phylogenetic relationships among passiflora species based...
TRANSCRIPT

MOLECULARPHYLOGENETICSAND
Molecular Phylogenetics and Evolution 31 (2004) 379–396
EVOLUTION
www.elsevier.com/locate/ympev
Phylogenetic relationships among Passiflora species basedon the glutamine synthetase nuclear gene expressed
in chloroplast (ncpGS)
Roxana Yockteng* and Sophie Nadot
Laboratoire �Ecologie, Syst�ematique et �Evolution, Universit�e Paris-XI, CNRS UMR 8079, Orsay, France
Received 21 April 2003; revised 10 July 2003
Abstract
This paper presents the first molecular phylogeny of the genus Passiflora encompassing almost all sections of this large genus. The
nuclear-encoded chloroplast-expressed glutamine synthetase gene (ncpGS) was used to examine the relationships among Passiflora
species (passionflowers), which was then compared with the new classification proposed by Feuillet and MacDougal. The resulting
Bayesian, likelihood, and parsimony trees are congruent and well supported. The 90 Passiflora species examined apparently split
into eight main subgenera: Plectostemma, Granadilla, Astrophea, Deidamioides, Polyanthea, Dysosmia, Tetrapathea, and Trypho-
stemmatoides. These results are in overall agreement with the Feuillet and MacDougal�s classification but here we propose that three
additional subgenera, Polyanthea, Dysosmia, and Tetrapathea, should be maintained. We observe a striking overall correlation
between the phylogenetic position of the different species and their chromosome number. The first clade contains the arborescent
species of the subgenus Astrophea, with n ¼ 12. The second clade, subgenus Plectostemma, includes species from four subgenera of
Killip�s classification with n ¼ 6 chromosomes. The last clade, subgenus Granadilla, includes species of seven old subgenera with
n ¼ 9. Subgenus Dysosmia, with a variable chromosome number of n ¼ 9–11, is considered here as a separate subgenus closely
related to the subgenus Granadilla.
� 2003 Elsevier Inc. All rights reserved.
Keywords: Passiflora; Phylogeny; ncpGS
1. Introduction
The genus Passiflora L. is well known for its com-
mercial uses. Many species are widely cultivated for fruit
production such as P. edulis Sims. (passion fruit),
P. tripartita var. mollissima (Kunth) Holm-Niel andJørg (banana passion fruit) or P. ligularis Juss. (sweet
granadilla). The sedative properties of P. incarnata L.
are exploited in the pharmaceutical industry. Several
other species are used as ornamental plants for their
intricate and richly coloured flowers. Furthermore, the
specific relationships with their herbivores, butterflies
of the Heliconiine group, have been widely studied
(Benson, 1978; Benson et al., 1975; Gilbert, 1980, 1982,1983; Spencer, 1986).
* Corresponding author. Fax: +33-1-69-15-73-53.
E-mail address: [email protected] (R. Yockteng).
1055-7903/$ - see front matter � 2003 Elsevier Inc. All rights reserved.
doi:10.1016/S1055-7903(03)00277-X
The Passifloraceae are distributed throughout the
tropics and many warm temperate areas, from humid
rain forests to deserts. It is classified in the order Mal-
phigiales and appear closely related to the Violaceae and
Flacourtiaceae (Chase et al., 2002; Soltis et al., 1999).
Passifloraceae are divided into two tribes, the Africantribe Pariopsae with 6 genera and the worldwide tribe
Passiflorae with 11 genera. Passiflora L. (tribe Passiflo-
rae) is the largest genus of the family with over 450
herbaceous, woody, and climbing plant species. The
genus Passiflora is mainly distributed in tropical
America with a few species in Asia and Oceania.
Passionflowers are usually vines, with alternate and
simple leaves, extrafloral nectaries on the petioles or theleaf surfaces. The large hermaphroditic flowers are
generally colourful and composed of five stamens, five
sepals, generally five petals, and three styles. The genus
is assumed to be monophyletic on the basis of three

380 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
diagnostic characters: the series of the corona filaments,axillary tendrils, and specialized flowers (Judd et al.,
1999).
The genus Passiflora was established in 1735 by
Linnaeus who described 22 species. There are now be-
tween 475 and 485 recognized species of Passiflora
(Vanderplanck, 2000). The first infrageneric classifica-
tion encompassing 355 species of the genus recognized
22 subgenera (Killip, 1938) (Table 1) based on variationin the following characters: shape and size of operculum,
number of series of filaments in the corona, size and
shape of calyx tube, and finally position, size, and shape
of the glands in petioles. A new subgenus and several
sections were subsequently added to the genus (Escobar,
1988, 1989). The old world Passiflora species were
studied by Cusset (1967) and deWilde (1972). More re-
cently, Feuillet and MacDougal (1999) proposed a newinfrageneric classification with only four subgenera:
Astrophea, Deidamioides, Passiflora, and Plectostemma.
They argued that many species are linked by strong
affinities and hence must be grouped. At present, the
number and nature of the subdivisions of this genus, as
well as the position of the monospecific Tetrapathea
group of New Zealand classified either as an indepen-
dent genus or as a subgenus of Passiflora, remain to beresolved.
Although studies of molecular genetic variation based
on RAPD and AFLP markers have been carried out on
the cultivated species of subgenera Tacsonia and Gra-
nadilla (Fajardo et al., 1998; S�anchez et al., 1999), no
molecular phylogenetic analysis encompassing all sub-
genera of Passiflora has been yet published. Therefore,
in order to clarify the evolutionary history of Passiflora,we sequenced the nuclear-encoded glutamine synthetase
gene expressed in the chloroplast (ncpGS EC 6.3.1.2)
and we constructed a molecular phylogeny of this gene
using parsimony, likelihood, and Bayesian methods.
Though variation of chloroplast genes and the widely
used nuclear ITS region (Internal Transcribed Spacers)
is often insufficient for resolving relationships among
closely related species, ncpGS is comparable and hasbeen proven useful for phylogenetic reconstruction and
identification of processes of reticulate evolution (Ems-
hwiller and Doyle, 1999, 2002). This gene belongs to a
multigene family responsible for the nitrogen metabo-
lism. It diverged 250–300Myr ago from the cytosolic-
expressed glutamine synthetase genes (Kumada et al.,
1993; Pesole et al., 1991) and plays an independent role
in the assimilation of the ammonia produced duringphotorespiration. Although the coding region is ex-
tremely conserved among angiosperms (1365 bp), the
total length of the gene ncpGS is highly variable due to
the presence of 11 introns distributed along the gene.
Our main objectives are: (a) to examine the subgen-
eric boundaries in Passiflora; (b) to test the monophyly
of the subgenera proposed by Killip (1938) and Feuillet
and MacDougal (1999); and finally (c) to propose apreliminary scenario of the evolutionary history of the
genus.
2. Materials and methods
2.1. Plant samples
For this study, 91 Passiflora species (138 individuals)
were selected, representing 17 of 23 subgenera recog-
nized by Killip (1938) and Escobar (1989) (Table 1). All
non-monospecific subgenera were represented by at least
two species with the exception of subgenera Psilanthus
and Deidamioides. We used two Adenia species and one
Dilkea species of the tribe Passiflorae (Passifloraceae) as
outgroups to root the trees. Specimens were obtainedfrom the wild, from botanical gardens, and from several
national collections (Table 2). Sampled leaves were
preserved in silica gel. Herbarium specimens were de-
posited at the Herbarium of the Museum National
d�Histoire Naturelle (Paris, France). To avoid confusion
between the section and subgenus Decaloba, we used the
name Plectostemma (Killip, 1938) for the subgenus. In
the same way, we used Granadilla for the also calledPassiflora subgenus and Incarnatae for the Passiflora
series (Table 2) (Killip, 1938). Since, the new classifica-
tion of Feuillet and MacDougal (1999) is not yet pub-
lished in full, the discussion is based on Killip�sclassification.
2.2. DNA extraction, amplification, and sequencing
For each sample, DNA was isolated from 0.2 g of dry
leaves. Extractions were completed using the Dneasy
Plant Mini Kit (Qiagen) following manufacturer�s in-
structions.
The primers designed by Emshwiller and Doyle
(1999) were tested on a selected subset of Passiflora
DNAs. These primers (GScp687f and GScp994r) were
designed to amplify the 50 region between positions 687and 994 bp of the coding region, which includes four
introns of variable lengths. We found that these primers
were not specific for the region ncpGS in Passiflora since
in some species the cytosolic glutamine synthetase was
amplified instead. The sequences of ncpGS obtained
were used to design a new specific pair of primers,
named GScp839f (50 CAC CAA TGG GGA GGT TAT
GC 30) and GScp1056r (50 CAT CTT CCC TCA TGCTCT TTG T 30). Fig. 1 represents the sequenced region
of the ncpGS gene, localized between positions 836 and
1061 of the coding region in Juglans nigra.
PCR amplifications were performed in a 40 ll volume
reaction using 1� buffer (Q-Biogene), 0.2U of Taq
polymerase (Q-Biogene), 1.5mM of MgCl2, 250 lMof dNTPs (Promega), and 0.2 lM of each primer.

Table 1
Infrageneric classification in alphabetic order of Passiflora proposed by Killip (1938) and modified by Escobar (1988, 1989)
Subgenus Section Series Number of species
Adenosepala, Killip, 1938 1
Apodogyne, Killip, 1938 1
Astephia, Killip, 1938 1
Astrophea (DC.) Masters, 1871 Botryastrophea 11
Cirrhipes 1
Dolichostemma 2
Euastrophea 15
Leptopoda 1
Pseudoastrophea 16
Calopathanthus (Harms) Killip, 1938 1
Chloropathanthus (Harms) Killip, 1938 2
Deidamioides (Harms) Killip, 1938 2
Distephana (Juss.) Killip, 1938 11
Dysosmia (DC.) Killip, 1938 11
Dysosmioides, Killip, 1938 4
Granadilla, (Medic.) Mast., 1871 Quadrangulares 2
Digitatae 1
Tiliafoliae 10
Marginatae 1
Laurifoliae 13
Serratifoliae 4
Setaceae 1
Pedatae 1
Incarnatae 6
Palmatisectae 1
Kermesinae 8
Imbricatae 2
Simplicifoliae 14
Lobatae 29
Menispermifoliae 4
Manicata (Harms) Escobar, 1988 5
Murucuja (Medic.) Mast, 1871 4
Plectostemma, Masters, 1871
Cieca 25
Decaloba
Auriculatae 2
Heterophyllae 2
Sexflorae 4
Ap�etalae 2
Luteae 3
Organenses 7
Miserae 5
Punctatae 47
Eudecaloba* 19
Polyanthea* 16
Hahniopathanthus 3
Hollrungiella* 1
Mayapathanthus 1
Pseudodysosmia 18
Pseudogranadilla 6
Xerogona 8
Polyanthea (DC.) Killip, 1938 1
Porphyropathantus Escobar, 1989 1
Pseudomurucuja (Harms) Killip, 1938 4
Psilanthus, Killip, 1938 4
Rathea (Karst.) Killip, 1938 2
Tacsonia (Juss.) Tr. & Planch, 1873 Ampullacea 1
Boliviana 1
Bracteogama 12
Colombiana Leptomischae 7
Colombiana Colombianae 8
Fimbriatistipula 2
Parritana 1
R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 381
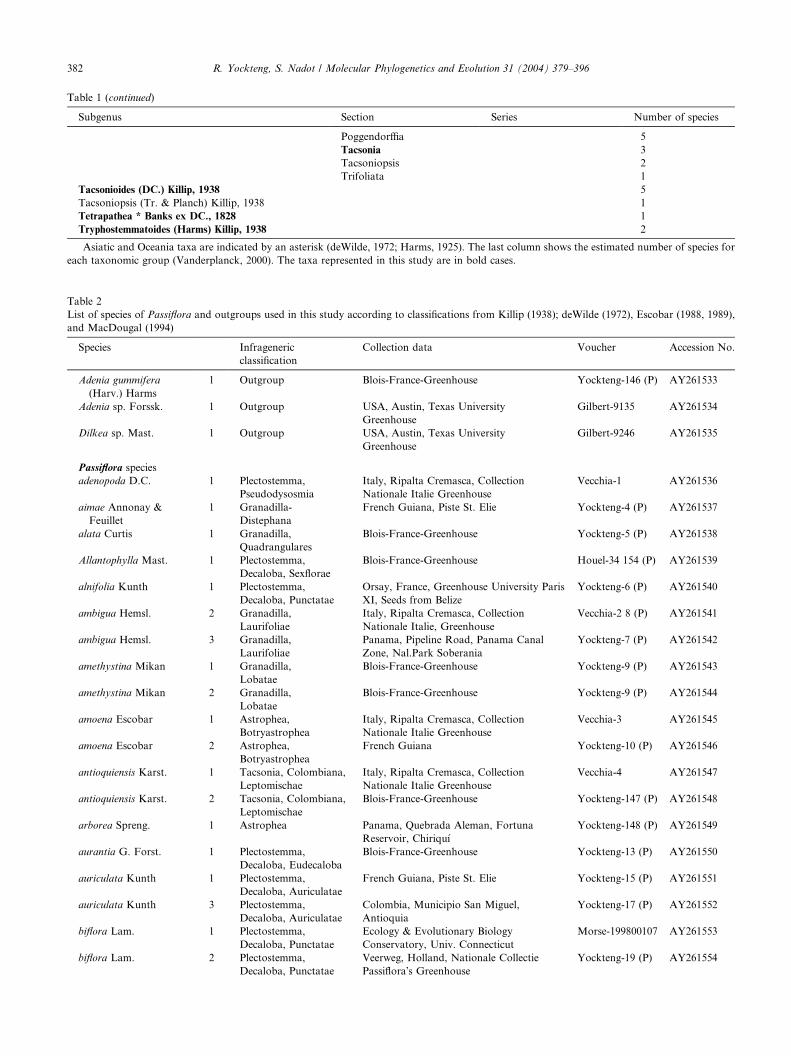
Table 1 (continued)
Subgenus Section Series Number of species
Poggendorffia 5
Tacsonia 3
Tacsoniopsis 2
Trifoliata 1
Tacsonioides (DC.) Killip, 1938 5
Tacsoniopsis (Tr. & Planch) Killip, 1938 1
Tetrapathea * Banks ex DC., 1828 1
Tryphostemmatoides (Harms) Killip, 1938 2
Asiatic and Oceania taxa are indicated by an asterisk (deWilde, 1972; Harms, 1925). The last column shows the estimated number of species for
each taxonomic group (Vanderplanck, 2000). The taxa represented in this study are in bold cases.
Table 2
List of species of Passiflora and outgroups used in this study according to classifications from Killip (1938); deWilde (1972), Escobar (1988, 1989),
and MacDougal (1994)
Species Infrageneric
classification
Collection data Voucher Accession No.
Adenia gummifera
(Harv.) Harms
1 Outgroup Blois-France-Greenhouse Yockteng-146 (P) AY261533
Adenia sp. Forssk. 1 Outgroup USA, Austin, Texas University
Greenhouse
Gilbert-9135 AY261534
Dilkea sp. Mast. 1 Outgroup USA, Austin, Texas University
Greenhouse
Gilbert-9246 AY261535
Passiflora species
adenopoda D.C. 1 Plectostemma,
Pseudodysosmia
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-1 AY261536
aimae Annonay &
Feuillet
1 Granadilla-
Distephana
French Guiana, Piste St. Elie Yockteng-4 (P) AY261537
alata Curtis 1 Granadilla,
Quadrangulares
Blois-France-Greenhouse Yockteng-5 (P) AY261538
Allantophylla Mast. 1 Plectostemma,
Decaloba, Sexflorae
Blois-France-Greenhouse Houel-34 154 (P) AY261539
alnifolia Kunth 1 Plectostemma,
Decaloba, Punctatae
Orsay, France, Greenhouse University Paris
XI, Seeds from Belize
Yockteng-6 (P) AY261540
ambigua Hemsl. 2 Granadilla,
Laurifoliae
Italy, Ripalta Cremasca, Collection
Nationale Italie, Greenhouse
Vecchia-2 8 (P) AY261541
ambigua Hemsl. 3 Granadilla,
Laurifoliae
Panama, Pipeline Road, Panama Canal
Zone, Nal.Park Soberania
Yockteng-7 (P) AY261542
amethystina Mikan 1 Granadilla,
Lobatae
Blois-France-Greenhouse Yockteng-9 (P) AY261543
amethystina Mikan 2 Granadilla,
Lobatae
Blois-France-Greenhouse Yockteng-9 (P) AY261544
amoena Escobar 1 Astrophea,
Botryastrophea
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-3 AY261545
amoena Escobar 2 Astrophea,
Botryastrophea
French Guiana Yockteng-10 (P) AY261546
antioquiensis Karst. 1 Tacsonia, Colombiana,
Leptomischae
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-4 AY261547
antioquiensis Karst. 2 Tacsonia, Colombiana,
Leptomischae
Blois-France-Greenhouse Yockteng-147 (P) AY261548
arborea Spreng. 1 Astrophea Panama, Quebrada Aleman, Fortuna
Reservoir, Chiriqu�ıYockteng-148 (P) AY261549
aurantia G. Forst. 1 Plectostemma,
Decaloba, Eudecaloba
Blois-France-Greenhouse Yockteng-13 (P) AY261550
auriculata Kunth 1 Plectostemma,
Decaloba, Auriculatae
French Guiana, Piste St. Elie Yockteng-15 (P) AY261551
auriculata Kunth 3 Plectostemma,
Decaloba, Auriculatae
Colombia, Municipio San Miguel,
Antioquia
Yockteng-17 (P) AY261552
biflora Lam. 1 Plectostemma,
Decaloba, Punctatae
Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-199800107 AY261553
biflora Lam. 2 Plectostemma,
Decaloba, Punctatae
Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-19 (P) AY261554
382 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
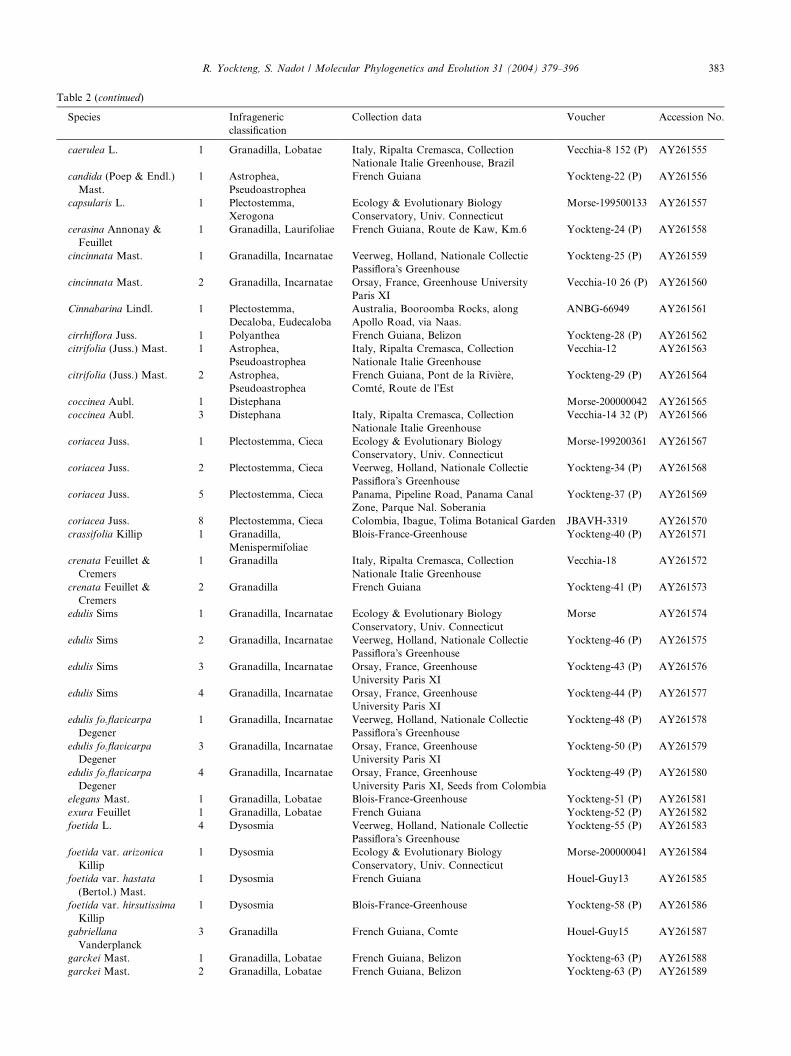
Table 2 (continued)
Species Infrageneric
classification
Collection data Voucher Accession No.
caerulea L. 1 Granadilla, Lobatae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse, Brazil
Vecchia-8 152 (P) AY261555
candida (Poep & Endl.)
Mast.
1 Astrophea,
Pseudoastrophea
French Guiana Yockteng-22 (P) AY261556
capsularis L. 1 Plectostemma,
Xerogona
Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-199500133 AY261557
cerasina Annonay &
Feuillet
1 Granadilla, Laurifoliae French Guiana, Route de Kaw, Km.6 Yockteng-24 (P) AY261558
cincinnata Mast. 1 Granadilla, Incarnatae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-25 (P) AY261559
cincinnata Mast. 2 Granadilla, Incarnatae Orsay, France, Greenhouse University
Paris XI
Vecchia-10 26 (P) AY261560
Cinnabarina Lindl. 1 Plectostemma,
Decaloba, Eudecaloba
Australia, Booroomba Rocks, along
Apollo Road, via Naas.
ANBG-66949 AY261561
cirrhiflora Juss. 1 Polyanthea French Guiana, Belizon Yockteng-28 (P) AY261562
citrifolia (Juss.) Mast. 1 Astrophea,
Pseudoastrophea
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-12 AY261563
citrifolia (Juss.) Mast. 2 Astrophea,
Pseudoastrophea
French Guiana, Pont de la Rivi�ere,
Comt�e, Route de l�EstYockteng-29 (P) AY261564
coccinea Aubl. 1 Distephana Morse-200000042 AY261565
coccinea Aubl. 3 Distephana Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-14 32 (P) AY261566
coriacea Juss. 1 Plectostemma, Cieca Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-199200361 AY261567
coriacea Juss. 2 Plectostemma, Cieca Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-34 (P) AY261568
coriacea Juss. 5 Plectostemma, Cieca Panama, Pipeline Road, Panama Canal
Zone, Parque Nal. Soberania
Yockteng-37 (P) AY261569
coriacea Juss. 8 Plectostemma, Cieca Colombia, Ibague, Tolima Botanical Garden JBAVH-3319 AY261570
crassifolia Killip 1 Granadilla,
Menispermifoliae
Blois-France-Greenhouse Yockteng-40 (P) AY261571
crenata Feuillet &
Cremers
1 Granadilla Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-18 AY261572
crenata Feuillet &
Cremers
2 Granadilla French Guiana Yockteng-41 (P) AY261573
edulis Sims 1 Granadilla, Incarnatae Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse AY261574
edulis Sims 2 Granadilla, Incarnatae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-46 (P) AY261575
edulis Sims 3 Granadilla, Incarnatae Orsay, France, Greenhouse
University Paris XI
Yockteng-43 (P) AY261576
edulis Sims 4 Granadilla, Incarnatae Orsay, France, Greenhouse
University Paris XI
Yockteng-44 (P) AY261577
edulis fo.flavicarpa
Degener
1 Granadilla, Incarnatae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-48 (P) AY261578
edulis fo.flavicarpa
Degener
3 Granadilla, Incarnatae Orsay, France, Greenhouse
University Paris XI
Yockteng-50 (P) AY261579
edulis fo.flavicarpa
Degener
4 Granadilla, Incarnatae Orsay, France, Greenhouse
University Paris XI, Seeds from Colombia
Yockteng-49 (P) AY261580
elegans Mast. 1 Granadilla, Lobatae Blois-France-Greenhouse Yockteng-51 (P) AY261581
exura Feuillet 1 Granadilla, Lobatae French Guiana Yockteng-52 (P) AY261582
foetida L. 4 Dysosmia Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-55 (P) AY261583
foetida var. arizonica
Killip
1 Dysosmia Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-200000041 AY261584
foetida var. hastata
(Bertol.) Mast.
1 Dysosmia French Guiana Houel-Guy13 AY261585
foetida var. hirsutissima
Killip
1 Dysosmia Blois-France-Greenhouse Yockteng-58 (P) AY261586
gabriellana
Vanderplanck
3 Granadilla French Guiana, Comte Houel-Guy15 AY261587
garckei Mast. 1 Granadilla, Lobatae French Guiana, Belizon Yockteng-63 (P) AY261588
garckei Mast. 2 Granadilla, Lobatae French Guiana, Belizon Yockteng-63 (P) AY261589
R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 383
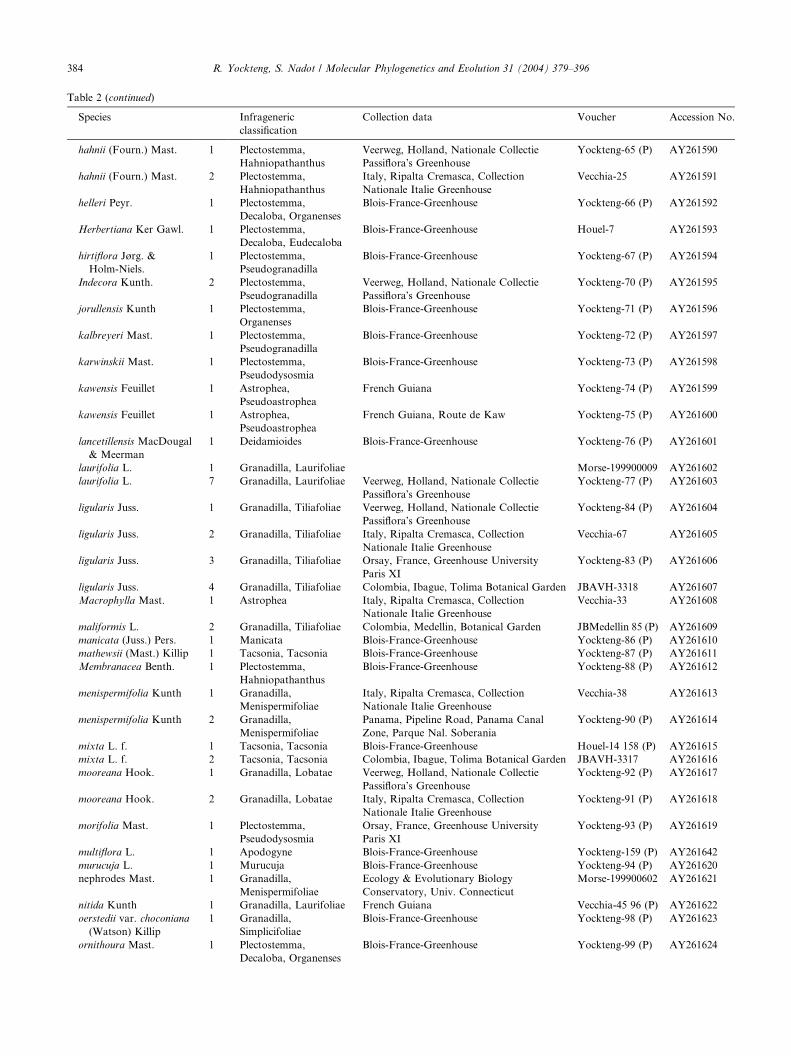
Table 2 (continued)
Species Infrageneric
classification
Collection data Voucher Accession No.
hahnii (Fourn.) Mast. 1 Plectostemma,
Hahniopathanthus
Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-65 (P) AY261590
hahnii (Fourn.) Mast. 2 Plectostemma,
Hahniopathanthus
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-25 AY261591
helleri Peyr. 1 Plectostemma,
Decaloba, Organenses
Blois-France-Greenhouse Yockteng-66 (P) AY261592
Herbertiana Ker Gawl. 1 Plectostemma,
Decaloba, Eudecaloba
Blois-France-Greenhouse Houel-7 AY261593
hirtiflora Jørg. &
Holm-Niels.
1 Plectostemma,
Pseudogranadilla
Blois-France-Greenhouse Yockteng-67 (P) AY261594
Indecora Kunth. 2 Plectostemma,
Pseudogranadilla
Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-70 (P) AY261595
jorullensis Kunth 1 Plectostemma,
Organenses
Blois-France-Greenhouse Yockteng-71 (P) AY261596
kalbreyeri Mast. 1 Plectostemma,
Pseudogranadilla
Blois-France-Greenhouse Yockteng-72 (P) AY261597
karwinskii Mast. 1 Plectostemma,
Pseudodysosmia
Blois-France-Greenhouse Yockteng-73 (P) AY261598
kawensis Feuillet 1 Astrophea,
Pseudoastrophea
French Guiana Yockteng-74 (P) AY261599
kawensis Feuillet 1 Astrophea,
Pseudoastrophea
French Guiana, Route de Kaw Yockteng-75 (P) AY261600
lancetillensis MacDougal
& Meerman
1 Deidamioides Blois-France-Greenhouse Yockteng-76 (P) AY261601
laurifolia L. 1 Granadilla, Laurifoliae Morse-199900009 AY261602
laurifolia L. 7 Granadilla, Laurifoliae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-77 (P) AY261603
ligularis Juss. 1 Granadilla, Tiliafoliae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-84 (P) AY261604
ligularis Juss. 2 Granadilla, Tiliafoliae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-67 AY261605
ligularis Juss. 3 Granadilla, Tiliafoliae Orsay, France, Greenhouse University
Paris XI
Yockteng-83 (P) AY261606
ligularis Juss. 4 Granadilla, Tiliafoliae Colombia, Ibague, Tolima Botanical Garden JBAVH-3318 AY261607
Macrophylla Mast. 1 Astrophea Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-33 AY261608
maliformis L. 2 Granadilla, Tiliafoliae Colombia, Medellin, Botanical Garden JBMedellin 85 (P) AY261609
manicata (Juss.) Pers. 1 Manicata Blois-France-Greenhouse Yockteng-86 (P) AY261610
mathewsii (Mast.) Killip 1 Tacsonia, Tacsonia Blois-France-Greenhouse Yockteng-87 (P) AY261611
Membranacea Benth. 1 Plectostemma,
Hahniopathanthus
Blois-France-Greenhouse Yockteng-88 (P) AY261612
menispermifolia Kunth 1 Granadilla,
Menispermifoliae
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-38 AY261613
menispermifolia Kunth 2 Granadilla,
Menispermifoliae
Panama, Pipeline Road, Panama Canal
Zone, Parque Nal. Soberania
Yockteng-90 (P) AY261614
mixta L. f. 1 Tacsonia, Tacsonia Blois-France-Greenhouse Houel-14 158 (P) AY261615
mixta L. f. 2 Tacsonia, Tacsonia Colombia, Ibague, Tolima Botanical Garden JBAVH-3317 AY261616
mooreana Hook. 1 Granadilla, Lobatae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-92 (P) AY261617
mooreana Hook. 2 Granadilla, Lobatae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Yockteng-91 (P) AY261618
morifolia Mast. 1 Plectostemma,
Pseudodysosmia
Orsay, France, Greenhouse University
Paris XI
Yockteng-93 (P) AY261619
multiflora L. 1 Apodogyne Blois-France-Greenhouse Yockteng-159 (P) AY261642
murucuja L. 1 Murucuja Blois-France-Greenhouse Yockteng-94 (P) AY261620
nephrodes Mast. 1 Granadilla,
Menispermifoliae
Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-199900602 AY261621
nitida Kunth 1 Granadilla, Laurifoliae French Guiana Vecchia-45 96 (P) AY261622
oerstedii var. choconiana
(Watson) Killip
1 Granadilla,
Simplicifoliae
Blois-France-Greenhouse Yockteng-98 (P) AY261623
ornithoura Mast. 1 Plectostemma,
Decaloba, Organenses
Blois-France-Greenhouse Yockteng-99 (P) AY261624
384 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
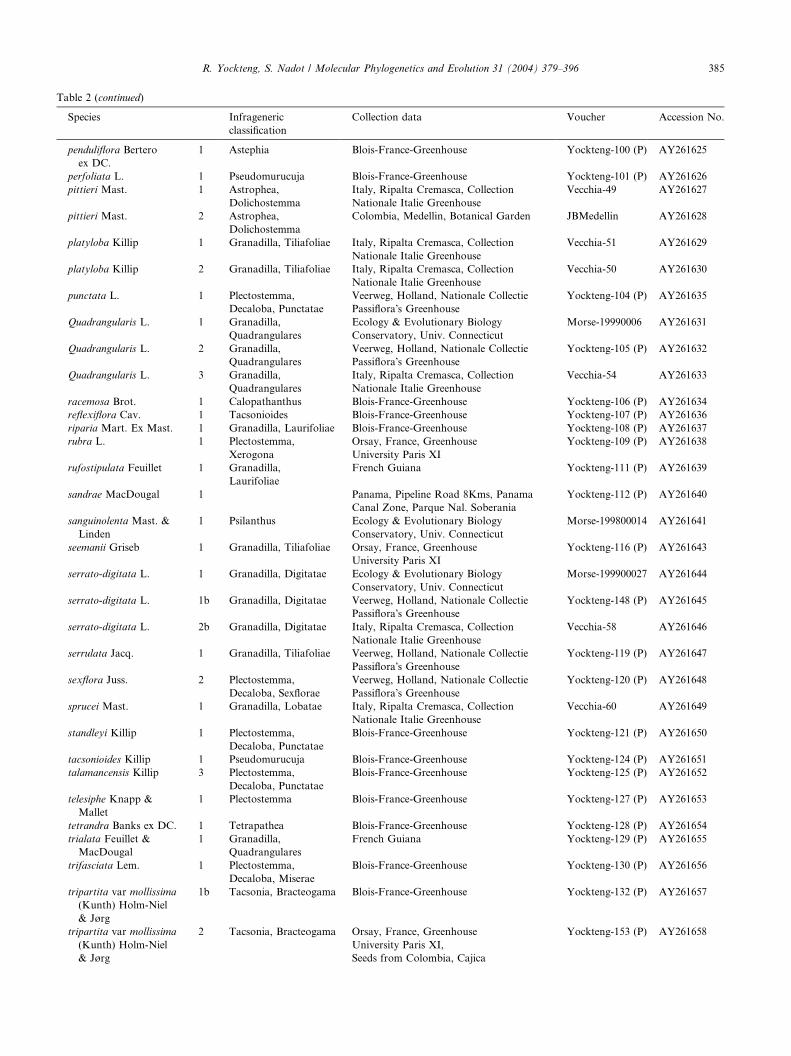
Table 2 (continued)
Species Infrageneric
classification
Collection data Voucher Accession No.
penduliflora Bertero
ex DC.
1 Astephia Blois-France-Greenhouse Yockteng-100 (P) AY261625
perfoliata L. 1 Pseudomurucuja Blois-France-Greenhouse Yockteng-101 (P) AY261626
pittieri Mast. 1 Astrophea,
Dolichostemma
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-49 AY261627
pittieri Mast. 2 Astrophea,
Dolichostemma
Colombia, Medellin, Botanical Garden JBMedellin AY261628
platyloba Killip 1 Granadilla, Tiliafoliae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-51 AY261629
platyloba Killip 2 Granadilla, Tiliafoliae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-50 AY261630
punctata L. 1 Plectostemma,
Decaloba, Punctatae
Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-104 (P) AY261635
Quadrangularis L. 1 Granadilla,
Quadrangulares
Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-19990006 AY261631
Quadrangularis L. 2 Granadilla,
Quadrangulares
Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-105 (P) AY261632
Quadrangularis L. 3 Granadilla,
Quadrangulares
Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-54 AY261633
racemosa Brot. 1 Calopathanthus Blois-France-Greenhouse Yockteng-106 (P) AY261634
reflexiflora Cav. 1 Tacsonioides Blois-France-Greenhouse Yockteng-107 (P) AY261636
riparia Mart. Ex Mast. 1 Granadilla, Laurifoliae Blois-France-Greenhouse Yockteng-108 (P) AY261637
rubra L. 1 Plectostemma,
Xerogona
Orsay, France, Greenhouse
University Paris XI
Yockteng-109 (P) AY261638
rufostipulata Feuillet 1 Granadilla,
Laurifoliae
French Guiana Yockteng-111 (P) AY261639
sandrae MacDougal 1 Panama, Pipeline Road 8Kms, Panama
Canal Zone, Parque Nal. Soberania
Yockteng-112 (P) AY261640
sanguinolenta Mast. &
Linden
1 Psilanthus Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-199800014 AY261641
seemanii Griseb 1 Granadilla, Tiliafoliae Orsay, France, Greenhouse
University Paris XI
Yockteng-116 (P) AY261643
serrato-digitata L. 1 Granadilla, Digitatae Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-199900027 AY261644
serrato-digitata L. 1b Granadilla, Digitatae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-148 (P) AY261645
serrato-digitata L. 2b Granadilla, Digitatae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-58 AY261646
serrulata Jacq. 1 Granadilla, Tiliafoliae Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-119 (P) AY261647
sexflora Juss. 2 Plectostemma,
Decaloba, Sexflorae
Veerweg, Holland, Nationale Collectie
Passiflora�s Greenhouse
Yockteng-120 (P) AY261648
sprucei Mast. 1 Granadilla, Lobatae Italy, Ripalta Cremasca, Collection
Nationale Italie Greenhouse
Vecchia-60 AY261649
standleyi Killip 1 Plectostemma,
Decaloba, Punctatae
Blois-France-Greenhouse Yockteng-121 (P) AY261650
tacsonioides Killip 1 Pseudomurucuja Blois-France-Greenhouse Yockteng-124 (P) AY261651
talamancensis Killip 3 Plectostemma,
Decaloba, Punctatae
Blois-France-Greenhouse Yockteng-125 (P) AY261652
telesiphe Knapp &
Mallet
1 Plectostemma Blois-France-Greenhouse Yockteng-127 (P) AY261653
tetrandra Banks ex DC. 1 Tetrapathea Blois-France-Greenhouse Yockteng-128 (P) AY261654
trialata Feuillet &
MacDougal
1 Granadilla,
Quadrangulares
French Guiana Yockteng-129 (P) AY261655
trifasciata Lem. 1 Plectostemma,
Decaloba, Miserae
Blois-France-Greenhouse Yockteng-130 (P) AY261656
tripartita var mollissima
(Kunth) Holm-Niel
& Jørg
1b Tacsonia, Bracteogama Blois-France-Greenhouse Yockteng-132 (P) AY261657
tripartita var mollissima
(Kunth) Holm-Niel
& Jørg
2 Tacsonia, Bracteogama Orsay, France, Greenhouse
University Paris XI,
Seeds from Colombia, Cajica
Yockteng-153 (P) AY261658
R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 385

Table 2 (continued)
Species Infrageneric
classification
Collection data Voucher Accession No.
tripartita var mollissima
(Kunth) Holm-Niel
& Jørg
3 Tacsonia, Bracteogama Colombia, La Calera Yockteng-131 (P) AY261659
tripartita var mollissima
(Kunth) Holm-Niel
& Jørg
4 Tacsonia, Bracteogama Colombia, La Calera Yockteng-131 (P) AY261660
tripartita var mollissima
(Kunth) Holm-Niel
& Jørg
1 Tacsonia, Bracteogama Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-Student AY261661
trisecta Mast. 1 Manicata Blois-France-Greenhouse Yockteng-133 (P) AY261662
tryphostemmatoides
Harms
2 Tryphostemmatoides Colombia-Valle del Cauca Yumbo
Monta~nitas KM6 Via Pavitas San Jose
Yockteng-134 (P) AY261663
tulae Urb 1 Murucuja Blois-France-Greenhouse Yockteng-136 (P) AY261664
umbilicata (Griseb.)
Harms
1 Tacsonioides Blois-France-Greenhouse Houel-27 157 (P) AY261665
variolata Poep & Endl. 1 Distephana Blois-France-Greenhouse Houel-30 AY261666
vespertilio L. 3 Plectostemma,
Decaloba, Punctatae
French Guiana, Crique Plomb Yockteng-137 (P) AY261667
vitifolia (Harv.) Harms 1 Distephana Ecology & Evolutionary Biology
Conservatory, Univ. Connecticut
Morse-198700236 AY261668
vitifolia (Harv.) Harms 5 Distephana Colombia, Ibague, Tolima Botanical Garden JBAVH-3320 AY261669
Collection data, voucher, and GenBank accessions numbers are given. Specimens are deposited at the Herbarium of the Museum National
d�Histoire Naturelle (Paris, France (P)). JBMedellin, Botanical Garden Medellin, Colombia; JBAVH, Botanical Garden Alejandro Von Humboldt,
Tolima, Colombia; Morse, University of Connecticut, USA; Houel, Passiflora National Collection, Blois, France; Vecchia, Passiflora National
Collection, Italy; ANBG, Australian National Botanical Gardens; Gilbert, Passiflora collection, Greenhouse University Texas, Austin.
Fig. 1. Location of the amplified and sequenced region of the chloroplast-expressed glutamine synthetase (ncpGS) for Passiflora. The intron-exon
distribution is with respect to the ncpGS sequence of Juglans nigra (GenBank Accession No. AF169795).
386 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
The thermal cycler (PTC-100 MJ-Research) was pro-
grammed for an initial step of 5min at 94 �C, followedby 35 cycles of 1min at 94 �C, 45 s at 55 �C, 1.5min at
72 �C, and a final step of 7min at 72 �C. PCR productswere visualized on a 2% agarose gel, purified using
the PEG precipitation protocol (Rosenthal et al., 1993),
and both strands were sequenced using the same
primer combination as for PCR amplifications. Cycle
sequencing products were run on ABI capillary se-
quencers (MWG Biotech). Sequences were deposited in
GenBank.
2.3. Phylogenetic analyses
Clustal X (Thompson et al., 1997) was used to pro-
duce a first alignment, which was corrected manually
using the Bioedit program (Hall, 1999). The indels of the
non-coding regions were coded using GapCoder, a
program developed by Neil Young and based on the
Simmons and Ochoterena�s method that codes indels as
separate characters in a data matrix (Simmons and
Ochoterena, 2000).
We examined nucleotide and translated sequences tosearch for patterns characteristic of specific taxonomic
groups.
The significance of the taxonomic group effect on
intron length was tested by one-way ANOVA (Excel,
Microsoft).
Phylogenetic reconstruction was carried out using
either the whole alignment or two subsets of this align-
ment, one consisting of the coding regions, the otherconsisting of the non-coding regions (introns).
2.4. Parsimony
Maximum parsimony (MP) analyses were conducted
using PAUP 4.0b10a* (Swofford, 2001). Heuristic sear-
ches were conducted with tree-bisection reconnection

R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 387
(TBR) branch swapping, multiple trees ON, and 1000random taxon addition replicates limiting the rear-
rangements to 100,000 per replicate and saving all most
parsimonious trees. Support for internal nodes was as-
sessed by bootstrapping (100 replicates). Decay indexes
were calculated using the Autodecay 4.0 program (Eri-
ksson, 1998).
2.5. Maximum likelihood
Due to the large number of taxa included in our study
we conducted the maximum likelihood (ML) analyses
using the Quartet Puzzling method (Strimmer and vo-
nHaeseler, 1996) implemented in PAUP 4.0b10a*
(Swofford, 2001), which takes a reasonable calculation
time. We used 1000 steps and the model of evolution
GTR+ I+G with a gamma shape parameter equal to1.9758, estimated using Modeltest version 3.06 (Posada,
2001). This model was used in the analyses based on the
whole alignment and on the intron subset. For the exon
subset, we specified that the sequence corresponded to a
coding region.
2.6. Bayesian approach
Additionally, a Bayesian analysis was conducted
with MrBayes 2.01 program (Huelsenbeck and Ron-
quist, 2001). We used uniform, prior probabilities, the
general time reversible + I+G model of molecular
evolution, and a random starting tree. Four chains of
the Markov Chain Monte Carlo were run simulta-
neously, sampled every 100 generations for a total of
150,000 generations. Stationarity conditions werereached around generation 82,000; thus the first 820
trees were burnt in of the chain (eliminated). The
majority rule consensus tree was calculated with PAUP
4.0b10a* (Swofford, 2001). Phylogenetic inferences
were based on this consensus.
2.7. Character evolution
The patterns of phenotypic evolution based on 19
morphological characters and the divergence in the
chromosome numbers were assessed by overlaying the
different states on the subgenera and outgroups bran-
ches. The character-state evolution was reconstructed
using parsimony method in MacClade 3.08 program
(Maddison and Maddison, 1992). The morphological
characters selected consist principally of the charactersused by Killip (1938) to establish the different subgen-
era. The character states of different subgenera were
established based on Killip (1938), deWilde (1974),
MacDougal (1994), Escobar (1988, 1994), Cervi (1997),
Vanderplanck (2000), and several floras. We used the
outgroup states as the ancestral states (deWilde, 1974;
Killip, 1938).
3. Results
3.1. Characteristics of ncpGS sequences in Passiflora
The primers used by Emshwiller and Doyle (1999)
resulted in the amplification, in several Passiflora spe-
cies, of the cytosolic-expressed glutamine synthetase
(cytGS) instead of the expected chloroplast-expressed
glutamine synthetase (ncpGS). These preliminary resultsrevealed a first division within the genus Passiflora. The
cytosolic glutamine synthetase was amplified for species
belonging to two subgenera: Murucuja and Plec-
tostemma. The chloroplast-expressed glutamine synthe-
tase (ncpGS) was amplified for all other species
belonging to the subgenera Distephana, Granadilla,
Tacsonia, and Tacsonoides.
The newly designed pair of primers allowed specificamplification of the ncpGS gene for all Passiflora spe-
cies. The sequences length ranges between 401 and
659 bp. The final alignment comprises 814 positions. The
coding subset includes 226 characters, corresponding to
75 amino acids and the intron subset comprises 587
positions.
The exons of the ncpGS region analyzed are quite
conserved but nonetheless they present motifs contain-ing taxonomic information. As an example, the 46th
amino acid splits the species into two groups. The first
group, holding all species of the subgenera Deidamio-
ides, Distephana, Granadilla, Polyanthea, Tacsonia, and
Tetrapathea, excepting the species representing the series
Lobatae and Menispermifoliae (subgenus Granadilla),
present a Serine (S). At this position, all the other species
except P. rubra and P. sexflora, and the 13 species ofLobatae and Menispermifoliae share a Threonine (T). A
similar pattern is observed with positions 22, 27, and 44.
The introns, with their numerous indels, display a
reasonable variability, suitable for phylogenetic infer-
ence. The mean pairwise intron divergence of 0.147 is
three times higher than the exon divergence (0.043).
Only a few indels were due to the outgroup. The ma-
jority were produced by the ingroup alignment. ANO-VA tests show that the differences in total intron length
among subgenera are significant (Table 3). The non-
coding region was significantly longer in subgenera
Astrophea, Distephana, Dysosmia, Granadilla, Manicata,
Tacsonia, and Tacsonoides, than in Murucuja, Plec-
tostemma, and Pseudomurucuja. Moreover, the length of
the first intron of Astrophea was significantly different
from that of the other subgenera.The analyses did not include the individuals of sub-
genera Calopathanthus, Psilanthus, Polyanthea, Deida-
mioides, Tetrapathea, Apodogyne, and Astephia because
the number of individuals available in these subgenera
was less than two. However, the intron lengths of Cal-
opathanthus, Psilanthus, Polyanthea, Deidamioides, Tet-
rapathea, Tryphostemmatoides, and the outgroup were

Table 3
ANOVA test of the significance of total intron length differences among the different subgenera
N ¼ 108 df Mean square F P > F
Subgenera 9 14400.59
Subgenera� total intron length 98 230.75 62.4 1.27E) 36
Error 107 152219.07
388 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
similar to the third group. In contrast, the intron length
of Apodogyne and Astephia species was closer to that of
the second group.
3.2. Phylogenetic analysis
The total alignment displayed 329 parsimony-infor-
mative sites and 356 invariable sites. Most informativesites were located in the intron regions (272 sites). The
number of informative sites rose to 412 when the indels
were coded.
MP analyses of the whole alignment (coding and non-
coding regions) yielded 32,573 most parsimonious trees
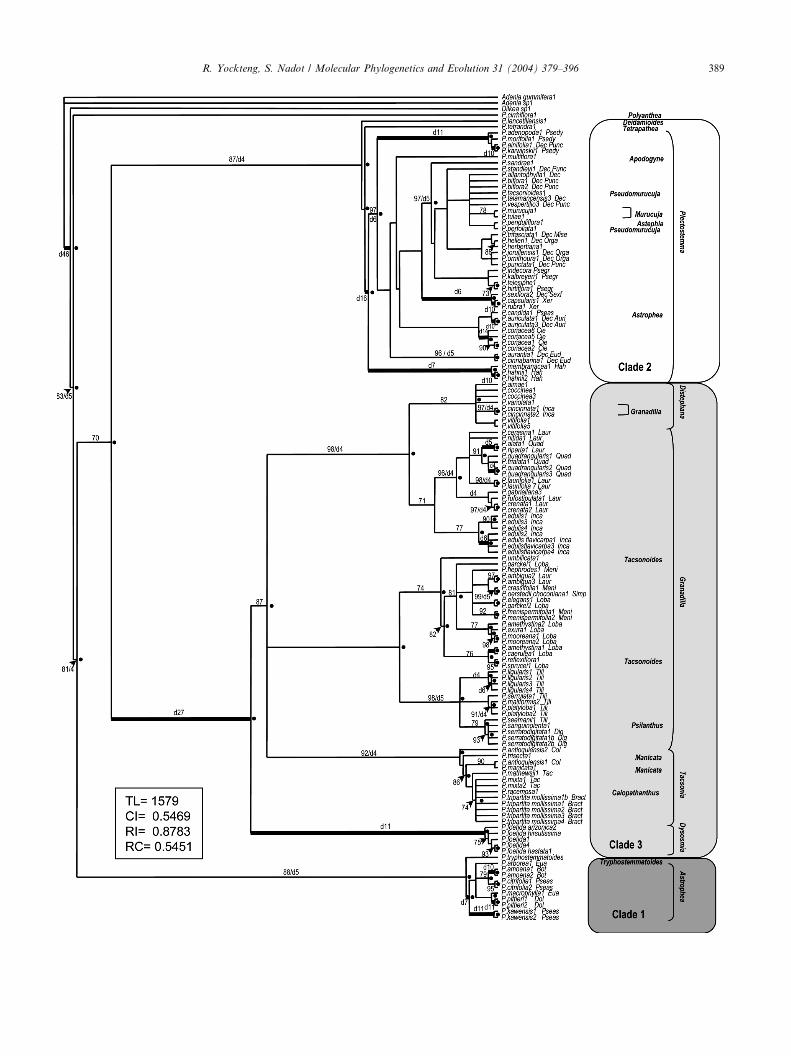
of 1579 steps. The strict consensus tree (Fig. 2) is highly
resolved and most of the nodes are well supported, as
shown both by the bootstrap values and decay indexes.The topology did not change using coded or not coded
indels. However, the indels were important to identify
the main groups, and for this reason the analyses were
performed with coded indels. Both maximum likelihood
and Bayesian analyses resulted in trees well supported
and highly congruent with the MP consensus tree.
Separate analyses of the coding and non-coding
regions produced less resolved trees. For each datasetthe different methods used yielded similar topologies.
The non-coding region produced topologies very
similar to the trees obtained using the entire align-
ment, although some nodes were less supported. When
using the coding region, only the internal nodes de-
fining the main taxonomic groups were well sup-
ported.
The individuals of the same species are branchedtogether or they are at least in the same clade in the tree.
The overall topology revealed three well-supported
clades (Fig. 2), each including one of the three largest
subgenera: Plectostemma, Granadilla and Astrophea.
Clade 1 is composed of the subgenera Astrophea and
Tryphostemmatoides; clade 2 contains the species of the
subgenera Astephia, Deidamioides, Murucuja, Plec-
tostemma, Pseudomurucuja, and Tetrapathea; and clade
Fig. 2. Strict consensus tree of 32,573 most parsimonious trees based on the
quences. Bootstrap support values (BS) greater than 70% and decay indices
values of 100%, dots indicate nodes supported by 100% in the Bayesian tree. T
The sections and series names are abbreviated as follows: Cie, Cieca; Dec, D
Miserae; Punc, Punctatae; Eud, Eudecaloba; Xero, Xerogona; Psedy, Pseu
Leptomischae; Col, Colombianae; Bract, Bracteogama; Tac, Tacsonia; Quad
Inca, Incarnatae; Simp, Simplicifoliae; Loba, Lobatae; Meni, Menispermifoli
The three clades emerging from the analysis are indicated on the right.
3 consists of the subgenera Calopathanthus, Distephana,
Dysosmia, Granadilla, Manicata, Psilanthus, Tacsonia,
and Tacsonoides. P. cirrhiflora Juss., from the mono-
specific Polyanthea subgenus, is sister group to all other
Passiflora species.
Figs. 3a–c present the evolution of 19 morphological
characters and the chromosome numbers in different
subgenera. These reconstructions helped us to supportor reject the different clades.
4. Discussion
The chloroplast-expressed glutamine synthetase gene
proved its usefulness for resolving the relationships
among species of the genus Passiflora. In spite of therelatively short length of the ncpGS alignment, the level
of variation was satisfactory with 329 parsimony-infor-
mative sites (40%) compared to other DNA regions.
For example, the alignment of ITS sequences for 36
Passiflora species (GenBank Accession Nos. AY1023461–
AY1023831, AY0328231–AY0328431, AY0327811–
AY0328011) presents 21% of informative sites. The
alignment of 38 sequences of the chloroplast trnL–trnFregion from Passiflora species (GenBank Accession Nos.
AY0327601–AY0327801, AY1023861–AY1024031)
displays 11% of informative sites. The comparison of
ncpGS, cytosol-expressed glutamine synthetase (cytGS),
and the chloroplastic gene matK shows that the ncpGS
has a higher level of divergence than matK and gives an
acceptable phylogenetic signal (Yockteng and Nadot,
2003). As expected at this taxonomic level, the non-coding region provides most of the phylogenetic infor-
mation. Interestingly, the length of the introns proved
particularly informative, creating indels in the alignment
that had a great impact on the resolution of the trees.
Although the coding region was highly conserved, the
information it contained allowed identification of clear,
well supported major taxonomic divisions within the
genus.
alignment of chloroplast-expressed glutamine synthetase (ncpGS) se-
greater than 4 are indicated above branches. Thick lines indicate BS
he subdivisions according to Killip (1938) are indicated for each taxon.
ecaloba; Auri, Auriculatae; Sexf, Sexflorae; Orga, Organenses; Mise,
dodysosmia; Psegr, Pseudogranadilla; Hah, Hahniopathanthus; Lept,
, Quadrangulares; Digi, Digitatae; Tili, Tiliafoliae; Laur, Laurifoliae;
ae; Doli, Dolichostemma; Euas, Euastrophea; Pseas, Pseudoastrophea.
c
R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 389

Fig. 3. Character evolution of 19 morphological characters and chromosome numbers for different subgenera. The different states are represented by
black tick marks, densely dotted tick marks, dispersed dotted tick marks, and double tick marks. Double pattern tick mark indicates a polymorphic
character state (two different states). The character states indicated on the basal branch are the outgroup states. (a) Vegetative characters: 1. tendrils;
2. tendrils terminating the peduncle; 3. leaves; 4. polymorphic leaves (two different leaf shapes in the same individual); 5. hooked trichomes or
uncinate trichomes in the leaf blade; 6. petiolar glands; 7. bracteal glands with three different states absent, present at margin or present at base; 8.
bracts; (b) anatomical characters and chromosome numbers: 9. tree species: yes or no; 10. dioecious plants; 11. seed ridges; 12. pollen type: type 1
(reticulum with less than 5 lm wide and exine with 2 12–4lm wide) or type 2 (reticulum with more than 7lm wide and exine with more than 6lm
wide) (Presting, 1965); 13. chromosome number (n); and (c). floral characters: 14. inflorescences; 13. colour flower: light (white, green, pinkish,
yellowish) or hot (red, rose, violet, yellow carmine); 14. calyx tube: not tubular (bowl-shaped, funnel-shaped, patelliform), short tubular (less than
2 cm long) or long tubular (more than 2 cm long); 15. petals; 16. corona filaments: linear/filiform, verrucose, forming a tubular membrane or tu-
bercles-shaped; 17. operculum, and 18. ovary shape.
390 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
All the phylogenetic analyses conducted resulted in
congruent trees that have well resolved and well sup-
ported topologies. The strongest support values wereobtained with the Bayesian analysis, which has recently
become popular in phylogenetics, and accessible
through the MrBayes program (Huelsenbeck and Ron-
quist, 2001). Although Suzuki et al. (2002) pointed out
that this method creates an overestimation of the
probabilities of the internal nodes with respect to other
methods, the strong support values obtained with our
Bayesian analysis were corroborated by the bootstrap-ping assessment and the decay indexes conducted in the
MP analysis (Fig. 2).
The general topology presents three main well sup-
ported monophyletic groups (Fig. 2). The first clade,
which consists of the species representing subgeneraAstrophea and Tryphostemmatoides, is sister group to
two derived clades. These two clades regroup 15 of the
17 subgenera represented in this study.
The most basal group appears to be the monotypic
subgenus Polyanthea, with its one species P. cirrhiflora
from French Guiana. On the basis of morphological
characteristics such as the presence of compound leaves
and glands at the margin of bracts (Fig. 3a), Killipcreated a separate subgenus for this species in spite
noting its apparent relationship to the subgenus

Fig. 3. (continued)
R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 391
Astrophea because of the shared presence of verrucose
outer corona filaments and a truncate and 3-angled
ovary (Fig. 3c). Our results show that P. cirrhiflora and
Astrophea are sister groups to the rest of the genus, and
paraphyletic, indicating that the characters shared by
these two taxa could be considered to be plesiomorphic.Their paraphyly is strongly supported in all trees: 14
changes separate the two groups. We were unable to
include SE Asian Passiflora species, but it would have
been interesting to examine the relationship between
species from this geographical region and P. cirrhiflora,
since both groups have been considered as subsections
of the section Decaloba by Harms (1925). So far, our
results support the classification of Killip (1938) inwhich P. cirrhiflora forms an independent group.
4.1. Clade 1
The subgenus Astrophea forms a very well supported
clade with bootstrap support of 100. This result is
in agreement with the new classification of Passiflora
proposed by Feuillet and MacDougal (1999), in whichAstrophea is considered as one of the four subgenera.
Morphological features such as their arboreal habit and
scar-like petiolar glands (Killip, 1938) (Figs. 3a and c)
constitute therefore synapomorphies of this group. The
basal position of Astrophea suggests that the arborescent
habit could be ancestral in the genus, as assumed Ben-
son et al. (1975). Its pollen type shared with Plec-
tostemma, Polyanthea, and the outgroups (Presting,
1965) (Fig. 3b) could then considered as a pleisiomor-phy. The position of P. candida (Poep & Endl.) Mast.,
separated from the Astrophea clade in our trees, is un-
explained. This position is possibly the result of a wrong
identification of this species.
Passiflora tryphostemmatoides Harms appears closely
related to the Astrophea clade. This species has been
classified in an independent subgenus called Trypho-
stemmatoides on the basis of the presence of a non-pli-cate operculum, of a tendril terminating the peduncle
and finally the presence of specialized branching-tendril
(Killip, 1938; Vanderplanck, 2000) (Fig. 3a and c).
Several species from the subgenus Plectostemma were
described as presenting also a non-plicate operculum
(Holm-Nielsen and Jørgensen, 1986; Killip, 1938),
leading the authors to argue that all members of the
subgenus Tryphostemmatoides should be included inPlectostemma. However, the relationship between P.
tryphostemmatoides and Astrophea subgenus is well
supported in our phylogeny (Bootstrap Support¼ 88).

Fig. 3. (continued)
392 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
Therefore, we consider subgenus Tryphostemmatoides as
the sister group of subgenus Astrophea.
4.2. Clade 2
Clade 2 consists of a well supported group (BS¼ 87)
of 36 species belonging to six different subgenera
identified by Killip (1938). Two small subgenera are
found at the base of this clade: Deidamioides, (repre-
sented by P. lancetellensis MacDougal & Meerman),
and the monospecific Tetrapathea from New Zealand
(P. tetrandra Banks ex DC). Most of this clade consists
of species representing the subgenus Plectostemma ofFeuillet and MacDougal (1999), which includes the
subgenera Apodogyne, Astephia, Murucuja, Plec-
tostemma, and Pseudomurucuja recognized by Killip
(1938). The status of Deidamioides as an independent
subgenus, suggested by Harms (1923) and accepted
by Killip (1938) and Feuillet and MacDougal (1999), is
supported by our results, as shown by the presence of
21 changes between P. lancetellensis and the Plec-
tostemma group. The monophyly of this subgenus is
supported by a rare combination of features like the
plicate operculum, trifoliate leaves and peduncles ter-
minating in a 2-flowered tendril (Figs. 3a and c). The
position of P. tetrandra, intermediate between P.
lancetellensis and the Plectostemma group, indicatesthat Tetrapathea is not an independent monotypic ge-
nus. This result suggests that the particular reproduc-
tive features of this species such as dioecy (Fig. 3b),
never observed elsewhere in the genus Passiflora, are
derived character states and do not reflect an inde-
pendent origin from Passiflora.
Likewise, this clade includes five subgenera initially
considered by Killip (1938) as independent. Our resultsshow that these subgenera are embedded within Plec-
tostemma, making this group non-monophyletic. We
suggest therefore that only one subgenus should be
maintained: Plectostemma. In fact, P. multiflora, P.
penduliflora, P. perfoliata, and P. tacsonoides were re-
garded in the past as belonging to Plectostemma until
they were placed in three different subgenera for pos-
sessing many segregating characters (Killip, 1938). Forexample, P. multiflora was excluded from Plectostemma

R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 393
and moved to a new monotypic subgenus Apodogyne onthe basis of distinctive characters such as glandular
petioles and inflorescences in fascicles (Figs. 3a and c).
Our results shed a new light on the interpretation of
morphological features. For example, P. multiflora and
Plectostemma species closely related in our trees share a
plicate operculum and also transversely sulcate seeds
(Figs. 3b and c). These arguments support the inclusion
of the subgenus Apodogyne into Plectostemma. Simi-larly, characters like the absence of glands in petioles
and transversely sulcate seeds argue for the inclusion of
P. perfoliata L., P. tacsonoides Killip (Pseudomurucuja),
P. tulae Urb., and P. murucuja L. (Murucuja) in Plec-
tostemma (Figs. 3a and b). These latter species were
classified in a separate subgenus on the basis on their
coloured flowers and the corona forming a tubular
membrane forMurucuja (Fig. 3c). Passiflora pendulifloraBertero ex DC. was also considered as a separate sub-
genus by Killip (1938) because of its lack of an oper-
culum (Fig. 3c). The position of P. murucuja, P. tulae, P.
penduliflora, P. tacsonoides, and P. perfoliata in a same
sub-clade is not unexpected since all these species are
mainly distributed in the West Indies.
Furthermore, the trees confirmed the position of the
three Australian species, P. herbertiana Ker Gawl., P.aurantia G. Forst., and P. cinnabarina Lindl. described
before as belonging to Plectostemma. These were
grouped in a subsection called Eudecaloba within section
Decaloba of Plectostemma (deWilde, 1972; Harms, 1925)
(Table 1). To confirm the hypothesis of deWilde (1972)
that old world indigenous species belong to Plec-
tostemma, it would be necessary, in a further study of
the genus Passiflora, to include the Asian and NewGuianan species regrouped in the three sections Deca-
loba subsect. Polyanthea, Holrungiella, and Octandran-
thus (deWilde, 1974).
On the basis of these results, we suggest retention of
the three following subgenera in clade 2: Deidamioides,
Tetrapathea, and Plectostemma. We also suggest a
subdivision of subgenus Plectostemma into three differ-
ent sections. One section would be composed of all theseries of Decaloba, sections Xerogona, Cieca and
Pseudogranadilla and also the species P. murucuja, P.
tulae, P. penduliflora, P. tacsonoides, and P. perfoliata.
The newly described species from Panama, P. sandrae
(ined.) should be included in this section, according to
our results. The second section would be composed of
the section Pseudodysosmia and also include P. alnifolia
Kunth., initially classified in a different series, namelyPuntactae but appearing derived from Pseudodysosmia
in our trees. This last result is in agreement with Mac-
Dougal (1994) who described this section as monophy-
letic because of the presence uncinate trichomes in the
shoot apex: consequently he named the section the
‘‘hooked trichomes group’’ (Fig. 3a). Finally, the species
from the series Hahniopathanthus with their reticulate
sulcate seeds (Fig. 3b) would form the last section,which is sister to the rest of Plectostemma species.
4.3. Clade 3
This strongly supported clade (BS¼ 100) includes all
the species representing the subgenera Calopathanthus,
Distephana, Dysosmia, Granadilla, Manicata, Psilanthus,
Tacsonia, and Tacsonoides. The subgenus Dysosmia isthe sister clade to the rest of the species. The monophyly
of the species P. foetida L. is well supported (BS¼ 100).
The presence of unique characters such as pinnatisect
bracts, petiolar glands modified in hairs, and absence of
tendrils (Fig. 3a) supports the maintenance of this spe-
cies in a separate subgenus.
The rest of the species are distributed in three
well-supported groups of unresolved relationship. Thepresence of P. sanguinolenta Mast. & Linden from the
subgenus Psilanthus is unexpected in clade 3. In fact,
several morphological characters such as the pollen type
and the absence of bracts (Figs. 3a and b) suggest an
affinity of P. sanguinolenta with the species of clade 2
(Killip, 1938; Snow and MacDougal, 1993). However,
the presence of a red tubular calyx and a non-plicate
operculum of P. sanguinolenta distinguish it from clade2 (Fig. 3c). Further analyses including more species
of the subgenus Psilanthus are necessary to clarify its
relationship with the species of the Granadilla group.
Since the species representing subgenera Calopa-
thanthus, Distephana, Manicata, Psilanthus, Tacsonia,
and Tacsonoides are nested within Granadilla in the
trees, we suggest that this group should be united in one
subgenus. We also propose that Granadilla should besubdivided into three different sections. The first section
(BS¼ 92) would correspond to the subgenera Calopa-
thanthus, Manicata, and Tacsonia. Both latter subgenera
are mainly distributed in the Andes Mountains and
share a long calyx tube (Fig. 3c). In contrast, P. race-
mosa Brot. (Calopathanthus) is mainly distributed in
Brazil and has an operculum that forms a cylindrical
tube. However, its relationship with Manicata andTacsonia is strongly supported, suggesting that these
characters are derived. The second section would cor-
respond to the series Quadrangulares, Incarnatae, and
Laurifoliae (subgenus Granadilla) and the subgenus Di-
stephana. This latter subgenus consists of species with
scarlet flowers and corona filaments forming a tubular
membrane suggesting a monophyletic origin for this
group of species (Fig. 3c). The position of P. cincinnataMast. (subgenus Granadilla) within Distephana is unex-
pected, since this species does not display these specific
morphological features. As this subclade consists of a
large polytomy, an explanation could be that P. cincin-
nata is actually the sister group to Distephana. The po-
sition of the recently described French Guiana species
P. aimae confirms its relationship with Distephana, as

394 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
suggested by Annonay and Feuillet (1998) P. gabriell-
ana, also a French Guiana species recently described as
being part of Granadilla subgenus (Vanderplanck, 2000),
appears to be related to P. laurifolia inside this group
confirming its classification.
The last section that we propose can be split into two
subsections. The first subsection is formed by the series
Tiliafoliae and Digitatae (subgenus Granadilla). The
presence of united bracts at the base enveloping the budswould represent a synapomorphy of this sub-clade. The
other subsection contains the series Lobatae, Menispe-
rmifoliae, and Simplicifoliae (subgenus Granadilla),
which share foliaceous and verticillate bracts. The sub-
genus Tacsonoides, represented by P. umbilicata and P.
reflexiflora, is paraphyletic, in a basal position within
this subsection. The classification of P. reflexiflora and
P. umbilicata has varied over time. Their long calyx tubefavours their inclusion in Tacsonia (Harms, 1925)
whereas the 2–4 seriate corona supports their inclusion
in the disappeared section Tacsonoides (subgenus Gra-
nadilla) (De Candolle, 1828). Our results validate this
latter suggestion. Since Tacsonoides appears paraphy-
letic in the trees, the similarity between the two species
could be due to the retention of plesiomorphies inherited
from a common ancestor or a convergent adaptation tohummingbird pollination (MacDougal, 1994).
To summarize, we suggest that two subgenera should
be recognized in clade 3: Dysosmia and Granadilla, and
we propose to subdivide Granadilla into three sections.
4.4. Chromosome number
The division of Passiflora into three main cladesbased on DNA sequence data from the glutamine syn-
thetase nuclear gene expressed in the chloroplast, and
our subsequent suggestion to group 10 subgenera in
three subgenera, complies with variation in chromosome
numbers. Each of our three clades is characterized by a
different base chromosome number: x ¼ 12 chromo-
somes for clade 1 (Astrophea s.l.), x ¼ 6 for clade 2
(Plectostemma s.l.), and x ¼ 9 for clade 3 (Granadillas.l.) (De Melo et al., 2001; Snow and MacDougal, 1993)
(Fig. 3b).
The only incongruence between the classification and
the chromosome number concerns P. sanguinolenta. The
single chromosome count available for this species gave
n ¼ 6 chromosomes, leading Snow and MacDougal
(1993) to place it with Plectostemma, whereas our results
support its inclusion within Granadilla, in agreementwith Killip (1938). The similarity of chromosome num-
ber between P. sanguinolenta and Plectostemma could
result from convergence.
Furthermore, the unique chromosome count avail-
able for the Deidamioides subgenus (Gilbert and Mac-
Dougal, 2000) gave n ¼ 9 chromosomes (Fig. 3b),
relating it to clade 3. The difference in the chromosome
number of Deidamioides and Plectostemma (s.l) wouldsupport the independent origin and the monophyly of
Deidamioides.
Dysosmia, considered here as a separate subgenus
within clade 3, has variable chromosome numbers from
n ¼ 9 to n ¼ 11 (Fig. 3b). The high morphological var-
iability displayed by the species belonging to Dysosmia
and reflected by their subdivision in several subspecies
and varieties has been suggested to be the product of aconstant hybridization process (Killip, 1938; Vanderp-
lanck, 2000), a likely explanation for the chromosomal
variability of this group. Despite this variability, the
chromosome numbers observed support the sister group
relationship between Granadilla and Dysosmia.
Raven (1975) affirmed that the base number for
Passiflora is x ¼ 9. The position of Astrophea in the
phylogeny suggests in contrast that n ¼ 12 is the an-cestral chromosome number of Passiflora. The presence
of n ¼ 12 chromosomes in the outgroup Adenia (Passi-
floraceae) supports this possibility. To confirm this hy-
pothesis, chromosome counts for P. cirrhiflora, sister
group to all other Passiflora species, and for P. trypho-
stemmatoides (basal in clade 1) are necessary.
5. Conclusion
The study presented here is the first comprehensive
molecular phylogenetic analysis of the large genus Pas-
siflora. It allowed the identification of eight clades within
Passiflora, and we suggest that each of them should be
considered as a separate subgenus, namely Astrophea,
Deidamioides, Dysosmia, Granadilla, Plectostemma,Polyanthea, Tetrapathea, and Tryphostemmatoides. Our
phylogeny is in overall agreement with the new infra-
generic classification proposed by Feuillet and Mac-
Dougal (1999), in which four subgenera are recognized
(Plectostemma, Astrophea, Deidamioides, and Grana-
dilla), but our results give evidence for the existence of
three additional subgenera. In spite of the good resolu-
tion provided by the ncpGS gene, further studies usingadditional molecular markers, such as chloroplast re-
gions, must be undertaken in order to confirm our re-
sults. It would also be necessary to increase the number
of species and in particular to include the Asian mem-
bers of Passiflora as well as species of subgenera not
represented in this analysis.
Acknowledgments
We are extremely grateful to Christian Houel (Pas-
siflora National Collection, Blois, France) who provided
a great part of the plant material. We deeply thank his
continuous help and availability. We are also indebted
to Catalina Estrada (University of Texas, Austin, USA),

R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396 395
Cor Laurens (Passiflora National Collection, Holland),Maurizio Vecchia (Passiflora National Collection, It-
aly), Clinton Morse (University of Connecticut, USA),
Chris Jiggins (Smithsonian Tropical Institute, Panama),
Margarita Beltr�an (Smithsonian Tropical Institute,
Panama), Alejandro Merch�an (Universidad de los An-
des, Bogota, Colombia), H�ector Eduardo Esquivel
(Botanical Garden Alejandro Von Humboldt, Tolima,
Colombia), Alexandra Hiller (Giessen University, Ger-many), Lawrence Gilbert (University of Texas, Austin,
USA), and the Australian National Botanical Gardens
for generously providing leaf and seed material for
DNA extractions. We thank C�eline Devaux, Bernard
Lejeune, Jacqui Shykoff, and two anonymous reviewers
for their helpful comments on the manuscript.
References
Annonay, H., Feuillet, C., 1998. Passiflora aimae (Passifloraceae), une
esp�ece nouvelle de Guyane Franc�aise. Adansonia 20 (3), 295–298.
Benson, W.W., 1978. Resource partitioning in passion vine butterflies.
Evolution 32, 493–518.
Benson, W.W., Brown, K.S., Gilbert, L.E., 1975. Coevolution of
plants and herbivores: passion flower butterflies. Evolution 29,
659–680.
Cervi, A.C., 1997. Passifloraceae do Brasil. Estudo do genero
Passiflora L., subgenero Passiflora, Fontqueria, Cyanus, S.L.,
Madrid.
Chase, M.W., Zmarzty, S., Ll�edo, M.D., Wurdack, K.J., Swensen,
S.M., Fay, M.F., 2002. When in doubt, put in Flacourtiaceae: a
molecular phylogenetic analysis based on plastid rcbL DNA
sequences. Kew Bulletin 37, 141–181.
Cusset, G., 1967. Les passiflorac�ees asiatiques. Adansonia 7 (2), 371–
385.
De Candolle, A.P., 1828. Passifloreae. Prodr. Syst. Nat. 3, 321–338.
De Melo, N.F., Cervi, A.C., Guerra, M., 2001. Karyology and
cytotaxonomy of the genus Passiflora. Plant Syst. Evol. 226, 69–84.
deWilde, W.J., 1972. The indigenous old world passifloras. Blumea 20,
227–250.
deWilde, W.J., 1974. The genera of tribe Passifloreae (Passifloraceae),
with species reference to flower morphology. Blumea 22, 37–50.
Emshwiller, E., Doyle, J.J., 1999. Chloroplast-expressed glutamine
synthetase (ncpGS): potential utility for phylogenetic studies with
an example from Oxalis (Oxalidaceae). Mol. Phylogenet. Evol. 12,
310–319.
Emshwiller, E., Doyle, J.J., 2002. Origins of domestication and
polyploidy in oca (Oxalis tuberosa: Oxalidaceae). Chloroplast-
expressed glutamine synthetase data. Am. J. Bot. 89, 1042–1056.
Eriksson, T., 1998. AutoDecay 4.0, Program distributed by the author.
Department of Botany, Stockholm University, Stockholm.
Escobar, L.K., 1988. Passifloraceae. Passiflora, subg�eneros: Tacsonia,
Rathea, Manicata y Distephana. Flora de Colombia 10, 1–138.
Escobar, L.K., 1989. A new subgenus and five new species in Passiflora
(Passifloraceae) from South America. Ann. Missouri Bot. Gard. 76,
877–885.
Escobar, L.K., 1994. Two new species and a key to Passiflora subg.
Astrophea. Syst. Bot. 19, 203–210.
Fajardo, D., Angel, F., Grum, M., Tohme, J., Lobo, M., Roca, W.,
S�anchez, I., 1998. Genetic variation analysis of the genus Passiflora
L. using RAPD markers. Euphytica 101, 341–347.
Feuillet, C., MacDougal, J., 1999. XVI International Botanical
Congress, Raven, P., Saint Louis, Missouri, USA, Abstract 4295.
Gilbert, L.E., 1980. Butterflies and plants. In: Gilbert, L., Raven, P.
(Eds.), Coevolution of Animals and Plants. University of Texas,
Austin, pp. 210–240.
Gilbert, L.E., 1982. The coevolution of a butterfly and a vine.
Heliconius butterflies, Passiflora. Sci. Am. 247, 110–116, 119–121
ill.
Gilbert, L.E., 1983. Coevolution and mimicry. Coevolution, 263–
281.
Gilbert, L.E., MacDougal, J.M., 2000. Passiflora microstipula, a new
species of Passifloraceae from Southeast Mexico. Lundellia 3, 1–5.
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence align-
ment editor and analysis program for Windows 95/98/NT. Nucleic
Acids Symp. Ser. 41, 95–98.
Harms, 1923. Beitrage zur Kenntnis der amerikanischen Passiflorac-
een—I. Feddes Repertorium Specierum Novarum Regni Vegetabi-
lis 19, 58.
Harms, 1925. Passiflora. In: Engler, A., Prantl, K. (Ed.), Die
nat€urlichen pflanzenfamilien, vol. 21. Duncker and Humblot, pp.
470–507.
Holm-Nielsen, L., Jørgensen, P.M., 1986. Passiflora tryphostemmato-
ides and its allies. Phytologia 60, 119–124.
Huelsenbeck, J.P., Ronquist, F., 2001. MrBayes: Bayesian inference of
phylogenetic trees. Bioinformatics Appl. Notes 17, 754–755.
Judd, W.S., Campbell, C.S., Kellogg, E.A., Stevens, P.E., 1999. Plant
Systematics—A Phylogenetic Approach. Sinauer, Sunderland, MA.
Killip, E.W., 1938. The American Species of Passifloraceae. Field
Museum of Natural History, Chicago.
Kumada, Y., Benson, D.R., Hilleman, D., Hosted, T.J., Rochefort,
D.A., Thompson, C.J., Wohlleben, W., Tateno, Y., 1993. Evolu-
tion of the glutamine synthetase gene, one of the oldest existing and
functioning genes. Proc. Natl. Acad. Sci. USA 90, 3009–3013.
MacDougal, J.M., 1994. Revision of Passiflora subgenus Decaloba
section Pseudodysosmia (Passifloraceae). Syst. Bot. Monogr. 41, 1–
146.
Maddison, W.P., Maddison, D.R., 1992. MacClade: Analysis of
Phylogeny and Character Evolution 3.08. Sinauer Associates,
Sunderland, MA.
Pesole, G., Bozsetti, M.P., Lanave, C., Preparata, G., Saccone, C.,
1991. Glutamine synthetase gene evolution: a good molecular
clock. Proc. Natl. Acad. Sci. USA 88, 522–526.
Posada, D., 2001. ModelTest 3.06, Department of Zoology Brigham
Young University, Provo.
Presting, D., 1965. Zur morphologie der pollenk€orner der Passiflor-
aceae. Pollen et Spores 7, 193–247.
Raven, P.H., 1975. The bases of the angiosperm phylogeny: cytology.
Ann. Mo. Bot. Gard. 62, 724–764.
Rosenthal, A., Coutelle, O., Craxton, M., 1993. Large-scale produc-
tion of DNA sequencing templates by microtitre format PCR.
Nucleic Acids Res. 21, 173–174.
S�anchez, I., Angel, F., Grum, M., Duque, M.C., Lobo, M., Tohme, J.,
Roca, W., 1999. Variability of chloroplast DNA in the genus
Passiflora L. Euphytica 106, 15–26.
Simmons, M.P., Ochoterena, H., 2000. Gaps as characters in sequence-
based phylogenetic analyses. Syst. Biol. 49, 369–381.
Snow, N., MacDougal, J.M., 1993. New chromosome reports in
Passiflora (Passifloraceae). Syst. Bot. 18, 261–273.
Soltis, D.E., Mort, M.E., Soltis, P.S., Hibsch-Jetter, C., Zimmer, E.A.,
Morgan, D., 1999. Phylogenetic relationships of the enigmatic
angiosperm family Podostemaceae inferred from 18S rDNA and
rbcL sequence data. Mol. Phylogenet. Evol. 11, 261–272.
Spencer, K.C., 1986. Chemical mediation of coevolution in the
Passiflora–Heliconius interaction. In: Spencer, K.C. (Ed.), Chem-
ical Mediation of Coevolution. Academic Press, Chicago, pp. 167–
240.
Strimmer, K., vonHaeseler, A., 1996. Quartet puzzling: a quartet
Maximum likelihood method for reconstructing tree topologies.
Mol. Biol. Evol. 13, 964–969.

396 R. Yockteng, S. Nadot / Molecular Phylogenetics and Evolution 31 (2004) 379–396
Suzuki, Y., Glazko, G.V., Nei, M., 2002. Overcredibility of molecular
phylogenies obtained by Bayesian phylogenetics. Proc. Natl. Acad.
Sci. USA 99, 16138–16143.
Swofford, D.L., 2001. PAUP*. Phylogenetic Analysis Using Parsi-
mony and other methods 4.0b10. Sinauer Associates, Sunderland,
MA.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins,
D.G., 1997. The Clustal_X windows interface: flexible strategies for
multiple sequence alignment aided by quality analysis tools.
Nucleic Acids Res. 25, 4876–4882.
Vanderplanck, J., 2000. Passion Flowers. MIT Press, Cambridge,
MA.
Yockteng, R., Nadot, S., 2003. Infrageneric phylogenies: a comparison
of chloroplast-expressed glutamine synthetase, cytosol-expressed
glutamine synthetase and cpDNA maturase K in Passiflora, Mol.
Phylogenet. Evol., in press.