persistent phytoplankton bloom in lake st. lucia (isimangaliso
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 2011, p. 5888–5896 Vol. 77, No. 170099-2240/11/$12.00 doi:10.1128/AEM.00460-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Persistent Phytoplankton Bloom in Lake St. Lucia (iSimangalisoWetland Park, South Africa) Caused by a Cyanobacterium
Closely Associated with the Genus Cyanothece(Synechococcaceae, Chroococcales)�
David G. Muir* and Renzo Perissinotto*School of Biological and Conservation Sciences, University of KwaZulu-Natal,
Westville Campus, Durban 4000, South Africa
Received 1 March 2011/Accepted 29 June 2011
Lake St. Lucia, iSimangaliso Wetland Park, South Africa, is the largest estuarine lake in Africa. Extensiveuse and manipulation of the rivers flowing into it have reduced freshwater inflow, and the lake has also beensubject to a drought of 10 years. For much of this time, the estuary has been closed to the Indian Ocean, andsalinities have progressively risen throughout the system, impacting the biotic components of the ecosystem,reducing zooplankton and macrobenthic biomass and diversity in particular. In June 2009, a bloom of ared/orange planktonic microorganism was noted throughout the upper reaches of Lake St. Lucia. The bloompersisted for at least 18 months, making it the longest such bloom on record. The causative organism wascharacterized by light and electron microscopy and by 16S rRNA sequencing and was shown to be a large,unicellular cyanobacterium most strongly associated with the genus Cyanothece. The extent and persistence ofthe bloom appears to be unique to Lake St. Lucia, and it is suggested that the organism’s resistance to hightemperatures, to intense insolation, and to hypersalinity as well as the absence of grazing pressure bysalinity-sensitive zooplankton all contributed to its persistence as a bloom organism until a freshwater influx,due to exceptionally heavy summer rains in 2011, reduced the salinity for a sufficient length of time to producea crash in the cyanobacterium population as a complex, low-salinity biota redeveloped.
Persistent phytoplankton blooms in estuaries and coastalhypersaline waters have been reported on a few occasions (7,13, 34, 35, 41, 49), but the dynamics of their development andduration are not always clear (5, 21). Cyanobacterial blooms inparticular are becoming more frequent worldwide, and whileusually associated with eutrophication (5, 41), they have alsobeen linked to climatic change, including its effects on salina-tion (36).
In June 2009, a widespread bloom of an orange-pigmentedplanktonic organism appeared in the upper reaches of theLake St. Lucia estuary, South Africa, in conjunction with thedevelopment of extreme hypersaline conditions in the area.Lake St. Lucia forms part of the iSimangaliso Wetland Park, aWorld Heritage Site, and is the largest estuarine lake in Africa(19, 20). It has been subject to at least a century of indirectanthropogenic alteration, which has dramatically reduced thefreshwater inflow to it. Extensive areas of wetlands that for-merly supplied the estuary have been drained for agriculturaluse, and water diversion from the rivers feeding into the estu-ary has significantly reduced freshwater inputs (29, 52, 58). Inaddition, the structure of the mouth is managed to prevent thelargest freshwater inflow, the Mfolozi River, from breaching
fully into the estuary, ostensibly to prevent siltation, but thishas resulted in prolonged closure of the mouth (58).
Climatically, the area is subject to alternating drought andwet conditions varying over decadal scales. The current 10-year-long drought is one of the longest on record, however,and has produced profound changes in the ecological condi-tions of the lake (4, 9, 20, 37, 39, 40, 52). Since 1992 the mouthhas opened to the Indian Ocean on only two occasions: brieflyin January 2004 and then for a 6-month period in 2007, whenwaves generated by Cyclone Gamede, combined with highequinoctial spring tides, breached the berm from the oceanside (9). This infrequent opening of the estuary mouth, cou-pled with very low freshwater inflow, has resulted in low waterlevels throughout the system and progressively rising salinities,particularly in the shallow North Lake and False Bay, wherehigh insolation rates, shallow water levels, and high tempera-tures result in rapid evaporation (58). As a result, hypersalin-ity, at times in excess of 200 (on the practical salinity scale) hasbecome the norm for much of the northern reaches, and areverse salinity gradient has been established, with the lowestsalinities near the mouth and hypersaline conditions to thenorth (20).
Under normal conditions, the whole St. Lucia system sup-ports a diverse biotic community (20, 39, 40). However, as aresult of rising salinity, many species have been eliminatedfrom large areas of the lake, and elsewhere only the mostresistant eukaryotes appear to survive (9, 37, 39, 40). Resultsalso suggest that under hypersaline conditions, trophic systemsbegin to be truncated, as grazers are particularly susceptible torising salinity, and that microalgal biomass, both benthic and
* Corresponding author. Mailing address: School of Biological andConservation Sciences, University of KwaZulu-Natal, Westville Cam-pus, P. Bag X54001, Durban 4000, South Africa. Phone for D. G. Muir:27 31 260 1592. Fax: 27 31 260 2029. E-mail: [email protected]. Phonefor R. Perissinotto: 27 31 260 1194. Fax: 27 31 260 2029. E-mail:[email protected].
� Published ahead of print on 8 July 2011.
5888
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

planktonic, initially rises as a result (37). Above 140 on thesalinity scale, however, only prokaryotes appear able to survive,and eukaryotes are eliminated from both the plankton andbenthos (9).
In July 2009, a major fish kill, attributed to exceptionallycold weather, occurred throughout the St. Lucia system, in-cluding False Bay and the North Lake. At the same time,routine sampling showed that salinities had reached levels of�150 in the northern reaches and that zooplankton stocks hadbeen almost entirely eliminated, while megafauna, such aswaterbirds, crocodiles, and hippopotami, disappeared from vir-tually the entire region as they migrated toward the lowersalinities nearer the mouth. At the same time, a widespreadbloom of an orange-pigmented planktonic organism was ob-served throughout the North Lake and False Bay. Provisionaldiagnoses suggested that this bloom might be due to prolifer-ation of a protist and algal genera, such as Chlamydamonas,Dunaliella, or a dinoflagellate, seemed likely candidates, sinceGrindley and Heydorn (23), for example, had reported a2-month-long bloom of Noctiluca scintillans in the False Bayregion during the winter of 1969, which was also a period ofdrought, hypersalinity, and an associated (although more mi-nor) fish kill.
The 2009 bloom persisted for nearly 18 months and changedvery little in appearance. When episodic heavy rains occurred,the bloom would become somewhat diluted with the addedwater, but it reappeared once evaporation again reduced thewater volume. Finally, in January 2011, very heavy rains re-sulted in flooding of the entire lake system, including thenorthern reaches, and led to the apparent final dissolution ofthe bloom. The current work was initiated to provide a defin-itive identification of the bloom organism and preliminary in-formation on the ecological drivers which had caused thebloom to occur and which were responsible for its lengthypersistence.
MATERIALS AND METHODS
Sample site. Lake St. Lucia is located on the subtropical east coast of SouthAfrica, in a summer rainfall area where yearly rainfall totals are normally inexcess of 700 mm (29, 58). The estuary consists of four physically distinct regions(Fig. 1). From the mouth, a narrow riverine channel (The Narrows) leads to theSouth Lake, which is tidally influenced under open-mouth conditions. This isnarrowly connected across a shallow bottom to two large northern embayments(False Bay and North Lake), which are themselves shallow (�2-m depth) andonly minimally tidally influenced (52, 58).
The sample site was located in the False Bay region at Lister’s Point (Fig. 1).The sample site is typical of the northern reaches, with shallow water that reacheshigh daytime temperatures and which is subject to seiching (wide-scale move-ment due to wind action). A comparison site at Esengeni in The Narrows (Fig.1) was used to compare physico-chemical parameters.
Physico-chemical variables. Depth, temperature (°C), pH, dissolved oxygen(mg liter�1) and salinity (on the practical salinity scale) were measured by usinga YSI 6920 water quality logger fit with the appropriate probes.
Water sample collection. Samples of 450 ml were taken in acid-washed, sterile,1.0-liter Schott bottles. These were transported to the laboratory and maintainedwith loose caps in front of a bank of warm fluorescent lighting (140 �mol m�2
s�1 at the bottle surface) under a 14-h light, 10-h dark regimen at room tem-perature at a 30-cm distance.
Chl a determinations. Known volumes of water were filtered onto WhatmanGF/F filters under gentle vacuum and stored frozen. Chlorophyll a (Chl a) wasextracted by immersing filters in cold (�5°C) 90% acetone for 24 h. The Chl aconcentration was determined using a Turner Designs 10-AU fluorometer witha narrow-band, nonacidification system (57).
Cell counts. Cell counts of the bloom organism were determined with ahemocytometer, following the methods of Guillard and Sieracki (24). Historicalcounts were determined from Lugol’s iodine-fixed samples taken prior to Feb-ruary 2010. After that date, cell counts were determined from live samples withina day of collection.
Culture and enrichment responses. Employing nutrient amounts recom-mended for F/2 marine algal culture medium (2), duplicate 450-ml samples oflake water were fertilized with 8.83 � 10�4 M (final concentration [f.c.]) NaNO3
and 3.63 � 10�5 M NaH2PO4 � H2O (f.c.). Duplicate controls were left unfer-tilized. Chl a content was determined each week for a period of 5 weeks usingmethods described above.
Attempts were made to purify the organism by dilution plating and streakingonto SN medium (2) solidified with 1.0% (f.c.) Noble agar, but even whereapparently isolated colonies arose, these never proved to be axenic, and theorganism has proved recalcitrant to all attempts to establish pure culture usingpublished methods (55).
Light microscopy. Sample aliquots were examined on glass slides with a NikonEclipse 80i microscope, using phase contrast and Nomarski differential interfer-ence contrast (DIFC). To examine capsule and slime structures, cells werestained using the Anthony direct stain modification of the Hiss method (3), inwhich dried smears of cell suspensions were stained for 2 min with 1% (wt/vol)aqueous crystal violet and destained by rinsing with 20% (wt/vol) CuSO4 � 7H2O,followed by brief rinsing in distilled water to reduce salt crystal formation.Capsules were also demonstrated with a negative stain modification of theAnthony method, in which cell suspensions were mixed with skim milk (0.1%[vol/vol]) to serve as a background. The Duguid negative stain method (17) couldnot be used, because nigrosin and India ink both precipitated and clumped athigh salinities and failed to show capsules adequately.
FIG. 1. St. Lucia Estuary, iSimangaliso Wetland Park, KwaZulu-Natal, South Africa, showing both sites (Lister’s Point and Esengeni)for which physico-chemical data are presented. Adapted from refer-ence 9 with permission of the publisher.
VOL. 77, 2011 CYANOTHECE BLOOM IN HYPERSALINE LAKE ST. LUCIA, SA 5889
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from
Epifluorescence microscopy. Five-milliliter sample aliquots were stained with4�,6-diaimidino-2-phenylindole (DAPI; 5 � 10�5 mg ml�1 f.c.), and subsampleswere mounted on glass slides. Examination was with a Nikon Eclipse 80i, utilizingNikon filter sets UV-2a (excitation, 330 to 380 nm; barrier filter, 420 nm) forDAPI visualization and the FITC (G2A) filter set (excitation, 510 to 560 nm;barrier filter, 590 nm) for visualization of Chl a autofluorescence.
For photomicrography we used Nikon imaging software (NIS-D) for digitalimage capture using a Nikon digital sight DS-Fli digital camera. Where neces-sary, pictures were cropped and enhanced for color and contrast, etc., using theGNU image manipulation program (GIMP-2; http://www.gimp.org; last accessed6/23/2011).
Transmission electron microscopy (TEM). Fresh samples were fixed in 2.5%glutaraldehyde (f.c.) before proceeding through preparation solutions. Aftereach solution change, the samples were lightly centrifuged to form a pellet. Thesupernatant was then carefully removed before being replaced by the next solu-tion in the process.
After several rinses with 0.1 M phosphate buffer, the material was postfixed in1% osmium tetroxide solution for 1 h, washed several times with phosphatebuffer, and dehydrated in a graded acetone series. The samples were infiltratedwith 50:50 acetone-epoxy resin (48) overnight and further infiltrated with 100%epoxy resin for 24 h. Samples were placed in resin-filled Beem capsules andpolymerized at 70°C for 8 h.
Sectioning was carried out using a Reichert Ultracut E microtome. Ultrastruc-tural analysis was performed using ultrathin sections (60 to 100 nm), which werepoststained with saturated uranyl acetate solution (10 min) followed by leadcitrate (44) for 10 min. Sections were viewed with a Jeol 1010 transmissionelectron microscope, and images were captured using a Megaview 3 Soft imagingsystem (SIS).
16S rRNA amplification and sequencing. To reduce the proportion of DNAfrom other organisms in the bulk DNA extraction, a centrifugation method wasused to amplify the relative proportion of large cyanobacterial cells beforesubmission for 16S rRNA sequencing. After centrifugation for 30 min at 5,000 �g, the cells under consideration failed to form a firm pellet, but other organisms,such as eubacteria and smaller cyanobacteria, did, as determined by phase-contrast microscopic examination of centrifugal pellets. As a result, repeatedcentrifugation and decantation of supernatant containing cells of interest led tothe progressive, albeit crude amplification of large cyanobacteria relative to otherorganisms.
Samples of concentrated cells were sent to the Inqaba Laboratories (InqabaBiotech, Hatfield 0028, Pretoria, South Africa) for DNA extraction, 16S rRNAgene amplification by PCR, and sequencing of 26 randomly picked clones usingstandard methods. Amplification and sequencing in both directions were per-formed using universal primers pJET-F (5�-CGACTCACTATAGGGAGAGCGGC), pJET-r (5�-AAGAACATCGATTTTCCATGGCAG), 27F (5�-AGAGTTTGATCMTGGCTCAG), and 1492r (5�-TACGGYTACCTTGTTACGACTT). Contigs for each of 26 randomly selected clones were compiled and joined,and sequences for all selected clones were aligned using the CLC GenomicsWorkbench 4.0.2 program (CLC-Bio, Aarhus, Denmark). The alignment wasedited to attempt to match all 26 isolated clone sequences, a total of 1,400 bases,or 91% of the complete 16S rRNA gene. Three sequences were not used becausethe forward and reverse contigs could not be joined, as the complete sequenceswere excessively short. All 23 sequences used in subsequent phylogenetic ana-lyses were designated environmental clone isolates Saint Lucia (SL) 01 to 26 andhave been submitted to the GenBank database (http://www.ncbi.nlm.nih.gov/GenBank/) with designations detailed below. An initial Basic Local AlignmentSearch Tool (BLAST) search was done using the consensus sequence to deter-mine approximate phylogenetic relationships, and each individual sequence wasalso subjected to BLAST analysis. Sequence information for organisms associ-ated on the BLAST searches was extracted from the GenBank database fordetermination of phylogenetic relationships (see Fig. 7, below). The marineeubacterium Oceanobacterium iheyi was used as an outgroup, and the completealignment of 35 sequences was again compiled using the CLC Genomics work-bench.
These sequence data were analyzed for phylogenetic relatedness by using aneighbor-joining method (46, 50) of Mega-4 (51) with a bootstrap consensus treeinferred from 1,000 replicates. Phylogenetic distances were computed using themaximum composite likelihood method, where the unit of distance is the numberof base substitutions per site.
Nucleotide sequence accession numbers. All sequences used for phylogeneticanalysis were deposited in the GenBank database (National Center for Biotech-nology Information; http://www.ncbi.nlm.nih.gov) in January 2011 and assignedaccession numbers HQ914216 to HQ14239.
RESULTS
Water chemistry. At our initial sampling (19 February 2010),the water at the sample site was distinctly pigmented with abright orange/red color. The water was shallow (maximumdepth, 50 cm) and recorded mid-day water temperatures werehigh (55°C), with a pH of 8.1. The oxygen content was 8.17 mgliter�1 (100% saturation). Large banks of brown, stable foamwere present on the beaches, and foam could be seen formingon the water surface as increasing wind speed caused watermovement across the shallow embayment. The photosyntheti-cally available radiation (PAR) at the surface at the time ofsampling was 530.4 �mol m�2 s�1. Because of the shallownessof the water, it was not possible to calculate the Kd, but Peris-sinotto et al. (37) have recorded even higher PAR values(�2,000 �mol m�2 s�1) in the system, although the Kd mayalso be extremely high (0.9 to 36.1 m�1), depending on depth,turbidity, and phytoplankton density.
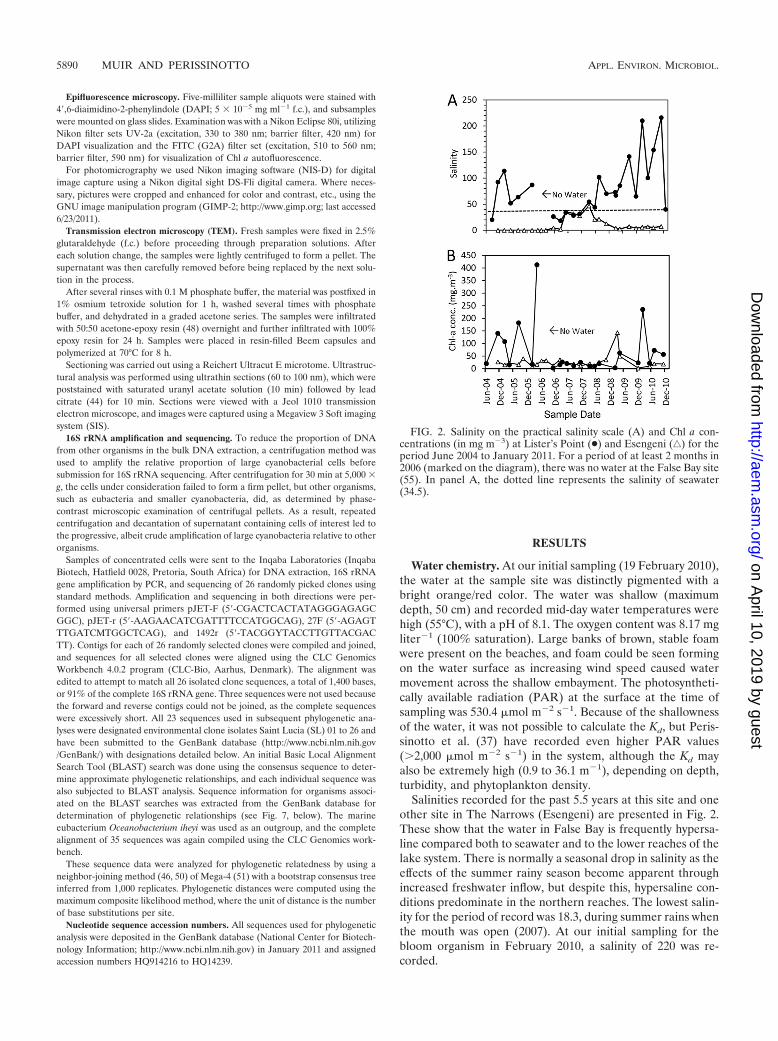
Salinities recorded for the past 5.5 years at this site and oneother site in The Narrows (Esengeni) are presented in Fig. 2.These show that the water in False Bay is frequently hypersa-line compared both to seawater and to the lower reaches of thelake system. There is normally a seasonal drop in salinity as theeffects of the summer rainy season become apparent throughincreased freshwater inflow, but despite this, hypersaline con-ditions predominate in the northern reaches. The lowest salin-ity for the period of record was 18.3, during summer rains whenthe mouth was open (2007). At our initial sampling for thebloom organism in February 2010, a salinity of 220 was re-corded.
FIG. 2. Salinity on the practical salinity scale (A) and Chl a con-centrations (in mg m�3) at Lister’s Point (●) and Esengeni (‚) for theperiod June 2004 to January 2011. For a period of at least 2 months in2006 (marked on the diagram), there was no water at the False Bay site(55). In panel A, the dotted line represents the salinity of seawater(34.5).
5890 MUIR AND PERISSINOTTO APPL. ENVIRON. MICROBIOL.
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from
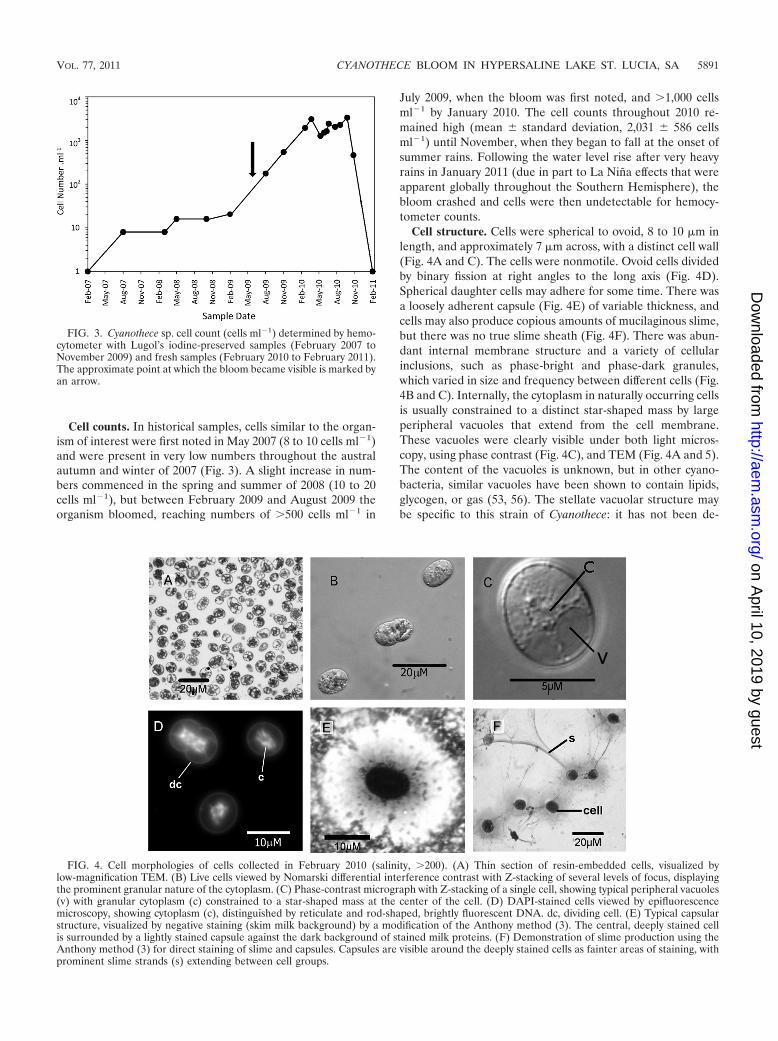
Cell counts. In historical samples, cells similar to the organ-ism of interest were first noted in May 2007 (8 to 10 cells ml�1)and were present in very low numbers throughout the australautumn and winter of 2007 (Fig. 3). A slight increase in num-bers commenced in the spring and summer of 2008 (10 to 20cells ml�1), but between February 2009 and August 2009 theorganism bloomed, reaching numbers of �500 cells ml�1 in
July 2009, when the bloom was first noted, and �1,000 cellsml�1 by January 2010. The cell counts throughout 2010 re-mained high (mean � standard deviation, 2,031 � 586 cellsml�1) until November, when they began to fall at the onset ofsummer rains. Following the water level rise after very heavyrains in January 2011 (due in part to La Nina effects that wereapparent globally throughout the Southern Hemisphere), thebloom crashed and cells were then undetectable for hemocy-tometer counts.
Cell structure. Cells were spherical to ovoid, 8 to 10 �m inlength, and approximately 7 �m across, with a distinct cell wall(Fig. 4A and C). The cells were nonmotile. Ovoid cells dividedby binary fission at right angles to the long axis (Fig. 4D).Spherical daughter cells may adhere for some time. There wasa loosely adherent capsule (Fig. 4E) of variable thickness, andcells may also produce copious amounts of mucilaginous slime,but there was no true slime sheath (Fig. 4F). There was abun-dant internal membrane structure and a variety of cellularinclusions, such as phase-bright and phase-dark granules,which varied in size and frequency between different cells (Fig.4B and C). Internally, the cytoplasm in naturally occurring cellsis usually constrained to a distinct star-shaped mass by largeperipheral vacuoles that extend from the cell membrane.These vacuoles were clearly visible under both light micros-copy, using phase contrast (Fig. 4C), and TEM (Fig. 4A and 5).The content of the vacuoles is unknown, but in other cyano-bacteria, similar vacuoles have been shown to contain lipids,glycogen, or gas (53, 56). The stellate vacuolar structure maybe specific to this strain of Cyanothece: it has not been de-
FIG. 3. Cyanothece sp. cell count (cells ml�1) determined by hemo-cytometer with Lugol’s iodine-preserved samples (February 2007 toNovember 2009) and fresh samples (February 2010 to February 2011).The approximate point at which the bloom became visible is marked byan arrow.
FIG. 4. Cell morphologies of cells collected in February 2010 (salinity, �200). (A) Thin section of resin-embedded cells, visualized bylow-magnification TEM. (B) Live cells viewed by Nomarski differential interference contrast with Z-stacking of several levels of focus, displayingthe prominent granular nature of the cytoplasm. (C) Phase-contrast micrograph with Z-stacking of a single cell, showing typical peripheral vacuoles(v) with granular cytoplasm (c) constrained to a star-shaped mass at the center of the cell. (D) DAPI-stained cells viewed by epifluorescencemicroscopy, showing cytoplasm (c), distinguished by reticulate and rod-shaped, brightly fluorescent DNA. dc, dividing cell. (E) Typical capsularstructure, visualized by negative staining (skim milk background) by a modification of the Anthony method (3). The central, deeply stained cellis surrounded by a lightly stained capsule against the dark background of stained milk proteins. (F) Demonstration of slime production using theAnthony method (3) for direct staining of slime and capsules. Capsules are visible around the deeply stained cells as fainter areas of staining, withprominent slime strands (s) extending between cell groups.
VOL. 77, 2011 CYANOTHECE BLOOM IN HYPERSALINE LAKE ST. LUCIA, SA 5891
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

scribed for C. aeruginosa, the type species of the genus (26),nor for strains BH63 and BH68 (43), C. halobia (10), or C.minervae/cedrorum (28), three strains described by Porta et al.(42), nor for any others we have considered from the literature,such as the study reported by Garcia-Pichel et al. (22).
Nuclear staining with DAPI showed that there was no dis-tinct nucleoid but that DNA was distributed throughout thecentral cytoplasm mass, forming distinct thick filaments con-strained to the cytoplasm within the boundaries of the periph-eral vacuoles (Fig. 4D). There were regional areas where theDNA is more brightly fluorescent than elsewhere. There waslittle change in DNA distribution during cell division, as re-vealed by DAPI staining (Fig. 4D).
Viewed under an epifluorescent wavelength of 510 to 560nm, freshly sampled cells were autofluorescent with a variablebrightness that was red, typical of chlorophyll. The autofluo-rescence showed filaments and plates distributed throughoutthe cytoplasm, in a manner strongly suggestive of the thyla-koids characteristic of other unicellular, diazotrophic cyano-bacteria (28, 43). They are typically arranged in a net-likefashion (Fig. 5), described as “keritomization” by Komarekand Cepak (28).
We noted that after cells had been maintained for 2 monthswithout addition of nutrients, the culture gradually paled incolor from the original red/orange to a straw yellow/orange.Cells from such cultures also lost the bright autofluorescenceof freshly sampled cells, although autofluorescence was neverentirely lost, and the volume of cytoplasm decreased dramat-ically (Fig. 5A) with a concomitant increase in vacuole vol-umes. Further, cells that had been fertilized developed a bril-liant red autofluorescence as their pigmentation changed fromred to blue-green. This could be maintained for more than 6months even without further fertilization, and only then slowlyfading to resemble environmentally collected cells.
TEM showed that cells were uniformly circular in cross-section with a thin but distinct cell wall (Fig. 5). The largevacuoles previously described were clearly visible, bounded bythe cell membrane and thylakoids on the outer periphery andby a stellate arrangement of cytoplasm and thylakoids that ex-tended inwards to the central cytoplasm itself (Fig. 4A and 5).
Effects of fertilization. Both electron microscopy (Fig. 5A toD) and Chl a results (Fig. 6) showed that there was an almostimmediate response to nutrient addition as the internal struc-tures of the cells of interest became substantially reorganizedwithin a week: the amount of cytoplasm increased, vacuolesbegan to be subdivided by production of dense and elaborate
thylakoid structures (Fig. 5A to D), and cells developed abright blue-green pigmentation. There was a 42% increase inChl a content that was maintained for at least another month(Fig. 6). It was evident that thylakoid development continuedfor at least 3 weeks (Fig. 5D).
Nutrient dynamics of this organism are the subject of ongo-ing research, since these results reflect the addition of highlevels of nutrient at very high salinity. Responses to morerealistic levels of nutrient at low salinities remain to be deter-mined.
16S rRNA sequence analysis. BLAST searching with eachSt. Lucia isolate sequence against the NCBI database of bac-terial 16S rRNA sequences and genomes showed similar re-sults. The organisms were placed within the Cyanobacteria,with each being most closely associated with identified Cyan-othece sequences, with some partial identification to the genusDactylococcopsis and with a more remote relatedness to thegenera Halothece and Euhalothece. Similarity cluster analysisby the neighbor-joining method (46, 50) of 26 cloned se-quences suggested that a population of very closely relatedorganisms had been sampled, albeit with some degree of poly-morphism. Nonetheless, all 26 clones showed a high degree ofsimilarity, and a preliminary consensus sequence was derived,which was itself subjected to BLAST searching. This confirmedthe placement of the organism within the cyanobacteria, mostclosely associated with members of the genus Cyanothece andto other cyanobacteria associated with hypersaline conditions,such as in salt ponds (Dactylococcopsis sp.).
The 23 most complete sequences were then comparedagainst selected identified sequences that BLAST searchinghad shown to have a range of relatedness (Fig. 7). This re-
FIG. 5. Micrographs of thin sections of cells maintained at a salinity of 200, at various times after addition of nutrient (NaNO3 or NaH2PO4).(A) Nutrient-starved cell; (B) 1 week after nutrient addition; (C) 2 weeks after nutrient addition; (D) 3 weeks after nutrient addition. Bar, 1 �M.
FIG. 6. Chlorophyll a content (mg m�3) after addition of NaNo3and NaH2PO4 � H2O. F, experimental; E, control.
5892 MUIR AND PERISSINOTTO APPL. ENVIRON. MICROBIOL.
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from
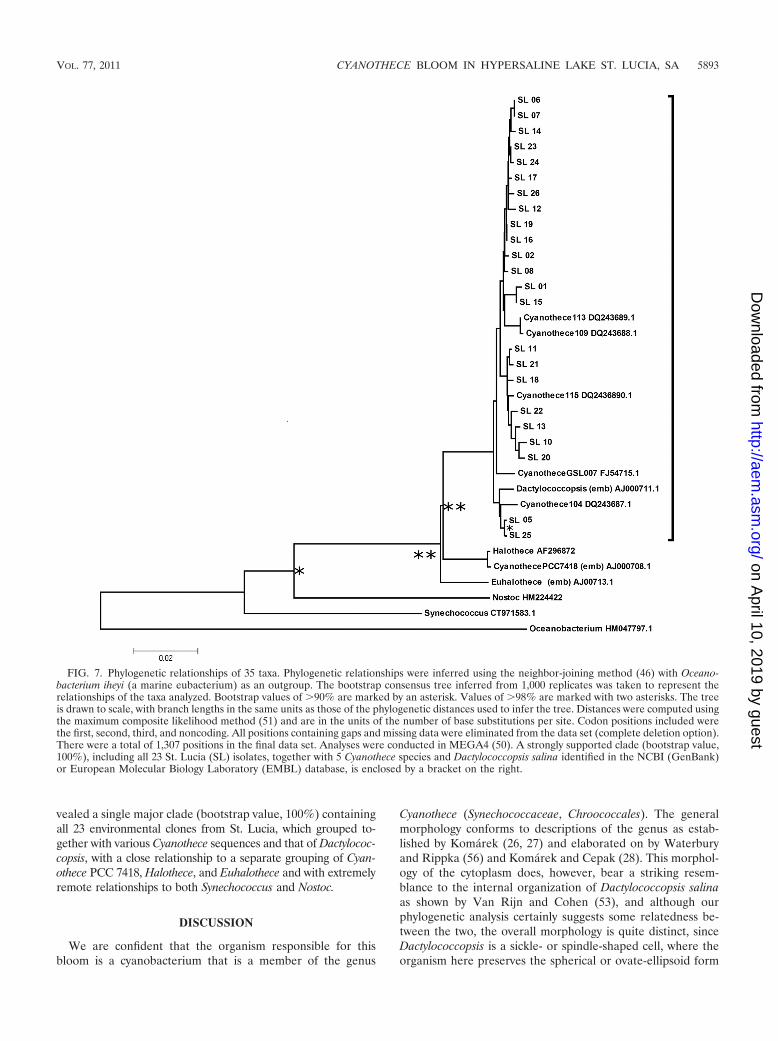
vealed a single major clade (bootstrap value, 100%) containingall 23 environmental clones from St. Lucia, which grouped to-gether with various Cyanothece sequences and that of Dactylococ-copsis, with a close relationship to a separate grouping of Cyan-othece PCC 7418, Halothece, and Euhalothece and with extremelyremote relationships to both Synechococcus and Nostoc.
DISCUSSION
We are confident that the organism responsible for thisbloom is a cyanobacterium that is a member of the genus
Cyanothece (Synechococcaceae, Chroococcales). The generalmorphology conforms to descriptions of the genus as estab-lished by Komarek (26, 27) and elaborated on by Waterburyand Rippka (56) and Komarek and Cepak (28). This morphol-ogy of the cytoplasm does, however, bear a striking resem-blance to the internal organization of Dactylococcopsis salinaas shown by Van Rijn and Cohen (53), and although ourphylogenetic analysis certainly suggests some relatedness be-tween the two, the overall morphology is quite distinct, sinceDactylococcopsis is a sickle- or spindle-shaped cell, where theorganism here preserves the spherical or ovate-ellipsoid form
FIG. 7. Phylogenetic relationships of 35 taxa. Phylogenetic relationships were inferred using the neighbor-joining method (46) with Oceano-bacterium iheyi (a marine eubacterium) as an outgroup. The bootstrap consensus tree inferred from 1,000 replicates was taken to represent therelationships of the taxa analyzed. Bootstrap values of �90% are marked by an asterisk. Values of �98% are marked with two asterisks. The treeis drawn to scale, with branch lengths in the same units as those of the phylogenetic distances used to infer the tree. Distances were computed usingthe maximum composite likelihood method (51) and are in the units of the number of base substitutions per site. Codon positions included werethe first, second, third, and noncoding. All positions containing gaps and missing data were eliminated from the data set (complete deletion option).There were a total of 1,307 positions in the final data set. Analyses were conducted in MEGA4 (50). A strongly supported clade (bootstrap value,100%), including all 23 St. Lucia (SL) isolates, together with 5 Cyanothece species and Dactylococcopsis salina identified in the NCBI (GenBank)or European Molecular Biology Laboratory (EMBL) database, is enclosed by a bracket on the right.
VOL. 77, 2011 CYANOTHECE BLOOM IN HYPERSALINE LAKE ST. LUCIA, SA 5893
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

typical of Cyanothece. In Dactylococcopsis salina, the corre-sponding vacuolar structures are gas filled and provide a sen-sitive flotation control, which maintains cells under optimumconditions in the water column (54). The content and functionof vacuoles in Cyanothece recovered from Lake St. Lucia re-main to be determined.
Although cells have no true slime tube, there is a prominentcapsule, and cells produce a large amount of a mucilaginousslime that remains only loosely adherent to the cells but whichcan be demonstrated by staining using the Anthony method(Fig. 4F). With wind or wave action, this slime contributes tothe formation of large amounts of a stable foam, which ischaracteristic of those areas of Lake St. Lucia where the bloomis present. Similarly, prolific exopolysaccharide production hasbeen demonstrated for several other Cyanothece species andstrains and may account for a substantial proportion of cellularproduction (11, 15, 16). Not only may exopolymers provide adirect trophic resource (14), but also the resulting foam mayhave an additional important ecological function. Blown uponto the beaches, it remains as a stable layer that is continuallyreplenished as wind action moves fresh foam onshore. If thesefoam banks are moved aside, a distinctly wet layer of substra-tum with a prominent, active microbial mat is revealed beneaththem. While extremely hypersaline, the environment beneaththe foam banks is moist and shielded from the intense heat andinsolation characteristic of the dry, bare mudflats that com-prise the uncovered beach. Bate and Smailes (4) have pointedout the importance of refugia such as these in maintainingdiatom populations in particular, and we are certain that closeexamination would show a complex associated bacterial floraas well. We also believe that the capsule and associated slimecontribute to the difficulty of establishing axenic cultures of thisorganism, since other bacteria and smaller cyanobacteria arevery frequently observed to associate with the enveloping poly-saccharides.
Members of the genus Cyanothece are commonly associatedwith extreme conditions similar to those in Lake St. Lucia (22).For example, C. halobia was isolated from saltworks (45) andDe Philippis et al. (16) isolated the genus from a Somali salt-pan and subsequently described 14 other strains from similarhabitats (15), while Margheri et al. (32) described a further 11strains isolated from geographically disparate saltworks, hyper-saline ponds, tidal pools, solar lakes, and the Dead Sea.
Cells similar to those of the bloom were present in lownumbers in water samples as early as June 2007, a year beforethe bloom began, when salinities, while high (�70), were farbelow the maximum observed later (�200). Routine samplingshowed that a diverse planktonic and benthic trophic commu-nity was still present at these lower salinities (9). As salinitiesrose above 100 in 2009; however, this community was trun-cated. In particular, eukaryotes, such as ciliates, phytoflagel-lates, and the entire zooplankton community, disappeared (al-though a complex prokaryotic community survived, in additionto the bloom organisms). With the loss of the predatory com-munity, the number of cyanobacterial cells began to increase.The fish kill, due to cold weather in July 2009, occurred whenwater levels were reduced at the end of the lengthy dry winterseason in a year where rainfall had already been lowered bydrought. Although hypersaline conditions were present, thesalinity at first notice of the bloom was 100 or less, conditions
much more conducive to rapid cell division than the latersalinities of �150, and other authors have noted that nutrientrequirements of phytoplankton increase at higher salinitiesand that even organisms tolerant of hypersalinity experienceincreased metabolic demands (27, 29).
We are unable to say what sparked the onset of the bloomconditions, but we speculate that a transient nutrient pulse ledto the initial increase in numbers of Cyanothece cells sufficientto produce bloom conditions by late June 2009 and that thelater fish kill further augmented nutrients. We have noted thateven a brief exposure to high nutrient levels results in a veryrapid restructuring of the cell architecture and swift upregula-tion of photosynthetic pigments, which then persist for manymonths (Fig. 5 and 6), and that even environmental, unfertil-ized samples can be maintained, virtually unchanged, for morethan a year. Perissinotto et al. (33) have also shown that dis-solved inorganic nitrate and phosphate are seldom at limitingconcentrations in Lake St. Lucia and may thus be high enoughfor maintenance of the bloom through efficient regeneration ofnutrients. Without any grazing pressure from ciliates or zoo-plankton, and with resistance to hypersalinity and extremelight and heat conditions, the bloom organism, once estab-lished, would survive in the absence of other competitive or-ganisms.
Although Cyanothece has been isolated from marine andhypersaline environments frequently (11, 12, 15, 32, 43) andthere is considerable interest in the diazotrophic activity ofseveral of the members of the genus (43), there is very littleinformation in the literature regarding its general ecology orthat of any of the other large, nanoplanktonic unicellular cya-nobacteria. The only similar reports are from work with Dac-tylococcopsis salina recovered from Solar Lake, Sinai, whichundergoes seasonal population expansions and becomes spa-tially concentrated in the lake, forming a dense narrow bandconstrained by its intolerance of H2S in deeper water and highinsolation and zooplankton predation in the upper layers (49,53, 54). We believe that ours is the first recorded bloom of thegenus Cyanothece. Other members of the Chroococcales, suchas Synechococcus and Prochlorococcus, have been documentedto form blooms in Florida Bay (6, 25, 38), Australian coastallagoons (47), and also recorded in Brazilian coastal lagoons(12), but these are extremely small, picoplanktonic cells. Syn-echococcus, for example, seldom exceeds 2 �m in diameter,and even at its largest has a surface area/volume (SA/V) ratioof approximately 1.5, whereas the cells found in Lake St. Luciahad a biovolume about 25 times that of Synechococcus and anSA/V ratio of only 0.89. We suggest that the nutrient uptakedynamics of nanoplanktonic cells must be very different andthat the high uptake efficiencies of the picoplanktonic cells,which result from their high SA/V ratio (1) and which accountfor their tendency to bloom even at low nutrient levels, do notapply in this case. The nutrient dynamics of our isolate remainthe subject of ongoing research, but it should be noted that thisis a mesotrophic system (37) and that it is likely that nutrienttransfer from sediments to the very shallow water is extremelyefficient, especially during windy periods, when rapid watermovement across the sediments is seen. It must be borne inmind, however, that halotolerant cells may become nutrientstressed even at relatively high nutrient levels as salinity rises
5894 MUIR AND PERISSINOTTO APPL. ENVIRON. MICROBIOL.
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

(31, 33), and thus cellular metabolic demands at high salinitiesmay be much greater.
The cause of the onset of the bloom thus remains unclear,but we believe that the loss of grazing organisms at high salinitywas the most significant factor in establishment of the bloom.An analogous situation, albeit for a protistan population, hasbeen described by Buskey et al. (7, 8) and Stockwell et al. (49)for the Laguna Madre, an estuarine system in Texas also char-acterized by periodic development of hypersaline conditions.There, the onset of an 8-year bloom of Aureoumbra lagunensiswas ascribed to the collapse of grazer populations due to theonset of hypersaline conditions as well as a fish kill due to coldweather, and its persistence was due to elimination of compet-itors. After 8 years, the bloom collapsed when salinities fellbelow 20 after a period of heavy rain. At this time, a complexplanktonic community of ciliates, dinoflagellates, and zoo-plankters was reestablished, only to be lost again as hypersa-linity reestablished and the Aureoumbra bloom returned toprominence (7, 8). We suggest that a similar dynamic operatesin the northern areas of Lake St. Lucia. A further, tellingsimilarity between the two systems is that Buskey et al. (7, 8)suggested that slow water turnover is a major contributor tomaintenance of the bloom. In Lake St. Lucia, not only are thenorthern reaches currently receiving minimal freshwater input,but low water levels have resulted in their physical separationfrom the lower reaches, where salinity is far lower (�35) (29).
Long-term climate prediction models for the KwaZulu-Na-tal region are pertinent to this discussion. Overall, it has beensuggested that the area will become somewhat wetter, but themodels also predict that the frequency and duration of ex-treme climate events, such as floods and droughts, will alsoincrease (18, 30). If the status quo in St. Lucia is maintainedand the main freshwater input, the Mfolozi River, is keptseparate, as it is at present, it is likely that the frequency oflengthy hypersaline periods in the lake will increase, inter-rupted by periods of flood. These conditions would stronglyfavor the development of halotolerant blooms in the lake: ifnot of Cyanothece, then of other cyanobacteria or even ofhalotolerant protists, such as Dunaliella.
The Cyanothece bloom described here persisted throughoutthe northern reaches of Lake St. Lucia for 18 months as thedominant phytoplankter, in a system of markedly reducedtrophic complexity. It appears, from our examination of theliterature, that this was one of the most persistent cyanobac-terial blooms in a natural system on record, serving as a markerof the biological crisis of hypersalinity in which this system iscurrently placed. The current very low salinities throughout thesystem are, unfortunately, a temporary respite for Lake St.Lucia. Without substantial and repeated opening of the estuarymouth, the onset of the lengthy winter dry seasons will againsee the rapid reduction of water volumes, with concomitantrises in salinities. We wait, with interest, to see if these will beaccompanied by the reappearance of the identified Cyanothecesp. cells.
ACKNOWLEDGMENTS
Funding for this project was provided by the National ResearchFoundation (NRF Pretoria grant number 71051).
We thank the iSimangaliso Wetland Park Authority and the staffand management of Ezemvelo KwaZulu-Natal Wildlife (EKZN Wild-
life). We are particularly grateful to Ricky Taylor and Caroline Fox fortheir help with logistics and field collections. Guy Bate and Pat Smaileskindly provided samples from early collections undertaken in 2007 to2009. Ursula Scharler, David Dyer, Nelson Miranda, Nicola Carrasco,Sarah Bownes, Katrin Tirok, Holly Nel, and Nasreen Peer are grate-fully acknowledged for their help in the field and Jennifer Godlontonfor lab work. Angus MacDonald provided invaluable assistance withanalysis of molecular data. We also thank James Wesley-Smith andPriscilla Maartens of the UKZN electron microscopy unit for theirhelp with TEM and light microscopy work.
REFERENCES
1. Agawin, N. S. R., C. M. Duarte, and S. Agusti. 2000. Response of Mediter-ranean Synechococcus growth and loss rates to experimental nutrient inputs.Mar. Ecol. Prog. Ser. 206:97–106.
2. Andersen, R. A., J. A. Berges, P. J. Harrison, and M. M. Watanabe. 2005.Recipes for freshwater and seawater media, p. 429–538. In R. A. Andersen(ed.), Algal culturing techniques. Elsevier, Amsterdam, Netherlands.
3. Anthony, E. E. 1931. A note on capsule staining. Science 73:319–320.4. Bate, G. C., and P. A. Smailes. 2008. The response of the diatom flora of St.
Lucia Lake and estuary, South Africa, to a severe drought. Afr. J. Aquat. Sci.33:1–15.
5. Bianchi, T. S., et. al. 2000. Cyanobacterial blooms in the Baltic Sea: naturalor human-induced? Limnol. Oceanogr. 45:716–726.
6. Boyer, J. N., C. R. Kelble, P. B. Ortner, and D. T. Rudnick. 2009. Phyto-plankton bloom status: chlorophyll a biomass as an indicator of water qualitycondition in the southern estuaries of Florida, USA. Ecol. Indic. 9S:S56–S67.
7. Buskey, E. J., H. Liu, C. Collumb, and J. G. F. Bersano. 2001. The declineand recovery of a persistent Texas brown tide algal bloom in the LagunaMadre (Texas USA). Estuaries 24:337–346.
8. Buskey, E. J., B. Wysor, and C. Hyatt. 1998. The role of hypersalinity in thepersistance of the Texas ‘brown tide’ in the Laguna Madre. J. Plankton Res.20:1553–1565.
9. Carrasco, N. K., R. Perissinotto, and D. Pillay. 2010. Zooplankton of the St.Lucia Estuary during the current drought cycle: a comparison between open-and closed-mouth conditions. Mar. Ecol. Prog. Ser. 399:157–171.
10. Cepak, V. 1996. Nucleoid morphology in the coccal cyanophyte Cyanothecehalobia (Chroococcales, Cyanophyta): a DAPI fluorescence study. Phycolo-gia 35:523–527.
11. Chi, Z., C. D. Su, and W. D. Lu. 2007. A new exopolysaccharide produced bymarine Cyanothece sp. 113. Bioresour. Technol. 98:1329–1332.
12. Clementino, M. M., et al. 2008. Prokaryotic diversity in one of the largesthypersaline coastal lagoons in the world. Extremophiles 12:595–604.
13. da Rosa, C. E., et al. 2005. Cyanobacterial blooms in estuarine ecosystems:characteristics and effects on Laonereis acuta (Polychaeta, Nereididae). Mar.Pollut. Bull. 50:956–964.
14. Decho, A. W., and G. R. Lopez. 1993. Exopolymer microenvironments ofmicrobial flora: multiple and interactive effects on trophic relationships.Limnol. Oceanogr. 38:1633–1645.
15. De Philippis, R. D., M. C. Margheri, R. Materassi, and M. Vincenzini. 1998.Potential of unicellular cyanobacteria from saline environments as exopoly-saccharide producers. Appl. Environ. Microbiol. 64:1130–1132.
16. De Philippis, R., M. C. Margheri, E. Pelosi, and S. Ventura. 1993. Exopoly-saccharide production by a unicellular cyanobacterium isolated from a hy-persaline habitat. J. Appl. Phycol. 5:387–394.
17. Duguid, J. P. 1951. The demonstration of bacterial capsules and slime.J. Pathol. Bacteriol. 63:673.
18. Fauchereau, N., S. Trzaska, M. Roault, and Y. Richard. 2003. Rainfallvariability and changes in Southern Africa during the 20th century in theglobal warming context. Nat. Hazards 29:139–154.
19. Fielding, A. T., A. T. Forbes, and N. T. Demetriades. 1991. Chlorophyllconcentrations and suspended particulate loads in St. Lucia, a turbid estuaryon the east coast of South Africa. Afr. J. Mar. Sci. 11:491–498.
20. Forbes, A. T., and D. P. Cyrus. 1993. Biological effects of salinity gradientreversals in a southeast African estuarine lake. Neth. J. Aquat. Ecol. 27:483–488.
21. Galat, D. L., and J. P. Verdin. 1989. Patchiness, collapse and succession of acyanobacterial bloom evaluated by synoptic sampling and remote sensing. J.Plankton Res. 11:925–948.
22. Garcia-Pichel, F., U. Nubel, and G. Muyzer. 1998. The phylogeny of unicel-lular, extremely halotolerant cyanobacteria. Arch. Microbiol. 169:469–482.
23. Grindley, J. R., and A. E. F. Heydorn. 1970. Red water and associatedphenomena in St. Lucia. S. Afr. J. Sci. 66:210–213.
24. Guillard, R. R. L., and M. S. Sieracki. 2005. Counting cells in cultures withthe light microscope, p. 239–252. In R. A. Andersen (ed.), Algal culturetechniques. Elsevier, Amsterdam, Netherlands.
25. Hitchcock, G. E., L. Brand, and D. Morrison. 2007. Plankton blooms, p.77–91. In J. H. Hunt and W. Nuttle (ed.), Florida Bay science program: asynthesis of research on Florida Bay. Fish and Wildlife Research InstituteTechnical Report TR-11. Fish and Wildlife Research Institute, St. Peters-burg, FL.
VOL. 77, 2011 CYANOTHECE BLOOM IN HYPERSALINE LAKE ST. LUCIA, SA 5895
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

26. Komarek, J. 1976. Taxonomic review of the genera SynechocystisSAUV.1892, Synechococcus NAG.1849, and Cyanothece gen. nov. (Cyano-phyceae). Arch. Protist. 118:119–179.
27. Komarek, J., and K. Anagnostidis. 1986. Modern approach to the classifi-cation system of cyanophytes. 2. Chroococcales. Arch. Hydrobiol. Algolog.Studies 43(Suppl. 73):157–226.
28. Komarek, J., and V. Cepak. 1998. Cytomorphological characters supportingthe taxonomic validity of Cyanothece (Cyanoprokaryota). Plant Syst. Evol.210:25–39.
29. Lawrie, R. A., and D. D. Stretch. 2011. Anthropogenic impacts on the waterand salt budgets of St. Lucia estuarine lake in South Africa. Estuar. Coast.Shelf Sci. 93:58–67.
30. Lumsden, T. G., R. E. Schulze, and B. C. Hewitson. 2008. Evaluation ofpotential changes in hydrologically relevant statistics of rainfall in SouthernAfrica under conditions of climate change. Water SA 35:649–656.
31. Marcarelli, A. M., W. A. Wurtsbaugh, and O. Griset. 2006. Salinity controlsphytoplankton response to nutrient enrichment in the Great Salt Lake, Utah,USA. Can. J. Fish. Aquat. Sci. 63:2236–2248.
32. Margheri, M. C., M. Bosco, L. Giovannetti, and S. Ventura. 1999. Assess-ment of the genetic diversity of halotolerant coccoid cyanobacteria usingamplified 16S rDNA restriction analysis. FEMS Microbiol. Lett. 173:9–16.
33. Moisander, P. H., E. McClinton III, and H. W. Paerl. 2002. Salinity effects ongrowth, photosynthetic parameters, and nitrogenase activity in estuarineplanktonic cyanobacteria. Microb. Ecol. 43:432–442.
34. Paerl, H. W. 1996. A comparison of cyanobacterial bloom dynamics infreshwater, estuarine and marine environments. Phycologia 35:25–35.
35. Paerl, H. W. 1988. Nuisance phytoplankton blooms in coastal, estuarine, andinland waters. Limnol. Oceanogr. 33:823–847.
36. Paerl, H. W., and J. Huisman. 2009. Climate change: a catalyst for globalexpansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep.1:27–37.
37. Perissinotto, R., D. Pillay, and G. Bate. 2010. Microalgal biomass in the St.Lucia Estuary during the 2004 to 2007 drought period. Mar. Ecol. Prog. Ser.405:147–161.
38. Philps, E. T., S. Badylak, and T. C. Lynch. 1999. Blooms of the picoplank-tonic cyanobacterium Synechococcus in Florida Bay, a sub-tropical inner-shelf lagoon. Limnol. Oceanogr. 44:1166–1175.
39. Pillay, D., and R. Perissinotto. 2008. The benthic macrofauna of the St.Lucia estuary during the 2005 drought year. Estuar. Coast. Shelf Sci.77:35–46.
40. Pillay, D., and R. Perissinotto. 2009. Community structure of epibenthicmeiofauna in the St. Lucia estuarine lake during a drought cycle. Estuar.Coast Shelf Sci. 81:94–104.
41. Plinski, M., H. Mazur-Marzec, T. Jozwiak, and J. Kobos. 2007. The potentialcauses of cyanobacterial blooms in Baltic Sea estuaries. Int. J. Oceanogr.Hydrobiol. 36:125–137.
42. Porta, D., R. Rippka, and M. Hernandez-Marine. 2000. Unusual ultrastruc-tural features in three strains of Cyanothece (cyanobacteria). Arch. Micro-biol. 173:154–163.
43. Reddy, K. J., J. B. Haskell, D. M. Sherman, and L. A. Sherman. 1993.Unicellular, aerobic nitrogen-fixing cyanobacteria of the genus Cyanothece.J. Bacteriol. 175:1284–1292.
44. Reynolds, E. S. 1963. The use of lead citrate at high pH as an electronopaque stain in electron microscopy. J. Cell Biol. 17:208–212.
45. Roussomoustakaki, M., and K. Anagnostidis. 1991. Cyanothece halobia, anew planktic chroococcalean cyanophyte from Hellenic heliothermal salt-works. Arch. Hydrobiol. Suppl. Algolog. Stud. 64:71–95.
46. Saitou, N., and M. Nei. 1987. The neighbor-joining method: a new methodfor reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406–425.
47. Schapira, M., M.-J. Buscot, T. Pollet, S. C. Leterme, and L. Seuront. 2010.Distribution of picophytoplankton communities from brackish to hypersalinecommunities in a South Australian lagoon. Saline Systems 6:2–15.
48. Spurr, A. R. 1969. A low viscosity epoxy resin embedding medium forelectron microscopy. J. Ultrastruct. Res. 26:31–43.
49. Stockwell, D. A., E. J. Buskey, and T. E. Whitledge. 1993. Studies on condi-tions conducive to the development and maintenance of a persistent “browntide” in Laguna Mardre, Texas, p. 693–698. In T. J. Smayda and Y. Shimizu(ed.), Toxic phytoplankton blooms in the sea. 5th International Conferenceon Toxic Marine Phytoplankton. Elsevier, Amsterdam, Netherlands.
50. Tamura, K., J. Dudley, M. Nei, and S. Kumar. 2007. MEGA4: molecularevolutionary genetics analysis (MEGA) software version 4. Mol. Biol. Evol.24:1569–1599.
51. Tamura, K., M. Nei, and S. Kumar. 2004. Prospects for inferring very largephylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci.U. S. A. 101:11030–11035.
52. Taylor, R. H., J. B. Adams, and S. Haldorsen. 2006. Primary habitats of theSt. Lucia estuarine system, South Africa, and their responses to mouthmanagement. Afr. J. Aquat. Sci. 31:31–41.
53. Van Rijn, J., and Y. Cohen. 1983. Ecophysiology of the cyanobacteriumDactylococcopsis salina: effect of light intensity, sulphide and temperature.J. Gen. Microbiol. 129:1849–1856.
54. Walsby, A. E., J. Van Rijn, and Y. Cohen. 1983. The biology of a newgas-vacuolate cyanobacterium, Dactylococcopsis salina sp. nov., in SolarLake. Proc. Roy. Soc. Lond. B Biol. Sci. 217:417–447.
55. Waterbury, J. B. 2006. The cyanobacteria: isolation, purification, and iden-tification, p. 1053–1073. In M. Dworkin, S. Falkow, E. Rosenberg, K.-H.Schleifer, and E. Stackebrandt (ed.), The prokaryotes: a handbook on thebiology of bacteria, vol. 4. Springer, New York, NY.
56. Waterbury, J. B., and R. Rippka. 1989. The order Chroococcales Wettstein1924, emend. Rippka et al., 1979, p. 1728–1746. In J. T. Staley, M. P. Bryant,N. Pfennig, and J. G. Holts (ed.), Bergey’s manual of systematic bacteriology,vol. 3. Williams and Wilkins, Baltimore, MD.
57. Welschmeyer, N. A. 1994. Fluorometric analysis of chlorophyll a in thepresence of chlorophyll b and phaeopigments. Limnol. Oceanogr. 39:1985–1992.
58. Whitfield, A., and R. H. Taylor. 2009. A review of the importance of fresh-water inflow to the future conservation of Lake St. Lucia. Aquat. Conserv.Mar. Freshw. Ecoyst. 19:838–848.
5896 MUIR AND PERISSINOTTO APPL. ENVIRON. MICROBIOL.
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from