principles of biochemistry - unifr.ch ch12.pdf · turn over numbers d’enzymes catalase h 2o 2...
TRANSCRIPT

Principles of Biochemistry Third Edition
International Student Edition
Chapter 12 Enzyme Kinetics, Inhibition, and Control
Copyright © 2008 by John Wiley & Sons, Inc.
Donald Voet • Judith G. Voet • Charlotte W. Pratt
Chapter 12 Opener

Cinetique de Catalyse Rappel de notions de cinétique
Réaction simple, irréversible • 1/ la vitesse d’une réaction est proportionnelle à la concentration x constante de vitesse:
• Pour A -> P v= k(A) réaction de 1er ordre
• Pour A+B -> P v=k(A)(B) réaction de 2ème ordre (1er ordre pour A et 1er ordre pour B)
• Pour 2A -> P v=k(A)2 réaction de 2ème ordre
Réaction réversible:
• A+B <=> P v=k1(A)(B) - k-1(P)
Réaction catalysée:
• A+cat <=> A-cat <=> P-cat <=> P + cat
k1
k-1
k
k
k
k1
k-1

Figure 12-1
Diagramme de vitesse de 1er ordre dA/A = dlnA = -kdt A = A0e-kt

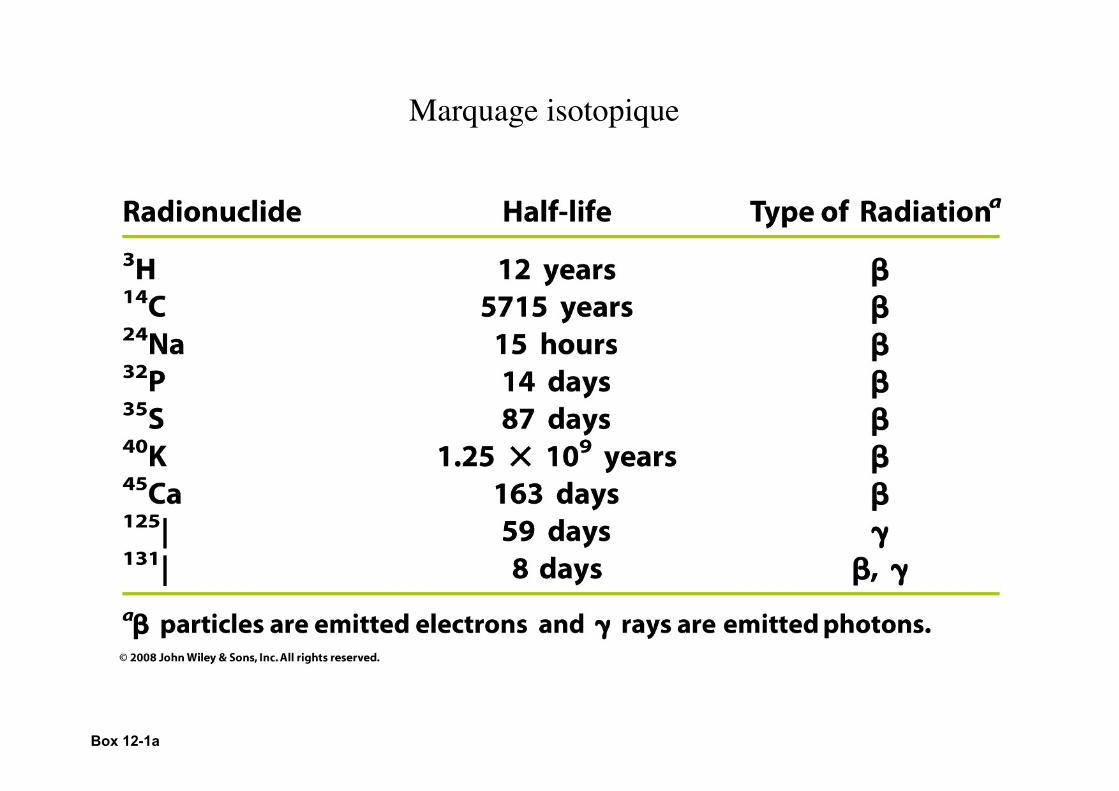
Box 12-1a
Marquage isotopique

Box 12-1b

Figure 12-2
Etat stationnaire

Figure 12-3
Cinétique de Michaelis
Vmax * S vo = _________________
KM + S
k2 + k3 KM= __________
k1
k1
k2
k3

Table 12-1

Figure 12-4
Diagramme de Lineweaver-Burk
k2 + k3 KM= __________
k1
k1
k2
k3

k1[E][S] = k-1[ES] + k2[ES]
KM = (k-1 + k2)/ k1 • KM mesure la tendance de ES à se dissocier dans chaque direction en fonction de la tendance à se former. • donc un KM élevé signifie - rapide formation du produit (grand k2)
OU - rapide dissociation du complexe (grand k-1).
E + S ES E + Pk1
k- 1
k2
Que signifie KM? Cinétique

si v = 1/2 Vmax = Vmax [S] KM + [S]
Vmax 2
=>: 2[S] = KM + [S] [S] = KM
Que signifie KM?
KM
Vmax
Initialrate, V
Substrate concentration, [S]
Vmax/2
Plot of rate (V) against substrate concentration ([S])
E + S ES E + Pk1
k- 1
k2
=>:
=>: v = Vmax [S] KM + [S]
Cinétique

• Pour toute réaction selon Michaelis-Menten:
KM = [S] => v=Vmax/2
• donc, KM est une mesure de [S] requise pour une catalyse effective
• Plus KM est grand, plus [S] est élevée pour réaliser une certaine vitesse de réaction.
NB: la valeur KM ne donne pas la force de liaison du substrat à l’enzyme (sauf cas limite où k2 << k-1) ni la specificité ou l’efficacité de l’enzyme.
Ceci est fourni par kcat/KM.
E + S ES E + Pk1
k- 1
k2
Que signifie KM? Cinétique

Turn Over Number, kcat
vo = Vmax [S] Km + [S]
(vo) E + S ES E + P
k2
k1 k3
Si [substrat] en excès , k3 = kcat, turn over number (t.o.n)
Si [substrat] faible
= k3 [E][S] Km + [S]
= Km
k3 [E][S]
[substrat] négligeable specificité de Substrat Equation de Michaelis-Menten
2ème ordre (IV)
Juang RH (2004) BCbasics

Turn Over Numbers d’Enzymes
Catalase H2O2
Carbonic anhydrase HCO3-
Acetylcholinesterase Acetylcholine
40,000,000
400,000
140,000
b-Lactamase Benzylpenicillin 2,000
Fumarase Fumarate 800
RecA protein (ATPase) ATP 0.4
Enzymes Substrat kcat (s-1)
Le nombre de molécules de produit transformées par mol. de substrate par une molecule d’enzyme en une seconde Adapted from Nelson & Cox (2000) Lehninger Principles of Biochemistry (3e) p.263

kcat
• kcat est une mesure directe de la production catalytique du produit sous conditions saturantes de substrat.
• L’unité de kcat est s-1. • kcat mesure le nombre molecules de substrat
transformées par molecule d’enzyme par seconde (turn-over).
E + S ES E + Pk1
k- 1
k2
Cinétique

kcat/KM • kcat/KM est une mesure directe de l’efficience et de la
specificité de l’ enzyme.
• indique ce que l’enzyme peut accomplir si un nombre abondant de sites sur l’enzyme sont accessibles.
• kcat/KM permet une comparison directe de la spécificité d’un enzyme pour un substrate.
• L’efficience maximum possible serait si toutes les collisions moleculaires resultent en une reaction. Selon la theorie de diffusion, cette valeur serait ca. 108 –109 (mol/L)-1 s-1.
E + S ES E + Pk1
k- 1
k2
Cinétique

Chymotrypsine a des kcat / Km distinct pour Different Substrats
O R O H3C–C–N–C–C–O–CH3
H H
= – = – –
–H Glycine
kcat / Km
1.3 ╳ 10-1
–CH2–CH2–CH3 Norvaline 3.6 ╳ 102
–CH2–CH2–CH2–CH3 Norleucine 3.0 ╳ 103
–CH2– Phenylalanine 1.0 ╳ 105
(M-1 s-1)
R =
Adapted from Mathews et al (2000) Biochemistry (3e) p.379

Determiner kcat/KM • Effectuer une série de mesures, avec même [E]t mais en
variant [S]0 et mesurer l’augmentation de [P] ou la diminution de [S] à divers moments.
• Vitesses initiales, V0 est linéaire = [S]/t
[S]
time 3 6 9 15 30
[S]01
[S]02
[S]03
Cinétique

kcat/KM On peut estimer KM et Vmax par le diagramme v vs. [S]. 1.Estimer Vmax (asymptote).
2.Calculer Vmax/2
3.lire KM du graphique.
Vmax difficile à determiner sur le diagramme v/S (hyperbole). Difficile d’extrapole [S] infini et Vmax.
E + S ES E + Pk1
k- 1
k2
Cinétique

Page 374
L’étude d’état stationnaire n’est pas univoque (notion de Black Box)

Cinétique à plusieurs substrats
États de l’enzyme
A, B, C, D…. = substrats P, Q, R, S… = produits
A B C P Q R
E EA EAB EABC<=>EPQR EQR ER E Mécanisme séquentiel ter, ter
Selon nombre de réactants: uni, bi, ter, quad, etc
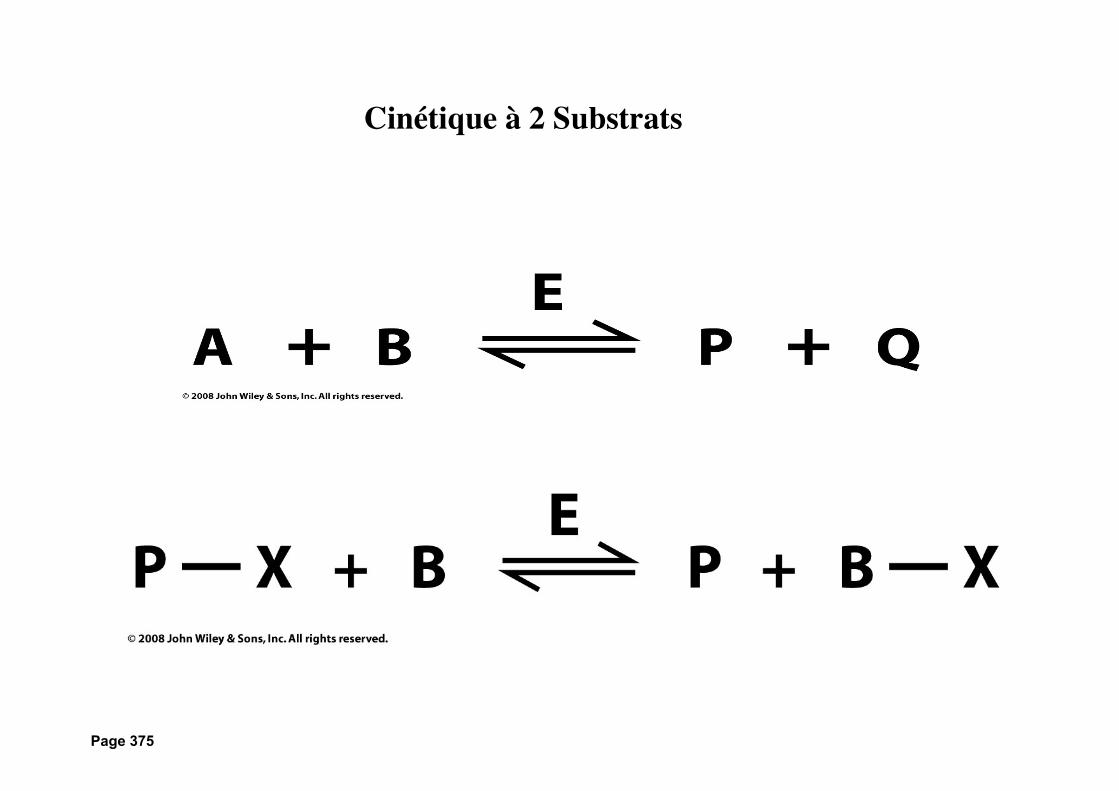
Page 375
Cinétique à 2 Substrats

Figure 12-5
Cinétique à 2 Substrats: exemples

Page 376
Cinétique à 2 Substrats: Mécanismes
1. Mécanisme ordonné bi-bi

Page 376
2. Mécanisme aléatoire bi-bi
Cinétique à 2 Substrats: Mécanismes

Page 376
3. Mécanisme séquentiel bi-bi
Cinétique à 2 Substrats: Mécanismes



Page 377
Inhibition Enzymatique
Exemple : inhibiteur compétitif

Page 378
Inhibition Enzymatique
Exemple : inhibiteur d’état de transistion

Page 378
Inhibition Enzymatique Compétitive
Le degré d’inhibition compétitive varie avec le taux d’enzyme qui lie l’inhibiteur
Vmax * S vo = ______________
αKM + S

Figure 12-6
Inhibition Enzymatique Compétitive
Le degré d’inhibition compétitive varie avec le taux d’enzyme qui lie l’inhibiteur

Page 379
Inhibition Enzymatique Compétitive

Figure 12-7
Inhibition Enzymatique Compétitive
Le degré d’inhibition compétitive varie avec le taux d’enzyme qui lie l’inhibiteur

Page 381
Inhibition Enzymatique Incompétitive
L’inhibiteur incompétitf se lie sur le complexe enzyme-substrat, mais pas sur l’enzyme libre

Figure 12-8
Inhibition Enzymatique Incompétitive
L’inhibiteur incompétitf se lie sur le complexe enzyme-substrat, mais pas sur l’enzyme libre

Page 382
Inhibition Enzymatique Mixte
L’inhibiteur mixte se lie aussi bien sur le complexe enzyme-substrat, que sur l’enzyme libre

Figure 12-9
Inhibition Enzymatique Mixte
L’inhibiteur mixte se lie aussi bien sur le complexe enzyme-substrat, que sur l’enzyme libre
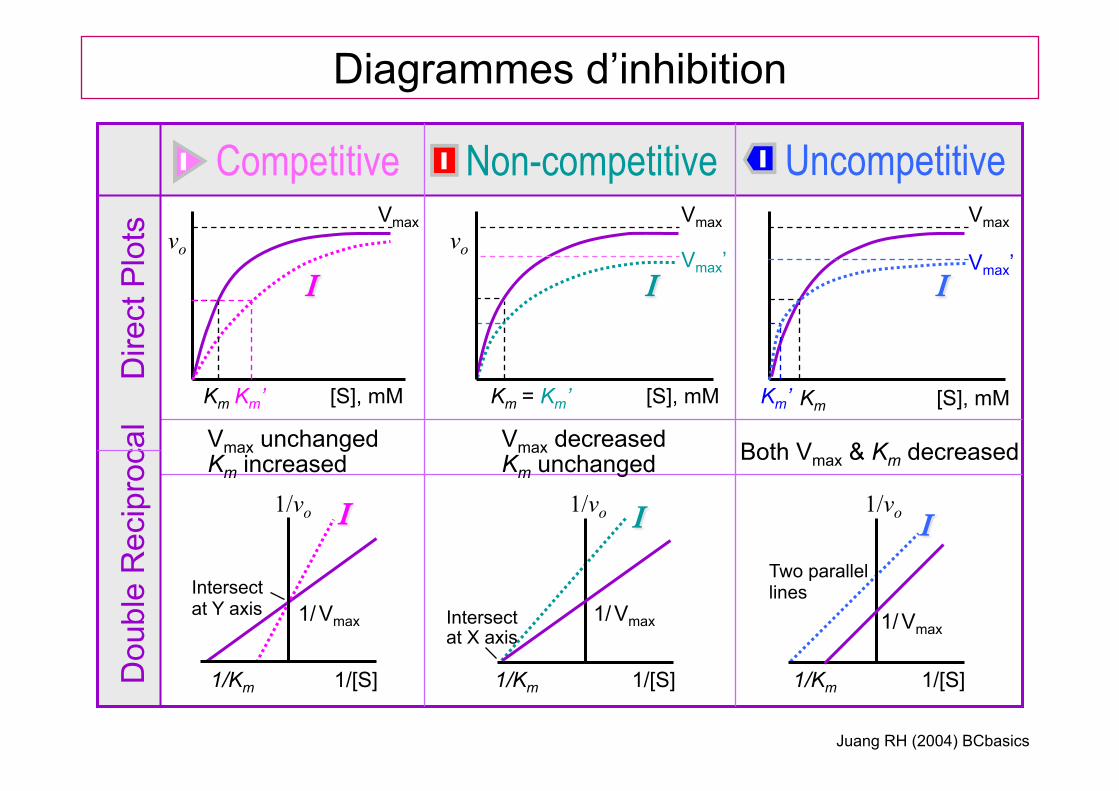
Km
Diagrammes d’inhibition
Competitive Non-competitive Uncompetitive D
irect
Plo
ts
Dou
ble
Rec
ipro
cal
Vmax Vmax
Km Km’ [S], mM
vo
[S], mM
vo
Km [S], mM
Vmax
Km’
Vmax’ Vmax’
Vmax unchanged Km increased
Vmax decreased Km unchanged Both Vmax & Km decreased
1/[S] 1/Km
1/vo
1/ Vmax
Two parallel lines
Intersect at X axis
1/vo
1/ Vmax
1/[S] 1/Km 1/[S] 1/Km
1/ Vmax
1/vo
Intersect at Y axis
= Km’
Juang RH (2004) BCbasics

Table 12-2
Effets des Inhibiteurs
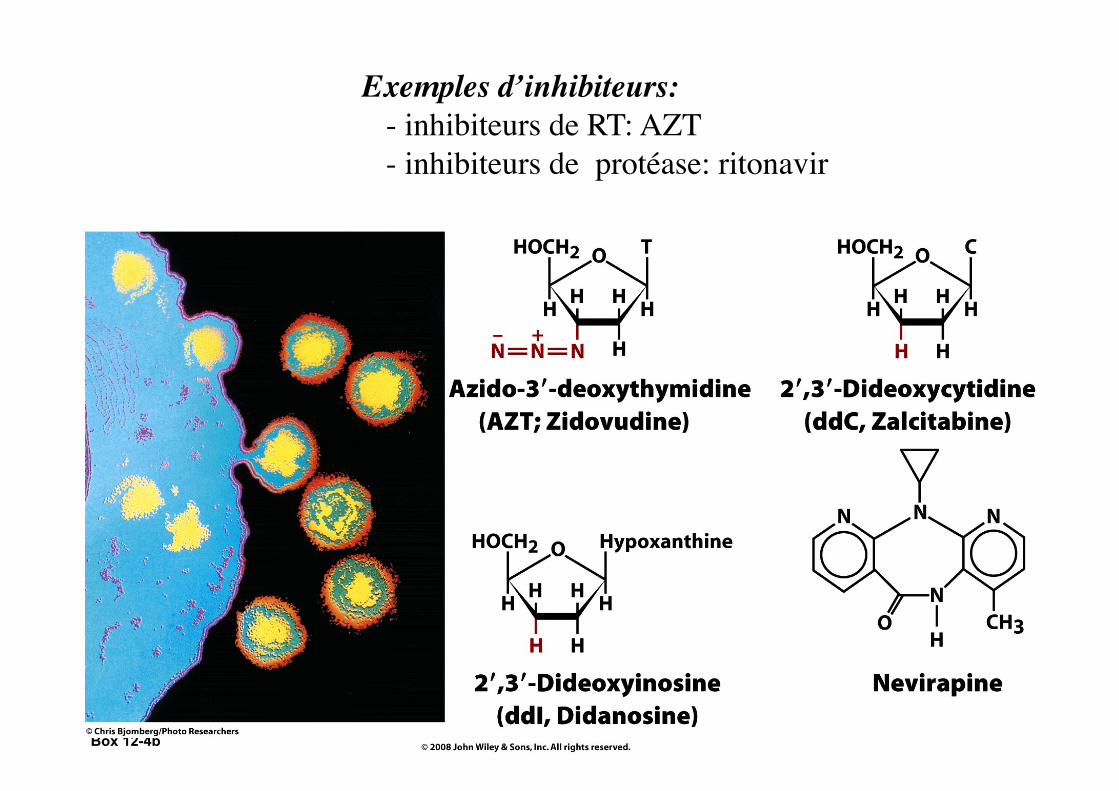
Box 12-4b
Exemples d’inhibiteurs: - inhibiteurs de RT: AZT - inhibiteurs de protéase: ritonavir

Box 12-4c
Exemples d’inhibiteurs: - inhibiteurs de RT: AZT - inhibiteurs de protéase: ritonavir


Figure 12-11
Régulation enzymatique : Allostérie
Exemple: Asp Transcarbamoylase

Page 386
Régulation enzymatique : Allostérie
Exemple: Asp Transcarbamoylase

Figure 12-10
Régulation enzymatique : Allostérie
Exemple: Asp Transcarbamoylase

Figure 12-12
Régulation enzymatique : Allostérie
C6r6 : 6 ss-unités catalytiques + 6 ss-unités régulatrices
Exemple: Asp Transcarbamoylase

PALA est un analogue bi-substrat qui mime l’intermediaire de reaction formant le carbamoyl aspartate
Il se lie au site active de la sous-unité catalytique
Régulation enzymatique : Allostérie
Exemple: Asp Transcarbamoylase

Figure 12-13
Exemple: Asp Transcarbamoylase
Régulation enzymatique : Allostérie

Propriétés Générales des Enzymes Allosteriques 1. L’activité est modifiée par des inhibiteurs ou activateurs
(modulateurs)
2. Les modulateurs allosteriques se lient de manière non-covalente à l’enzyme
3. Les enzymes ont une structure quaternaire
4. Les enzymes présentent une courbe sigmoidale vo versus [S] (cooperativité positive de sites multiples de liaison du substrat)
5. Il y a transition rapide entre la conformation active (R) et la conformation inactive (T)
6. Substrats et activateurs ne peuvent se lier que dans la configuration R et les inhibiteurs seulement dans la configuration T.
Régulation Allosterique

Transition rapide entre états R et T • L’addition de S: augmente la concentration de l’état R • L’addition de I: augmente la concentration de l’état T • L’activateur se lie de preference à R, donc augmente le rapport R/T
Role de la cooperativité de liaison sur la régulation:
• L’addition de l’agent allostérique modifie l’activité de l’enzyme
• L’activateur peut diminuer Km, l’inhibiteur peut augmenter Km
Régulation Allosterique

Deux Theories de Régulation allosterique
Modèle Concerté Monod-Wyman-Changeux (theorie basée sur la symmetrie)
• Il n’existe que 2 conformations: R et T
• la symmetrie est maintenue lors de la transition entre l’état R et l’état T)
• Les sous-unités sont toutes soit R soit T
• R = forte affinité pour S T = faible affinité pour S
• La liaison de S déplace l’équilibre vers l’état R
• La liaison de I déplace l’équilibre vers l’état T
Régulation Allosterique

Deux Theories de Régulation allosterique
• un ligand peut induire un changement de la structure de la sous-unité sur laquelle il se lie
• Le changement conformationel d’une sous-unité affecte la conformation de la sous-unité voisine
• Un mélange des deux formes de sous-unité R (haute affinité pour S) et T (faible affinité pour S) peut exister
=> la symmetrie ne doit pas nécessairement être conservée
Modèle Sequentiel (theorie d’induction par le ligand)
Régulation Allosterique

(b) Modèle séquentiel : Mélange possible de
sous-unités T et de sous-unités R. Liaison
de S convertit seulement la sous-unité concernée de T vers R
(a) Modèle concerté Monod-Wyman-Changeux: sous-unités toutes à l’état T
ou toutes à l’état R
Régulation Allosterique

Modèle séquentiel: Exemple de l’Hémoglobine Changement conformationel
lors de la liaison de O2 sur l’hémoglobine
• Oxygène se lie à Hb
• correspond en partie au modèle sequentiel et en partie au modèle concerté
Régulation Allosterique

Page 390
Régulation Enzymatique: phosphorylation
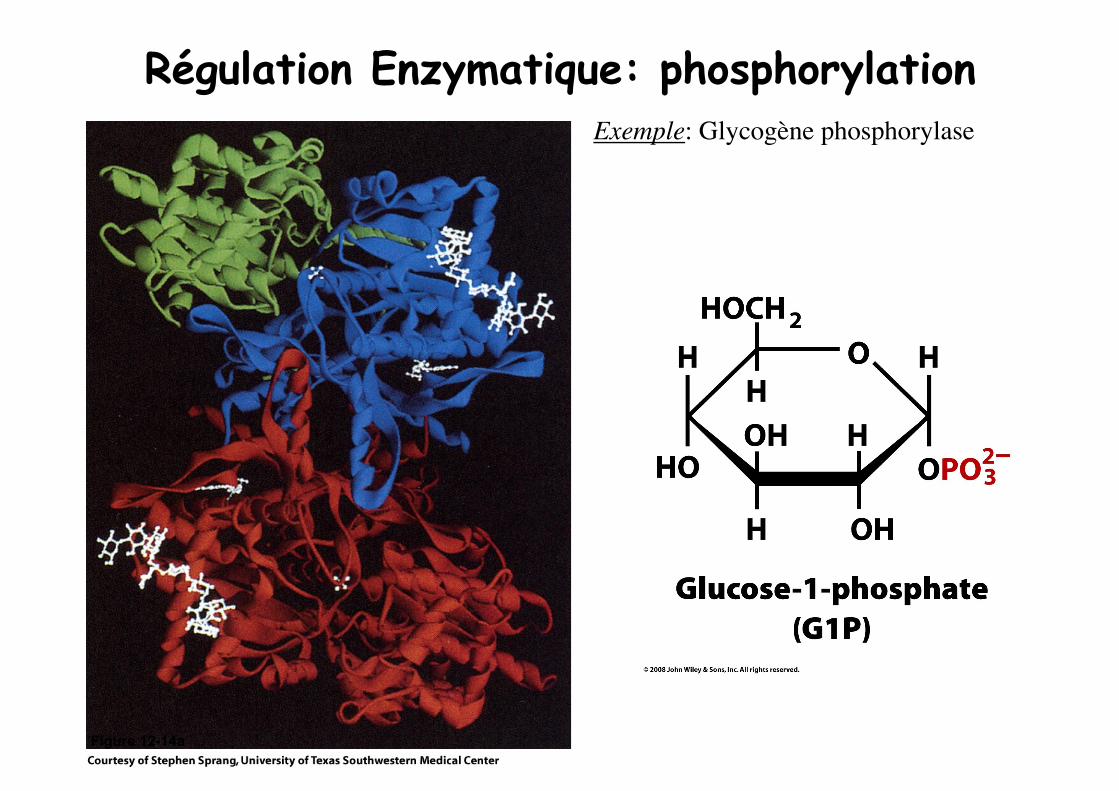
Figure 12-14a
Régulation Enzymatique: phosphorylation Exemple: Glycogène phosphorylase
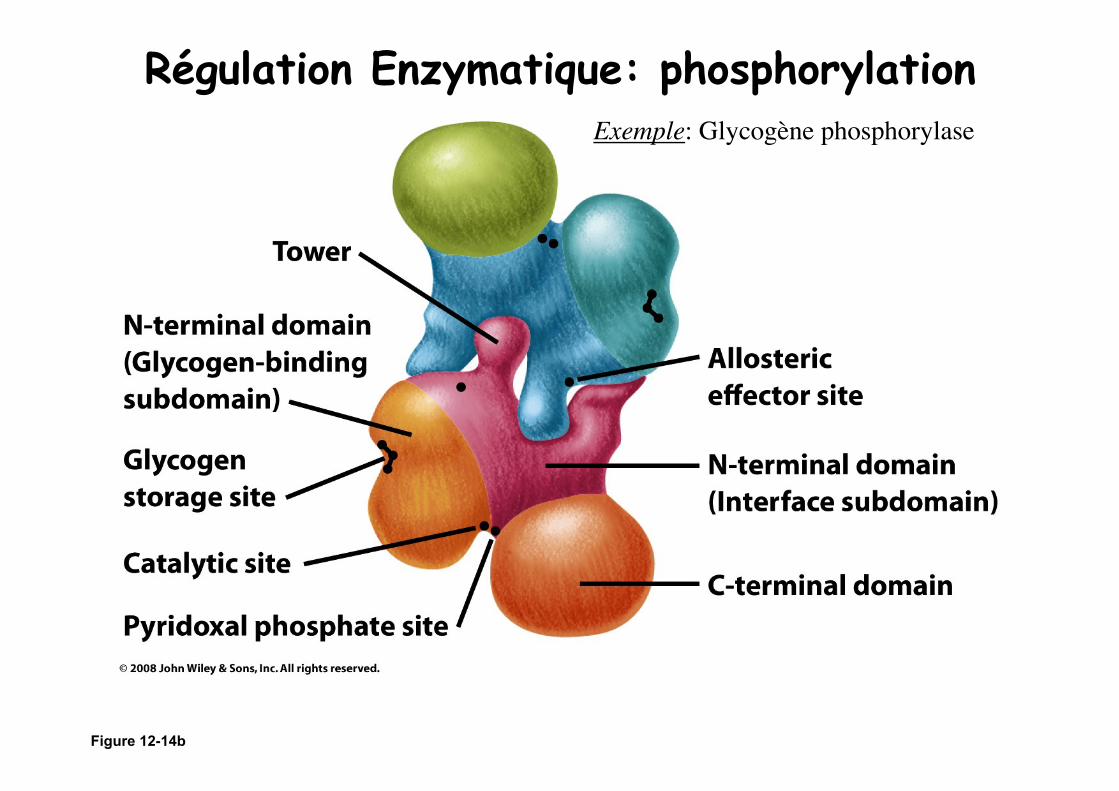
Figure 12-14b
Régulation Enzymatique: phosphorylation Exemple: Glycogène phosphorylase

Figure 12-15
Régulation Enzymatique: phosphorylation Exemple: Glycogène phosphorylase

Figure 12-16
Régulation Enzymatique: phosphorylation Exemple: Glycogène phosphorylase

Figure 12-17
Drug Design

Détoxification: rôle du Cytochrome P450 (une mono-oxygénase)