post-harvest physiology of tetraploid banana fruit: response to storage and ripening
TRANSCRIPT

Ann. appl. Biol. (I974), 78, 193-204 Printed in Great Britain
Post-harvest physiology of tetraploid banana fruit : response to storage and ripening
BY S. NEW AND J. MARRIOTT Tropical Products Institute, 56-62 Gray's Inn Road, London WCI X 8LU
(Accepted 28 March 1974)
SUMMARY
Aspects of the post-harvest physiology relating to storage and ripening of the fruit of tetraploid banana clones resistant to Sigatoka disease, have been compared with fruit of Valery, an important commercial triploid cultivar. Significant differences in susceptibility to low temperature injury, duration of the preclimacteric period, the texture of pulp and peel and ethylene evolution have been found between tetraploid and Valery fruit and also between tetraploid fruit of different clones.
Fruit of Valery and one tetraploid clone developed serious chilling injury during storage at 12 "C whereas that of five other tetraploid clones showed only slight damage. The preclimacteric period for fruit of two tetraploid clones was 30-45% less than for Valery fruit at an equivalent stage of physical development. Pulp firmness of preclimacteric tetraploid fruit was 20-30 % less than that of Valery fruit and the differences persisted through ripening. The softening response to applied ethylene was up to 15 h earlier in fruit of tetraploid clones than of Valery but respiratory patterns, colour development and starch-to-sugar conversion were similar. Unlike Valery fruit, ripe tetra- ploid fruit did not develop senescent spotting, and shelf life was terminated by rapid deterioration of peel strength to a state of severe finger drop. Temporal and quantitative differences occurred between fruit of tetraploid clones and Valery in production of ethylene and these may relate to the observed differences in control of softening in both pulp and peel.
I N T R O D U C T I O N
Banana production in the Caribbean is currently derived from cultivars of the Cavendish subgroup which are resistant to the race of Panama disease which destroyed Gros Michel production but are susceptible to Sigatoka (Leaf Spot) disease. The annual cost of control of Sigatoka disease in Jamaica and the Windward Islands is over EI m, making the development of resistant cultivars very desirable. New cultivars are being bred by crossing commercial triploid bananas with selected diploids and screening of the tetraploid progeny (Shepherd, 1968). Clones are selected first for resistance to Panama disease and Sigatoka disease, then for yield and for the shape and uniformity of the banana bunch. The Banana Breeding Research Scheme in Jamaica has developed a number of promising tetraploid clones from crosses between Highgate (a dwarf mutant of Gros Michel) and synthetic diploids, which are bred from crosses involving wild seeded diploids and edible diploids. The most promising

I94 S. NEW AND J. MARRIOTT tetraploid clones are resistant to Sigatoka and the Gros Michel race of Panama disease and their agronomic performance compares favourably with triploid cultivars of the Cavendish subgroup. As little information was available on those aspects of the post- harvest physiology of tetraploid bananas relating to commercial quality requirements, the present studies were initiated.
The quality requirements for bananas grown in the West Indies for export to Britain are:
(a) Storage life must be sufficient to allow 2-4 days at tropical ambient temperature between harvesting and loading, 10-12 days a t sea under refrigeration and up to I wk between discharge at a port and the initiation of ripening. The fruit must still be preclimacteric when ripening commences, as otherwise synchronization of ripening is impossible. Consequently the preclimacteric phase or 'green life' (Peacock & Blake, 1970) of the bananas needs to be at least 20 days.
(b) It must be possible to obtain uniform and controlled ripening in response to alteration of storage conditions. Most Cavendish cultivars are ripened at 9035 % r.h. by raising their temperature to 18-20 "C and 'gassing' with ethylene to 1000 ppm for 24 h then storing at 16-18 "C. The fruit are marketed when yellow-green; a stage which is reached about 5 days from the start of ripening. The ripening period can be conveniently manipulated by slight changes in temperature. I t is essential that partially ripe fruit should be distributed before softening is complete, so as to avoid undue mechanical damage during handling.
(c) Fully ripe fruit must have a good colour, flavour and texture and these must be maintained for about 3 days.
An early tetraploid clone, ICz, compared favourably with Gros Michel in its shipping and ripening characteristics (Wardlaw, Leonard & Barnell, 1939) but no information was available on these aspects of fruit quality for recently selected tetra- ploid clones.
Fruit supply M A T E R I A L S A N D M E T H O D S
Fruit was supplied from trial plots by the Banana Breeding Research Scheme in Jamaica and by the Windward Island Banana Growers Association in St Lucia, Windward Islands. Although eight tetraploid clones (designated A to H) were used in various aspects of this study, comprehensive data are available for only two clones (A and B). However, the data for other clones are presented as an indication that most of the post-harvest characteristics of the tetraploid clones were similar. These were compared with the triploid Cavendish cultivar Valery. Maturity for harvest was deter- mined by measuring with calipers the diameter of the middle finger of the outer whorl on the second proximal hand on each bunch: this measurement is subsequently termed the caliper grade. Bunches were cut at caliper grade 31-35 mm except for measurement of green life where they were cut at 33-34mm. Normal commercial practice with Valery is to cut at 32-34 mm. After harvest, hands on the bunch were numbered, cut off the stem, washed, dipped in fungicide (400 ppm Thiabendazole or 250ppm Benlate) and packed into cardboard cartons. Fruit was held overnight at a tropical ambient temperature of 2-30 "C and then air-freighted to London where it was normally received within 48 h of harvest.

Post-harvest physiology of tetraploid banana fruit '95
Fruit storage and r2pening Fruit was stored and ripened in cabinets (capacity 20001) at 85-95% r.h.
For storage experiments each bunch was divided into two parts and stored at 12.0 and 140 5 0.5 "C. During ripening fruit colour was assessed on a 1-8 colour index scale (Loesecke, 1949) and at colour index no. 4 (more yellow than green) it was transferred to an air-conditioned room at 20 "C and low humidity. Whenever possible, samples for physical or chemical tests were taken from the third proximal hand of each bunch.
Chilling injury This was assessed by visual inspection of underpeel injury (Stover, 1972) in green
fruit after 10 days storage at low temperature. Injury was scored as nil (o), slight (I), moderate (2) or severe (3). Fruit which was already ripening was excluded from this assessment as turning fruit is more susceptible to chilling than unripe fruit (Simmonds, 1966).
Duration of the preclimacteric phase This was taken as the time from harvest until softening of the pulp was first evident.
Fruit stored at 12 or 14 "C was sampled daily for measurement of the pulp firmness of two fingers. When softening was first recorded it was confirmed after storage for one more day and ripening fruit was then removed from the cabinet. Data from other experiments, not yet published, has shown that rapid softening, which proceeds regularly down the bunch and is complete in about 4 days, coincides with the onset of the climacteric in bananas at these storage temperatures.
Gas evolution Carbon dioxide was measured using an infra-red gas analyser (Series 200, Analytical
Development Co., Hoddesden, Herts) and ethylene using a gas chromatograph (Series 1400, Varian Aerograph, Walton-on-Thames, Surrey) with a flame ionization detector. Fruit was incubated in an air stream of measured rate, but when a flow- through system could not be used because of the need to remove samples for other tests fingers were incubated at the temperature of the ripening cabinet in sealed con- tainers and the rate of gas accumulation calculated.
Firmness measurements A compression force was applied to the tissue at a speed of I cm/min using an
Instron Universal Testing Machine (Model TMI) fitted with a 6 mm diameter probe. For measurements on the pulp, a transverse section I cm thick was cut across each finger, the probe inserted axially into the starchy cortex, avoiding vascular strands, and the maximum force on the force-distance curve recorded. For measurements on peel, a section of peel was cut from the fruit, placed on a platform containing a hole aligned with the probe and slightly larger than it, and the force required to shear a disk out of the peel measured.
7 . 4 P B 78

196 S. NEW AND J. MARRIOTT
Cheinical measurements Moisture content of the pulp was measured by freezing the tissue rapidly with liquid
nitrogen and freeze-drying to constant weight. Alcohol insoluble solids were deter- mined gravimetrically by refluxing the freeze-dried pulp (250 mg) with 80 yo (v/v) aqueous ethanol. The extract was concentrated, clarified with ion exchange resins, and an aliquot inverted with hydrochloric acid (Anon, 1965). Total sugars and reducing sugars in the inverted and non-inverted extracts were determined colorimetrically by reduction of alkaline ferricyanide, using glucose standards (Snell & Snell, 1953).
Analysis of results Bunches of suitable maturity for harvesting were only available regularly for \'alerv
and tetraploid clones A and B; bunches from other clones were received intermittentlv and in differing quantities depending on availability from the experimental plots. Consequently, for analysis of differences between clones, results have been grouped and the standard error of the mean reported irrespective of place of origin or date of harvest. The data presented here are derived from fruit from nine shipments from St Lucia and eight from Jamaica sent between January and September 1972. Differ- ences between Valery and each tetraploid clone and large differences between tetraploid clones have been tested by a t test. Differences have been reported when significant at the level P = 0.05.
Storage life R E S U L T S
During storage at 14.0 ~f: 0.5 "C only Valery showed more than a trace of chilling injury (Table I). At 12.0 k 0.5 "C Valery and one tetraploid clone (F) showed moderate to severe damage and the five other tetraploid clones showed significantly lower mean damage scores (0.6-1.3). The green lives of tetraploid clones A and B were 30-45 "/b less than that of Valery (Table I ) and these differences were highly significant (P = 0.01). Preliminary work with clones C to H suggested that their storage behaviour closely resembled that of clones A and B. Although the tetraploid clones were generally less susceptible to chilling injury, the increase in green life gained by storage at the lower temperature of 12 "C was small. When paired differences for individual bunches stored at 12 "C and 14 "C were analysed the extension in green life of 1.90 days for clone B was significant, but that of 0.56 days for clone A was not.
Spontaneous ripening When tetraploid fruit of any of the clones was incubated at 20 "C in a flow-through
system for measurement of respiration, ripening behaviour was very similar to that of Valery and is typified by that of clone A in Fig. I. The shorter preclimacteric period in the tetraploid fruit is consistent with its behaviour at the storage temperatures. However, after commencement of the climacteric the rates of change in both ethylene and carbon dioxide evolution were similar in the two clones. The rates of coloration were also similar, both clones taking 5 days from the beginning of the climacteric to reach colour index no. 6 (all yellow). The maximum rate of ethylene evolution

Tab
le I
. Ch
illin
g in
jury
, gre
en I
ije, f
irmne
ss a
nd d
ry m
atte
r co
nten
t of
the
culti
var
Val
ery
and
vario
us t
etra
ploi
d ba
nana
clo
nes
Clo
ne
...
Chi
lling
inju
ry s
core
at
12
"C
Chi
lling
inju
ry s
core
at
14 "
C
Gre
en li
fet a
t 12 "
C (
days
)
Gre
en li
fe a
t 14
"C (
days
)
Prec
limac
teri
c pe
el r
uptu
re
forc
e (k
g)
Prec
limac
teri
c pu
lp r
uptu
re
forc
e (k
g)
Prec
limac
teri
c dr
y m
atte
r co
nten
t of
Jam
aica
n fr
uit
(yo f
resh
wei
ght)
Pr
eclim
acte
ric
dry
mat
ter
cont
ent
of S
t Luc
ian
frui
t (y
o fre
sh w
eigh
t)
F G
-
0'0
f 0'
00
(6)
*:
Num
bers
of
bunc
hes
mea
sure
d sh
own
in p
aren
thes
es.
t G
reen
life
mea
sure
d fo
r bu
nche
s at
com
mer
cial
cal
iper
gra
de o
f 33
and
34
mm
.

198 S. NEW AND J. MARRIOTT
1 8
- 6 d 5
- 4 - s - 2
- 0
Tetraploid A
0
Valery
I I I I I - 8 12 16 20 24 8 12 16 20 24
Time (days)
Fig. I . Physiological changes during spontaneous ripening of bananas of two clones. Bananas (about I kg) from the third proximal hand of a bunch incubated at 20 "C in a constant flow of air (about 3 l/h). 0,Ethylene production; 0 , carbon dioxide produc- tion; A , colour index number.
I I I I I I I I I I I 7 8 9 10 11 12 13 14 15 16 17
Days from harvest
Fig. 2. Carbon dioxide production during induced ripening of bananas of different clones transferred from storage at 14 to 16.5 "C eight days after harvest and treated with ethylene (1000 ppm) for 24 h. When fruit ripened to colour index no. 4, it was transferred to storage at 20 "C. Carbon dioxide production (measured by incubation of sample fingers in sealed containers): 0, Valery; 0 , tetraploid clone A ; A, tetra- ploid clone B; 0, colour index number for all clones. Broken lines show carbon dioxide production in fruit transferred to 16.5 "C but not treated with ethylene.

Post-harvest physiology of tetraploid banana fruit 199 was three to four times higher for the tetraploid clone than for Valery but the rates of carbon dioxide evolution were similar between the clones before and during the climacteric.
Ethylene-induced ripening When ripening was induced by treatment with ethylene (1000 ppm) for 24 h at
16.5 "C, the rates of coloration were the same for Valery and for tetraploid clones A and B, and so are represented by a single line in Fig. 2. All clones reached colour index no. 6 eight days after the commencement of the climacteric. The appear- ance of ripe tetraploid fruit was excellent, as they ripened to a deeper yellow than Valery, resembling Gros Michel rather than Cavendish cultivars (Simmonds, 1966). The data on carbon dioxide evolution (Fig. 2) are complicated by the temperature
9 8 7
6 8 5 2 4 3
3 2
7 8 9 10 11 12 13 14 15 16 17 Days from harvest
Fig. 3 . Ethylene production during induced ripening of bananas of different clones. Conditions and symbols as for Fig. 2.
increase which occurred when fruit were transferred from the ripening cabinet to shelf conditions. Except at the climacteric maximum, carbon dioxide production rates were very similar and, at the maximum, the production by Valery was within the range covered by the tetraploid clones. Measurement of climacteric ethylene production was not possible because of release of absorbed ethylene. Post-climacteric evolution of ethylene was similar for each clone until transfer to shelf conditions when the increase in ethylene evolution accompanying senescence was greater in the tetraploid clones, the rate being two to three times that for Valery (Fig. 3).

200 S. NEW AND J. MARRIOTT
Texture changes during r$ening The pulp of firm, preclimacteric tetraploid fruit was 20-30% softer than unripe
Valery pulp (Table I). A small but significant difference occurred between clone F and clones A and B. Fewer data were available from peel measurements but it appears that the peel of tetraploid fruit is marginally softer than that of Valery although the differences for this sample were not quite significant.
*I -16:s OC-1- 19.8 "C - 173.9 "C
I I I I I I I 7 8 9 10 11 12 13 14 15 16 17
9 8
6 3 7,
4 3 3 2
Days after harvest
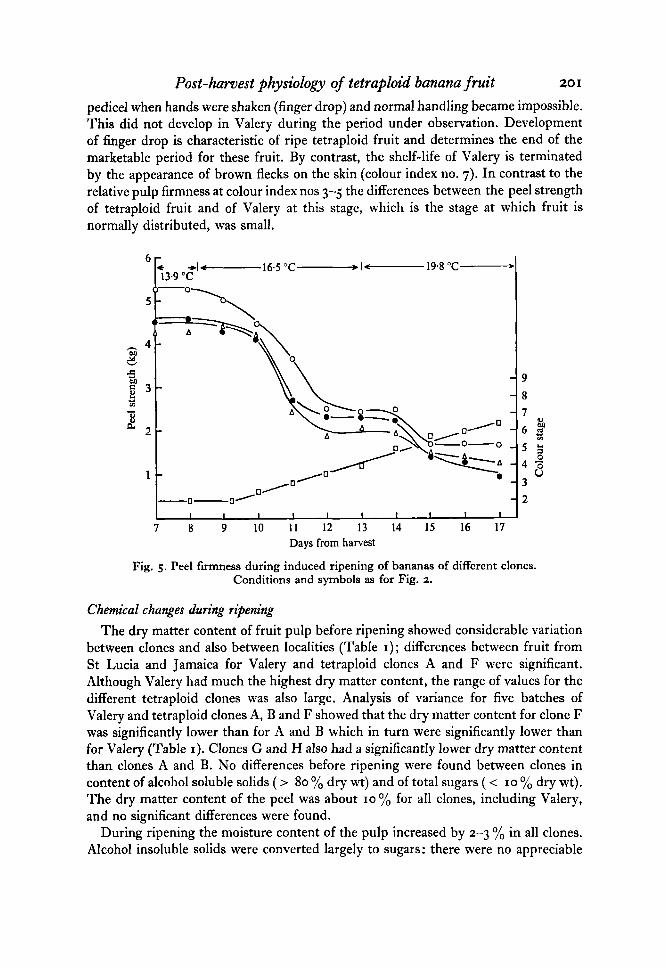
Fig. 4. Pulp firmness during induced ripening of bananas of different clones. Conditions and symbols as for Fig. 2.
Within 20 h of ethylene treatment at 16.5 "C the pulp of tetraploid fruit commenced softening and over a period of 48 h a tenfold reduction in firmness occurred which was almost complete at colour index no. 3, after 3 days ripening (Fig. 4). Valery responded more slowly to ethylene, taking 12-15 h longer to begin rapid softening. At colour index no. 3 the rupture force was two to three times greater in Valery than in tetraploid fruit and softening was not complete until colour index no. 5 , after 6 days ripening. After colour index no. 5 , firmness in all fruit was nearly constant but Valery was 20-30 yo firmer than tetraploid fruit.
The softening of the peel occurred at similar rates for tetraploid fruit and for Valery until colour index nos 5-6, after which the tetraploid fruit continued to soften whilst Valery remained almost constant (Fig. 5) . This change coincided with the develop- ment of much higher ethylene evolution by the tetraploid fruit (Fig. 3). Soon after reaching colour index no. 6 the tetraploid fruit peel became soft enough to break at the
Post-harvest physiology of tetraploid banana fruit 20 I
pedicel when hands were shaken (finger drop) and normal handling became impossible. This did not develop in Valery during the period under observation. Development of finger drop is characteristic of ripe tetraploid fruit and determines the end of the marketable period for these fruit. By contrast, the shelf-life of Valery is terminated by the appearance of brown flecks on the skin (colour index no. 7). In contrast to the relative pulp firmness at colour index nos 3-5 the differences between the peel strength of tetraploid fruit and of Valery at this stage, which is the stage at which fruit is normally distributed, was small.
6 4-16.5 " C - I - 1 9 ~ 8 " C ~
b 3 . 9 "C
I I I I I I I I I 1 I 7 8 9 10 11 12 13 14 15 16 17
Days from harvest
Fig. 5 . Peel firmness during induced ripening of bananas of different clones. Conditions and symbols as for Fig. 2.
Chemical changes during ripening The dry matter content of fruit pulp before ripening showed considerable variation
between clones and also between localities (Table I ) ; differences between fruit from St Lucia and Jamaica for Valery and tetraploid clones A and F were significant. Although Valery had much the highest dry matter content, the range of values for the different tetraploid clones was also large. Analysis of variance for five batches of Valery and tetraploid clones A, B and F showed that the dry matter content for clone F was significantly lower than for A and B which in turn were significantly lower than for Valery (Table I). Clones G and H also had a significantly lower dry matter content than clones A and B. No differences before ripening were found between clones in content of alcohol soluble solids ( > 80 % dry wt) and of total sugars ( < 10 yo dry wt). The dry matter content of the peel was about 10 yo for all clones, including Valery, and no significant differences were found.
During ripening the moisture content of the pulp increased by 2-3 yo in all clones. Alcohol insoluble solids were converted largely to sugars: there were no appreciable
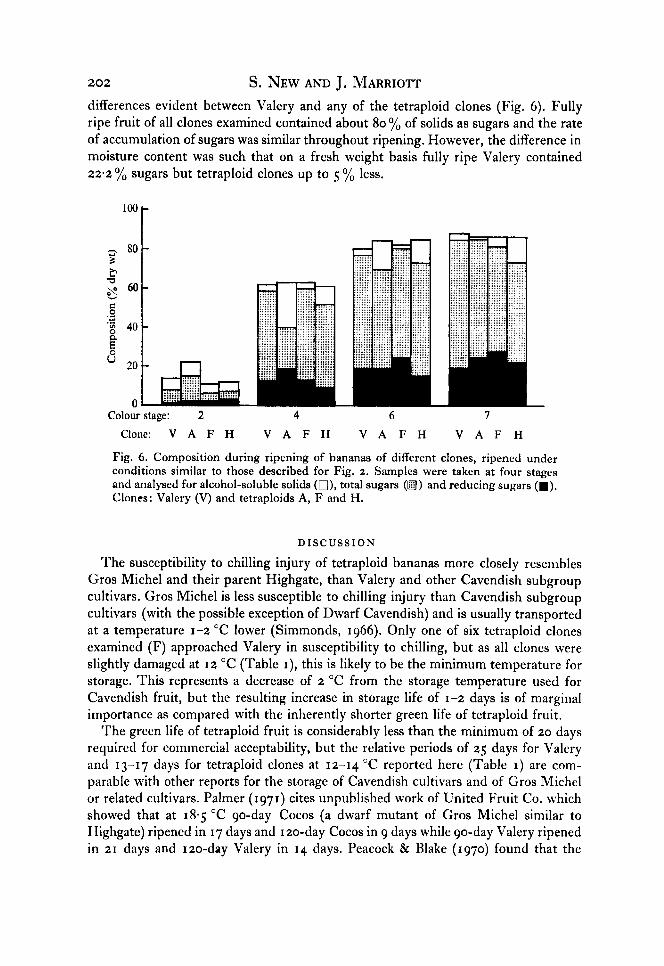
202 S. NEW AND J. MARRIOTT differences evident between Valery and any of the tetraploid clones (Fig. 6). Fully ripe fruit of all clones examined contained about 80 yo of solids as sugars and the rate of accumulation of sugars was similar throughout ripening. However, the difference in moisture content was such that on a fresh weight basis fully ripe Valery contained 22-2 % sugars but tetraploid clones up to 5 yo less.
Coloir stage: 2 4 6 7 Clone: V A F H V A F H V A F H V A F H
Fig. 6. Composition during ripening of bananas of different clones, ripened under conditions similar to those described for Fig. 2. Samples were taken at four stages and analysed for alcohol-soluble solids (a), total sugars (a) and reducing sugars (m). Clones: Valery (V) and tetraploids A, F and H.
D I S C U S S I O N
The susceptibility to chilling injury of tetraploid bananas more closely resembles Gros Michel and their parent Highgate, than Valery and other Cavendish subgroup cultivars. Gros Michel is less susceptible to chilling injury than Cavendish subgroup cultivars (with the possible exception of Dwarf Cavendish) and is usually transported at a temperature 1-2 "C lower (Simmonds, 1966). Only one of six tetraploid clones examined (F) approached Valery in susceptibility to chilling, but as all clones were slightly damaged at 12 "C (Table I), this is likely to be the minimum temperature for storage. This represents a decrease of 2 "C from the storage temperature used for Cavendish fruit, but the resulting increase in storage life of 1-2 days is of marginal importance as compared with the inherently shorter green life of tetraploid fruit.
The green life of tetraploid fruit is considerably less than the minimum of 20 days required for commercial acceptability, but the relative periods of 25 days for Valcry and 13-17 days for tetraploid clones at 12-14 "C reported here (Table I ) are com- parable with other reports for the storage of Cavendish cultivars and of Gros Michel or related cultivars. Palmer (1971) cites unpublished work of United Fruit Co. which showed that at 18.5 "C go-day Cocos (a dwarf mutant of Gros Michel similar to Highgate) ripened in 17 days and 120-day Cocos in 9 days while go-day Valery ripened in 21 days and 120-day Valery in 14 days. Peacock & Blake (1970) found that the

Post-harvest physiology of tetraploid banana fruit 203
green life of Giant Cavendish bananas at 15'5 "C was between 14 and 45 days, depend- ing on maturity. Wardlaw et aE. (1939) found that at 11.5 "C a high proportion of 'heavy three-quarters full' bunches of Gros Michel and the tetraploid ICZ were beginning to ripen after 15 days. It therefore seems likely that the tetraploid clones examined so far have inherited adverse green life characteristics from their triploid parent Highgate.
During both spontaneous and induced ripening the climacteric changes in carbon dioxide and the rates of coloration were similar in fruit of Valery and tetraploid clones (Figs I , z) , and starch-sugar conversion was comparable during induced ripening (Fig. 6). In contrast the quantities of ethylene produced were very different both during the climacteric (Fig. I ) and during senescence (Fig. 3). These results are com- parable with those of Mapson & Robinson (1966) who found that for fruit of Gros Michel and Lacatan the rates of change of ethylene production and absolute rates of carbon dioxide production were similar throughout the climacteric but that Gros Michel fruit produced more ethylene than Lacatan fruit.
The softening of the pulp of tetraploid fruit commences 10-15 h earlier than in Valery fruit (Fig. 4) and this may relate to the much greater endogenous production of ethylene at this time (Fig. I). Similarly during the post-climacteric period the peel of tetraploid fruit continued to soften whilst that of Valery fruit became constant (Fig. 5 ) and this divergence coincided with an increase in production of ethylene by the tetraploid fruit to three times the rate found in Valery fruit (Fig. 3). Respiration and softening are known to be differently affected by ethylene treatment in Anjou pears (Wang, Mellenthin & Hansen, 1972) and it is possible that there is a causal relationship in bananas between the rates of endogenous ethylene synthesis and of texture changes.
Although rapid softening of the pulp is not desirable, the peel probably plays a more important role in the physical protection of thc fruit during commercial handling. Differences in the peel strength of Valery and the tetraploids are small at the distribu- tion stage and handling problems are not apparent on the laboratory scale until the fruit becomes senescent. When scored by taste panels for both texture and overall acceptibility (G. E. Howard & J. Baldry, personal communication), all tetraploid clones were scored softer than Valery but this was not the over-riding organoleptic consideration of the panel, which scored two tetraploids equal to Valery on a hedonic scale. Compositional data (Fig. 6) show that starch-sugar conversion in tetraploid fruit is neither slower nor less complete than in Valery but that ripe tetraploid fruit have a higher moisture content and so may taste less sweet.
The tetraploid clones examined were all derived from crosses between Highgate and a single diploid so that the differences observed between these tetraploids and the triploid Valery were not necessarily caused by the difference in ploidy. Consistent differences do exist between these tetraploid clones and Valery in green life, chilling susceptibility, pulp strength, peel strength in senescence and pulp dry matter content although most of the physiological changes during ripening were similar. The ripe fruit of all clones tested had satisfactory colour and texture, removal of starch was similar in all clones and the flavour of some of the clones was as acceptable as Valery (G. E. Howard & J. Baldry, personal communication). Before commercial shipment of

204 S. NEW AND J. MARRIOTT fruit of any tetraploid clone is feasible it will be necessary to extend green life, either by selection of an improved clone or by modification of the storage method, possibly using sealed packs to create a modified atmosphere (Scott et al. 1971). Extension of green life is essential because if the climacteric begins in storage, ripening cannot be controlled and fruit is over-ripe and very soft at the end of normal ripening. Develop- ment of a ripening schedule which will maintain the strength of the peel to give an adequate shelf-life is also essential. However, the shelf-life of bananas is sensitive to manipulation of temperature and humidity during ripening (Hicks, 1934; Anon, 1964) and it is possible that considerable improvement in firmness will result from use of optimum conditions.
We thank K. Shepherd and E. A. Dixon of the Banana Breeding Research Scheme, research staff of the Banana Board, Jamaica, J. Seeyave of WINBAN Research Scheme and 0. J. Burden of TPI/WINBAN Fruit Quality Scheme, St Lucia for provision of the experimental material and Mrs H. C. Aldridge, T. Powlesland, Miss Z. Vatcher and A. Vellacott for technical assistance.
R E F E R E N C E S
ANON (1964). In Banana ripening manual. Boston, Mass. : United Fruit Sales Corporation. ANON (1965). In Oficial methods of analysis, Association of Official Agricultural Chemists, 10th
HICKS, E. W. (1934). Finger dropping from bunches of Australian Cavendish bananas.
LOESECKE, H. VON (1949). Bananas, p. 108. New York: Interscience. MAPSON, L. W. & ROBINSON, J. E. (1966). Relationship between oxygen tension, biosynthesis of
ethylene, respiration and ripening changes in banana fruit. Journal of Food Technology I, 215-225.
PALMER, J. K. (1971). In The biochemistry of fruits and their products, vol. 2, p. 74. Ed. A. C. Hulme. London: Academic Press.
PEACOCK, B. C. & BLAKE, J. R. (1970). Some effects of non-damaging temperatures on the life and respiration behaviour of bananas. Queensland Journal of Agriculture and Animal Sciences 27, 147-168.
SCOTT, K. J., BLAKE, J. R., STRACHAN, G., TUGWELL, B. L. & MCGLASSON, W. B. (1971). Transport of bananas at ambient temperatures using polyethylene bags. Tropical Agriculture (Trinidad) 4, 245-254.
SHEPHERD, K. (1968). Banana Breeding in the West Indies. Pesticides Abstracts Section B 14, 370-379.
SIMMONDS, N. W. (1966). In Bananas, p. 411. London: Longmans. SNELL, F. D. & SNELL, C. T. (1953). Colorimetric methods of analysis, vol. 3, p. 204. Princeton:
Van Nostrand. STOVER, R. H. (1972). In Banana, Plantain and Abaca diseases, p. 279. England: Common-
wealth Mycological Institute. WANG, C. Y., MELLBNTHIN, W. M. & HANSEN, E. (1972). Maturation of ‘Anjou’ pears in rela-
tion to chemical composition and reaction to ethylene. Journal of the American Society of Horticultural Scientists 97, 9-1 2.
WARDLAW, C. W., LEONARD, E. R. & BARNELL, H. R. (1939). Metabolic and Storage investiga- tions on the banana. I C T A Low Temperature Research Station Memorandum, no. I I, p. 42.
edition, pp. 88 and 424. Washington, D.C.
Australian Journal of the Council for Scient$c and Industrial Research 7, 165-168.