polymorphisms in dna regulation - · pdf filepolymorphisms in dna regulation - ... marco...
TRANSCRIPT

POLYMORPHISMS IN DNA REGULATION -ASSOCIATED GENES AND GASTRIC CANCER DEVELOPMENT MSc dissertation in Medicine and Molecular Oncology submitted to the FACULTY OF MEDICINE, UNIVERSITY OF PORTO
Marco António Assunção das Neves
Molecular Oncology and Viral Pathology Group (CI-IPOP),
IPO Porto FG EPE, Porto, Portugal
SUPERVISION
Hugo Manuel Lopes de Sousa, MD, PhD
Virology Service & Molecular Oncology and Viral Pathology Group (CI-IPOP),
IPO Porto FG EPE, Porto, Portugal
CO-SUPERVISION
Joana Patrícia Costa Ribeiro, MSc
Molecular Oncology and Viral Pathology Group (CI-IPOP),
IPO Porto FG EPE, Porto, Portugal
Rui Manuel de Medeiros Melo Silva
Invited Professor, PharmD PhD
Virology Service & Molecular Oncology and Viral Pathology Group (CI-IPOP),
IPO Porto FG EPE, Porto, Portugal
Department of Molecular Pathology and Immunology,
ICBAS - UP, Porto, Portugal


AKNOWLEDGMENTS


AKNOWLEDGMENTS
Polymorphisms in DNA regulation-associated genes and GC development |iii
Ao Professor Doutor Hugo Sousa, meu orientador, muito obrigado pelo apoio
e por tudo que me ensinou. Praticamente tudo o que aprendi nesta etapa de
aspirante a investigador foi graças ao Dr. Hugo. Se as coisas não estão a
funcionar então somos nós que temos de as resolver, há sempre solução
para um problema.
À Joana Ribeiro, minha co-orientadora, pela disponibilidade para ajudar em
tudo aquilo que eu precisei. Obrigado por partilhares a tua expêriencia no
laboratório comigo, aprendi imenso contigo.
Ao Professor Doutor Rui Medeiros, meu co-orientador, pelos conselhos e
sabedoria que me transmitiu. É sempre bom poder contar com a ajuda de
alguém com a experiência do Dr. Rui nos momentos em que as coisas não
correm como esperado.
Ao Professor Doutor Henrique Almeida, coordenador do Mestrado, pela
disponibilidade para resolver todos os problemas que foram surgindo ao
longo deste ano e pela oportunidade que me deu para poder alcançar um
dos objetivos da minha vida, realizar este mestrado que tanto me ensinou
nestes últimos dois anos.
Aos co-autores dos projetos de investigação: Dra. Áurea Lima e Mónica
Silva, pela ajuda crucial na realização de parte desta dissertação. Áurea,
obrigado por tudo o que disponibilizaste para que fosse possível realizar esta
dissertação. Agradeço imenso o apoio que me deste na fase mais
importante e que mais precisava. Mónica foste uma excelente colega de
laboratório, parte deste trabalho foi realizado graças à tua ajuda. Muito
obrigado pela disponibilidade e pela companhia. O trabalho é sempre mais
agradável quando temos alguém para conversar.
Ao Grupo de Oncologia Molecular, pela disponibilidade de todos para ajudar
quando foi necessário e pelo bom “ambiente científico” que proporcionam.
É mais fácil “fazer” ciência quando se trabalha num grupo com o vosso.

AKNOWLEDGMENTS
iv| Polymorphisms in DNA regulation-associated genes and GC development
Aos meus amigos, por todo apoio que me deram. O trabalho de investigação
não é fácil e é sempre bom poder contar com os nossos amigos para
desanuviar e melhorar um dia que não estava a correr tão bem. Um obrigado
especial Fernandes e Gonçalo, vocês sabem como me fazer sentir bem.
Muito obrigado pelo apoio meus meninos.
E por último, mas sempre em primeiro, à minha família e à minha namorada,
o meu maior apoio e a minha maior fonte de motivação. Paizinhos muito
obrigado por tudo o que fazem por mim. Eu sei o esforço que fizeram para
que eu pudesse chegar a este momento. Tudo se torna mais fácil com o
vosso apoio incondicional. Dedico-vos este trabalho como pequena forma
de agradecer por tudo.
Mariana, meu amor, tu sabes o quão importante és para mim e que sem ti
eu não encontraria forças para me superar. Obrigado por seres a minha
companheira e por saberes sempre o que é melhor para mim.
You are the love of my life.
Por último, quero agradecer à minha Avó, a minha maior inspiração. Eu sei
que a Avó é a pessoa que fica mais feliz por eu terminar esta etapa da minha
vida. Essa felicidade que me transmite é a minha maior fonte de força de
vontade para fazer mais e melhor. Obrigado pelo carinho, obrigado pelo
apoio, obrigado por tudo Avó.

ABBREVIATIONS LIST


ABBREVIATIONS LIST
Polymorphisms in DNA regulation-associated genes and GC development |vi
A
A – Adenine
Arg – Arginine
C
C – Cytosine
CI – Confidence Interval
CGI – CpG Island
CpG – 5'-C-phosphate-G-3'
D
DNA – Deoxyribonucleic Acid
DNMT – DNA Methyltransferase
DNMT1 – DNA (cytosine-5)-methyltransferase 1
DNMT2 – TRNA (cytosine38-C5)-methyltransferase
DNMT3 – DNA (cytosine-5)-methyltransferase 3
DNMT3A – DNA (cytosine-5)-methyltransferase 3 Alpha
DNMT3B – DNA (cytosine-5-)-methyltransferase 3 Beta
dNTP – deoxyribose Nucleoside Triphosphate
G
G – Guanine
GC – Gastric Cancer
GLOBOCAN – International Agency for Research on Cancer’s online database
GWAS – Genome-Wide Association Studies
H
His – Histidine
HP – Helicobacter pylori
HRM – High Resolution Melting
HWE – Hardy–Weinberg Equilibrium
I
IARC – International Agency for Research on Cancer

ABBREVIATIONS LIST
vii| Polymorphisms in DNA regulation-associated genes and GC development
M
MALDI-TOF – Matrix Assisted Laser Desorption/Ionization - Time of Flight
miRNA – micro Ribonucleic Acid
MGMT – O6-methylguanine-DNA Methyltransferase
MTHF – Methylenetetrahydrofolate
MTHFR – Methylenetetrahydrofolate Reductase
O
OR – Odds Ratio
P
PCR – Polymerase Chain Reaction
R
Real-Time PCR – Real-Time Polymerase Chain Reaction
RFC1 – Reduced Folate Carrier 1
RFLP – Restriction Fragment Length Polymorphism
RNA – Ribonucleic Acid
S
SAM – S-adenosylmethionine
SLC19A1 – Solute Carrier family 19, Member 1
SNP – Single Nucleotide Polymorphism
SPSS – Statistical Package for Social Sciences
T
T – Thymine
tRNA – transfer Ribonucleic Acid
Tyr – Tyrosine

OUTLINE OF THE THESIS


OUTLINE OF THE THESIS
Polymorphisms in DNA regulation-associated genes and GC development |ix
OUTLINE OF THE THESIS
In Chapter 1, the rational for this study will be presented focusing on explain the potential
impact of Single Nucleotide Polymorphisms (SNPs), in genes involved in DNA regulation,
on Gastric Cancer susceptibility.
In Chapter 2, it will be presented a systematic review and meta-analysis regarding the
association of SNPs of DNA methyltransferases (DNMTs) with gastric cancer. This
chapter is presented as the following article:
Neves M, Ribeiro J, Medeiros R, Sousa H. Genetic polymorphism in DNMTs and
gastric cancer: a systematic review and meta-analysis. (prepared to submit to
Porto Biomedical Journal)
In Chapter 3, it will be characterized the association of DNMT2 and DNMT3A SNPs with
the development of gastric lesions and gastric adenocarcinoma in a population from the
northern region of Portugal. This chapter is presented as the following article:
Neves M, Ribeiro J, Dinis-Ribeiro M, Medeiros R, Sousa H. Single Nucleotide
polymorphisms in DNA methyltransferases and gastric cancer. (prepared to
submit to Acta Medica)
In Chapter 4, it will be shown the association of SNPs in SLC19A1 and MTHFR genes
with the susceptibility to GC. This chapter is presented as the following article:
Neves M, Francisco M, Ribeiro J, Lima A, Dinis-Ribeiro M, Medeiros R, Sousa H.
SLC19A1 G80A and MTHFR C677T as risk factors for gastric cancer
development. (prepared to submit to Tumor Biology)
In Chapter 5, it will be summarized the importance of the results obtained in this thesis.
Up to our knowledge, this is the first study to evaluate the association of SNPs in
SLC19A1, MTHFR and DNMTs with GC, in the Portuguese population.
Finally, in Chapter 6, it will be presented the references of the thesis.


INDEX


INDEX
Polymorphisms in DNA regulation-associated genes and GC development |xi
INDEX OF CONTENTS
AKNOWLEDGMENTS .................................................................................................. ii
ABBREVIATIONS LIST ................................................................................................ v
OUTLINE OF THE THESIS ........................................................................................ viii
INDEX ......................................................................................................................... xi
ABSTRACT ................................................................................................................ xiii
RESUMO .................................................................................................................... xiv
CHAPTER 1 ................................................................................................................. 1
CHAPTER 2 ............................................................................................................... 10
CHAPTER 3 ............................................................................................................... 29
CHAPTER 4 ............................................................................................................... 40
CHAPTER 5 ............................................................................................................... 55
CHAPTER 6 ............................................................................................................... 62
ATTACHMENTS ......................................................................................................... 66


ABSTRACT


ABSTRACT
Polymorphisms in DNA regulation-associated genes and GC development |xiii
Gastric cancer (GC) constitutes a serious public health problem, representing the fifth
most common diagnosed cancer with approximately 3 000 new diagnoses and 2 200
deaths per year, in Portugal. GC carcinogenesis is associated with several risk factors,
such as, environmental factors (Helicobacter pylori, diet, obesity, smoking), epigenetic
alterations and genetic susceptibility. Regarding epigenetic alterations, studies have
shown that aberrant DNA methylation have a role in GC carcinogenesis. Therefore,
genetic alterations in genes responsible for DNA methylation (DNA methyltransferases)
may have an impact on GC development. Other risk factor that has been associated with
GC is low folate intake. Folates are important for DNA regulation, thereby alterations in
genes important for folate metabolism might affect individual’s susceptibility for GC.
Genetic polymorphisms, such as single nucleotide polymorphisms (SNPs), have been
studied and associated with altered protein functions and, consequently, with the
development of cancer. Several studies suggest that SNPs in genes important for DNA
methylation might have an influence on the susceptibility for GC.
We developed a study to analyze the role of some SNPs in GC development. The study
was performed with a population from the northern region of Portugal with gastric lesions
(n=43), GC (n=137) and a cancer-free control group (n=250). SNPs were selected based
on literature and genetic characterization was performed with different methodologies.
Firstly, we performed a systematic review and meta-analysis to evaluate which SNPs in
DNA methyltransferases (DNMTs) may be associated to GC, according to literature. The
results showed that DNMT2 rs11254413, DNMT3A rs7560488, DNMT3A rs36012910
and, specially, DNMT1 rs16999593 seem to be good candidates to study the role of
SNPs in DNMTs in GC development. Then, we aimed to characterize DNMT2
rs11254413 and DNMT3A rs7560488 genotypes and their association with GC, in our
population. Our results showed that there is no association between both SNPs and GC
development. Finally, another aim of our study was to analyze the association between
SLC19A1 G80A and MTHFR C677T with GC development. Regarding the results
obtained for SLC19A1 G80A genotypes, we observed a protective effect for gastric
lesion (OR=0.373; 95%CI=0.189-0.736; p=0.006), but not with GC. On the other hand,
MTHFR C677T was associated with an increased risk for GC development (OR=2.046;
CI 95%=1.323-3.165; p=0.001), but it was not associated with gastric lesions.

ABSTRACT
Polymorphisms in DNA regulation-associated genes and GC development |xiv
This study revealed that DNMT2 rs11254413 and DNMT3A rs7560488 polymorphisms
are not associated with GC in our population, nevertheless SLC19A1 G80A may have a
protective role in gastric lesions development and MTHFR C677T may be associated
with an increased risk for GC development.

RESUMO


RESUMO
Polymorphisms in DNA regulation-associated genes and GC development |xv
O cancro gástrico (CG) constitui um grave problema de saúde pública, representando o
5º cancro mais diagnosticado com cerca de 3 000 novos diagnósticos e 2 200 mortes
por ano, em Portugal. A carcinogénese do CG está associada a vários fatores de risco,
tais como, fatores ambientais (Helicobacter pylori, dieta, obesidade, tabaco), alterações
epigenéticas e suscetibilidade genética. Em relação às alterações epigenéticas, estudos
têm mostrado que a metilação anormal do DNA tem um papel na carcinogénese do CG.
Portanto, alterações genéticas em genes responsáveis pela metilação do DNA (DNA
metiltransferases) podem ter um impacto no desenvolvimento do CG. Outro fator de
risco que tem sido associado ao CG é a baixa ingestão de folatos. Os folatos são
importantes para a regulação do DNA, pelo que alterações nos genes importantes para
o metabolismo dos folatos podem afetar a suscetibilidade individual para o CG. Os
polimorfismos genéticos, tais como single nucleotide polymorphisms (SNPs), têm sido
estudados e associados a alterações nas funções das proteínas e, consequentemente,
com o desenvolvimento de cancro. Muitos estudos sugerem que SNPs nos genes
responsáveis pela metilação podem ter influência na suscetibilidade para o CG.
Nós desenvolvemos um estudo para analisar o papel de alguns SNPs no
desenvolvimento de CG. O estudo foi realizado com uma população da região Norte de
Portugal com lesões gástricas (n=43), CG (n=137) e um grupo sem cancro (n=250). Os
SNPs foram selecionados com base na literatura e a caracterização genética foi feita
através de diferentes metodologias. Primeiramente, nós realizamos uma revisão
sistemática e uma meta-análise para avaliar que SNPs em DNA metiltransferases
(DNMTs) podem ser associados ao CG, de acordo com a literatura. Os resultados
mostraram que os DNMT2 rs11254413, DNMT3A rs7560488, DNMT3A rs36012910 e,
especialmente, o DNMT1 rs16999593 parecem ser bons candidatos para estudar o
papel de SNPs em DNMTs no desenvolvimento de CG. De seguida, o nosso objetivo foi
caracterizar os genótipos para DNMT2 rs11254413 e DNMT3A rs7560488 na nossa
população. Os nossos resultados mostraram que não existe associação entre os dois
SNPs e o desenvolvimento de CG. Por fim, outro objetivo do nosso estudo era analisar
a associação entre SLC19A1 G80A e MTHFR C677T com o desenvolvimento de CG.
De acordo com os resultados obtidos para os genótipos do SLC19A1 G80A,
observámos um efeito protetor para a lesões gástricas (OR = 0.373; 95%CI = 0.189-
0.736; p = 0.006), mas não para o CG. Por outro lado, MTHFR C677T foi associado a
um risco aumentado de desenvolvimento de CG (OR = 2.046; CI 95% = 1.323-3.165; p
= 0.001), mas não associado a lesões gástricas.

RESUMO
Polymorphisms in DNA regulation-associated genes and GC development |xvi
Este estudo revelou que os polimorfismos DNMT2 rs11254413 e DNMT3A rs7560488
não estão associados com CG na nossa população, porém o SLC19A1 G80A pode ter
um papel protetor no desenvolvimento de lesões gástricas e o MTHFR C677T pode
estar associado a um risco aumentado de desenvolvimento de CG.

CHAPTER 1


CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |2
I. CANCER
Cancer could be defined as a disease where cells, with “damaged” DNA, grow without
control and gain the potential to invade other tissues and spread to other parts of the
body. The emergence of conditions for the development of cancer is dependent on many
factors, such as genetic, epigenetic and environmental factors, which shows the
complexity of this disease (1).
According to the International Agency for Research on Cancer (IARC), cancer is one of
the leading causes of mortality worldwide, with 8.2 million deaths in 2012 (13% of all
deaths) (2,3). In the same year, 14 million new cases of cancer were reported and it is
expected a 70% increase of new cases of cancer in the next 20 years (3). The top 5
cancers worldwide, and in Portugal, are Breast, Prostate, Lung, Colorectal and Stomach,
being breast cancer the more incident and lung cancer the one with higher mortality –
figure 1.
Figure 1. Incidence and mortality of top 5 cancer sites (both sexes) in 2012. (Adapted from
GLOBOCAN 2012 (4))

CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |3
II. GASTRIC CANCER
i. Epidemiology
Gastric Cancer (GC) is the third leading cause of death by cancer, worldwide (4). It
represents 6.8% of all incident cancer cases and 8.8% of all cancer deaths. The highest
mortality rates are in eastern Asia (China is the country with higher mortality rates) and
the lowers in north America. Central and eastern Europe and central and south America
have also high mortality rates (4). In Portugal, GC represents 9.5% of all cancer deaths
(2265 deaths) and 6.1% of all incident cases (3018 new cases) (4).
The most frequent histological types of GC are adenocarcinomas and, according to
Lauren classification, are divided in two sub-types: diffuse (poorly differentiated) and
intestinal (well/moderately differentiated). Additionally, tumors that exhibit both intestinal
and diffuse characteristics, and undifferentiated tumors are classified as indeterminate
(5). The most sensitive and specific diagnostic test for GC is endoscopic screening,
nevertheless the majority of GC patients are diagnosed at an advanced stage because
of the lack of early symptoms (6). GC initiation and progression are a result of complex
genetic and environmental interactions. GC carcinogenesis differs according to the
histological type: the intestinal type carcinogenesis is a sequential step process,
characterized by a start with a premalignant gastric change, such as atrophic gastritis,
followed by intestinal metaplasia, dysplasia and, finally, gastric carcinoma; and the
diffuse type carcinogenesis is more often associated with some genetic changes and
there is not a consistent sequence of events, such as in the intestinal type. The diffuse
type is associated with different pathological characteristics, such as signet ring cells and
loss of cell cohesion (6–8).
ii. Risk factors
Tumorigenesis of GC is a multistep process and involve multiple factors (9,10).The major
risk factor for the development of gastric cancer (especially the intestinal type) is the
infection by Helicobacter pylori (HP), nevertheless, GC can also be the result of specific
genetic alterations (7). The majority of GC cases arise sporadically and only less than
3% are hereditary cancer syndromes (8). There are several sporadic genetic changes
that influences GC pathogenesis, such as chromosomal instability, microsatellite
instability, epigenetic changes, somatic gene mutations or single nucleotide
polymorphisms (SNPs) (8).

CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |4
The incidence of gastric cancer have been reduced since the identification of HP as a
key factor in carcinogenesis, but other factors associated to our modern lifestyle and
aging of population are also influencing the incidence of GC worldwide (10). The risk for
GC increases with age and incidence rates of GC are twice higher in men comparing to
women (4). Unhealthy habits, such as heavy alcohol drinking (four or more drinks per
day) and smoking increases risk for GC (10). Obesity and diet are two major factors
affecting GC incidence, nowadays. Literature shows that low consumption of fruits and
vegetables, intake of salty and smoked food and obesity are associated with higher risk
for GC (9–14) - figure 2.
Figure 2. Gastric cancer risk factors. Environmental and genetic/epigenetic factors are
associated with gastric carcinogenesis. (9–17).
GASTRIC CANCER RISK
FACTORS
Helicobacter Pylori Epstein-Barr
Virus
Age
Sex
(Male)
Smoking
Alcohol
Obesity
High intake of salty, nitrated and smoked
food
Low consumption of
fruits and vegetables
Hereditary cancer
syndromes
Pre-cancerous lesions
Chromosomal and
microsatellite instability
Somatic mutations
Genetic susceptibility
Epigenetic alterations

CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |5
III. DNA REGULATION AND CANCER
i. Folate intake – Folates and DNA regulation
Folates are found naturally in a wide variety of foods, specially vegetables like spinach,
kale, brussels sprouts and broccoli. Folates are water soluble B-vitamins essential for
intracellular transmethylation reactions including those involved in DNA methylation and
DNA synthesis (17). The active form of folate for cells is reduced folate 5-
methyltetrahydrofolate (5-MTHF). Reduced folates are transported into the cell by
Reduced Folate Carrier 1 (RFC1 or SLC19A1), or metabolized by Methylene
tetrahydrofolate reductase (MTHFR) – Figure 3 (18). MTHFR has a key role by
converting dietary folates and folic acid (artificially form of folate) to 5-MTHF (18,19). The
5-MTHF works as a single carbon donor used to convert homocysteine in to methionine,
by removing a methyl group from 5-MTHF, using vitamin B12 as coenzyme. Methionine
serves as a methyl group donor through conversion to S-Adenosyl methionine (SAM), a
key agent for DNA methylation (18,20).
Figure 3. Folate metabolism mechanism. (Adapted from Ulrich 2003 (21))

CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |6
Multiple studies have demonstrated that alterations in folate metabolism, such as folate
deficiency, low levels of folate in serum or genetic factors affecting genes in folate
metabolism are associated with cancer, including GC (22–25). Alterations in RFC1
activity affects the amount of reduced folates, leading to a lower concentration in the
cells (23). MTHFR alterations and consequent reduced enzymatic activity leads to higher
levels of homocysteine and the cycle homocysteine – methionine – SAM is disrupted,
affecting DNA methylation and leading in to changes in the epigenetic profile (24).
ii. Epigenetics and control of gene expression
Epigenetics is the study of heritable and transient/reversible changes in gene expression
that are not accompanied by a change in the DNA sequence (1). The most extensively
studied epigenetic modification is DNA methylation, which is characterized by an addition
of a methyl group in to the fifth carbon position of a cytosine residue in a CpG dinucleotide
(26). Clusters of CpG dinucleotides in GC rich regions of the genome called CpG islands
(CGI) frequently occur in the 5′-flanking promoter areas of genes, which are often
responsible for the control of protein translation (15). Epigenetic control of gene
expression by cytosine methylation is facilitated by the activity of DNA
methyltransferases (DNMTs). These enzymes recognize CpGs and catalyze the
transmethylation of cytosine by transferring methyl groups from S-adenosylmethionine
(SAM) (26,27).
DNA methylation was the first epigenetic mark shown to be critically involved in cancer
development. Alterations in DNA methylation are associated with silencing of tumor
suppressor genes and the activation of oncogenes involved in cancer (17,1). Literature
have reported that global hypomethylation has been associated to early stages of
carcinogenesis, causing genome-wide allelic instability (28). Paradoxically, the genomes
of cancer cells are also characterized by promoter genes hypermethylation, typically in
the CpG island of tumor suppressor genes and microRNA (miRNA). In fact, the
inactivation of tumor suppressor genes through the hypermethylation of CpG islands
within promoter methylation has been considered an important hallmark of cancer
(17,26,29,30).

CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |7
IV. GENETIC SUSCEPTIBILITY AND CANCER
The human genome sequence of two individuals is only 99% similar, and 90% of the
variations in the genome are Single Nucleotide Polymorphisms (31). SNPs are
differences in single bases of DNA that turned out to be stable through evolution and
where the minor allelic frequency is over 1% in the population. These variants are
present in almost all population but some are specific from one region or population (32).
Several studies have reported the association of SNPs with differential function of
proteins and therefore contributing for the development of several diseases, including
cancer (31,33,34).
A project called Genome-Wide Association Studies (GWAS) have emerged as a
powerful approach to identify risk alleles for common diseases (32,35). These studies
compare two large groups of individuals, one healthy control group and one case group
affected by a disease, including cancer (35). The enormous variety of data produced by
GWAS and other studies regarding potential genomic markers in cancer contributes to
population screening for genetic susceptibility to cancer. All this information may help to
select groups of individuals who are at increased risk of cancer development and provide
to them a preventive strategy (36).
Figure 4. Preventive strategy on increased risk groups may help to reduce mortality rates by
reducing the risk of cancer development or by proving better prognosis and personalized therapy
for the populations. (Source: National Cancer Institute, USA)

CHAPTER 1
Polymorphisms in DNA regulation-associated genes and GC development |8
V. AIM OF THE DISSERTATION
The main aim of the dissertation was to evaluate different SNPs on genes that are
important for cellular methylation and DNA regulation and study an association with
gastric cancer. SNPs in genes of folate metabolism and DNMTs have already been
described in GC, nevertheless these were not studied in our population, and there are
only few studies in eastern/Caucasian populations. By evaluating SNPs in gastric cancer
patients and comparing with healthy controls, we expected to find associations between
SNPs and gastric cancer risk.
Throughout the dissertation we tried to answer specific aims through different studies:
1) Systematic review and meta-analysis to understand witch SNPs in DNMTs may
be associated to GC;
2) Case-control study to understand the association of some SNPs in DNMTs with
GC in our population;
3) Case-control study to understand the association of SNPs in SLC19A1 and
MTHFR with GC in our population.


CHAPTER 2


CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |10
TITLE:
GENETIC POLYMORPHISM IN DNMTs AND GASTRIC CANCER: A SYSTEMATIC
REVIEW AND META-ANALYSIS
AUTHORS
Neves M1,2, Ribeiro J1,2,3, Medeiros R1,2,3,4,5, Sousa H1,2,5
Affiliations
1 Molecular Oncology and Viral Pathology Group, Research Centre (CI-IPOP),
Portuguese Oncology Institute of Porto (IPO-Porto), Rua Dr. António Bernardino de
Almeida, 4200-072 Porto, PORTUGAL
2 Faculty of Medicine of University of Porto, Alameda Prof. Hernâni Monteiro, 4200-319
Porto, PORTUGAL
3 Research Department, Portuguese League Against Cancer (LPCC-NRNorte), Estrada
Interior da Circunvalação 6657, 4200 Porto, PORTUGAL
4 Abel Salazar Institute for the Biomedical Sciences (ICBAS), University of Porto, Rua de
Jorge Viterbo Ferreira n.º 228, 4050-313 Porto, PORTUGAL.
5 Virology Service, Portuguese Oncology Institute of Porto (IPO-Porto), Rua Dr. António
Bernardino de Almeida, 4200-072 Porto, PORTUGAL
Correspondence to:
Hugo Sousa, MD PhD
Serviço de Virologia, Laboratórios 4º Piso
Instituto Português de Oncologia do Porto FG EPE
Rua Dr. António Bernardino Almeida
4200-072 Porto, Portugal
Phone: +351 22 508 4000 (ext 5410)
Fax: +351 22 508 4001
E-mail: [email protected] or [email protected]

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |11
ABSTRACT
Epigenetics alterations, such as aberrant DNA methylation, have been associated with
gastric carcinogenesis. Single Nucleotide Polymorphisms (SNPs) in DNA
methyltransferases (DNMTs) may influence protein expression and therefore affect DNA
regulation and susceptibility for Gastric Cancer (GC).
We performed a systematic review and meta-analysis involving 11 studies and a total of
24 SNPs in DNMTs were analysed. According to literature, only 4 SNPs, DNMT1
rs16999593, DNMT2 rs11254413 and DNMT3A rs7560488 and DNMT3A rs36012910,
were associated with GC. DNMT1 rs16999593 and DNMT3A rs7560488 C allele and
DNMT3A rs36012910 G allele showed an increased risk for GC. On the other hand,
DNMT2 rs11254413 G allele presented a protective effect for GC. Additionally, the meta-
analysis evaluated the SNPs analysed in more than one study (n=6). Results revealed
that only DNMT1 rs16999593 had a statistically significant association with GC
development (OR = 1.31; 95% CI = 1.08-1.60; p = 0.006 for TC+CC genotypes).
Our study suggests that DNMT2 rs11254413, DNMT3A rs7560488, DNMT3A
rs36012910 and, specially, rs16999593 may have an association with GC development
and more studies with different populations are needed to verify if this SNPs are related
with GC.
KEY WORDS
DNA methylation; DNA Methyltransferase; Single Nucleotide Polymorphism; Gastric
Cancer

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |12
INTRODUCTION
Epigenetics, and more extensively DNA methylation, has been studied in the past
decades and associated with several diseases, including cancer. Literature shows that
altered epigenetic control of gene expression has an important role in carcinogenesis
(Ziogas and Roukos, 2009). DNA methylation is an important epigenetic mark in
tumorigenesis, in which a methyl group is added to the 5’ position of cytosine residue in
a CpG dinucleotide (CpG islands (CGI)), in the 5′-flanking promoter’s genes (Feinberg
and Vogelstein, 1983). Aberrant methylation of promoter’s genes is an important
hallmark of cancer cells and, in Gastric Cancer (GC), a large number of genes involved
in carcinogenesis and clinical outcome accumulate aberrant methylation. Promoter
methylation is an important mechanism of inactivation of tumor suppressor genes, in
cancer cells (Feinberg and Vogelstein, 1983).
The process of promoter methylation is catalysed by DNA Methyltransferases (DNMTs)
which organize, regulate and maintain properly mammalian genomes (Yang et al., 2012).
There are three types of DNMTs: DNMT1, over-expressed in human cancers including
GC, that catalyse post-replication DNA methylation and maintain the methylation
patterns during cell divisions (Kanai et al., 2003; Saito et al., 2001; Yang et al., 2011);
DNMT2 which is the most conserved and has a role in both DNA and RNA methylation
(Schaefer and Lyko, 2010); and DNMT3, which is divided in two types (DNMT3a and
DNMT3b) that are responsible for de novo methylation during gametogenesis and
embryogenesis, and that also seem to be over-expressed in human cancers, including
GC (Yang et al., 2011, 2012). It is well known that gene expression can be influenced by
single nucleotide polymorphisms (SNPs) and, in DNMTs genes, they may have
functional consequences affecting methylation and therefore individual’s susceptibility to
cancer development.
Several studies have recently explored the potential association of SNPs in DNMTs with
the susceptibility to GC, nevertheless, the observed associations of these studies are
inconsistent and some studies are not large enough to take conclusions on the effect of
DNMTs SNPs on GC. As a result, we have performed a meta-analysis of all eligible
studies to understand the possible association of DNMTs SNPs with GC.

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |13
MATERIALS AND METHODS
Search strategy
The present systematic review and meta-analysis follows PRISMA guidelines for
Systematic Review and Meta-analyses. Pubmed was searched till the end of March 2016
and literature searching was performed by applying the following query: "dna
modification methylases"[MeSH Terms] OR ("dna"[All Fields] AND "modification"[All
Fields] AND "methylases"[All Fields]) OR "dna modification methylases"[All Fields] OR
("dna"[All Fields] AND "methyltransferase"[All Fields]) OR "dna methyltransferase"[All
Fields] OR DNMT[All Fields]) AND ("polymorphism, genetic"[MeSH Terms] OR
("polymorphism"[All Fields] AND "genetic"[All Fields]) OR "genetic polymorphism"[All
Fields] OR "polymorphism"[All Fields]) AND ("stomach neoplasms"[MeSH Terms] OR
("stomach"[All Fields] AND "neoplasms"[All Fields]) OR "stomach neoplasms"[All Fields]
OR ("gastric"[All Fields] AND "cancer"[All Fields]) OR "gastric cancer"[All Fields].
Inclusion and exclusion criteria
The literature search was limited to original studies performed in humans and no
publication year restriction was applied. Only case-control studies with histologically-
confirmed adenocarcinoma cases were included in this analysis. All DNMTs
polymorphisms were selected for this analysis. Regarding the language of the papers,
only studies written in English, Portuguese or Spanish were selected. Reviews, Meta-
analysis, Systematic Reviews and studies not related to gastric cancer were excluded
for this analysis.
Data extraction
Two authors performed the data extraction and all disagreements were resolved with the
opinion of a third author. For each study, the following items were collected: country, type
of study, ethnicity, matching criteria, age (years), polymorphisms in study, genotype
method, number of cases and controls. Data concerning genotype distribution and the
relative risk were also extracted for all polymorphisms included in the studies.
Statistical analysis
The evidence of Hardy Weinberg Equilibrium (HWE) in controls was recalculated in the
present meta-analysis through the application of the online software
(http://www.had2know.com/academics/hardy-weinberg-equilibrium-calculator-2-
alleles.html). A p–value less than 0.05 of HWE was considered significant.

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |14
Meta-analysis for DNMTs SNPs was conducted by using the software Review Manager,
version 5.3.5 (Copenhagen: The Nordic Cochrane Centre, The Cochrane Collaboration,
2014). The calculated OR (95% CI) was performed with Mantel-Haenszel statistical
method and Random effects analysis model. A p–value less than 0.05 was considered
significant.
RESULTS
Literature search
A total of 35 papers were evaluated from which only 11 matched with inclusion criteria.
Seventeen articles were excluded due to the exclusion criteria by screening the titles and
abstracts. After a comprehensive evaluation of the remaining 18 articles, 7 were
excluded: 4 articles didn’t evaluate polymorphisms of DNMTs, only O6-methylguanine-
DNA methyltransferase (MGMT) (Chikan et al., 2015; Imai et al., 1995; Lin et al., 2014;
Xiong et al., 2013); 2 articles only reported mutations on DNMTs (Kanai et al., 2003; Oh
et al., 2014); 1 study was about allelic loss in gastric cancer (Kim et al., 2013). Finally,
11 studies involving a total of 3049 cases and 5185 controls were included in this
analysis – figure 1.
Study characteristics
Table 1 describes the principal baseline characteristics of included studies. The majority
of the studies were performed in China (Cao et al., 2013; Fan et al., 2010; Hu et al.,
2010; Jiang et al., 2012; Wang et al., 2015; Y. Wang et al., 2005; Wu et al., 2014, 2012;
Yang et al., 2012), one study in Iran (Khatami et al., 2009) and one in Japan (Aung et
al., 2005). Almost all studies have selected the controls and cases using age and gender
as matching criteria, but some studies also use match with Helicobacter Pylori infection
and Ethnicity. The number of cases and controls varies between the studies. A total of
24 polymorphisms were evaluated, 10 of DNMT1, 1 of DNMT2, 6 of DNMT3A and 7 of
DNMT3B. The majority of studies were performed using Polymerase Chain Reaction –
Restriction Fragment Length Polymorphism (PCR-RFLP) methodology, but other
methods of genotyping were used like TaqMan assay, High Resolution Melting (HRM)
and Matrix Assisted Laser Desorption/Ionization - Time of Flight (MALDI-TOF).
Supplementary table 1 shows the characteristics of all polymorphisms included in the
studies. The re-calculated HWE for controls is also shown in supplementary table 2.

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |15
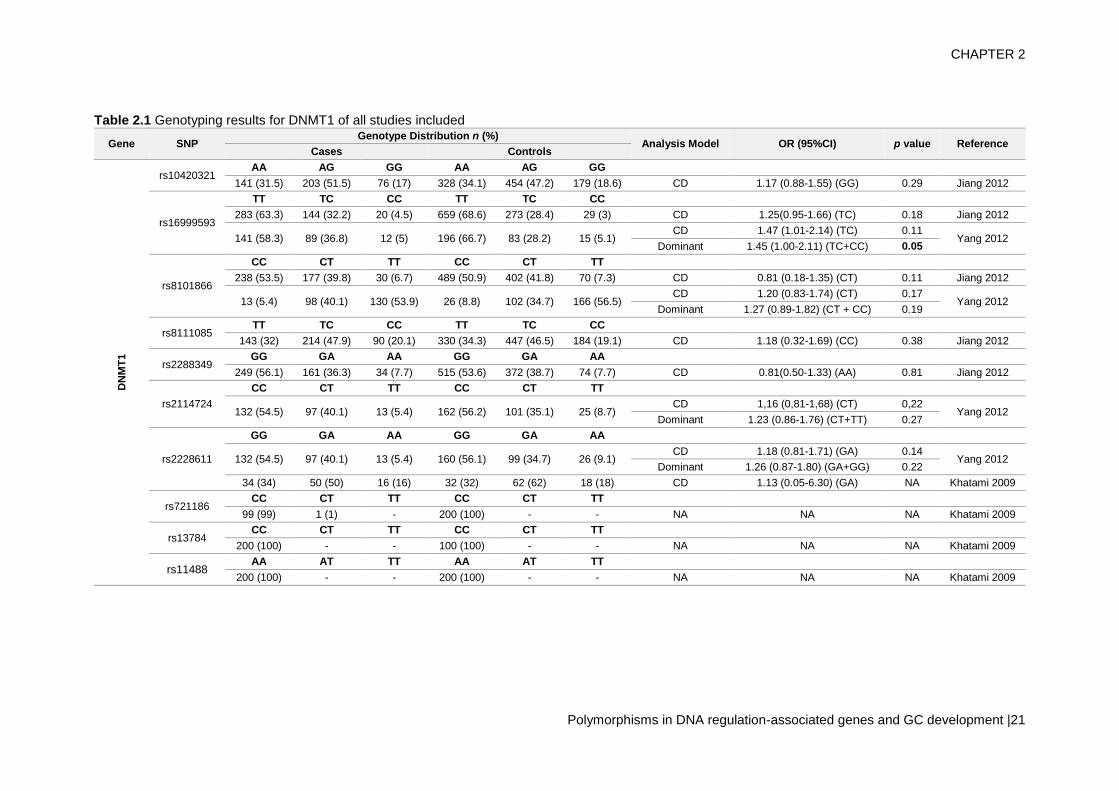
Genotyping results
The genotyping results of all studies are described in table 2. In this study, we have
included data from 24 SNPs in DNMTs from the 11 studies.
A total of 10 SNPs in DNMT1 were studied by 4 different studies (Jiang et al., 2012;
Khatami et al., 2009; Yang et al., 2012) – table 2.1. Of these SNPs, only rs16999593,
rs8101866 and rs2228611 have data from more than one population. Regarding the risk
analysis, there was only one study that reported a significant risk association of one SNP
(rs16999593) with increased risk of GC development (OR=1.45; 95%CI = 1.00–2.11;
p = 0.05, for C allele). All other SNPs do not show statistical significance.
Only one study evaluated SNPs in DNMT2 (Yang et al., 2012) revealing an association
between rs112254413 SNP and protection for the development of GC, especially
associated with A allele (OR = 0.15; 95%CI = 0.08-0.27; p <0.01) – table 2.2.
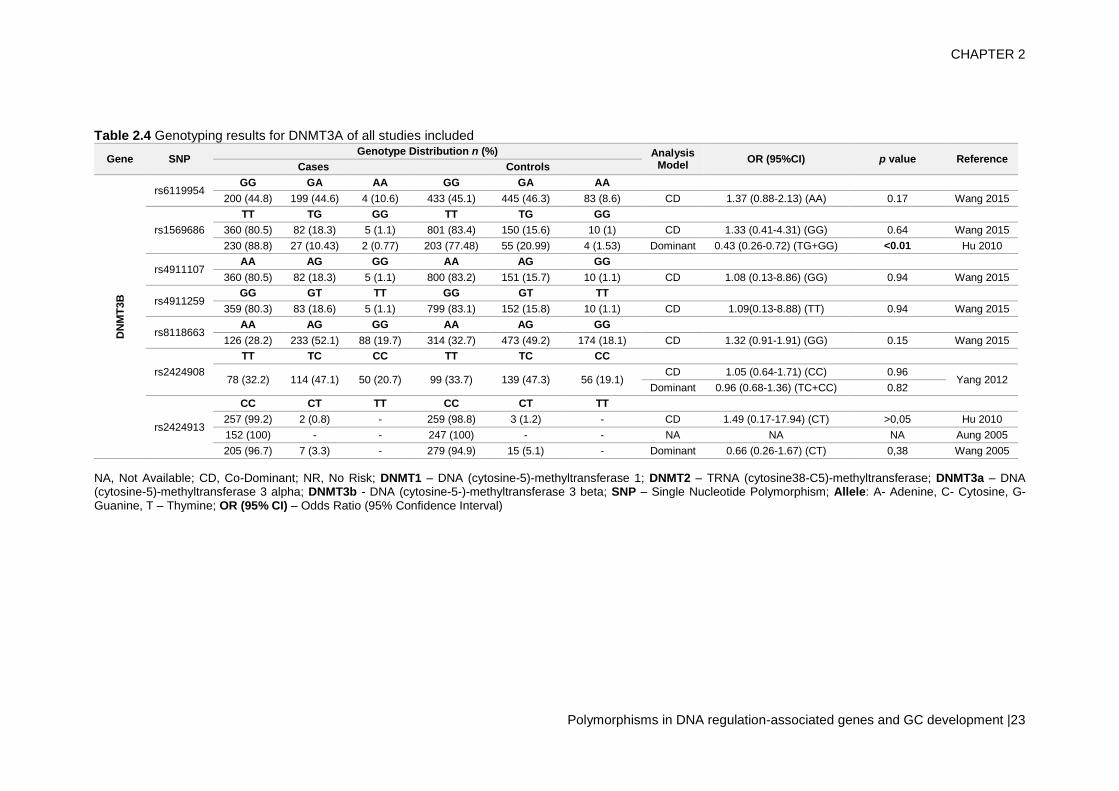
Thirteen SNPs of DNMT3, 6 of DNMT3A and 7 of DNMT3B, were evaluated by 9 different
studies, with 2 SNPs of each gene with more than one study (Aung et al., 2005; Cao et
al., 2013; Fan et al., 2010; Hu et al., 2010; Wang et al., 2015, 2005, Wu et al., 2014,
2012; Yang et al., 2012). Regarding the risk analysis, DNMT3A rs7560488 (OR = 1.65;
95%CI = 1.19–2.29; p = 0.002, for C allele) and rs36012910 (OR = 2.36; 95%CI = 1.35–
4.14; p = 0.002, for allele G) seem to be associated with an increased risk for GC (Wu
et al., 2014, 2012), and rs1550117 and rs13420867 show contradictory findings between
the reported studies. In DNMT3B, only rs1569686 showed a statistically significant
association (p<0.01) with GC, nevertheless the results are contradictory among different
studies – tables 2.3/2.4.
Meta-analysis results
A meta-analysis was performed with data from all SNPs evaluated in more than one
study - Figures 2-4. A total of 6 SNPs were analysed: DNMT1 rs16999593, rs8101866
and rs2228611; DNMT3A rs1550117 and rs13420827; and DNMT3B rs1569686. For the
analysis, we have used the dominant genetic model, considering the risk genotype as
the presence of any copy of the less frequent allele. The majority of studies showed no
significant heterogeneity in the data reported (p>0.05).
DNMT1: The analysis revealed that there was no heterogeneity (p>0.05) amongst the
three SNPs tested. We found that only rs16999593 had a statistically significant
association (p = 0.006), revealing a 31% increased risk of GC development for TC+CC

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |16
genotypes (OR = 1.31; 95%CI = 1.08-1.60). The rs8101866 and rs2228011 showed no
significant differences (p>0.05) and moreover no significant impact on the risk (OR =
1.08 and OR = 0.93, respectively) – figure 2.
DNMT3A: Data showed that in addition to differences in the heterogeneity of rs1550117
(p = 0.03), there is no impact on the risk association of GC development for either
rs1550117 and rs13420827 (p = 0.20 and p = 0.19, respectively) – figure 3.
DNMT3B: One SNP (rs2424913) was excluded from our meta-analysis because it was
not possible to estimate the OR value. The lack of data for all genotypes in the three
studies makes it impossible to evaluate any association (Aung et al., 2005; Hu et al.,
2010; Wang et al., 2005). The analysis revealed that the data from studies regarding
rs1569686 are very contradictory, which result in a high heterogeneity on studies
(p<0.001) and no risk association with GC – figure 4.
We have also tested all data considering the Recessive Model for each SNP, and no
significant change was observed in the reported data (data not shown).
DISCUSSION
Gastric carcinogenesis is a multistep process, where different factors are involved and
epigenetic alterations, including DNA methylation of CGI, seem to play an important role
in early mechanisms of carcinogenesis (Yang et al., 2011). The process of CpG
methylation is catalysed by DNMTs and modified DNA methylation has direct effect on
the regulation of gene expression, thus altered activity of DNMTs can affect DNA
methylation. Literature reveals that DNMTs are frequently overexpressed in GC (Lin et
al., 2014). Considering the significant impact of genetic susceptibility based on the
genetic sequences of individuals, functional SNPs may affect DNMTs expression, and
therefore, it is important to study SNPs in DNMTs and understand the effect that
functional SNPs can have on their activity.
Recently, several studies have been performed to evaluate the impact of DNMTs
polymorphisms in GC susceptibility (Aung et al., 2005; Cao et al., 2013; Fan et al., 2010;
Hu et al., 2010; Jiang et al., 2012; Khatami et al., 2009; Wang et al., 2015, 2005, Wu et
al., 2014, 2012; Yang et al., 2012). Hence, this systematic review intends to resume the
data published regarding the association of SNPs in DNMTs with GC. By searching
literature, we have found 11 studies matching our inclusion criteria. One of the first

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |17
findings was that the majority of included studies were performed in Asiatic populations,
where GC is extremely common (Karimi et al., 2014). This constitutes a major limitation
for this systematic review since that is not possible to extrapolate the conclusions to
others populations.
There are very few studies considering DNMTs SNPs and GC susceptibility and very few
data on different SNPs. A total of 24 SNPs were found in this systematic review, 10 in
DNMT1, 1 in DNMT2 and 13 in DNMT3. Possible limitations of some of the included
studies are not only population bias but also the different genotyping methods. The
genotyping methods used were different and with different specificity: PCR-RFLP
method has some limitations to genotype correctly the three genotypes, specially
heterozygote genotypes (Davis et al., 2007); while TaqMan real-time PCR assays are
more accurate and have less limitations and are a better candidate for SNP genotyping
(Shen et al., 2009). Furthermore, the number of cases and controls included are
extremely different among studies and therefore more studies with bigger populations
and more precise genotyping methods are still required.
The literature report that DNMT1 rs16999593, DNMT2 rs11254413 and DNMT3A
rs7560488 and rs36012910 were the only associated with GC within published studies
(Wu et al., 2014, 2012; Yang et al., 2012). The DNMT1 rs16999593 is characterized by
a C>T variation, resulting in an Arg to His amino acid substitution at position 97 of the
protein, leading to missense mutation that may affect the structure and function of
DNMT1. Literature revealed that rs16999593 is significantly associated to some
malignancies: Tao et. al showed that individuals with TC genotype presented a 4-fold
increased risk for sporadic triple-negative breast carcinoma (Tao et al., 2015); Xiang et.
al also revealed that rs16999593 can provide protective effect for ductal breast
carcinoma (Xiang et al., 2010); and Yang et. al reported a 1.45-fold increased risk for
TC+CC carriers and gastric cancer (Yang et al., 2012). Although these evidences, other
studies do not show any association (Jiang et al., 2012; Ye et al., 2010). DNMT2
rs11254413 is characterized by a G>A substitution leading to an amino acid change, His
to Tyr at position 101 of the protein that may affect DNMT2 function (Yang et al., 2012).
The A-allele was associated with protection risk for GC by Yang et al, nevertheless, there
is no functional study that support the biological role of this SNP on DNMT2 function
(Yang et al., 2012). The DNMT3A rs7560488 (variation T>C) is a tagSNP that represents
SNPs of the DNMT3A1 promoter with high linkage disequilibrium and has been
associated with higher risk for GC in Asiatic population (Wu et al., 2014). Authors
suggested that T to C change influences the binding of transcriptional factors and

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |18
consequently expression levels of DNMT3a (Wu et al., 2014). The DNMT3A
rs36012910, which is characterized by a A>G substitution located in the promoter region
of DNMT3A and the substitution has been associated with DNMT3A activity (Wu et al.,
2012). Wu et al reveals that AG+GG genotypes were associated with a higher risk for
GC (Wu et al., 2012).
Regarding the data from the meta-analysis, we only were able to compare the
information regarding 6 SNPs, which were described in more than one study. For DNMT1
analysis, 3 SNPs were studied (rs16999593, rs8101866 and rs2228611) and only
rs16999593 showed statistically significant data from the information of two different
studies (Khatami et al., 2009; Yang et al., 2012) – figure 2. The results showed an
increased risk for TC+CC genotypes of GC development. Nevertheless, the low number
of studies and the lack of studies from populations apart from China make it necessary
to perform more studies with different populations to confirm the association of
rs16999593 with GC. Concerning DNMT3A, 2 SNPs were included in meta-analysis
(rs1550117 and rs13420827) – figure 3. It was not found association of both SNPs with
GC in the analysis. The heterogeneity analysis revealed differences only in rs1550117
populations, because the ORs of Fan et al. were different comparing to the other studies
(Fan et al., 2010). By removing Fan et al. of analysis, it became homogeneously but we
did not find a significantly association with GC. The rs1550117 was the only SNP
evaluated in three studies but analysis didn’t show an association with GC. The results
in the studies were different, probably because of different genotype methods used in
the three studies – table 1.
Finally, we included one SNP in DNMT3B on the meta-analysis (rs1569686) – figure 4.
We found no association of rs1569686 with GC. The analysis revealed heterogeneity
because OR values of two studies were too different (0.43 in Hu et al. and 1.21 in Wang
et al.) (Hu et al., 2010; Wang et al., 2015).

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |19
CONCLUSIONS
We found 11 articles studying SNPs in DNMTs but the variety of SNPs and the reduced
number of studies per each SNP made this analysis more complicated allowing us to
draw only some remarks and not strong conclusions. The data from studies revealed
that rs11254413, rs7560488, rs36012910 and, specially, rs16999593 seem to be good
candidates to study the role of SNPs in DNMTs and GC development. Nevertheless, our
meta-analysis revealed that only one SNP (rs16999593) in DNMT1 was statistically
associated with GC development, especially TC+CC carriers. More studies with different
populations are needed to prove if the SNPs selected in our analysis are associated with
GC. Nevertheless, studies regarding biological importance of functional SNPs in DNMTs
activity and expression are also need to fully understand the impact of SNPs in DNMTs
on GC.
REFERENCES
Aung, P.P., Matsumura, S., Kuraoka, K., Kunimitsu, K., Yoshida, K., Matsusaki, K., Nakayama, H., Yasui, W., 2005. No evidence of correlation between the single nucleotide polymorphism of DNMT3B promoter and gastric cancer risk in a Japanese population. Oncol. Rep. 14, 1151–1154.
Cao, X.-Y., Jia, Z.-F., Cao, D.-H., Kong, F., Jin, M.-S., Suo, J., Jiang, J., 2013. DNMT3a rs1550117 polymorphism association with increased risk of Helicobacter pylori infection. Asian Pac. J. Cancer Prev. 14, 5713–5718.
Chikan, N.A., Bukhari, S., Shabir, N., Amin, A., Shafi, S., Qadri, R.A., Patel, T.N.C., 2015. Atomic Insight into the Altered O6-Methylguanine-DNA Methyltransferase Protein Architecture in Gastric Cancer. PLoS One 10, e0127741. doi:10.1371/journal.pone.0127741
Davis, A.H.T., Jianhua Wang, Tsang, T.C., Harris, D.T., 2007. Direct sequencing is more accurate and feasible in detecting single nucleotide polymorphisms than RFLP: using human vascular endothelial growth factor gene as a model. Biol. Res. Nurs. 9, 170–8. doi:10.1177/1099800407308083
Fan, H., Liu, D., Qiu, X., Qiao, F., Wu, Q., Su, X., Zhang, F., Song, Y., Zhao, Z., Xie, W., 2010. A functional polymorphism in the DNA methyltransferase-3A promoter modifies the susceptibility in gastric cancer but not in esophageal carcinoma. BMC Med. 8, 12. doi:10.1186/1741-7015-8-12
Feinberg, A.P., Vogelstein, B., 1983. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature 301, 89–92. doi:10.1038/301089a0
Hu, J., Fan, H., Liu, D., Zhang, S., Zhang, F., Xu, H., 2010. DNMT3B promoter polymorphism and risk of gastric cancer. Dig. Dis. Sci. 55, 1011–1016. doi:10.1007/s10620-009-0831-3
Imai, Y., Oda, H., Nakatsuru, Y., Ishikawa, T., 1995. A polymorphism at codon 160 of human O6-methylguanine-DNA methyltransferase gene in young patients with adult type cancers and functional assay. Carcinogenesis 16, 2441–5.
Jiang, J., Jia, Z., Cao, D., Jin, M.-S., Kong, F., Suo, J., Cao, X., 2012. Polymorphisms of the DNA methyltransferase 1 associated with reduced risks of Helicobacter pylori infection and increased risks of gastric atrophy. PLoS One 7, e46058.

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |20
doi:10.1371/journal.pone.0046058 Kanai, Y., Ushijima, S., Nakanishi, Y., Sakamoto, M., Hirohashi, S., 2003. Mutation of
the DNA methyltransferase (DNMT) 1 gene in human colorectal cancers. Cancer Lett. 192, 75–82.
Khatami, F., Noorinayer, B., Ghiasi, S., Mohebi, R., Hashemi, M., Zali, M.R., 2009. Lack of effects of single nucleotide polymorphisms of the DNA methyltransferase 1 gene on gastric cancer in Iranian patients: a case control study. Asian Pac. J. Cancer Prev. 10, 1177–1182.
Kim, M.S., Kim, Y.R., Yoo, N.J., Lee, S.H., 2013. Mutational analysis of DNMT3A gene in acute leukemias and common solid cancers. APMIS 121, 85–94. doi:10.1111/j.1600-0463.2012.02940.x
Lin, J., Zeng, R.M., Li, R.N., Cao, W.H., 2014. Aberrant DNA methylation of the P16, MGMT,and hMLH1 genes in combination with the methylenetetrahydrofolate reductase C677T genetic polymorphism and folate intake in gastric cancer. Genet. Mol. Res. 13, 2060–2068. doi:10.4238/2014.March.24.10
Oh, H.R., An, C.H., Yoo, N.J., Lee, S.H., 2014. Somatic mutations of amino acid metabolism-related genes in gastric and colorectal cancers and their regional heterogeneity--a short report. Cell. Oncol. (Dordr). 37, 455–61. doi:10.1007/s13402-014-0209-1
Saito, Y., Kanai, Y., Sakamoto, M., Saito, H., Ishii, H., Hirohashi, S., 2001. Expression of mRNA for DNA methyltransferases and methyl-CpG-binding proteins and DNA methylation status on CpG islands and pericentromeric satellite regions during human hepatocarcinogenesis. Hepatology 33, 561–8. doi:10.1053/jhep.2001.22507
Schaefer, M., Lyko, F., 2010. Solving the Dnmt2 enigma. Chromosoma 119, 35–40. doi:10.1007/s00412-009-0240-6
Shen, G.-Q., Abdullah, K.G., Wang, Q.K., 2009. The TaqMan method for SNP genotyping. Methods Mol. Biol. 578, 293–306. doi:10.1007/978-1-60327-411-1_19
Tao, R., Chen, Z., Wu, P., Liu, C., Peng, Y., Zhao, W., Hu, C., Feng, J., 2015. The possible role of EZH2 and DNMT1 polymorphisms in sporadic triple-negative breast carcinoma in southern Chinese females. Tumour Biol. 36, 9849–55. doi:10.1007/s13277-015-3754-y
Wang, C., Jia, Z., Cao, D., You, L., Jin, M., Wu, X., Wen, S., Cao, X., Jiang, J., 2015. Polymorphism of DNA Methyltransferase 3b and Association with Development and Prognosis in Gastric Cancer. PLoS One 10, e0134059. doi:10.1371/journal.pone.0134059
Wang, Y., Lou, B., Yan, L.P., Huang, B.H., Zhao, P., 2005. Relationship between Epstein-Barr virus-encoded proteins with cell proliferation, apoptosis, and apoptosis-related proteins in gastric carcinoma. World J. Gastroenterol. 11, 3234–3239.
Wang, Y.-M., Wang, R., Wen, D.-G., Li, Y., Guo, W., Wang, N., Wei, L.-Z., He, Y.-T., Chen, Z.-F., Zhang, X.-F., Zhang, J.-H., 2005. Single nucleotide polymorphism in DNA methyltransferase 3B promoter and its association with gastric cardiac adenocarcinoma in North China. World J. Gastroenterol. 11, 3623–3627.
Wu, H., Zhang, K., Gong, P., Qiao, F., Wang, L., Cui, H., Sui, X., Gao, J., Fan, H., 2014. A novel functional TagSNP Rs7560488 in the DNMT3A1 promoter is associated with susceptibility to gastric cancer by modulating promoter activity. PLoS One 9, e92911. doi:10.1371/journal.pone.0092911
Wu, Q., Lu, S., Wang, L., Hu, J., Qiao, F., Qiu, X., Zhao, C., Lao, Y., Song, Y., Fan, H., 2012. DNMT3A rs36012910 A>G polymorphism and gastric cancer susceptibility in a Chinese population. Mol. Biol. Rep. 39, 10949–10955. doi:10.1007/s11033-012-1996-y
Xiang, G., Zhenkun, F., Shuang, C., Jie, Z., Hua, Z., Wei, J., Da, P., Dianjun, L., 2010. Association of DNMT1 gene polymorphisms in exons with sporadic infiltrating ductal breast carcinoma among Chinese Han women in the Heilongjiang Province. Clin.

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |21
Breast Cancer 10, 373–7. doi:10.3816/CBC.2010.n.049 Xiong, H.-L., Liu, X.-Q., Sun, A.-H., He, Y., Li, J., Xia, Y., 2013. Aberrant DNA methylation
of P16, MGMT, hMLH1 and hMSH2 genes in combination with the MTHFR C677T genetic polymorphism in gastric cancer. Asian Pac. J. Cancer Prev. 14, 3139–3142.
Yang, J., Wei, X., Wu, Q., Xu, Z., Gu, D., Jin, Y., Shen, Y., Huang, H., Fan, H., Chen, J., 2011. Clinical significance of the expression of DNA methyltransferase proteins in gastric cancer. Mol. Med. Rep. 4, 1139–1143. doi:10.3892/mmr.2011.578
Yang, X.-X., He, X.-Q., Li, F.-X., Wu, Y.-S., Gao, Y., Li, M., 2012. Risk-association of DNA methyltransferases polymorphisms with gastric cancer in the southern chinese population. Int. J. Mol. Sci. 13, 8364–8378. doi:10.3390/ijms13078364
Ye, C., Beeghly-Fadiel, A., Lu, W., Long, J., Shu, X.O., Gao, Y.-T., Zheng, W., Cai, Q., 2010. Two-stage case-control study of DNMT-1 and DNMT-3B gene variants and breast cancer risk. Breast Cancer Res. Treat. 121, 765–9. doi:10.1007/s10549-009-0569-9
Ziogas, D., Roukos, D., 2009. Epigenetics in gastric cancer: challenges for clinical implications. Ann. Surg. Oncol. 16, 2077–8. doi:10.1245/s10434-009-0472-y

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |20
Table 1. Characteristics of all studies included in the analysis
Reference Country
(Ethnicity) Matching Criteria
Cases Controls Gene SNPS (rs)
Genotype method n Age n Age
Wang et al.,2015 China (Asian) Age, gender and H. pylori 447 NA 961 NA DNMT3B rs6119954, rs1569686,
rs4911107, rs4911259, rs8118663 TaqMan assay
Wu et al., 2014 China (Asian) Age and gender 405 NA 408 NA DNMT3A rs7560488 HRM
Cao et al., 2013 China (Asian) Age, gender and H. pylori 447 61.6 961 50.6 DNMT3A rs1550117, rs13420827, TaqMan assay
Wu et al., 2012 China (Asian) Age and gender 340 62.2* 251 62.7* DNMT3A rs36012910 PCR-RFLP
Jiang et al., 2012 China (Asian) Age, gender and H. Pylori 447 61.6 961 50.6 DNMT1 rs10420321, rs16999593,
rs8101866, rs8111085, rs2288349 TaqMan assay
Yang et al., 2012 China (Asian) Age and gender 242 54.9 ± 12.5
294 58.4 ± 16.4
DNMT3A DNMT3B DNMT1 DNMT2
rs16999593, rs8101866, rs2228611, rs2114724,
rs11254413, rs1550117, rs13420827, rs11887120, rs13428812, rs2424908
MALDI-TOF
Fan et al., 2010 China (Asian) Age and gender 208 65 346 71 DNMT3A rs1550117 PCR-RFLP
Hu et al., 2010 China (Asian) Age and gender 259 64.0 262 65.1 DNMT3B rs1569686, rs2424913 PCR-RFLP
Khatami et al., 2009
Iran (Turk and Fars)
Age, gender and ethnicity 200 50±13 200 58±13 DNMT1 rs721186, rs13784, rs2228611,
rs11488 PCR-RFLP
Aung et al., 2005 Japan (Asian) NA 152 64,1* 247 43,9* DNMT3B rs2424913 PCR-RFLP
Wang et al., 2005 China (Asian) Age and gender 212 60.4 294 59.1 DNMT3B rs2424913 PCR-RFLP
NA, Not Available; * Median (range); H. Pylori - Helicobacter Pylori; DNMT1 - DNA (cytosine-5)-methyltransferase 1; DNMT2 - TRNA (cytosine38-C5)-methyltransferase; DNMT3A - DNA (cytosine-5)-methyltransferase 3 alpha; DNMT3B - DNA (cytosine-5-)-methyltransferase 3 beta; MALDI-TOF - Matrix-Assisted Laser Desorption/Ionization- Time of Flight; PCR-RFLP - Polymerase Chain Reaction-Restriction Fragment Length Polymorphism; HRM - High Resolution Melting.
CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |21
Table 2.1 Genotyping results for DNMT1 of all studies included
Gene SNP Genotype Distribution n (%)
Analysis Model OR (95%CI) p value Reference Cases Controls
DN
MT
1
rs10420321 AA AG GG AA AG GG
141 (31.5) 203 (51.5) 76 (17) 328 (34.1) 454 (47.2) 179 (18.6) CD 1.17 (0.88-1.55) (GG) 0.29 Jiang 2012
rs16999593
TT TC CC TT TC CC
283 (63.3) 144 (32.2) 20 (4.5) 659 (68.6) 273 (28.4) 29 (3) CD 1.25(0.95-1.66) (TC) 0.18 Jiang 2012
141 (58.3) 89 (36.8) 12 (5) 196 (66.7) 83 (28.2) 15 (5.1) CD 1.47 (1.01-2.14) (TC) 0.11
Yang 2012 Dominant 1.45 (1.00-2.11) (TC+CC) 0.05
rs8101866
CC CT TT CC CT TT
238 (53.5) 177 (39.8) 30 (6.7) 489 (50.9) 402 (41.8) 70 (7.3) CD 0.81 (0.18-1.35) (CT) 0.11 Jiang 2012
13 (5.4) 98 (40.1) 130 (53.9) 26 (8.8) 102 (34.7) 166 (56.5) CD 1.20 (0.83-1.74) (CT) 0.17
Yang 2012 Dominant 1.27 (0.89-1.82) (CT + CC) 0.19
rs8111085 TT TC CC TT TC CC
143 (32) 214 (47.9) 90 (20.1) 330 (34.3) 447 (46.5) 184 (19.1) CD 1.18 (0.32-1.69) (CC) 0.38 Jiang 2012
rs2288349 GG GA AA GG GA AA
249 (56.1) 161 (36.3) 34 (7.7) 515 (53.6) 372 (38.7) 74 (7.7) CD 0.81(0.50-1.33) (AA) 0.81 Jiang 2012
rs2114724
CC CT TT CC CT TT
132 (54.5) 97 (40.1) 13 (5.4) 162 (56.2) 101 (35.1) 25 (8.7) CD 1,16 (0,81-1,68) (CT) 0,22
Yang 2012 Dominant 1.23 (0.86-1.76) (CT+TT) 0.27
rs2228611
GG GA AA GG GA AA
132 (54.5) 97 (40.1) 13 (5.4) 160 (56.1) 99 (34.7) 26 (9.1) CD 1.18 (0.81-1.71) (GA) 0.14
Yang 2012 Dominant 1.26 (0.87-1.80) (GA+GG) 0.22
34 (34) 50 (50) 16 (16) 32 (32) 62 (62) 18 (18) CD 1.13 (0.05-6.30) (GA) NA Khatami 2009
rs721186 CC CT TT CC CT TT
99 (99) 1 (1) - 200 (100) - - NA NA NA Khatami 2009
rs13784 CC CT TT CC CT TT
200 (100) - - 100 (100) - - NA NA NA Khatami 2009
rs11488 AA AT TT AA AT TT
200 (100) - - 200 (100) - - NA NA NA Khatami 2009

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |22
Table 2.2 Genotyping results for DNMT2 of all studies included
Gene SNP Genotype Distribution n (%) Analysis
Model OR (95%CI) p value Reference
Cases Controls
DNMT2 rs11254413
GG GA AA GG GA AA
204 (84.3) 15 (6.2) 23 (9.5) 187 (63.8) 91 (31.1) 15 (5.1) CD 0.16 (0.09-0.28) (GA) <0.01
Yang 2012 Dominant 0.15 (0.08-0.27) (GA+AA) <0.01
Table 2.3 Genotyping results for DNMT3A of all studies included
Gene SNP Genotype Distribution n (%) Analysis
Model OR (95%CI) p value Reference
Cases Controls
DN
MT
3A
rs7560488
TT TC CC TT TC CC
279 (68.9) 111 (27.4) 15 (3.7) 326 (79.9) 75 (18.4) 7 (1.7) CD
1.73(1.24-2.41) (TC); 2.50(1.01-6.23) (CC)
0.01; 0.04 Wu 2014
Dominant 1.65(1.19-2.29) (TC + CC) 0.002
rs1550117
GG GA AA GG GA AA
289 (64.7) 142 (31.8) 16 (3.6) 640 (66.6) 288 (30) 33 (3.4) CD 1.10 (0.83-1.47) (AA) 0.51
Cao 2013 Dominant 1.06 (0.83-1.32) (GA+AA) 0.51
157 (64.9) 74 (30.6) 11 (4.5) 191 (65) 93 (31.6) 10 (3.4) CD 1.34 (0.55-3.29) (AA) 0.74
Yang 2012 Dominant 0.92 (0.63-1.34) (GA+AA) 0.68
102 (49) 75 (36.1) 31 (14.9) 218(63) 118 (34.1) 10 (2.9) CD 6.63 (3.13-14.03) (AA) 0.00 Fan 2010
rs13420827
CC CG GG CC CG GG
295 (66) 133 (29.8) 19 (4.3) 618 (64.3) 304 (31.6) 39 (4.1) CD 1.06 (0.79-1.41) (GG) 0.71
Cao 2013 Dominant 1.05(0.83-1.33) (CG+GG) 0.64
167 (69) 61 (25.2) 14 (5.8) 183 (62.7) 99 (33.9) 10 (3.4) CD 0.68 (0.46-1.01) (CG) 0.05
Yang 2012 Dominant 0.66 (0.45-0.97) (CG+GG) 0.03
rs36012910
AA AG GG AA AG GG
288 (84.71) 51 (15) 1 (0.29) 234 (93.23) 17 (6.77) 0 CD 2.44 (1.37-4.33) (AG) 0.002
Wu 2012 Dominant 2.36 (1.35-4.14) (AG+GG) 0.002
rs11887120 TT TC CC TT TC CC
57 (23.6) 121 (50) 64 (26.4) 74 (25,3) 155 (53,1) 63 (21,6) CD 1.37 (0.88-2.13) (AA) 0.17 Wang 2015
rs13428812
AA AG GG AA AG GG
137 (56.6) 84 (34.7) 21 (8.7) 160 (55.4) 106 (36.7) 23 (8) CD 1.11 (0.58-2.12) (GG) 0.84
Yang 2012 Dominant 0.91 (0.64-1.31) (AG+GG) 0.63
CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |23
Table 2.4 Genotyping results for DNMT3A of all studies included
Gene SNP Genotype Distribution n (%) Analysis
Model OR (95%CI) p value Reference
Cases Controls
DN
MT
3B
rs6119954 GG GA AA GG GA AA
200 (44.8) 199 (44.6) 4 (10.6) 433 (45.1) 445 (46.3) 83 (8.6) CD 1.37 (0.88-2.13) (AA) 0.17 Wang 2015
rs1569686
TT TG GG TT TG GG
360 (80.5) 82 (18.3) 5 (1.1) 801 (83.4) 150 (15.6) 10 (1) CD 1.33 (0.41-4.31) (GG) 0.64 Wang 2015
230 (88.8) 27 (10.43) 2 (0.77) 203 (77.48) 55 (20.99) 4 (1.53) Dominant 0.43 (0.26-0.72) (TG+GG) <0.01 Hu 2010
rs4911107 AA AG GG AA AG GG
360 (80.5) 82 (18.3) 5 (1.1) 800 (83.2) 151 (15.7) 10 (1.1) CD 1.08 (0.13-8.86) (GG) 0.94 Wang 2015
rs4911259 GG GT TT GG GT TT
359 (80.3) 83 (18.6) 5 (1.1) 799 (83.1) 152 (15.8) 10 (1.1) CD 1.09(0.13-8.88) (TT) 0.94 Wang 2015
rs8118663 AA AG GG AA AG GG
126 (28.2) 233 (52.1) 88 (19.7) 314 (32.7) 473 (49.2) 174 (18.1) CD 1.32 (0.91-1.91) (GG) 0.15 Wang 2015
rs2424908
TT TC CC TT TC CC
78 (32.2) 114 (47.1) 50 (20.7) 99 (33.7) 139 (47.3) 56 (19.1) CD 1.05 (0.64-1.71) (CC) 0.96
Yang 2012 Dominant 0.96 (0.68-1.36) (TC+CC) 0.82
rs2424913
CC CT TT CC CT TT
257 (99.2) 2 (0.8) - 259 (98.8) 3 (1.2) - CD 1.49 (0.17-17.94) (CT) >0,05 Hu 2010
152 (100) - - 247 (100) - - NA NA NA Aung 2005
205 (96.7) 7 (3.3) - 279 (94.9) 15 (5.1) - Dominant 0.66 (0.26-1.67) (CT) 0,38 Wang 2005
NA, Not Available; CD, Co-Dominant; NR, No Risk; DNMT1 – DNA (cytosine-5)-methyltransferase 1; DNMT2 – TRNA (cytosine38-C5)-methyltransferase; DNMT3a – DNA (cytosine-5)-methyltransferase 3 alpha; DNMT3b - DNA (cytosine-5-)-methyltransferase 3 beta; SNP – Single Nucleotide Polymorphism; Allele: A- Adenine, C- Cytosine, G- Guanine, T – Thymine; OR (95% CI) – Odds Ratio (95% Confidence Interval)

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |25
Figure 1. Flow chart of studies identification, exclusion and inclusion. Adapted from: Moher D,
Liberati A, Tetzlaff J, Altman DG, The PRISMA Group (2009). Preferred Reporting Items for
Systematic Reviews and Meta-analyses: The PRISMA Statement. PLoS Med 6(6): e1000097.
doi:10.1371/journal.pmed1000097
Records identified through
database searching
(n =35)
Sc
ree
nin
g
Inclu
de
d
Eli
gib
ilit
y
Iden
tifi
cati
on
Additional records identified
through other sources
(n =0)
Records after duplicates removed
(n =35)
Records screened
(n =35)
Full-text articles
assessed for eligibility
(n =18)
Full-text articles
excluded (n =7)
MGMT polymorphisms (n=4)
Only evaluate DNMTs mutations (n=2)
Allelic loss in gastric cancer (n=1) Studies included in
qualitative synthesis
(n = 11)
Records excluded (n =17)
Do not evaluate DNMTs polymorphisms (n=9)
Do not evaluate polymorphisms (n=3)
Not related with gastric cancer (n=3)
Review or meta-analysis (n=2)

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |26
Figure 2. Forest-plot for DNMT1 SNPs
Figure 3. Forest-plot for DNMT3A SNPs
Figure 4. Forest-plot for DNMT3B SNPs

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |27
Supplementary Table 1. Description of DNMTs SNPS included in the studies Gene
(Chromosome) SNP Location
Allele Change
Amino Acid Change
Function
DNMT1 (Chromosome 19)
(19p13.2)
rs11488 10133368 A/T NA 3' UTR
rs13784 10133520 C/T NA 3' UTR
rs2288349 10146569 G/A NA Intron
rs2114724 10154572 C/T NA Intron
rs721186 10154636 C/T Thr594 Synonymous codon
rs2228611 10156401 G/A Pro463 Synonymous codon
rs8111085 10162696 T/C Ile327Val Missense
rs8101866 10164984 C/T NA Intron
rs16999593 10180505 T/C His97Arg Missense
rs10420321 10189741 A/G NA Intron
DNMT2 (Chromosome 10)
(10p15.1) rs11254413 17162188 G/A His101Tyr Missense
DNMT3A (Chromosome 2)
(2p23)
rs7560488 25345952 T/C NA NA
rs1550117 25343038 G/A NA NearGene-5
rs13420827 25231099 C/G NA NA
rs36012910 25345310 A/G NA Promoter
rs11887120 25262866 T/C NA Intron
rs13428812 25269598 A/G NA Intron
DNMT3B (Chromosome 20)
(20q11.2)
rs6119954 32776360 G/A NA Intron
rs1569686 32779273 T/G NA Intron
rs4911107 32787185 A/G NA Intron
rs4911259 32788476 G/T NA Intron
rs8118663 32811070 A/G NA NA
rs2424908 32772577 T/C NA Intron
rs2424913 32786453 C/T NA Intron
NA, Not Available; DNMT1 – DNA (cytosine-5)-methyltransferase 1; DNMT2 – TRNA (cytosine38-C5)-methyltransferase; DNMT3a – DNA (cytosine-5)-methyltransferase 3 alpha; DNMT3b - DNA (cytosine-5-)-methyltransferase 3 beta; SNP – Single Nucleotide Polymorphism; Allele: A- Adenine, C- Cytosine, G- Guanine, T – Thymine; UTR – Untranslated Region

CHAPTER 2
Polymorphisms in DNA regulation-associated genes and GC development |28
Supplementary Table 2. HWE in controls in all studies
Gene SNP Reference HWE (p)
DNMT1
rs10420321 Jiang 2012 0.323
rs16999593 Jiang 2012 0.910
Yang 2012 0.120
rs8101866 Jiang 2012 0.306
Yang 2012 0.078
rs8111085 Jiang 2012 0.139
rs2288349 Jiang 2012 0.550
rs721186 Khatami 2009 0.550
rs13784 Khatami 2009 0.550
rs2228611 Khatami 2009 0.187
Yang 2012 0.068
rs11488 Khatami 2009 0.550
rs2114724 Yang 2012 0.113
DNMT2 rs11254413 Yang 2012 0.371
DNMT3A
rs7560488 Wu 2014 0.274
rs1550117
Cao 2013 0.932
Yang 2012 0.747
Fan 2010 0.206
rs13420827 Cao 2013 0.833
Yang 2012 0.444
rs36012910 Wu 2012 0.579
rs11887120 Yang 2012 0.280
rs13428812 Yang 2012 0.450
DNMT3B
rs6119954 Wang 2015 0.036
rs1569686 Wang 2015 0.321
Hu 2010 0.901
rs4911107 Wang 2015 0.341
rs4911259 Wang 2015 0.362
rs8118663 Wang 2015 0.859
rs2424908 Yang 2012 0.563
rs2424913
Hu 2010 0.926
Aung 2005 0.824
Wang 2005 0.654
DNMT1 – DNA (cytosine-5)-methyltransferase 1; DNMT2 – TRNA (cytosine38-C5)-methyltransferase; DNMT3A – DNA (cytosine-5)-methyltransferase 3 alpha; DNMT3B - DNA (cytosine-5-)-methyltransferase 3 beta; HWE - Hardy–Weinberg equilibrium (http://www.had2know.com/academics/hardy-weinberg-
equilibrium-calculator-2-alleles.html)

CHAPTER 3


CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |30
TITLE
SINGLE NUCLEOTIDE POLYMORPHISMS IN DNA METHYLTRANSFERASES AND
GASTRIC CANCER
AUTHORS
Neves M1,2, Ribeiro J1,2,3, Dinis-Ribeiro M,4,5, Medeiros R1,3,6,7, Sousa H1, 6
AFILIATIONS
1 Molecular Oncology and Viral Pathology Group, CI-IPOP, Portuguese Oncology
Institute of Porto (IPO-Porto), Porto, Portugal.
2 Faculty of Medicine of University of Porto, Porto, Portugal.
3 Research Department, Portuguese League Against Cancer, Porto, Portugal.
4 Center for Health Technology and Services Research (CINTESIS), Faculty of Medicine,
University of Porto, Porto, Portugal.
5Gastroenterology Department, Portuguese Oncology Institute of Porto, Porto, Portugal.
6 Abel Salazar Institute for the Biomedical Sciences (ICBAS), University of Porto, Rua de
Jorge Viterbo Ferreira n.º 228, 4050-313 Porto, Portugal.
7Virology Service, Portuguese Oncology Institute of Porto, Porto, Portugal.
Correspondence to:
Hugo Sousa, MD PhD
Serviço de Virologia, Laboratórios 4º Piso
Instituto Português de Oncologia do Porto FG EPE
Rua Dr. António Bernardino Almeida
4200-072 Porto, Portugal
Phone: +351 22 508 4000 (ext 5410)
Fax: +351 22 508 4001
E-mail: [email protected] or [email protected]

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |31
ABSTRACT
AIM: To investigate the relationship between the DNMT2 rs11254413 and DNMT3A1
rs7560488 and the development of gastric lesions and gastric adenocarcinoma.
METHODS: A case-control study was performed involving 380 individuals, 220 without
evidence of neoplastic disease (control), 43 with gastric lesion and 117 patients with
gastric adenocarcinomas. Genotyping was executed with TaqMan® SNP Genotyping
Assay.
RESULTS: The genotyping results for DNMT2 rs11254413 and DNMT3A1 rs7560488
revealed no association with gastric lesion and gastric cancer. DNMT2 rs11254413 GG,
GA and AA genotypes frequencies were 27.3%, 51.8% and 20.9%, respectively in
controls; 25.6%, 48.8% and 25.6% in patients with gastric lesions; and 28.4%, 45.7%
and 25.9% in patients with gastric adenocarcinoma. Regarding DNMT3A1 rs7560488
analysis the frequencies of TT, TC and CC genotypes were 29.3%, 51.4% and 19.3%,
respectively in controls, 35.7%, 47.6% and 16.7% in patients with gastric lesions and
28.2%, 50.9% and 20.9% in patients with gastric adenocarcinoma.
CONCLUSION: DNMT2 rs11254413 and DNMT3A1 rs7560488 may not be associated
with the development of gastric lesions and gastric cancer. More studies, with different
populations, are needed to confirm our findings.
KEY WORDS
DNA Methyltransferase; DNMT2; DNMT3A; Single Nucleotide polymorphism; Gastric
Cancer

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |32
INTRODUTION
Gastric cancer (GC) is one of the most common type of malignancies worldwide. It was
the fifth common malignancy in 2012 worldwide (952,000 cases, 6.8% of the total) and
the third leading cause of cancer deaths worldwide (723,000 deaths, 8.8% of the total)
(1). According to Lauren’s classification, GC can be divided in two types: intestinal and
diffuse type (2). The intestinal type is characterized by a stepwise carcinogenesis from
normal mucosa through atrophic gastritis, atrophy, intestinal metaplasia and dysplasia
to invasive adenocarcinoma (3,4). On the other hand, diffuse type is characterized by a
less differentiated malignant neoplasia, without a well-marked multiple step process
(3,4).
The major risk factor for the development of GC is Helicobacter pylori infection
(especially the intestinal type), nevertheless, GC can also be the result of specific genetic
alterations. Another factor recently associated to CG has been epigenetic changes (5,6).
Alterations in DNA methyltransferases (DNMTs), more precisely, DNMT1, DNMT2 and
DNMT3A have been studied and have been suggested to have a role in carcinogenesis
of GC (7,8,6,9). DNMTs are enzymes that catalyze the process of promoter methylation.
There are three types of DNMTs: DNMT1, DNMT2 and DNMT3 (10). DNMT1 function is
catalyze DNA post-replication methylation and maintain DNA methylation patterns during
cell divisions (11,10,12). DNMT2 is the most conserved DNMT and their function is
catalyze DNA and aspartic acid tRNA methylation (13). DNMT3 is divided in two types:
DNMT3A and DNMT3B and they are responsible for de novo methylation during
gametogenesis and embryogenesis (8).
The most common form of genetic variants of the human genome are Single Nucleotide
Polymorphisms (SNPs) (14) . Some SNPs have functional effects on the susceptibility to
the development of human cancers by modifying the transcriptional expression
(5,15,16). Several SNPs in DNMTs have been identified and some have been associated
to GC risk (17,8,18). For example, SNP rs11254413 of DNMT2 leads to missense
mutation of His to Tyr, and may also alter the function of the protein encoded by the
DNMT2 gene. Yang 2012 demonstrated that GA genotype of rs11254413 has a
potentially protective effect on GC risk (8). A study produced by Wu 2014 concluded that
DNMT3A1 rs7560488 can be a promising biomarker to evaluate GC susceptibility (6).
Other studies didn’t find an association between DNMTs SNPs and susceptibility for GC
(17,18).

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |33
The aim of this study was to evaluate the association of DNMT2 and DNMT3A SNPs
(rs11254413 and rs7560488, respectively) with the development of gastric lesions and
gastric adenocarcinoma, in a population from the northern region of Portugal.
METHODS
Study Participants
A retrospective hospital-based case-control study was developed with 43 patients with
gastric lesion, 117 histologically confirmed GC patients and 220 cancer-free controls
from the northern region of Portugal recruited at the Portuguese Institute of Oncology,
Porto. Controls are healthy individuals without any clinical evidence of cancer selected
from a database of blood donors. The characteristics of our population are presented in
Table 1. An adjustment of gender and age it was taken into account to select controls.
The study was submitted to the approval of Ethical Committee from host institution.
Sample Collection and Processing
In this study we used peripheral blood samples collected with standard venipuncture
techniques in EDTA-containing tubes during routine clinical visits and stored in the
institution archives. Total nucleic acids extraction was performed with QIAamp DNA
Blood Mini Kit (QIAGEN, Venlo, Netherlands) according to manufacturer's instructions.
DNA quality was assessed by measuring the absorbance at 260/280nm using an
UV/Visible spectrophotometer and its purity assessed by the ratio of the values of
absorbance at 260/280nm. The presence of amplifiable genomic DNA was tested with a
PCR protocol for amplification of beta-globin gene (19).
Genotyping of DNMTs SNPs
The DNMTs SNPs, rs11254413 and rs7560488, were genotyped using the TaqMan®
SNP Genotyping Assay C___2822597_10 and C__26104534_10, respectively (Applied
Biosystems, Foster City CA, USA).
Reactions were performed in a 5μL master containing 1x TaqMan Genotyping Master
Mix (Applied Biosystems, Foster City CA, USA), 1x TaqMan® SNP Genotyping Assay
(Applied Biosystems, Foster City CA, USA) and 10ng of DNA from each sample.
Amplification was performed on the Applied Biosystems 7300 Real Time PCR System
(Applied Biosystems, Foster City CA, USA) according to the manufacturer instructions.

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |34
Allelic discrimination was performed by measuring end-point fluorescence and analyzed
by two authors. Negative controls were used using double distilled water and 10% of all
samples were randomly selected and re-submitted to genotyping.
Statistical analysis
The Hardy–Weinberg equilibrium (HWE) was tested by calculated through the
application of the online software (http://www.had2know.com/academics/hardy-
weinberg-equilibrium-calculator-2-alleles.html) to compare the observed versus the
expected genotype frequencies. A p–value less than 0.05 of HWE was considered
significant.
Data analysis was performed using the computer software IBM SPSS Statistics for
Windows, Version 22.0 (IBM Corp, Armonk NY, USA). Chi-square analysis was used to
compare categorical variables, with a 5% level of significance. Multivariate logistic
regression analysis was used to estimate odds ratio (OR) and its 95% confidence interval
(CI) as a measure of the association between variant allele carriers and the risk for the
development of gastric cancer. Potential confounding variables (age, gender) were
addressed either by being included as covariates in the multivariate analysis or/and
through data stratification. Homozygotes for the allele with the highest frequency was
used as the reference group for each OR estimation. Variant allele carriers were defined
as the heterozygous and minor allele homozygous genotype carriers pooled together
(dominant model).
RESULTS
Allelic distribution of DNMT2 and DNMT3A SNPs
The distribution of rs11254413 genotypes is shown in table 2. The frequency of the GG,
GA and AA genotypes were 27.3%, 51.8% and 20.9%, respectively in controls; and
25.6%, 48.8% and 25.6% in patients with gastric lesions and 28.4%, 45.7% and 25.9%
in patients with gastric adenocarcinoma. The controls genotypic distribution was in HWE
(p = 0.547).
Regarding the distribution of rs7560488 genotypes, the frequencies of TT, TC and CC
genotypes were 29.3%, 51.4% and 19.3%, respectively in controls, 35.7%, 47.6% and
16.7% in patients with gastric lesions and 28.2%, 50.9% and 20.9% in patients with

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |35
gastric adenocarcinoma – table 3. The distribution of genetic variants in controls was in
HWE (p = 0.431).
Risk estimates for associated lesions and gastric adenocarcinoma
Considering DNMT2 rs11254413 status, our results revealed no association with the
development of gastric lesion and adenocarcinoma. We did not found statistically
significant association with the development of gastric lesions and cancer (p = 0.793 and
p = 0.488, respectively) – table 2.
DNMT3A rs7560488 results revealed no statistically significant association with the
development of gastric lesions or GC (p = 0.724 and p = 0.946, respectively) – table 3.
DISCUSION
DNA methylation was the first epigenetic mark linked directly with tumorigenesis.
Aberrant CpG island methylation is associated to gene silencing and to cancer risk,
including GC (16,20). DNMTs are responsible for DNA methylation and literature reveals
that DNMTs are overexpressed in GC (21). Functional SNPs could affect DNMTs
expression, therefore, it is important to study SNPs in DNMTs.
DNMT2 rs11254413, leads to an amino acid change, His to Tyr, that may affect the
function of DNMT2 and it has been described as associated with protection for GC (8).
The other SNP analysed in this study, rs7560488, is a tagSNP representing different
SNPs of DNMT3A promoter with high linkage disequilibrium (5). Nevertheless, this
polymorphism seems to affect transcriptional activity of DNMT3A, resulting in altered
expression of protein and TC+CC genotypes have been associated with increased risk
for GC (5). Both these SNPs have only been studied in GC in one study each (5,8), and
up to our knowledge, this is the first study within a southern European population from
the northern region of Portugal reporting data regarding their role in GC development.
Our results suggest that rs11254413 and rs7560488 have no relation with the
development of gastric lesion and GC. These results are not in concordance with the
results observed in the two studies regarding the two SNPs in study (5,8). The
differences in the allele frequency, between regions, may explain the discordance of our
results with the ones find in the literature, since the two studies were performed with
Asiatic populations. We have compared the genetic distribution in our population with
other populations as described in the HapMap Project. Table 4 describe the allele

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |36
frequency, in control population, of the two SNPs in 3 different regions, Asia, Europe and
Africa. After analyse the data, it is possible to conclude that allele frequency of the two
SNPs is similar comparing Europe and Africa, but major differences are found when
comparing to Asiatic population: in rs11254413, the GA genotype is the most common
genotype in Europe while GG genotype is the most common in Asia; and in rs7560488,
the TC genotype is most common in Europe and the genotype with higher percentage in
Asia is TT. Thus we can conclude that the genotypic distribution in our population is
similar to those already described in Caucasian populations, which may represent a
major genetic difference between these populations.
CONCLUSION
In conclusion, contribution of genetic polymorphisms to the risk of GC may be dependent
on the population in study, as well on several environmental and dietary factors that
influence that population. Our findings suggest these two SNPs (rs11254413 and
rs7560488) do not seem to be associated with GC, nevertheless more studies, with
larger Caucasian populations, are needed to clarify its role in GC susceptibility.
REFERENCES
1. Ferlay J, Soerjomataram I, Ervik M, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide. IARC CancerBase No 11 [Internet] [Internet]. 2013;Lyon, Fran. Available from: http://globocan.iarc.fr
2. Lauren P. The two histological main types of gastric carcinoma: diffuse and so-called intestinal-type carcinoma. an attempt at a histo-clinical classification. Acta Pathol Microbiol Scand [Internet]. 1965 [cited 2016 Jul 26];64:31–49. Available from: http://www.ncbi.nlm.nih.gov/pubmed/14320675
3. Hu B, El Hajj N, Sittler S, Lammert N, Barnes R, Meloni-Ehrig A. Gastric cancer: Classification, histology and application of molecular pathology. J Gastrointest Oncol [Internet]. 2012;3(3):251–61. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3418539&tool=pmcentrez&rendertype=abstract
4. McLean MH, El-Omar EM. Genetics of gastric cancer. Nat Rev Gastroenterol Hepatol [Internet]. 2014 Nov [cited 2016 Aug 17];11(11):664–74. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25134511
5. Fu D-G. Epigenetic alterations in gastric cancer (Review). Mol Med Rep [Internet]. Spandidos Publications; 2015 Sep [cited 2016 Jul 30];12(3):3223–30. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25997695
6. Wu H, Zhang K, Gong P, Qiao F, Wang L, Cui H, et al. A novel functional TagSNP Rs7560488 in the DNMT3A1 promoter is associated with susceptibility to gastric

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |37
cancer by modulating promoter activity. PLoS One. United States; 2014;9(3):e92911.
7. Qu Y, Dang S, Hou P. Gene methylation in gastric cancer. Clin Chim Acta [Internet]. 2013 Sep 23 [cited 2016 Jan 12];424:53–65. Available from: http://www.sciencedirect.com/science/article/pii/S0009898113001964
8. Yang X-X, He X-Q, Li F-X, Wu Y-S, Gao Y, Li M. Risk-association of DNA methyltransferases polymorphisms with gastric cancer in the southern chinese population. Int J Mol Sci. Switzerland; 2012;13(7):8364–78.
9. Mutze K, Langer R, Schumacher F, Becker K, Ott K, Novotny A, et al. DNA methyltransferase 1 as a predictive biomarker and potential therapeutic target for chemotherapy in gastric cancer. Eur J Cancer. England; 2011 Aug;47(12):1817–25.
10. Yang J, Wei X, Wu Q, Xu Z, Gu D, Jin Y, et al. Clinical significance of the expression of DNA methyltransferase proteins in gastric cancer. Mol Med Rep [Internet]. Greece: Spandidos Publications; 2011 Nov 1 [cited 2016 Feb 25];4(6):1139–43. Available from: http://www.spandidos-publications.com/mmr/4/6/1139/abstract
11. Saito Y, Kanai Y, Sakamoto M, Saito H, Ishii H, Hirohashi S. Expression of mRNA for DNA methyltransferases and methyl-CpG-binding proteins and DNA methylation status on CpG islands and pericentromeric satellite regions during human hepatocarcinogenesis. Hepatology [Internet]. 2001 Mar [cited 2016 Feb 25];33(3):561–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11230735
12. Kanai Y, Ushijima S, Kondo Y, Nakanishi Y, Hirohashi S. DNA methyltransferase expression and DNA methylation of CPG islands and peri-centromeric satellite regions in human colorectal and stomach cancers. Int J Cancer [Internet]. 2001 Jan 15 [cited 2016 Feb 25];91(2):205–12. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11146446
13. Schaefer M, Lyko F. Solving the Dnmt2 enigma. Chromosoma [Internet]. 2010 Feb [cited 2016 Feb 14];119(1):35–40. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19730874
14. Collins FS, Brooks LD, Chakravarti A. A DNA Polymorphism Discovery Resource for Research on Human Genetic Variation. Genome Res. Cold Spring Harbor Laboratory Press; 1998;8(12):1229–31.
15. Kanwal R, Gupta K, Gupta S. Cancer epigenetics: an introduction. Methods Mol Biol [Internet]. 2015 [cited 2016 Jul 16];1238:3–25. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25421652
16. Brookes AJ. The essence of SNPs. Gene [Internet]. 1999 Jul [cited 2016 Aug 17];234(2):177–86. Available from: http://linkinghub.elsevier.com/retrieve/pii/S037811199900219X
17. Jiang J, Jia Z, Cao D, Jin M-S, Kong F, Suo J, et al. Polymorphisms of the DNA methyltransferase 1 associated with reduced risks of Helicobacter pylori infection and increased risks of gastric atrophy. PLoS One [Internet]. United States; 2012 Jan [cited 2016 Apr 19];7(9):e46058. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3457938&tool=pmcentrez&rendertype=abstract
18. Cao X-Y, Jia Z-F, Cao D-H, Kong F, Jin M-S, Suo J, et al. DNMT3a rs1550117 polymorphism association with increased risk of Helicobacter pylori infection. Asian Pac J Cancer Prev. Thailand; 2013;14(10):5713–8.
19. Ribeiro J, Teixeira D, Marinho-Dias J, Monteiro P, Loureiro J, Baldaque I, et al. Characterization of human papillomavirus genotypes and HPV-16 physical status in cervical neoplasias of women from northern Portugal. Int J Gynaecol Obstet. Ireland; 2014 May;125(2):107–10.
20. Guo M, Yan W. Epigenetics of gastric cancer. Methods Mol Biol [Internet]. 2015 [cited 2016 Jul 16];1238:783–99. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25421692

CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |38
21. Lin J, Zeng RM, Li RN, Cao WH. Aberrant DNA methylation of the P16, MGMT,and hMLH1 genes in combination with the methylenetetrahydrofolate reductase C677T genetic polymorphism and folate intake in gastric cancer. Genet Mol Res. Brazil; 2014;13(1):2060–8.
22. International HapMap Consortium. The International HapMap Project. Nature [Internet]. 2003 Dec 18 [cited 2016 Aug 29];426(6968):789–96. Available from: http://www.ncbi.nlm.nih.gov/pubmed/14685227
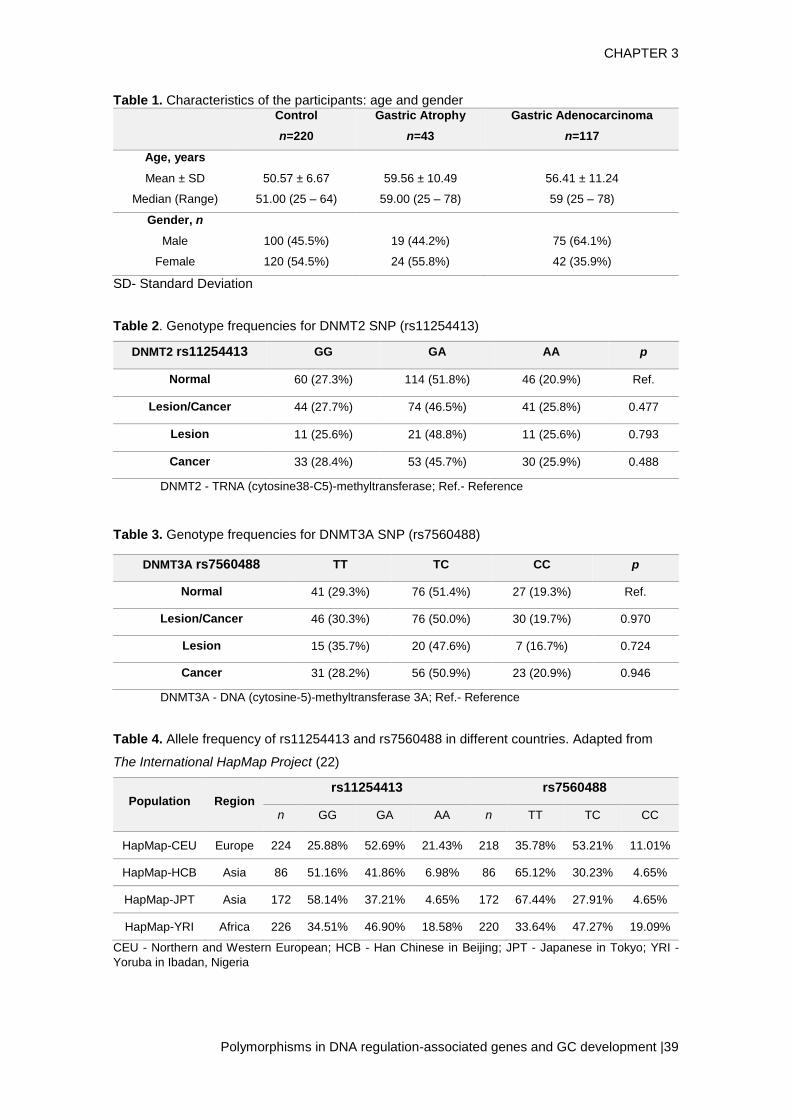
CHAPTER 3
Polymorphisms in DNA regulation-associated genes and GC development |39
Table 1. Characteristics of the participants: age and gender
Control
n=220
Gastric Atrophy
n=43
Gastric Adenocarcinoma
n=117
Age, years
Mean ± SD 50.57 ± 6.67 59.56 ± 10.49 56.41 ± 11.24
Median (Range) 51.00 (25 – 64) 59.00 (25 – 78) 59 (25 – 78)
Gender, n
Male 100 (45.5%) 19 (44.2%) 75 (64.1%)
Female 120 (54.5%) 24 (55.8%) 42 (35.9%)
SD- Standard Deviation
Table 2. Genotype frequencies for DNMT2 SNP (rs11254413)
DNMT2 rs11254413 GG GA AA p
Normal 60 (27.3%) 114 (51.8%) 46 (20.9%) Ref.
Lesion/Cancer 44 (27.7%) 74 (46.5%) 41 (25.8%) 0.477
Lesion 11 (25.6%) 21 (48.8%) 11 (25.6%) 0.793
Cancer 33 (28.4%) 53 (45.7%) 30 (25.9%) 0.488
DNMT2 - TRNA (cytosine38-C5)-methyltransferase; Ref.- Reference
Table 3. Genotype frequencies for DNMT3A SNP (rs7560488)
DNMT3A rs7560488 TT TC CC p
Normal 41 (29.3%) 76 (51.4%) 27 (19.3%) Ref.
Lesion/Cancer 46 (30.3%) 76 (50.0%) 30 (19.7%) 0.970
Lesion 15 (35.7%) 20 (47.6%) 7 (16.7%) 0.724
Cancer 31 (28.2%) 56 (50.9%) 23 (20.9%) 0.946
DNMT3A - DNA (cytosine-5)-methyltransferase 3A; Ref.- Reference
Table 4. Allele frequency of rs11254413 and rs7560488 in different countries. Adapted from
The International HapMap Project (22)
Population Region rs11254413 rs7560488
n GG GA AA n TT TC CC
HapMap-CEU Europe 224 25.88% 52.69% 21.43% 218 35.78% 53.21% 11.01%
HapMap-HCB Asia 86 51.16% 41.86% 6.98% 86 65.12% 30.23% 4.65%
HapMap-JPT Asia 172 58.14% 37.21% 4.65% 172 67.44% 27.91% 4.65%
HapMap-YRI Africa 226 34.51% 46.90% 18.58% 220 33.64% 47.27% 19.09%
CEU - Northern and Western European; HCB - Han Chinese in Beijing; JPT - Japanese in Tokyo; YRI -
Yoruba in Ibadan, Nigeria

CHAPTER 4


CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |41
TITLE
SLC19A1 G80A AND MTHFR C677T AS PREDICTORS OF GASTRIC CANCER
DEVELOPMENT
AUTHORS
Neves M1,2, Francisco M3,4, Ribeiro J1,2,5, Lima A1,4,6, Dinis-Ribeiro M7,8 Medeiros R1,5,6,9,
Sousa H1,9
AFILIATIONS
1 Molecular Oncology and Viral Pathology Group, CI-IPOP, Portuguese Oncology
Institute of Porto (IPO-Porto), Porto, Portugal.
2 Faculty of Medicine of University of Porto, Porto, Portugal.
3 Faculty of Medicine of University of Coimbra, Coimbra, Portugal.
4 CESPU Institute of Research and Advanced Training in Health Sciences and
Technologies, Department of Pharmaceutical Sciences, Gandra PRD, Portugal.
5 Research Department, Portuguese League Against Cancer, Porto, Portugal.
6 Abel Salazar Institute for the Biomedical Sciences (ICBAS), University of Porto, Porto,
Portugal.
7Center for Health Technology and Services Research (CINTESIS), Faculty of Medicine,
University of Porto, Porto, Portugal.
8Gastroenterology Department, Portuguese Oncology Institute of Porto, Porto, Portugal.
9 Virology Service, Portuguese Oncology Institute of Porto, Porto, Portugal.
Correspondence to:
Hugo Sousa, MD PhD
Serviço de Virologia, Laboratórios 4º Piso
Instituto Português de Oncologia do Porto FG EPE
Rua Dr. António Bernardino Almeida
4200-072 Porto, Portugal
Phone: +351 22 508 4000 (ext 5410)
Fax: +351 22 508 4001
E-mail: [email protected] or [email protected]

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |42
ABSTRACT
AIM: To evaluate the relationship between SLC19A1 G80A and MTHFR C677T and the
development of gastric lesions and gastric cancer.
METHODS: We performed a case-control study including 427 Portuguese individuals:
250 without evidence of neoplastic disease, 43 with gastric lesion and 137 patients with
gastric adenocarcinomas). For genotyping proceeding was used a PCR-RFLP method.
RESULTS: SLC19A1 G80A genotyping results revealed a protective effect for gastric
lesion (OR = 0.373; 95%CI = 0.189-0.736; p = 0.006), but not for gastric cancer.
Regarding MTHFR C677T results, we found an increased risk for GC development (OR
= 2.046; CI 95% = 1.323-3.165; p = 0.001), but no association with gastric lesions.
CONCLUSION: SLC19A1 G80A may have a protective role in gastric lesion
development and MTHFR C677T may be associated with an increased risk for GC
development, but more studies with Caucasian populations are needed to prove our
findings.
KEY WORDS
Folates; SLC19A1; MTHFR; Single Nucleotide Polymorphism; Gastric Cancer

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |43
INTRODUTION
Gastric cancer (GC) is one of the most common type of malignancies worldwide,
representing the fifth common malignancy and the third leading cause of cancer deaths
(1). GC can be divided in two types: intestinal type defined by a sequence of events from
normal mucosa through atrophic gastritis, atrophy, intestinal metaplasia and dysplasia
to invasive adenocarcinoma; and diffuse type characterized by a less differentiated
malignant neoplasia, without a well-marked multiple step process (2,3,4). The
development of GC is associated with several risk factors and the major one, especially
for the development of the intestinal type, is Helicobacter pylori infection. GC can also
be the result of specific genetic alterations, epigenetic alterations and other
environmental factors, such as the low consumption of fruits and vegetables and, more
specifically, folate deficiency. Literature reveals that low folate levels are correlated with
development and invasiveness of GC (5).
It is well known that gene expression can be influenced by a single nucleotide
polymorphism (SNP). Polymorphisms in genes involved in folate metabolism, like
SLC19A1 and MTHFR, may have functional consequences affecting levels of folate in
plasma and therefore individual’s susceptibility to cancer.
SLC19A1 80G>A is characterized by a substitution of a G by an A at position 80, leading
to the change of arginine (Arg) into histidine (His) at codon 27 in the first transmembrane
domain, a region implicated in carrier function (1,4). This polymorphism has been
associated with differential activity of SLC19A1 and literature shows that it may modulate
risk of gastric cancer (GC) (6,7).
MTHFR C677T variant is characterized by a substitution of a C by a T at position 677,
leading to a change of Alanine (Ala) into valine (Val) at codon 222, resulting in a
thermolabile enzyme with reduced specific activity. Several studies have explored the
potential association of this SNP with the susceptibility to GC (8,9).
The aim of this case-study was to evaluate the association of SLC19A1 and MTHFR
SNPs with the development of gastric lesions and gastric adenocarcinoma, in a
population from the northern region of Portugal.

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |44
METHODS
Study Participants
A retrospective hospital-based case-control study was developed with 43 patients with
gastric lesion, 137 histologically confirmed GC patients and 250 cancer-free controls
from the northern region of Portugal recruited at the Portuguese Institute of Oncology,
Porto. Controls are healthy individuals without any clinical evidence of cancer selected
from a database of blood donors. The characteristics of our population are presented in
Table 1. An adjustment for gender and age it was taken into account to select controls.
The study was submitted to the approval of Ethical Committee from host institution.
Sample Collection and Processing
In this study, we used peripheral blood samples collected with standard venipuncture
techniques in EDTA-containing tubes during routine clinical visits and stored in the
institution archives. Total nucleic acids extraction was performed with QIAamp DNA
Blood Mini Kit (QIAGEN, Venlo, Netherlands) according to manufacturer's instructions.
DNA quality was assessed by measuring the absorbance at 260/280nm using an
UV/Visible spectrophotometer and its purity assessed by the ratio of the values of
absorbance at 260/280nm. The presence of amplifiable genomic DNA was tested with a
PCR protocol for amplification of beta-globin gene (10).
Polymorphisms genotyping
The SLC19A1 G80A polymorphism (rs1051266) was genotyped by PCR-RFLP as
previously described (11). PCR reaction was performed in a final volume of 50μl
containing 1× DreamTaq® Green master mix (Thermo Fisher Scientific, MA USA), 0.3μM
of each primer (forward 5´-AGTGTCACCTTCGTCCCCTC-3´ and reverse 5´-
CTCCCGCGTGAAGTTCTT-3´), 1 unit of DreamTaq® (Thermo Fisher Scientific, MA
USA) and 50–100ng of genomic DNA. The PCR amplification consisted of initial
denaturation at 95°C during 5min followed by 40 cycles with denaturation for 15s at 95°C,
annealing/extension for 1min at 58°C and a final extension at 72°C for 7min. PCR
product of 230bp was digested by CfoI (HhaI) (Thermo Fisher Scientific, MA USA)
restriction endonuclease at 37°C for 1h. Individuals with the SLC19A1 80GG genotype
presented two fragments (68 bp and 125 bp), individuals with the SLC19A1 80AA also
presented two fragments (68 bp and 162 bp), whereas individuals with the SLC19A1
80AG genotype presented three fragments (68 bp, 125 bp and 162 bp).

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |45
The MTHFR C677T polymorphism (rs1801133) was genotyped by PCR-RFLP as
previously shown (12). PCR reaction was performed in a final volume of 50μl containing
1× DreamTaq® Green master mix (Thermo Fisher Scientific, MA USA), 0.3μM of each
primer (forward 5´-TGAAGGAGAAGGTGTCTGCGGGA-3´ and reverse 5´-
AGGACGGTGCGGTGAGAGTG-3´), 1 unit of DreamTaq® (Thermo Fisher Scientific,
MA USA) and 50–100ng of genomic DNA. The PCR amplification consisted of initial
denaturation at 94°C during 5min followed by 30 cycles with denaturation for 1min at
94°C, annealing for 1min at 57°C, extension for 15s at 72°C, and a final extension at
72°C for 10min. PCR product of 198bp was digested by HinfI (Thermo Fisher Scientific,
MA USA) restriction endonuclease at 37°C for 1h. Individuals with the MTHFR 677CC
genotype presented one fragment with 198bp, individuals with the MTHFR 677TT
presented one fragment with 175bp, whereas individuals with the MTHFR 677CT
genotype presented two fragments (198bp and 175bp).
Genotypes were accessed by the interpretation of two authors. Negative controls were
added to each PCR reaction using double distilled water to replace template DNA and
10% of all samples were randomly selected and re-submitted to genotyping.
Statistical analysis
The Hardy–Weinberg equilibrium was calculated through online software
(http://www.had2know.com/academics/hardy-weinberg-equilibrium-calculator-2-
alleles.html) to compare the observed versus the expected genotype frequencies and
was considered significant a p–value less than 0.05. Data analysis was performed using
the computer software IBM SPSS Statistics for Windows, Version 22.0 (IBM Corp,
Armonk NY, USA). Chi-square analysis was applied to compare categorical variables,
using a 5% level of significance. Multivariate logistic regression analysis was used to
estimate odds ratio (OR) and its 95% confidence interval (CI) as a measure of the
association between variant allele carriers and the risk for the development of GC.
Possible confounding variables, such as age and gender, were addressed either by
being included as covariates in the multivariate analysis or/and through data
stratification. Homozygotes for the allele with the highest frequency was used as the
reference group for each OR estimation. Variant allele carriers were defined as the
heterozygous and minor allele homozygous genotype carriers pooled together through
a statistically dominant model.

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |46
RESULTS
Allelic distribution of SLC19A1 G80A polymorphism
The distribution of rs1051266 genotypes is shown in table 2. The frequency of the AA,
AG, GG and Gcarrier genotypes were 21.2%, 50.2%, 28.6% and 78.8%, respectively in
controls, 41.9%, 44.2%, 14.0% and 58.1% in patients with gastric lesions and 27.7%,
48.2%, 24.1% and 72.3% in patients with gastric adenocarcinoma. The controls
genotypic distribution was in HWE (p = 0.88).
Allelic distribution of MTHFR C677T polymorphism
Regarding the distribution of rs1801133 genotypes, the frequencies of CC, CT, TT and
Tcarrier genotypes were 50.8%, 33.2%, 16.0% and 49.2%, respectively in controls,
41.9%, 41.9%, 16.3% and 58.1% in patients with gastric lesions and 33.6%, 47.2%,
17.2% and 66.4% in patients with gastric adenocarcinoma – table 3. The HWE was
presented in controls genotypic distribution (p = 0.15).
Risk estimates for associated lesions and gastric adenocarcinoma
Table 2 describes the Odds Ratio (OR) for the development of gastric lesion and
adenocarcinoma considering SLC19A1 G80A status. We found statistically significant
risk for the development of gastric lesion (OR = 0.373; 95%CI = 0.189-0.736; p = 0.006).
We observed a possible protective role for 80G carriers, and when the same analysis
was adjusted for age and gender we also observed this protective role (OR = 0.347;
95%CI = 0.158-0.762; p = 0.008). In contrast, we found no statistically significant risk for
the development of GC (OR = 0.699; 95%CI = 0.430-1.136; p = 0.166). When the same
analysis was adjusted for age and gender we also found the same no statistically
significant results (OR = 0.692; 95%CI = 0.410-1.166; p = 0.167).
Table 3 describes the OR for the development of gastric lesion and GC considering
MTHFR C677T status. We found no statistically risk for the development of lesion (OR
= 1.436; CI 95% = 0.745-2.771; p = 0.322). When the same analysis was adjusted for
age and gender we also found the same no statistically significant results (OR = 1.155;
CI 95% = 0.556-2.400; p = 0.700). In contrast, we found statistically significant risk for
the development of GC (OR = 2.046; CI 95% = 1.323-3.165; p = 0.001). We observed a
2-fold increased risk for the development of GC, and when the same analysis was
adjusted for age and gender we also found a statistically significant risk for the
development of GC (OR = 1.918; CI 95% = 1.209-3.044; p = 0.006).

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |47
DISCUSSION
Literature revealed that lower serum folate levels are associated with development and
invasiveness of gastric cancer (5). Folates are important for DNA methylation and
nucleotide biosynthesis, and low levels of folate may result in DNA global
hypomethylation, DNA damage, impaired DNA repair and altered oncogene and tumor
suppressor gene expressions (5,13). Functional SNPs, in genes involved in folate
metabolism, may affect folate levels, therefore, it is important to study SNPs in genes
important for folate metabolism, such as, SLC19A1 and MTHFR.
The SLC19A1 G80A polymorphism leads to an amino acid change of Arg into His at
codon 27 in the first transmembrane domain, a region implicated in carrier function (1,4).
In literature, we found only one study evaluating the association of SLC19A1 G80A
polymorphism with GC development (7). Wang and colleagues reported a 1.43-fold
increased risk for GC development (7).
Literature reports other SNP in a protein involved in the folates metabolism that is often
associated with other diseases (9,13–16). The MTHFR C677T, characterized by an
amino acid change of Ala into Val at codon 222, results in a thermolabile enzyme with
reduced specific activity (9). Indeed, this polymorphism has been shown to significantly
increase susceptibility to gastric cancer (9,13–16).
A study conducted by Chango et. al demonstrated that individuals with SLC19A1 80AA
and MTHFR 677TT genotypes had significantly lower plasma folate levels and
homocysteinemia (high level of homocysteine in blood) than individuals with wild-type
SLC19A1 80GG and MTHFR 677CC genotypes (17). This evidence suggests a role for
both SLC19A1 G80A and MTHFR C677T in the carcinogenesis of GC.
SLC19A1 G80A and development of gastric lesion and GC
SLC19A1 G allele seems to be extremely frequent (78.8%) in our population, which by
comparison with data reported from other studies and from HapMap project is different
among the different populations (Asia, Europe and Africa). The G allele is more frequent
in Europe than in Asia and Africa. Moreover, our results for the genotype frequencies are
in concordance with reported data in Caucasian populations – table 4.
Our results suggest that SLC19A1 G80A is associated with a protective effect for gastric
lesion (OR = 0.373; 95%CI = 0.189-0.736; p = 0.006), but not with GC. After an analysis
adjusted for age and gender, the results remain statistically significant for gastric lesion

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |48
but not for GC – Table 1. Up to our knowledge, there is no study regarding the association
of this SNP with gastric lesion. A meta-analysis performed by Huang et al. revealed a
protective association with GC (GA+GG: OR = 0.89, 95%CI = 0.81-0.99, p = 0.030) (6).
Our results also demonstrated the protective effect of G allele with GC but the results
were not statically significant (OR = 0.699; 95%CI = 0.430-1.136; p = 0.166).
MTHFR C677T and development of gastric lesion and GC
Regarding MTHFR C677T allele frequency, our results shown that is no big difference
between allele frequencies. The frequencies of this variant seem to diverge among
different ethnic populations: C allele is more frequent in Europe and Africa, with the
particularity that in Africa, the TT genotype is not present; in Asia the frequencies differ
between countries, being the C allele more common in Japan and the T allele more
frequent in China – Table 4. Furthermore, our results are not in concordance with the
reported data in Caucasian populations, being more similar to those from Chinese allele
frequencies. This results may be one factor to explain different risk susceptibilities of
populations and may approximate us from those countries with higher incidences of GC
when comparing to the European average rates (18).
MTHFR C677T is associated with an increased risk for GC development (OR = 2.046;
CI 95% = 1.323-3.165; p = 0.001), but was not associated with gastric lesions. The
analysis adjusted for age and gender presented the same results – Table 2. Our results
are in concordance with the results observed in the literature concerning the association
of this SNP with GC (9,13–16). A meta-analysis presented by Yan et al. showed that,
after stratified analyses according to ethnicity, MTHFR C677T may be a risk factor for
gastric cancer among Asians and Caucasians (16). Regarding the association of MTHFR
C677T with gastric lesion, to our knowledge, there is no study demonstrating this
association, and despite we report a tendency for an increased risk for the development
of gastric lesion (OR = 1.436; 95%CI = 0.745-2.771; p = 0.322) results were not statically
significant.

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |49
CONCLUSIONS
In conclusion, our findings suggest that SLC19A1 G80A may have a protective role in
gastric lesion development and MTHFR C677T may be associated with an increased
risk for GC development. Up to our knowledge, this is the first study within a southern
European population, from the north region of Portugal evaluating the association of
SLC19A1 G80A and MTHFR C677T polymorphisms. The contribution of genetic
polymorphisms to the risk of GC may be dependent on the population in study, as well
on several environmental and dietary factors that influence that population. More studies
are needed, with Caucasian populations, to understand the relation of both SNPs with
GC.
REFERENCES
1. Ferlay J, Soerjomataram I, Ervik M, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide. IARC CancerBase No 11 [Internet] [Internet]. 2013;Lyon, Fran. Available from: http://globocan.iarc.fr
2. Lauren P. The two histological main types of gastric carcinoma: diffuse and so-called intestinal-type carcinoma. an attempt at a histo-clinical classification. Acta Pathol Microbiol Scand [Internet]. 1965 [cited 2016 Jul 26];64:31–49. Available from: http://www.ncbi.nlm.nih.gov/pubmed/14320675
3. Hu B, El Hajj N, Sittler S, Lammert N, Barnes R, Meloni-Ehrig A. Gastric cancer: Classification, histology and application of molecular pathology. J Gastrointest Oncol [Internet]. 2012;3(3):251–61. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3418539&tool=pmcentrez&rendertype=abstract
4. McLean MH, El-Omar EM. Genetics of gastric cancer. Nat Rev Gastroenterol Hepatol [Internet]. 2014 Nov [cited 2016 Aug 17];11(11):664–74. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25134511
5. Lee TY, Chiang EP, Shih YT, Lane HY, Lin JT, Wu CY. Lower serum folate is associated with development and invasiveness of gastric cancer. World J Gastroenterol. 2014;20(32):11313–20.
6. Huang X, Gao Y, He J, Cai J, Ta N, Jiang H, et al. The association between RFC1 G80A polymorphism and cancer susceptibility: Evidence from 33 studies. J Cancer [Internet]. 2016;7(2):144–52. Available from: http://www.jcancer.org/v07p0144.htm
7. Wang L, Chen W, Wang J, Tan Y, Zhou Y, Ding W, et al. Reduced folate carrier gene G80A polymorphism is associated with an increased risk of gastroesophageal cancers in a chinese population. Eur J Cancer. 2006;42(18):3206–11.
8. Gotze T, Rocken C, Rohl FW, Wex T, Hoffmann J, Westphal S, et al. Gene polymorphisms of folate metabolizing enzymes and the risk of gastric cancer. Cancer Lett. 2007;251(2):228–36.
9. Xiong H-L, Liu X-Q, Sun A-H, He Y, Li J, Xia Y. Aberrant DNA methylation of P16, MGMT, hMLH1 and hMSH2 genes in combination with the MTHFR C677T genetic polymorphism in gastric cancer. Asian Pac J Cancer Prev. Thailand; 2013;14(5):3139–42.
10. Ribeiro J, Teixeira D, Marinho-Dias J, Monteiro P, Loureiro J, Baldaque I, et al.

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |50
Characterization of human papillomavirus genotypes and HPV-16 physical status in cervical neoplasias of women from northern Portugal. Int J Gynaecol Obstet. Ireland; 2014 May;125(2):107–10.
11. Lima A, Bernardes M, Sousa H, Azevedo R, Costa LL, Ventura F, et al. SLC19A1 80G allele as a biomarker of methotrexate-related gastrointestinal toxicity in Portuguese rheumatoid arthritis patients. Pharmacogenomics [Internet]. England; 2014 Apr;15(6):807–20. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24350725
12. Lima A, Monteiro J, Bernardes M, Sousa H, Azevedo R, Seabra V, et al. Prediction of methotrexate clinical response in Portuguese rheumatoid arthritis patients: Implication of MTHFR RS1801133 and ATIC RS4673993 polymorphisms. Biomed Res Int. Hindawi Publishing Corporation; 2014;2014.
13. Gao S, Ding L-H, Wang J-W, Li C-B, Wang Z-Y. Diet folate, DNA methylation and polymorphisms in methylenetetrahydrofolate reductase in association with the susceptibility to gastric cancer. Asian Pac J Cancer Prev. Thailand; 2013;14(1):299–302.
14. Xia L-Z, Liu Y, Xu X-Z, Jiang P-C, Ma G, Bu X-F, et al. Methylenetetrahydrofolate reductase C677T and A1298C polymorphisms and gastric cancer susceptibility. World J Gastroenterol [Internet]. 2014 Aug 28 [cited 2016 Sep 9];20(32):11429–38. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25170232
15. Lv L, Wang P, Sun B, Chen G. The polymorphism of methylenetetrahydrofolate reductase C677T but not A1298C contributes to gastric cancer. Tumour Biol [Internet]. 2014 Jan [cited 2016 Sep 9];35(1):227–37. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23897558
16. Yan S, Xu D, Wang P, Wang P, Liu C, Hua C, et al. MTHFR C677T polymorphism contributes to the risk for gastric cancer. Tumour Biol [Internet]. 2014 Mar [cited 2016 Sep 9];35(3):2123–32. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24122207
17. Chango A, Emery-Fillon N, de Courcy GP, Lambert D, Pfister M, Rosenblatt DS, et al. A Polymorphism (80G->A) in the Reduced Folate Carrier Gene and Its Associations with Folate Status and Homocysteinemia. Mol Genet Metab [Internet]. 2000 Aug [cited 2016 Aug 10];70(4):310–5. Available from: http://linkinghub.elsevier.com/retrieve/pii/S1096719200930348
18. Ferlay J, Steliarova-Foucher E, Lortet-Tieulent J, Rosso S, Coebergh JWW, Comber H, et al. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries in 2012. Eur J Cancer [Internet]. 2013 [cited 2016 Sep 10];49:1374–403. Available from: http://dx.doi.org/10.1016/j.ejca.2012.12.027

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |51
Table 1. Characteristics of the participants: age and gender
Control
n=250
Gastric Atrophy
n=43
Gastric Adenocarcinoma
n=117
Age, years
Mean ± SD 50.80 ± 6.86 59.56 ± 10.49 56.91 ± 11.14
Median (Range) 51 (25 – 64) 59(25 – 78) 59 (25 – 78)
Gender, n
Male 124 (50.4%) 19 (44.2%) 84 (61.3%)
Female 126 (49.6%) 24 (55.8%) 53 (38.7%)
SD – Standard Deviation

CHAPTER 4
Polymorphisms in DNA regulation-associated genes and GC development |53
Table 2. Genotype frequencies for SLC19A1 G80A polymorphism
a adjusted for age and gender; OR- Odds Ratio; CI- Confidence Interval
Table 3. Genotype frequencies for MTHFR C677T polymorphism
MTHFR C677T CC CT TT Tcarrier OR CI (95%) p ORa CI (95%)a pa
Normal 121 (50.8%) 79 (33.2%) 38 (16.0%) 117 (49.2%) 1.000 Reference 1.000 Reference
Lesion/Cancer 64 (35.6%) 85 (47.2%) 31 (17.2%) 116 (64.4%) 1.874 1.260-2.788 0.002 1.693 1.108-2.587 0.015
Lesion 18 (41.9%) 18 (41.9%) 7 (16.3%) 25 (58.1%) 1.436 0.745-2.771 0.322 1.155 0.556-2.400 0.700
Cancer 46 (33.6%) 85 (47.2%) 31 (17.2%) 91 (66.4%) 2.046 1.323-3.165 0.001 1.918 1.209-3.044 0.006
a adjusted for age and gender; OR- Odds Ratio; CI- Confidence Interval
SLC19A1 G80A GG GA AA Gcarrier OR CI (95%) p ORa CI (95%)a pa
Normal 69 (28.6%) 121 (50.2%) 51 (21.2%) 190 (78.8%) 1.000 Reference 1.000 Reference
Lesion/Cancer 39 (21.7%) 85 (47.2%) 56 (31.1%) 124 (68.9%) 0.594 0.382-0.924 0.024 0.551 0.340-0.894 0.016
Lesion 6 (14.0%) 19 (44.2%) 18 (41.9%) 25 (58.1%) 0.373 0.189-0.736 0.006 0.347 0.158-0.762 0.008
Cancer 33 (24.1%) 66 (48.2%) 38 (27.7%) 99 (72.3%) 0.699 0.430-1.136 0.166 0.692 0.410-1.166 0.167

Table 4. Allele frequency of SLC19A1 G80A and MTHFR C677TT in different regions. Adapted from The International HapMap Project (1)
Population Region n SLC19A1 G80A MTHFR C677TT
GG AG AA Gcarrier CC CT TT Tcarrier
HapMap-CEU Europe 224 28.57% 55.36% 16.07% 56.25% 46.90% 44.25% 8.85% 30.97%
HapMap-HCB Asia 86 25.58% 46.51% 27.91% 48.84% 27.91% 41.86% 30.23% 51.16%
HapMap-JPT Asia 172 19.77% 48.84% 31.39% 44.19% 39.53% 48.84% 11.63% 36.05%
HapMap-YRI Africa 226 13.27% 35.40% 51.33% 30.97% 81.42% 18.58% - 9.29%
CEU - Northern and Western European; HCB - Han Chinese in Beijing; JPT - Japanese in Tokyo; YRI - Yoruba in Ibadan, Nigeria


CHAPTER 5


CHAPTER 5
Polymorphisms in DNA regulation-associated genes and GC development |56
FINAL DISCUSSION AND CONCLUSIONS
Gastric Cancer is a major cause of cancer-related deaths worldwide (4). In Portugal, GC
is the fifth cause of death by cancer (4). The majority of patients with GC are diagnosed
in advanced stages, leading to a poor prognosis (37).Therefore, it is important to
implement preventive strategies, such as population screening and genetic susceptibility
tests, to allow the detection of GC at early stages.
In this thesis we have been focused on genetic risk markers of GC susceptibility
associated with two of the risk factors for GC development: altered DNA methylation and
low folate intake. Literature revealed that genes involved in carcinogenesis accumulate
aberrant methylation patterns (26) and additionally, studies have also found an
association between low folate levels and development and invasiveness of GC (22).
Genetic alterations, such as SNPs, have been described as important predictors of
cancer development and a great number of them have the ability to modulate gene
expression. Therefore, the study of SNPs in genes responsible for DNA methylation and
folate metabolism may contribute for a better knowledge of genetic influence on
individual’s susceptibility to GC development.
DNA Methylation
We started by performing a systematic review and meta-analysis to understand what has
been described in literature regarding the role of SNPs in DNMTs and GC development,
in order to attempt to clarify which SNPs could be important to study. From a total of 24
SNPs reported, we found that only 4 SNPs seem to have some association with GC. The
DNMT2 rs11254413, DNMT3A rs7560488 and rs36012910 and, specially, DNMT1
rs16999593 seemed to be good candidates to study the role of SNPs in DNMTs with
GC – figure 5.
The DNMT1 rs16999593 C>T variation results in a missense mutation caused by a His
to Arg amino acid substitution at position 97 of DNMT1 promoter (33). A study performed
by Yang and colleagues revealed that DNMT1 rs16999593 C allele was associated with
an increased risk of GC (OR = 1.45; 95%CI = 1.00-2.11; p = 0.05) (38). Moreover, Jiang
et al. results showed an association of DNMT1 rs16999593 in GC but the results were
not statistically significant (33). Nevertheless, the influence of this SNP on DNMT1
function is still unknown. We hypothesized that DNMT1 rs16999593 variation may have
an effect on structure and function of the protein.

CHAPTER 5
Polymorphisms in DNA regulation-associated genes and GC development |57
Figure 5. SNPs in DNMTs candidates for association with GC.
DNMT1 is responsible for the methylation of newly biosynthesized DNA strands after
replication (39). Literature showed that DNMT1 expression levels are significantly higher
in GC patients (40–42). DNMT1 increased expression levels are associated with
hypermethylation of CGI and, therefore, tumor suppressor genes silencing (43). Genetic
variants in DNMT1 promoter, such as rs16999593, may have an impact in GC
carcinogenesis and contribute to genetic susceptibility to GC.
The literature systematic review also revealed that DNMT2 rs11254413 A allele was
associated to a protective effect against GC (OR = 0.15; 95%CI = 0.08-0.27); p < 0.01)
(38). DNMT2 rs11254413 is characterized by a G>A allele change leading to a His to
Tyr missense mutation in position 101. DNMT2 is responsible for aspartic acid tRNA and
residual DNA (cytosine-C5) methylation, nevertheless, there are needed future studies
to understand the impact of DNMT2 in cellular pathways (44). The results presented by
Yang et al. suggest a potentially protective effect of DNMT2 rs11254413 on GC risk, but
more studies are needed to prove this association, since this was the only study
analyzing an association of DNMT2 rs11254413 with GC.
Finally, regarding DNMT3A literature analysis, we found two SNPs with a statistically
significant association with GC: rs7560488 and rs36012910. The C allele of DNMT3A1
rs7560488 was associated with an increased risk for GC (OR = 1.65; 95%CI = 1.19-2.29;
p = 0.002) (45). DNMT3A rs7560488 T>C variation is a putative functional tagSNP, which
means that represents all SNPs from DNMT3A promoter with high linkage disequilibrium
(35,45). Wu and colleagues also performed assays to understand the impact of this
RISK Protection

CHAPTER 5
Polymorphisms in DNA regulation-associated genes and GC development |58
tagSNP in DNMT3A expression. The results revealed that the expression levels of
DNMT3A1 mRNA was lower in individuals with C allele (p<0.05) (45). DNMT3A
rs7560488 T>C variation have an effect on the binding of transcriptional factors, which
results in an altered transcriptional activity of DNMT3A1 gene (45). Therefore, these
findings suggest that DNMT3A rs7560488 may regulate DNMT3A expression and
contribute for GC susceptibility. Another SNP in DNMT3A statistically significant
associated with GC was rs36012910. Individuals with the G allele of DNMT3A
rs36012910 presented an increased risk for GC when compared to controls (OR = 2.36;
95%CI = 1.35-4.14; p = 0.002) (46). This SNP is localized on the transcription start site
in the promoter region of DNMT3A and is characterized by an A to G allele change (46).
By bioinformatics analyses, Wu and his colleagues hypothesized that DNMT3A is a
functional SNP with impact on the DNMT3A promoter activity, and, therefore, may be
associated with GC (46). Future studies are needed to confirm the biological impact of
DNMT3A rs rs36012910 and the association with GC.
Furthermore, we have attempted to summarize the data in literature with a meta-
analysis, and we found that from the 24 SNPs, only 6 SNPs were evaluated in more than
one study, which clearly reduces the data available for the meta-analysis. Our results
revealed that only rs16999593 was associated with GC, showing statistically significant
increased risk (OR = 1.31; 95% CI = 1.08-1.60; p=0.006) for TC+CC genotypes. Only
two studies evaluate this association, both in Asiatic populations. As described before,
more studies are needed to understand the functional impact of this SNP. Besides, this
results suggest that Chinese individuals carrying DNMT1 rs16999593 C allele have an
increased risk for GC, and more studies with different populations are needed to confirm
the association of DNMT1 rs16999593 with GC.
After the literature systematic review and meta-analysis, we decided to evaluate 3
different SNPs in DNMTs: DNMT1 rs16999593, DNMT2 rs11254413 and DNMT3A
rs7560488. We decided to study one SNP for each DNMT, according to systematic
review and meta-analysis results. All chosen SNPs presented promising associations
with GC and we aimed to characterize the genotyping distribution for the three SNPs in
our population. Unfortunately, we only were capable to studied DNMT2 rs11254413 and
DNMT3A rs7560488 because of technical issues with the assay for the other SNP that
did not allowed us to do a correct interpretation of genotypes and therefore we have
decided to exclude it from the study. Our results revealed no association of both SNPs
with gastric lesions and cancer development. Nevertheless, the findings of our study are
not in agreement with the literature available. Only two studies evaluate the association

CHAPTER 5
Polymorphisms in DNA regulation-associated genes and GC development |59
of DNMT2 rs11254413 and DNMT3A rs7560488 with GC (38,45). The results, also
explored before, showed that DNMT2 rs11254413 A allele may have a protective effect
for GC and DNMT3A rs7560488 C allele may be associated with an increased risk for
GC. The majority of studies regarding SNPs in DNMTs and association with GC were
performed with Asian populations and there are no studies with Caucasian populations.
We found no association with gastric lesion and GC in our population, nevertheless, this
was the first study evaluating the association of DNMT2 rs11254413 and DNMT3A
rs7560488 in a European population and more studies are needed to confirm our
findings.
Folate Metabolism
Finally, we also performed a study to understand the association of SNPs in genes
essential for folate metabolism with gastric lesions and GC. We selected two important
genes that have been studied: SLC19A1 and MTHFR (47,48). Our study demonstrated
an association between SLC19A1 G80A and gastric lesions development and MTHFR
C677T and GC development.
SLC19A1 G80 is associated with a protective effect for gastric lesions (OR = 0.373;
95%CI = 0.189-0.736; p = 0.006), but not with GC. According to literature, there is only
one study evaluating the association of SLC19A1 G80A with GC (49). The results
showed a protective effect of G allele for GC, which is in line with our results although
we do not find a statistically significant association. This is the first study to evaluate the
association of this SNP with gastric lesions.
On the other hand, MTHFR C677T is associated with an increased risk for GC
development (OR = 2.046; 95%CI = 1.323-3.165; p = 0.001 for T allele), but was not
associated with gastric lesions. Up to our knowledge, there is no studies to demonstrate
the association between MTHFR C677T and gastric lesions, still, several studies have
been performed concerning the association of this SNP with GC (50–54). All studies
suggest that T allele is associated with an increased risk for GC development. Therefore,
MTHFR C677T seem like a good biomarker for GC susceptibility.
Folates are B vitamins naturally present in many foods, important for DNA regulation,
including DNA methylation (51). In folate metabolism, SLC19A1 or RFC1 is responsible
for transport of active form of folate 5-MTHF and MTHFR has the capacity to convert
folates in to the reduced and active form 5-MTHF. Without 5-MTHF, there is no
conversion of homocysteine in methionine and, therefore, no formation of SAM, a methyl
group donor crucial for DNA methylation catalysed by DNMTs – figure 6.

CHAPTER 5
Polymorphisms in DNA regulation-associated genes and GC development |60
Figure 6. Homocysteine cycle. The 5-MTHF is a methyl donor for the conversion of homocysteine
into methionine and SAM is a methyl donor for DNA methylation. (5-MTHF – 5-Methylene
tetrahydrofolate; SAM - S-Adenosyl methionine)
Literature showed that individuals with SLC19A1 80AA and MTHFR 677TT genotypes
had significantly lower plasma folate levels and homocysteinemia (high level of
homocysteine in blood), comparing to individuals with wild-type SLC19A1 GG80 and
MTHFR CC677 genotypes (17). The results of Chango and his colleagues suggests that
SLC19A1 G80A and MTHFR C677T influence protein function and, therefore,
carcinogenesis of GC (17).
Conclusions
In conclusion, our study demonstrated that DNMT2 rs11254413 and DNMT3A
rs7560488 are not associated with gastric lesions and GC in our population, but,
otherwise, SLC19A1 G80A and MTHFR C677T seem to have an impact on susceptibility
for gastric lesions and GC, respectively. These results suggest that SLC19A1 G80A and
MTHFR C677T may have a role in gastric carcinogenesis, though this observation needs
to be complemented with further studies, including larger studies with different ethnic
populations and functional studies regarding the effect of both SNPs in gene expression.
Homocysteine
Methionine
SAM
DNA Methylation
5-MTFH


CHAPTER 6


CHAPTER 6
Polymorphisms in DNA regulation-associated genes and GC development |62
REFERENCES
1. Kanwal R, Gupta K, Gupta S. Cancer epigenetics: an introduction. Methods Mol Biol [Internet]. 2015 [cited 2016 Jul 16];1238:3–25. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25421652
2. Stewart BW, Wild C, International Agency for Research on Cancer., World Health Organization. World cancer report 2014. International Agency for Research on Cancer; 2014. 916 p.
3. Ferlay J, Soerjomataram I, Ervik M, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide. IARC CancerBase No 11 [Internet] [Internet]. 2013;Lyon, Fran. Available from: http://globocan.iarc.fr
4. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J cancer [Internet]. 2015 Mar 1 [cited 2016 Jul 16];136(5):E359-86. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25220842
5. Lauren P. The two histological main types of gastric carcinoma: diffuse and so-called intestinal-type carcinoma. an attempt at a histo-clinical classification. Acta Pathol Microbiol Scand [Internet]. 1965 [cited 2016 Jul 26];64:31–49. Available from: http://www.ncbi.nlm.nih.gov/pubmed/14320675
6. Carl-Mcgrath S, Ebert M, Röcken C. Gastric adenocarcinoma: epidemiology, pathology and pathogenesis. Cancer Ther Vol Cancer Ther. 2007;5(5):877–94.
7. Hu B, El Hajj N, Sittler S, Lammert N, Barnes R, Meloni-Ehrig A. Gastric cancer: Classification, histology and application of molecular pathology. J Gastrointest Oncol [Internet]. 2012;3(3):251–61. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3418539&tool=pmcentrez&rendertype=abstract
8. McLean MH, El-Omar EM. Genetics of gastric cancer. Nat Rev Gastroenterol Hepatol [Internet]. 2014 Nov [cited 2016 Aug 17];11(11):664–74. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25134511
9. Karimi P, Islami F, Anandasabapathy S, Freedman ND, Kamangar F. Gastric cancer: descriptive epidemiology, risk factors, screening, and prevention. Am Assoc Cancer Res [Internet]. NIH Public Access; 2014 May [cited 2016 Jul 26];23(5):700–13. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24618998
10. Yoon H, Kim N. Diagnosis and management of high risk group for gastric cancer. Gut Liver [Internet]. Editorial Office of Gut and Liver; 2015 Jan [cited 2016 Jul 26];9(1):5–17. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25547086
11. D’Elia L, Rossi G, Ippolito R, Cappuccio FP, Strazzullo P. Habitual salt intake and risk of gastric cancer: a meta-analysis of prospective studies. Clin Nutr [Internet]. Elsevier; 2012 Aug [cited 2016 Jul 30];31(4):489–98. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22296873
12. Wiseman M. The second World Cancer Research Fund/American Institute for Cancer Research expert report. Food, nutrition, physical activity, and the prevention of cancer: a global perspective. Proc Nutr Soc [Internet]. 2008 Aug [cited 2016 Jul 30];67(3):253–6. Available from: http://www.ncbi.nlm.nih.gov/pubmed/18452640
13. Nagini S. Carcinoma of the stomach: A review of epidemiology, pathogenesis, molecular genetics and chemoprevention. World J Gastrointest Oncol [Internet]. 2012 Jul 15 [cited 2016 Jul 30];4(7):156–69. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22844547
14. Wogan GN, Hecht SS, Felton JS, Conney AH, Loeb LA. Environmental and chemical carcinogenesis. Semin Cancer Biol. 2004;14(6):473–86.
15. Sousa H, Pinto-Correia A-LL, Medeiros R, Dinis-Ribeiro M. Epstein-Barr virus is

CHAPTER 6
Polymorphisms in DNA regulation-associated genes and GC development |63
associated with gastric carcinoma: The question is what is the significance? World J Gastroenterol [Internet]. 2008 Jul 21 [cited 2016 Jul 30];14(27):4347–51. Available from: http://www.ncbi.nlm.nih.gov/pubmed/18666324
16. Fukayama M. Epstein-Barr virus and gastric carcinoma. Pathol Int [Internet]. 2010 May [cited 2016 Jul 30];60(5):337–50. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20518883
17. Fu D-G. Epigenetic alterations in gastric cancer (Review). Mol Med Rep [Internet]. Spandidos Publications; 2015 Sep [cited 2016 Jul 30];12(3):3223–30. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25997695
18. Levine AJ, Lee W, Figueiredo JC, Conti D V., Vandenberg DJ, Davis BD, et al. Variation in folate pathway genes and distal colorectal adenoma risk: A sigmoidoscopy-based case-control study. Cancer Causes Control. 2011;22(4):541–52.
19. Zacho J, Yazdanyar S, Bojesen SE, Tybjærg-Hansen A, Nordestgaard BG. Hyperhomocysteinemia, methylenetetrahydrofolate reductase c.677C>T polymorphism and risk of cancer: Cross-sectional and prospective studies and meta-analyses of 75,000 cases and 93,000 controls. Int J Cancer. 2011;128(3):644–52.
20. Mayne ST, Risch HA, Dubrow R, Chow WH, Gammon MD, Vaughan TL, et al. Nutrient intake and risk of subtypes of esophageal and gastric cancer. Cancer Epidemiol Biomarkers Prev [Internet]. 2001;10(10):1055–62. Available from: http://libaccess.mcmaster.ca/login?url=http://ovidsp.ovid.com/ovidweb.cgi?T=JS&CSC=Y&NEWS=N&PAGE=fulltext&D=med4&AN=11588131\nhttp://sfx.scholarsportal.info/mcmaster?sid=OVID:medline&id=pmid:11588131&id=doi:&issn=1055-9965&isbn=&volume=10&issue=10&spage=1
21. Ulrich CM, Robien K, McLeod HL. Cancer pharmacogenetics: polymorphisms, pathways and beyond. Nat Rev Cancer [Internet]. 2003 Dec [cited 2016 Sep 7];3(12):912–20. Available from: http://www.nature.com/doifinder/10.1038/nrc1233
22. Lee TY, Chiang EP, Shih YT, Lane HY, Lin JT, Wu CY. Lower serum folate is associated with development and invasiveness of gastric cancer. World J Gastroenterol. 2014;20(32):11313–20.
23. Liang S, Zhou Y, Wang H, Qian Y, Ma D, Tian W, et al. The effect of multiple single nucleotide polymorphisms in the folic acid pathway genes on homocysteine metabolism. Biomed Res Int. Hindawi Publishing Corporation; 2014;2014.
24. Zhang D, Wen X, Wu W, Guo Y, Cui W. Elevated homocysteine level and folate deficiency associated with increased overall risk of carcinogenesis: Meta-analysis of 83 case-control studies involving 35,758 individuals. PLoS One. 2015;10(5):1–16.
25. Gotze T, Rocken C, Rohl FW, Wex T, Hoffmann J, Westphal S, et al. Gene polymorphisms of folate metabolizing enzymes and the risk of gastric cancer. Cancer Lett. 2007;251(2):228–36.
26. Qu Y, Dang S, Hou P. Gene methylation in gastric cancer. Clin Chim Acta [Internet]. 2013 Sep 23 [cited 2016 Jan 12];424:53–65. Available from: http://www.sciencedirect.com/science/article/pii/S0009898113001964
27. Hitchler MJ, Domann FE. An epigenetic perspective on the free radical theory of development. Free Radic Biol Med [Internet]. NIH Public Access; 2007 Oct 1 [cited 2016 Jul 29];43(7):1023–36. Available from: http://www.ncbi.nlm.nih.gov/pubmed/17761298
28. Tie J, Zhang X, Fan D. Epigenetic roles in the malignant transformation of gastric mucosal cells. Cell Mol Life Sci [Internet]. Springer International Publishing; 2016 Jul 27 [cited 2016 Aug 16];1–12. Available from: http://link.springer.com/10.1007/s00018-016-2308-9
29. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell [Internet]. Elsevier; 2011 Mar 4 [cited 2016 Jul 26];144(5):646–74. Available from:

CHAPTER 6
Polymorphisms in DNA regulation-associated genes and GC development |64
http://www.ncbi.nlm.nih.gov/pubmed/21376230 30. Calcagno DQ, de Arruda Cardoso Smith M, Burbano RR. Cancer type-specific
epigenetic changes: gastric cancer. Methods Mol Biol [Internet]. 2015 [cited 2016 Jul 16];1238:79–101. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25421656
31. Brookes AJ. The essence of SNPs. Gene [Internet]. 1999 Jul [cited 2016 Aug 17];234(2):177–86. Available from: http://linkinghub.elsevier.com/retrieve/pii/S037811199900219X
32. Haiman CA, Stram DO. Exploring genetic susceptibility to cancer in diverse populations. Curr Opin Genet Dev [Internet]. NIH Public Access; 2010 Jun [cited 2016 Sep 8];20(3):330–5. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20359883
33. Jiang J, Jia Z, Cao D, Jin M-S, Kong F, Suo J, et al. Polymorphisms of the DNA methyltransferase 1 associated with reduced risks of Helicobacter pylori infection and increased risks of gastric atrophy. PLoS One [Internet]. United States; 2012 Jan [cited 2016 Apr 19];7(9):e46058. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3457938&tool=pmcentrez&rendertype=abstract
34. Saradalekshmi KR, Neetha NV, Sathyan S, Nair I V, Nair CM, Banerjee M. DNA methyl transferase (DNMT) gene polymorphisms could be a primary event in epigenetic susceptibility to schizophrenia. PLoS One [Internet]. 2014 Jan [cited 2016 Apr 19];9(5):e98182. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=4032286&tool=pmcentrez&rendertype=abstract
35. Easton DF, Eeles RA. Genome-wide association studies in cancer. Hum Mol Genet [Internet]. 2008 Oct 15 [cited 2016 Sep 8];17(R2):R109-15. Available from: http://www.ncbi.nlm.nih.gov/pubmed/18852198
36. Wooster R, Bachman KE. Catalogue, cause, complexity and cure; the many uses of cancer genome sequence. Curr Opin Genet Dev [Internet]. 2010 Jun [cited 2016 Sep 8];20(3):336–41. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20382522
37. Waddell T, Verheij M, Allum W, Cunningham D, Cervantes a., Arnold D. Gastric cancer+: ESMO-ESSO-ESTRO clinical practice guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2013;24(SUPPL.6).
38. Yang X-X, He X-Q, Li F-X, Wu Y-S, Gao Y, Li M. Risk-association of DNA methyltransferases polymorphisms with gastric cancer in the southern chinese population. Int J Mol Sci. Switzerland; 2012;13(7):8364–78.
39. Hermann A, Goyal R, Jeltsch A. The Dnmt1 DNA-(cytosine-C5)-methyltransferase methylates DNA processively with high preference for hemimethylated target sites. J Biol Chem. 2004;279(46):48350–9.
40. Kanai Y, Ushijima S, Kondo Y, Nakanishi Y, Hirohashi S. DNA methyltransferase expression and DNA methylation of CPG islands and peri-centromeric satellite regions in human colorectal and stomach cancers. Int J Cancer [Internet]. 2001 Jan 15 [cited 2016 Feb 25];91(2):205–12. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11146446
41. Saito Y, Kanai Y, Sakamoto M, Saito H, Ishii H, Hirohashi S. Expression of mRNA for DNA methyltransferases and methyl-CpG-binding proteins and DNA methylation status on CpG islands and pericentromeric satellite regions during human hepatocarcinogenesis. Hepatology [Internet]. 2001 Mar [cited 2016 Feb 25];33(3):561–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11230735
42. Etoh T, Kanai Y, Ushijima S, Nakagawa T, Nakanishi Y, Sasako M, et al. Increased DNA methyltransferase 1 (DNMT1) protein expression correlates significantly with poorer tumor differentiation and frequent DNA hypermethylation of multiple CpG islands in gastric cancers. Am J Pathol. 2004;164(2):689–99.
43. Guo M, Yan W. Epigenetics of gastric cancer. Methods Mol Biol [Internet]. 2015

CHAPTER 6
Polymorphisms in DNA regulation-associated genes and GC development |65
[cited 2016 Jul 16];1238:783–99. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25421692
44. Goll MG, Kirpekar F, Maggert K a, Yoder J a, Hsieh C-L, Zhang X, et al. Methylation of tRNAAsp by the DNA methyltransferase homolog Dnmt2. Science (80- ) [Internet]. 2006;311(5759):395–8. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16424344
45. Wu H, Zhang K, Gong P, Qiao F, Wang L, Cui H, et al. A novel functional TagSNP Rs7560488 in the DNMT3A1 promoter is associated with susceptibility to gastric cancer by modulating promoter activity. PLoS One. United States; 2014;9(3):e92911.
46. Wu Q, Lu S, Wang L, Hu J, Qiao F, Qiu X, et al. DNMT3A rs36012910 A>G polymorphism and gastric cancer susceptibility in a Chinese population. Mol Biol Rep. Netherlands; 2012 Dec;39(12):10949–55.
47. Ganapathy V, Smith SB, Prasad PD. SLC19: The folate/thiamine transporter family. Pflugers Arch Eur J Physiol. 2004;447(5):641–6.
48. DeRoo LA, Bolick SCE, Xu Z, Umbach DM, Shore D, Weinberg CR, et al. Global DNA methylation and one-carbon metabolism gene polymorphisms and the risk of breast cancer in the Sister Study. Carcinogenesis. 2014;35(2):333–8.
49. Wang L, Chen W, Wang J, Tan Y, Zhou Y, Ding W, et al. Reduced folate carrier gene G80A polymorphism is associated with an increased risk of gastroesophageal cancers in a chinese population. Eur J Cancer. 2006;42(18):3206–11.
50. Xiong H-L, Liu X-Q, Sun A-H, He Y, Li J, Xia Y. Aberrant DNA methylation of P16, MGMT, hMLH1 and hMSH2 genes in combination with the MTHFR C677T genetic polymorphism in gastric cancer. Asian Pac J Cancer Prev. Thailand; 2013;14(5):3139–42.
51. Gao S, Ding L-H, Wang J-W, Li C-B, Wang Z-Y. Diet folate, DNA methylation and polymorphisms in methylenetetrahydrofolate reductase in association with the susceptibility to gastric cancer. Asian Pac J Cancer Prev. Thailand; 2013;14(1):299–302.
52. Xia L-Z, Liu Y, Xu X-Z, Jiang P-C, Ma G, Bu X-F, et al. Methylenetetrahydrofolate reductase C677T and A1298C polymorphisms and gastric cancer susceptibility. World J Gastroenterol [Internet]. 2014 Aug 28 [cited 2016 Sep 9];20(32):11429–38. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25170232
53. Lv L, Wang P, Sun B, Chen G. The polymorphism of methylenetetrahydrofolate reductase C677T but not A1298C contributes to gastric cancer. Tumour Biol [Internet]. 2014 Jan [cited 2016 Sep 9];35(1):227–37. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23897558
54. Yan S, Xu D, Wang P, Wang P, Liu C, Hua C, et al. MTHFR C677T polymorphism contributes to the risk for gastric cancer. Tumour Biol [Internet]. 2014 Mar [cited 2016 Sep 9];35(3):2123–32. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24122207

ATTACHMENTS