postnatal addition of satellite cells to parasympathetic neurons
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 375:518-525 (1996)
Postnatal Addition of Satellite Cells to Parasympathetic Neurons
SCOTT L. POMEROY, DAVID ZURAKOWSKI, SOMMALA KHOXAYO, MIRE1 ENDARA, AND PIETER DIKKES
Departments of Neurology (S.L.P., S.K., M.E., P.D.) and Research Computing (D.Z.), Children’s Hospital, Harvard Medical School, Boston, Massachusetts 02115;
Departments of Anatomy and Neurobiology and Neurology and Neurosurgery, Washington University School of Medicine, St. Louis, Missouri 631 10 (S.L.P.)
ABSTRACT We have examined the postnatal development of satellite cells associated with parasympa-
thetic neurons of mouse salivary duct ganglia. The number of satellite cells associated with each neuron was found to increase during the first 8 weeks after birth but remained constant thereafter. This corresponds to the period of maximal growth of the salivary gland that serves as the target organ innervated by these neurons. At all ages examined, the number of satellite cells associated with each neuron was found to be highly correlated with neuronal volume. The development of satellite cells associated with individual identified neurons was followed directly by in vivo video microscopy over several months, and the number of satellite cell nuclei was found to increase in regions of the neuronal surface with increasing numbers of synaptic boutons. These results indicate that the postnatal addition of satellite cells to parasympathetic neurons is linked to neuronal enlargement and that synaptic remodeling occurs in concert with satellite cell development. D 1996 Wiley-Liss, Inc.
Indexing terms: autonomic ganglion, satellite cells, glia, mice, synaptic remodeling
Neurons and glial cells that make up the peripheral nervous system must undergo a variety of changes during postnatal development to accommodate an increasing body size. Because their number does not increase significantly after birth, peripheral neurons respond to an expanding field of innervation by growth of the neuronal cell body and extension of a dendritic arbor (Voyvodic, 1987, 1989). Relatively little is known about the postnatal development of satellite cells that ensheathe peripheral neurons (Ga- bella, 1976; Pannese, 1981; Peters et al., 1991). During embryonic development, glial precursors arise from cells that have migrated from the neural crest. These cells become committed to the nonneuronal lineage as early as E13.5 in rats, but continue to proliferate in clusters of glial progenitors after the peak of neurogenesis at E15-16 (Hall and Landis, 1991, 1992). They become associated with neurons during late fetal development. It is not known, however, whether the number of satellite cells associated with peripheral neurons continues to increase in the postna- tal period, nor how the final number of glial cells is regulated. Because peripheral neurons and synaptic connec- tions are invariably encased by glial cells, this issue is of central importance in understanding the cellular basis of postnatal neural development.
In this study we examined the postnatal development of satellite cells in parasympathetic ganglia that innervate the
submandibular salivary glands of mice. There are several reasons for choosing to study this ganglion. First, the neurons in this ganglion have no dendrites (Purves and Lichtman, 19871, allowing accurate identification of all satellite cells associated with each neuron. This, in addi- tion, simplifies neuronal measurement, because there are no dendrites contributing to neuronal volume. The ganglia are also readily accessible for in vivo videomicroscopy, allowing direct observation of satellite cell development. Because our earlier work demonstrated that synaptic bou- tons are localized near satellite cell nuclei (Pomeroy and Purves, 1988), we viewed synapses and satellite cells associ- ated with identified neurons over an interval of 2-3 months to determine whether synaptic remodeling might be related to satellite cell development.
Our results show that satellite cells continue to be added to parasympathetic neurons before 8 weeks of age. After that, the number of glia per neuron is constant. Further- more, the number of satellite cells per neuron is highly correlated with the volume of the associated neuron. We
Accepted July 2,1996 Address reprint requests to Scott L. Pomeroy, Department of Neurology,
Children’s Hospital, 300 Longmood Avenue, Boston, MA 02115. E-mail: Pomeroy(aA1.TCH.Hard Edu
o 1996 WILEY-LISS, INC.

SATELLITE CELL DEVELOPMENT 519
propose that the addition of glial cells to parasympathetic neurons is regulated by neuronal growth.
MATERIALS AND METHODS Quantitation of satellite cells and neurons
Male mice of known age (CF-1 strain; 1-16 weeks old) were anesthetized (pentobarbital, 50 mg/kg, i.p.) and per- fused by intracardiac injection of saline followed by 4% paraformaldehyde in phosphate-buffered saline. The sub- mandibular salivary duct and accompanying ganglia were removed and postfixed in the same fixative for 4 hours. Serial paraffin sections (14-16 p,m) were prepared and stained with cresyl violet. All neuronal and satellite cell nuclei within each ganglion were viewed with a x50 objective (Leitz 1.0 NA) and counted. The counts obtained for each ganglion were corrected according to the method of Abercrombie (1946). Pilot studies were performed to deter- mine the average nuclear diameters of both neurons and satellite cells at each of the postnatal ages. Serial paraffin sections were then cut at a thickness approximately 1.6 times the diameter of the neuronal nucleus diameter at each age. To corroborate the evidence obtained from serial sections, the number of satellite cells per neuron was also determined by counting the number of satellite cell nuclei associated with parasympathetic neurons in salivary duct ganglion whole mounts. Thus, in another set of animals, fresh whole mounts of ganglia were prepared from animals (1, 12, and 99 weeks old) anesthetized with pentobarbital. Neurons on the surface of these ganglia were viewed with a x 100 objective (Leitz 1.2 NA) using differential interfer- ence contrast. The maximum length and width of each neuron were measured with an ocular micrometer, and the number of associated satellite nuclei was determined by counting them while focusing up and down through the entire depth of the neuron (Pomeroy and Purves, 1988).
Microscopy of synaptic boutons and satellite cell nuclei in vivo
Male mice (CF-1 strain; 25-30 g) were used for all experiments. To stain synaptic boutons, 0.2 ml of 1 mM 4-(4-dimethylaminostyryl)-N-methylpyridinium iodide (4- Di-1-ASP; Molecular Probes Inc., Eugene, OR) in saline was injected into a tail vein. After 30-45 minutes, the mice were anesthetized (0.5 mgikg diazepam and 1.5 mg/kg ketamine hydrochloride i.p.), the right submandibular and sublingual salivary glands and ducts exposed, and the salivary duct parasympathetic ganglia viewed with a ~ 5 0 objective (Leitz 1.0 NA) as described previously (Purves et al., 1987; Pomeroy and Purves, 1988). Brightfield video images (Dage-MTI Newvicon camera) of cells and fluores- cence video images (Dage-MTI SIT camera) of synaptic boutons were analyzed on an image processing system (Trapix 5500; Recognition Concepts, Inc., Incline Village, NV) driven by a MicroVAX I1 computer. A series of brightfield and fluorescent focal planes through the depth of the neuron (approximately 20-25 p,m) was obtained and stored, and the position and number of glial cell nuclei were noted.
To view the same cells again, the procedure was repeated 2-3 months later. The interval between observations ranged from 62 to 113 days (mean 93 days). Over this time, the mice grew substantially, their average weight increasing from 28.9 f 2.9 g (mean * S.D.) to 34.7 ? 3.0 g. Reference images from the first viewing were used to locate the cells of
Fig. 1. Cresyl violet stained 15-km-thick cryostat section of a salivary duct ganglion from a 3-week-old mouse. Neuronal nuclei (small arrows) are larger than the satellite cell nuclei (large arrows), which are located on the perimeter of the ganglion neurons. Scale bar = 10 Fm.
interest. A second set of images was then obtained and stored for later comparison to the original images. In total 50 neurons from 44 mice were successfully studied in this way. Subsequent analysis was performed with the aid of the image processing program IMAGR (Voyvodic, 1986). Maps of satellite cell nuclei and synaptic boutons were made and compared as described in Figure 5.
Statistical analysis and modeling Statistical comparisons of means were made by two-
tailed Student’s t tests and paired t tests using a Bonferroni correction for multiple comparisons (Snedecor and Coch- ran, 1980). Poisson regression was used to determine whether the number of satellite cells could be estimated from neuronal size or postnatal age. In the model fitting process, the number of satellite cells was specified to follow an exponential distribution with a log link function. Goodness of fit of the model was assessed by Pearson’s x2 statistic (McCullagh and Nelder, 1995). The statistical modeling was performed using the GENMOD procedure in SAS (Release 6.10; SAS Institute, Inc, Cary, NC).
RESULTS The satellite cell sheath of salivary duct ganglion neurons
is one cell thick and cannot be resolved by light microscopy except for the glial nucleus and perinuclear cytoplasm (Pomeroy and Purves, 1988). Because the boundaries be- tween the glial cells cannot be resolved by light microscopy, quantitative estimates are made by counting the number of satellite cell nuclei (see Fig. 1). Therefore, these estimates must be considered to be maximum possible values, because the estimates cannot take into account the possibility of multinucleate satellite cells.
Numbers of satellite cells per parasympathetic neuron increase with postnatal age
Satellite cell and neuronal nuclear profiles were counted in serial sections of parasympathetic ganglia, defined as discrete clusters of neurons and glia adjacent to the salivary duct, from male mice 1 week (10 ganglia, three mice), 3 weeks (21 ganglia; three mice), 8 weeks (14 ganglia; three mice), and 16 weeks (14 ganglia; three mice) after birth. We

520
Postnatal age (weeks) 0
S.L. POMEROY ET AL.
1[
I I I I I I I 1
IT
.15 - A 0) 3 .10 - Gl 6 .05 - E
Y
*
.I
0
U I I I I I I I I I
0 2 4 6 8 10 12 14 16
I
3 , I I I I I I 1
A 40
A 30
B : I €
I
counted nuclear profiles for 1,445 neurons and 1,253 satellite cells from parasympathetic ganglia of 1-week-old mice, 5,714 neurons and 6,367 satellite cells from 3-week- old mice, 1,680 neurons and 2,911 satellite cells from 8-week-old mice, and 1,874 neurons and 3,668 satellite cells from 16-week-old mice. In total, 24,912 nuclear profiles were counted.
After the counts were corrected (Abercrombie, 19461, the number of satellite cell nuclei per neuronal nucleus was calculated for each ganglion. The ratio of satellite cells to neurons (Fig. 2) increased during postnatal development, reaching a plateau by 8 weeks of age. The increases of ratio between 1 and 3 weeks, as well as between 3 and 8 weeks, are of high statistical significance (Student’s t test with Bonferroni correction for multiple comparisons; P < .001). There was no difference detected between the ages of 8 weeks and 16 weeks (P > .35). Thus, the number of satellite cell nuclei for each neuronal nucleus in the gan- glion increased during the first 8 weeks of postnatal life. This corresponds to the interval of greatest growth of the salivary gland innervated by the parasympathetic neurons as well as of the entire animal (Fig. 3). The central nervous system of this species parallels this growth pattern as well (Pomeroy et al., 1990).
Number of satellite cells per neuron is directly correlated with neuronal size
To determine how satellite development is related to neuronal growth, we asked next whether the increasing ratio of satellite cells to neurons during postnatal develop- ment reflected a net gain of glia per neuron and how the number of satellite cells was related to neuronal size. The number of satellite cells associated with a neuron was counted after measuring the longest neuronal length and width in whole mounts of parasympathetic ganglia. By using these dimensions, an index of neuronal size was derived by calculating the volume of a spheroid with the following equation (Selby, 1969):
€
neuron was thus found to increase during postnatal devel- opment in a pattern similar to that demonstrated in counts taken from serial sections. Furthermore, as is shown in Figure 4, at all ages the number of satellite cells associated with a neuron was highly correlated with the neuronal size index. Small neurons had the least number of satellite cells, whereas increasing numbers of glia were found on succes- sively larger neurons. Likelihood ratio statistics revealed that the neuronal size index was the most significant predictor of satellite cell number (x2 = 19.26, 1 df, P < .0001), although postnatal age predicted the number of satellite cells associated with each neuron as well (9 = 10.81, 2 df, P = .004). Further analysis revealed a two-way interaction between neuronal size index and postnatal age (interaction test, x2 = 6.80, 2 df, P = .033), indicating that the addition of satellite cells to neurons of increasing size varies according to the postnatal age of the animal.
V = 4/3rrab2
where a and b are the measurements in microns of the maximal length and width of the neuron.

SATELLITE CELL DEVELOPMENT 521
160 140 1 .% p1 1004 T
1 2 3 1 1 2
I 2: 3
I
E 4 5
1 wk 12 wks
rf 1 2
r
3 4 5 99 wks
Number of satellite cells per neuron
Fig. 4. Association of satellite cell number with neuronal size. The number of satellite cells associated with each neuron was determined from fresh whole mounts of ganglia obtained at postnatal ages 1, 12, and 99 weeks. The number of satellite cells associated with each neuron was determined while focusing through the depth of the neuron. Dimensions of the neuron were then determined with an ocular micrometer. Satellite cell number is highly correlated with neuronal size, denoted here by an index calculated from the maximum length and width of the neuron (see text). The number at the base of each bar indicates the number of neurons in that category. The error bars indicate 1 S.D.
Synaptic bouton remodeling reflects the addition of satellite cell nuclei
In our previous investigations (Pomeroy and Purves, 19881, we determined that satellite cells change in position and number over time on the surface of salivary duct ganglion neurons and that synaptic boutons are clustered near the nucleus of satellite cells. Considered together, these findings predict that the pattern of synaptic boutons should change to reflect the addition of satellite cells during the postnatal development of salivary duct ganglia. Satellite cells and synaptic boutons associated with identified neu- rons were observed in situ over several months using in vivo video microscopy. At the initial viewing, the number of satellite cells associated with each neuron was determined by counting satellite nuclei while focusing through the depth of the neuron. The procedure was repeated when the same neuron was viewed 2-3 months later. Over this interval, 21 of 50 (42%) neurons gained satellite cells, and 28 (56%) had no change in number. Only one neuron lost a satellite cell. As a result of the additions, the average number of satellite cells per neuron increased significantly from 2.1 -+ 0.8 (mean k S.D.) to 2.6 2 1.0 (paired t test; P < .001) over the interval between observations.
At the same time when satellite cell imaging was done, synaptic boutons stained with the vital fluorescent dye 4-di-1-ASP were also viewed. When maps of synaptic bou- tons were superimposed on those of glial nuclei, regions of the salivary duct ganglion neuron surface that had gained glial nuclei also had increased synaptic bouton staining, whereas those regions with fewer nuclei had less bouton staining (Figs. 5,6; Table 1).
DISCUSSION In contrast to the case with most neurons, the number of
glial cells in the nervous system increases during postnatal development (Dobbing and Sands, 1973; Volpe, 1995). Although the increase of glial number contributes signifi- cantly to growth of the nervous system, the cellular mecha- nisms regulating glial number after fetal development are not known. Here, we demonstrate that a postnatal increase of satellite cells occurs in parasympathetic ganglia during the period of maximal somatic growth of the animal and that the number of glial cells per neuron is highly correlated with neuronal size.
Neuronal growth has been shown by several studies to be regulated by an increasing target size. Investigations of the effects of limb bud ablation in developing chick embryos or of tail regeneration in reptiles demonstrated that neurons atrophy or enlarge in direct proportion to their target of innervation (Hamburger and Levi-Montalcini, 1949; Terni, 1920). Homologous neurons of increasingly larger animals have been found to extend more complex dendritic arbors that can accommodate a greater number of innervating axons (Purves and Lichtman, 1985; Purves et al., 1986; Snider, 19871, and motor neuron growth has been shown to be coupled to enlargement of myofibers (Balice-Gordon and Lichtman, 1990). Voyvodic (1989) experimentally altered the relative target size of rat superior cervical ganglion neurons and demonstrated that the neurons had increased cell body diameter and dendritic complexity after expansion of their target size but had decreased neuronal size and dendritic complexity with smaller relative target size. Tar- get size has been hypothesized to regulate neuronal size and dendritic complexity through target derived trophic sup- port, ultimately influencing the convergence of synaptic inputs to efferent neurons of the peripheral nervous system (Purves, 1988). Similar mechanisms might be operative in the development of synaptic connectivity of the central nervous system as well (Pomeroy et al., 1990; Purves, 1994; Purves and LaMantia, 1993; Riddle et al., 1992).
Other investigations have linked neuronal size with satellite cell number. Neuronal measurements and quanti- tative estimates of satellite cells associated with dorsal root ganglion neurons have demonstrated that satellite cell number is directly proportional to neuronal volume and surface area (Humbertson et al., 1969; Pannese, 1960; Pannese et al., 1972). This relationship is also retained for homologous neurons across species (Pannese et al., 1972). In principle, it is possible that glial cells determine neuronal size. The fact that target size also governs neuronal size suggests that this would have to be mediated either by a soluble signal from the target to the glial cells or by a signal transferred to glia through the neuron.
A neuron-derived signal, glial growth factor (GGF), that regulates glial survival and proliferation in the peripheral nervous system has been described (Gassmann et al., 1995; Lee et al., 1995; Meyer and Birchmeier, 1995; Verdi et al., 1996). A member of the epidermal growth factor superfam- ily of peptide growth factors known collectively as the neuregulins, GGF serves as a ligand for the receptor tyrosine kinases p185erbB2/HER2 and p180erbB4/HER4 (Pe- les et al., 1992; Plowman et al., 1993). The growth factor, expressed by neurons as a membrane-associated protein, has been found to promote Schwann cell proliferation and survival, or skeletal muscle development, by activation of receptors that are expressed primarily by glia or muscle

Fig. 5. Concurrent localization of glial nuclei and synaptic boutons. A Representative brightfield image demonstrating a single focal plane of a parasympathetic ganglion neuron and associated glial nuclei (arrows). B: Representative enhanced fluorescent image shows the same parasympathetic neuron with associated synaptic boutons that have been stained with 4-Di-1-ASP. C: The image from the focus- through series of brightfield images with the largest cross-sectional area was traced with a digitizing tablet, and a reference guide was made by using the centerpoint of the neuron and a point outside the neuron. Glial nuclei from all focal planes were then traced. In this case, a glial nucleus from a deeper focal plane is not evident in the more superficial focal plane shown in A. D For synaptic bouton maps, a guide was made by using the same points of reference. Primary fluorescent images (B) were processed by a contrast-detection convolution, which maps areas
of maximum fluorescent intensity (Purves et al., 1987). The staining patterns from all focal planes were combined and displayed graphically along with the reference guide. In this example, the pattern of fluorescent staining shown in B was combined with patterns from five other focal planes to create a synaptic bouton map (displayed in red). E The patterns of synaptic boutons and glial nuclei were then compared by superimposing the maps and computing the proportion of pixels in each reference quadrant occupied by the map depicting fluorescent bouton staining. The numerals beside each quadrant indicate the proportion of pixels representing boutons. Boutons outside the cell boundary, defined by tracing the brightfield neuron image with the largest cross-sectional area, are not included in this calculation. Scale bar = 10 pm.
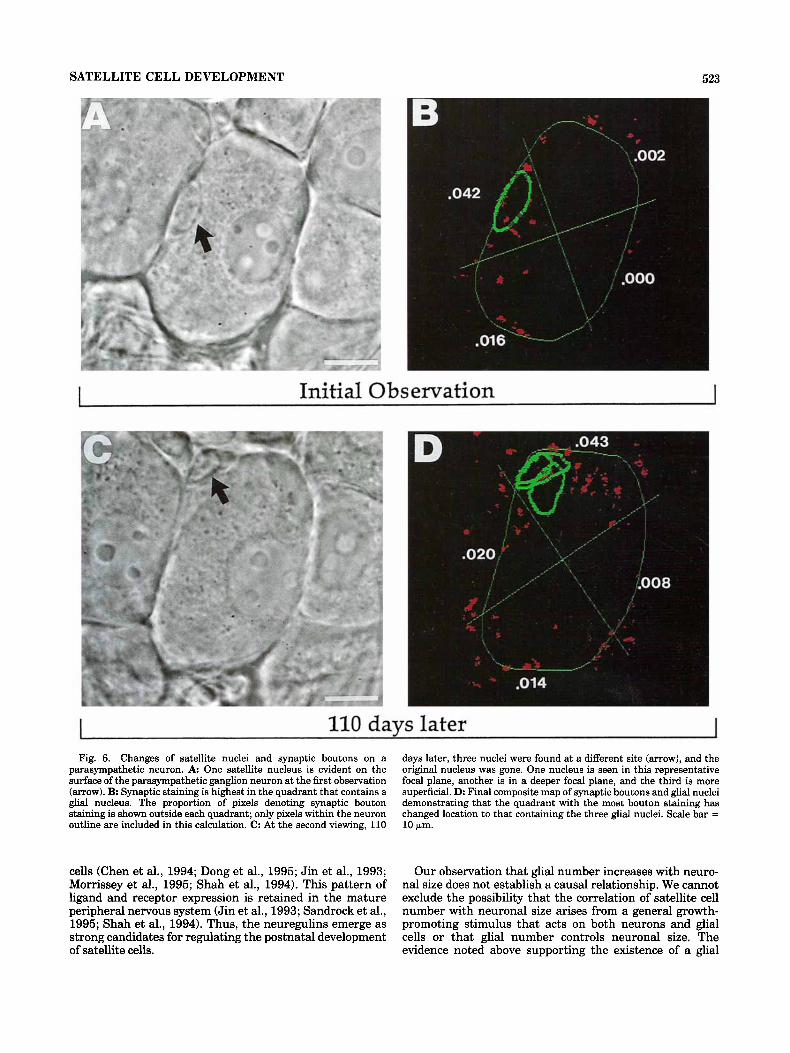
SATELLITE CELL DEVELOPMENT 523
Initial Observation I
I 110 davs later I
Fig. 6. Changes of satellite nuclei and synaptic boutons on a parasympathetic neuron. A One satellite nucleus is evident on the surface of the parasympathetic ganglion neuron at the first observation (arrow). B Synaptic staining is highest in the quadrant that contains a glial nucleus. The proportion of pixels denoting synaptic bouton staining is shown outside each quadrant; only pixels within the neuron outline are included in this calculation. C: At the second viewing, 110
days later, three nuclei were found at a different site (arrow), and the original nucleus was gone. One nucleus is seen in this representative focal plane, another is in a deeper focal plane, and the third is more superficial. D: Final composite map of synaptic boutons and glial nuclei demonstrating that the quadrant with the most bouton staining has changed location to that containing the three glial nuclei. Scale bar = 10 Fm.
cells (Chen et al., 1994; Dong et al., 1995; Jin et al., 1993; Morrissey et al., 1995; Shah et al., 1994). This pattern of ligand and receptor expression is retained in the mature peripheral nervous system (Jin et al., 1993; Sandrock et al., 1995; Shah et al., 1994). Thus, the neuregulins emerge as strong candidates for regulating the postnatal development of satellite cells.
Our observation that glial number increases with neuro- nal size does not establish a causal relationship. We cannot exclude the possibility that the correlation of satellite cell number with neuronal size arises from a general growth- promoting stimulus that acts on both neurons and glial cells or that glial number controls neuronal size. The evidence noted above supporting the existence of a glial

S.L. POMEROY ET AL. 524
TABLE 1. Synaptic Bouton Staining Compared to Changes of Glial Nucleus Number Over the 2-3 Month Study Interval
Initial Final synpatic synaptic
n staining' staining' P2
Regions with increased glial nuclei at
Regions with decreased glial nuclei at
Regions with no change of glial nuclei 130 ,055 2 ,042 ,055 f ,043 N.S.
'Proportion of pixels denoting synaptic bouton staining (mean f standard deviation). 2Paired t-test with Bonferroni correction.
final viewing 48 ,042 2 .028 .068 f .042 <.001
final viewing 22 ,053 -t ,038 .039 -t ,033 < ,001
growth factor on the cell surface of peripheral neurons suggests a neuron-derived stimulus for glial development. With this in mind, we propose that neuronal growth regulates satellite cell development during postnatal devel- opment and that the addition of satellite cells to salivary duct ganglia is a consequence of neuronal enlargement. Glial cells may, in turn, provide neurotrophic support to the neurons with which they are associated. For example, satellite cells in autonomic ganglia have been shown to participate in the synaptic response to postganglionic axotomy (Matthews and Nelson, 1975; Purves, 1975,19761, and Schwann cells, which are quite similar to satellite cells, have been found to express neurotrophic factors in re- sponse to axonal injury (Heumann et al., 1987; Taniuchi et al., 1988). Reciprocal cell-cell interactions, with neural cells expressing neuregulins that promote the expression of neurotrophic factors by surrounding nonneural cells, have been described for the embryonic development of sympa- thetic ganglia (Verdi et al., 1996). Our observation of the positional coincidence of synaptic boutons and satellite cell nuclei during postnatal development may be a manifesta- tion of a similar interdependence of neurons and glia. This result extends earlier observations (Pomeroy and Purves, 1988; Purves and Lichtman, 1987; Purves et al., 1987) by demonstrating that synaptic remodeling occurs in concert with satellite cell development.
Estimation of nuclear number using the method of Abercrombie (1946) provides an accurate assessment of cell number if several assumptions are met. These include making accurate measurements of nuclear height, cutting even sections at least 1.5 times the thickness of the nuclear height, and identifying all nuclear fragments in all sections (Abercrombie, 1946; Coggeshall and Lekan, 1996). We have conformed to these assumptions and have verified the counts obtained from serial sections by focusing through the entire depth of parasympathetic neurons and counting their associated glia. These results, obtained both in whole mounts and by direct observation in living animals, confirm that the number of satellite cells per neuron increases during postnatal development. Thus, our experiments pro- vide consistent estimates of the number of satellite cells associated with parasympathetic neurons and are also consistent in supporting the conclusion that satellite cells are added to neurons during postnatal development.
Considered together with earlier investigations of auto- nomic ganglion development, the evidence presented here shows that the postnatal increase of satellite cells associ- ated with parasympathetic neurons parallels neuronal devel- opment. We propose that neuronal growth promotes glial cell development and that both ultimately are regulated by an increasing target of innervation in postnatal life. From this perspective, the cellular response of the nervous sys-
tem to somatic growth extends to glial cells that surround and support neurons during postnatal development.
ACKNOWLEDGMENTS We thank Pat Newton, Ladan Hedayati, and Steve Tur-
ney for help with technical aspects of the project and Anthony LaMantia, Jeff Lichtman, and Dale Purves for advice and comments on the manuscript. This project was supported by NIH grant NS27773 and a Biomedical Re- search Support Grant.
LITERATURE CITED Abercrombie, M. (1946) Estimation of nuclear population from microtome
sections. Anat. Rec. 94239-247. Balice-Gordon, R.J., and J.W. Lichtman (1990) In vivo visualization of the
growth of pre and postsynaptic elements of neuromuscular junctions in the mouse. J. Neurosci. 103394-908.
Chen, M.S., 0. Bermingham-McDonogh, F.T.Y. Danehy, Jr., C. Nolan, S.S. Scherer, J. Locas, D. Gwynne, and M.A. Marchionni (1994) Expression of multiple neuregulin transcripts in postnatal rat brains. J. Comp. Neurol. 349:389400.
Coggeshall, R.E., and H.A. Lekan (1996) Methods for determining numbers of cells and synapses: A case for more uniform standards. J. Comp. Neurol. 364:&15.
Dobbing, J., and J. Sands (1973) Quantitative growth and development of human brain. Arch. Dis. Child. 48:757-767.
Dong, Z., A. Brennan, N. Liu, Y. Yarden, G. Lefkowitz, R. Mirsky, R., and K.R. Jessen (1995) Neu differentiation factor is a neuron-glia signal and regulates survival, proliferation, and maturation of rat Schwann cell precursors. Neuron 15:585-596.
Gabella, G. (1976) Structure of the Autonomic Nervous System. London: Chapman and Hall. p. 214.
Gassmann, M., F. Casagranda, D. Orioli, H. Simon, C. Lai, R. Klein, and G. Lemke (1995) Aberrant neural and cardiac development in mice lacking the ErbB4 neuregulin receptor. Nature 378t390-394.
Hall, A.K., and S.C. Landis (1991) Early commitment of precursor cells from the rat superior cervical ganglion to neuronal or nonneuronal fates. Neuron 6341-752.
Hall, A.K., and S.C. Landis (1992) Division and migration of satellite glia in the embryonic rat superior cervical ganglion. J. Neurocytol. 21:635-647.
Hamburger, V., and R. Levi-Montalcini (1949) Proliferation, differentiation, and degeneration in the spinal ganglia of the chick embryo under normal and experimental conditions. J. Exp. Zool. 111:457-501.
Heumann, R., S. Korsching, C. Bandtlow, and H. Thoenen (1987) Changes of nerve growth factor synthesis in nonneuronal cells in response to sciatic nerve transection. J. Cell Biol. 104:1623-1631.
Humbertson, A,, Jr., E. Zimmermann, and M. Leedy (1969) A chronological study of mitotic activity in satellite cell hyperplasia associated with chromatolytic neurons. Z. Zellforsch. 100:507-515.
Jin, J.-J., A.Y. Nikitin, and M.F. Rajewsky (1993) Schwann cell lineage- specific neu (erbB2) gene expression in the developing rat nervous system. Cell Growth Differ. 4:227-237.
Lee, K.-F., H. Simon, H. Chen, B. Bates, M.-C. Hung, and C. Hauser (1995) Requirement for neuregulin receptor erbB2 in neural and cardiac development. Nature 378r394-398.
Matthews, M.R., and V.H. Nelson (1975) Detachment of structurally intact nerve endings from chromatolytic neurones of rat superior cervical ganglion during the depression of synaptic transmission induced by postganglionic axotomy. J. Physiol. (London) 245:91-135.
McCullagh, P. and J.A. Nelder (1995) Generalized Linear Models, 2nd Ed. London: Chapman and Hall, pp. 2 1 4 3 .
Meyer, D., and C. Birchmeier (1995) Multiple essential functions of neuregu- lin in development. Nature 378t386-390.
Morrissey, T.K., A.D.O. Levi, A. Nuijens, M.X. Sliwkowski, and R.P. Bunge (1995) Axon-induced mitogenesis of human Schwann cells involves
Pannese, E. (1960) Observations on the morphology, submicroscopic struc- ture, and biological properties of satellite cells in sensory ganglia of mammals. Z. Zellforsch. 52567-597.
Pannese, E. (1981) The satellite cells of the sensory ganglia. In: Adv. Anat. Embryo. Cell. Biol., Vol. 65. Berlin: Springer-Verlag, pp 1-107.
neuregulin and p185erbB2. Roc. Natl. Acad. SC1. USA 921431-1435,

SATELLITE CELL DEVELOPMENT 525
Pannese, E., R. Bianchi, B. Calligaris, R. Ventura, and E.R. Weihel (1972) Quantitative relationships between nerve and satellite cells in spinal ganglia. An electron microscopical study. I. Mammals. Brain Res. 46215-234.
Peles, E., S.S. Bacus, R.A. Koski, H.S. Lu, D. Wen, S.G. Ogden, R.B. Levy, and Y. Yarden (1992) Isolation of the neuiHER2 stimulatory ligand: A 44 kd glycoprotein that induces differentiation of mammary tumor cells. Cell 69t205-216.
Peters, A,, S.L. Palay, and H. DeF. Wehster (1991) The Fine Structure of the Nervous System. New York Oxford, pp. 213-218.
Plowman, G.D., J.-M. Culouscou, G.S. Whitney, J.M. Green, G.W. Carlton, L. Foy, M.G. Neubauer, and M. Shoyab (1993) Ligand-specific activation of HER4ip18OerhB4, a fourth member of the epidermal growth factor receptor family. Proc. Natl. Acad. Sci. USA 90t1746-1750.
Pomeroy, S.L., and D. Purves (1988) Neuroniglial relationships observed over intervals of several months in living mice. J. Cell Biol. 107t1167- 1175.
Pomeroy, S.L., A.3. LaMantia, and D. Purves (1990) Postnatal construction of neural circuitry in the mouse olfactory bulb. J. Neurosci. 10:1952- 1966.
Purves, D. (1975) Functional and structural changes in mammalian sympa- thetic neurones following interruption of their axons. J. Physiol. (Lon- don) 252t405-426.
Purves, D. (1976) Functional and structural changes in mammalian sympa- thetic neurones following colchicine application to postganglionic nerves. J. Physiol. (London) 259t159-175.
Purves, D. (1988) Body and Brain: A Trophic Theory of Neural Connections. Cambridge, MA: Harvard University Press.
Purves, D. (1994) Neural Activity and the Growth of the Brain. New York: Cambridge University Press.
Purves, D., and A.-S. LaMantia (1993) The development of blobs in the monkey visual cortex. J. Comp. Neurol. 334t169-175.
Purves, D., and J.W. Lichtman (1985) Geometrical differences among homologous neurons in mammals. Science 228t298-302.
Purves, D., and J.W. Lichtman (1987) Synaptic sites on reinnervated nerve cells visualized at two different times in living mice. J. Neurosci. 7t1492-1497.
Purves, D., E. Rubin, W.D. Snider, and J.W. Lichtman (1986) Relation of animal size to convergence, divergence, and neuronal number in periph- eral nervous sympathetic pathways. J. Neurosci. 6158-163.
Purves, D., J.T. Voyvodic, L. Magrassi, and H. Yaw0 (1987) Nerve terminal remodeling visualized in living mice by repeated examination of the same neuron. Science 238: 1122-1 126.
Riddle, D.R., A. Richards, F. Zsuppan, and D. Purves (1992) Growth of the rat somatic sensory cortex and its constituent parts during postnatal development. J. Neurosci. 12t3509-3524.
Sandrock, A.W., A.D.J. Goodearl, &.-W. Yin, D. Chang, and G.D. Fischbach (1995) J. Neurosci. 15t6124-6136.
Selhy, S.M. (1969) Standard Mathematical Tables. ARIA is concentrated in nerve terminals at neuromuscular junctions and at other synapses. Cleveland: Chemical Rubber Co., p. 18.
Shah, N.M., M.A. Marchionni, I. Isaacs, P. Stroobant, and D.J. Anderson (1994) Glial growth factor restricts mammalian neural crest stem cells to a glia fate. Cell 77t349-360.
Snedecor, G.W., and W.G. Cochran (1980) Statistical Methods, 7th Ed. Ames, IA: Iowa State University Press, pp. 93-97.
Snider, W.D. (1987) A comparison of the dendritic complexity and the innervation of suhmandihular neurons in five species of small animals. J. Neurosci. 7; 1760-1 768.
Taniuchi, M., H.B. Clark, J.B. Schweitzer, and E.M. Johnson, Jr. (1988) Expression of nerve growth factor receptors hy Schwann cells of axotomized peripheral nerves: Ultrastructural location, suppression by axonal contact, and binding properties. J. Neurosci. 8t664-681.
Terni, T. (1920) Sulla correlazione fra ampiezza del territorio di innervazi- one e grandezza delle cellule ganglionari. 11. Ricerchi sui gangli spinali che innervano la coda rigenerata, nei Sauri (Gongylus ocellatus). Arch. Ital. Anat. Emhriol. 17t507-543.
Verdi, J.M., A.K. Groves, I. Farifias, K. Jones, M.A. Marchionni, L.F. Reichardt, and D.J. Anderson (1996) A reciprocal cell-cell interaction mediated by NT-3 and neuregulins controls the early survival and development of sympathetic neuroblasts. Neuron 16t515-527.
Volpe, J.J. (1995) Neurology of the Newborn, 3rd Ed. Philadelphia: W.B. Saunders, pp. 43-92.
Voyvodic, J.T. (1986) A general purpose image processing language (IMAGR) facilitates visualizing neuronal structure in fixed tissue and in vivo. SOC. Neurosci. Abstr. 12t390.
Voyvodic, J.T. (1987) Development and regulation of dendrites in the rat superior cervical ganglion. J. Neurosci. 7t904-912.
Voyvodic, J.T. (1989) Peripheral target regulation of dendritic geometry in the rat superior cervical ganglion. J. Neurosci. 9t1997-2010.