potential distribution of the alien invasive brown tree ... · potential distribution of the alien...
TRANSCRIPT

11
Potential Distribution of the Alien Invasive Brown Tree Snake,Boiga irregularis (Reptilia: Colubridae)1
Dennis Rodder2,3,4 and Stefan Lotters3
Abstract: The Brown Tree Snake (Boiga irregularis) is native to Southeast Asiaand Australia and has been introduced to Guam. There it causes major ecologi-cal and socioeconomic problems and is considered to belong to the 100 worstalien invasive species worldwide. We used a maximum entropy-based ClimateEnvelope Model to identify worldwide areas outside the species’ known rangethat are potentially suitable under current climatic conditions. Projections re-vealed that this invasive alien species potentially occurs in tropical and somesubtropical regions. In the closer vicinity of the snake’s known distribution,highest suitability was found for the Northern Mariana Islands, HawaiianIslands, Madagascar, New Caledonia, and Fiji Islands. If predictions are inter-preted as depicting invasiveness potential of B. irregularis, strategies to preventinvasion should focus on these regions. An analysis of potential distributionsunder different future anthropogenic climate-change scenarios showed thatthe Fiji Islands, Hawaiian Islands, and Northern Mariana Islands will remainoverall most suitable habitat for the Brown Tree Snake. In addition, we notedan increase of suitability in New Zealand.
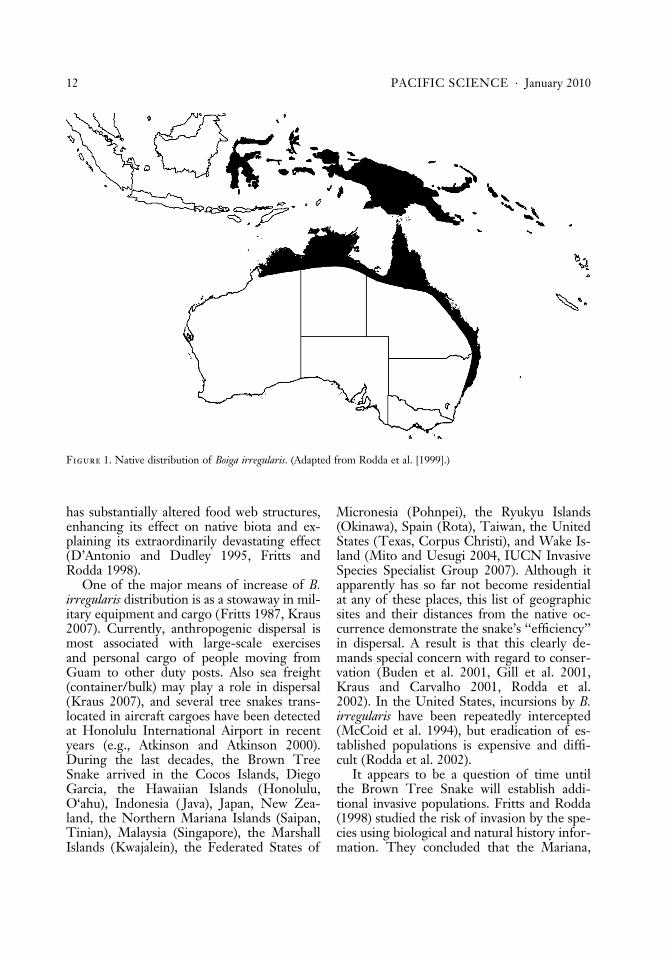
Alien invasive species are a concern innature conservation because they may havenegative impact on native biodiversity andcan have major socioeconomic impacts. A re-markable example is the Brown Tree Snake,Boiga irregularis (Merrem, 1802) (Savidge1987, Rodda and Fritts 1992, Fritts andRodda 1998, Wiles et al. 2003). This venom-ous, 1- to 3-m-long arboreal colubrid snake isnative to Papua New Guinea, the SolomonIslands, and the northern and eastern coastsof Australia (Figure 1) (Rodda et al. 1999).
Its status in Sulawesi (Indonesia) is uncertain.Rodda et al. (1999) listed populations fromSulawesi as ‘‘native,’’ but Iskandar and Tjan(1996) suggested that B. irregularis may havereached the island only via trade. Ecologi-cally, this snake can be regarded a ‘‘general-ist’’ concerning habitat requirements andprey selectivity. Boiga irregularis is knownfrom natural forest and grassland as well asagricultural and urban areas and does notshow particular food preferences (Rodda etal. 1999).
The Brown Tree Snake was brought ac-cidentally to several islands in the Pacificshortly after World War II where it success-fully established local populations due to ob-viously unoccupied niches and plentiful naiveprey (Rodda et al. 1992). As a result, B. irreg-ularis today is listed among the 100 worstalien invasive species worldwide (Lowe et al.2000). The most devastating consequencesof its introduction are known from Guam,where it led to decimation of vertebrate spe-cies including flying foxes, several small ter-restrial mammals, and lizards as well as theextinction of eight of the 11 endemic birdspecies in the 1980s (Savidge 1987, Wileset al. 2003). It is suggested that B. irregularis
Pacific Science (2010), vol. 64, no. 1:11–22doi: 10.2984/64.1.011: 2010 by University of Hawai‘i PressAll rights reserved
1 This work benefited from a grant by the Graduier-tenforderung des Landes Nordrhein-Westfalen to D.R.Manuscript accepted 1 February 2009.
2 Herpetology Department, Zoologisches For-schungsmuseum Alexander Koenig, Adenauerallee 160,53113 Bonn, Germany.
3 Biogeography Department, Trier University, 54286Trier, Germany.
4 Corresponding author: (e-mail: [email protected]).
has substantially altered food web structures,enhancing its effect on native biota and ex-plaining its extraordinarily devastating effect(D’Antonio and Dudley 1995, Fritts andRodda 1998).
One of the major means of increase of B.irregularis distribution is as a stowaway in mil-itary equipment and cargo (Fritts 1987, Kraus2007). Currently, anthropogenic dispersal ismost associated with large-scale exercisesand personal cargo of people moving fromGuam to other duty posts. Also sea freight(container/bulk) may play a role in dispersal(Kraus 2007), and several tree snakes trans-located in aircraft cargoes have been detectedat Honolulu International Airport in recentyears (e.g., Atkinson and Atkinson 2000).During the last decades, the Brown TreeSnake arrived in the Cocos Islands, DiegoGarcia, the Hawaiian Islands (Honolulu,O‘ahu), Indonesia ( Java), Japan, New Zea-land, the Northern Mariana Islands (Saipan,Tinian), Malaysia (Singapore), the MarshallIslands (Kwajalein), the Federated States of
Micronesia (Pohnpei), the Ryukyu Islands(Okinawa), Spain (Rota), Taiwan, the UnitedStates (Texas, Corpus Christi), and Wake Is-land (Mito and Uesugi 2004, IUCN InvasiveSpecies Specialist Group 2007). Although itapparently has so far not become residentialat any of these places, this list of geographicsites and their distances from the native oc-currence demonstrate the snake’s ‘‘efficiency’’in dispersal. A result is that this clearly de-mands special concern with regard to conser-vation (Buden et al. 2001, Gill et al. 2001,Kraus and Carvalho 2001, Rodda et al.2002). In the United States, incursions by B.irregularis have been repeatedly intercepted(McCoid et al. 1994), but eradication of es-tablished populations is expensive and diffi-cult (Rodda et al. 2002).
It appears to be a question of time untilthe Brown Tree Snake will establish addi-tional invasive populations. Fritts and Rodda(1998) studied the risk of invasion by the spe-cies using biological and natural history infor-mation. They concluded that the Mariana,
Figure 1. Native distribution of Boiga irregularis. (Adapted from Rodda et al. [1999].)
12 PACIFIC SCIENCE . January 2010

Hawaiian, and Caroline islands are most atrisk because a native snake fauna is absent andpotential prey density is high. Invasion riskwill depend on the arrival of founder individ-uals over time and also on environmentalconditions making more areas available forthe ectothermic species.
We see an urgent need to identify thoseregions in which this species finds suitableconditions for successful establishment of in-vasive populations. Climate Envelope Modelsare a useful tool to study potential distribu-tions of species under past, current, and fu-ture climatic scenarios (e.g., Hijmans andGraham 2006, Malcom et al. 2006, Petersonand Nyari 2007, Carnaval and Moritz 2008,Rodder et al. 2008, Rodder 2009). The scopeof the study reported here is to assess theworldwide potential distribution of B. irregu-laris under current climatic conditions toidentify regions with high potential for inva-sions.
materials and methods
Species Records and Climate Data
A total of 2,679 Boiga irregularis recordswas available through the Global BiodiversityInformation Facility (GBIF, www.gbif.org)and HerpNet databases (www.herpnet.org).In addition, records of invasive populationswere obtained from the IUCN Invasive Spe-cies Specialist Group (www.issg.org), where-by only records within areas with confirmedreproduction were included. For georeferenc-ing, the Alexandria Digital Library GazetteerServer Client (www.middleware.alexandria.ucsb.edu/client/gaz/adl/index.jsp) was used.
We used DIVA-GIS 5.4 (Hijmans et al.2001) to test the accuracy of coordinates(Check Coordinates tool) by comparing in-formation accompanying the species recordsand locality data extracted from an adminis-trative boundaries database at the smallestpossible level (country/state/city). This in-formation should be the same, and any mis-matches may reflect errors (see Hijmans et al.1999). In addition, we used altitudinal infor-mation to spot likely errors in the coordinatedata when this information was provided withthe records used. Altitude was compared with
the altitude of the locality in a digital eleva-tion model, using the Extract Values byPoints function of DIVA-GIS. Of the totalrecords available, 337 were situated in uniquegrid cells within the species’ native range,classified as being accurate and hence suitablefor model building (see later section).
Information on current climate was ob-tained from the WorldClim database, version1.4 (www.worldclim.org), which is based onweather conditions recorded between 1950and 2000 with a grid cell resolution of 2.5min (Hijmans et al. 2005). It was created byinterpolation using a thin-plate smoothingspline of observed climate at weather sta-tions, with latitude, longitude, and elevationas independent variables (Hutchinson 1995,2004).
Variable Selection
Rodda et al. (2007) investigated the climateenvelope of the Brown Tree Snake in its na-tive geographic range regarding annual meanmonthly temperature and precipitation. Theyfound that the amount of precipitation was agood predictor for its distribution. Such a re-lationship appears to be further supported bynatural history observations because the sea-sonal activity of the Brown Tree Snake islimited to the warmer and wetter summermonths in its Australian range (Covacbvichand Limpus 1973, Shine 1991a). It was alsoobserved that the snake is unable to shedproperly when the relative humidity is lowerthan 60% (Rodda et al. 1999). Within the na-tive distribution of the target species, this de-pendency is reflected in the geographic rangein Australia, where B. irregularis inhabitsmainly humid areas close to the sea. There-fore, next to the mean annual values, mini-mum and maximum annual values may beimportant, suggesting that ‘‘annual precipita-tion,’’ ‘‘precipitation of the wettest month,’’and ‘‘precipitation of the driest month’’ aresuitable predictor variables.
Temperature is a key factor influencingectothermic species; therefore the ‘‘annualmean temperature’’ is an important variablerelated to energetic balances and digestiveturnover rates. Mathies and Miller (2002)
Potential Distribution of Boiga irregularis . Rodder and Lotters 13

showed that temperature seasonality is physi-ologically important in influencing reproduc-tion. Those authors investigated the effect oftwo different temperature regimes for elicit-ing reproduction in male and female B. irreg-ularis. They found that specimens maintainedat 24�C followed by a 60-day cool period at19�C exhibited substantial reproductive ac-tivity. Clutches were produced by femalesshortly after returning to 24�C. In contrast,individuals maintained at 28�C followed byan identical 19�C cooling period exhibitedrelatively little reproductive activity, andnone of the females produced eggs. Con-sidering these effects, we added the ‘‘maxi-mum temperature of the warmest month’’and ‘‘minimum temperature of the coldestmonth’’ as predictor variables for model com-putation.
Climate Envelope Models
Maxent 3.2.1 (www.cs.princeton.edu/~shapire/maxent) (Phillips et al. 2006) wasapplied for Climate Envelope Model cal-culation to assess the potential distributionof the Brown Tree Snake and to map it intogeographic space. Maxent is a grid-basedmachine-learning algorithm following theprinciples of maximum entropy ( Jaynes1957). The general concept is to find a prob-ability distribution covering the study areathat satisfies a set of constraints derived fromoccurrence data. Each constraint requires thatthe expected value of an environmental vari-able or a function thereof must be within aconfidence interval of its empirical meanover the presence records. The programchooses the distribution that is closest to uni-form and therefore maximizes entropy withinall distributions that satisfy the constraintsbecause any other choice would representconstraints on the distribution that are notjustified by the data. Maxent is able to incor-porate complex dependencies between pre-dictor variables and has been shown to revealbetter results than other comparable methods(e.g., Elith et al. 2006, Wisz et al. 2008). Thereliability of the results obtained from Max-ent models has been confirmed by its capacityto predict novel presence localities for poorly
known species (Pearson et al. 2007) and theoutcome of introductions of alien invasivespecies outside the native distribution (e.g.,Peterson and Vieglais 2001, Ficetola et al.2007, Jeschke and Strayer 2008, Rodder etal. 2008, Rodder 2009).
Runs used herein were conducted usingthe default values for all program settings,whereby randomly chosen background pointswere restricted to an area defined by a mini-mum convex polygon including all native rec-ords. Herein, the logistic output format withsuitability values ranging from 0 (unsuitable)to 1 (optimal) was used, and areas where‘‘clamping’’ (i.e., nonanalogous climatic con-dition present in the training area) occurredduring projections were subsequently ex-cluded (Philips and Dudık 2008).
Maxent allows for model testing by calcu-lation of the Area Under the Curve (AUC),referring to the ROC (Receiver OperationCharacteristic) curve; herein we assessed theability of the model to distinguish back-ground points from training points (Hanleyand McNeil 1982, Phillips et al. 2006). Thismethod is recommended for ecological appli-cations because it is nonparametric (Pearceand Ferrier 2000, but see Lobo et al. 2008).Values of AUC range from 0.5 (random) formodels with no predictive ability to 1.0 formodels giving perfect predictions. Accordingto the classification of Swets (1988), AUCvalues >0.9 describe ‘‘very good,’’ >0.8‘‘good,’’ and >0.7 ‘‘useful’’ discriminationabilities. Maxent allows for an assessment ofthe relative contribution of variables includedusing a jackknifing approach.
results
Current Potential Distribution
We obtained ‘‘very good’’ AUC values in themodel (AUC ¼ 0.971), and the known inva-sive range of Boiga irregularis on Guam is sit-uated within higher Maxent classes (>0.780),also confirming the predictive power of themodel. The lowest observed Maxent value atthe presence point used for model trainingwas 0.201. Analyses of variable contributionsin the model revealed that ‘‘annual precipita-
14 PACIFIC SCIENCE . January 2010

tion’’ with 33.3% has the highest explanativepower, followed by ‘‘maximum temperatureof the warmest month’’ (28.5%), ‘‘annualmean temperature’’ (13.8%), ‘‘precipitationof the driest month’’ (11.7%), ‘‘precipitationof the wettest month’’ (7.7%), and the ‘‘min-imum temperature of the coldest month’’(4.9%). The relative variable importance ob-tained via jackknifing from the Maxent modelappears to be consistent with natural historyobservations (see earlier). ‘‘Clamping’’ oc-curred at very few sites, such as at a very smallridge on the west coast of India. Those areaswere excluded from further analyses.
Under current climatic conditions, theBrown Tree Snake has a geographically widepotential distribution throughout almost allthe tropics and adjacent subtropical regions,especially widespread within the SouthernHemisphere but also extending to NorthAmerica including Florida and coastal areasof the Gulf of Mexico (Figure 2). Regardingthe general region in which the Brown TreeSnake occurs (i.e., Southeast Asia and Austra-lia), major parts of New Caledonia, NewZealand, the Fiji Islands, and Vanuatu wereidentified as highly suitable for B. irregularisby the Climate Envelope Model (Figure 3).Guam, the Caroline Islands, and the North-ern Mariana Islands including Saipan are allsuitable for B. irregularis (Figure 3). In addi-tion to regions in the vicinity of the species’native geographic range in Southeast Asia, itsclimate envelope is mirrored in both lowland
(e.g., Congo and Amazon basins) and mon-tane (e.g., southern coastal Brazil, Ethiopianhighlands) regions. In addition, high suit-ability was found in the Hawaiian Islands(lower elevations) (Figure 4) and Madagas-car (mainly east coast and higher elevations)(Figure 5).
discussion
When interpreting results and assessing aninvasion risk, it is important to evaluate possi-ble discrepancies between the realized andfundamental climatic niche of B. irregularisand the relative contribution of ecologicalfactors other than climate, which may limitthe species’ current distribution. In additionto a species’ climate envelope, these are acces-sibility limitations and/or biotic interactionssuch as competition or predation (see also So-beron and Peterson 2005).
Biotic Interactions and/or Accessibility
Although the modeled potential distributionin the eastern portion of the native rangeof Boiga irregularis is coincident with thereported distribution (i.e., current species rec-ords), that of the western portion is not (Fig-ure 3). Climatically, the Brown Tree Snakecan find suitable areas on Borneo, Java, andSumatra west of Wallace’s line, but none ofthese islands has been successfully invaded,as far as is known (Rodda et al. 1999, IUCN
Figure 2. Potential distribution of Boiga irregularis under current climatic conditions. Any area with a Maxent valueabove 0.201 may be climatically suitable for B. irregularis, although higher Maxent values suggest higher climatic suit-ability.
Potential Distribution of Boiga irregularis . Rodder and Lotters 15

Figure 3. Potential distribution of Boiga irregularis under current climatic conditions within Southeast Asia and Aus-tralia. Higher Maxent values suggest higher climatic suitability. Native-distribution records are indicated as opencircles and invasive ones as open triangles.
Figure 4. Potential distribution of Boiga irregularis under current climatic conditions in the Hawaiian Islands. HigherMaxent values suggest higher climatic suitability.

Invasive Species Specialist Group 2007). Pos-sible explanations could be a nonequilibriumof the actual range with climate due tolimited accessibility or ongoing spreading,limited prey availability, or the presence ofpredators or competitors (e.g., Rodda et al.1999, Araujo and Pearson 2005, Rodder et al.2008).
Accessibility highly influences the numberof jurisdictions where a species can be intro-duced, which is an important predictor of theprobability that a species can establish inva-sive populations (Bomford et al. 2008). Lim-ited accessibility appears to be an unlikelyexplanation for the absence of B. irregulariswest of Wallace’s line. Although faunal ex-change between the westernmost populationsin Sulawesi and New Guinea and those of ad-jacent Indonesian islands in past geologicaltimes was rather restricted (e.g., Inger andVoris 2001), cargo traffic within Indonesiaand adjacent islands increased during the last
century and is generally high today (Espadaand Kumazawa 2005). Therefore, the chancefor accidental introduction of specimens isexpected to be high considering that cargo isone of the major means of spreading this spe-cies (Kraus 2007).
In the case of the Brown Tree Snake,presence of competitors perhaps best explainsthe observed pattern. Shine (1991a,b) sug-gested that colubrid snakes such as B. irregu-laris are not in noteworthy competition withthe endemic elapid snakes and pythons inAustralia because they have invaded the con-tinent with ecological specializations that arerare among the endemics. Differences inforaging habitats and in the preferred preyspectrum in comparison with the endemicAustralian snakes especially may have en-hanced their success (Shine 1991b). Thismight not be true for the snake fauna westof Wallace’s line. The genus Boiga currentlycomprises 34 species; 11 of them inhabit allo-patric ranges adjacent to the native range ofB. irregularis (B. angulata, B. bengkuluensis, B.cynodon, B. dendrophila, B. drapiezii, B. jaspidea,B. multimaculata, B. nigriceps, B. philippina,B. schultzei, B. tanahjampeana) (Uetz et al.2007), but none of them is known to be inva-sive (IUCN Invasive Species Specialist Group2007). It is remarkable that although othersnakes occur in sympatry with the BrownTree Snake, other Boiga species commonlydo not. Only at Sulawesi B. dendrophila andB. multimaculata occur together with theBrown Tree Snake, but there B. irregulariswas only reported from the immediate neigh-borhood of seaports, where it might havebeen introduced (Iskandar and Tjan 1996). Itis unclear if Sulawesi inland populations actu-ally do exist (Inger and Voris 2001).
It is interesting that the Brown Tree Snakeis the only member of the genus known tohave established invasive populations. Differ-ences in behavioral traits, which might pro-vide superior abilities to establish invasivepopulations, are not known. In our opinion,the most likely explanation is that the nativerange of the Brown Tree Snake is muchcloser to areas lacking a native terrestrialsnake fauna, enhancing the chance of translo-cation.
Figure 5. Potential distribution of Boiga irregularisunder current climatic conditions in Madagascar. HigherMaxent values suggest higher climatic suitability.
Potential Distribution of Boiga irregularis . Rodder and Lotters 17

Climate
Mathies and Miller (2002) showed that a pe-riod of cool temperatures elicits reproductiveactivity in both sexes. Those authors pointedout that temperatures experienced during theartificial hibernation were much lower thanthe snakes would experience on Guam, wheretemperature seasonality is relatively invari-ant throughout the year (G1�C [Rodda et al.1999]). Comparing native populations in Aus-tralia and invasive populations on Guam,Moore et al. (2005) found that snake speci-mens from Guam exhibited substantiallyreduced body conditions compared with indi-viduals from Australia. Those authors sug-gested that Brown Tree Snakes on Guamwere living under stressful conditions, possi-bly due to overcrowding and overexploitationof food resources, resulting in decreased adultsize/weight and suppressed reproduction.The findings of Mathies and Miller (2002)might provide an additional explanation fortheir finding because environmental stressmay cause absence of hibernation and dis-turbed seasonal reproductive cycles. How-ever, although reproductive activity onGuam may be suppressed and desynchro-nized, only minor enhanced fluctuations intemperature throughout the year may be suf-ficient to elicit reproduction; therefore fluctu-ations and/or extreme weather events due toanthropogenic climate change might enhancethe snake problem. Furthermore, the obser-vation that moderate hibernation enhancesthe reproductive cycle allows the hypothesisthat Boiga irregularis may also find climati-cally suitable habitats in more temperate re-gions with a higher degree of seasonality,such as the northern parts of New Zealand,Japan, or Taiwan. Generally, climatic similar-ities of a novel environment compared withthat of the native range enhance the probabil-ity of successful establishment of alien inva-sive species (Bomford et al. 2008).
Which Regions Are at High Risk of Invasionby the Brown Tree Snake?
Fritts and Rodda (1998) studied the risk ofinvasion by the Brown Tree Snake using bio-
logical and natural history information. Theyconcluded that the Northern Mariana, Ha-waiian, and Caroline islands are most at riskbecause a native (competing) snake fauna isabsent and prey is available. Because the Mar-iana and Caroline islands are comparable interms of faunal composition with Guam, aninvasion in those islands may have similarlydramatic ecological consequences. The faunalcomposition of the Hawaiian Islands mightalso generally support the establishment ofBoiga irregularis (Kraus and Carvalho 2001).However, predictions based on natural his-tory and biological information appear tobear a higher degree of uncertainty there be-cause the faunal composition of the HawaiianIslands is distinctly different from that foundon Guam (Fritts and Rodda 1998).
Climate Envelope Model predictions ob-tained in this paper confirm the expected cli-matic suitability of the Northern Mariana,Hawaiian, and Caroline islands. In addition,our models highlight major parts of centralAfrica, Central America, South America, abroad swath of the southeastern UnitedStates coastal plain (coincident with the pre-diction presented by Rodda et al. [2007]),New Caledonia, New Zealand, and Madagas-car as having high climatic suitability for B.irregularis. Records of Brown Tree Snakesthat were accidentally imported to Hawai‘iand New Zealand already exist (Gill et al.2001, IUCN Invasive Species SpecialistGroup 2007), and it may be feared that theBrown Tree Snake will establish there. Im-pacts may be enhanced due to high availabil-ity of prey affecting population dynamics ofthe invader. Establishment of B. irregularis inNew Zealand, in addition, may be facilitateddue to the absence of competitors, becauseonly sea snakes occur there (Hydrophiidae).Similarly, for New Caledonia and the Fiji Is-lands only 19 and seven snake species areknown, respectively (Uetz et al. 2007). Six-teen of the New Caledonian species also arehydrophiids, two are fossorial Thyphlopida,but Candoia bibroni (Boidae) could be a poten-tial competitor to the Brown Tree Snake. AtFiji, there are three species each of hydro-phiids and thyphlopids plus Candoia bibroni.
It must be noted that possible discrepan-
18 PACIFIC SCIENCE . January 2010

cies between the realized climatic niche (cli-matic conditions within realized distribution)and the fundamental climatic niche of B. ir-regularis may lead to an underestimation ofthreat in some regions. Unfortunately, the re-lationship between realized and fundamentalniches can be addressed only experimentallybut not with climate envelope model ap-proaches, leaving some degree of uncertainty.However, it is interesting that the potentialdistribution of B. irregularis derived from ourmodel is remarkably coincident with the real-ized distributions of all other members ofthe genus Boiga, implying that climatic nichesare rather conservative within the genus (atleast regarding the variables chosen herein)and that we captured a great part of its funda-mental niche with our model. However, areasoutside the proposed potential distributionmay exhibit climatic conditions differentfrom those present within the realized distri-bution of B. irregularis, but they may not nec-essarily be unsuitable.
conclusions
We conclude that Boiga irregularis is a speciescausing a high extinction risk to endemicfaunas in regions where specimens are fre-quently translocated via military shipmentand cargo (Fritts 1987, Kraus 2007, Bomfordet al. 2008) and that are suitable under cur-rent climatic conditions and lack competitors,namely the Northern Mariana and Hawaiianislands, New Caledonia, and the Fiji Islands.Climatically suitable regions harboring mini-mal or lacking native snake species are likelyto be most impacted. There, availability ofresources may be enhanced compared withan ecosystem already harboring competingsnakes.
Under future anthropogenic climatechange, the situation will not change essen-tially, but climatic suitability for B. irregulariswill generally remain high in Madagascar andNew Caledonia and even increase in NewZealand (D.R., unpubl. data). Possible routesfor spreading of the species to Madagascar,New Caledonia, and the Fiji Islands includesea cargo, military equipment (Fritts 1987,Kraus 2007), and also accidental transloca-
tions via aircraft cargoes (Atkinson and Atkin-son 2000). Preventing further spreading tothe areas highlighted may depend on main-taining strict cargo checks of boats andsearches at airports.
acknowledgments
We are grateful to Marcelo R. Duarte, TomC. Mathies, and Mirco Sole, who helpedwith literature used herein. Three anony-mous reviewers helped improve this paper.
Literature Cited
Araujo, M. B., and R. G. Pearson. 2005.Equilibrium of species’ distribution withclimate. Ecography 28:693–695.
Atkinson, I. A. E., and T. J. Atkinson. 2000.Land vertebrates as invasive species on theislands of the South Pacific Regional Envi-ronment Programme. Pages 19–84 in G.Sherely, ed. Invasive species in the Pacific:A technical review and draft regional strat-egy. South Pacific Regional EnvironmentProgram, Samoa.
Bomford, M., F. Kraus, S. C. Barry, and E.Lawrence. 2008. Predicting establishmentsuccess for alien reptiles and amphibians:The role of climate matching. Biol. Inva-sions 11:713–724.
Buden, D. W., D. B. Lynch, and G. R. Zug.2001. Recent records of exotic reptiles onPohnpei, eastern Caroline Islands, Micro-nesia. Pac. Sci. 55:65–70.
Carnaval, A. C., and C. Moritz. 2008. Histor-ical climate modelling predicts patterns ofcurrent biodiversity in the Brazilian Atlan-tic forest. J. Biogeogr. 25:1187–1201.
Covacbvich, J., and C. Limpus. 1973. Twolarge winter aggregations of three speciesof tree-climbing snakes in south-easternQueensland. Herpetofauna 6:16–21.
D’Antonio, C. M., and T. L. Dudley. 1995.Biological invasions as agents of changeon islands versus mainlands. Pages 103–121 in P. M. Vitousek, L. L. Loope, andH. Andersen, eds. Islands: Biological di-versity and ecosystem function. EcologicalStudies. Springer, Berlin.
Potential Distribution of Boiga irregularis . Rodder and Lotters 19

Elith, J., C. H. Graham, R. P. Anderson, M.Dudik, S. Ferrier, A. Guisan, R. J. Hij-mans, F. Huettmann, J. R. Leathwick, A.Lehmann, J. Li, L. G. Lohmann, B. A.Loiselle, G. Manion, C. Moritz, M. Naka-mura, Y. Nakazawa, J. M. M. Overton,A. T. Perterson, S. J. Phillips, K. Richard-son, R. Scachetti-Pereira, R. E. Shapire,J. Soberon, S. Williams, M. S. Wisz, andN. E. Zimmermann. 2006. Novel methodsimprove prediction of species’ distribu-tions from occurrence data. Ecography29:129–151.
Espada, I. C., and K. Kumazawa. 2005. O-Dstructure of domestic maritime traffic inIndonesia. Proc. East. Asia Soc. Transport.Stud. 5:647–660.
Ficetola, G. F., W. Thuiller, and C. Miaud.2007. Prediction and validation of the po-tential global distribution of a problematicalien species: The American Bullfrog.Divers. Distrib. 13:476–485.
Fritts, T. H. 1987. Movements of snakesvia cargo in the Pacific region. Elepaio47:17–18.
Fritts, T. H., and G. H. Rodda. 1998. Therole of introduced species in the degrada-tion of island ecosystems. Annu. Rev.Ecol. Syst. 29:113–140.
Gill, B. J., D. Bejakovich, and A. H. Whi-taker. 2001. Records of foreign reptilesand amphibians accidentally imported toNew Zealand. N. Z. J. Zool. 28:351–359.
Hanley, J., and B. McNeil. 1982. The mean-ing of the use of the area under a receiveroperating characteristic (ROC) curve. Ra-diology 143:29–36.
Hijmans, R. J., S. E. Cameron, J. L. Parra,P. G. Jones, and A. Jarvis. 2005. Veryhigh resolution interpolated climate sur-faces for global land areas. Int. J. Climatol.25:1965–1978.
Hijmans, R. J., J. M. Cruz, R. Rojas, and L.Guarino. 2001. DIVA-GIS, version 1.4.A geographic information system for themanagement and analysis of genetic re-sources data. Manual. International PotatoCenter and International Plant GeneticResources Institute, Lima, Peru.
Hijmans, R. J., and C. H. Graham. 2006.The ability of climate envelope models topredict the effect of climate change on
species distributions. Global Change Biol.12:2272–2281.
Hijmans, R. J., M. Schreuder, J. De la Cruz,and L. Guarino. 1999. Using GIS tocheck coordinates of genebank accessions.Genet. Resour. Crop Evol. 46:291–296.
Hutchinson, M. F. 1995. Interpolating meanrainfall using thin plate smoothing splines.Int. J. Geogr. Inf. Sys. 9:385–403.
———. 2004. Anusplin, version 4.3. Centrefor Resource and Environment Studies,The Australian National University, Can-berra.
Inger, R. F., and H. K. Voris. 2001. The bio-geographical relations of the frogs andsnakes of Sundaland. J. Biogeogr. 28:863–891.
Iskandar, D. T., and K.-N. Tjan. 1996. Theamphibians and reptiles of Sulawesi, withnotes on the distribution and chromosomenumber of frogs. Pages 39–46 in D. J.Kitchener and A. Suyanto, eds. Proceed-ings of the 1st International Conferenceon Eastern Indonesian-Australian Verte-brate Fauna, Mandao. Western AustralianMuseum for lembaga Ilmu Penetahuan In-donesia, Perth, Australia.
IUCN Invasive Species Specialist Group.2007. www.issg.org.
Jaynes, E. T. 1957. Information theory andstatistical mechanics. Phys. Rev. 106:620–630.
Jeschke, J. M., and D. L. Strayer. 2008. Use-fulness of bioclimatic models for studyingclimate change and invasive species. Ann.N. Y. Acad. Sci. 1134:1–24.
Kraus, F. 2007. Using pathway analysis to in-form prevention strategies for alien rep-tiles and amphibians. Pages 94–103 inG. W. Witmer, W. C. Pitt, and K. A.Fagerstone, eds. Managing vertebrate in-vasive species: Proceedings of an interna-tional symposium. USDA/APHIS/WS,National Wildlife Research Center, FortCollins, Colorado.
Kraus, F., and D. Carvalho. 2001. The risk toHawai‘i from snakes. Pac. Sci. 55:409–417.
Lobo, J. M., A. Jimenez-Valverde, and R.Real. 2008. AUC: A misleading measureof the performance of predictive distri-bution models. Global Ecol. Biogeogr.17:145–151.
20 PACIFIC SCIENCE . January 2010

Lowe, S., M. Browne, S. Boudjelas, and M.De Poorter. 2000. 100 of the world’s worstinvasive alien species: A selection from theGlobal Invasive Species Database. The In-vasive Species Specialist Group (ISSG),Auckland.
Malcom, J. R., D. Liu, R. P. Neilson, L. Han-sen, and L. Hannah. 2006. Global warm-ing and extinction of endemic speciesfrom biodiversity hotspots. Conserv. Biol.20:538–548.
Mathies, T., and L. A. Miller. 2002. Cooltemperatures elicit reproduction in a bio-logically invasive predator, the Browntreesnake. Zoo Biol. 22:227–238.
McCoid, M. J., T. H. Fritts, and E. W.Campbell. 1994. A Brown tree snake(Colubridae: Boiga irregularis) sighting inTexas. Tex. J. Sci. 46:365–368.
Mito, T., and T. Uesugi. 2004. Invasive alienspecies in Japan: The status quo and newregulation for prevention of their adverseeffects. Global Environ. Res. 8:171–191.
Moore, I. T., M. J. Greene, A. T. Lerner,C. E. Asher, R. W. Krohmer, D. L. Hess,J. Whittier, and R. T. Mason. 2005. Phys-iological evidence for reproductive sup-pression in the introduced population ofbrown tree snakes (Boiga irregularis) onGuam. Biol. Conserv. 121:91–98.
Pearce, J., and S. Ferrier. 2000. An evaluationof alternative algorithms for fitting speciesdistribution models using logistic regres-sion. Ecol. Modell. 128:128–147.
Pearson, R. G., C. J. Raxworthy, M. Na-kamura, and A. T. Peterson. 2007. Pre-dicting species distributions from smallnumbers of occurrence records: A testcase using cryptic geckos in Madagascar.J. Biogeogr. 34:102–117.
Peterson, A. T., and A. S. Nyari. 2007. Eco-logical niche conservatism and Pleistocenerefugia in the Thrush-like Mourner, Shif-forinis sp., in the Neotropics. Evolution62:173–183.
Peterson, A. T., and D. A. Vieglais. 2001.Predicting species invasions using ecologi-cal niche modeling: New approaches frombioinformatics attack a pressing problem.BioScience 51:363–371.
Phillips, S. J., R. P. Anderson, and R. E.Schapire. 2006. Maximum entropy model-
ling of species geographic distributions.Ecol. Modell. 190:231–259.
Phillips, S. J., and M. Dudık. 2008. Modelingof species distributions with Maxent: Newextensions and comprehensive evaluation.Ecography 31:161–175.
Rodda, G. H., and T. H. Fritts. 1992. Theimpact of the introduction of the colubridsnake Boiga irregularis on Guam’s lizards.J. Herpetol. 26:166–174.
Rodda, G. H., T. H. Fritts, E. W. Campbell,K. Dean-Bradley, G. Perry, and C. P.Qualls. 2002. Practical concerns in theeradication of island snakes. Pages 260–265 in C. R. Veitch and M. N. Clout, eds.Turning the tide: The eradication of in-vasive species. Proceedings of the Inter-national Conference on Eradication ofIsland Invasives. Occasional Paper of theIUCN Species Survival Commission,Gland, Switzerland, and Cambridge,United Kingdom.
Rodda, G. H., T. H. Fritts, and P. J. Conry.1992. Origin and population growth ofthe Brown tree snake, Boiga irregularis, onGuam. Pac. Sci. 46:46–57.
Rodda, G. H., T. H. Fritts, M. J. McCoid,and E. W. Campbell. 1999. An overviewof the biology of the Brown tree snake,Boiga irregularis, a costly introduced peston Pacific islands. Pages 44–80 in G. H.Rodda, Y. Sawai, D. Chiszar, and H. Ta-naka, eds. Problem snake management:The Habu and the Brown Tree Snake.Cornell University Press, Ithaca, NewYork.
Rodda, G. H., R. N. Reed, and C. S. Jarne-vich. 2007. Climate matching as a tool forpredicting potential North Americanspread of Brown treesnakes. Pages 138–145 in G. W. Witmer, W. C. Pitt, andK. A. Fangerstone, eds. Managing verte-brate invasive species: Proceedings of aninternational symposium. USDA/APHIS/WS, National Wildlife Research Center,Fort Collins, Colorado.
Rodder, D. 2009. ‘Sleepless in Hawaii’: Doesanthropogenic climate change enhanceecological and socioeconomic impacts ofthe alien invasive Eleutherodactylus coquiThomas, 1966 (Anura: Eleutherodactyli-dae)? North-West. J. Zool. 5:16–25.
Potential Distribution of Boiga irregularis . Rodder and Lotters 21

Rodder, D., M. Sole, and W. Bohme. 2008.Predicting the potential distribution oftwo alien invasive housegeckos (Gekkoni-dae: Hemidactylus frenatus, Hemidactylusmabouia). North-West. J. Zool. 4:236–246.
Savidge, J. A. 1987. Extinction of an islandforest avifauna by an introduced snake.Ecology 68:660–668.
Shine, R. 1991a. Australian snakes: A naturalhistory. Cornell University Press, Ithaca,New York.
———. 1991b. Strangers in a strange land:Ecology of the Australian colubrid snakes.Copeia 1991:120–131.
Soberon, J., and A. T. Peterson. 2005. Inter-pretation of models of fundamental eco-logical niches and species’ distributionalareas. Biodiv. Inf. 2:1–10.
Swets, K. 1988. Measuring the accuracy ofdiagnostic systems. Science (Washington,D.C.) 240:1285–1293.
Uetz, P., J. Goll, and J. Hallermann. 2007.Die TIGR-Reptiliendatenbank. Elaphe15:22–25.
Wiles, G. J., J. Bart, R. E. Beck, and C. F.Aguon. 2003. Impacts of the Brown treesnake: Patterns of decline and species per-sistence in Guam’s avifauna. Conserv. Biol.17:1350–1360.
Wisz, M. S., R. J. Hijmans, A. T. Peterson,C. H. Graham, A. Guisan, and NPSDWGroup. 2008. Effects of sample size onthe performance of species distributionmodels. Divers. Distrib. 14:763–773.
22 PACIFIC SCIENCE . January 2010