population genetic structure and natural variation of the model plant, arabidopsis thaliana, in its...
TRANSCRIPT

POPULATION GENETIC STRUCTURE AND NATURAL VARIATION OF THE MODEL PLANT, ARABIDOPSIS THALIANA, IN ITS NATIVE SOUTHERN RANGE EXTREME
AC Brennan, B Méndez-Vigo, A Haddioui,
JM Martínez-Zapater, FX Picó, C Alonso-Blanco

A. thaliana: a model plant for genetics and molecular biology research
Small genome consisting of 5 chromosomes, 125 Mbp, 27,000 genes, 35,000 proteins
Much known about on gene structure, gene products, protein function and expression
Extensive research collections of mutants and wild ecotypes
www.rikenresearch.riken.jp

A. thaliana: a model plant for ecological research?
A small selfing annual of ruderal habitats and openings of natural vegetation
Native to Europe and Central Asia and now found across much of the northern hemisphere
Exhibits both overwintering and spring germination life histories

A. thaliana: a model plant for ecological research?
A small selfing annual of ruderal habitats and openings of natural vegetation
Native to Europe and Central Asia and now found across much of the northern hemisphere
Exhibits both overwintering and spring germination life histories

Recent examples of ecological studies
Hoffmann 2002. J. Biogeography 29: 125-134

Recent examples of ecological studies
Hoffmann 2002. J. Biogeography 29: 125-134

Recent examples of ecological studies
Cao et al. 2011. Nat. Genet. 43: 956-963

Recent examples of ecological studies
Lasky et al. 2012. Mol. Ecol. 21: 5512-5529
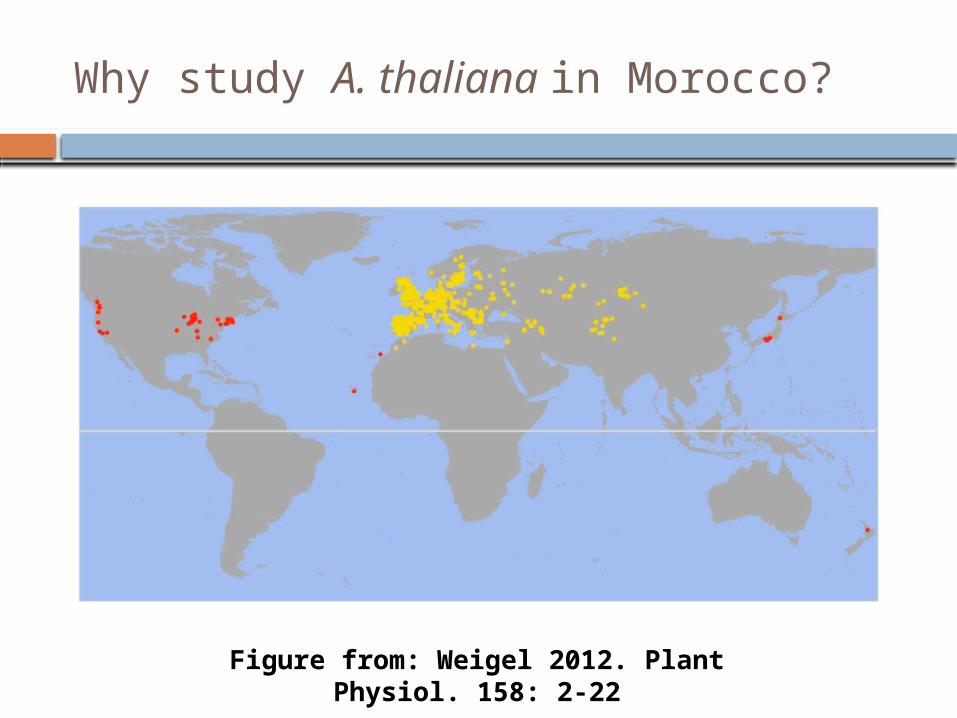
Why study A. thaliana in Morocco?
Figure from: Weigel 2012. Plant Physiol. 158: 2-22

Why study A. thaliana in Morocco?
Figure from: Weigel 2012. Plant Physiol. 158: 2-22
Previously, only three A. thaliana accessions from the whole of North Africa!

Why study A. thaliana in Morocco?
Does A. thaliana in Morocco share a common population history with Iberia?Or was it colonized from Central East Asia?
Figure from: Weigel 2012. Plant Physiol. 158: 2-22

Why study A. thaliana in Morocco?
Natural variation across the whole native latitudinal range.
Figure from: Weigel 2012. Plant Physiol. 158: 2-22

A new collection of Moroccan A. thaliana

A. thaliana samples
472 accessions: selfed seed lines from separate wild-sampled individuals 154 from Morocco (20 different populations) 182 from the Iberian peninsula 136 from the rest of the worldwide range

A. thaliana samples
North Africa Iberia British Isles Central Europe Northern Europe Southern Europe Balkans West Asia Caucasus Central Asia
North America (left panel) Other African (centre panel) East Asia (right panel)

Molecular genetic tools
SNPlex and VeraCode high-throughput single nucleotide polymorphism (SNP) genotyping at CEGEN, (www.cegen.org)
249/343 SNPs produced high quality, polymorphic genotypes for analysis 55 from central Europe 80 from Iberia 114 from worldwide distribution

Genome-wide coverage with single nucleotide polymorphism (SNP) markers
AtMSQTsnp20.1MASC037710.2IP10280105 IP132298500.3AtMSQTsnp91.0SNP1-022110332.2MASC03758 AtMSQTsnp882.3IP1253757802.5MASC039302.8AtMSQTsnp183.9SNP1-042488724.2MASC024754.8AtMSQTsnp265.0IP156291255.6AtMSQTsnp315.9MASC039117.1AtMSQTsnp407.8SNP1-08015166 AtMSQTsnp418.0MASC050298.2AtMSQTsnp458.9MASC053039.4AtMSQTsnp5110.0SNP1-1009592110.1AtMSQTsnp5310.4AtMSQTsnp5410.7MASC0922310.9SNP1-12093455 AtMSQTsnp5812.1IP11354181913.5IP11416282714.2SNP1-16568457 MASC0420916.6AtMSQTsnp6617.0IP11757220217.6MASC0054518.9MASC0375420.0SNP1-2021615720.2MASC0920520.9IP12166935421.7AtMSQTsnp7621.9MASC0417022.3SNP1-2290426622.9AtMSQTsnp8923.8MASC0344724.0AtMSQTsnp9124.1SNP1-2627841326.3AtMSQTsnp10527.3SNP1-2803461328.0AtMSQTsnp10728.2AtMSQTsnp10828.7SNP1-2907617129.1AtMSQTsnp11029.2IP1296964529.7MASC0920630.3
C1
MASC055020.1IP215251700 SNP2-001726680.2NW23476940.3IP20694128 AtMSQTsnp1200.7NW27645280.8MASC054771.1SNP2-195589402.0SNP2-021722342.2NW230534563.1NW246283534.6IP247969694.8SNP2-066542276.6AtMSQTsnp1327.1AtMSQTsnp1347.5SNP2-080418688.0AtMSQTsnp138 MASC027478.4AtMSQTsnp1429.2MASC02600 AtMSQTsnp1439.4MASC0264410.4AtMSQTsnp14510.6MASC0922111.4AtMSQTsnp15011.6SNP2-12296907 NW21229690712.3AtMSQTsnp15512.7AtMSQTsnp159 MASC0544113.3IP21347156513.5AtMSQTsnp16213.7IP21435071714.3MASC0922214.4IP21664068316.6AtMSQTsnp17116.7AtMSQTsnp17216.8IP21705142217.0AtMSQTsnp17717.9SNP2-18143027 AtMSQTsnp18018.1MASC0037118.4AtMSQTsnp18318.7IP21895442918.9NW21957391819.6MASC0281219.7
C2
AtMSQTsnp1870.1AtMSQTsnp2280.2SNP3-00297894 MASC033440.3MASC03001 MASC045231.0IP318041581.8AtMSQTsnp188 MASC053122.1SNP3-022329332.2AtMSQTsnp1943.7SNP3-041948594.2MASC029474.6AtMSQTsnp1974.8AtMSQTsnp1985.2SNP3-06501776 MASC042796.5AtMSQTsnp2036.9AtMSQTsnp2047.3MASC028417.8AtMSQTsnp2078.0SNP3-083470488.3AtMSQTsnp2118.7IP389713839.0AtMSQTsnp2139.3SNP3-1079073210.8IP31139820811.4MASC0426212.5NW31349634513.5NW31476306914.8SNP3-1502819615.0NW31523348315.2IP31638035716.4AtMSQTsnp22516.7SNP3-1727750417.3AtMSQTsnp232 IP31904329519.0AtMSQTsnp23319.1MASC03218 AtMSQTsnp23519.9MASC0278820.7AtMSQTsnp23820.8SNP3-2123008621.2AtMSQTsnp24221.6MASC0921821.8AtMSQTsnp24322.1MASC0492522.6IP32305411923.0AtMSQTsnp24523.2AtMSQTsnp246 MASC0921923.4
C3
MASC046420.1SNP4-004286590.4MASC03275 AtMSQTsnp2780.9AtMSQTsnp283 MASC047251.1AtMSQTsnp2981.5MASC092161.8MASC052581.9SNP4-026115862.6NW426623222.7MASC092252.9NW430069153.0NW44197278 IP441976214.2SNP4-04877304 AtMSQTsnp2584.9IP452917885.3IP457757075.8NW462728076.3AtMSQTsnp2626.9SNP4-070458017.0IP475862817.6AtMSQTsnp2668.1IP487891878.8AtMSQTsnp2799.6MASC032639.8SNP4-1058244510.6AtMSQTsnp28411.2IP41127750811.3AtMSQTsnp28611.6AtMSQTsnp28912.1SNP4-1219617312.2AtMSQTsnp29212.4MASC0921412.7NW41279466912.8NW41310140013.1AtMSQTsnp294 MASC0921513.6SNP4-1407577514.1AtMSQTsnp29514.2MASC03154 AtMSQTsnp29614.7IP41508723315.1AtMSQTsnp300 MASC0400515.8AtMSQTsnp30115.9AtMSQTsnp30416.3IP4160416016.4AtMSQTsnp30616.7SNP4-1744296917.4AtMSQTsnp31518.6
C4
IP500436910.0IP56114900 NW5732970.1SNP5-003760930.4AtMSQTsnp320 MASC001440.5AtMSQTsnp3230.9MASC04860 AtMSQTsnp3251.2AtMSQTsnp3261.4MASC045312.1SNP5-021530682.2AtMSQTsnp3383.0IP532276353.2MASC051274.2AtMSQTsnp3494.7AtMSQTsnp3515.3SNP5-066207366.6AtMSQTsnp3577.0AtMSQTsnp3587.3SNP5-088219688.8AtMSQTsnp3649.7IP598204179.8NW51042785510.4IP51090986410.9MASC0117412.1NW51327236613.3SNP5-1361548313.6MASC0136113.8AtMSQTsnp36814.2MASC04275 IP515065196AtMSQTsnp37015.0
AtMSQTsnp37215.6SNP5-1612280816.1AtMSQTsnp37716.9MASC0312817.2AtMSQTsnp38017.7SNP5-1825853518.2AtMSQTsnp38518.4IP5191238019.1AtMSQTsnp38919.5SNP5-2002178620.0MASC02675 AtMSQTsnp39020.2AtMSQTsnp39320.5SNP5-2231803622.3MASC0439422.4AtMSQTsnp40423.3MASC0154523.4MASC0457624.4MASC0921125.6SNP5-2581995625.8AtMSQTsnp41326.4
C5
One SNP per 0.46 Mbp genome

Strong A. thaliana population structure in Morocco
Percentage genetic group assignment results

Strong A. thaliana population structure in Morocco
Principal components plot and genetic distance neighbour joining tree

Moroccan A. thaliana is genetically similar to an Iberian genetic group
Majority genetic group assignment results
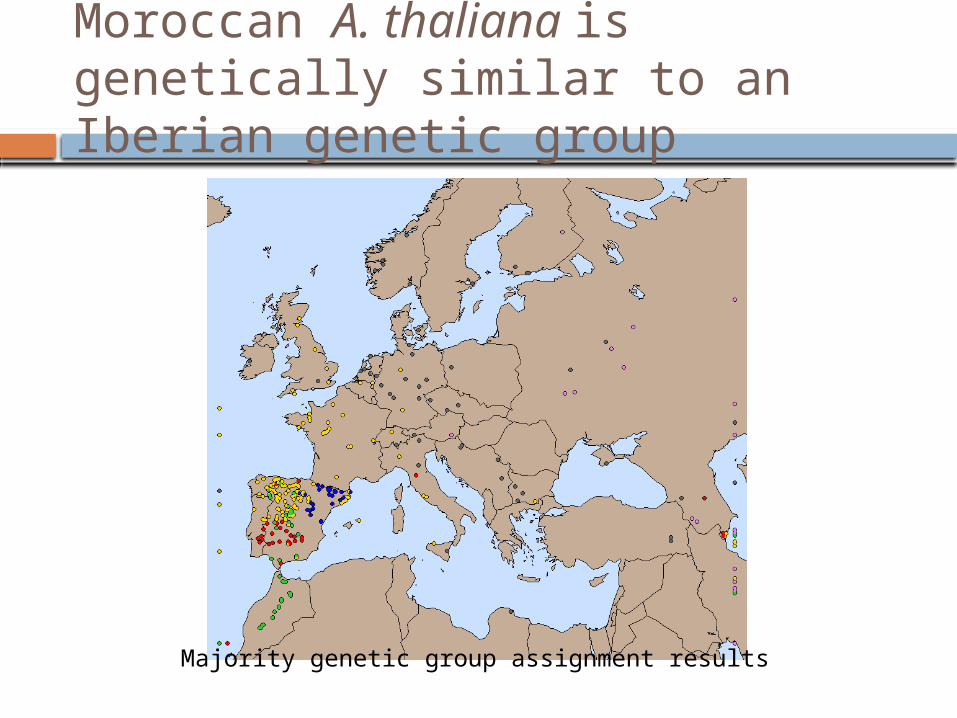
Moroccan A. thaliana is genetically similar to an Iberian genetic group
Majority genetic group assignment results

Stronger isolation by distance in Morocco than Iberia

Expectations for population structure and genetic diversity across the range
Figure from: Hampe & Petit 2005 Ecology Letters 8: 461-467

Changes in A. thaliana genetic diversity across latitude
Additional genotype data from: Platt et al. 2010 PLoS Genet. 6: e1000843

Changes in A. thaliana population differentiation across latitude
Additional genotype data from: Platt et al. 2010 PLoS Genet. 6: e1000843
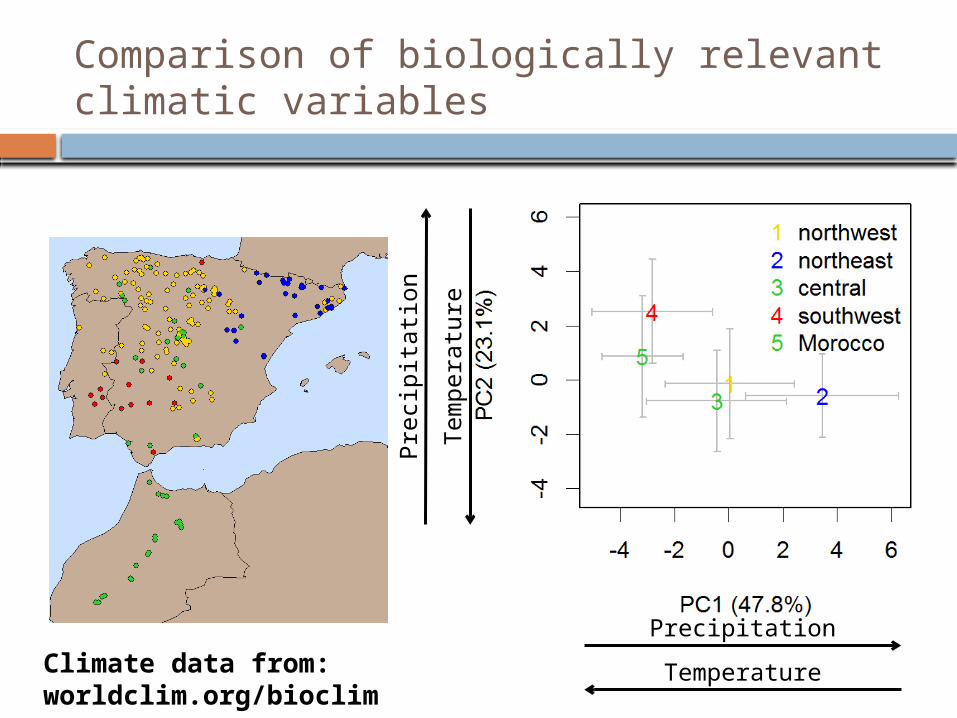
Comparison of biologically relevant climatic variables
Precipitation
Temperature
Pre
cip
itati
on
Tem
pera
ture
Climate data from: worldclim.org/bioclim

Flowering time associations with climate
Filled points indicate climatic significant associations
Climate data from: worldclim.org/bioclim

Flowering time variation matches neutral genetic affinities
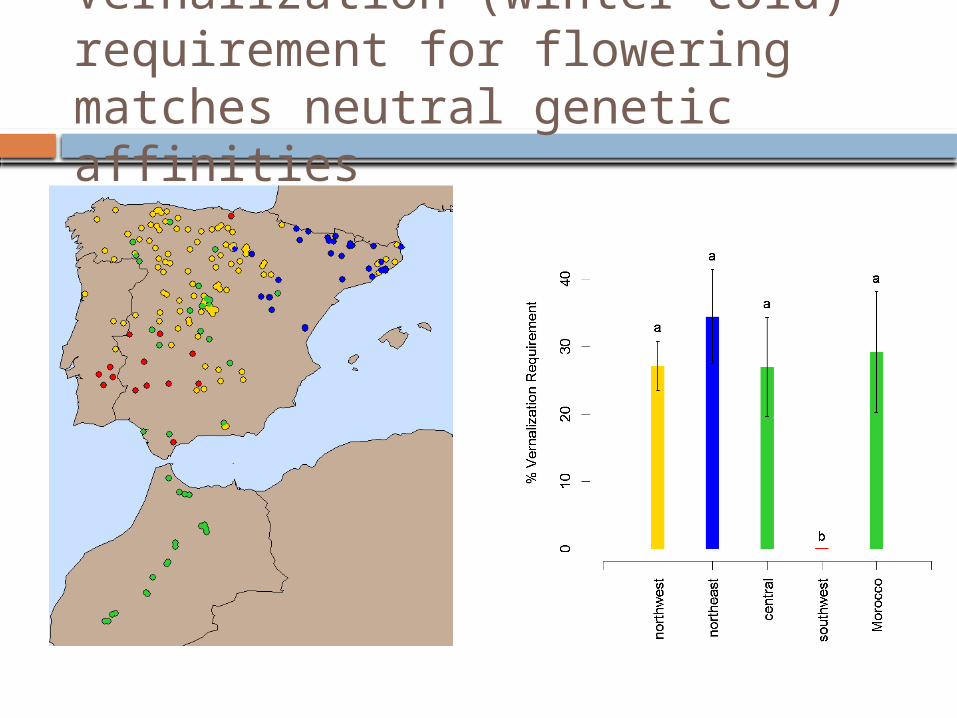
Vernalization (winter cold) requirement for flowering matches neutral genetic affinities

Distribution of functional genetic variation
Functional genetic relationships match neutral genetic variation
PhyC: flowering time response to light

Distribution of functional genetic variation
Functional genetic relationships match climatic similarity
FLC: flowering time, germination, and water use efficiency responses
to temperature

Distribution of functional genetic variation
Functional genetic relationships show aspects of both neutral genetic relatedness and climate similarity FRI: flowering time response to
temperature
CRY2: flowering time response to light

Conclusions: population structure and genetic affinities
A. thaliana in Morocco displayed relatively low but highly structured genetic diversity supporting a native history in North Africa
Morocco is genetically related to the Iberian peninsula indicating a shared demographc history between Southwest Europe and North Africa
Functional genetic variation reflects aspects of both population history and climatic associations

Conclusions: opportunities to study adaptation across the species range New populations representing the
southern trailing edge of A. thaliana’s native range have been genetically characterized
This new resource raises many possibilities for future research of population dynamics, local adaptation, and new functional genetic variation

Acknowledgments
Colleagues and friends at Estación Biológica de Doñana (EBD-CSIC), Sevilla