presenter; dr. desderius b.m (md) resident 2012/13 mmed. internal medicine muhas facilitator; prof....
TRANSCRIPT

Presenter; Dr. Desderius B.M (MD)Resident 2012/13
Mmed. Internal MedicineMUHAS
Facilitator; Prof. Maselle
Bacterial & Fungal Pathogenicity and virulence factors.

A pathogen is a microorganism that is able to cause disease in a plant, animal or insect
Microbes express their pathogenicity by means of their virulence, a term which refers to the degree of pathogenicity of the microbe. Hence the determinants of virulence of a pathogen are any of its genetic, biochemical or structural features that enable it to produce disease in a host.

a) Pathogenicity – ability of an infectious agent to cause disease.b)Invasion – Process whereby organisms enter host cell or tissue and spread in the body.c)Infectivity – ability to breach host defensesd)Toxigenicity – Ability to produce a toxin that contribute to the development of diseasee)Virulence – Extent to which an organism can inflict disease. N.B; Virulence involves invasiveness and toxigenicity.f) Lethal dose 50 (LD 50) Number of microorganism which kill 50% of host tissue cells and cause ill effect. It is a measure of virulence. The higher the lethal dose the lower the virulence

Adhesin -A surface structure or macromolecule that binds a bacterium to a specific surface
Receptor-A complementary macromolecular binding site on a (eukaryotic) surface that binds specific adhesins.
Common pili -Same as fimbriae Lipopolysaccharide (LPS) -A distinct cell wall
component of the outer membrane of Gram-negative bacteria with the potential structural diversity to mediate specific adherence. Probably functions as an adhesin
Teichoic acids and lipoteichoic acids (LTA)-Cell wall components of Gram-positive bacteria that may be involved in nonspecific or specific adherence

Fimbriae -Filamentous proteins on the surface of bacterial cells that may behave as adhesins for specific adherence
Type 1 fimbriae-Fimbriae in Enterobacteriaceae which bind specifically to mannose terminated glycoproteins on eukaryotic cell surfaces
Glycocalyx -A layer of exopolysaccharide fibers on the surface of bacterial cells which may be involved in adherence to a surface
Capsule-A detectable layer of polysaccharide (rarely polypeptide) on the surface of a bacterial cell which may mediate specific or nonspecific attachment

EXPOSURE TO PATHOGENS
ADHERENCE (ADHESION), COLONIZATION, GROWTH
INVASION
TISSUE DAMAGES, DISEASE
ESCAPE HOST DEFENSES
TOXICITY

Pathogenesis : is a multistep process involving
1) Transmission;- Pathogen is transmitted to host-
either through:-- Food- Air- Water- Sexual intercourse

2) AttachmentPathogens must be able to bind to some
receptor molecule on cell surface.Most diseases are tissue specific because
only certain tissue have receptor molecule needed eg. HIV need only certain lymphocyte ie. CD4 cells
Fimbriae or pilli are used by some bacteria to attach selectively to certain tissues.
Eg. N. gonnorrhoea binds to genital epithelia by fimbriae.

In mutant cells without fimbriae, infectivity and pathogenicity are lost
3) Penetration & Spread ( Invasiveness)- There are bacteria that cause disease without penetration Eg. Vibrio cholerae- After penetration organisms should be able to spread through. There are some that don’t require to spread.
Many bacteria have specific enzymes that allow them to penetrate tissues

STEP REQUIREMENT PHENOMENON
Attachment +/- entry into body
Evade natural protective and cleansing mechanisms
Entry (infection)
Local or general spread in the body
Evade immediate local defenses
spread
Multiplication Increase numbers (many will die in the host, or en route to new hosts)
Multiplication
Evasion of host defenses Evade immune and other defenses long enough for the full cycle in the host to be completed
Microbial answer to host defenses
Shedding from body (exit) Leave body at a site and on a scale that ensures spread to fresh hosts
transmission
Cause damage in host Not strictly necessary but often occurs
Pathology, disease

Adherence factors Invasins - invasion of host cells and tissues Toxins Enzymes Anti-phagocytic factors Intracellular pathogenicity Antigenic heterogeneity Bacterial biofilms Requirement for iron

Two broad qualities of pathogenic bacteria underlie the means by which they cause disease:
1. The ability to invade tissues: Invasiveness, which encompasses mechanisms for colonization (adherence and initial multiplication), ability to bypass or overcome host defense mechanisms, and the production of extracellular substances which facilitate invasion.
2. The ability to produce toxins is termed as Toxigenesis - Bacteria produce two types of toxins called exotoxins and endotoxins.
Exotoxins are released from bacterial cells and may act at tissue sites removed from the site of bacterial growth. Endotoxins are cell-associated

The first stage of microbial infection is colonization: the establishment of the pathogen at the appropriate portal of entry. Pathogens usually colonize host tissues that are in contact with the external environment. -Adherence and colonization: Bacteria need to avoid being washed away by non-specific host defense mechanisms (e.g., saliva).
Most bacteria possess specific adherence factors which bind receptors on the surface of the host cells or tissue.

Bacterial Adherence to Mucosal Surfaces. In its simplest form, bacterial adherence or attachment to a eukaryotic cell or tissue surface requires the participation of two factors: a receptor and an adhesin. The receptors so far defined are usually specific carbohydrate or peptide residues on the eukaryotic cell surface. The bacterial adhesin is typically a macromolecular component of the bacterial cell surface which interacts with the host cell receptor. Adhesins and receptors usually interact in a complementary and specific fashion

The specific interaction between bacterial adhesins and host receptors explains the observed tissue specificity of bacterial infections. For example, N. gonorrhoeae adheres better to human urogenital epithelia than to other tissue.
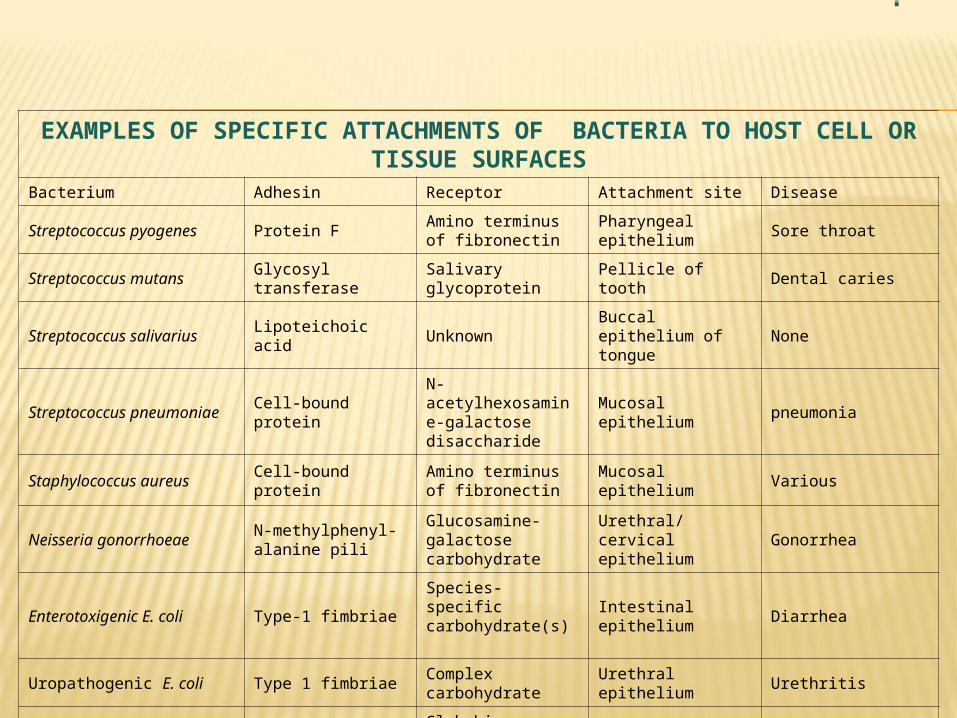
EXAMPLES OF SPECIFIC ATTACHMENTS OF BACTERIA TO HOST CELL OR TISSUE SURFACES
Bacterium Adhesin Receptor Attachment site Disease
Streptococcus pyogenes Protein FAmino terminus of fibronectin
Pharyngeal epithelium Sore throat
Streptococcus mutans Glycosyl transferase Salivary glycoprotein Pellicle of tooth Dental caries
Streptococcus salivarius Lipoteichoic acid Unknown Buccal epithelium of tongue
None
Streptococcus pneumoniae Cell-bound proteinN-acetylhexosamine-galactose disaccharide
Mucosal epithelium pneumonia
Staphylococcus aureus Cell-bound proteinAmino terminus of fibronectin
Mucosal epithelium Various
Neisseria gonorrhoeaeN-methylphenyl- alanine pili
Glucosamine-galactose carbohydrate
Urethral/cervical epithelium
Gonorrhea
Enterotoxigenic E. coli Type-1 fimbriaeSpecies-specific carbohydrate(s)
Intestinal epithelium Diarrhea
Uropathogenic E. coli Type 1 fimbriae Complex carbohydrate Urethral epithelium Urethritis
Uropathogenic E. coli P-pili (pap)Globobiose linked to ceramide lipid
Upper urinary tract Pyelonephritis
Bordetella pertussisFimbriae ("filamentous hemagglutinin")
Galactose on sulfated glycolipids
Respiratory epithelium Whooping cough
Vibrio choleraeN-methylphenylalanine pili
Fucose and mannose carbohydrate
Intestinal epithelium Cholera
Treponema pallidumPeptide in outer membrane
Surface protein(fibronectin)
Mucosal epithelium Syphilis
Mycoplasma Membrane protein Sialic acid Respiratory epithelium Pneumonia
Chlamydia Unknown Sialic acidConjunctival or urethral epithelium
Conjunctivitis or urethritis
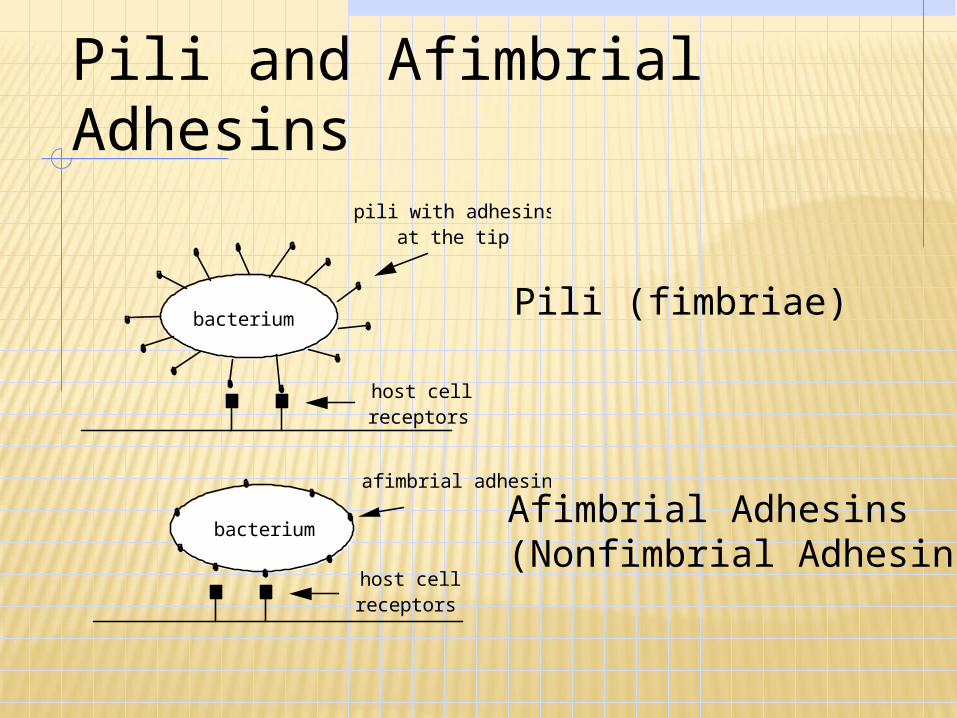
Pili and Afimbrial Adhesins
bacterium
host cell receptors
pili with adhesins at the tip
afimbrial adhesins
host cell receptors
bacterium
Pili (fimbriae)
Afimbrial Adhesins (Nonfimbrial Adhesins)

S. aureus: Lipoteichoid acid Staphylococcus: Slime Group A streptococci: F protein (non-
fibrillar, mediates attachment to fibronectin).
Streptococcus pneumonia: Capsule

The invasion of a host by a pathogen may be aided by the production of bacterial extracellular substances which act against the host by breaking down primary or secondary defenses of the body. These are referred to as Invasins.
Invasins are proteins (enzymes) that act locally to damage host cells and/or have the immediate effect of facilitating the growth and spread of the pathogen. The damage to the host as a result of this invasive activity may become part of the pathology of an infectious disease.
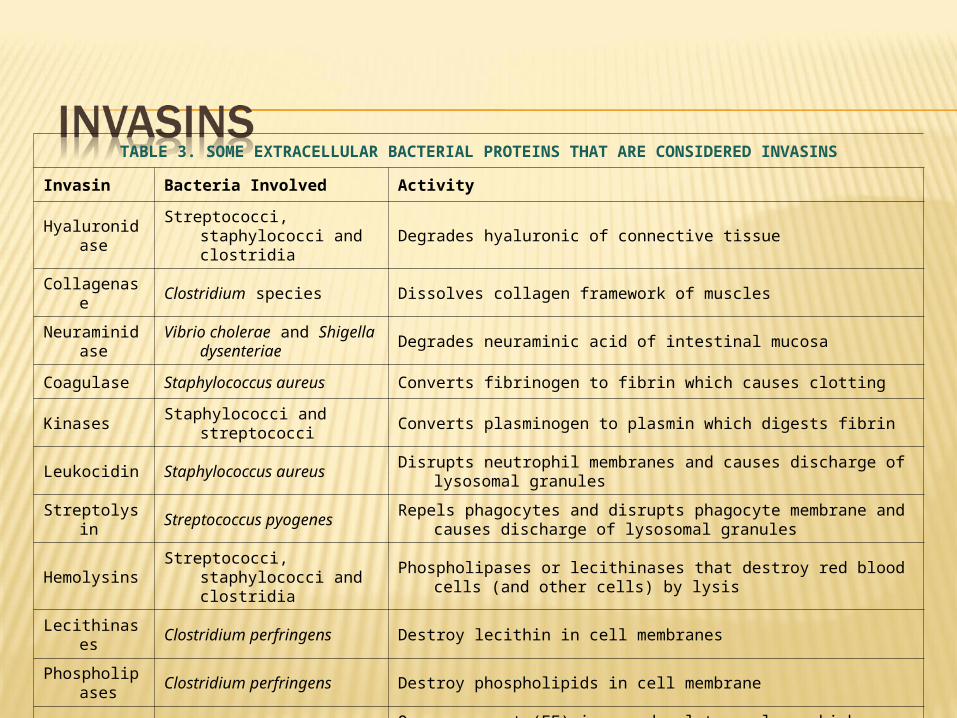
TABLE 3. SOME EXTRACELLULAR BACTERIAL PROTEINS THAT ARE CONSIDERED INVASINS
Invasin Bacteria Involved Activity
HyaluronidaseStreptococci, staphylococci and
clostridiaDegrades hyaluronic of connective tissue
Collagenase Clostridium species Dissolves collagen framework of muscles
NeuraminidaseVibrio cholerae and Shigella
dysenteriaeDegrades neuraminic acid of intestinal mucosa
Coagulase Staphylococcus aureus Converts fibrinogen to fibrin which causes clotting
Kinases Staphylococci and streptococci Converts plasminogen to plasmin which digests fibrin
Leukocidin Staphylococcus aureus Disrupts neutrophil membranes and causes discharge of lysosomal granules
Streptolysin Streptococcus pyogenesRepels phagocytes and disrupts phagocyte membrane and causes discharge of
lysosomal granules
HemolysinsStreptococci, staphylococci and
clostridia Phospholipases or lecithinases that destroy red blood cells (and other cells) by
lysis
Lecithinases Clostridium perfringens Destroy lecithin in cell membranes
Phospholipases
Clostridium perfringens Destroy phospholipids in cell membrane
Anthrax EF Bacillus anthracisOne component (EF) is an adenylate cyclase which causes increased levels of
intracellular cyclic AMP
Pertussis AC Bordetella pertussisOne toxin component is an adenylate cyclase that acts locally producing an
increase in intracellular cyclic AMP

Some pathogenic bacteria are inherently able to resist the bactericidal components of host tissues. For example, the poly-D-glutamate capsule of Bacillus anthracis protects the organisms against cell lysis by cationic proteins in sera or in phagocytes.
The outer membrane of Gram-negative bacteria is a formidable permeability barrier that is not easily penetrated by hydrophobic compounds such as bile salts which are harmful to the bacteria.
Pathogenic mycobacteria have a waxy cell wall that resists attack or digestion by most tissue bactericides. And intact lipopolysaccharides (LPS) of Gram-negative pathogens may protect the cells from complement-mediated lysis or the action of lysozyme.

Microbial strategies to avoid phagocytic killing are numerous and diverse, but are usually aimed at blocking one or of more steps in the phagocytic process. Recall the steps in phagocytosis:
1. Contact between phagocyte and microbial cell
2. Engulfment 3. Phagosome formation 4. Phagosome- lysosome fusion 5. Killing and digestion

Invade or remain confined in regions inaccessible to phagocytes
Avoid provoking an overwhelming inflammatory response
Inhibit phagocyte chemotaxis Hide the antigenic surface of bacterial
cell.

Capsules
Bacterium
Encapsulated organisms: Few C3b formed on the surface; the few C3b formed under capsule are not accessible to phagocytes. Also fewer MAC formed; reduces MAC killing in G- organisms
C3b Capsule

Many important bacteria bear on their surfaces substances that inhibit phagocytic adsorption and engulfment. Examples are polysaccharide capsule of S. pneumoniae, H. influenza.
Fimbriae of group A streptococci O antigen associated with
lipopolysaccharide of E. coli.

Inhibition of Phagosome-lysosome fusion: Salmonella, M. TB, Legionella and Chlamydiae
survival inside the phagolysosome: resistance to inhibition and killing by the lysosomal constituents- B. anthracis, M.TB,S. aureus.
Early escape from the Phagosome vacuole e.g. rickettsias
Killing phagocytes before ingestion e.g. P. aureginosa
Killing phagocytes after ingestion e.g. Mycobacteria, Brucella, Listeria.

Two types of bacterial toxins At a chemical level there are two types of
bacterial toxins: lipopolysaccharides, which are associated
with the cell walls of Gram-negative bacteria. proteins, which may be released into the
extracellular environment of pathogenic bacteria.
The lipopolysaccharide (LPS) component of the Gram-negative bacterial outer membrane bears the name endotoxin because of its association with the cell wall of bacteria.
Most of the protein toxins are thought of as exotoxins, since they are "released" from the bacteria and act on host cells at a distance.

PROPERTY ENDOTOXIN EXOTOXIN
CHEMICAL NATURE Lipopolysaccharide(mw = 10kDa) Protein (mw = 50-1000kDa)
RELATIONSHIP TO CELL Part of outer membrane Extracellular, diffusible
DENATURED BY BOILING No Usually
ANTIGENIC Yes Yes
FORM TOXOID No Yes
POTENCY Relatively low (>100ug) Relatively high (1 ug)
SPECIFICITY Low degree High degree
ENZYMATIC ACTIVITY No Usually
PYROGENICITY YesOccasionally

NAME OF TOXINBACTERIUM
INVOLVED
ACTIVITY
Anthrax toxin (EF)Bacillus
anthracis
Edema Factor (EF) is an adenylate cyclase that causes increased levels in intracellular cyclic AMP in phagocytes and formation of ion-permeable pores in membranes (hemolysis)
Cholera enterotoxin Vibrio choleraeADP ribosylation of G proteins stimulates adenlyate cyclase and increases cAMP in
cells of the GI tract, causing secretion of water and electrolytes
E. coli LT toxin Escherichia coli Similar to cholera toxin
Shiga toxinShigella
dysenteriae
Enzymatically cleaves rRNA resulting in inhibition of protein synthesis in susceptible cells
Botulinum toxinClostridium
botulinumZn++ dependent protease that inhibits neurotransmission at neuromuscular synapses
resulting in flaccid paralysis
Tetanus toxinClostridium
tetaniZn++ dependent protease that inhibits neurotransmission at inhibitory synapses
resulting in spastic paralysis
Staphylococcus enterotoxins*
Staphylococcus aureus
Massive activation of the immune system, including lymphocytes and macrophages, leads to emesis (vomiting)
Toxic shock syndrome toxin (TSST-1)*
Staphylococcus aureus
Acts on the vascular system causing inflammation, fever and shock

The surface structure of many bacteria have considerable antigenic heterogeneity. The antigenic type of the bacteria may be a marker for virulence. Some bacteria have the ability to make frequent shifts in the antigenic form of their surface structures, leading to the observed resistance to drugs.

Biofilms are bacterial communities which are enclosed within a matrix of polysaccharides produced by the bacteria and adhere to a living or inert surface. In nature, biofilms constitute a protected growth modality that allows the bacteria to survive in hostile environments.
The structures which make up these micro colonies contain canals through which nutrients circulate, and in different zones of the biofilm the cells express different genes, as if they were part of an organized structure.

The general idea is that the individualized bacteria are exposed to the agents found in the environment while inside the biofilm these cells are protected, hence are very resistant to normal host defense mechanisms and drugs.
Examples include Staphylococcus epidermidis and Staphylococcus aureus infections of central venous catheters, eye infections such as with contact lenses and intraocular lenses.

The speed with which host adaptive responses can be mobilized is crucial.
Every infection is a race between the capacity of the microorganism to multiply, spread and cause disease; and the ability of the host to control and finally terminate the infection.

4 types1. Microorganisms with specific mechanisms
for attaching to, or penetrating, the body surfaces of normal healthy hosts (most viruses and certain bacteria)
2. Those introduced into normal healthy hosts by biting arthropods e.g. malaria
3. Those introduced into otherwise normal healthy hosts via skin wounds or animal bites e.g. rabies
4. Microorganisms able to infect a normal healthy host only when the surface or systemic defenses are impaired as occurs in burns, catheters

The microbe must be present in every case of the disease
The microbe must be isolated from the diseased host and grown in pure culture
The disease must be reproduced when a pure culture is introduced into a non-diseased susceptible host
The microbe must be recoverable from an experimentally infected host

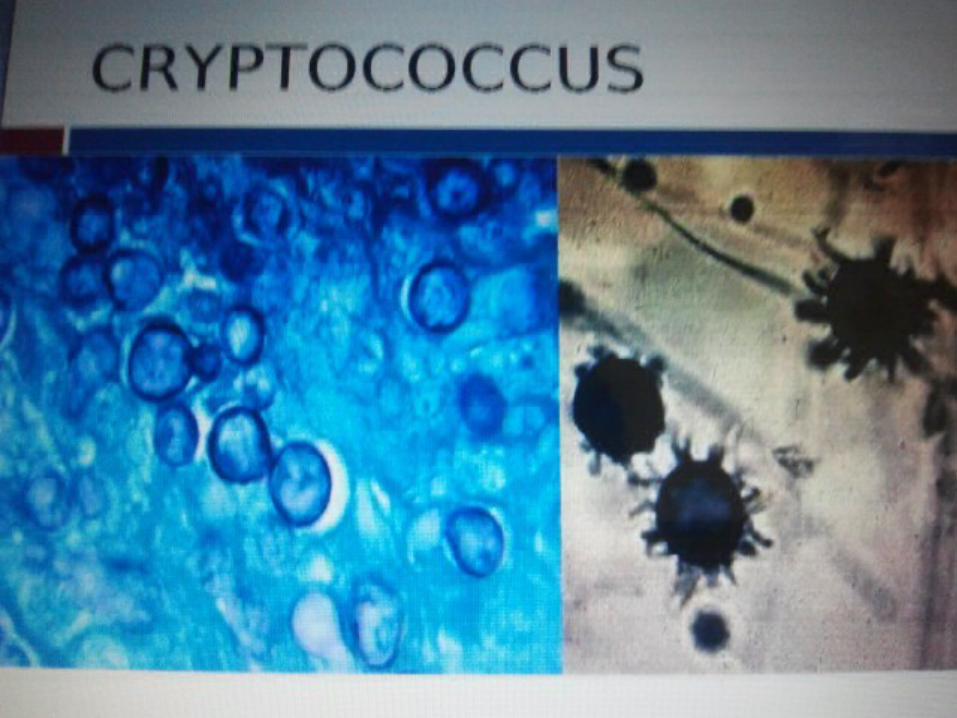
FUNGAL PATHOGENESIS AND VIRULENCE FACTORS

The complex interplay between fungal virulence factors and host defence factors will determine if a fungal infection will cause a disease
Infection depends on Inoculum size and the general immunity of the host.
Fungal infection is divided into; A) Primary Pathogens B) Opportunistic pathogens

Primary pathogens are those fungi that cause disease in non-compromised patients.
This distinction is necessarily a gray one, because C. neoformans, a model opportunist, sometimes causes disease in healthy individuals and the primary pathogens such as C. immitis are much more virulent in immunocompromised patients.

Furthermore, infection by primary pathogen often leads to subclinical disease. However, the distinction is worth making in the effort to understand general mechanisms of pathogenesis

Opportunistic pathogens incite disease in hosts whose local or systemic immune attributes have been impaired, damaged, or are innately dysfunctional
The pathogenesis of opportunistic infections involves production of virulence factors that allow individual organisms to be commensals during times when humans have normal immune systems.

Because of the complex nature of the host-fungus interaction, there are few factors that are absolutely required for fungal virulence.
Because pathogenesis is a complex, possession of singal putative virulence factor is not likely to render a fungus pathogenic; a complex mix of properties is usually required.

Ability to adhere to host cells by way of cell wall glycoproteins.
Production of capsules allow them to resist phagocytosis.
Production of a Cytokine called GM-CSR by Candida albicans that suppress the production of complement.
Growth at Elevated Temperature.

Ability to acquire Iron from red blood cells as in Candida albicans.
Ability to damage host by secreting enzymes such as keratinase, elastase, collagenase
Ability to resist killing by phagocytes as in dimorphic fungi.
Ability to secrete mycotoxins.

Having a unique enzymatic capacity. Exhibiting thermal dimorphism. Ability to block the cell-mediated
immune defences of the host. Surface hydrophobicity.

The ability to grow at body temperature, 37oC, and within the fever range, 38-42oC of the human host is clearly an important requirement for systemic infection.
This property is not a simple genetic one, experiments with Saccharomyces cerevisiae variants able to grow at physiological temperatures suggest that is a multigenic trait.

S. cerevisiae is normally not pathogenic, but McCusker and co-workers have found that, strains which are able to grow at 42oC and form pseudohyphae, are able to infect and persist in mice. This property is highly correlated with the high-temperature growth phenotype.

For most fungal infections, the ability of the host to resist the physical clearing of the infectious agent is important. E.g the lungs have effective means of clearing foreign particles, but Aspegillus species, H.capsulatum, and C.neoformans all infect via the bronchial route and must avoid clearance.
C. albicans also must adhere to various host surfaces both as a commensal to avoid being washed out of its various niches and as a pathogen during the onset of haematogenous infections

Adhesion of C. albicans has been the focus of much investigation and several gene products have been implicated.
Three of the most intensely studied adhesion mechanisms involve the HW gene product, the ALS gene family, and the INT1 gene product.

In various pathogens, a 120-kDa cell wall adhesin, Wl-1, which contains 34 copies of a 25-amino acid tandem repeat, has been isolated from the surface of Blastomyces dermatitidis.
This adhesin mediates attachement to human monocyte-derived macrophages mainly through binding complement type 3 receptors.

This protein is both released into the growth medium and found on the cell wall; free Wl-1 seems to be recaptured in the fungus and binds with the cell wall via covalent and noncovalent interactions.
The disruption of the gene prevents binding to and infection of macrophage diminishes adhesion and attenuates the virulence of the fungus.

Hence, in blastomycosis a single molecule affects adhesion both at the site of entry and during later stages of infection.

The first step in fungal infection is introduction of the agent to the host.
Infection may be limited to portal of entry or they may become systemic, disseminating either via hematogenous or contiguous routes.
Movement from the infecting surface into the blood stream requires tissue damage.
This damage can be preexisting or can occur either by mechanical penetration or new tissue necrosis.

Therefore, the ability of fungi to penetrate host cells is crucial for progression of infection in the setting of intact skin or gut barriers.
For Candida, it is the ability of hyphae to grow through host cell walls that is proposed to account for the importance of polymorphism in virulence.
A.fumigatus and other true molds are able to penetrate blood vessels and grow along the vessel lumen as they invade tissue.

Hyphae respond thigmotropically (movement toward or away from a touch stimulus) and morphologically to cues such as the presence of a surface, pores, grooves, and ridges.
Hyphae follow grooves and ridges of inert substrates and penetrate pores of filtration membranes.
Thus, thigmotropism may enhance the ability of a hyphato invade epithelia of a host at sites of weakened integrity or to follow vasculature.

Fungi may also spread from the site of infection throughout the host by such mechanisms as host Phagocytosis.
C. albicans invades endothelial cells through being phagocytosed.
H. capsulatum is phagocytosed by macrophages but does not seem to be killed and multiplies within the phagosome.

In order to flourish in the host, fungi need to be able to carry out biosynthetic reactions while concentrating relatively scarce nutrients like Ca2+ and Fe2+.Experiments with auxotrophic mutants of C. albicans (auxotrophs require a nutrient that the parent organism, the prototroph, does not require) have shown that the inability to synthesize purines, pyrimidines, or heme de novo significantly diminishes virulence

Necrotic factors are vehicles of virulence because they allow the fungus to overcome structural barriers that the human host uses to prevent invasive infection.
Most necrotic factors are enzymes. Because the majority of fungal pathogens are opportunists, these enzymes may have evolved for saprophytic purposes and might be considered nutritional factors, but it seems more likely that their major role in infection is degradation of tissue.

Among the factors that are thought to contribute to this damage are extracellular degradative enzymes such as Proteinases, Phosphatases and DNAses.
The earliest identification of a potential necrotic factor was the extracellular proteinase of C. albicans, given several different names to the gene of this enzyme such as Opa and EPR but all correspond to member of the Secreted aspartyl proteinase(SAP) gene family.

A. fumigatus secretes at least two proteinases, both of which are active on elastin, a protein that constitutes about 30% of lung tissue.
C. albicans is known to secrete phospholipases and to possess a phospholipase gene family.
Extracellular phospholipases have a role in the pathogenicity of C. albicans, as blood isolates produce significantly more extracellular phospholipase activity than do commensal strains.

MORPHOLOGICAL VERSALITY Almost all pathogenic fungi can grow in
more than one form. Aspergillus species, which are classical
filamentous molds, a form that they are infectious.
The major exception is C.neoformans which apparently exists only in the yeast form in vivo. In vitro it also grows mostly as a yeast; however, it does form filaments during the mating process.

H. capsulatum, blastomyces dermatitidis and several (but not all) species of candida can grow both as yeasts and as hyphae.
In C.albicans, both the yeast and hyphal forms are found at the site of infection, in Histoplasma and Blastomyces, the yeast form seems to be the major, if not the exclusive parasitic form.

The transition to the parasitic form in H.capsulatum leads to a specific pattern of gene expression; this pattern facilitates many of the steps important in infection , including blocking acidification of the phagolysosome and synthesis of the calcium-binding protein, Cbp1p.
In contrast to C.albicans , the H.capsulatum hypha seems to play no role in vivo.

Dimorphic fungi regulate their cellular morphology in response to environmental conditions. E.g. ellipsoidal single cells of C.albicans (blastospores) predominate in rich media, where as filaments composed of elongated cells attached end to end form in response to starvation, serum and other conditions.
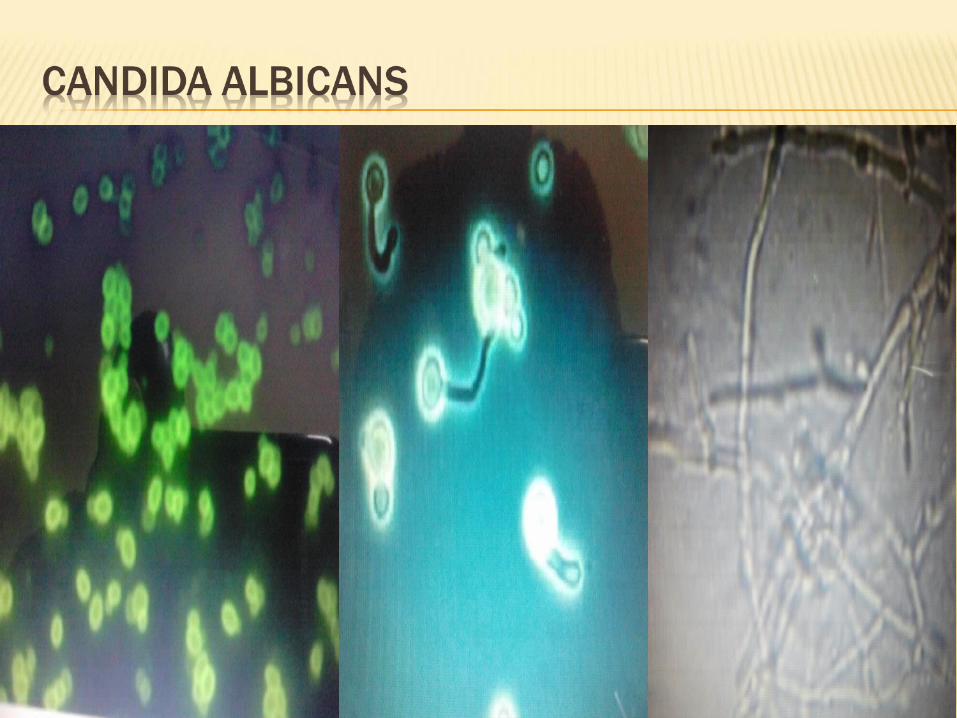

A shift of environmental changes, including a shift from an aerobic to a fermentative metabolism or growth on particular compounds such as N-acetyl glucosamine, cause C.albicans to switch from yeast to filamentous growth.
This change is accomplished by changes in carbohydrate metabolism and an interruption of electron transfer within the cell. Both temperature ( a shift to 37oC) and pH can regulate C.albicans dimorphism

Many molds form Conidia, or vegetative spores; scattered by wind or water, these small resistant cells serve as a mode of dissemination. In the case of aspergillosis, conidia serve as the propagule that infects debilitated patients.
Hydrophobicity is thought to contribute to the efficacy of Aspergillus conidia, already an ideal size for deposition into alveoli to disperse in air.
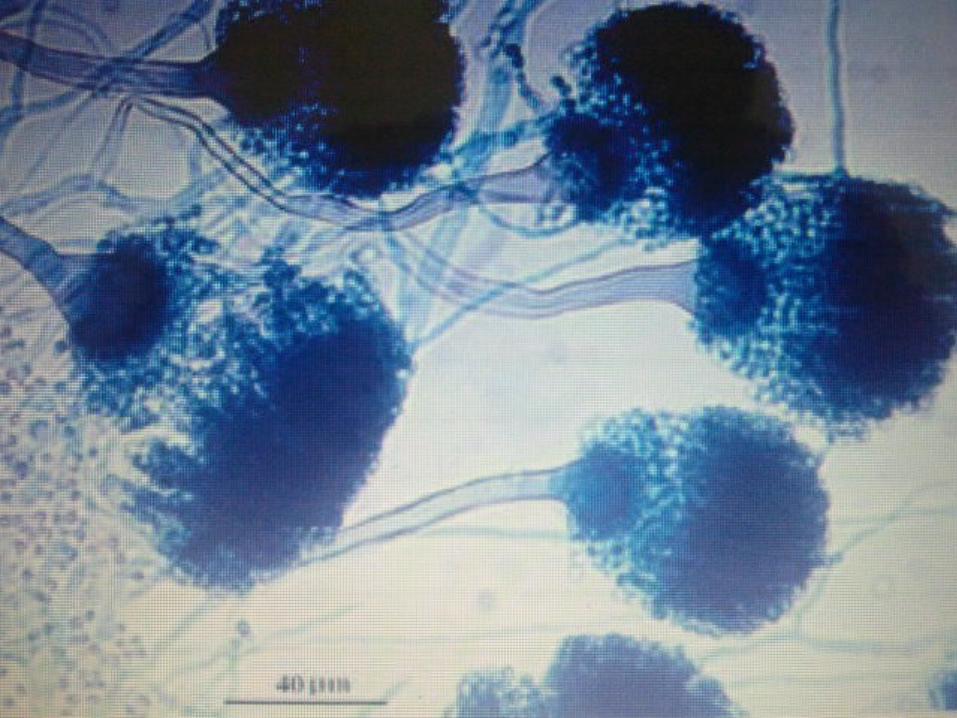

The capacity of fungi to undergo an epigenetic change (regulation of expression of gene activity without alteration of genetic structure) in colony morphology has come to be called phenotypic switching.
This phenomenon was often observed in C. albicans.
Phenotypic switching is characterised by reversible change, usually occurring between 10-3 and 10-5 per cell division.

A particular strain, WO-1 that has two major colony phenotypes, “white” and “Opaque”.
White colonies almost exclusively contain the classical yeast cells. Opaque colonies contain cells that are bean shaped and differ from white cells in a variety of ways including surface properties, gene expression, and temperature sensitivity.
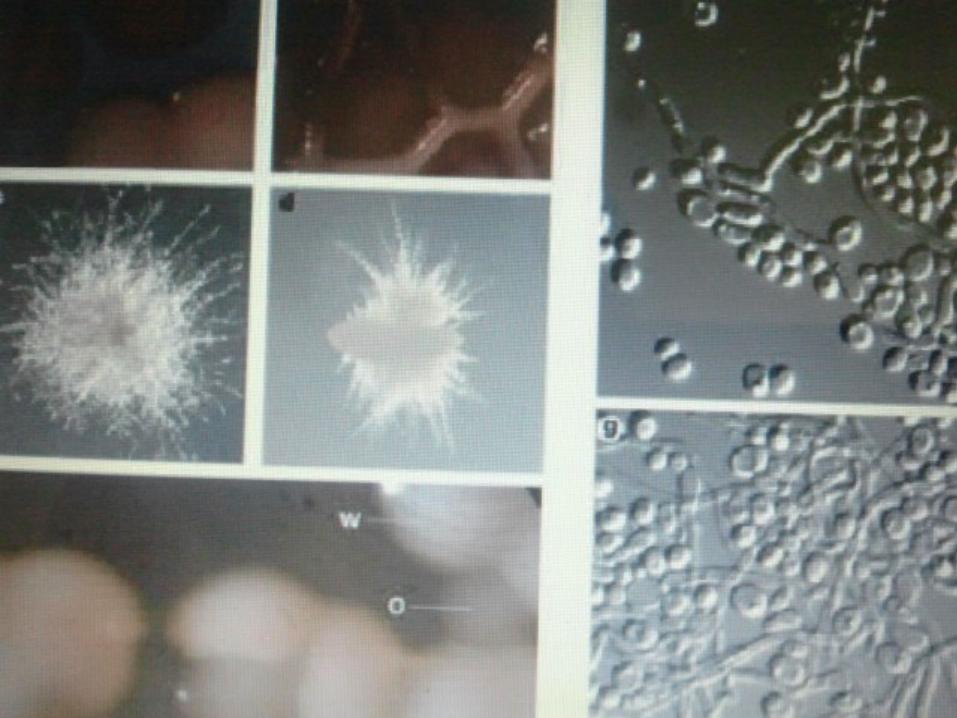

Opaque cells switch to White cells within one generation at 37oc.
The white phenotype is more virulent in a systemic mouse model of infection, where as opaque cells are more effective in colonization in a mouse cutaneous infection model.

GROWTH AT DIFFERENT pHsC.albicans can grow both at acid and at basic
pHs, a reflection of its ability to colonize several niches, ranging from the acid vagina to the neutral oropharyngeal tract.
MELANINMelanins are scavengers of reactive oxygen
intermediaries, making organisms relatively resistant to leukocyte attack.

IRON AND CALCIUM Iron is an essential element for the growth
and metabolism of fungi.C. Neoformans capsular polysaccharide
synthesis is increased by limitation of ferric iron.
Most pathogenic microbes elaborate siderophores (Molecules that can bind iron) to mobilize iron from ferric ligands.

Hereditary or acquired hemocromatosis, the human disease with progressive iron overload leading to fibrosis and organ failure, poses an increased risk for the development of fungal infection.

The best known of the fungal immuno-evasion system is the capsule of C.neoformans. A viscous polysaccharide capsule composed of glucuronoxyomannan and other components, the capsule is believed to present a surface not recognised by phagocytes, down regulate cytokine secretion, inhibit leukocyte accumulation, induce suppressive T-cells, inhibit antigen presentation, and inhibit lymphoproliferations.

Hence, it serves as a barrier to host defences in a variety of ways.

Many fungi produce biologically active compounds, several of which are toxic to animals or plants in and are therefore called mycotoxins.
Of particular relevance to humans are mycotoxins produced by molds causing food spoilage and poisonous mushrooms.
Particularly infamous are the lethal amatoxins in some Amanita mushrooms, and ergot alkaloids, which have a long history of causing serious epidemics of ergotism.


Other notable mycotoxins include aflatoxins, which are insidious liver toxins and highly carcinogenic metabolites produced by certain Aspergillus species often growing in or on grains and nuts consumed by humans.
Mycotoxins are secondary metabolites (or natural products).
Mycotoxins may provide fitness benefits in terms of physiological adaptation, competition with other microbes and fungi, and protection from consumption (Fungivory)

Physical barriers, such as skin and mucus membranes.
The fatty acid content of the skin. The pH of the, mucosal surfaces and
body fluids. Epithelial cell turnover Normal flora

Prolonged antibiotic therapy Underlying disease (HIV Infection, cancer, diabetes etc.) Age Surgical procedures. Immunosuppressive drugs Irradiation therapy Indwelling catheters Obesity Drug addiction Obesity Drug addiction Transplants Occupation

To establish infection in the host, microbes must attach to, or pass across body surfaces
Many microbes have developed chemical or mechanical mechanisms to attach themselves to different surfaces and to bypass host defense mechanisms; and in turn host defenses have had to be modified
Speed matters in causation of disease It is sometimes difficult to incriminate a
specific microbe as the cause of disease Microbes do not necessary cause the same
disease in all infected individuals. A biologic response gradient causes a spectrum that can range from an asymptomatic to a lethal infection.

With examples, Compare and contrast bacterial and fungal pathogenicity for human diseases.
Out line the virulence factors for bacterial and fungal infections.

Thank you