physiol. res. 40:481-491, 1991 the beta-adrenergic
TRANSCRIPT

Physiol. Res. 40:481-491, 1991
The Beta-Adrenergic Receptor-Regulated 1,4-Dihydropyridine Calcium Channel Receptor Sites in Coronary Artery Smooth M uscle
J. DŘÍMAL
Institute o f Experimental Pharmacology, Slovak Academy of Sciences, Bratislava
Received December 10, 1990Accepted June 26, 1991
Summary __________________ ;________________________________The cross-regulatory communication from /3-adrenergic receptors to1,4-dihydropyridine (DHP) Ca2+ channel agonist and antagonist binding sites and cooperativity between DHP binding sites were studied in microsomal membranes of canine coronary artery (purified to a factor 2.9 for DHPs). The maximal number of binding sites (Bmax) identified in coronary artery microsomal membranes (CAM) with Ca2+ channel agonist ( - )-S-(3H)BAY K 8644 was two times higher than Bmax °f sites labelled with Ca2* channel antagonist ( + )-(3H)PN 200-110. The exposure of CAM to isoprénaline was accompanied with down-regulation of /3-adrenergic receptors and with increase in binding capacity for DHPs. The increase in B^ax was proportional in both groups of experiments and was related to increased affinity of DHPs. The 1,4-DHP binding sites identified in vascular smooth muscle showed characteristics typical for classification of specific 1,4-DHP receptor on Ca2+ channels. The binding was of high affinity, saturable and reversible, it showed stereoselectivity and it was positively modulated by /3-adrenergic stimulation and it showed cAMP and GTP sensitivity. The results support the hypothesis that /3-receptors also regulate the mode of Ca2+ channels in coronary artery smooth muscle.
Key wordsCoronary artery - Smooth muscle - Microsomal membranes - L-type Ca2+ channels - (~)-S-(3H)BAYK8644 - (+)-(3H)PN 200-110 - (-)-3-(125I)iodocyanopindolol - Ligand binding
Introduction
Two types of calcium channels, similar to the Ca2+ channels in heart muscle have been found in cells from a variety of mammalian arteries (Bean et al 1986, Benham et aL 1987) and in aortic cell lines (Cohen and McCarthy 1987, Friedman etaL 1986).
Considerable experimental evidence has accumulated in the last few years indicating that voltage-sensitive and possibly also receptor-regulated Ca2+ channels are regulated by cAMP and protein phosphorylation (Armstrong and Eckert 1987, Trautwein and Kameyama 1986). The Ca2+ channels in cardiac and in vascular smooth muscle, however, may not be precisely identical since the Ca2+ channels in

482 Dřímal Vol. 40
vascular smooth muscle are much more sensitive to 1,4-dihydropyridine Ca2+ channel antagonists (Bean et al. 1986) and possibly do not gain possession of that type of Ca2+ channel that is phosphorylated by cAMP-dependent protein kinase (Worley et al 1986). Evidence has also emerged for direct coupling of G protein to Ca2+ channel (Trautwein and Herscheller 1990).
Recent studies have demonstrated the presence of specific, saturable and high-affinity binding sites for 1,4-dihydropyridines also in porcine (DePover et al. 1982, Muramatsu et al 1985, Nishimura et al 1987, Yamada et al. 1988, 1990) and in canine coronary arteries (Dřímal et al 1988).
The purpose of the present study was to evaluate the cross-regulatory communication from ̂ -adrenergic to 1,4-dihydropyridine Ca2+ channel binding sites in microsomal membranes of coronary artery smooth muscle. These studies were performed in an effort to examine further the effects of /3-adrenergic receptor activation on cooperativity of 1,4-dihydropyridine Ca2+ channel agonist and Ca2+ channel antagonist receptor sites in microsomal membranes of coronary artery smooth muscle.
Methods
Male mongrel dogs (20 - 25 kg) were anaesthetized with sodium pentobarbital (30 mg/kg intravenously) and the hearts were rapidly removed. Segments of the large coronary arteries (ramus anterior descendens and ramus circumflexus of the left coronary artery) were dissected, cleaned from myocardial adipose and connective tissue, opened longitudinally and the intima and adventitia were carefully removed. The cleaned medial layers of the coronary artery were minced with scissors and homogenized in nine volumes of 0.35 mol/1 of ice cold sucrose and 5 mmol/1 of Tris-HCl (pH = 7.51).
Membrane preparation
Microsomal membranes of vascular smooth muscles were prepared according to Kwan et al. (1983), with the exception that the medial layers of large coronary arteries were prepared from postnuclear supernatant in accordance with Dřímal et al. (1987). Preparations were treated in an Ultraturrax homogenizer. Homogenates of the media were pressed under vacuum through stainless wire mesh and centrifuged further at 1500 x g for 10 min at 4 °C. The supernatant was centrifuged further at 105 000 x g for 45 min on a MOM Ultracentrifuge. The resulting pellet was resuspended in an assay buffer (50 mmol/1 Tris-HCl, pH = 7.51) in a Potter-Elvehjem homogenizer and used as crude microsomal fraction. Refined microsomal fractions were prepared from the supernatant which sedimented in a sucrose buffer (8-60 % w/v sucrose gradient) at 130 000 x g using fixed angle rotor and centrifugation time of 60 min.
Enzyme assays
The measurements of 5’-nucleotidase and Mg2+-ATPase activity were accomplished according to Kwan et al. (1984) and Pullman (1967). Spectrophotometric determinations were performed on a Beckman Model 25 double-beam spectrophotometer. The assay mixture for the Mg2+-ATPase contained (in mmol/1): sucrose (0.25), Tris-acetate (50.0), EDTA (0.5), glucose (25.0), MgCh (5.0), potassium-phosphate (20.0) and 1.5 x 104 cpm of 32P (specific activity 5 mCi/g) and hexokinase Type 1 0 - 3 Mg/mg of protein (70 Mg)- Protein was assayed by the method of Bradford (1976).

483
measured by incubating (25 °C) microsomal membranes with the refined microsomal fraction for30 min at various concentrations of /¡-adrenergic ligands (from 0.1 to 30 nmol/1) of (3H)DHA and (~ )-3-(125I)iodocyanopindolol (up to 1.0 nmol/1); c-AMP content was assayed by the (^l)-method (IRAPRA, Prague) as described previously (DHmal 1989,1991).
8*S(-)-S-(*H)BAY K 8,
0 ;(+)-(3H)PN 200-110 (125I)iodocyanopindolol
X) Ci/mihydrocft'oride (Fluka); BAY K 8644 (Calbiochem); verapamil (Knoll).
Results
Membrane characteristics and enzyme activity
The refined microsomal fractions (RM-1, RM-2 and RM-3) obtained from the continuous sucrose density gradient were morphologically more homogeneous than crude microsomes. When examined by electron microscopy, these fractions contained morphologically sealed membrane vesicles of various sizes, membrane fragments derived from the endoplasmatic reticulum (RM-1 and RM-3) and mitochondrial and other fragments of intracellular membranes. The RM-2 was much more homogeneous.

484 Dřímal Vol. 40
Table 1Enzyme activity of postnur.lear and microsomal membrane fractions isolated from
the media of canine coronary arteries by differential centrifugation and sucrose density gradient centrifugation
Fraction PNS Sucrose density gradient
P CM RM-110/15
RM-2 RM-3 20/30 30/40
SOL
5’-nudeotidase 1.5 16 3.8 44 8.9 1.2 0.8±0.04 ±0.09 ±0.1 ±0.9 ±1.7 ±0.5 ±0.6
Mg2+-ATPase 8.0 10.0 25.0 33.0 96.0 6.0 0.5(/¿mol/mg/h) +0.1 +0.9 +9.0 + 17.0 + 19.0 +4.0 +4.3
Values are Means ± S.E.M. (n=5); PNS - postnuclear supernatant, P - particulate, CM - crudemicrosomal fraction, RM-1 to RM-3 - refined microsomal fractions, SOL - soluble fractions
Table 2Effect o f isoprenaline of cAMP content ir,t microsomal membranes o f coronary artery
cAMP content (pmol/mg protein)
Control 0.85 ±CIsoprenaline 5.08 ±0.50
Values are Means ± S.E.M. (n=8)
Enzymatically RM-2 fraction (Tab. 1) showed the highest marker enzyme activity of two major membrane enzyme markers 5’-nucleotidase and Mg2+-ATPase. The adenylate-cyclase activity is summarized in Tab. 2. Short-term incubation of the left coronary artery preparation (ramus circumflexus) with isoprenaline (1.0/xmol/l; n = 4) significantly increased the cAMP content in RM-2 fraction obtained from that preparation.
P-adrenoreceptor numbers in microsomal membranes following exposure to isoproterenol
To elucidate the effects of prolonged /3-adrenoreceptor stimulation in the coronary artery on the number of microsomal membrane receptors we compared the ability of ̂ -adrenergic ligands ((3H)DHA and (-)-3-(125I)iodocyanopindolol) to bind to microsomal membranes prepared from control unexposed coronary arteries and from those exposed to isoprenaline for one hour.
We have studied /S-adrenergic receptor in the coronary artery using two /S-adrenergic ligands. Saturation binding experiments with (3H)DHA under control
1991 Coronary Artery 1,4-DHP Receptor 485
conditions showed Kj 5.2±0.2 nmol/1 and Bmax9.60±0.90 fmol/mg protein. The exposure of coronary artery preparations to isoprénaline significantly reduced (-)-3-(125I)iodocyanopindolol specific binding from 13.4 ±0.88 fmol/mg protein to 8.9 ±1.2 fmol/mg protein (n = 10) while the ( - )-3-(125I)iodocyanopindolol affinity for this binding site was unaltered averaging 6.21 ±0.16 nmol/1.
Identification o f 1,4-dihydropyridine Ca2+ channel antagonist binding sites in microsomal membranes
Specific binding of Ca2+ channel agonist to microsomal membranes (hereafter referred to as membrane inasmuch as only the RM-2 fraction was used in1.4- dihydropyridine binding experiments) was rapid, reaching steady state within 20 min (at 25 °C). This steady state was maintained for at least 70 min. The calculated association rate measured in kinetic experiments using 0.90 nmol/1 of ( - )-S-(3H)BAY K 8644 was 0.31 min'1. The microsomal membranes in our experiments had a purification factor 2.1 for /3-adrenergic radioligands and 2.9 for1.4- dihydropyridines.
(pmol.mp-lprdttlii) (p mol. raj 4 prot.in)
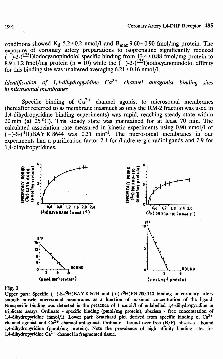
Fig. 1Upper part: Specific (-)-S-(3H)BAY K8644 and (+)-(3H)PN 200-110 binding in coronary artery smooth muscle microsomal membranes as a function of maximal concentration of the ligand. Nonspecific binding was detected in the presence of 1 /rmol/1 of unlabelled 1,4-dihydropyridine in triplicate assays. Ordinate - specific binding (pmol/mg protein), abscissa - free concentration of1.4- dihydropyridine (nmol/1). Lower part: Scatchard plot derived from specific binding of Ca2+ channel agonist and Ca2+ channel antagonist. Ordinate - bound over free (B/F), abscissa - bound1.4- dihydropyridine (pmol/mg protein). Note the prevalence of high affinity binding sites for1.4- dihydropyridine Ca2+ channel in fragmented tissue.
486 Dřímal Vol. 40
The binding of ( - )-S-(3H)BAY K 8644 and ( + )-(3H)PN 200-110 to partially purified microsomal membranes was specific and saturable over a concentration range from 0.14 to 2.4 (or 0.14 to 3.0) nmol/1, respectively (Fig. 1). The linear regression analysis of untransformed experimental data showed adequate affinity and relatively high capacity of binding sites with the corresponding Bmax values 7.15±0.04 and 3.40±0.18 pmol/mg protein and Kd values 18.0±0.1 and 9.0±0.8 nmol/1, respectively. The Scatchard analysis in each group of experiments waslinear, indicating that a single population of high affinity binding si
The displacement studies in competition experiments showed effective displacement of nitrendipine by unlabelled nitredipine and by an analogue of nimodipine DHPS-2 VULM (Fig. 2). The K; values calculated from concentrations that inhibited specific (-)-S-(3H)BAY K 8644 binding by 50 % for optical isomers of BAY K 8644 (data not shown) displayed stereoselectivity for the optic isomer of ( - )-S-(3H)BAY K 8644.
Effects o f drugs on ( - )-S-(^H)BAYK8644 and (+)-(iH)PN 200-110 specific binding in microsomal membranes
The effects of structurally unrelated Ca2+ channel antagonist and of pharmacologically active drugs on 1,4-dihydropyridine Ca2+ channel agonist and Ca2+ channel antagonist binding were studied to investigate further the cooperativity of interactions between 1,4-dihydropyridine binding sites and modulatory effects of different pharmacologically active drugs on1,4-dihydropyridine binding. Additional evidence for the existence of the

1991 Coronary Artery 1,4-DHP Receptor 487
modulatory role of basic cellular signals in the regulations of 1,4-dihydropyridine Ca2+ channel receptor was obtained in experiments with four different procedures: ^-adrenergic receptor stimulation, GTP, cAMP and EDTA presence in the system.
CORONARY SMOOTH MUSCLE MICROSOMAL MEMBRANES
(♦)
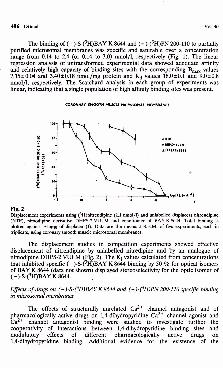
Fig. 3Pharmacology of 1,4-dihydropyridine Ca2+ channel antagonist (+)-(3H)PN 200-110 and Ca2+ channel agonist (-)-S-(3H)BAYK8644 binding in coronary artery smooth muscle microsomal membranes. The concentration of ( )-(3H)PN 200-110 and (-)-S-(3H)BAY K 8644 binding sites after incubation of coronary artery preparation with isoprenaline (P, 1.0); dibutyryl-cAMP (db cAMP, 1.0); guanylylimidodiphosphate (Gpp(NH)p, 10.0); nitrendipine (NTP, 1.0); d-cis-dilthiazem (DIL, 10.0) and dipyridamole (DIPY, 100.0); all in/rmol/1.
As is shown in Fig 3, the exposure of a coronary artery preparation to isoprenaline was accompanied by increased density of ( - )-S-(3H)BAY K 8644 and (+)-(3H)PN 200-110 binding. The increase in Bmax was proportional in both groups of experiments (+56 ±7 and +40 ±9 %) and was mostly due to the increased affinity of both 1,4-dihydropyridines for its receptors (Kd = 10.9 ±1.1 and 7.0 ±0.2 nmol/1, i.e. a reduction by -39±8 and -22±6 %, respectively, P<0.05). The involvement of G protein in observed response was characterized after pretreatment of membranes with guanylylimidodiphosphate, a nonhydrolyzable analogue of GTP. This pretreatment significantly decreased the maximal capacity of (-)-S-(3H)BAY K 8644 binding (-37± 8 %, P<0.05). The nonhydrolyzable analogue of cAMP, dibutyryl-cAMP, significantly increased ( - )-S-(3H)BAY K 8644 maximal binding and increased affinity of Ca2+ channel agonist for its receptor (32 ±6 %). The non-1,4-DHP Ca2+ channel antagonist d-cis-dilthiazem significantly
Vol. 40
increased 1,4-dihydropyridine Ca2+ channel receptor binding ( + 81 ±11 %). The presence of chelator EDTA in the system significantly reduced 1,4-dihydropyridine binding (-8 7 ±5 %, P<0.05). Adenosine uptake inhibitor, dipyridamole,significantly reduced maximal capacity of ( - )-S-(3H)BAY K 8644 binding (-5 9 ±5 %, P<0.05).
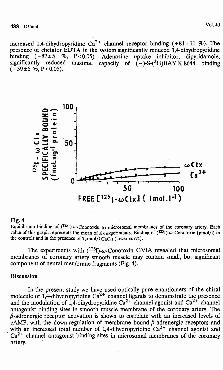
Fig. 4Equilibrium binding of (125I)-cu-Conotoxin to microsomal membranes of the coronary artery. Each value of this graph represents the mean of six experiments. Binding of (12sI)-<u-Conotoxm (pmol/1) in the controls and in the presence of 1 mol/1 CaCfe (lower curve).
The experiments with (125I)+y-Conotoxm GVIA revealed that microsomal membranes of coronary artery smooth muscle may contain small, but significant component of neural membrane fragments (Fig. 4).
Discussion
In the present study we have used optically pure enantiomers of the chiral molecule of 1,4-dihydropyridine Ca2+ channel ligands to demonstrate the presence and the modulation of 1,4-dihydropyridine Ca2+ channel agonist and Ca2+ channel antagonist binding sites in smooth muscle membrane of the coronary artery. The /3-adrenergic receptor activation is shown to correlate with an increased levels of cAMP, with the down-regulation of membrane bound ^-adrenergic receptors and with an increased total number of 1,4-dihydropyridine Ca2+ channel agonist and Ca2+ channel antagonist binding sites in microsomal membranes of the coronary artery.

1991 Coronary Artery 1,4-DHP Receptor 489
The total number of binding sites labelled with the Ca2+ channel agonist in microsomal membranes was more than two times higher that the number of binding sites identified with the Ca2+ channel antagonist. Binding sites identified with1.4- dihydropyridine Ca2+ channel agonist ( - )-S-(3H)BAY K 8644 showed a high affinity, saturability and reversibility, were stereoselective and showed a positive modulation with ^-adrenergic activation of the system, and displayed GTP and cAMP sensitivity. The Ca2+ channel antagonist ( + )-(3H)PN 200-110 binding showed a high affinity, saturability in microsomal membranes and positive modulation with ^-adrenergic receptor stimulation but it did not display GTP and cAMP sensitivity.
Compared to our data on 1,4-dihydropyridine binding in the coronary artery membrane preparation, lower Bmax values were reported for (3H)nitrendipine (561 fmol/mg protein) by Nishimura et al. (1987) and for (+)-(3H)PN 200-110 in porcine coronary artery membrane (Yamada et al. 1988, 1990, Muramatsu et aL 1985). These discrepancies could be due to species differences and/or the employment of different preparative procedures.
The goal of the present report was to study the systematic difference in the regulatory response of Ca2+ channel agonist and antagonist binding in the presence of high levels of GTP and cAMP in microsomal membranes. Contrary to the /l-adrenergic receptor stimulation resulting in a positive modulation of1.4- dihydropyridine binding in both our groups, the presence of nonhydrolyzable analogue of cAMP and GTP in other experiments failed to induce any significant change in the Bmax of antagonist. This finding may further support the hypothesis of a mudal mode of Ca2+ channel.
The /3-adrenergic receptor stimulation will increase both 1,4-dihydropyridine Ca2+ channel agonist and Ca2+ channel antagonist binding sites in microsomal membranes but in the presence of high levels of cAMP in microsomal membranes only 1,4-dihydropyridine Ca2+ channel agonist binding is increased by the "activation" of Ca2+ channels.
As in the cardiac muscle, the slowly inactivating type of Ca2+ channel in coronary vascular smooth muscle is sensitive to the activator BAY K 8644. Hughes et al. (1990) recently reported that agonist and antagonist enantiomers of1.4- dihydropyridine PN 202-791 may act at different sites on the voltage-dependent calcium channel of vascular smooth muscle.
A small but significant component of specific co-Conotoxin binding in microsomal membranes suggests the complexity of different "subtypes" of Ca2+ channels expressed in the coronary artery.
The allosteric regulation of 1,4-dihydropyridine binding with d-cis-dilthiazem, namely the stimulation of 1,4-dihydropyridine binding, has been demonstrated in various membrane preparations in different laboratories (DePover et aL 1982, Yamamura et al. 1982, Ferry et al 1982, Vaghy et al. 1987). In the present study we document very prominent stimulation of 1,4-dihydropyridine Ca2+ channel agonist binding in smooth muscle membranes of coronary arteries.
With findings of a profound inhibition of 1,4-dihydropyridine binding in microsomal membranes after pretreatment with an adenosine uptake blocker, dipyridamole, we confirm our recent results (Dřímal 1988) suggesting that the pharmacological receptor for 1,4-dihydropyridines in coronary artery smooth muscle membranes is associated with a nucleoside carrier.

490 Dřímal Vol. 40
The findings that 1,4-dihydropyridine Ca2+ channel antagonist stabilizes Ca2+ channel binding site? in the coronary artery smooth muscle membrane in an inactive mode and that /3-adrenergic receptor activation exerts modulatory effects on 1,4-dihydropyridine Ca2+ channel binding, may have important implications for understanding the mechanism of action of 1,4-dihydropyridine drugs on coronary artery smooth muscle.
Acknowledgement
The author would like to thank Mrs. B. Sedlarova and Mrs. Z. Veghova for their technical assistance.
References
ARMSTRONG D., ECKERT R.: Voltage-activated calcium channels that must be phosphorylated to respond to membrane depolarization. Proc. Nall. Acad. Sci. USA 84:2518-2522,1987.
BEAN B.P., STUREK M., PUGA A., HERMSMEYER K.: Calcium channels in muscle cells isolated from rat mesenteric arteries. Modulation by dihydropyridine drugs. Circ. Res. 59: 229 - 235, 1986.
BENHAM CA., TS1EN R.W.: A novel-receptor-operated Ca2+ permeable channel activated by ATP in smooth muscle. Nature 328:275-278,1987.
BRADFORD M.M.: A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 72: 248-254,1976.
COHEN C.J., MCCARTHY R.T.: Nimodipine block of calcium channels in rat vascular smooth muscle cells./. Gen. Physiol. 387:2412- 2416,1987.
DePOVER A., MATL1B MA, LEE S.W., DUBE G.P., GRUPP G., SCHWARTZ A.: Specific binding of (3H)nitrendipine to membranes from coronary arteries and heart in relation to pharmacological effects. Paradoxical stimulation by dilthiazem. Biochem. Biophys. Res. Commun. 108:110-117,1982.
DŘÍMAL J.: Regulation of /3-adrenergic receptors and calcium channel agonist binding sites in cultured human embryonal smooth muscle cells. Gen. Physiol. Biophys. 8:341-350,1989.
DŘÍMAL J.: Heterogeneity of calcium channel agonist binding sites in the coronary artery. Gen. Physiol. Biophys. 7:135-142,1988.
DŘÍMAL J.: Regulation of 1,4-dihydropyridine and ^-adrenergic receptor sites in coronary artery smooth muscle membranes. Cell Signalling 3:225 - 232,1991.
DŘÍMAL J., KNEZL V., MAGNA D., STŘÍŽOVÁ K.: External transport of /3-adrenergic binding sites in ischemic myocardium. Gen. Physiol. Biophys. 6:583 - 591,1987.
DŘÍMAL J , MAGNA D.,KNEZL V, SOTN1KOVÁ R.: Evidence that high affinity (3H)clonidine binding cooperates with ^-receptors in the canine coronary smooth muscle membrane. Agents Action 23:263 - 265,1988.
FERRY D.R., GLOSSMANN H.: Evidence for multiple receptor sites within the putative calcium channel. Naunyn-Schmiedeberg’s Arch. Pharmacol. 321:80 - 83,1982.
FRIEDMAN M.E., SUAREZ-KURTZ S„ KAIZOWSKI G., KATZ G.M., REUBEN J.P.: Two calcium currents in a smooth muscle cell line. Am. J. Physiol. 250: H699-H703,1986.
HUGHES A.D., HERING S., BOLTON T.B.: Evidence that agonist and antagonist enantiomers of the dihydropyridine PN 202-709 act at different sites on the voltage-dependent calcium channel of vascular smooth muscle. Br. J. Pharmacol. 101:3 - 5,1990.
KWAN C.Y., TRIGGLE C.R., GROOVER A.K., LEE R.M.K.W.: An analytical approach to the preparation and characterization of subcellular membranes from canine mesenteric arteries. Preparat. Biochem. 13:275 - 314,1983.
KWAN C.Y., TRIGGLE C.R., GROOVER A.K., LEE R.M.K.W., DANIEL E.E.: Membrane fractionation of canine aortic smooth muscle: subcellular distribution of calcium transport activity./. Mol. Cell. Cardiol. 16:747 - 764,1984.

1991 Coronary Artery 1,4-DHP Receptor 491
MURAMATSU M, FXJJITA A., TANAKA M., AIHARA H.: Effects of neuraminidase on dilthiazem-mediated alteration of nitrendipine binding in the coronary artery. Jpn. I. Pharmacol. 39:217-223,1985.
NISHIMURA J., KANAIDE H„ NAKAMURA M.: Characteristics of adrenoceptors and ^^nitrendipine receptors of porcine vascular smooth muscle. Differences between coronary artery and aorta. Circ. Res. 60:837-844,1987.
PULLMANN M.E.: Measurement of ATPase, 14C-ADP-ATP, and 32Pj-ATP exchange reactions. Meth. Enzymol. 10:57-60,1967.
ROSENTHAL H.E.: Graphic method for the determination and presentation of binding parameters in a complex system. Anal. Biochem. 20:525 - 531,1967.
TOBLER HJ., ENGEL J.: Affinity spectra: a novel way for the evaluation of equilibrium binding experiments. Naunyn-Schmiedeberg’s Arch. Pharmacol. 322:187-192,1983.
TRAUTWEIN W., HERSCHELLER J.: Regulation of cardiac L-type of calcium current by phosphorylation and G proteins. Annu. Rev. Physiol. 52:257-274,1990.
TRAUTWEIN W„ KAMEYAMA M.: Beta-adrenergic control of calcium channels in cardiac myocytes. In: Calcium Electrogenesis and Neuronal Functioning. U. HEINEMANN, M. KLEE, E. NEHER, W. SINGER (eds), Exp. Brain Res. Ser. Vol. 14, Springer, Berlin, 1986, pp. 185-195.
VAGHY P.L., WILLIAMS J.S., SCHWARTZ A.: Receptor pharmacology of calcium entry blocking agents. Am. J. Cardiol. 59:4A-17A, 1987.
WORLEY J.F., DEITMAR J.W., NELSON M.T.: Single nisoldipine-sensitive calcium channels in smooth muscle cells isolated from rabbit mesenteric artery. Proc. Natl. Acad. Sci. USA 83:5746- 5750,1986.
YAMADA S., HARADA Y., NAKAYAMA K.: Characterization of Ca2+ channel antagonist binding sites labelled by (3H)nitrendipine in coronary artery and aorta. Eur. J. Pharmacol. 154:203 - 208,1988.
YAM ADA S, KIMURA K., HARADA Y., NAKAYAMA K.: Ca2+ channel receptor sites for (+)-(3H)PN 200-110 in coronary artery./. Pharmacol. Exp. Ther. 252:327-332,1990.
YAMAMURA H.I., SCHOEMAKER H., BOLES R.G., ROESKE W.R.: Diltiazem enhancement of [3H]nitrendipine binding to calcium channel associated drug receptor sites in rat brain synaptosomes. Biochem. Biophys. Res. Commun. 108:640-646,1982.
Reprint requestsDr. J. Dřímal, Institute of Experimental Pharmacology, Slovak Academy of Sciences, CS-84216 Bratislava, Dúbravská cesta 2.