premotor neurons for vertical eye movements in the rostral mesencephalon of monkey and human:...
TRANSCRIPT

Premotor Neurons for Vertical EyeMovements in the Rostral Mesencephalon
of Monkey and Human: HistologicIdentification by Parvalbumin
Immunostaining
ANJA K.E. HORN* AND JEAN A. BUTTNER-ENNEVER
Institute of Anatomy, LMU Munich, D-80336 Munich, Germany
ABSTRACTIn the monkey, premotor neurons for vertical gaze are located in the mesencephalic
reticular formation: the rostral interstitial nucleus of the medial longitudinal fascicle (riMLF)contains medium-lead burst neurons, and the interstitial nucleus of Cajal (iC) acts asintegrator for the eye-velocity signals to eye-position signals. Both nuclei lie adjacent to eachother and are similar in appearance at the transition zone in Nissl-stained sections, whichmakes a delineation of the functionally different nuclei difficult in human. For a neuropatho-logic analysis of degenerative changes in saccadic disorders of patients, the histologicidentification of the riMLF and the iC is important. The aim of this study is to identify bothnuclei in human by using parvalbumin as a histologic marker. First, in monkeys the premotorneurons in riMLF and iC were identified by trans-synaptic labelling after injections of tetanustoxin fragment C into vertical-pulling eye muscles. Premotor neurons were found in theriMLF mainly ipsilateral to the corresponding eye muscle motoneurons and on both sideswithin the iC, but here the labelled cell populations differed: the contralateral side containedmore medium-sized cells compared with the mainly small-sized cell population on theipsilateral side. Double labelling showed that almost all premotor neurons in the iC and allpremotor neurons in the riMLF were parvalbumin-immunoreactive. The immunocytochemi-cal staining of human brainstem sections revealed the riMLF as a cluster of medium-sized,elongated parvalbumin-positive cells, with a similar appearance and at a similar location asthat in monkey: a wing-shaped nucleus dorsomedial to the red nucleus, rostral to thetraversing tractus retroflexus, dorsally bordered by the thalamo-subthalamic paramedianartery. The adjacent iC could be distinguished easily by its more densely packed, roundparvalbumin-immunoreactive neurons. The exact identification of premotor neurons of thevertical system in the normal human brain provides a reference basis for the neuropathologicanalysis of vertical gaze disorders at a cellular level. J. Comp. Neurol. 392:413–427, 1998.r 1998 Wiley-Liss, Inc.
Indexing terms: mesencephalic reticular formation; rostral interstitial nucleus of the medial
longitudinal fascicle; interstitial nucleus of Cajal; burst neurons; trans-synaptic
labelling
Neurons essential for the generation of vertical saccadiceye movements lie in the rostral interstitial nucleus of thefasciculus longitudinalis medialis (riMLF) in the mesence-phalic reticular formation. Recording experiments in mon-keys have shown that the riMLF contains medium-leadburst neurons (also called short-lead burst neurons) thatdischarge with high-frequency bursts 8 to 15 ms beforeand during upward or downward saccades (Buttner et al.,1977; Hepp et al., 1989). Recent physiologic studies apply-
ing three-dimensional eye position recordings demon-strated that burst neurons in the riMLF in addition encode
Grant sponsor: Deutsche Forschungsgemeinschaft; Grant number: Ho1639/3-1/3-2.
*Correspondence to: Dr. Anja Horn, Institute of Anatomy III, LMUMunich, Pettenkoferstrasse 11, D-80336 Munich, Germany.E-mail: [email protected]
Received 15 April 1997; Revised 3 October 1997; Accepted 7 October 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 392:413–427 (1998)
r 1998 WILEY-LISS, INC.

the torsional component of saccadic eye movements (Viliset al., 1989; Crawford and Vilis, 1992). These studiessuggest that the right riMLF contains up- and down-burstneurons with a clockwise torsional component, and the leftriMLF up- and down-burst neurons with a counterclock-wise torsional component (Vilis et al., 1989). Accordingly,unilateral lesions of the riMLF produce deficits of torsionalsaccades in the ipsitorsional direction, whereas saccadesin the vertical direction are less affected (Vilis et al., 1989;Crawford and Vilis, 1992; Suzuki et al., 1995). In addition,unilateral riMLF lesions produce a spontaneous torsionalnystagmus with fast phases to the contralateral side, andipsilesional fast phases of the torsional vestibular-ocularreflex are lost or slowed (Crawford and Vilis, 1992; Suzukiet al., 1995; Helmchen et al., 1996b). Bilateral lesionsresult in a complete vertical paralysis (Kompf et al., 1979;Suzuki et al., 1995). Clinically, the rare, pure isolateddowngaze paralysis is only seen after bilateral discretelesions of the riMLF, whereas a combined up- and down-gaze paralysis can be observed after unilateral riMLFlesions mostly with involvement of the posterior commis-sure (Christoff, 1974; Cogan, 1974; Trojanowski and Lafon-taine, 1981; Buttner-Ennever et al., 1982; Pierrot-Deseil-ligny et al., 1982), but in some unilateral cases, theposterior commissure is spared from the lesion (Ranalli etal., 1988; Riordan-Eva et al., 1996; Helmchen et al. 1996b).
Anatomical studies showed direct projections of theriMLF to the motoneurons of vertical-pulling eye muscles(Buttner-Ennever and Buttner, 1978; Steiger and Buttner-Ennever, 1979; Buttner-Ennever et al., 1981; Moschovakiset al., 1991a, 1991b; Wang and Spencer, 1996a).
The adjacent interstitial nucleus of Cajal (iC) is alsorelated to vertical and torsional eye movements but ismore involved in the integration of eye-velocity signalsinto eye-position signals (Crawford et al., 1991; Fuku-shima et al., 1992) and eye-head coordination (Fukushima,1987). Unilateral lesions of the iC result in a contralateralhead tilt and torsion of the eyes to the contralateral side(as riMLF), but a torsional spontanous nystagmus to theipsilateral side (opposite to riMLF) (Halmagyi et al., 1994).The target sites of the iC projections are the motoneuronsof the vertical extraocular muscles, the vestibular nuclei,and the motoneurons of the neck muscles in the spinal cord(Holstege and Cowie, 1989; Kokkoroyannis et al., 1996).
In the monkey, the cells of the riMLF are embedded inthe fibres of the medial longitudinal fascicle (MLF) andform a wing-shaped nucleus in the reticular formation ofthe rostral mesencephalon. It lies dorsomedial to theparvocellular portion of the red nucleus. The riMLF ad-joins directly the rostral end of the iC from which it isseparated by the traversing fibres of the tractus retro-flexus (TR). Its rostral end is roughly demarcated by thetraversing fibres of the mamillo-thalamic tract (MT;Buttner-Ennever and Buttner, 1988).
Although the riMLF and iC were already identified inhuman, it is still difficult to delineate both nuclei clearly inNissl-stained sections (Buttner-Ennever et al., 1982; Cross-land et al., 1994). The riMLF and iC lie in close proximityto each other but play different roles in the generation ofvertical eye movements. For a neuropathologic analysis ofhuman brains with saccadic disorders, it is important toidentify the riMLF and the burst neurons, delineate themfrom the neighbouring iC, and correlate the individualdamage to each with functional deficits.
Recently, the saccadic omnipause neurons and the premo-tor burst neurons of the horizontal saccadic system wereidentified in monkey and human on the basis of theirparvalbumin immunostaining (Horn et al., 1994, 1995).The present paper provides a similar histologic identifica-tion of the premotor burst neurons of the vertical saccadicsystem in human by parvalbumin immunostaining and, inaddition, clearly delineates the iC from riMLF.
MATERIALS AND METHODS
All animal experiments were performed in accordancewith state regulations and with approval of universityanimal care committees.
Eye muscle injection with fragment C
Premotor neurons of the vertical system within therostral mesencephalon were specifically labelled after injec-tions of the nontoxic fragment C of tetanus toxin (TTC;15%) into vertical-pulling eye muscles of macaque mon-keys. Under deep anesthesia (Nembutal; 40 mg/kg of bodyweight) and under sterile conditions, one monkey receivedan injection into the inferior rectus muscle (A4), one in the
Abbreviations
3 third ventricleIII oculomotor nucleusIV trochlear nucleusVI nucleus abducensXII hypoglossal nucleusam anteromedian nucleusBC brachium conjunctivumccn central caudal nucleus of the oculomotor nucleuscm nucleus centromedianus thalamiEBN excitatory burst neuronhb nucleus habenularisiC interstitial nucleus of Cajalio inferior oliveld nucleus lateralis dorsalismb mamillary bodiesmd nucleus medialis dorsalis thalamiMLF fasciculus longitudinalis medialisMT tractus mamillo-thalamicusNIII oculomotor nervenB medial accessory nucleus of BechterewnD nucleus of Darkschewitsch
ol pretectal olivary nucleusPAV parvalbuminPB phosphate bufferPC posterior commissurepcn nucleus paracentralis thalamipf nucleus parafascicularispul pulvinarrn red nucleusriMLF rostral interstitial nucleus of the medial longitudinal fas-
ciclesc superior colliculussf nucleus subfascicularissn substantia nigrast nucleus subthalamicusTR tractus retroflexusTTC tetanus toxin fragment Cvlm nucleus ventralis lateralisvpm nucleus ventralis posterior medialisWGA-HRP wheat germ agglutinin conjugated to horseradish peroxi-
dase
414 A.K.E. HORN AND J.A. BUTTNER-ENNEVER

superior oblique muscle (A5), one in the inferior obliquemuscle (B28), and two animals in the superior rectusmuscle (A9, A3). The preparation of the TTC and thesurgical procedure were described earlier (Buttner-Enn-ever and Akert, 1981; Horn and Buttner-Ennever, 1990).After 4 days of survival time, the animals were killed withan overdose of pentobarbital (Nembutal; 80 mg/kg of bodyweight) and transcardially perfused with saline (37°C)followed by 4% paraformaldehyde in 0.1 M phosphatebuffer (PB), pH 7.4. The brains were post-fixed for another3 to 5 hours at 4°C in the same fixative. Four brains weretransferred through increasing concentrations of sucrose(10–30%) for freeze cutting. Transverse sections (40 µm)were cut with the Vibratome (Oxford Biomedical Research,Oxford, MI) or freezing microtome. One set of free-floatingsections was immunocytochemically treated with a mono-clonal antibody directed against TTC (1:3,000) and stainedwith the avidin-biotin method (Evinger and Erichsen,1986; Horn and Buttner-Ennever, 1990). A series of neigh-bouring sections was stained with cresyl violet.
Double labelling of retrogradely labelledpremotor neurons and parvalbumin
To prove whether parvalbumin immunoreactive neuronswithin the riMLF and iC are premotor neurons, oneanesthetized (Nembutal; 40 mg/kg of body weight) ma-caque monkey (B47) received an injection of 0.4 µl of wheatgerm agglutinin conjugated to horseradish peroxidase(WGA-HRP, 2.5%; Sigma Chemical Co., St. Louis, MO) intothe right oculomotor nucleus. The injection site was lo-cated by microstimulation (3–10 µA) with insulated tung-sten electrodes as described earlier (Horn et al., 1995).After 2 days of survival time, the animal was killed with anoverdose of pentobarbital (80 mg/kg of body weight) andtranscardially perfused with saline (37°C) followed by 4%paraformaldehyde in 0.1 M PB. The brain was equilibratedin increasing concentrations (10–30%) of sucrose for freezecutting. In transverse sections (40 µm) the WGA-HRP wasvisualized with the tetramethyl-benzidine method fol-lowed by the stabilization procedure described in Horn andHoffmann (1987). For the subsequent immunocytochemi-cal detection of parvalbumin on free-floating sections withthe avidin-biotin method, see Horn et al. (1994). Therostral mesencephalon of two monkey brains (B43, Z25)were stained only for PAV.
Parvalbumin immunocytochemistryof the human tissue
For the parvalbumin (PAV)-immunolabelling in human,the brainstems of two humans, who had not suffered froma neurologic disease, were obtained 24 hours postmortem(NP 526/94; GS1317) and immersed in 10% formalin for 6days. The brainstem containing the rostral mesencepha-lon was blocked in 1-cm-thick slices perpendicular to thelong axis of the forebrain, which corresponds approxi-mately to sections in the frontal plane (Fig. 6). The tissuewas embedded in paraffin, and 10-µm serial paraffinsections were cut and processed for the immunocytochemi-cal detection of PAV. After deparaffination and rehydratingin aqua bidest, the sections were incubated in a targetunmasking fluid (TUF, Dianova, Hamburg, Germany)diluted 1:3 in aqua bidest at 95°C for 10 minutes andwashed in aqua bidest for 15 minutes. After treatmentwith 7.5% H2O2, the sections were washed and incubatedin 0.1% Triton-X 100 for 10 minutes followed by blockade of
nonspecific antigenic sites with 2% normal horse serum in0.1 PB for 20 minutes at room temperature. The sectionswere then treated with a monoclonal PAV antibody (Sigma;1:500) at 4°C overnight. After three washes (10 minutes),sections were incubated in biotinylated anti-mouse serum(Vector Laboratories, 1:200) for 30 minutes, followed bythree washes and an 1-hour incubation in streptavidin-biotin complex (Dakopatts, Hamburg, Germany; 1:50).After three washes, the antigenic site was visualized by areaction in 0.1% diaminobenzidine and 0.01% H2O2 for 10minutes. Then the slides were dehydrated and cover-slipped in DEPEX. For orientation and analysis, alternatesets of sections were counterstained with cresyl violet.
Analysis
Labelled neurons were analysed and photographed us-ing a light microscope (Nikon; Biophot). Images of Figures3 and 8 were digitalized by using a cooled 3 CCD videocam-era (Hamamatsu; C5810) mounted on a microscope (LeicaDMRB). The images were captured on a computer (200Pentium Pro) with 4.0 Adobe Photoshop software. Afterconversion into black and white pictures, the sharpness,contrast, and brightness were adjusted only to reflect theappearance of the sections through the light microscope.The figures were printed on a dye sublimation printer(Sony UP-D8800SC). Cell measurements and counts oftrans-synaptically labelled and PAV-immunoreactive neu-rons in the iC and riMLF and of cresyl violet-stainedneurons in corresponding neighbouring sections were per-formed with a graphic tablet (Minimop; Kontron, Eching,Germany).
RESULTS
Labelling of premotor neurons
After the TTC injection into vertical-pulling eye muscles,the motoneurons of the corresponding eye muscles showedstrong direct labelling in the oculomotor or trochlearnuclei. Weaker retrograde trans-synaptic labelling wasfound in all regions, which are known to synapse directlyon inferior rectus, inferior oblique, superior rectus, orsuperior oblique motoneurons, e.g., in the vestibular nucleiincluding the y-group, the interstitial nucleus of Cajal, andthe interstitial nucleus of the medial longitudinal fascicle(Steiger and Buttner-Ennever, 1979). Despite the precau-tions that had been taken, there was always some spreadto neighbouring eye muscles as judged from the directlabelling pattern of the various subgroups in the oculomo-tor, the trochlear nucleus, and abducens nucleus (Buttner-Ennever, 1988). Although in some cases the medial andlateral recti muscles, which pull the eyes in the horizontalplane, were contaminated as well, the retrograde trans-synaptic labelling did not interfere with this study, be-cause their premotor saccadic burst neurons are wellknown and located in the pontine reticular formation(Horn et al., 1995).
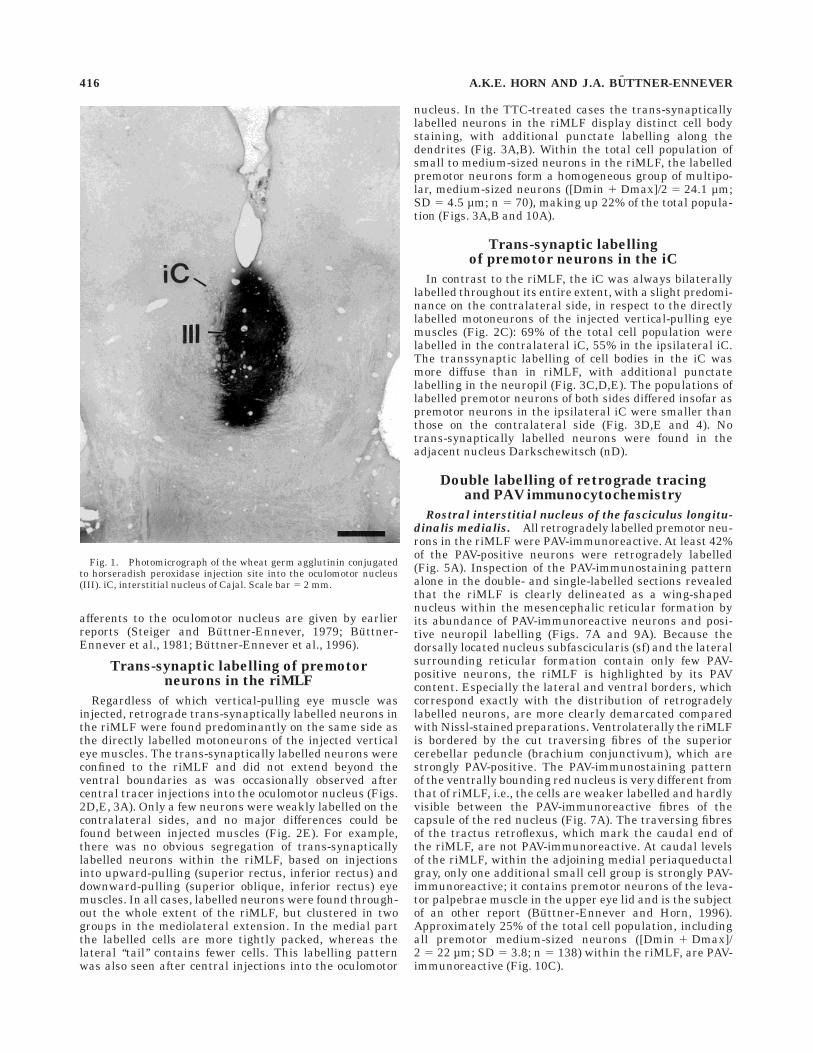
The central WGA-HRP injection site was centered in theright oculomotor nucleus with minor involvement of thesurrounding reticular formation and the left oculomotornucleus (Fig. 1). But the effective uptake area appearedrestricted to the right oculomotor nucleus as judged by thelack of retrograde labelling of internuclear neurons in theright abducens nucleus (Buttner-Ennever andAkert, 1981).
In this paper only the premotor neurons in the iC andriMLF are analysed in detail; extensive descriptions of the
PREMOTOR NEURONS FOR VERTICAL SACCADES IN MONKEY AND MAN 415
afferents to the oculomotor nucleus are given by earlierreports (Steiger and Buttner-Ennever, 1979; Buttner-Ennever et al., 1981; Buttner-Ennever et al., 1996).
Trans-synaptic labelling of premotorneurons in the riMLF
Regardless of which vertical-pulling eye muscle wasinjected, retrograde trans-synaptically labelled neurons inthe riMLF were found predominantly on the same side asthe directly labelled motoneurons of the injected verticaleye muscles. The trans-synaptically labelled neurons wereconfined to the riMLF and did not extend beyond theventral boundaries as was occasionally observed aftercentral tracer injections into the oculomotor nucleus (Figs.2D,E, 3A). Only a few neurons were weakly labelled on thecontralateral sides, and no major differences could befound between injected muscles (Fig. 2E). For example,there was no obvious segregation of trans-synapticallylabelled neurons within the riMLF, based on injectionsinto upward-pulling (superior rectus, inferior rectus) anddownward-pulling (superior oblique, inferior rectus) eyemuscles. In all cases, labelled neurons were found through-out the whole extent of the riMLF, but clustered in twogroups in the mediolateral extension. In the medial partthe labelled cells are more tightly packed, whereas thelateral ‘‘tail’’ contains fewer cells. This labelling patternwas also seen after central injections into the oculomotor
nucleus. In the TTC-treated cases the trans-synapticallylabelled neurons in the riMLF display distinct cell bodystaining, with additional punctate labelling along thedendrites (Fig. 3A,B). Within the total cell population ofsmall to medium-sized neurons in the riMLF, the labelledpremotor neurons form a homogeneous group of multipo-lar, medium-sized neurons ([Dmin 1 Dmax]/2 5 24.1 µm;SD 5 4.5 µm; n 5 70), making up 22% of the total popula-tion (Figs. 3A,B and 10A).
Trans-synaptic labellingof premotor neurons in the iC
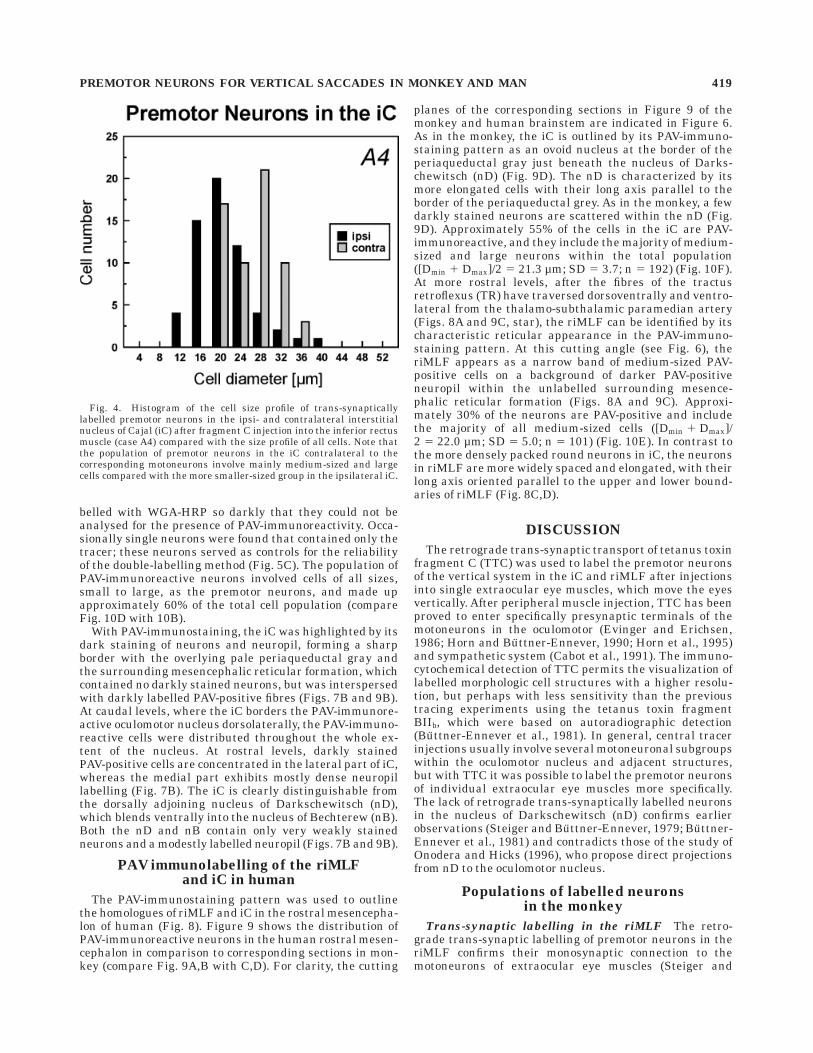
In contrast to the riMLF, the iC was always bilaterallylabelled throughout its entire extent, with a slight predomi-nance on the contralateral side, in respect to the directlylabelled motoneurons of the injected vertical-pulling eyemuscles (Fig. 2C): 69% of the total cell population werelabelled in the contralateral iC, 55% in the ipsilateral iC.The transsynaptic labelling of cell bodies in the iC wasmore diffuse than in riMLF, with additional punctatelabelling in the neuropil (Fig. 3C,D,E). The populations oflabelled premotor neurons of both sides differed insofar aspremotor neurons in the ipsilateral iC were smaller thanthose on the contralateral side (Fig. 3D,E and 4). Notrans-synaptically labelled neurons were found in theadjacent nucleus Darkschewitsch (nD).
Double labelling of retrograde tracingand PAV immunocytochemistry
Rostral interstitial nucleus of the fasciculus longitu-
dinalis medialis. All retrogradely labelled premotor neu-rons in the riMLF were PAV-immunoreactive. At least 42%of the PAV-positive neurons were retrogradely labelled(Fig. 5A). Inspection of the PAV-immunostaining patternalone in the double- and single-labelled sections revealedthat the riMLF is clearly delineated as a wing-shapednucleus within the mesencephalic reticular formation byits abundance of PAV-immunoreactive neurons and posi-tive neuropil labelling (Figs. 7A and 9A). Because thedorsally located nucleus subfascicularis (sf) and the lateralsurrounding reticular formation contain only few PAV-positive neurons, the riMLF is highlighted by its PAVcontent. Especially the lateral and ventral borders, whichcorrespond exactly with the distribution of retrogradelylabelled neurons, are more clearly demarcated comparedwith Nissl-stained preparations. Ventrolaterally the riMLFis bordered by the cut traversing fibres of the superiorcerebellar peduncle (brachium conjunctivum), which arestrongly PAV-positive. The PAV-immunostaining patternof the ventrally bounding red nucleus is very different fromthat of riMLF, i.e., the cells are weaker labelled and hardlyvisible between the PAV-immunoreactive fibres of thecapsule of the red nucleus (Fig. 7A). The traversing fibresof the tractus retroflexus, which mark the caudal end ofthe riMLF, are not PAV-immunoreactive. At caudal levelsof the riMLF, within the adjoining medial periaqueductalgray, only one additional small cell group is strongly PAV-immunoreactive; it contains premotor neurons of the leva-tor palpebrae muscle in the upper eye lid and is the subjectof an other report (Buttner-Ennever and Horn, 1996).Approximately 25% of the total cell population, includingall premotor medium-sized neurons ([Dmin 1 Dmax]/2 5 22 µm; SD 5 3.8; n 5 138) within the riMLF, are PAV-immunoreactive (Fig. 10C).
Fig. 1. Photomicrograph of the wheat germ agglutinin conjugatedto horseradish peroxidase injection site into the oculomotor nucleus(III). iC, interstitial nucleus of Cajal. Scale bar 5 2 mm.
416 A.K.E. HORN AND J.A. BUTTNER-ENNEVER
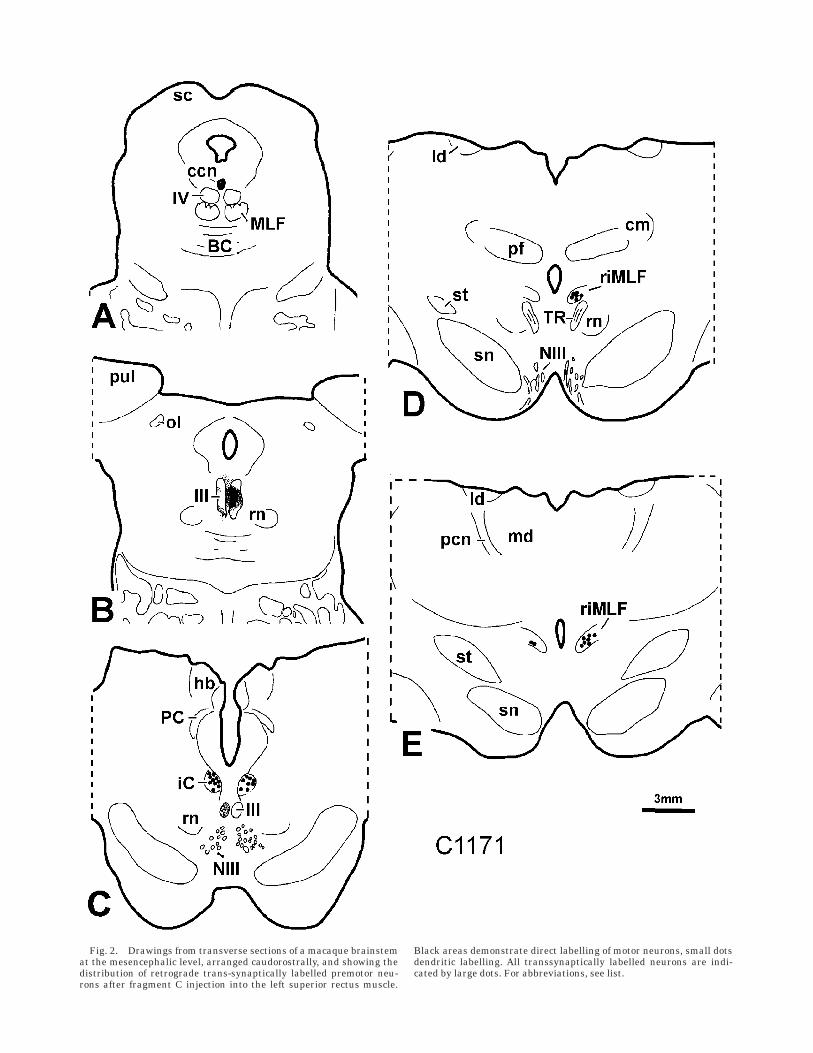
Fig. 2. Drawings from transverse sections of a macaque brainstemat the mesencephalic level, arranged caudorostrally, and showing thedistribution of retrograde trans-synaptically labelled premotor neu-rons after fragment C injection into the left superior rectus muscle.
Black areas demonstrate direct labelling of motor neurons, small dotsdendritic labelling. All transsynaptically labelled neurons are indi-cated by large dots. For abbreviations, see list.

Interstitial nucleus of Cajal. At least 90% of theretrogradely labelled neurons were PAV-immunoreactive,and approximately 50% of the PAV-positive cells in theipsilateral and 61% in the contralateral iC were retro-
gradely labelled (Fig. 5B). These percentages must beconsidered as minimum values, because some of theretrograde labelling might be lost at the stabilization stepfor the double-labelling protocol. Some neurons were la-
Fig. 3. Photomicrograph showing the pattern of the trans-synapticlabelling in rostral interstitial nucleus of the medial longitudinalfascicle (riMLF) (A,B) and the interstitial nucleus of Cajal (iC) (C–E).A: In the riMLF, the premotor neurons are more distinctly stainedwith punctate labelling confined to the dendrites. The star indicatesthe thalamo-subthalamic paramedian artery. The arrow points to atrans-synaptically labelled premotor neuron, which is shown in high-
power magnification in B. C: Trans-synaptic labelling of the iCipsilateral to the labelled motoneurons. The iC shows more diffuse cellbody staining compared with riMLF, with strong additional labeling ofthe neuropil (C,D). Note that additional large cells are labelled in thecontralateral iC (E, arrows) compared with the smaller cells ipsilat-eral iC (D, arrows). Scale bars 5 30 µm in B,E (applies to B,D,E), 250µm in C (applies to A,C).
418 A.K.E. HORN AND J.A. BUTTNER-ENNEVER
belled with WGA-HRP so darkly that they could not beanalysed for the presence of PAV-immunoreactivity. Occa-sionally single neurons were found that contained only thetracer; these neurons served as controls for the reliabilityof the double-labelling method (Fig. 5C). The population ofPAV-immunoreactive neurons involved cells of all sizes,small to large, as the premotor neurons, and made upapproximately 60% of the total cell population (compareFig. 10D with 10B).
With PAV-immunostaining, the iC was highlighted by itsdark staining of neurons and neuropil, forming a sharpborder with the overlying pale periaqueductal gray andthe surrounding mesencephalic reticular formation, whichcontained no darkly stained neurons, but was interspersedwith darkly labelled PAV-positive fibres (Figs. 7B and 9B).At caudal levels, where the iC borders the PAV-immunore-active oculomotor nucleus dorsolaterally, the PAV-immuno-reactive cells were distributed throughout the whole ex-tent of the nucleus. At rostral levels, darkly stainedPAV-positive cells are concentrated in the lateral part of iC,whereas the medial part exhibits mostly dense neuropillabelling (Fig. 7B). The iC is clearly distinguishable fromthe dorsally adjoining nucleus of Darkschewitsch (nD),which blends ventrally into the nucleus of Bechterew (nB).Both the nD and nB contain only very weakly stainedneurons and a modestly labelled neuropil (Figs. 7B and 9B).
PAV immunolabelling of the riMLFand iC in human
The PAV-immunostaining pattern was used to outlinethe homologues of riMLF and iC in the rostral mesencepha-lon of human (Fig. 8). Figure 9 shows the distribution ofPAV-immunoreactive neurons in the human rostral mesen-cephalon in comparison to corresponding sections in mon-key (compare Fig. 9A,B with C,D). For clarity, the cutting
planes of the corresponding sections in Figure 9 of themonkey and human brainstem are indicated in Figure 6.As in the monkey, the iC is outlined by its PAV-immuno-staining pattern as an ovoid nucleus at the border of theperiaqueductal gray just beneath the nucleus of Darks-chewitsch (nD) (Fig. 9D). The nD is characterized by itsmore elongated cells with their long axis parallel to theborder of the periaqueductal grey. As in the monkey, a fewdarkly stained neurons are scattered within the nD (Fig.9D). Approximately 55% of the cells in the iC are PAV-immunoreactive, and they include the majority of medium-sized and large neurons within the total population([Dmin 1 Dmax]/2 5 21.3 µm; SD 5 3.7; n 5 192) (Fig. 10F).At more rostral levels, after the fibres of the tractusretroflexus (TR) have traversed dorsoventrally and ventro-lateral from the thalamo-subthalamic paramedian artery(Figs. 8A and 9C, star), the riMLF can be identified by itscharacteristic reticular appearance in the PAV-immuno-staining pattern. At this cutting angle (see Fig. 6), theriMLF appears as a narrow band of medium-sized PAV-positive cells on a background of darker PAV-positiveneuropil within the unlabelled surrounding mesence-phalic reticular formation (Figs. 8A and 9C). Approxi-mately 30% of the neurons are PAV-positive and includethe majority of all medium-sized cells ([Dmin 1 Dmax]/2 5 22.0 µm; SD 5 5.0; n 5 101) (Fig. 10E). In contrast tothe more densely packed round neurons in iC, the neuronsin riMLF are more widely spaced and elongated, with theirlong axis oriented parallel to the upper and lower bound-aries of riMLF (Fig. 8C,D).
DISCUSSION
The retrograde trans-synaptic transport of tetanus toxinfragment C (TTC) was used to label the premotor neuronsof the vertical system in the iC and riMLF after injectionsinto single extraocular eye muscles, which move the eyesvertically. After peripheral muscle injection, TTC has beenproved to enter specifically presynaptic terminals of themotoneurons in the oculomotor (Evinger and Erichsen,1986; Horn and Buttner-Ennever, 1990; Horn et al., 1995)and sympathetic system (Cabot et al., 1991). The immuno-cytochemical detection of TTC permits the visualization oflabelled morphologic cell structures with a higher resolu-tion, but perhaps with less sensitivity than the previoustracing experiments using the tetanus toxin fragmentBIIb, which were based on autoradiographic detection(Buttner-Ennever et al., 1981). In general, central tracerinjections usually involve several motoneuronal subgroupswithin the oculomotor nucleus and adjacent structures,but with TTC it was possible to label the premotor neuronsof individual extraocular eye muscles more specifically.The lack of retrograde trans-synaptically labelled neuronsin the nucleus of Darkschewitsch (nD) confirms earlierobservations (Steiger and Buttner-Ennever, 1979; Buttner-Ennever et al., 1981) and contradicts those of the study ofOnodera and Hicks (1996), who propose direct projectionsfrom nD to the oculomotor nucleus.
Populations of labelled neuronsin the monkey
Trans-synaptic labelling in the riMLF The retro-grade trans-synaptic labelling of premotor neurons in theriMLF confirms their monosynaptic connection to themotoneurons of extraocular eye muscles (Steiger and
Fig. 4. Histogram of the cell size profile of trans-synapticallylabelled premotor neurons in the ipsi- and contralateral interstitialnucleus of Cajal (iC) after fragment C injection into the inferior rectusmuscle (case A4) compared with the size profile of all cells. Note thatthe population of premotor neurons in the iC contralateral to thecorresponding motoneurons involve mainly medium-sized and largecells compared with the more smaller-sized group in the ipsilateral iC.
PREMOTOR NEURONS FOR VERTICAL SACCADES IN MONKEY AND MAN 419
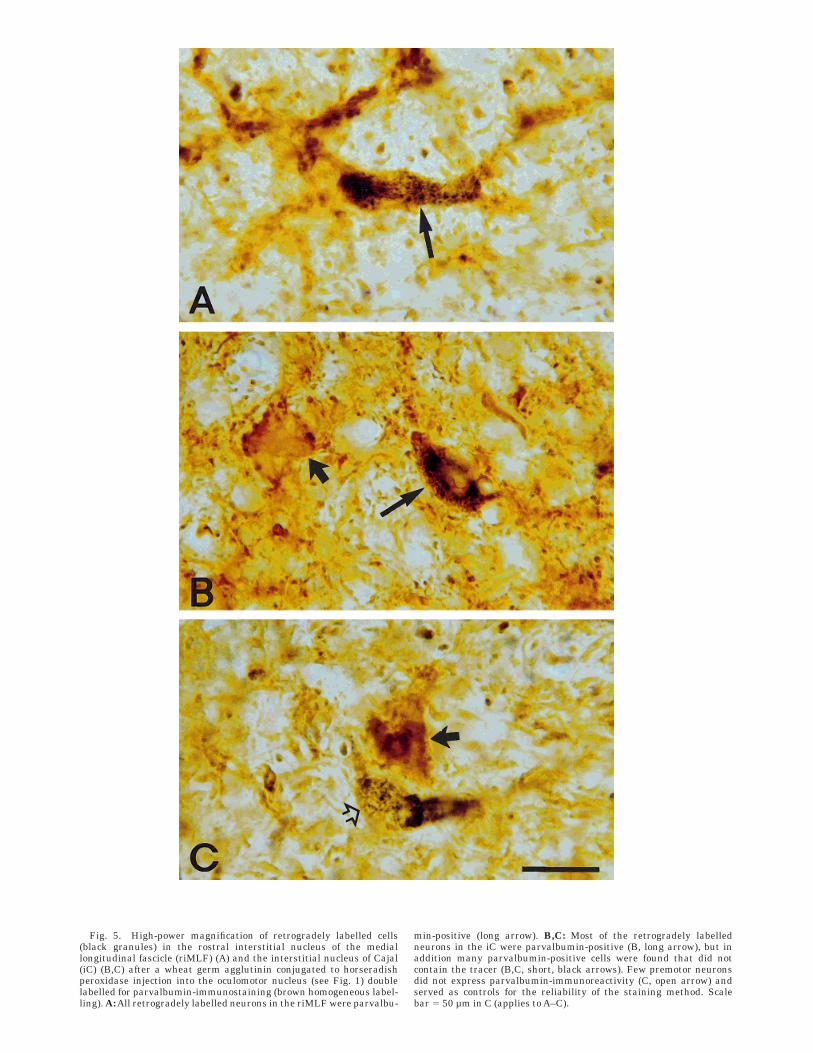
Fig. 5. High-power magnification of retrogradely labelled cells(black granules) in the rostral interstitial nucleus of the mediallongitudinal fascicle (riMLF) (A) and the interstitial nucleus of Cajal(iC) (B,C) after a wheat germ agglutinin conjugated to horseradishperoxidase injection into the oculomotor nucleus (see Fig. 1) doublelabelled for parvalbumin-immunostaining (brown homogeneous label-ling). A: All retrogradely labelled neurons in the riMLF were parvalbu-
min-positive (long arrow). B,C: Most of the retrogradely labelledneurons in the iC were parvalbumin-positive (B, long arrow), but inaddition many parvalbumin-positive cells were found that did notcontain the tracer (B,C, short, black arrows). Few premotor neuronsdid not express parvalbumin-immunoreactivity (C, open arrow) andserved as controls for the reliability of the staining method. Scalebar 5 50 µm in C (applies to A–C).

Buttner-Ennever, 1979; Moschovakis et al., 1991a, 1991b,Wang and Spencer, 1996a). The morphometric analysisshows that the majority of, if not all, medium-sized neu-rons within the riMLF are premotor neurons, taking intoaccount that not all vertical premotor neurons are labelledfrom the TTC injection in one single experiment, that is,the burst neurons encoding vertical and torsional saccades
(Buttner et al., 1977; King and Fuchs, 1979; Vilis et al.,1989). The predominantly ipsilateral projection to thecorresponding motoneurons is in accordance with otherstudies applying central tracer injections into the oculomo-tor nucleus (Steiger and Buttner-Ennever, 1979; Wang andSpencer, 1996b). Additional weakly labelled cells werefound in the contralateral riMLF in all cases. Because
Fig. 6. Schematic sagittal sections through a macaque (left) and ahuman (right) brain to show the planes of cutting (dashed lines) for thetransverse sections in Figures 9 (A,B) and the photographs in Figures7 and 8. The cutting plane (dashed line) of the human rostral
brainstem is approximately perpendicular to the long axis of theforebrain (solid line 0°), which forms an angle of 60° (solid line 60°)with the long axis of the brainstem. ri, rostral interstitial nucleus ofmedial longitudinal fascicle. For other abbreviations, see list.
Fig. 7. Photomicrographs of transverse sections through the ros-tral mesencephalon of a macaque monkey (see also Fig. 6, left)immunocytochemically labelled with parvalbumin (PAV) antibodies.A: The rostral interstitial nucleus of the medial longitudinal fascicle(riMLF), dorsally bordered by the thalamo-subthalamic paramedianartery (star), is clearly delineated within the mesencephalic reticularformation by its high content of parvalbumin. The parvalbumin-
positive neurons in the riMLF are elongated with long dendrites.B: The interstitial nucleus of Cajal (iC) is also highlighted by its PAVcontent, the labelled cells are round and medium-sized. Note that theadjacent nucleus of Darkschewitsch is pale and contains only fewstrong PAV-immunoreactive neurons. Scale bar 5 500 µm in B (appliesto A).
PREMOTOR NEURONS FOR VERTICAL SACCADES IN MONKEY AND MAN 421

tracer involvement of neighbouring eye muscle motoneu-rons could not be avoided, the contralateral trans-synapticlabelling in our cases could at least partly be due tolabelling of premotor neurons of motoneuronal groupswith crossing axons, i.e., superior oblique or superiorrectus muscle. Based on the reconstructions of physiologi-cally characterized intracellularly HRP-filled riMLF-burstneurons (Moschovakis et al., 1991a,b), the authors proposebilateral projections of upward burst neurons to the oculo-motor nucleus vs. purely ipsilateral projections of down-ward burst neurons to the oculomotor and trochlear nucleusin the squirrel monkey. Although the authors found bilat-eral terminal labelling from upward burst neurons withinthe oculomotor nucleus, their conclusion of a functionalbilateral projection needs more proof, because the motoneu-rons of the superior rectus muscle of one eye are locatedbilaterally around the midline as well (Buttner-Ennever,1988). Unilateral stimulation of upward burst neuronsdoes not result in purely upward eye movements withouttorsion, which suggests at least a weaker efficacy of thecontralateral projection, if it is functionally existent (Craw-ford and Vilis, 1992). A possible weaker activity of thecontralateral projections of upward burst neurons to themotoneurons in the oculomotor nucleus could have af-
fected the trans-synaptic tracer uptake into the premotorterminals, which would explain the weak labelling and thelow number of trans-synaptically labelled neurons in thecontralateral riMLF in the present study (Wellhorner etal., 1973).
The single eye muscle injections of TTC did not reveal anobvious spatial segregation between upward premotorneurons that innervate superior rectus and/or inferioroblique motoneurons, and downward premotor neuronsthat innervate superior oblique and/or inferior rectusmotoneurons within the riMLF, which could form theneuroanatomical basis for an isolated upgaze or downgazeparalysis in patients (Cogan, 1974; Pierrot-Deseilligny etal., 1982; Buttner-Ennever et al., 1982; Ranalli et al.,1988). Our findings are in accordance with physiologicrecording experiments in monkey and cat that foundupward and downward burst neurons intermingled withinthe riMLF (Buttner et al., 1977; King and Fuchs, 1979;Moschovakis et al., 1991a,b; Li et al., 1993). Anatomicalexperiments in cats revealed a tendency of upward neu-rons lying more caudally and downward neurons morerostrally (Wang and Spencer, 1996b). The authors explainthe discrepancy of physiologic and anatomical results withthe existence of excitatory and inhibitory premotor neu-
Fig. 8. Microphotographs of transverse sections through a humanbrainstem (see Fig. 6) comparable to Figure 7, immunocytochemicallylabelled with parvalbumin (PAV) antibodies. A,C: The rostral intersti-tial nucleus of the medial longitudinal fascicle (riMLF) is dorsallybordered by the thalamo-subthalamic paramedian artery (star) and isoutlined by the PAV-immunolabelling of neurons and the neuropil. The
labelled neurons, the presumed premotor burst neurons (arrows), areelongated and oriented parallel to the mediolateral axis of the riMLF.B,D: The interstitial nucleus of Cajal (iC) is highlighted by its PAVcontent and forms a compact nucleus. The labelled neurons (arrows)are round and more densely packed. Scale bar 5 500 µm in B (appliesto A), 30 µm in D (applies to C).
422 A.K.E. HORN AND J.A. BUTTNER-ENNEVER

Fig. 9. Drawings from transverse sections through the interstitialnucleus of Cajal (iC) (A,B) and rostral interstitial nucleus of themedial longitudinal fascicle (riMLF) (C,D) of monkey (A,C) and man(B,D) at the planes shown in Figure 6 and showing the distribution ofparvalbumin (PAV)-immunoreactive neurons. Each dot indicates onePAV-immunoreactive neurons. The iC (A,B) and riMLF (C,D) in
monkey (A,C) and man (B,D) are outlined within the mesencephalicreticular formation by their relative abundance of PAV-immunoreac-tive cells. In both species, the riMLF forms a wing-shaped nucleusdorsally bordered by the thalamo-subthalamic paramedian artery(star) (A,C). For abbreviations, see list.
PREMOTOR NEURONS FOR VERTICAL SACCADES IN MONKEY AND MAN 423
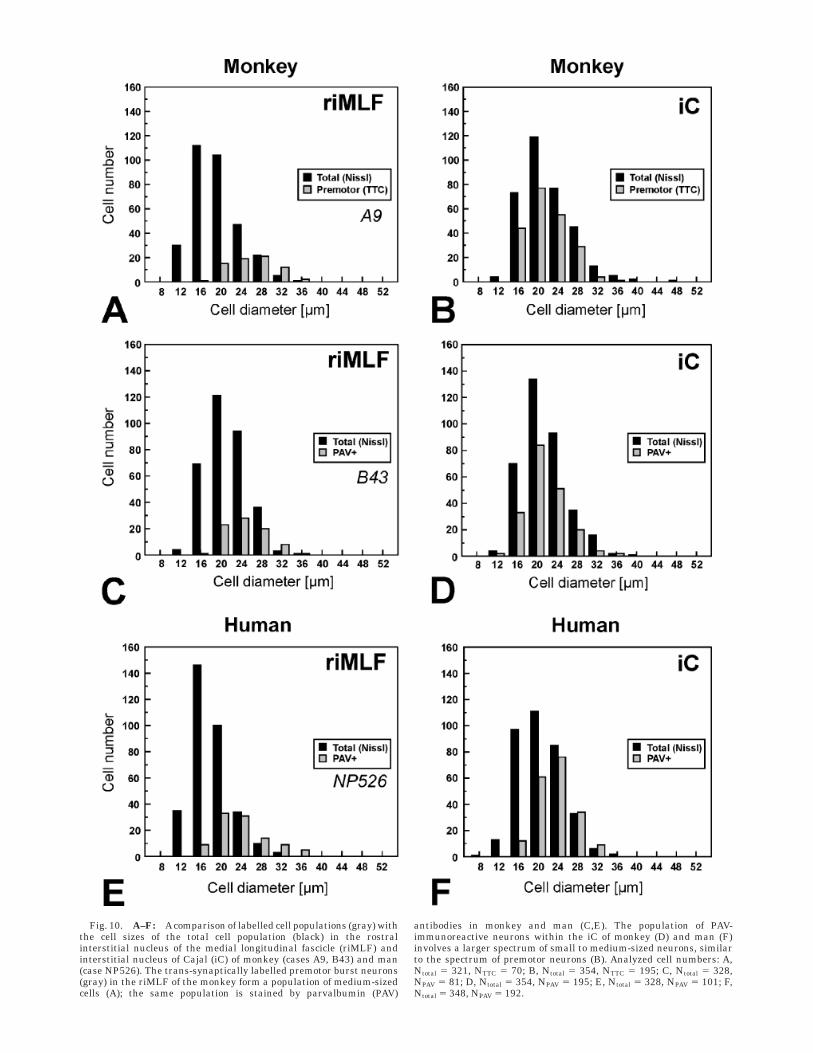
Fig. 10. A–F: Acomparison of labelled cell populations (gray) withthe cell sizes of the total cell population (black) in the rostralinterstitial nucleus of the medial longitudinal fascicle (riMLF) andinterstitial nucleus of Cajal (iC) of monkey (cases A9, B43) and man(case NP526). The trans-synaptically labelled premotor burst neurons(gray) in the riMLF of the monkey form a population of medium-sizedcells (A); the same population is stained by parvalbumin (PAV)
antibodies in monkey and man (C,E). The population of PAV-immunoreactive neurons within the iC of monkey (D) and man (F)involves a larger spectrum of small to medium-sized neurons, similarto the spectrum of premotor neurons (B). Analyzed cell numbers: A,Ntotal 5 321, NTTC 5 70; B, Ntotal 5 354, NTTC 5 195; C, Ntotal 5 328,NPAV 5 81; D, Ntotal 5 354, NPAV 5 195; E, Ntotal 5 328, NPAV 5 101; F,Ntotal 5 348, NPAV 5 192.

rons with the same projection targets, but with oppositemoving vectors. Evidence for the existence of inhibitorypremotor neurons in the riMLF is not strong. It comesfrom stimulation studies that evoke monosynaptic inhibi-tory postsynaptic potentials in the trochlear (Schwindt etal., 1974) and oculomotor nucleus (Nakao and Shiraishi,1983, 1985), although it cannot be excluded that iCneurons were stimulated. Anterograde and retrogradedouble-labelling experiments revealed GABAergic neu-rons in the riMLF that project to the motoneurons in theoculomotor and trochlear nuclei of the cat (Spencer andWang, 1996), although no such GABAergic projection wasseen in monkey (Carpenter et al., 1992).
Trans-synaptic labelling in the iC. The retrogradetrans-synaptic TTC labelling of neurons within the iCconfirms its monosynaptic input to the motoneurons ofvertical-pulling extraocular muscles (Schwindt et al., 1974;Steiger and Buttner-Ennever, 1979; Buttner-Ennever etal., 1981; Nakao and Shiraishi, 1985). In contrast toriMLF, the iC of both sides were strongly labelled aftersingle muscle injections with TTC. This observation is inaccordance with physiologic experiments (Schwindt et al.,1974) and recent anatomical studies applying biocytin andPhaseolus-lectin as anterograde tracers in the monkey(Kokkoroyannis et al., 1996). Electrophysiologic recordingrevealed several functional cell groups within the iC inalert cats and monkeys: burst-tonic, tonic, saccade-relatedburst neurons, burster-driving neurons, and vestibularneurons (for review, see Fukushima et al., 1992; Helmchenet al., 1996a). Unfortunately no conclusive correlativestudies of functional cell groups within the iC and theirprojection targets are available so far. The tonic andburst-tonic neurons encode the eye position in the verticalplane, and it is assumed that they receive afferents fromthe burst neurons within the riMLF (Moschovakis et al.,1991a, 1991b) and project monosynaptically to the moto-neurons of vertical eye muscles, implying their involve-ment in the vertical integrator function (review in Fuku-shima et al., 1992). The function of saccade-related burstneurons within the iC is not fully understood yet (Helm-chen et al., 1996a), but apparently they do not project toeye muscle motoneurons (Moschovakis et al., 1991b).Based on these studies the labelled premotor neurons inthe iC of our experiments most probably include tonic andburst-tonic neurons.
Anatomically, the iC contains two main classes of neu-rons: small to medium-sized neurons and a small popula-tion of large multipolar cells (Carpenter and Peter, 1970;Zuk et al., 1982). Interestingly, the present anatomicalresults revealed differences between the premotor popula-tions in the iC of both sides: more premotor neurons, 69%,were labelled in the contralateral iC, including additionallarger-sized cells, compared with the ipsilateral premotorneurons, which made up 55% of the total cell population ofthe iC.
Anterograde tracing experiments showed that efferentfibres of the iC to the contralateral oculomotor and troch-lear nucleus run through the posterior commissure (Kok-koroyannis et al., 1996). A lesion of the posterior commis-sure results in the inability to hold eccentric gaze aftervertical saccades (Partsalis et al., 1994). In comparisonwith the horizontal saccadic system, the correlate ofinhibitory burst neurons, which inhibit the motoneuronsof the antagonistic eye muscles during vertical saccades isnot known so far. Both the riMLF and the iC are possiblelocations, which remain to be studied in the monkey.
Another hypothesis suggests the existence of vertical‘‘internuclear’’ neurons in the iC, which should activate themotoneurons of agonistic eye muscles during verticalsaccades, similar to that of the horizontal system (Buttner-Ennever and Buttner, 1988). Taken together, the presentresults suggest at least partly functional different monosyn-aptic inputs to eye muscle motoneurons from the iC of bothsides.
PAV-immunoreactivity of premotor neurons in the
riMLF. The present study confirms the observation madein cat and rat that the riMLF is highlighted with PAV-immunostaining (Spencer and Wang, 1996; Celio 1990).Moreover, it is shown that the premotor saccadic burstneurons themselves in the riMLF are PAV immunoreac-tive, and this finding was taken as evidence that parvalbu-min can be used as a marker for premotor burst neurons.These results are in accordance with the notion that thecalcium-binding protein, parvalbumin, is associated withfast-firing neurons and cells with high oxidative metabo-lism (Celio, 1990). The premotor saccadic neurons in theriMLF exhibit a high-frequency burst of up to 600 spikes/second, shortly before and during saccades, depending ontheir amplitude and direction (Buttner et al., 1977). Al-though it cannot be ruled out that some of the premotorneurons in the riMLF are GABAergic (Spencer and Wang,1996), this is another example of parvalbumin associatedwith excitatory as well as inhibitory neurons in thebrainstem (Horn et al., 1994; Horn et al., 1995).
PAV-immunoreactivity of premotor neurons in the
iC. Approximately 60% of the total cell population withinthe iC are PAV immunoreactive, including two popula-tions, e.g., double-labelled premotor neurons and thosecontaining only PAV immunoreactivity. The double-labelled premotor neurons make up at least 61% of thePAV-immunoreactive cell population, as judged from thecontralateral projection, and most probably constitute thetonic and burst-tonic neurons judging by their projectionto the oculomotor neurons as mentioned above. The parval-bumin content of these neurons can be explained by theirfiring properties. In the monkey, burst-tonic neurons dis-charge a burst of spikes during a saccade and show asteady firing, depending on the eye position during fixa-tion; tonic neurons lack the saccade-related burst (forreview: Fukushima et al., 1992). Based on the assumptionthat parvalbumin is also expressed in cells with high firingfrequencies, the PAV-only cell population might includethe saccade-related burst neurons, which exhibit a similardischarge pattern with more than 600 spikes/second likethe PAV-immunoreactive premotor burst neurons withinthe riMLF. The saccade-related burst neurons are inter-mingled with tonic and burst-tonic neurons and constituteat least one-third of eye movement-related cells within theiC (Helmchen et al., 1996a). In contrast to tonic andburst-tonic neurons, they do not project to oculomotorneurons and are thought to relay an inhibitory feedbacksignal to the riMLF (Moschovakis et al., 1991b, 1996).
Identification of vertical premotorneurons in human
Although the locations of the riMLF and iC in humanare already described in earlier papers based on Nissl andfibre stainings, it was also clear from this paper that therostral part of the iC is very similar in its cellularappearance in Nissl-stained sections to the caudal riMLF(Buttner-Ennever et al., 1982). Both nuclei appear reticu-lated by the interspersed fibres of the MLF and are
PREMOTOR NEURONS FOR VERTICAL SACCADES IN MONKEY AND MAN 425

roughly separated from each other by the traversing fibresof the tractus retroflexus (TR). In addition, the riMLF isdorsally bordered by the thalamo-subthalamic parame-dian artery. All these landmarks are helpful but do notexactly define the location of iC and riMLF and dependstrongly of the plane of the section. That the long axis ofthe brainstem and of the forebrain form an angle ofapproximately 60° (Fig. 6), the cutting angle of the rostralmesencephalon can vary greatly. For this study, we chosean angle of section roughly perpendicular to the long axisof the forebrain, which corresponds most closely to theappearance of the critical structures in transverse sectionsof the macaque brain. The differential PAV-immunolabel-ling pattern of the iC and riMLF allows a more accuratedelineation of both nuclei from each other (independent ofthe plane of section): the riMLF contains more scatteredelongated PAV-immunoreactive cells, which are orientedparallel to the surface of the nucleus; the iC consists ofmore densely packed, round and randomly oriented PAV-immunoreactive neurons.
Comparison of premotor burst neurons inthe horizontal and vertical saccadic system
Retrograde trans-synaptically labelling in the riMLFrevealed a portion of approximately 22% of the total cellpopulation as premotor neurons of one injected verticalpulling eye muscle. A similar portion of 20% had beenlabelled using the same technique within the excitatoryburst neuron (EBN) area of the horizontal saccadic system(Horn et al., 1995). Whereas premotor inhibitory burstneurons of the horizontal system lie in a separate nucleus,the nucleus paragigantocellularis dorsalis, the location ofvertical inhibitory burst neurons within the riMLF cannotbe ruled out and might be included in the labelled premo-tor population in riMLF (Spencer and Wang, 1996). Takinginto consideration that the riMLF contains at least twodifferent sets of premotor neurons for two agonistic pairs ofeye muscles, the total portion of premotor neurons withinriMLF might be expected to be at least two times larger. Inthe horizontal and vertical saccadic system, all premotorburst neurons express PAV-immunoreactivity, which canbe used as a histologic marker of premotor burst neuronsin both systems. Similar large portions, 25% in the riMLFand 30% in the EBN area, within the total cell populationcontain PAV, but should be considered as minimum valuesdependent on the penetration capabilities of the antibodiesused in the immunocytochemical staining. Within thePAV-immunoreactive neuron populations, 65% were doublelabelled in the EBN area of the horizontal saccadic systemand 42% in the riMLF after tracer injection in the abdu-cens and oculomotor nucleus, respectively. The differencesin the double-labelled portions of both systems are mostprobably due to methologic reasons: first, the tracer injec-tion into the oculomotor nucleus did not involve all moto-neurons (the trochlear nucleus was not involved at all);second, different tracer and double-labelling procedureswere used. The double-labelling of premotor neurons in thehorizontal system was based on injections with choleratoxin subunit B and two consecutive immunocytochemicalprotocols. The double-labelling of premotor neurons in thevertical system in the present paper was based on injec-tions with WGA-HRP, which was visualized with thetetramethyl-benzidine method, followed by the immunocy-tochemical detection of PAV. Some of the retrograde label-ling might be lost at the stabilization step for the double-labelling protocol in the latter case (see Materials and
Methods section). In general, the PAV-immunoreactivepopulation of the riMLF appears functionally more homo-geneous representing mostly premotor burst neurons forthe vertical saccadic system. In contrast, the nucleusreticularis pontis caudalis (nrpc) of the paramedian pon-tine reticular formation in monkey contains only one set ofocular premotor neurons, those of the lateral rectus muscle.In addition, there are other functional oculomotor cellgroups, such as reticulospinal neurons or long-lead burstneurons, which might be PAV-immunoreactive as well(Robinson et al., 1994; Horn et al., 1995; Scudder et al.,1996).
In conclusion, the present paper shows that in additionto the previously described omnipause neurons and thehorizontal burst neurons, two other premotor neuronalpopulations of the saccadic system can be identified inhuman with the help of PAV-immunocytochemistry (Hornet al., 1994, 1995). It is now possible in human postmortemmaterial to delineate riMLF and iC clearly from eachother; the results form the neuroanatomic basis for anaccurate neuropathologic analysis of the vertical eye-movement–related structures in the rostral mesencepha-lon. Moreover, it is possible to perform an analysis at thecellular level, because the saccadic premotor neurons areprobably all medium-sized PAV-positive neurons in theriMLF. Experimental and clinical observations suggestdifferent deficits in the torsional plane after unilateral iCor unilateral riMLF lesions (Halmagyi et al., 1994; Helm-chen et al., 1996b; Riordan-Eva et al., 1996), which wouldserve as a useful tool for the topographic diagnosis inpatients by appropriate eye-movement recordings. Withthe results of this paper, it is now possible to test thehypothesis and perhaps establish a useful diagnostic toolfor lesions of the rostral mesencephalon.
ACKNOWLEDGMENTS
We are grateful to Prof. P. Mehraein for his valuablesupport. We thank Drs. U. Buttner, B. Cohen, and V. Henn,who allowed us to process the brainstem tissue of theirexperimental animals, which helped us to reduce thenumber of experimental animals in this study. We thankU. Schneider, M. Phil, and S. Messoudi for their excellenttechnical assistance.
LITERATURE CITED
Buttner, U., J.A. Buttner-Ennever, and V. Henn (1977) Vertical eyemovement related activity in the rostral mesencephalic reticular forma-tion of the alert monkey. Brain Res. 130:239–252.
Buttner-Ennever, J.A. (1988) Anatomy of the ocular motor nuclei. In C.Kennard and F.C. Rose (eds): Physiological Aspects of Clinical Neuro-Ophthalmology. London: Chapman and Hall, pp. 199–208.
Buttner-Ennever, J.A. and K. Akert (1981) Medial rectus subgroups of theoculomotor nucleus and their abducens internuclear input in themonkey. J. Comp. Neurol. 197:17–27.
Buttner-Ennever, J.A. and U. Buttner (1978) A cell group associated withvertical eye movements in the rostral mesencephalic reticular forma-tion of the monkey. Brain Res. 151:31–47.
Buttner-Ennever, J.A. and U. Buttner (1988) Neuroanatomy of the oculomo-tor system. The reticular formation. Rev. Oculomot. Res. 2:119–176.
Buttner-Ennever, J.A., P. Grob, K. Akert, and B. Bizzini (1981) Transsynap-tic retrograde labeling in the oculomotor system of the monkey withtetanus toxin BIIb fragment. Neurosci. Lett. 26:233–238.
Buttner-Ennever, J.A., U. Buttner, B. Cohen, and G. Baumgartner (1982)Vertical gaze paralysis and the rostral interstitial nucleus of the mediallongitudinal fasciculus. Brain 105:125–149.
Buttner-Ennever, J.A., B. Cohen, A.K.E. Horn, and H. Reisine (1996)Pretectal projections to the oculomotor complex of the monkey and theirrole in eye movements. J. Comp. Neurol. 366:348–359.
426 A.K.E. HORN AND J.A. BUTTNER-ENNEVER

Buttner-Ennever, J.A. and A.K.E. Horn (1996) Upper eyelid premotorneurons in the rostral mesencephalon of the primate. Soc. Neurosci.Abstr. 22:2035.
Cabot, J.B., A. Mennone, N. Bogan, J. Carrol, C. Evinger, and J.T. Erichsen(1991) Retrograde, trans-synaptic and transneuronal transport of frag-ment C of tetanus toxin by sympathetic preganglionic neurons. Neuro-science 40:805–823.
Carpenter, M.B. and P. Peter (1970) Accessory oculomotor nuclei in themonkey. J. Hirnforsch. 12:405–418.
Carpenter, M.B., A.B. Periera, and N. Guha (1992) Immunocytochemistryof oculomotor afferents in the squirrel monkey (Saimiri sciureus). J.Hirnforsch. 33:151–167.
Celio, M.R. (1990) Calbindin and parvalbumin in the rat nervous system.Neuroscience 35:375–475.
Christoff, N. (1974) A clinicopathological study of vertical eye movements.Arch. Neurol. 31:1–8.
Cogan, D.C. (1974) Paralysis of down-gaze. Arch. Ophthalmol. 91:192–200.Crawford, J.D., W. Cadera, and T. Vilis (1991) Generation of torsional and
vertical eye position signals by the interstitial nucleus of Cajal. Science252:1551–1553.
Crawford, J.D. and T. Vilis (1992) Symmetry of oculomotor burst neuroncoordinates about Listing’s plane. J. Neurophysiol. 68:432–448.
Crossland, W.J., X.J. Hu, and J.A. Rafols (1994) Morphological study of therostral interstitial nucleus of the medial longitudinal fasciculus in themonkey, Macaca mulatta, by Nissl, Golgi, and computer reconstructionand rotation methods. J. Comp. Neurol. 347:47–63.
Evinger, C. and J.T. Erichsen (1986) Transsynaptic retrograde transport offragment C of tetanus toxin demonstrated by immunohistochemicallocalization. Brain Res. 380:383–388.
Fukushima, K. (1987) The interstitial nucleus of Cajal and its role in thecontrol of movements of head and eyes. Prog. Neurobiol. 29:107–192.
Fukushima, K., C.R.S. Kaneko, and A.F. Fuchs (1992) The neuronalsubstrate of integration in the oculomotor system. Prog. Neurobiol.39:609–639.
Halmagyi, G.M., I. Dehaene, I.S. Curthoys, and M.J. Todd (1994) Jerk-waveform see-saw nystagmus due to a unilateral mesodiencephaliclesion. Brain 117:789–803.
Helmchen, C., H. Rambold, and U. Buttner (1996a) Saccade-related burstneurons with torsional and vertical on-directions in the interstitialnucleus of cajal of the alert monkey. Exp. Brain Res. 112:63–78.
Helmchen, C., S. Glasauer, K. Bartl, and U. Buttner (1996b) Contralesion-ally beating torsional nystagmus in a unilateral rostral midbrain lesion.Neurology 47:482–486.
Hepp, K., V. Henn, T. Vilis, and B. Cohen (1989) Brainstem regions relatedto saccade generation. Rev. Oculomot. Res. 3:105–212.
Holstege, G. and R.J. Cowie (1989) Projections from the rostral mesence-phalic reticular formation to the spinal cord. Exp. Brain Res. 75:265–279.
Horn, A.K.E. and J.A. Buttner-Ennever (1990) The time course of retro-grade transsynaptic transport of tetanus toxin fragment C in theoculomotor system of the rabbit after injection into extraocular eyemuscles. Exp. Brain Res. 81:353–362.
Horn,A.K.E. and K.P. Hoffmann (1987) Combined GABA-immunocytochem-istry and TMB-HRP histochemistry of pretectal nuclei projecting to theinferior olive in rats, cats and monkeys. Brain Res. 409:133–138.
Horn, A.K.E., J.A. Buttner-Ennever, Y. Suzuki, and V. Henn (1995) Histo-logical identification of premotor neurons for horizontal saccades inmonkey and man by parvalbumin immunostaining. J. Comp. Neurol.359:350–363.
Horn, A.K.E., J.A. Buttner-Ennever, P. Wahle, and I. Reichenberger (1994)Neurotransmitter profile of saccadic omnipause neurons in nucleusraphe interpositus. J. Neurosci. 14:2032–2046.
King, W.M. and A.F. Fuchs (1979) Reticular control of vertical saccadic eyemovements by mesencephalic burst neurons. J. Neurophysiol. 42:861–876.
Kompf, D., T. Pasik, P. Pasik, and M.B. Bender (1979) Downward gaze inmonkeys: Stimulation and lesion studies. Brain 102:527–558.
Kokkoroyannis, T., C.A. Scudder, C.D. Balaban, S.M. Highstein, and A.K.Moschovakis (1996) Anatomy and physiology of the primate interstitialnucleus of Cajal: I. Efferent projections. J. Neurophysiol. 75:725–739.
Li, W.B., Y. Shiraishi, and S. Nakao (1993) Topographical organization ofcat mesodiencephalic areas for monosynaptic activation of verticaloculomotoneurons. Exp. Brain Res. 94:43–52.
Moschovakis, A.K., C.A. Scudder, and S.M. Highstein (1991a) Structure ofthe primate oculomotor burst generator: I. Medium-lead burst neuronswith upward on-directions. J. Neurophysiol. 65:203–217.
Moschovakis, A.K., C.A. Scudder, S.M. Highstein, and J.D. Warren (1991b)Structure of the primate oculomotor burst generator: II. Medium-leadburst neurons with downward on-directions. J. Neurophysiol. 65:218–229.
Moschovakis, A.K., C.A. Scudder, and S.M. Highstein (1996) The micro-scopic anatomy and physiology of the mammalian saccadic system.Prog. Neurobiol. 50:133–254.
Nakao, S. and Y. Shiraishi (1983) Excitatory and inhibitory synaptic inputsfrom the medial mesodiencephalic junction to vertical eye movement-related motoneurons in the cat oculomotor nucleus. Neurosci. Lett.42:125–130.
Nakao, S. and Y. Shiraishi (1985) Direct excitatory and inhibitory synapticinputs from the mesodiencephalic junction to motoneurons innervatingextraocular oblique muscles in the cat. Exp. Brain Res. 61:62–72.
Onodera, S. and T.P. Hicks (1996) A projection linking motor cortex with theLM-suprageniculate nuclear complex through the periaqueductal grayarea which surrounds the nucleus of Darkschewitsch in the cat. Prog.Brain Res. 112:85–98.
Partsalis, A.M., S.M. Highstein, and A.K. Moschovakis (1994) Lesions of theposterior commissure disable the vertical neural integrator of theprimate oculomotor system. J. Neurophysiol. 71:2582–2585.
Pierrot-Deseilligny, C.H., F. Chain, F. Gray, M. Serdaru, R. Escourolle, andF. Lhermitte (1982) Parinaud’s syndrome: Electro-oculographic andanatomical analyses of six vascular cases with deductions aboutvertical gaze organization in the premotor structures. Brain 105:667–696.
Ranalli, P.J., J.A. Sharpe, and W.A. Fletcher (1988) Palsy of upward anddownward saccadic, pursuit and vestibular movements with a unilat-eral midbrain lesion. Neurology 38:114–122.
Riordan-Eva, P., M. Faldon, J.A. Buttner-Ennever, A. Gass, A.M. Bronstein,and M.A. Gresty (1996) Abnormalities of torsional fast phase eyemovements in unilateral rostral midbrain disease. Neurology 47:201–207.
Robinson, F.R., J.O. Phillips, and A.F. Fuchs (1994) Coordination of gazeshifts in primates: Brainstem inputs to neck and extraocular motoneu-ron pools. J. Comp. Neurol. 346:43–62.
Schwindt, P.C., W. Precht, and A. Richter (1974) Monosynaptic excitatoryand inhibitory pathways from medial midbrain nuclei to trochlearmotoneurons. Exp. Brain Res. 20:223–238.
Scudder, C.A., A.K. Moschovakis, A.B. Karabelas, and S.M. Highstein(1996) Anatomy and physiology of saccadic long-lead burst neuronsrecorded in the alert squirrel monkey: II. Pontine neurons. J. Neuro-physiol. 76:353–370.
Spencer, R.F. and S.F. Wang (1996) Immunohistochemical localization ofneurotransmitters utilized by neurons in the rostral interstitial nucleusof the medial longitudinal fasciculus (riMLF) that project to theoculomotor and trochlear nuclei in the cat. J. Comp. Neurol. 366:134–148.
Steiger, H.J. and J.A. Buttner-Ennever (1979) Oculomotor nucleus affer-ents in the monkey demonstrated with horseradish peroxidase. BrainRes. 160:1–15.
Suzuki, Y., J.A. Buttner-Ennever, D. Straumann, K. Hepp, B.J.M. Hess,and V. Henn (1995) Deficits in torsional and vertical rapid eye move-ments and shift of Listing’s plane after uni- and bilateral lesions of therostral interstitial nucleus of the medial longitudinal fasciculus. Exp.Brain Res. 106:215–232.
Trojanowski, J.Q. and M.H. Lafontaine (1981) Neuroanatomical correlatesof selective downgaze paralysis. J. Neurol. Sci. 52:91–101.
Vilis, T., K. Hepp, U. Schwarz, and V. Henn (1989) On the generation ofvertical and torsional rapid eye movements in the monkey. Exp. BrainRes. 77:1–11.
Wang, S.F. and R.F. Spencer (1996a) Morphology and soma-dendriticdistribution of synaptic endings from the rostral interstitial nucleus ofthe medial longitudinal fasciculus (riMLF) on motoneurons in theoculomotor and trochlear nuclei in the cat. J. Comp. Neurol. 366:149–162.
Wang, S.F. and R.F. Spencer (1996b) Spatial organization of premotorneurons related to vertical upward and downward saccadic eye move-ments in the rostral interstitial nucleus of the medial longitudinalfasciculus (riMLF) in the cat. J. Comp. Neurol. 366:163–180.
Wellhorner, H.H., U.C. Seib, and B. Hensel (1973) Local tetanus in cats:The influence of neuromuscular activity on spinal distribution of125I-labelled tetanus toxin. Naunyn-Schmiedebergs Arch. Pharmacol.276:387–394.
Zuk, A., D.G. Gwyn, and J.G. Rutherford (1982) Cytoarchitecture, neuronalmorphology, and some efferent connections of the interstitial nucleus ofCajal (INC) in the cat. J. Comp. Neurol. 212:278–292.
PREMOTOR NEURONS FOR VERTICAL SACCADES IN MONKEY AND MAN 427