phylogenetic relationships between pinworms (nematoda: enterobiinae) parasitising the critically...
TRANSCRIPT

ORIGINAL PAPER
Phylogenetic relationships between pinworms(Nematoda: Enterobiinae) parasitising the critically endangeredorang-utan, according to the characterisation of moleculargenomic and mitochondrial markers
Ivona Foitová & Kristína Civáňová & Vlastimil Baruš &
Wisnu Nurcahyo
Received: 9 October 2013 /Accepted: 9 April 2014 /Published online: 1 June 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Pinworms (Nematoda: Enterobiinae) include 52species parasitising primates throughout the world. In thepresent study, we performed the first ever molecular analysisto investigate the phylogenetic position of recently describedpinworms parasitising the Sumatran orang-utan. The phylo-genetic analysis of mitochondrial CO1 and chromosomal 18SrDNA and ITS1 regions could support the independent statusof several Nematoda species. Our molecular data clearly sug-gest that Enterobius (Colobenterobius) buckleyi andLemuricola (Protenterobius) pongoi together with Pongobiushugoti form separate clades among other studied species,which significantly supports the hypothesis of recently de-scribed new species parasitising the orang-utan (Pongo abeliiand Pongo pygmaeus). The phylogenetic tree based on cyto-chrome oxidase 1 (CO1) gene variability showed possibleclose relationships between L. (Protenterobius) pongoi andP. hugoti; thus, we can assume that these species could haveinitially diverged in sympatry from a common ancestor.
Keywords Molecular phylogeny . Cytochromoxidase 1 .
18S rDNA . ITS1 . Orang-utan pinworms .Pongo abelii
Introduction
The study of parasite infection in wild primate populations is afirst and important step toward gaining a better understandingof parasite infection dynamics as they relate to primateconservation.
Pinworms (Nematoda: Oxyuridae) are a parasite groupoccurring in most families and genera of the order Primatesincluding human. Enterobiasis caused by Enterobiusvermicularis is the most frequently worldwide gastrointestinaldiseases reported among preschool children and youngerschoolchildren (Song et al. 2003; Rim et al. 2003; Remm2006; Gonzáles-Moreno et al. 2011). The subfamilyEnterobiinae Hugot, Gardner et Morand, 1996 includes 47species parasitising primates, and 3 species parasitising ro-dents (survey in Quentin and Tenora 1975; Hugot 1984a, b,1999; Hugot et al. 1995, 1996; Sorci et al. 1997; Hugot andBaylac 2007). Recently, Hasegawa et al. (2003, 2005), Barušet al. (2007), Foitová at al. (2008, 2010), and Kuze et al.(2010) described five new primate pinworm species. Accord-ing to the current system of classification, pinworms(Enterobiinae) parasitising primates are classified under fourgenera (bold) [subgenera (underlined)] (Hugot 1999 Hugotet al. 1996 Hugot and Baylac 2007 Hasegawa et al. 2004Baruš et al. 2007): (1) Enterobius Leach, 1853 [Enterobius(Leach, 1853) Colobenterobius Quentin, Betterton etKrishasamy, 1979]; (2) Lemuricola Chabaud et Petter, 1959(syn. BiguetiusChabaud, Brygoo et Petter, 1965) [LemuricolaChabaud et Petter, 1959 Protenterobius Inglis 1961Madoxyuris Chabaud, Bryoo et Petter, 1959] (3) PongobiusBaruš, Foitová, Koubková, Hodová, Šimková et Nurcahyo,2007 and (4) Trypanoxyuris Vevers, 1923 (syn.Lobatorobius Skrjabin et Schikhobalova, 1959)[Trypanoxyuris (Vevers, 1923) Hapaloxyuris Inglis et
I. Foitová (*) :K. CiváňováDepartment of Botany and Zoology, Masaryk University, Kotlářská2, 611 37 Brno, Czech Republice-mail: [email protected]
V. BarušInstitute of Vertebrate Biology, Czech Academy of Sciences, Květná8, 603 65 Brno, Czech Republic
W. NurcahyoDepartment of Parasitology, Faculty of Veterinary Medicine, GadjahMada University, Yogyakarta, Indonesia
Parasitol Res (2014) 113:2455–2466DOI 10.1007/s00436-014-3892-y

Cosgrove, 1965 Oxyuronema Kreis, 1932 (syn.Paraoxyuronema Artigas, 1936 pro parte, sensu Hugot et al.1996) Buckleyenterobius Sandoshan, 1950 (syn.Odontorobius Skrjabin et Schikhobalova, 1951)Paraoxyuronema Artigas, 1936 pro parte, sensu Hugot et al.1996)]. The monotypic genus Ingloxyuris Chabaud, Petter etGolvan, 1961 containing the species Ingloxyuris inglisi doesnot share the characteristic of the subfamily Enterobiinae, andaccording to Hugot et al. (1996), it should be referred toOxyuridae sensu lato, until its precise classification.
It is believed that pinworms and their host primates havecoevolved (Cameron 1929; Sandosham 1950; Chabaud et al.1961; Inglis 1961; Petter et al. 1972; Brooks 1982; Glen andBrooks 1986; Hugot et al. 1996; Sorci et al. 1997; Hasegawa1999; Hasegawa et al. 2005). A cladistic analysis of 13 speciesof Enterobius based on 31 morphological characters (Brooks1982) supports the notion that pinworms and primates haveco-speciated. An exact detailed cladistic analysis of 46Enterobiinae species taxa based on 45 morphological charac-ters from various organ systems was also made by Hugot(1999). So far, only one study by Nakano et al. (2006) hasevaluated the sequences of the mitochondrial cytochrome coxidase subunit 1 gene and nuclear internal transcribed spacer2 region of ribosomal DNA (rDNA) in four pinworm speciesfrom the genera Enterobius (subgenus Enterobius) andTrypanoxyu r i s ( s ubgene r a Trypanoxyu r i s andBuckleyenterobius).
In the present study, we performed the first ever molecularanalysis to investigate the phylogenetic position of recentlydescribed pinworms parasitising the Sumatran orang-utan(Pongo abelii Lesson, 1827). The phylogenetic relationshipswere inferred according to the variability of partial sequencesof both mitochondrial and nuclear markers—the mitochondri-al cytochrome c oxidase 1 gene (CO1), the nuclear region ofthe small ribosomal subunit 18S rDNA (18S), and the internaltranscribed spacer 2 region of rDNA (ITS2).
Materials and methods
Parasite samples
The majority of the pinworms [Pongobius hugoti Baruš,Foitová, Koubková, Hodová, Šimková et Nurcahyo, 2007;Enterobius (Enterobius) buckleyi Sandosham 1950; andLemuricola (Protenterobius) pongoi Foitová, Baruš, Hodová,Koubková et Nurcahyo, 2008] were collected from freshfaeces from the Sumatran orang-utan (P. abelii). The apeslived in a semi-wild population in the Bukit Lawang locality(the former home to a rehabilitation centre for orang-utans),situated on the south-western border of the Gunung LeuserNational Park, province of North Sumatra, Indonesia. Sam-ples of E. (Enterobius) vermicularis (Linnaeus, 1758) were
collected from the faeces of an 11-year-old boy (Homosapiens L.) from Brno, the Czech Republic. These sampleswere provided by The Children’s Medical Center, The Uni-versity Hospital Brno, Czech Republic (hospital laboratorymaterial). All worms were identified by morphological anal-ysis and immediately fixed in 96 % ethanol.
Isolation of DNA
Genomic DNA was extracted separately from each adultworm (females only). The tissue samples fixed in 96 % etha-nol were used for DNA isolation using QIAamp DNA MiniKit (Qiagen) following protocol recommendations. The pro-cess was modified by means of overnight incubation at roomtemperature during lysis. The quality and yield of isolateswere verified by agarose gel electrophoresis on 1.5 % gelstained with GoldView dye (SBS Genetech Co. Ltd.). TheDNA concentration was determined using the Qubit fluorom-eter and Quant-iT dsDNA HS Assay Kit according to themanufacturer’s protocol (Invitrogen).
PCR amplification and automatic sequencing
All primers used in this study are presented in Table 1.To amplify the required DNA fragment, PCR was per-
formed in 30 μl of reaction volume using 50 ng of genomicDNA. Reaction mixtures for both CO1 and ITS2 were com-prised of 1× HF buffer, 0.2 mM of each dNTP, 0.3 μMof eachappropriate specific primer, and 1 U of Phusion high-fidelityDNA polymerase (Finnzymes). Amplification was run in35 cycles under the following conditions: initial denaturationat 98 °C/30 s; cycling at 98 °C/10 s, 55 °C/30 s, and 72 °C/30 s; and a final extension at 72 °C/10 min. Amplification ofthe 18S fragment was performed in a reaction mixtureconsisting of 1× standard PCR buffer without MgCl2, 2 mMMgCl2, 0.2 mM of each dNTP, 0.5 μM of each primer, and0.5 U of Taq DNA polymerase (Fermentas) in 39 cycles underthe following conditions: initial denaturation at 94 °C/5 min;cycling at 94 °C/30 s, 52 °C/30 s, and 72 °C/l min; and a finalextension at 72 °C/10 min. All PCR reactions were performedin a Mastercycler ep gradient thermocycler (Eppendorf). Thequality of PCR products was verified by gel electrophoresison 1.5 % agarose gel stained with GoldWiew dye.
For automatic sequencing, the obtained PCR products werepurified using the High Pure PCR Product Purification Kit(Roche), and amplification with the fluorescent chemistry ofBigDye Terminator v3.1 Ready reaction Cycle SequencingKit (Applied Biosystems by Life Technologies, Carlsbad, CA,USA) was performed. Subsequently, the products of the se-quencing reactions were purified using BigDye X-TerminatorPurification Kit (according to the manufacturer’s protocol)and sequenced in an ABI 3130 Genetic Analyser (AppliedBiosystems) under the appropriate module.
2456 Parasitol Res (2014) 113:2455–2466

Phylogenetic analysis and comparative data
The raw sequencing data were analysed by Sequencing Anal-ysis software v.5.2 (Applied Biosystems) and subjected toprocessing by Sequencher v.4.7. (Gene Codes Corporation)and SeqScape v2.5. (Applied Biosystems) software to obtaincontigs. Alignments of amino acid sequences and auxiliaryalignments were performed in BioEdit Sequence AlignmentEditor v.7.0.9.0. (Hall 1999). Pairwise alignments and phylo-genetic and evolutionary analyses (ML—maximum likeli-hood; MP—maximal parsimony) were conducted usingMEGA version 5 (Tamura et al. 2011). Bayesian analysis(BA) was performed in MrBayes v3.1.1 (Huelsenbeck andRonquist 2001) with the applied recommended model com-puted by the MrModeltest v. 2.2 (Nylander 2004) programme.
The representatives of obtained sequences from analysedgenes of E. vermicularis, E. buckleyi, L. (P.) pongoi, andP. hugoti used in our work were deposited at theEMBL/GenBank/DDBJ database (accession numbers for18S region FR687850–FR687853; for ITS2 regionFR687961–FR687964; and for CO1 gene FR687965–FR687979; FR687973 undetermined pinworm defined asEnterobiinae sp.) . Analysed sequences from theAscaridaefamily used as outgroup are not yet published, asthey will be used in further studies. However, this informationis available from the authors.
For comparison with our new data on the CO1 gene, weused sequences from five primate parasite species [Oxyuridae,Enterobinae: E. vermicularis (AB221457–AB221474),E. anthropopitheci (AB254450), Trypanoxyuris atelis(AB222177), Trypanoxyuris microon (AB222176),Lemuricola bauchoti (AB222178)] and two relative rodentparasite species [Oxyuridae, Oxyurinae: Syphacia montana(AB282581–AB282585) and Syphacia frederici (AB282586–AB282588; AB282593)) from the DDBJ/EMBL/GenBankdatabase.
Concerning the hosts, the above-mentioned sequenceswere obtained by analysing samples of (1)E. (E.) vermicularisfrom humans (Japan) and chimpanzees Pan troglogytes(Blumenbach, 1775) reared in zoos/institutions in Japan; (2)E.(E.) anthropopitheci (Gedoelst, 1916) from chimpanzees;(3) Trypanoxyuris (Trypanoxyuris) microon (Linstow, 1907)
from a southern night monkey (Aotus azarae Humboldt,1811); (4) T. (Buckleyenterobius) atelis (Cameron 1929) froma black-handed spider monkey (Atelles geoffroyi Kuhl, 1820)reared in a zoological gardens (Japan); (5) Lemuricola(Madoxyuris) bauchoti (Chabaud, Brygoo et Petter, 1965)from Hapalemursimus Gray, 1870 (Madagascar); (6)Syphacia montana Yamaguti, 1943 isolated from rodent hostsClethrionomys rufocanus (Sundevall, 1845) (Muridae,Microtinae); and (7) S. fredericiRoman, 1945 fromApodemusspeciosus (Temminck, 1844) (Muridae, Murinae).
All obtained phylogenetic trees were outgroup-rootedusing the nucleotide sequences from our unpublished samplesmorphologically determined as: (1) Ascaris sp. obtained froma Sumatran orang-utan, (2) A. suum (Goeze, 1782) obtainedfrom a pig (Sus scrofa f. domestica), and (3) A. columnaris(Leidy, 1856) obtained from a polecat (Mephitis mephitisSchreber, 1776).
Results
CO1 gene
A total of 19 successfully amplified specimens of four differ-ent species (the obtained nucleotide fragments of 441 bp) wereincluded in the analysis of the CO1 gene marker and used foralignment.
Evolutionary history was inferred using statistical analysesin MEGA5 according to the following parameters: (1) phy-logeny reconstruction by maximum parsimony test of phylog-eny—bootstrap method (1,000 replicates); nucleotide substi-tution model type using all sites; MP search method—close-neighbor interchange on random trees; CI=0.578059, RI=0.846272, RCI=0.489195 (for all sites); iCI=0.559471 (forparsimony informative sites); iRI=0.846272 (for parsimonyinformative sites); iRCI=0.473465 (for parsimony informa-tive sites); (2) phylogeny reconstruction by maximum likeli-hood test of phylogeny—bootstrap method (1,000 replicates);nucleotide substitution model type, model GTR+G+I (gen-eral time reversible model, rates among sites—gamma distrib-uted with invariant sites); number of discrete gamma
Table 1 Oligonucleotides usedfor PCR amplification andsequencing
AT optimised annealingtemperatures
Primer name Primer sequences (orientation 5′–3′) AT References
CO1-F TGGTTTTTTGTGCATCCTGAGGTTTA 55 °C Nakano et al. 2006
CO1-R AGAAAGAACGTAATGAAAATGAGCAAC
ITS2-F CGAGTATCGATGAAGAACGCAGC 55 °C Nakano et al. 2006
ITS2-R ATATGCTTAAGTTCAGCGGG
18S-F_P0 CGCGAATRGCTCATTACAACAGC 52 °C Floyd et al. 2005
18S-R_P1 GGGCGGTATCTGATCGCC
Parasitol Res (2014) 113:2455–2466 2457
2458 Parasitol Res (2014) 113:2455–2466

categories—6, using all sites; tree inference options—MLheuristic method—nearest-neighbor interchange (NNI).
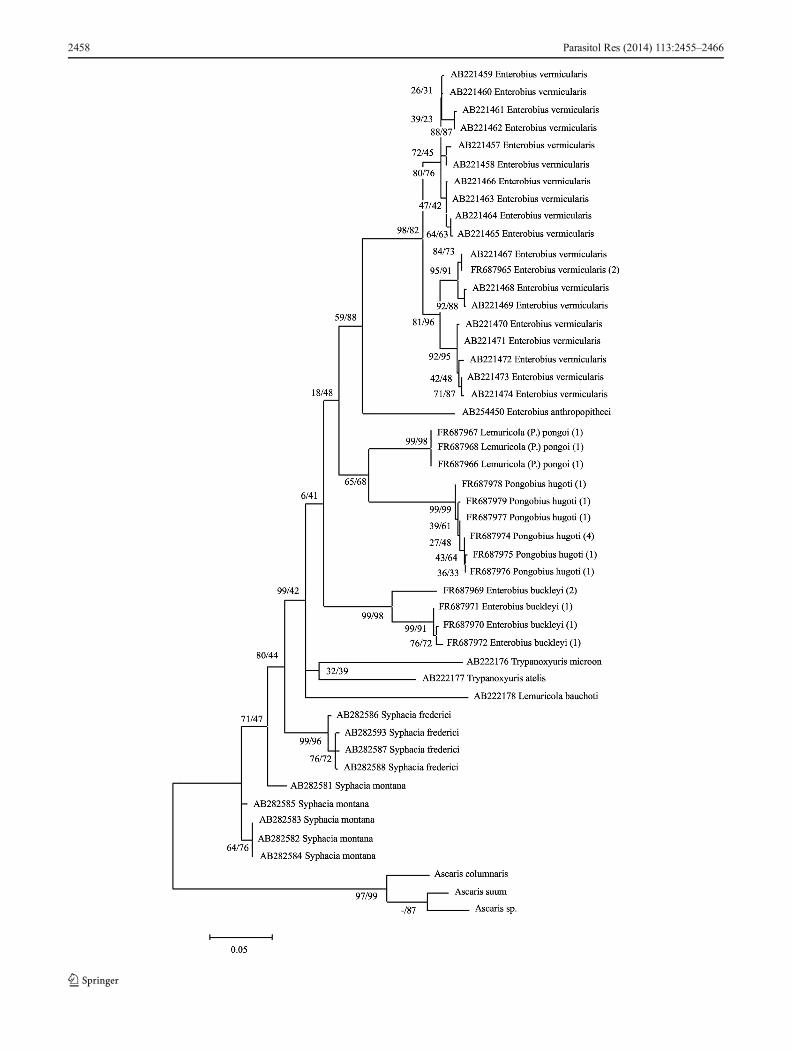
The resultant MP/ML tree containing 48 sequences isshown in Fig. 1.
To confirm the validity of the obtained nucleotide se-quences of E. vermicularis, the sequences were aligned andconverted to amino acid sequences using MEGA version 5software and BioEdit Sequence Alignment Editor v.7.0.9.0(Hall 1999). These were then aligned with three groups ofamino acid data from Nakano et al. (2006) [types and acces-sory database numbers: type A—AB221470; type B—AB221467; type C—AB221459]. We discovered that oursamples from humans appertain to Nakano’s CO1 aminoacid type B, which is characterised by threonine at position106 of the analysed amino acid chain. Previously, the authorsfound type B only in the ape parasite material from Aichi andHyogo, Honshu, Japan. Although Nakano et al. (2006) ob-served only type A in the human material, we confirmedamino acid type B in our human samples from the CzechRepublic (Fig. 2).
The numbers of base substitutions per site from aver-aging over all sequence pairs between groups and withineach group are shown in Tables 2 and 3. Analyses wereconducted using the Kimura two-parameter model(Kimura 1980) in MEGA5, and all results for CO1 werebased on the pairwise analysis of 48 sequences. Allcodon positions were included. Ambiguous positionswere removed for each sequence pair. There were a totalof 441 positions in the final dataset. The given estima-tion in Table 3 also confirmed the estimation of the netevolutionary divergence between groups of sequences bygiving almost the same values (data not shown).
ITS2 region
In the analyses of the ITS2 region, a total of 17 specimensdivided into four taxonomic groups were included. The phy-logenetic test conducted in MEGA5 confirmed the separatepositions of E. (E.) buckleyi, P. hugoti, L. (P.) pongoi, andE. (E.) vermicularis with high values of bootstraps (data notshown). In this region, a high degree of variability was noticedbetween groups. This region was excluded from the subse-quent combined BA.
Comparing our reproducible ITS2 data on E. (E.)vermicularis (551 bp long fragment) with the ITSmajor types(X, Y—subtypes Y-1, Y-2, Y-3, and Z; Nakano et al. (2006);database IDs: AB222054– AB222058), it was not possible tofind and confirm any of the previously determined types. Allof our samples isolated fromH. sapiens in the Czech Republichad 100 % identical ITS2 sequences and represented anidentical and unique type that had not been describedpreviously. The comparison of our ITS2 consensus se-quence with Nakano’s data from the database is shownin Fig. 3.
18S gene
The 18S gene is generally present in eukaryotes. As is wellknown, the 18S rDNA region is not suitable for phylogeneticanalysis as it amplifies a highly conserved region. However,our amplified fragment covered most of the 5′ half of the 18Sgene, where much sequence variability tends to be found,making it a useful region for the DNA barcoding approach(Floyd et al. 2005).
As it was not possible to obtain the complete sequence ofamplified PCR product (approximately 900 bp), we used onlythe 799–833-bp-long regions of reproducible sequences forfurther analysis (27 individuals). We decided to combine CO1data and the 18S region in combined phylogenetic analyses
�Fig. 1 Evolutionary relationship based onCO1 gene sequences and theiranalysis. MP/ML bootstrap values are shown near nodes. Numbers inparenthesis indicate the number of individuals representing the sequence
Fig. 2 Amino acid translation of CO1 DNA sequences of E. vermicularis compared with data of Nakano et al. 2006 (types A–C). Our samples arelabelled NM26 and NM25. The shown alignment was conducted using the BioEdit Sequence Alignment Editor v.7.0.9.0
Parasitol Res (2014) 113:2455–2466 2459

using Bayesian testing, comprising 13 individual sequenceswith complete data from both regions.
Before BA, the combined dataset (1,286 sites in total; theset for CO1: 1–441 bp; the set for 18S: 442–1,286 bp) wasanalysed in MrModeltest v. 2.2 (Nylander 2004), which sug-gested the best-fitting evolution model for the analysed DNAalignment. The parameters of BA were as follows: chosenmodel (GTR+I+G), gamma distribution shape parameterγ=0.4602, mcmc ngen=2,000,000, nchains=8, nruns=2,temp=0.05, swapfreq=1, nswap=2, mcmcdiagn=yes,diagnfreq=1,000, relburnin=yes, burninfrac=0.3, sumtdisplaygeq=0.5, burnin=300, and sump burnin=300. Theresulting tree is shown in Fig. 4.
Discussion
Hasegawa (1999) believed that the reconstruction of phylog-eny based only on DNA molecules or morphology may leadto an erroneous interpretation. Therefore, in the comparison,we discuss our results in the light of conclusions formulatedby Hugot (1999), Brooks (1982), and Nakano et al. (2006),which enable the combination of both aspects.
A cladistic analysis made by Hugot (1999) suggested themonophyly of the subfamily Enterobiinae, and also the mono-phyly of three main genera (or groups of genera and relatedsubgenera): Enterobius, Lemuricola and Trypanoxyuris.Combined phylogenetic analyses of our material exhibitedmonophyly only for E. (E.) vermicularis and E. (E.)anthropopitheci. According to the position of the speciesE. buckleyi in the resulting tree (Fig. 1), we can dispute themonophyletic origin of this genus. Considering the gamma-corrected pairwise genetic distances between the taxa includedin the phylogenetic analysis, differences of 13.9 % or more,and the position in the MP/ML tree, the results stronglydistinguish E. buckleyi as an independent subgenus of thegenus Enterobius or, indeed, also as an independent taxon.However, the divergences between E. buckleyi, L.(P.) pongoi,and P. hugoti (0.100, 0.162, and 0.148) indicate that thesethree species could be present on the same evolutionary lineand could potentially have the same common ancestor, as theMP/ML tree suggests.
The bootstrap and posterior probabilities in the resultingtrees do, in some cases, provide very little support for theclades and do not constitute significant evidence of speciesdifference, which may be the result of the “long branchattraction phenomenon” (Wägele and Mayer 2007) andwhich is hard to avoid with our small data set. However,obtaining this kind of material is very difficult. Thus, it canbe stated that we successfully completed a valuable sampleset, even if it may appear small. The combined phylogeneticBA of mitochondrial CO1 and genomic 18S rRNA genemarkers in our representatives of the phylum Nematoda alsosupports the separate position of E. buckleyi among the genusEnterobius and subgenus Enterobius. For strongerconfirmation of the validity of the genera, further analysesare required.
Quentin et al. (1979), who drew attention to the morpho-logical differences between E. buckleyi and the speciesparasitising hosts of the family Hominidae (Homininae) andCercopithecidae (Cercopithecinae), classified E. buckleyi inthe subgenus Enterobius. In addition, Hugot (1999) recom-mended keeping this species in the subgenus Enterobius.When Quentin and coauthors characterised the subgenusColobenterobius (1979), they did not include E. buckleyi inthe list of species of this subgenus (Quentin et al. 1979 Hugot1987). However, even though no species of the subgenusColobenterobius has been genetically studied so far, the
Table 2 Average evolutionary divergences (d) estimated by the Kimuratwo-parameter test over sequence pairs within defined systematic groups
D SE
Genus
Enterobius 0.083 0.009
Trypanoxyuris 0.144 0.022
Lemuricola 0.084 0.012
Syphacia 0.056 0.011
Pongobius 0.005 0.002
Ascaris 0.076 0.011
Subgenus
Enterobius 0.046 0.007
Trypanoxyuris n/c n/c
Buckleyenterobius n/c n/c
Lemuricola n/c n/c
Syphacia 0.056 0.01
Protenterobius 0.006 0.003
Colobenterobius 0.036 0.006
Pongobius 0.005 0.002
Ascaris 0.076 0.011
Species
Enterobius vermicularis 0.036 0.007
Trypanoxyuris microon n/c n/c
Trypanoxyuris atelis n/c n/c
Lemuricola bauchoti n/c n/c
Enterobius anthropopitheci n/c n/c
Syphacia montana 0.024 0.006
Syphacia frederici 0.009 0.004
Lemuricola pongoi 0.006 0.003
Enterobius buckleyi 0.036 0.006
Pongobius hugoti 0.005 0.002
Ascaris suum n/c n/c
Ascaris columnaris n/c n/c
Ascaris sp. n/c n/c
SE standard error, n/c cases in which it was not possible to estimateevolutionary distances
2460 Parasitol Res (2014) 113:2455–2466

classification of the species E. buckleyi under this subgenus issupported by morphological and zoogeographical aspectsalone. Moreover, since the morphological and genetic signssignificantly distinguish the taxon E. buckleyi and our phylo-genetic analyses demonstrate this, we support the opinion thatE. buckleyi should be classified as Colobenterobius. At thepresent time, the subgenus Colobenterobius comprises 12specialised species exclusively parasitising the subfamilyColobinae (family Cercopithecidae). Our results support thehypothesis of co-speciation with host-switching accounts ofparasite [E. (C.) buckleyi] speciation. A specialised study willbe necessary to establish whether the subgenera Enterobius(E. vermicularis) and Colobenterobius (E. buckleyi) should bepromoted to separate genera. This idea is supported by theresults of the phylogram produced by Bayesian analysis of thecombined data set (Fig. 4). According to both phylograms ofour results, the suggestion of the early separation of the cladeof E. buckleyi (Colobenterobius) from the Enterobius(Enterobius) branch is obvious. This course connotes thatthe clade of the genus and (subgenus) Enterobius (Enterobius)is phylogenetically closer to the Lemuricola (Protenterobius)and Pongobius groups.
Further branching of the phylogenetic tree conclusivelysupports the line to the genus Lemuricola (subgenusMadoxyuris). Its recently described species [P. hugoti, L. (P.)pongoi] are specialised parasites of orang-utans in Sumatra
and Borneo. The cladistics of these species have not beenstudied, and barcoding has been carried out only once and,therefore, cannot be compared with the data from Hugot(1999) and Nakano et al. (2006). The determined divergence(14.4±0.02 %) in the Trypanoxyuris group (also indicated inTables 2 and 3) is very high among individuals of one genusand could indicate that T. (Buckleyenterobius) atelis andT.(Trypanoxyuris) microon have a very ancient common an-cestor and should be assigned to two different subgenera (or,better, genera), as was suggested byHasegawa et al. (2004). Inour opinion, the 8 % divergence between sequences of poten-tial species represents the boundary value among genera andsubgenera. However, this large divergence in ourTrypanoxyuris group could be distorted by the use of onlyone database sequence from the particular species. The valid-ity of dividing this group into separate subgenera—e.g.,Trypanoxyuris 2 and Paraoxyuronema as indicated byHugot (1999)—should be confirmed by the analysis ofmore samples, even though our results correspond withhis hypothesis. According to the interpretation ofNakano et al. (2006), the species T. microon (subgenusTrypanoxyuris) and T. atelis (subgenus Buckleyenterobius)diverge from their common ancestral basis, which was re-vealed in our research, too. This difference suggests the pos-sibility of elevating the taxon from the subgenera level to thelevel of genus.
Table 3 Evolutionary divergence over sequence pairs between defined groups (shown in asterisk)
Parasitol Res (2014) 113:2455–2466 2461

Tree lineages of P. hugoti and Lemuricola pongoi (subge-nus Protenterobius) suggest the earlier divergence ofL. bauchoti (subgenus Madoxyuris) from their common an-cestor. The morphological characteristic of the group that alsosupports the affinity between Lemuricola, Pongobius, and
Trypanoxyuris is the presence of a tail appendix in males,which is reduced in the subgenus Madoxyuris (Hugot et al.1996 Hugot 1999).
Hasegawa et al. (2004) point out that the characteristicform of the oesophagus in both sexes is a significant
Fig. 3 Alignment comparison of partial ITS2 sequences with the treemajor ITS types defined by Nakano et al. (2006). EV_ITS2_consensusindicates the consensus sequence aligned from ITS2 data of
E. vermicularis from three Czech samples. The alignment was conductedusing the BioEdit Sequence Alignment Editor v.7.0.9.0
2462 Parasitol Res (2014) 113:2455–2466

systematic trait in primate pinworms (Fig. 5a, b). The mor-phological differences are very notable between P. hugoti andL. (P.) pongoi (cephalic and bucal morphology, length andshape of oesophagus and other characters), and these are welldocumented by Baruš et al. (2007) and Foitová et al. (2008,2010). In compared CO1 data on P. hugoti, little variability(four variable sites) was observed, and software evaluated thisas very little or zero divergence. The number of differences innucleotide sequences within the P. hugoti group was estimatedat 0.5±0.2 %. The estimation of evolutionary divergence oversequence pairs between groups (Table 3) also shows thatP. hugoti and L. (P.) pongoi had minimal (the lowest) substi-tutions per site (10 %). In earlier studies, the maximumpairwise CO1 sequence distance among pinworm species
was estimated at 6.5 and 6.0 % by Nakano et al. (2006) andBlouin (2002), respectively. This could be an indication thatthese two taxa [P. hugoti and L. (P.) pongoi] became morpho-logically differentiated only recently.
The CO1 tree diagram also confirms the monophyletic originof Pongobius and Lemuricola (Protenterobius). The affinity ofpinworms for parasitising the Asian Orang-utan, Nycticebuscoucang (Boddaert) and that of Madagascarian primate pin-worms for parasitising lemurids have already drawn considerableattention (Inglis and Dunn 1963), and Protenterobius is consid-ered to be a junior objective synonymum of Lemuricola(Chabaud et Petter, 1959). Baruš et al. (2007) and Foitová et al.(2010), in accordancewithHugot (1999) andHugot et al. (1996),recognise Protenterobius as a valid subgenus.
Fig. 4 Resulting phylogramproduced by Bayesian analysis ofthe combined gene dataset(CO1+18S). The posteriorprobabilities are shown above thebranches. Numbers in parenthesisindicate the number of individualsrepresenting the sequence
Parasitol Res (2014) 113:2455–2466 2463

The frequently discussed Cameron hypothesis (Cameron1929) was confirmed by our results at the level of rela-tions among the three main genera (with subgenera)groups:
Enterobius comprises the parasites of CatarrhiniLemuricola and Pongobius comprise the parasites ofStrepsini; and Pongo (Hominidae: Ponginae) +Nycticebus (Lorisidae) and Trypanoxyuris comprise theparasites of Platyrrhini. Contrary to the analysis carriedout by Hugot (1999) within each of the three maingroups, the subdivision of these parasitic groups is dif-ferent in terms of determining potentially very similarmonophylogenetic origins for Lemuricola (Pongobius,Protenterobius and Madoxyuris) and Trypanoxyuris
(Trypanoxyuris, Buckleyenterobius) (Fig. 6a, b). Howev-er, up to date, no supporting molecular evidence on thetaxa Colobenterobius, Oxyuronema and Hapaloxyurishas been collected.
Considering the described type B CO1 amino acid group,Nakano et al. (2006) reported the presence of the same mo-lecular variant and, in addition, the high similarity of CO1sequences in the E.(E.) vermicularis genome in both chim-panzees (captive) and humans in Japan. Our proof of theexistence of the B variant in humans supports the opinion ofNakano et al. (2006), which claims that these CO1 aminotypes were already present in the E. (E.) vermicularis popula-tion in humans, and that chimpanzees acquired them fromhumans following capture.
Fig. 6 a Schematic illustration of selected data (genera, subgenera, species) on Enterobiinae parasitizing Primates. According to Hugot (1999) cladisticanalysis, b the schema of the genetic tree (genera, subgenera, species) of Enterobiinae parasitizing Primates according to our analysis
Fig. 5 Oesophagus morphology.a P. hugoti—oesophagus with along pharyngeal part, corpusposteriorly slightly enlarged andcontinuing as a pyriform bulb,isthmus absent. b L.(P.) pongoi—oesophagus with a shortpharyngeal part, corpus slightlyenlarged and continuing in shortisthmus, bulb rounded
2464 Parasitol Res (2014) 113:2455–2466

Conclusions
Our study broadens current knowledge on Nematoda moleculardata and resolves the generic affiliation of the studied species.
The results of molecular analysis provide evidence that wehave discovered a new type of ITS2molecule in E. vermicularisthat was not previously described. Moreover, we provide evi-dence that pinworms can be transferred from apes to humansand vice versa, in virtue of our discovery ofE. vermicularis CO1molecules of type B in our samples from human hosts.
On the basis of the performed phylogenetical analyses, wesuggest separating Enterobius buckleyi from the subgenusEnterobius, and, on the basis of our further findings, classify-ing it under the subgenus Colobenterobius. For the proposedclassification of E. buckleyi as a separate genus, more detailedstudy with a larger number of samples is required.
Concerning the—in our opinion—incorrect categorisationof the genus Trypanoxyuris, we suggest more detailed study toconfirm the differences between T. microon and T. atelis at themolecular level.
Even though we performed only preliminary phylogeneticstudies by various means, the impact of using different ana-lytical methods on the obtained topology of the trees would beminimal. This should strengthen the validity of our outcomes.
Acknowledgements The authors would like to thank the State Ministryof Research and Technology (RISTEK) and the Directorate for General forNatural Conservation (PHKA) for their cooperation and for their permissionto conduct research in the Gunung Leuser National Park. The study wasfinancially supported by the UMI—Saving of Pongidae Foundation project“Parasites and Natural Antiparasitics in the Orang-utan” and by the CzechAcademy of Sciences, grant no. P505/11/1163. We thank Dr. MrkvicováMartina for help with molecular analysis and Dr. Tóthová Andrea forproviding the Bayesian analysis and phylogenetic consultations. We aregrateful to Matthew Nicholls for English correction of the draft.
References
BarušV, Foitová I, Koubková B, Hodová I, ŠimkováA, NurcahyoW (2007)A new nematode, Pongobius hugoti gen. et sp. n. from the orangutanPongo abelii (Primates: Hominidae). Helminthologia 44:162–169
Blouin MS (2002) Molecular prospecting for cryptic species of nema-todes: mitochondrial DNA versus internal transcribed spacer. Int JParasitol 32:527–531
Brooks D (1982) Pinworms and primates: a case study of coevolution.Proc Helminthol Soc Wash 49:76–85
Cameron TWM (1929) The species of Enterobius Leach in primates. JHelminthol 7:161–182
Chabaud AG, Petter AJ (1959) Les Nématodes parasites de LémuriensMalgaches II. Un nouvel Oxyure: Lemuricola contagiosus. MémInstitut Sci Madagascar 13:127–132
Chabaud AG, Petter AJ, Golvan Y (1961) Les Nématodes parasites deLémuriens Malgaches III. Collection récoltée par M. et MmeFrancis Petter. Ann Parasitol Hum Comp 36:113–126
Floyd RMA, Rogers D, Lambshead JD, Smith CR (2005) Nematode-specific PCR primers for the 18S small subunit rRNA gene. MolEcol Notes 5:611–612
Foitová I, Baruš V, Hodová I, Koubková B, Nurcahyo W (2008) Tworemarkable pinworms (Nematoda: Enterobiinae) parasitizing orangu-tan (Pongo abelii) in Sumatra (Indonesia). Helminthologia 45:162–168
Foitová I, Baruš V, Koubková B, Mašová Š, Nurcahyo W (2010)Description of Lemuricola (Lemuricola) pongoi–male (Nematoda:Enterobiinae) parasitizing orangutan (Pongo abelii). Parasitol Res106:817–820
Glen DR, Brooks DR (1986) Parasitological evidence pertaining to thephylogeny of the hominoid primates. Biol J Linn Soc Lond 27:331–354
Gonzáles-Moreno O, Domingo L, Teixidor J, Gracenae M (2011)Prevalence and associated factors of intestinal parasitisation: across-sectional study among outpatients with gastrointestinal symp-toms in Catalonia, Spain. Parasitol Res 108:87–93
Hall TA (1999) BioEdit: a user-friendly biological sequence alignmenteditor and analysis program for Windows 95/98/NT. Nucleic AcidsSymp 41:95–98
Hasegawa H (1999) Phylogeny, host–parasite relationship and zoogeog-raphy. Korean J Parasitol 37:197–213
Hasegawa H, Matsuo K, Onuma M (2003) Enterobius (Colobenterobius)serratus sp. nov. (Nematoda: Oxyuridae) from the Proboscis Monkey,Nasalis larvatus (Wurmb, 1787) (Primates: Cercopithecidae:Colobinae), in Sarawak, Borneo,Malaysia. CompParasitol 70:128–131
Hasegawa H, Ikeda Y, Diaz-Aquino DJJ, Fukui D (2004) Redescription oftwo pinworms from the black-handed spider monkey,Ateles geoffroyi,with reestablishment of Oxyuronema and Buckleyenterobius(Nematoda: Oxyuroidea). Comp Parasitol 71:166–174
Hasegawa H, Ikeda Y, Fujisaki A, Moscovice LR, Petrželková KJ, KaurT, Huffman MA (2005) Morphology of chimpanzee pinworms,Enterobius (Enterobius) anthropopitheci (Gedoelst, 1916)(Nematoda: Oxyuridae), collected from Chimpanzees, Pantroglodytes, on Rubondo Island, Tanzania. J Parasitol 91:1314–1317
Huelsenbeck JP, Ronquist FR (2001) MrBayes: Bayesian inference ofphylogeny. Biometrics 17:754–755
Hugot JP (1984a) Sur le genre Trypanoxyuris (Oxyuridae, Nematoda): I.Parasites de Sciuridés: sous-genre Rodentoxyuris. Bull Mus NatlHist Nat Sér A Zool 6:711–720
Hugot JP (1984b) Sur le genre Trypanoxyuris (Oxyuridae, Nematoda) II.Sous-genre Hapaloxyuris parasites de Primates Callitrichidae. BullMus Natl Hist Nat Sér A Zool 6:1007–1019
Hugot JP (1987) Sur le genre Enterobius (Oxyuridae, Nematoda): s.g.Colobenterobius I. Parasites de Primates Colobinae en regionÉthiopienne. Bull Mus Natl Hist Nat Sér A Zool 9:341–352
Hugot JP (1999) Primates and their pinworm parasites: Cameron hypoth-esis revisited. Syst Biol 48:523–546
Hugot JP, Baylac M (2007) Shape patterns of genital papillae in pin-worms (Enterobiinae, Oxyurida, Nematoda) parasite of primates: alandmark analysis. Inf Gen Evol 7:168–179
Hugot JP, Morand S, Gardner SL (1995)Morphology andmorphometricsof three oxyurids parasitic in primates with a description ofLemuricola microcebi n. sp. Int J Parasitol 25:1065–1075
Hugot JP, Gardner SL, Morand S (1996) The Enterobiinae subfam. nov.(Nematoda, Oxyurida) pinworm parasites of primates and rodents.Int J Parasitol 26:147–159
Inglis WG (1961) The oxyurids parasites (Nematoda) of primates. P ZoolSoc Lond 136:103–122
Inglis WG, Dunn FL (1963) The occurrence of Lemuricola (Nematoda:Oxyurida) in Malaya: with the description of a new species. ZParasitenk 23:354–359
Kimura M (1980) A simple method for estimating evolutionary rate ofbase substitutions through comparative studies of nucleotide se-quences. J Mol Evol 16:111–120
Kuze N, Kanamori T, Malim TP, Bernard H, Zamma K, Kooriyma T,Morimoto A, Hasegawa H (2010) Parasites found from the feces ofBornean Orangutans in Danum Valley, Sabah, Malaysia, with aredescription of Pongobius hugoti and the description of a newspecies ofPongobius (Nematoda: Oxyuridae). J Parasitol 96:954–960
Parasitol Res (2014) 113:2455–2466 2465

Nakano T, Okamoto M, Ikeda Y, Hasegawa H (2006) Mitochondrialcytochrome c oxidase subunit 1 gene and nuclear rDNA regions ofEnterobius vermicularis parasitic in captive chimpanzees with spe-cial reference to its relationship with pinworms in humans. ParasitolRes 100:51–57
Nylander JAA (2004) MrModeltest 2.2. Program distributed by theauthor. Evolutionary Biology Centre Uppsala University
Petter AJ, Chabaud AG, Delavenay R, Brygoo ER (1972) Une nouvelleespéce de Nématode du genre Lemuricola, parasite deDaubentoniamadagascariensis Gmelin, et considérations sur le genreLemuricola. Ann Parasitol Hum Comp 47:391–398
Quentin JC, Tenora F (1975) Morphologie et position systématique deLemuricola (Rodentoxyuris) sciuri (Cameron, 1933) nov. comb.,nov. subg., et Syphacia (Syphatineria) funambuli Johnson, 1967.Oxyures (Nematoda) parasites de Rongeurs Sciurides. Bull MusNatl Hist Natl Ser A Zool 178:1525–1535
Quentin JC, Betterton C, Krishnasamy M (1979) Oxyures nouveaux oupeu connus, parasites, de Primates, de Rongeurs et de Dermoptèresen Malaisie. Création du sous-genre Colobenterobius n. subgen.Bull Mus Natl Hist Nat Sér A Zool 1:1031–1050
Remm M (2006) Distribution of enterobiasis among nursery schoolchildren in SE Estonia and of other helminthiases in Estonia.Parasitol Res 99:729–736
Rim HJ, Chai JY, Min DY, Cho SY, Eom KS, Hong SJ, Sohn WM,Yong TS, Deodato G, Standgaard H, Phommasack B, Yun CH,Hoang EH (2003) Prevalence of intestinal parasite infections ona national scale among primary schoolchildren in Laos. ParasitolRes 91:267–272
SandoshamAA (1950) On Enterobius vermicularis (Linnaeus, 1758) andsome related species from primates and rodents. J Helminthol 24:171–204
Song HJ, Cho CH, Kim JS, Choi MH, Hong ST (2003) Prevalence andrisk factors for enterobiasis among preschool children in a metro-politan city in Korea. Parasitol Res 91:46–50
Sorci G, Morand S, Hugot JP (1997) Host parasites coevolution:comparative evidence for covariation of life history traits inprimates and oxyurid parasites. P Zool Soc Lond 264:285–289
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S(2011) MEGA5: molecular evolutionary genetics analysisusing maximum likelihood, evolutionary distance, andmaximum parsimony methods. Mol Biol Evol 28(10):2731–2739
Wägele JW, Mayer C (2007) Visualizing differences in phylogeneticinformation content of alignments and distinction of three classesof long-branch effects. BMC Evol Biol 7:147
2466 Parasitol Res (2014) 113:2455–2466