prog Brain Res. 2007;161:367–83.Urdzikova L, Jendelova P, Glogarova K, Burian M, Hajek M, 19. Sykova E. Transplantation of bone marrow stem cells as well as mobilization by granulocyte-colony stimulating factor promotes recovery after spinal cord injury in rats. J Neurotrauma. 2006;23:1379–91.Hofstetter CP, Schwarz EJ, Hess D, Widenfalk J, Manira AE, 20. Prockop DJ, et al. Marrow stromal cells form guiding strands in the injured spinal cord and promote recovery. Proc Natl Acad Sci USA. 2002;99:2199–204.Vanicky I, Urdzikova L, Saganova K, Cizkova D, Galik J. A 21. simple and reproducible model of spinal cord injury induced by epidural balloon infl ation in the rat. J Neurotrauma. 2001;18:1399–407.Sedy J, Urdzikova L, Likavcanova K, Hejcl A, Jendelova P, 22. Sykova Y. A new model of severe neurogenic pulmonary edema in spinal cord injured rat. Neurosci Lett. 2007;423:167–71.
Thus OEG and MSC could foster locomotor function by sparing the spinal cord, but their com-bined advantages provide no added benefi t to SCI therapy. Additional treatment might be required to facilitate their migration, leading to co-operation of the two types of cells, or different combinations of transplants and/or trophic factors might need to be considered for further functional improvement. In a previous study, the intravenous injection of MSC spared the white matter after SCI, but not the gray matter (19). Comparing the spared vol-ume of white and gray matter between the intra-venous injection used in our previous study and the intraspinal injection of MSC used in the cur-rent study, we found signifi cantly more gray matter sparing following intraspinal injection. If the spared volume in the control animals is considered 100%, the spared volume of the white matter following intravenous injection was 150% and 134% follow-ing intra spinal injection, while the spared volume of the gray matter was 137% following intravenous injection and 220% following intraspinal injection. In addition, recovery in the BBB test was fastest dur-ing the fi rst few weeks post-grafting in those animals that received OEG transplants. Taken together, we speculate that to maximize the advantageous effects of co-transplantation, direct implantation of OEG into the spinal cord lesion accompanied by intrave-nous injection of MSC might be another, perhaps more effi cacious, approach to the co-transplantation of OEG and MSC.
Acknowledgments
We thank James Dutt for critical reading of the manu-script. This work was supported by the following grants: AVOZ50390703, IAA500390902, GACR309/06/1246, LC554, 1M0538 and 1A8697-5. The EU FP6 project ENINET (LSHM-CT-2005–019063.
Declaration of interest: The authors report no confl icts of interest. The authors alone are respon-sible for the content and writing of the paper.
References
Verdu E, Garcia-Alias G, Fores J, Gudino-Cabrera G, 1. Muneton VC, Nieto-Sampedro M, et al. Effects of ensheath-ing cells transplanted into photochemically damaged spinal cord. Neuroreport. 2001;12:2303–9.Ramer LM, Au E, Richter MW, Liu J, Tetzlaff W, Roskams AJ. 2. Peripheral olfactory ensheathing cells reduce scar and cavity formation and promote regeneration after spinal cord injury. J Comp Neurol. 2004;473:1–15.Takami T, Oudega M, Bates ML, Woo PM, Kleitman N, 3. Bunge MB. Schwann cell but not olfactory ensheathing glia transplants improve hindlimb locomotor performance in the moderately contused adult rat thoracic spinal cord. J Neurosci. 2002;22:6670–81.

224 T. Amemori et al.
Dubreuil CI, Winton MJ, McKerracher L. Rho activation 41. patterns after spinal cord injury and the role of activated Rho in apoptosis in the central nervous system. J Cell Biol. 2003;162:233–43.Madura T, Yamashita T, Kubo T. Activation of Rho in the 42. injured axons following spinal cord injury. EMBO Rep. 2004;5:412–7.Meriane M, Duhamel S, Lejeune L, Galipeau J, Annabi B, 43. et al. Cooperation of matrix metalloproteinases with the RhoA/Rho kinase and mitogen-activated protein kinase kinase-1/extracellular signal-regulated kinase signaling path-ways is required for the sphingosine-1-phosphate-induced mobilization of marrow-derived stromal cells. Stem Cells. 2006;24:2557–65.Jaganathan BG, Ruester B, Dressel L, Stein S, Grez M, 44. Seifried E, et al. Rho inhibition induces migration of mesen-chymal stromal cells. Stem Cells. 2007;25:1966–74.McKerracher L, Higuchi H. Targeting Rho to stimulate repair 45. after spinal cord injury. J Neurotrauma. 2006;23:309–17.Fouad K, Pedersen V, Schwab ME. Cervical sprouting of 46. corticospinal fi bers after thoracic spinal cord injury accom-panies shifts in evoked motor responses. Curr Biol. 2001;11:1766–70.Deumens R, Koopmans GC, Joosten EAJ. Regeneration of 47. descending axon tracts spinal cord injury. Prog Neurobiol. 2005;77:57–89.Steward O, Zheng B, Tessier-Lavigne M, Hofstdter M, 48. Sharp K, Yee KM. Regeneration beyond the glial scar. J Neurosci. 2008;28:6836–47.Bareyre FM, Kerschensteiner M, Raineteau O, Mettenleiter 49. TC, Weinmann O, Schwab ME. The injured spinal cord spontaneously forms a new intraspinal circuits in adult rats. Nat Neurosci. 2004;7:269–77.Courtine G, Song B, Roy RR, Zhong H, Hermann JE, Ao Y, 50. et al. Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat Med. 2008;14:69–74.Oria M, Chatauret N, Raguer N, Cordoba J. A new method 51. for measuring motor evoked potentials in the awake rat: effects of anesthetics. J Neurotrauma. 2008;25:266–75.Ondrejcak T, Vanicky I, Galik J, Saganova K. Chronically 52. implanted electrodes for repeated stimulation and recording of spinal cord potentials. J Neurosci Methods. 2005;141:125–34.Smith HC, Savic G, Frankel HL, Ellawa PH, Maskill DW, 53. Jamous MA, et al. Corticospinal function studied over time following incomplete spinal cord injury. Spinal Cord. 2000;38:292–300.Fehlings MG, Tator CH. The relationships among the 54. severity of spinal cord injury, residual neurological function, axon counts, and counts of retrogradely labeled neurons after experimental spinal cord injury. Exp Neurol. 1995;132:220–8.Totoiu MO, Keirstead HS. Spinal cord injury is accompa-55. nied by chronic progressive demyelination. J Comp Neurol. 2005;486:373–83.Siegenthaler MM, Tu MK, Keirstead HS. The extent of 56. myelin pathology differs following contusion and transection spinal cord injury. J Neurotrauma. 2007;24:1631–46.Franklin RJM. Remyelination by transplanted olfactory 57. ensheathing cells. Anat Rec B New Anat. 2003;271:71–6.Boyd JG, Doucette R, Kawaja MD. Defi ning the role of the 58. olfactory ensheathing cells in facilitating axon remyelina-tion following damage to the spinal cord. FASEB J. 2005;19:694–703.Raineteau O, Schwab ME. Plasticity of motor systems after 59. incomplete spinal cord injury. Nat Rev Neurosci. 2001;2:263–73.
Bianco JI, Perry C, Harkin DG, Mackay-Sim A, Feron F. 23. Neurotrophin 3 promotes purifi cation and proliferation of olfactory ensheathing cells from human nose. Glia. 2004;45:111–23.Woodhall E, West AK, Vickers AK, Chuah MI. Olfactory 24. ensheathing cell phenotype following implantation in the lesioned spinal cord. Cell Mol Life Sci. 2003;60:2241–53.Azizi SA, Stokes D, Augelli BJ, Digirolamo C, Prockop DJ. 25. Engraftment and migration of human bone marrow stromal cells implanted in the brains of albino rats: similarities to astro-cyte grafts. Proc Natl Acad Sci USA. 1998;95:3908–18.Imaizumi T, Lankford KL, Kocsis JD. Transplantation of 26. olfactory ensheathing cells or Schwann cells restores rapid and secure conduction across the transected spinal cord. Brain Res. 2000;854:70–8.Li X, Dancausse H, Grijalva I, Oliveira M, Levi DO. 27. Labeling Schwann cells with CFSE: an in vitro and in vivo study. J Neurosci Methods. 2003;125:83–91.Okabe M, Ikawa M, Kominami K, Nakanishi T, 28. Nishimune Y. Green mice as a source of ubiquitous green cells. FASEB Lett. 1977;407:313–9.Basso DM, Beattie MS, Bresnahan JC. A sensitive and reli-29. able locomotor rating scale for open fi eld testing in rats. J Neurotrauma. 1995;12:1–12.Sykova E, Homola A, Mazanec R, Lachman H, Konradova SL, 30. Kobylka P, et al. Autologous bone marrow transplantation in patients with subacute and chronic spinal cord injury. Cell Transplant. 2006;15:675–87.Parr AM, Kulbatski I, Tator CH. Transplantation of adult 31. rat spinal cord stem/progenitor cells for spinal cord injury. J Neurotrauma. 2007;24:835–45.Park E, Velumian AA, Fehlings MG. The role of excitatoxic-32. ity in secondary mechanisms of spinal cord injury: a review with an emphasis on the implications for white matter degen-eration. J Neurotrauma. 2004;21:754–74.Rowland JW, Hawryluk GWJ, Kwon B, Fehlings MG. 33. Current status of acute spinal cord injury pathophysiology and emerging therapies: promise on the horizon. Neurosurg Focus. 2008;25[5]:e2.Rolls A, Shechter R, Londong A, Segev Y, Jacob-Hirsch J, 34. Amariglio N, et al. Two faces of chondroitin sulfate prote-oglycan in spinal cord repair: a role in microglia/macrophage activation. PLoS Med. 2008;5:e171.Morino T, Ogata T, Horiuchi H, Takeba J, Okumura H, 35. Miyazaki T, et al. Delayed neuronal dagame related to micro-glia proliferation after mild spinal cord compression injury. Neurosci Res. 2003;46:309–18.Richter MW, Fletcher PA, Liu J, Tetzlaff W, Roskams AJ. 36. Lamina propria and olfactory bulb ensheathing cells exhibit differential integration and migration and promote differen-tial axon sprouting in the lesioned spinal cord. J Neurosci. 2005;25:10700–11.Uccelli A, Pistia V, Moretta L. Mesenchymal stem cells: a 37. new strategy for immunosuppression? Trends Immunol. 2007;28:219–26.Sordi V, Malosio ML, Marchesi, F, Mercalli A, Melzi R, 38. Giordano T, et al. Bone marrow mesenchymal cells express a restricted set of functionally active chemokine receptors capable of promoting migration to pancreatic islets. Blood 2005;106:419–27.Lu P, Yang H, Culbertson M, Graham L, Roskams AJ, 39. Tuszynski MH. Olfactory ensheathing cells do not exhibit unique migratory or axonal growth-promoting properties after spinal cord injury. J Neurosci. 2006;26:11120–30.Su Z, Cao L, Zhu Y, Liu X, Huang Z, Huang A, et al. Nogo 40. enhances the adhesion of olfactory ensheathing cells and inhibits their migration. J Cell Sci. 2007;120:1877–87.

Co-transplantation of OEG and MSC 225
cord with reduced cavity formation. Exp Neurol. 2004;187:266–78.Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, 64. Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1998;284:143–7.Lu P, Blesch A, Tuszynski MH. Induction of bone marrow 65. stromal cells to neurons: differentiation, transdifferentia-tion, or artifact? J Neurosci Res. 2004;77:174–91.Krabbe C, Zimmer J, Meyer M. Neural trasndifferentiation 66. of mesenchymal stem cells: a critical review. APMIS. 2005;113:831–44.Ross JJ, Verfaillie CM. Evaluation of neural plasticity in adult 67. stem cells. Philos Trans R Sco Lond B Biol Sci. 2008;363:199–205.
Le Blanc K, Ringden O. Immunomodulation by mesen-60. chymal stem cells and clinical experience. J Intern Med. 2007;262:509–25.Firouzi M, Mashayedi P, Saberi H, Mobasheri H, 61. Abolhassani F, Jahanzad I, et al. Transplantation of Schwann cells to subarachnoid space induces repair in contused rat spinal cord. Neurosci Lett. 2006;402:66–70.Lin CR, Wu PC, Shih HC, Cheng JT, Lu CY, Chou AK, 62. et al. Intrathecal spinal progenitor cell transplantation for the treatment of neurotrophic pain. Cell Transplant. 2002;11:17–24.Ohta M, Suzuki Y, Noda T, Ejiri Y, Dezawa M, Kataoka K, 63. et al. Bone marrow stromal infused into the cerebrospinal fl uid promote functional recovery of the injured rat spinal

Copyright of Cytotherapy is the property of Taylor & Francis Ltd and its content may not be copied or emailed
to multiple sites or posted to a listserv without the copyright holder's express written permission. However,
users may print, download, or email articles for individual use.