probing non-selective cation binding in the hairpin ...rnapeopl/walterlabpub/walter(00b).pdf ·...
TRANSCRIPT

doi:10.1006/jmbi.2000.3691 available online at http://www.idealibrary.com on J. Mol. Biol. (2000) 298, 539±555
Probing Non-Selective Cation Binding in the HairpinRibozyme with Tb(III)
Nils G. Walter*, Ning Yang and John M. Burke*
Markey Center for MolecularGenetics, Department ofMicrobiology and MolecularGenetics, The University ofVermont, 306 Stafford HallBurlington, VT 05405, USA
Present address: N. G. Walter, DeChemistry, University of Michigan,48109-1055, USA.
Abbreviations used: FRET, ¯uoreenergy transfer.
E-mail addresses of the [email protected]; [email protected]
0022-2836/00/030539±17 $30.00/0
Catalysis by the hairpin ribozyme is stimulated by a wide range of bothsimple and complex metallic and organic cations. This independencefrom divalent metal ion binding unequivocally excludes inner-spherecoordination to RNA as an obligatory role for metal ions in catalysis.Hence, the hairpin ribozyme is a unique model to study the role ofouter-sphere coordinated cations in folding of a catalytically functionalRNA structure. Here, we demonstrate that micromolar concentrations ofa deprotonated aqueous complex of the lanthanide metal ion terbium(III),Tb(OH)(aq)2�, reversibly inhibit the ribozyme by competing for a crucial,yet non-selective cation binding site. Tb(OH)(aq)2� also reports a likelylocation of this binding site through backbone hydrolysis, and permitsthe analysis of metal binding through sensitized luminescence. We pro-pose that the critical cation-binding site is located at a position within thecatalytic core that displays an appropriately-sized pocket and a highnegative charge density. We show that cationic occupancy of this site isrequired for tertiary folding and catalysis, yet the site can be productivelyoccupied by a wide variety of cations. It is striking that micromolarTb(OH)(aq)2� concentrations are compatible with tertiary folding, yetinterfere with catalysis. The motif implicated here in cation-binding hasalso been found to organize the structure of multi-helix loops in evol-utionary ancient ribosomal RNAs. Our ®ndings, therefore, illuminategeneral principles of non-selective outer-sphere cation binding in RNAstructure and function that may have prevailed in primitive ribozymes ofan early ``RNA world''.
# 2000 Academic Press
Keywords: footprinting; hairpin ribozyme; lanthanide ¯uorescence;non-speci®c metal ion binding; outer-sphere cation binding pocket
*Corresponding authorsIntroduction
The discovery of catalytic function in certainRNAs, termed ribozymes, spurred great interest inthe underlying reaction mechanisms (Cech, 1993;Narlikar & Herschlag, 1997; Been & Wickham,1997; Sigurdsson et al., 1998; Wedekind & McKay,1998; Walter & Burke, 1998). Divalent metal ionswere soon identi®ed as essential cofactors for cata-lysis, and ribozymes were generally described asmetalloenzymes (Pan et al., 1993; Yarus, 1993;Steitz & Steitz, 1993; Smith, 1995; Pyle, 1993, 1996).
partment ofAnn Arbor, MI
scence resonance
ding authors:m.edu
Divalent metal ions are thought to play two majorroles in RNA catalysis: (1) an indirect role, inwhich they fold a functional RNA structure byneutralizing and bridging the negatively chargedphosphoribose backbone; and (2) a direct role inreaction chemistry, presumably activating reactantsby Lewis-acid type inner-sphere coordination.These two roles have proven dif®cult to dissect.
Recently, the hairpin ribozyme has been foundto maintain high levels of site-speci®c cleavageactivity independent of divalent metal ions, in thepresence of the kinetically inert metal ion complexcobalt(III) hexammine (Hampel & Cowan, 1997;Nesbitt et al., 1997; Young et al., 1997), high mono-valent cation concentrations (Murray et al., 1998;Nesbitt et al., 1999), or organic polyamines such asspermine (Earnshaw & Gait, 1998). These ®ndingsunequivocally exclude inner-sphere coordination toRNA as an obligatory role for metal ions in cataly-sis. Hence, the hairpin ribozyme can serve as a
# 2000 Academic Press

540 Cation Binding in the Hairpin Ribozyme
model to understand the role of a variety of outer-sphere coordinated cations in folding of a catalyti-cally functional RNA structure, independent of anydirect involvement in reaction chemistry.
Several ribozymes share this unexpected lack ofselectivity in cation cofactor, including the ham-merhead and VS ribozymes (Murray et al., 1998),as well as RNA and DNA enzymes found throughin vitro selection from random pools (Geyer & Sen,1997; Faulhammer & Famulok, 1997; Jayasena &Gold, 1997; Suga et al., 1998). In all of these cases,the catalytic motifs are small (30-150 nt), and cata-lysis comprises a relatively simple, albeit speci®cnucleophilic substitution reaction, typically a phos-phodiester transfer. Larger ribozymes with a morecomplex function, such as group I and group IIintrons or RNase P RNA, require more speci®cmetal ion cofactors (Celander & Cech, 1991; Smithet al., 1992; Cate et al., 1996, 1997; Cate & Doudna,1997; Basu et al., 1998). These observations raisethe possibility that primitive catalytic RNAs withnon-selective metal-ion binding sites may haveemerged ®rst in an ``RNA world'', subsequentlyevolving into larger catalysts with the ability tobury speci®c metal ions in their complex tertiarystructure, exploiting them for more elaborate tasks(Feig & Uhlenbeck, 1999). Studying the hairpinribozyme with its broad cofactor spectrum, there-fore, may allow a glimpse at the non-selectiveouter-sphere cation-binding sites of primitive ribo-zymes at the early stages of life.
The hairpin ribozyme is found in satellite RNAsassociated with plant viruses such as the tobaccoringspot virus (Buzayan et al., 1986; Hampel &Tritz, 1989). These virus satellites proliferate by
Figure 1. The hairpin ribozyme and its reaction pathwaymized construct SV5 EH4 Rz from Esteban et al. (1997) was ubinds the 14-nucleotide substrate (small letters) through its sconnected via a ¯exible hinge to domain B of the ribozymedomains are characterized by a highly conserved internal loarrow indicates the cleavage site. (b) Minimal reaction pathw(Walter et al., 1998). Substrate in trans (S) is bound by the ristructural isomer is ¯exible (curved double arrows) and, ustructure, enabling loops A and B to interact (short doublearrow), the complex undocks, and the 50 and 30 cleavage pcan be characterized by individual rate constants as indicateation, but not cleavage, where a subsequent, more local coet al., 1998). In the present work, we investigate the interfecomplex Tb(OH)(aq)2� with a post-docking step.
recruiting the viral replicase for a double rolling-circle replication of their circular positive-strandRNA genome. The hairpin ribozyme motif isrequired for self-cleavage and ligation of the repli-cated negative-strand polymers into cyclic mono-mers that can serve as templates in positive-strandsynthesis. In its natural con®guration, the hairpinribozyme is embedded in an RNA four-way junc-tion, that can be truncated into a hinged two-wayjunction to reversibly cleave a substrate in trans(Hampel & Tritz, 1989). The two essential domainscontribute determinants for substrate binding(domain A) and catalysis (domain B). Bothdomains fold independently, each forming a highlyconserved internal loop ¯anked by two helices(Figure 1(a)) (Walter & Burke, 1998). Site-speci®csubstrate cleavage and ligation reactions requireloops A and B to interact, and this docking eventinvolves a sharp bend about the interdomain junc-tion (Walter & Burke, 1998). The natural four-wayjunction serves as scaffold to stabilize docking ofthe two domains (Walter et al., 1999), presumablyrecruiting two multivalent metal ions, at least oneof which binds at the helical junction (Murchieet al., 1998). As activity requires higher metal ionconcentrations than docking, yet another, lower-af®nity metal-ion binding site has been proposedto be essential for activity of the four-way junctionribozyme (Thomson & Lilley, 1999).
In the two-way junction ribozyme, domain dock-ing and cleavage activity show parallel metal iondependencies (Walter et al., 1998, 1999), suggestingthat the same lower-af®nity metal ion bindingevent is responsible for both. As this junctiondesign is extensively used for targeted RNA inacti-
. (a) Ribozyme-substrate complex. The kinetically opti-sed throughout this study. The ribozyme (capital letters)ubstrate-binding strand to form domain A. Domain A iswhich contributes essential catalytic determinants. Bothop with ¯anking Watson-Crick base-paired helices. Theay of hairpin ribozyme catalysis, as derived previously
bozyme (Rz) into an open (extended) conformation. Thispon addition of cation Mn�, folds into a bent (docked)arrow). Subsequently, site-speci®c cleavage occurs (shortroducts (50P, 30P) dissociate. All steps are reversible andd. Domain docking was found to be rate-limiting for lig-nformational change might be the slowest step (Walterrence of micromolar concentrations of the deprotonated

Cation Binding in the Hairpin Ribozyme 541
vation in mammalian cells (Welsh et al., 1996;Earnshaw & Gait, 1997), there is considerable inter-est in understanding the mechanism of substratecleavage. We have used ¯uorescence based kineticassays to dissect the reaction pathway into severalconformational transitions (Walter & Burke, 1997;Walter et al., 1998). Domain docking was found tobe an essential global folding step, preceding therate-limiting event in catalysis (Figure 1(b)) (Walteret al., 1998). As the observed cleavage rate is pHindependent (Nesbitt et al., 1997), and since dockedribozyme-substrate and ribozyme-product com-plexes show the same solvent protected core(Hampel et al., 1998), the rate-limiting step maywell be a localized conformational transition fol-lowing docking.
Recently, the solution structures of the isolateddomains A and B of the ribozyme-substrate com-plex have been determined by NMR (Cai &Tinoco, 1996; Butcher et al., 1999). NMR spectrawere obtained, however, at low ionic strength, inthe absence of multivalent cations. Since domaindocking is not detectable under these conditions(Walter et al., 1998, 1999; Hampel et al., 1998), itseems reasonable to assume that multivalent metalion binding has an impact on these structures in amanner to favor interdomain interactions.Recently, a molecular model of these interactions,implementing a ``ribose zipper'' motif, was pro-posed based on cross-linking interference exper-iments (Earnshaw et al., 1997; Pinard et al., 1999a).However, we have evidence for a more intimate``induced ®t'' between loops A and B in the cataly-tic core of the hairpin ribozyme-substrate complexas a prerequisite for site-speci®c substrate cleavage(Pinard et al., 1999b).
Here, we have characterized the lanthanidemetal ion terbium(III) as a potent inhibitor of cata-lysis by the hairpin ribozyme. It functions as acompetitive inhibitor for all cations known to pro-mote activity, reversibly blocking a step in thereaction pathway following domain docking, poss-ibly the rate-limiting step. As inhibition is stronglypH dependent, only the deprotonated complexTb(OH)(aq)2� appears to inhibit. We were also ableto identify a high af®nity binding site ofTb(OH)(aq)2� in the catalytic core of the ribozymeby a slow backbone scission reaction, and proposethat this site is a likely candidate for inducing theobserved inhibition. In the NMR structure of loopB, this non-selective binding pocket for cations islocated in a region of high negative surface chargepotential.
Results
Terbium(III) inhibits the hairpin ribozyme bycompeting with all cations thatsupport catalysis
Terbium(III) is the most stable oxidation state ofthe eighth of the twelve lanthanide transitionmetals. It has been used, together with europiu-
m(III), as a probe for binding sites of metal ions,particularly Mg2� and Ca2�, in protein enzymes(Horrocks et al., 1977; Evans, 1990; Horrocks, 1993;Frey et al., 1996; Dickeson et al., 1998). Otherlanthanide ions, in particular samarium(III) andluthetium(III), have been used to solve the phaseproblem in X-ray diffraction analysis of tRNA crys-tals, by providing isomorphic heavy-atom deriva-tives (Jack et al., 1977). They were found to oftenbind to sites on RNA identical with Mg2�-bindingsites. More recent studies have shown that ter-bium(III) (Feig et al., 1998) and lanthanum(III)(Lott et al., 1998) interfere with activity of thehammerhead ribozyme, by competing with Mg2�
binding. These results prompted us to investigatethe impact of terbium(III) on hairpin ribozymeactivity to better understand the role of metal ionsin catalysis by RNA.
For activity interference studies we employed astructurally well-behaved and kinetically well-characterized variant of the hairpin ribozyme (SV5EH4 Rz as described by Esteban et al. (1997);Figure 1(a)). This construct has been shown toform a stably docked conformational intermediatebefore proceeding to the rate-limiting step of cata-lysis (Walter et al., 1998, 1999). Ribozyme andradiolabeled substrate were combined undersingle-turnover (pre-steady-state) conditions (ribo-zyme excess) at pH 7.5, in the presence of 200 mMNa�, to ensure that all substrate strands wereincorporated into yet inactive complexes. Cleavagewas initiated by the addition of a mixture of ter-bium(III) and a second, catalytically pro®cient cat-ion. Cleavage rate constants were determinedusing standard methods (see Materials andMethods), and their normalized values wereplotted as inhibition or activation curves over arange of cation concentrations (Figure 2). Resultsshow that terbium(III) is an ef®cient inhibitor of allcations that support catalysis, including the hardand soft divalent metal ions magnesium (Mg2�)and manganese (Mn2�), respectively, the kineticallyinert trivalent metal ion complex cobalt(III) hexam-mine (Co(NH3)6
3�), the organic triamine spermidine(spd���), and the monovalent metal ion sodium(Na�) (Figure 2, left panel). The observed inhibitionconstants Ki,app under the chosen conditions are inthe narrow range of 12 to 33 mM. In all cases, ter-bium(III)-mediated inhibition can be reversed byincreasing the concentration of the catalyticallypro®cient cation, and the apparent binding con-stant KD,app for the cation is higher in the presencethan in the absence (value in parenthesis) of terbiu-m(III) (Figure 2, right panel). These observationsstrongly suggest a competition between terbiu-m(III) and the catalytically pro®cient cations for acommon binding site. Consistent with this notion,activity in a reaction stalled in the presence of12 mM Mg2� and 50 mM terbium(III) could par-tially be rescued by subsequently increasing theMg2� concentration to 500 mM (data not shown).
Apparent cooperativity coef®cients for terbiu-m(III) are high (Figure 2), presumably indicative of
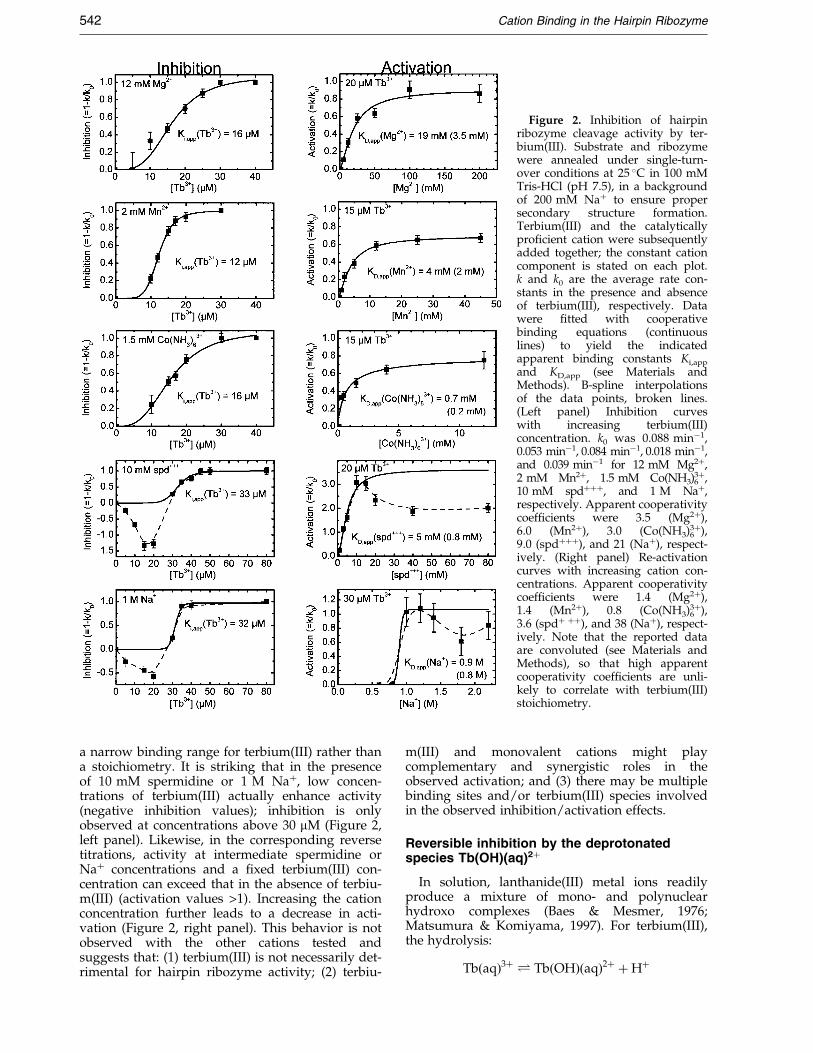
Figure 2. Inhibition of hairpinribozyme cleavage activity by ter-bium(III). Substrate and ribozymewere annealed under single-turn-over conditions at 25 �C in 100 mMTris-HCl (pH 7.5), in a backgroundof 200 mM Na� to ensure propersecondary structure formation.Terbium(III) and the catalyticallypro®cient cation were subsequentlyadded together; the constant cationcomponent is stated on each plot.k and k0 are the average rate con-stants in the presence and absenceof terbium(III), respectively. Datawere ®tted with cooperativebinding equations (continuouslines) to yield the indicatedapparent binding constants Ki,app
and KD,app (see Materials andMethods). B-spline interpolationsof the data points, broken lines.(Left panel) Inhibition curveswith increasing terbium(III)concentration. k0 was 0.088 minÿ1,0.053 minÿ1, 0.084 minÿ1, 0.018 minÿ1,and 0.039 minÿ1 for 12 mM Mg2�,2 mM Mn2�, 1.5 mM Co(NH3)6
3�,10 mM spd���, and 1 M Na�,respectively. Apparent cooperativitycoef®cients were 3.5 (Mg2�),6.0 (Mn2�), 3.0 (Co(NH3)6
3�),9.0 (spd���), and 21 (Na�), respect-ively. (Right panel) Re-activationcurves with increasing cation con-centrations. Apparent cooperativitycoef®cients were 1.4 (Mg2�),1.4 (Mn2�), 0.8 (Co(NH3)6
3�),3.6 (spd� ��), and 38 (Na�), respect-ively. Note that the reported dataare convoluted (see Materials andMethods), so that high apparentcooperativity coef®cients are unli-kely to correlate with terbium(III)stoichiometry.
542 Cation Binding in the Hairpin Ribozyme
a narrow binding range for terbium(III) rather thana stoichiometry. It is striking that in the presenceof 10 mM spermidine or 1 M Na�, low concen-trations of terbium(III) actually enhance activity(negative inhibition values); inhibition is onlyobserved at concentrations above 30 mM (Figure 2,left panel). Likewise, in the corresponding reversetitrations, activity at intermediate spermidine orNa� concentrations and a ®xed terbium(III) con-centration can exceed that in the absence of terbiu-m(III) (activation values >1). Increasing the cationconcentration further leads to a decrease in acti-vation (Figure 2, right panel). This behavior is notobserved with the other cations tested andsuggests that: (1) terbium(III) is not necessarily det-rimental for hairpin ribozyme activity; (2) terbiu-
m(III) and monovalent cations might playcomplementary and synergistic roles in theobserved activation; and (3) there may be multiplebinding sites and/or terbium(III) species involvedin the observed inhibition/activation effects.
Reversible inhibition by the deprotonatedspecies Tb(OH)(aq)2�
In solution, lanthanide(III) metal ions readilyproduce a mixture of mono- and polynuclearhydroxo complexes (Baes & Mesmer, 1976;Matsumura & Komiyama, 1997). For terbium(III),the hydrolysis:
Tb�aq�3� � Tb�OH��aq�2� �H�

Cation Binding in the Hairpin Ribozyme 543
has an equilibrium constant of Keq � 10ÿ7.9 M (atlow ionic strength) and yields signi®cant amountsof the deprotonated mononuclear speciesTb(OH)(aq)2� at pH 7.5 (Baes & Mesmer, 1976).From this intermediate, other species such asTb(OH)2(aq)�, Tb2(OH)2(aq)4�, or Tb3(OH)5(aq)4�
can form, ultimately leading to the precipitation ofterbium hydroxide, Tb(OH)3, at high pH and ter-bium(III) concentrations. In the handling of our sol-utions, we carefully avoided formation of anyhydroxide precipitate (see Materials and Methods).However, it seemed important to determine whichof the soluble hydroxo complex(es) mediates
inhibition is reversible. Two parallel reactions were initiatedpH 6.5, resulting in rapid accumulation of cleavage producwas subjected to a change in pH to 7.5, completely stallingwere changed back to pH 6.5, and catalysis resumed to similous line). B-spline interpolations of data points, lines. (e) Tconditions of terbium(III)-mediated inhibition of cleavage (1¯uorescence resonance energy transfer (FRET) assay (Waltesame level of domain docking (monitored as increase in reanalog) complex as does a reference reaction in the absence oto the data points, yielding docking rate constants kdock of 0.presence and absence of terbium(III), respectively.
inhibition, and whether the various terbium(III)complexes in solution might have distinct effectson hairpin ribozyme activity, a possibility raisedby our experiments with monovalent cations (seeabove).
Decreasing pH from 7.5 to 6.5 had a dramaticeffect on terbium(III)-mediated inhibition(Figure 3(a)); in the presence of 12 mM Mg2�,addition of 20 to 100 mM terbium(III) nowenhances activity nearly threefold. However, atconcentrations of 300 mM and above, completeinhibition was observed, as before. Terbium(III)can also promote hairpin ribozyme activity in the
Figure 3. Properties of terbiu-m(III)-mediated inhibition of thehairpin ribozyme at 25 �C. (a) Rateconstants k (see Materials andMethods) with increasing terbiu-m(III) concentration at pH 6.5 inthe presence (®lled squares, con-tinuous line) and absence (open cir-cles, broken line) of 12 mM Mg2�.Lines represent B-spline interpola-tions of data points. In the presenceof Mg2�, an inhibition constant forterbium(III) of 240 mM can be cal-culated from the data (seeMaterials and Methods). (b) Depen-dence of the inverse of the appar-ent inhibition constant ofterbium(III) on hydroxide ion con-centration in the presence of each12 mM Mg2� and terbium(III).Non-linear regression (continuousline) using the cooperativityequation (see Materials andMethods) yields a cooperativitycoef®cient of close to 1, and anapparent KOHÿ
D of 10ÿ6.4 M (equival-ent to a titration midpoint atpH 7.6). (c) Average rate constantsk (see Materials and Methods) withincreasing lanthanum(III) concen-tration at pH 7.5 in the presence(®lled squares, continuous line) andabsence (open circles, broken line)of 12 mM Mg2�. Lines represent B-spline interpolations of data points.In the presence of Mg2�, an inhi-bition constant for lanthanum(III)of 97 mM can be calculated fromthe data. (d) Terbium(III)-mediated
by the addition of 12 mM Mg2� and 50 mM Tb3�, att (see also (a)). One reaction (open circles, broken line)catalysis (see also Figure 2). After 20 minutes conditionsar levels as the reference reaction (®lled squares, continu-erbium(III) does not interfere with domain docking. At2 mM Mg2�, 50 mM Tb3� (pH 7.5); see also Figure 2) ar et al., 1998) (see Materials and Methods) reveals thelative FRET ef®ciency) in the ribozyme-substrate (dAÿ1
f Tb3�. Continuous lines represent single-exponential ®ts63 minÿ1 and 0.55 minÿ1 and identical amplitudes in the

544 Cation Binding in the Hairpin Ribozyme
absence of Mg2� at pH 6.5, with a similar asym-metric bell-shaped concentration dependence as inthe presence of Mg2� (Figure 3(a)).
Determining the apparent inhibition constantKi,app over a range of hydroxide ion concentrationsyields a cooperativity coef®cient with respect toOHÿ close to 1 and a titration midpoint of pH 7.6(Figure 3(b)). Both values strongly suggest thatinhibition is mediated by the most prevalentspecies in solution, Tb(OH)(aq)2�, as this complexcontains one OHÿ ligand with a titration midpointof pH 7.9 (see above).
To further test this hypothesis, we examinedlanthanum(III) as a potential inhibitor of the hair-pin ribozyme. As the ®rst in the row of lanthanideions, lanthanum(III) has a slightly larger ionicradius than terbium(III) (1.15 AÊ , compared to0.92 AÊ ). As a result, it has a slightly higherhydrolysis equilibrium constant of Keq � 10ÿ8.5 M(at low ionic strength) and is fourfold less deproto-nated at pH 7.5 than terbium(III). Accordingly,hairpin ribozyme activity showed a bell-shapeddependence on lanthanum(III) concentration atpH 7.5, both in the presence and absence of Mg2�
(Figure 3(c)), similar to the curves observed for ter-bium(III) at pH 6.5 (Figure 3(a)). The apparent inhi-bition constant for lanthanum(III) in the presenceof 12 mM Mg2� is sixfold higher than that of ter-bium(III) at the same pH (97 mM versus 16 mM), inreasonable agreement with their difference indeprotonation equilibrium, and consistent withLa(OH)(aq)2� as the inhibitory species. As anenhancement of hairpin ribozyme activity is onlyseen at low pH, it seems likely that the activationby low concentrations of either terbium(III) (pH 6.5or below) or lanthanum(III) (pH 7.5) is mediatedby the fully protonated aqueous complexesTb(aq)3� and La(aq)3�, respectively.
This strong effect of pH on the inhibition by ter-bium(III) allows for a simple reversibility test. Twoparallel reactions were initiated at pH 6.5, in thepresence of 12 mM Mg2� and 50 mM terbium(III),leading to the rapid accumulation of cleavage pro-duct. After two minutes, the pH of one of the tworeactions was changed to 7.5, completely suppres-sing cleavage. After 20 minutes, this reaction wasbrought back to pH 6.5, resulting in the recoveryof cleavage activity to similar extents as the controlreaction (Figure 3(d)) and proving that inhibitionindeed is readily reversible.
What is the mode of action for terbium(III)-mediated reversible inhibition of the hairpinribozyme? Terbium(III) at low concentrations haspreviously been found to alter the geometry of thesugar-phosphate backbone and the base stackinginteractions in nucleic acids (Gersanovski et al.,1985). This interference can be expected to bereversible and has the potential to impair essentialstructural transitions in the reaction pathway of acatalytic RNA. Under our experimental conditions,the hairpin ribozyme-substrate complex is formedprior to the addition of terbium(III). A subsequentand essential structural transition of the complex is
known to involve folding from an extended to adocked conformation, in which the substrate-bind-ing and catalytic domains interact, enabling cataly-sis (Figure 1(b)) (Walter et al., 1998, 1999; Hampelet al., 1998). A convenient way to monitor this glo-bal folding event is as an increase in ¯uorescenceresonance energy transfer (FRET) between adomain-terminal donor/acceptor ¯uorophore pair(Walter et al., 1998). Figure 3(e) shows a compari-son of FRET-monitored domain docking in the pre-sence and absence of 50 mM terbium(III), aconcentration suf®cient to completely stall cleavageactivity under these conditions (12 mM Mg2�,pH 7.5). Clearly, formation of the docked confor-mer is not inhibited by the presence of terbium(III),suggesting that the mechanism of inhibitioninvolves a step after domain docking and, hence,closer to the chemical transition state (Figure 1(b)).This result was con®rmed at terbium(III) concen-trations of up to 100 mM (above which the FRETdonor ¯uorescein becomes strongly quenched byterbium(III)) and by mapping the solvent-protectedcore of the ribozyme-substrate complex in the pre-sence and absence of terbium(III), employinghydroxyl radical footprinting (Hampel et al., 1998;Ken J. Hampel, personal communication).
Energy transfer from RNA bases enablesmonitoring terbium(III) binding to the hairpinribozyme-substrate complex
Terbium(III) has been known to bind to RNAand, as a result, to ¯uoresce upon excitation of theRNA, through energy transfer particularly fromnearby guanine bases (Yonushot et al., 1978; Topal& Fresco, 1980; Wenzel & Collette, 1988). Wefound that this effect can be used to monitor thekinetics and thermodynamics of terbium(III) bind-ing to the hairpin ribozyme-substrate complex in abackground of 12 mM Mg2�. Figure 4(a) shows thestrong enhancement of the ¯uorescence emissionspectrum of terbium(III) by addition of the RNAcomplex. In a stopped-¯ow kinetic experiment, therapid binding of a millimolar excess of terbium(III)to micromolar RNA concentrations can be resolvedinto at least three different processes with discern-able rate constants of 41 sÿ1, 2.9 sÿ1, and 0.25 sÿ1
(Figure 4(b)). 66 % of the ¯uorescence increase isassociated with the fastest rate, suggesting thatbinding is driven by electrostatic attraction of ter-bium(III) cations to areas on the RNA with highnegative charge potential.
An equilibrium titration of the ribozyme-sub-strate complex with terbium(III) in 12 mM Mg2�
indicates at least three distinct classes of bindingsites with af®nities of 9 mM, 120 mM, and 900 mM,respectively (Figure 4(c)), similar to previous obser-vations on lanthanide(III) ion binding to tRNA(Draper, 1985). Note that the highest af®nity bind-ing is observed at similar concentrations as theinhibition of ribozyme activity (Ki,app � 16 mMunder these conditions, see Figure 2).

Figure 4. Fluorescence detectionof terbium(III) binding to the hair-pin ribozyme-substrate complex in50 mM Tris-HCl (pH 7.5), 12 mMMg2�, at 25 �C. (a) When excited at290 nm, 2 mM terbium(III) boundto 1 mM ribozyme-substrate com-plex (dAÿ1 analog) exhibits acharacteristic ¯uorescence spectrum(continuous line). In the absence ofRNA, only a low background sig-nal is observed (broken line). (b) Ina stopped-¯ow ¯uorescence exper-iment (1 datum per millisecond;excitation, 290 nm; emission detec-tion: 545 nm; 29 individual timetraces averaged), the binding kin-etics of 2 mM terbium(III) to 1 mMribozyme-substrate complex (dAÿ1
analog) reveal at least three separ-ate rate constants (relative ampli-tudes in parenthesis; ®t: continuousline), not seen in the absence ofRNA. (c) Binding of terbium(III) to
1 mM ribozyme-substrate complex (dAÿ1 analog) can be monitored by ¯uorescence titration over several orders ofmagnitude of terbium(III) concentration (excitation, 290 nm; emission detection, 545 nm). Fits assuming a single bind-ing af®nity (broken lines) accurately represent the data only in narrow concentration ranges. The best ®t to the com-plete data set (continuous line in right panel) was obtained when ®tting to the sum of three independent Hillequations (see Materials and Methods), revealing apparent dissociation constants of 9 mM, 120 mM, and 900 mM.
Cation Binding in the Hairpin Ribozyme 545
Terbium(III) at millimolar concentrationsfootprints RNA secondary and tertiarystructure, while micromolar concentrationsreport only on the tightest binding site
Terbium(III), like other lanthanide and heavy-metal ions such as lead, has been used to detectspeci®c metal-ion binding sites in RNA (Jack et al.,1977; Ciesiolka et al., 1989; Evans, 1990; Gast et al.,1996; Feig & Uhlenbeck, 1999) or to degrade RNAfor biotechnological applications (Stein & Cohen,1988; Morrow, 1996; Matsumura & Komiyama,1997), taking advantage of a metal-ion-promotedhydrolytic backbone scission reaction. We foundthat high (12 mM) concentrations of terbium(III)produce a clear footprinting pattern of the second-ary and tertiary structure of (50-32P)-labeled hairpinribozyme. Under these conditions, terbium(III)cleaves the RNA backbone in a sequence-indepen-dent manner, preferentially cutting single-strandedor non-Watson-Crick base-paired regions such asloops A and B, or the GNRA tetraloop closinghelix 4 of the hairpin ribozyme (Figure 5). We usedorder-of-addition experiments to characterize thisstructure probing method.
At low concentrations of terbium(III), the pri-mary scission products co-migrate with the 20,30-cyclic phosphate products of the control alkali lad-der, while only higher terbium(III) concentrationsconvert the primary products into the higher-mobi-lity 20- or 30-phosphate fragments (best visible bycomparing region H1 of lanes 20 to 23 with that oflane ``OHÿ``, Figure 5(a)). This observation is con-sistent with the previously proposed two-step
mechanism for metal-ion-promoted RNA backbonescission (Pan & Uhlenbeck, 1992), involving, ®rst,deprotonation of a 20-hydroxyl group and nucleo-philic attack of the resulting oxyanion on the adja-cent 30,50-phosphodiester to form 20,30-cyclicphosphate and 50-hydroxyl termini, and, second,hydrolysis of the cyclic phosphate to yield the cor-responding mixture of 20- and 30-phosphomonoe-sters.
Addition of 12 mM terbium(III) to the hairpinribozyme prior to addition of substrate produced afootprint similar to that in the absence of substrate(compare lane 7 with lane 6, Figure 5(a)), with thesingle-stranded substrate-binding strand (compris-ing regions H1, loop A, and H2) strongly and uni-formly hit. Hence, millimolar concentrations ofterbium(III) can prevent substrate binding to thehairpin ribozyme. In contrast, when the substrateis allowed to bind before addition of terbium(III),the substrate-binding strand becomes strongly pro-tected from scission, in particular helices H1 andH2 (lane 8, Figure 5(a)). In this case, the footprint-ing pattern is independent of whether Mg2� isadded after or together with terbium(III) (comparelane 8 with lane 9, Figure 5(a)), indicating thathigh terbium(III) concentrations can interfere withtertiary structure formation of the ribozyme-sub-strate complex. However, if Mg2� is added beforeterbium(III) such that the ribozyme-substrate com-plex is tertiary structure folded when footprinted,the footprinting pattern is very different, demon-strating that terbium(III) does not disrupt a pre-formed tertiary structure. Under these conditions,

Figure 5. Terbium(III)-mediated footprinting of thehairpin ribozyme. (a) Footprint of (50-32P)-labeled hairpinribozyme (Rz) after incubation with terbium(III) overtwo hours at 25 �C. Lanes 1 and 2, controls; lanes 3 and4, sequencing ladders with RNase U2 (cutting after A)and RNase T1 (cutting after G); lane 5, partial alkalihydrolysis; lanes 6-19, order of addition experiments(numbers indicate order) with a 100-fold excess(500 nM) of non-cleavable substrate analog S(dAÿ1) orS(G�1A), 100 mM Na�, 12 mM Mg2�, 12 mM Co(N-H3)6
3�, and 12 mM spermidine (spd���), 12 mM Tb3�,and 120 mM EDTA as chelator; lanes 20-27, incubationof 10 mM ribozyme-substrate complex, containing traceamounts of radiolabeled ribozyme, in the presence of12 mM Mg2� with varying concentrations of Tb3�, asindicated, and with either non-cleavable substrate ana-log S(dAÿ1) or S(G�1A). (b) Superposition of backbonescission sites at 12 mM terbium(III) onto a schematic ofthe tertiary structure model of the ribozyme (Walter &Burke, 1998). Strong, medium, and weak hits, togetherwith changes caused by tertiary structure formation areindicated at the nucleotides 30 to which the scissionoccurs, as deduced from quantitative analysis of lanes 8(footprint of secondary structure) and 10 (footprint oftertiary structure) in (a). The only major hit at sub-milli-molar concentrations of terbium(III) is 30 to U37.
546 Cation Binding in the Hairpin Ribozyme
loop A, the 50-segment and parts of the 30-segmentof loop B become strongly protected from terbiu-m(III)-mediated scission, while A14 at the domainhinge and the GNRA tetraloop become moreexposed (compare lane 10 with lanes 8 and 9,Figure 5(a)). These changes are strictly dependenton tertiary structure formation, as shown by theirloss in the presence of a substrate analog witha G � 1! A mutation (S(G�1A)) (lane 11,Figure 5(a)), that inhibits docking (Walter et al.,1998, 1999; Hampel et al., 1998), or in the absenceof Mg2� (lane 12, Figure 5(a)) as an essential cofac-tor for docking (Walter et al., 1998). Chelation ofterbium(III) with EDTA prevents backbone scis-sion, as expected (lane 13, Figure 5(a)).
Similar results were obtained with cobalt(III)hexammine as the metal ion cofactor (lanes 14through 16, Figure 5(a)); only the docked ribo-zyme-substrate complex (lane 15) produces theunique footprinting/protection pattern indicativeof proper tertiary structure folding, while theundocked complex with the G�1A substrate analogdoes not (lane 16). Spermidine, despite promotinga signi®cant level of catalytic activity (with only a®ve times slower rate constant than magnesium,Figure 2), does not lead to a detectable level ofdocked complex (lanes 17 through 19, Figure 5(a)),consistent with previous ®ndings (Walter et al.,1998). Spermidine shares this behavior with mono-valent cations (Murray et al., 1998). Apparently,cleavage can occur in a transiently docked ribo-zyme-substrate complex (Pinard et al., 1999b).
Our observation, using ¯uorescence titration (seeabove), of at least three distinct af®nity classes ofterbium(III) binding sites prompted us to performfootprinting assays at different terbium(III) concen-trations, both with docked (lanes 20 through 23,Figure 5(a)) and undocked (lanes 24 through 27,Figure 5(a)) ribozyme-substrate complexes. Signi®-cantly, at 40 mM and 100 mM terbium(III), the onlysite saturated for backbone scission is at U37 in the30-segment of loop B of the ribozyme, both withthe docked (lanes 20 and 21) and undocked (lanes24 and 25) ribozyme-substrate complexes, respect-ively, suggesting a high-af®nity metal-ion bindingsite in that area. In contrast, millimolar concen-trations of terbium(III) bind to many additionalsites (lanes 22, 23, 26, and 27), generating theobserved footprinting patterns.
Figure 5(b) summarizes the footprinting infor-mation from quanti®cation of lanes 8 (before ter-tiary structure folding) and lane 10 (after tertiarystructure folding) of the gel in Figure 5(a) on aschematic of the proposed tertiary structure of thehairpin ribozyme-substrate complex (Earnshawet al., 1997; Walter & Burke, 1998). Terbium(III)strongly cuts non-Watson-Crick base-pairedregions of the ribozyme. Upon tertiary structurefolding, several regions, that coincide with regionsprotected from hydroxyl radical footprinting(Hampel et al., 1998), become partially protectedfrom terbium(III)-mediated backbone scission. Bycontrast, the backbone of A14 at the hinge between
Cation Binding in the Hairpin Ribozyme 547
domains A and B becomes more exposed, indicat-ing its role as an extended linker for proper align-ment of the two domains (Earnshaw et al., 1997).Overall, the information from footprinting at milli-molar concentrations of terbium(III) strongly sup-ports the previously proposed tertiary structuremodel of the hairpin ribozyme-substrate complex.
A high-affinity, but non-specific outer-sphereterbium(III) binding site is located in thecatalytic core of the hairpin ribozyme
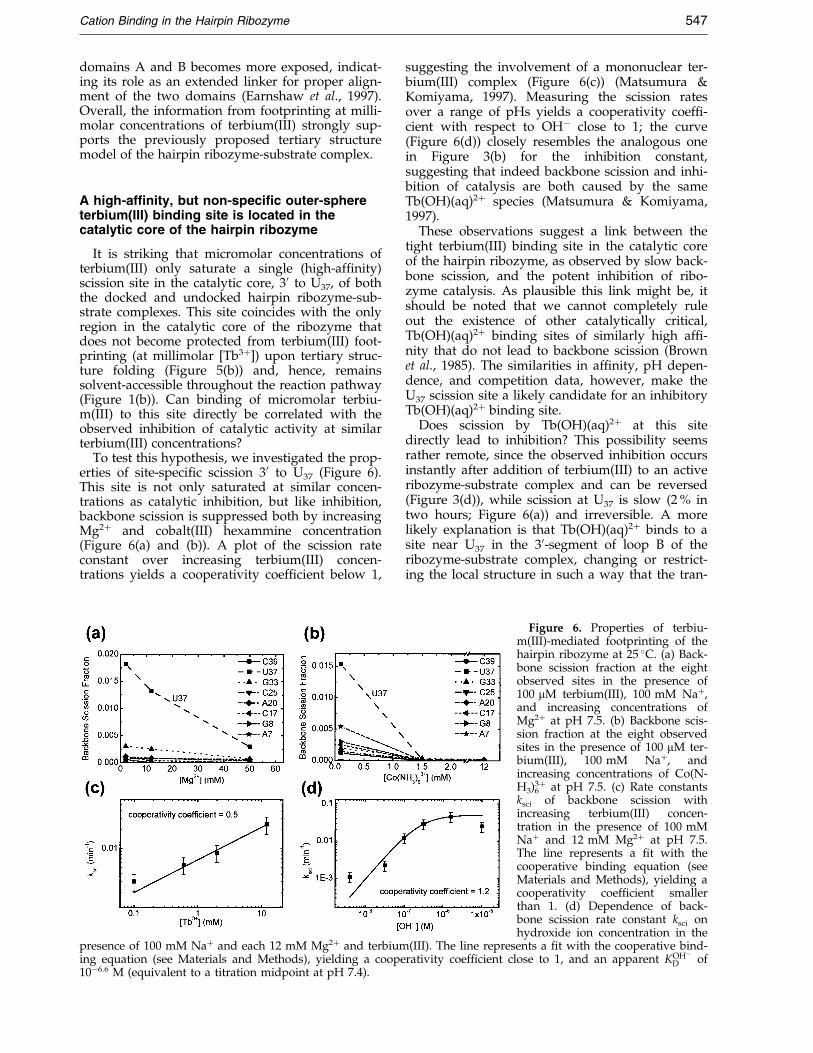
It is striking that micromolar concentrations ofterbium(III) only saturate a single (high-af®nity)scission site in the catalytic core, 30 to U37, of boththe docked and undocked hairpin ribozyme-sub-strate complexes. This site coincides with the onlyregion in the catalytic core of the ribozyme thatdoes not become protected from terbium(III) foot-printing (at millimolar [Tb3�]) upon tertiary struc-ture folding (Figure 5(b)) and, hence, remainssolvent-accessible throughout the reaction pathway(Figure 1(b)). Can binding of micromolar terbiu-m(III) to this site directly be correlated with theobserved inhibition of catalytic activity at similarterbium(III) concentrations?
To test this hypothesis, we investigated the prop-erties of site-speci®c scission 30 to U37 (Figure 6).This site is not only saturated at similar concen-trations as catalytic inhibition, but like inhibition,backbone scission is suppressed both by increasingMg2� and cobalt(III) hexammine concentration(Figure 6(a) and (b)). A plot of the scission rateconstant over increasing terbium(III) concen-trations yields a cooperativity coef®cient below 1,
presence of 100 mM Na� and each 12 mM Mg2� and terbiuming equation (see Materials and Methods), yielding a coope10ÿ6.6 M (equivalent to a titration midpoint at pH 7.4).
suggesting the involvement of a mononuclear ter-bium(III) complex (Figure 6(c)) (Matsumura &Komiyama, 1997). Measuring the scission ratesover a range of pHs yields a cooperativity coef®-cient with respect to OHÿ close to 1; the curve(Figure 6(d)) closely resembles the analogous onein Figure 3(b) for the inhibition constant,suggesting that indeed backbone scission and inhi-bition of catalysis are both caused by the sameTb(OH)(aq)2� species (Matsumura & Komiyama,1997).
These observations suggest a link between thetight terbium(III) binding site in the catalytic coreof the hairpin ribozyme, as observed by slow back-bone scission, and the potent inhibition of ribo-zyme catalysis. As plausible this link might be, itshould be noted that we cannot completely ruleout the existence of other catalytically critical,Tb(OH)(aq)2� binding sites of similarly high af®-nity that do not lead to backbone scission (Brownet al., 1985). The similarities in af®nity, pH depen-dence, and competition data, however, make theU37 scission site a likely candidate for an inhibitoryTb(OH)(aq)2� binding site.
Does scission by Tb(OH)(aq)2� at this sitedirectly lead to inhibition? This possibility seemsrather remote, since the observed inhibition occursinstantly after addition of terbium(III) to an activeribozyme-substrate complex and can be reversed(Figure 3(d)), while scission at U37 is slow (2 % intwo hours; Figure 6(a)) and irreversible. A morelikely explanation is that Tb(OH)(aq)2� binds to asite near U37 in the 30-segment of loop B of theribozyme-substrate complex, changing or restrict-ing the local structure in such a way that the tran-
Figure 6. Properties of terbiu-m(III)-mediated footprinting of thehairpin ribozyme at 25 �C. (a) Back-bone scission fraction at the eightobserved sites in the presence of100 mM terbium(III), 100 mM Na�,and increasing concentrations ofMg2� at pH 7.5. (b) Backbone scis-sion fraction at the eight observedsites in the presence of 100 mM ter-bium(III), 100 mM Na�, andincreasing concentrations of Co(N-H3)6
3� at pH 7.5. (c) Rate constantsksci of backbone scission withincreasing terbium(III) concen-tration in the presence of 100 mMNa� and 12 mM Mg2� at pH 7.5.The line represents a ®t with thecooperative binding equation (seeMaterials and Methods), yielding acooperativity coef®cient smallerthan 1. (d) Dependence of back-bone scission rate constant ksci onhydroxide ion concentration in the
(III). The line represents a ®t with the cooperative bind-rativity coef®cient close to 1, and an apparent KOHÿ
D of
548 Cation Binding in the Hairpin Ribozyme
sition state cannot be accessed and inhibition ofcatalysis is observed, as recently found for theKlenow fragment (Brautigam et al., 1999). Back-bone scission then occurs only in a small sub-popu-lation of complexes after an extended incubationtime, enabling identi®cation of the binding site.
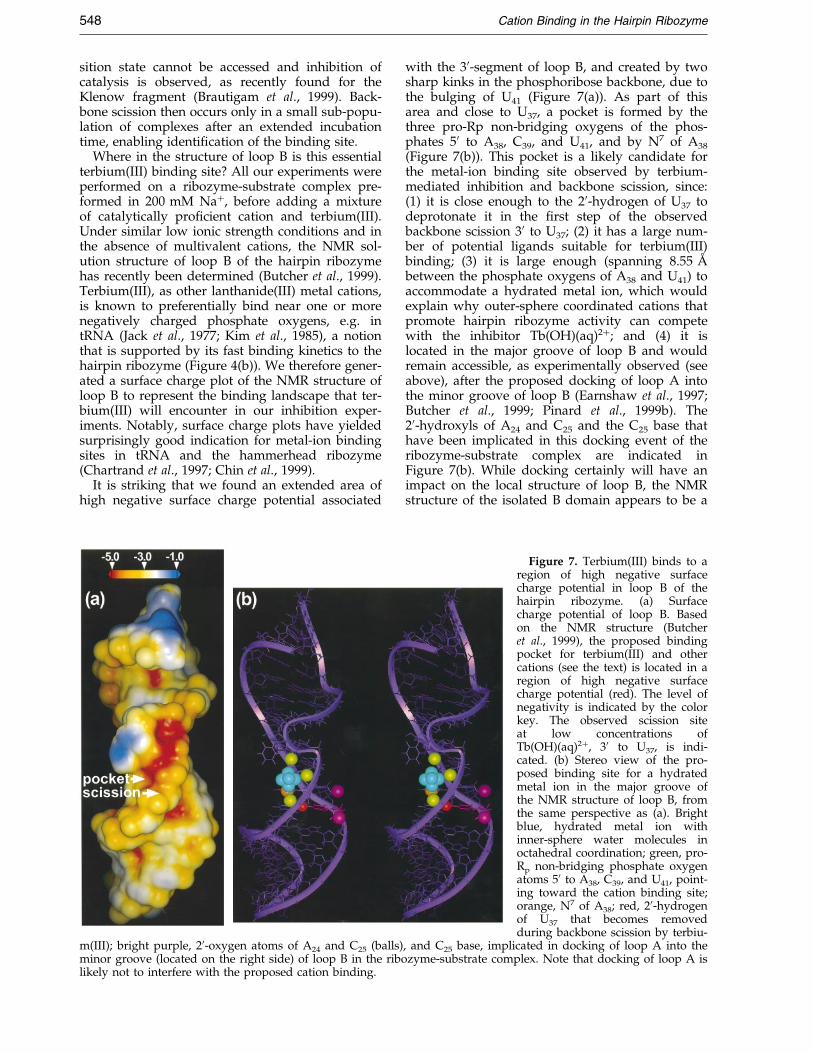
Where in the structure of loop B is this essentialterbium(III) binding site? All our experiments wereperformed on a ribozyme-substrate complex pre-formed in 200 mM Na�, before adding a mixtureof catalytically pro®cient cation and terbium(III).Under similar low ionic strength conditions and inthe absence of multivalent cations, the NMR sol-ution structure of loop B of the hairpin ribozymehas recently been determined (Butcher et al., 1999).Terbium(III), as other lanthanide(III) metal cations,is known to preferentially bind near one or morenegatively charged phosphate oxygens, e.g. intRNA (Jack et al., 1977; Kim et al., 1985), a notionthat is supported by its fast binding kinetics to thehairpin ribozyme (Figure 4(b)). We therefore gener-ated a surface charge plot of the NMR structure ofloop B to represent the binding landscape that ter-bium(III) will encounter in our inhibition exper-iments. Notably, surface charge plots have yieldedsurprisingly good indication for metal-ion bindingsites in tRNA and the hammerhead ribozyme(Chartrand et al., 1997; Chin et al., 1999).
It is striking that we found an extended area ofhigh negative surface charge potential associated
m(III); bright purple, 20-oxygen atoms of A24 and C25 (balls)minor groove (located on the right side) of loop B in the ribolikely not to interfere with the proposed cation binding.
with the 30-segment of loop B, and created by twosharp kinks in the phosphoribose backbone, due tothe bulging of U41 (Figure 7(a)). As part of thisarea and close to U37, a pocket is formed by thethree pro-Rp non-bridging oxygens of the phos-phates 50 to A38, C39, and U41, and by N7 of A38
(Figure 7(b)). This pocket is a likely candidate forthe metal-ion binding site observed by terbium-mediated inhibition and backbone scission, since:(1) it is close enough to the 20-hydrogen of U37 todeprotonate it in the ®rst step of the observedbackbone scission 30 to U37; (2) it has a large num-ber of potential ligands suitable for terbium(III)binding; (3) it is large enough (spanning 8.55 AÊ
between the phosphate oxygens of A38 and U41) toaccommodate a hydrated metal ion, which wouldexplain why outer-sphere coordinated cations thatpromote hairpin ribozyme activity can competewith the inhibitor Tb(OH)(aq)2�; and (4) it islocated in the major groove of loop B and wouldremain accessible, as experimentally observed (seeabove), after the proposed docking of loop A intothe minor groove of loop B (Earnshaw et al., 1997;Butcher et al., 1999; Pinard et al., 1999b). The20-hydroxyls of A24 and C25 and the C25 base thathave been implicated in this docking event of theribozyme-substrate complex are indicated inFigure 7(b). While docking certainly will have animpact on the local structure of loop B, the NMRstructure of the isolated B domain appears to be a
Figure 7. Terbium(III) binds to aregion of high negative surfacecharge potential in loop B of thehairpin ribozyme. (a) Surfacecharge potential of loop B. Basedon the NMR structure (Butcheret al., 1999), the proposed bindingpocket for terbium(III) and othercations (see the text) is located in aregion of high negative surfacecharge potential (red). The level ofnegativity is indicated by the colorkey. The observed scission siteat low concentrations ofTb(OH)(aq)2�, 30 to U37, is indi-cated. (b) Stereo view of the pro-posed binding site for a hydratedmetal ion in the major groove ofthe NMR structure of loop B, fromthe same perspective as (a). Brightblue, hydrated metal ion withinner-sphere water molecules inoctahedral coordination; green, pro-Rp non-bridging phosphate oxygenatoms 50 to A38, C39, and U41, point-ing toward the cation binding site;orange, N7 of A38; red, 20-hydrogenof U37 that becomes removedduring backbone scission by terbiu-
, and C25 base, implicated in docking of loop A into thezyme-substrate complex. Note that docking of loop A is

Cation Binding in the Hairpin Ribozyme 549
good starting point for discussing some of its struc-tural properties such as potential metal ion bindingpockets (Butcher et al., 1999).
To test whether the proposed metal ion bindingpocket is indeed an outer-sphere coordination site,we asked whether any of the potential RNAligands close to the observed Tb(OH)(aq)2� bindingsite are involved in direct coordination of either aMg2� or a terbium(III) metal ion. To this end, weindividually modi®ed the phosphates 50 to U37,A38, C39, A40, and U41 as phosphorothioates (RP
and Sp stereoisomer mixtures) and the N7 positionsof A38 and A40 as carbons. These modi®cationssubstitute soft sulfur for hard oxygen ligands andeliminate soft nitrogen ligands, respectively, sothat changes in the relative binding af®nities ofdirectly coordinated hard Mg2� versus soft terbiu-m(III) metal ions would be expected (Feig &Uhlenbeck, 1999). However, we did not observechanges in the terbium(III) inhibition constants inthe presence of 12 mM Mg2� for any of thesemodi®cations (data not shown), consistent with amodel in which a terbium(III) bound in the pro-posed binding pocket is not directly coordinated tothe RNA. The observed terbium(III) binding site,therefore, appears to be a catalytically essential,but rather non-selective outer-sphere cation bind-ing site, offering a plausible explanation for thelack in cation cofactor selectivity of the hairpinribozyme.
Discussion
Tb(OH)(aq)2� interferes with a late step inhairpin ribozyme catalysis
The hairpin ribozyme can cleave its substrateRNA under a variety of cationic conditions thatinclude monovalent cations, soft and hard divalentcations, kinetically non-exchangeable complexes oftrivalent metal ions, e.g. cobalt(III) hexammine,and organic polyamines (Walter & Burke, 1998;Murray et al., 1998; Earnshaw & Gait, 1998; Nesbittet al., 1999). In fact, only a few cations appear to beincompatible with catalytic activity, as long as theyare used in an appropriate concentration range.We now have characterized such an inhibitoryaqueous metal ion complex, Tb(OH)(aq)2�. Inhi-bition appears to be sensitive to the overall chargeof the terbium(III) complex, as protonation reversesinhibition. At pH values signi®cantly below thepKa of terbium(III) (7.9), a bell-shaped dependenceof hairpin ribozyme activity on the total lanthani-de(III) ion concentration is observed. An analogouseffect is seen in the presence of lanthanum(III) asthe inhibitory cation (pKa(La3�) � 8.5). These metalion dependencies resemble that of the hammer-head ribozyme on lanthanum(III) at a pH of 7.0,which was interpreted as evidence for a two-metalion mechanism (Lott et al., 1998). It is important tokeep in mind that not only two distinct metal-ionbinding sites, but also the presence of two differentaqueous lanthanide(III) ion complexes in solution,
such as Ln(aq)3� and Ln(OH)(aq)2�, can lead to abell-shaped activation curve, as shown here for thehairpin ribozyme.
Higher (millimolar) concentrations of terbiu-m(III) interfere with both formation and docking ofthe ribozyme-substrate complex, as shown by ter-bium(III)-mediated backbone scission (Figure 5(a)).However, we were also able to demonstrate, by¯uorescence resonance energy transfer (Figure 4(e))and hydroxyl radical footprinting, that lower ter-bium(III) concentrations (micromolar) do not inter-fere with domain docking of the complex, yet stillinhibit cleavage activity. Hence, inhibition atmicromolar terbium(III) concentration can beattributed to blockage of a post-docking step in thereaction pathway (Figure 1(b)).
How does Tb(OH)(aq)2� inhibit hairpin ribo-zyme activity? Inhibition occurs instantly and isreversible, as was shown previously for the ham-merhead ribozyme (Feig et al., 1998), ruling outslow and irreversible chemical damage to the RNAas mode of interference. The rate-limiting step ofhairpin ribozyme-catalyzed cleavage occurs afterglobal docking of the ribozyme-substrate complex(Walter et al., 1998; Hampel et al., 1998) and is pH-independent (Nesbitt et al., 1997), consistent withthe notion that a localized conformational changemight be the slowest step in the cleavage reactionpathway. In impairing cleavage, a proximalTb(OH)(aq)2� might alter the geometry of thesugar-phosphate backbone and the base-stackinginteractions required for this conformationalchange, or a tightly bound Tb(OH)(aq)2� mightconstrain the ¯exibility of the complex(Gersanovski et al., 1985; Tajmir-Riahi et al., 1993;Brautigam et al., 1999), to a degree that is incompa-tible with catalytic activity. The possibility thatterbium interferes with the as yet unde®nedrate-limiting step in hairpin ribozyme catalysismakes it a useful probe to further characterize thereaction mechanism of this and potentially otherRNA catalysts.
Terbium(III) is a versatile probe of RNAstructure, function, and metal-ion binding
We found that backbone scission with 12 mMterbium(III) at pH 7.5, after a two-hour incubationat room temperature, produces a distinct footprint-ing pattern of hairpin ribozyme secondary and ter-tiary structure, analogous to hydroxyl radicalfootprinting (Hampel et al., 1998). Terbium(III) con-centrations down to 2 mM, any pH above 7.0, orshorter incubation times (530 minutes) yield simi-lar patterns (data not shown). Unlike hydroxyl rad-icals, terbium(III) ions preferentially cut non-Watson-Crick base-paired regions, making them apowerful and easy-to-use probe for the solventaccessibility of structural elements in RNA thattypically are involved in long-range tertiary con-tacts or protein binding. Interestingly, terbium(III)at these high concentrations can interfere with theformation of both secondary and tertiary structure,

550 Cation Binding in the Hairpin Ribozyme
but it does not disrupt pre-formed structures, sothat RNA folding pathways can be revealed byorder-of-addition experiments. These applicationsexpand the previously described use of lanthani-de(III)-mediated backbone cleavage in detection ofspeci®c metal-ion binding sites in RNA (Ciesiolkaet al., 1989; Evans, 1990; Gast et al., 1996; Feig &Uhlenbeck, 1999), or in RNA degradation for bio-technological applications (Stein & Cohen, 1988;Morrow, 1996; Matsumura & Komiyama, 1997).Our results are also distinct from backbone scissionby other metal ions such as lead, that typically dis-play a much narrower range of scission sites(Ciesiolka et al., 1989).
Why does terbium(III) scission preferentiallycleave the backbone of non-Watson-Crick base-paired regions in RNA? The interaction of terbiu-m(III) ions with nucleic acids occurs both with thenegatively charged phosphate oxygens of the back-bone and the electron donor groups on the nucleo-side bases, in particular the N7 position of purines(Gross & Simpkins, 1981; Gersanovski et al., 1985;Tajmir-Riahi et al., 1993; Feig et al., 1998) and poss-ibly the base �-electron systems (McFail-Isom et al.,1998). It is well conceivable that the better accessi-bility of these groups in the widened major grooveof non-Watson-Crick base-paired regions in RNAstabilizes binding of terbium to these segmentsand, as a result, leads to a higher probability ofbackbone scission. The af®nity of the hairpin ribo-zyme for the most tightly binding terbium(III) ionsis about three orders of magnitude higher than theone for magnesium(II), as observed in our compe-tition experiments. Hence, the high binding freeenergy of terbium(III) may stabilize unstacked con-formations of weakly (non-Watson-Crick) base-paired nucleoside bases, which might explain theobserved inhibition of ribozyme activity (Feig et al.,1998; and results presented herein).
Terbium(III), like other lanthanide metal ions,particularly europium(III), permits ¯uorescenceanalysis of its binding sites on nucleic acids byenergy transfer from guanine bases (Yonushot et al.,1978; Topal & Fresco, 1980; Wenzel & Collette,1988). We have used this effect to distinguishclasses of binding sites with different af®nities aswell as distinct processes in the binding kinetics ofterbium(III) to the hairpin ribozyme. In addition,using more sophisticated spectroscopic equipment,the detailed ¯uorescence properties of nucleic acid-bound terbium(III) or europium(III) ions can yieldinformation on the number and nature of theirinner-sphere ligands (Horrocks, 1993; Feig et al.,1999). Taken together, these properties makelanthanide(III) ions versatile probes for the analysisof structure, function, and metal-ion binding sitesin RNA.
Terbium(III) reveals an outer-sphere metal-ionbinding site
RNA backbone scission at micromolar terbiu-m(III) concentrations was used to identify a high-
af®nity terbium(III) binding site of the hairpin ribo-zyme in an area of loop B with high negative sur-face charge potential. Both terbium(III) binding tothis site and inhibition of catalytic activity sharesimilar af®nities and can be competed by any cat-ion known to promote hairpin ribozyme activity.The involved functionally important binding pock-et(s), therefore, must be able to accommodate awide variety of cations, including Mg2�, Mn2�,Co(NH3)6
3�, spd���, and Na�. Although we cannotcompletely rule out inner-sphere coordination ofsome of these cations, the observations that cobal-t(III) hexammine and polyamines bind despitetheir inability to coordinate RNA ligands and thatTb3� binding is not in¯uenced by RNA modi®-cations designed to interfere with directly coordi-nated metal ions suggest that inner-spherecoordination is not a requirement for binding tothis site. Thus, the proposed binding pocket in themajor groove of loop B appears to be a member ofthe third class of the metal-ion binding sites asdescribed by Laing et al. (1994); binding occurssite-speci®cally, but non-selectively in an area ofRNA with unusually high charge density. Becauseof its variability and solvent accessibility, this typeof cation-binding site can be expected to be dif®-cult to identify by structure determination methodssuch as NMR and X-ray crystallography.
A cation site-speci®cally bound in an area ofhigh negative surface charge potential can beexpected to substantially contribute to the stabilityof RNA tertiary structure contacts close to thisarea. In the case of the hairpin ribozyme, the pro-posed cation binding pocket maps proximal to theproposed tertiary contacts between loops A and B(Figure 7(b)) (Earnshaw et al., 1997; Butcher et al.,1999; Pinard et al., 1999b) and stays occupiedbefore and after tertiary structure folding. Ourcompetition data suggest that it is indeed the sitetypically detected by metal-ion titration of cleavageactivity (Nesbitt et al., 1997, 1999; Walter et al.,1998; Thomson & Lilley, 1999). By contrast, manyadditional metal ions are more weakly bound tothe loop regions of the undocked conformationand become displaced upon tertiary structure for-mation, as observed by terbium(III)-mediated foot-printing. Hence, despite its lack in inner-spherecontacts, the proposed cation binding pocketshares some features with the classic ``speci®c''metal-ion binding sites that are observed in the ter-tiary structure of large RNA catalysts (Celander &Cech, 1991; Smith et al., 1992; Cate & Doudna,1996; Cate et al., 1996, 1997; Basu et al., 1998).
The unusually high negative charge density ofthe proposed cation binding pocket is created bytwo subsequent kinks in the backbone of the hair-pin ribozyme, due to two extra nucleotides in the30-segment of loop B (Figure 7) (Butcher et al.,1999). Interestingly, the associated motif has beenfound to also organize the structure of multi-helixloops in 16 S and 23 S ribosomal RNA (Leontis &Westhof, 1998), two of the most ancient RNAs,extensively used for phylogenetic classi®cation of

Cation Binding in the Hairpin Ribozyme 551
the ``tree of life'' (Doolittle, 1999). Other structuralelements in RNA that are able to bind outer-spherecoordinated metal ions in their major groove areG:U wobble or G:A sheared base-pairs, or GNRAtetraloops (Cate & Doudna, 1996; Kieft & Tinoco,1997; Suga et al., 1998; Wedekind & McKay, 1998;Feig & Uhlenbeck, 1999). These elements also havebeen identi®ed as building blocks for the morecomplex binding sites of speci®c metal ions in lar-ger RNAs (Cate et al., 1996, 1997). It is thereforetempting to propose that outer-sphere metal-ionbinding sites emerged ®rst in the primitive ribo-zymes of the early RNA world, ultimately becom-ing buried in the complex tertiary structures of thesubsequently evolving larger RNA catalysts. Nota-bly, even in these structures metal ions remain sig-ni®cantly hydrated (Feig & Uhlenbeck, 1999). Wenow have identi®ed such an outer-sphere metal-ion binding element in the hairpin ribozyme andhave related it to the function of this RNA catalyst.
Materials and Methods
Synthesis and purification of oligonucleotides
RNA oligonucleotides were synthesized by standardmethods using solid-phase phosphoramidite chemistryfrom Glen Research implemented on an Applied Biosys-tems 392 DNA/RNA synthesizer (1 mmol scale). 30-Fluor-escein and 50-hexachloro¯uorescein for FRET analysiswere attached during synthesis as described (Walter et al.,1998), oxidation with S8 was used to generate a stereoi-somer (RP and SP) mixture of site-speci®c phosphorothio-ates, and the N7-deaza-ribo adenosine phosphoramiditewas purchased from ChemGenes. Synthetic RNA wasdeprotected following the protocol of Wincott et al.(1995), utilizing a 3:1 mixture of concentrated aqueousammonia and ethanol to remove the exocyclic amineprotection groups and triethylamine trihydro¯uoride toremove the 20 OH silyl protection groups. Fully depro-tected, full-length RNA was isolated by denaturing, 8 Murea, 20 % (w/v) polyacrylamide gel electrophoresis, dif-fusion elution from the gel slices, ethanol precipitation,and subsequent C8-reversed phase HPLC with a gradientof 0-60 % acetonitrile in 0.1 M triethyl ammonium acetate(50 minutes, 1 ml/minute). Non-cleavable, chemicallyblocked substrate analog was obtained by introducing adeoxy-A (dA) modi®cation at the cleavage site (positionÿ1).
The ribozyme was transcribed from two synthetic,fully complementary and annealed DNA strands, basi-cally as described (Milligan & Uhlenbeck, 1989), exceptthat higher NTP concentrations (7.5 mM each) togetherwith inorganic pyrophosphatase were used to increaseyields (Cunningham & Ofengand, 1990). The full-lengthrun-off transcript was isolated after denaturing, 8 Murea, 10 % (w/v) polyacrylamide gel electrophoresis byUV shadowing, diffusion elution of the gel slices, andethanol precipitation.
Handling of cation stock solutions
For all salts, the highest available purity from Sigmaor Aldrich was used, with chloride as the common anio-nic component. Stock solutions of Tb3�, Mn2�, andCo(NH3)6
3� were kept in small aliquots at ÿ20 �C in the
dark to avoid oxidation or precipitation. TbCl3 was dis-solved to 1 mM, 10 mM, or 100 mM in a 5 mM sodiumcacodylate buffer at pH 5.5 to prevent formation of ter-bium(III) hydroxide precipitates at higher pH. Spermi-dine and spermine stock solutions (1 M and 100 mM,respectively) were brought to pH 7.5 by titrating the dis-solved hydrochloride salts with NaOH.
Radioactive cleavage reactions
(50-32P)-labeled substrate was prepared by phosphoryl-ation with T4 polynucleotide kinase and [g-32P]ATP, andwas puri®ed using CentriSep spin columns (PrincetonSeparations). Trace amounts (<1 nM) of radiolabeledsubstrate and 200 nM transcribed hairpin ribozyme(Figure 1(a)) were annealed in the absence of multivalentcations (in 100 mM Tris-HCl (pH 7.5), 200 mM NaCl;note that the subsequent addition of an equal volume ofcation stock solution (see below) lowered these concen-trations to 50 mM and 100 mM, respectively) at 25 �C for15 minutes. To obtain other pH values, the followingbuffers were used instead of Tris-HCl; MES-NaOH forpH 5.6 and 6.5; Pipes-NaOH for pH 7.0; TAPS-NaOH,for pH 8.2 and 9.0. To initiate cleavage, an equal volumeof cation stock solution (containing only the cationsrequired to give the ®nal concentrations as described inResults) at 25 �C was added. Typically ten 5-ml aliquotsof the reaction were taken over a two-hour period andstopped by mixing with at least an equal volume of ice-chilled loading buffer (80 % (v/v) formamide, 0.025 %(w/v) xylene cyanol, 0.025 % (w/v) bromophenol blue,50 mM EDTA). The 50 cleavage product was separatedfrom uncleaved substrate by denaturing, 8 M urea, 20 %polyacrylamide gel electrophoresis, quanti®ed, and nor-malized to the sum of the substrate and product bandsusing a Bio-Rad Molecular Imager System GS-525. Thetime trace of product formation was ®tted to the expo-nential growth equation y � y0 � A1(1 ÿ eÿt/t1) �A2(1 ÿ eÿt/t2), as described by Esteban et al. (1997). Thedata sets in the presence of terbium(III) were ®tted satis-factorily with single-exponentials (A1 6� 0, A2 � 0), whileother data sets, notably those in the presence of Mg2�
and Co(NH3)63�, required double-exponentials (A1,
A2 6� 0), as described by Esteban et al. (1997); in addition,the presence of terbium(III) often lowered the amplitudesrather than the rate constants. Therefore, a rate constantk was calculated from k � A1/t1 � A2/t2 to account forthe in¯uence of terbium(III) on cleavage rate and fractionof the two phases of hairpin ribozyme cleavage. Ratiosof the obtained rate constants were plotted over differentcation concentrations to analyze the data using the coop-erative-binding equation:
y � ymax�Mn��p
�Mn��p � KpD
yielding an apparent dissociation constant KDM for cation
Mn� (or the corresponding apparent inhibition constantKi for Tb3�) and a cooperativity or Hill coef®cient p. All®ts were calculated using MicrocalTM OriginTM 4.1 soft-ware employing Marquardt-Levenberg non-linear least-squares regression.
Steady-state fluorescence measurements
Steady-state ¯uorescence intensities of a doubly ¯uor-ophore-labeled two-piece hairpin ribozyme wererecorded on an Aminco-Bowman Series 2 (AB2) spectro-

552 Cation Binding in the Hairpin Ribozyme
¯uorometer from SLM (Rochester, NY) in a cuvette with3-mm excitation and emission path lengths (150 ml totalvolume), essentially as described (Walter et al., 1998).200 nM ribozyme-substrate complex were constituted in50 mM Tris-HCl (pH 7.5), 100 mM NaCl, 25 mM DTT(®nal concentration), by addition of each 500 nM non-cleavable (dAÿ1) substrate analog and unlabeled ribo-zyme 30-segment to 200 nM of the 50-hexachloro¯uores-cein and 30-¯uorescein doubly-labeled ribozyme 50-segment, heating to 70 �C for two minutes, and coolingto room temperature over ®ve minutes. The obtainedcomplex was pre-incubated at 25 �C for 15 minutes,before manually mixing 145 ml complex solution with5 ml cation stock solution in the ¯uorometer cuvette togive ®nal concentrations of 12 mM MgCl2 and up to100 mM Tb3�. Tertiary structure formation wasmonitored as steady-state ¯uorescence changes ofthe two ¯uorophores over time. Fluorescein was excitedat 485 nm (4 nm slit width), and ¯uorescence emissionwas monitored both at 515 nm and 545 nm (8 nm slitwidth) by shifting the emission monochromator backand forth, using the AB2 software package, and a ratioQ � F545/F515 re¯ecting the FRET ef®ciency was calcu-lated. Q was normalized with its value Q0 immediatelyafter formation of the substrate-ribozyme complex (i.e.(Q ÿ Q0)/Q0 was calculated). The resulting growthcurves were ®tted to the single-exponential functiony � y0 � A(1 ÿ eÿt/t) to yield the docking rate constantkdock � 1/t and the docking amplitude A.
Steady-state ¯uorescence spectra of Tb3� bound to1 mM of the pre-formed complex of ribozyme(Figure 1(a)) with non-cleavable (dA-1) substrate analogwere measured in standard reaction buffer (50 mM Tris-HCl (pH 7.5), 12 mM MgCl2) at 25 �C, by slowly titratingTbCl3 over several orders of magnitude from appropriatestock solutions. Excitation was at 290 nm (slit width4 nm), steady-state emission was scanned with a slitwidth of 8 nm. To extract the ¯uorescence intensity ofthe major peak at 545 nm, each peak was ®tted between530 nm and 555 nm with the Gaussian distributionfunction:
y � y0 � A
w��������p=2p eÿ2
�xÿx0 �2w2
to yield the peak height as the pre-exponential factor,from which the background value in the absence ofTb3� was subtracted. These ¯uorescence signals wereplotted over varying Tb3� concentration and were®tted with Hill equations as described above. For a®t over the complete Tb3� titration range, a sum ofthree independent Hill equations produced the bestresult.
Stopped-¯ow kinetic experiments were performedusing the Milli-Flow Reactor from SLM (Rochester, NY)on the AB2 spectro¯uorometer. A stock solution of4 mM Tb3� in standard reaction buffer (50 mM Tris-HCl(pH 7.5), 12 mM MgCl2) was mixed with an equalvolume of the same buffer, with or without 2 mM pre-formed complex of ribozyme (Figure 1(a)) with non-clea-vable (dAÿ1) substrate analog at 25 �C. Excitation was at290 nm (slit width 8 nm), emission was at 545 nm (slitwidth 8 nm), and changes in steady-state ¯uorescenceintensity were recorded for ten seconds in 1 ms timeintervals. In the presence of the ribozyme-substrate com-plex, a ¯uorescence increase was observed that couldonly be ®tted satisfactorily with a growth equation withat least three exponentials, y � y0 � A1(1 ÿ eÿt/t1) �
A2(1 ÿ eÿt/t2) � A3(1 ÿ eÿt/t3). Rate constants werecalculated from k � 1/t.
Terbium(III)-mediated footprinting
To observe the slow backbone scission reactionmediated by Tb(OH)(aq)2�, transcribed ribozyme(Figure 1(a)) was dephosphorylated with an excess ofcalf intestine alkaline phosphatase for one hour at 37 �C,phenol/chloroform extracted, recovered by ethanol pre-cipitation, (50-32P)-phosphorylated using T4 polynucleo-tide kinase and [g-32P]ATP, desalted using a CentriSepspin column (Princeton Separations), and puri®ed bydenaturing, 8 M urea, 10 % polyacrylamide gel electro-phoresis, followed by diffusion elution into 5 mMEDTA, ethanol precipitation in the presence of 1 mMATP as carrier, and desalting using a CentriSep spin col-umn. The labeled ribozyme (typically �500,000 dpm per10-ml reaction, or �5 nM) was incubated with varyingadditives in the presence of typically 12 mM TbCl3 fortwo hours at 25 �C. To fold the tertiary structure of theribozyme-substrate complex, Mg2� or cobalt(III) hexam-mine had to be added ®ve minutes prior to terbium(III).The scission reaction was stopped by addition of anequal volume of 80 % formamide, 50 mM EDTA, andanalyzed on an 8 M urea, 15 % wedged polyacrylamidesequencing gel, alongside sequencing ladders from par-tial digestion with ribonucleases T1 and U2, and alkalihydrolysis (Pinard et al., 1999a). For time courses, typi-cally eight aliquots were taken over three to six hours.Product bands were either visualized using autoradiog-raphy or quanti®ed and normalized to the sum of thesubstrate and product bands (after subtraction ofbackground degradation in the absence of terbium(III))using a Bio-Rad Molecular Imager System GS-525;scission rate constants ksci were calculated from a ®ty � y0 � A(1 ÿ eÿt/t) to the combined scission fractionsat all cut positions, with ksci � 1/t.
Molecular modeling
The NMR structure of loop B of the hairpin ribozymeas deposited in the RCSB Protein Data Bank, accessionnumber 1B36 (Butcher et al., 1999) with a boundhydrated metal ion was visualized using the programInsight II (MSI/Biosym). Electrostatic potentials werecalculated using the ®nite difference non-linear Poisson-Boltzmann equation implemented in the program Delphi(Molecular Simulations Inc., San Diego 1995) andmapped, using Insight II, onto the molecular surface ofloop B, created by rolling a spherical probe correspond-ing to the radius of a water molecule (1.4 AÊ ) around theexterior. The Amber-like partial charges for RNA weretaken from Tung et al. (1984).
Acknowledgments
We are grateful to David Pecchia for RNA synthesis,to Krishnan Venkataraman, Ken Hampel, and ErikaAlbinson for contribution of experimental data and help-ful discussions, and to Jeffrey Bond and his MolecularModeling Facility of the Vermont Cancer Center for helpwith Figure 7.

Cation Binding in the Hairpin Ribozyme 553
References
Baes, C. F. & Mesmer, R. E. (1976). The Hydrolysis of Cat-ions, pp. 129-146, Wiley-Interscience, New York.
Basu, S., Rambo, R. P., Strauss-Soukup, J., Cate, J. H.,FerreÂ-D'AmareÂ, A. R., Strobel, S. A. & Doudna, J. A.(1998). A speci®c monovalent metal ion integral tothe AA platform of the RNA tetraloop receptor.Nature Struct. Biol. 5, 986-992.
Been, M. D. & Wickham, G. S. (1997). Self-cleaving ribo-zymes of hepatitis delta virus RNA. Eur. J. Biochem.247, 741-753.
Brautigam, C. A., Aschheim, K. & Steitz, T. A. (1999).Structural elucidation of the binding and inhibitoryproperties of lanthanide (III) ions at the 30-50 exonu-cleolytic active site of the Klenow fragment. Chem.Biol. 6, 901-908.
Brown, R. S., Dewan, J. C. & Klug, A. (1985). Crystallo-graphic and biochemical investigation of the lea-d(II)-catalyzed hydrolysis of yeast phenylalaninetRNA. Biochemistry, 24, 4785-4801.
Butcher, S. E., Allain, F. H. T. & Feigon, J. (1999). Sol-ution structure of the loop B domain from the hair-pin ribozyme. Nature Struct. Biol. 6, 212-216.
Buzayan, J. M., Gerlach, W. L. & Bruening, G. (1986).Non-enzymatic cleavage and ligation of RNAscomplementary to a plant virus satellite RNA.Nature, 323, 349-353.
Cai, Z. & Tinoco, I. (1996). Solution structure of loop Afrom the hairpin ribozyme from tobacco ringspotvirus satellite. Biochemistry, 35, 6026-6036.
Cate, J. H. & Doudna, J. A. (1996). Metal-binding sites inthe major groove of a large ribozyme domain.Structure, 4, 1221-1229.
Cate, J. H., Gooding, A. R., Podell, E., Zhou, K., Golden,B. L., Kundrot, C. E., Cech, T. R. & Doudna, J. A.(1996). Crystal structure of a group I ribozymedomain: principles of RNA packing. Science, 273,1678-1685.
Cate, J. H., Hanna, R. L. & Doudna, J. A. (1997). A mag-nesium ion core at the heart of a ribozyme domain.Nature Struct. Biol. 4, 553-558.
Cech, T. (1993). Structure and Mechanism of the largecatalytic RNAs: group I and group II introns andribonuclease P. In RNA World (Gesteland, R. F. &Atkins, J. F., eds), pp. 239-269, Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
Celander, D. W. & Cech, T. R. (1991). Visualizing thehigher order folding of a catalytic RNA molecule.Science, 251, 401-407.
Chartrand, P., Leclerc, F. & Cedergren, R. (1997). Relat-ing conformation, Mg2� binding, and functionalgroup modi®cation in the hammerhead ribozyme.RNA, 3, 692-696.
Chin, K., Sharp, K. A., Honig, B. & Pyle, A. M. (1999).Calculating the electrostatic properties of RNA pro-vides new insights into molecular interactions andfunction. Nature Struct. Biol. 6, 1055-1061.
Ciesiolka, J., Marciniec, T. & Krzyzosiak, W. (1989).Probing the environment of lanthanide binding sitesin yeast tRNAPhe by speci®c metal-ion-promotedcleavages. Eur. J. Biochem. 182, 445-450.
Cunningham, P. R. & Ofengand, J. (1990). Use of inor-ganic pyrophosphatase to improve the yield ofin vitro transcription reactions catalyzed by T7 RNApolymerase. Biotechniques, 9, 713-714.
Dickeson, S. K., Bhattacharyya-Pakrasi, M., Mathis, N. L.,Schlesinger, P. H. & Santoro, S. A. (1998). Ligandbinding results in divalent cation displacement
from the a2b1 integrin I domain: evidence from ter-bium luminescence spectroscopy. Biochemistry, 37,11280-11288.
Doolittle, W. F. (1999). Phylogenetic classi®cation andthe universal tree. Science, 284, 2124-2129.
Draper, D. E. (1985). On the coordination properties ofEu3� bound to tRNA. Biophys. Chem. 21, 91-101.
Earnshaw, D. J. & Gait, M. J. (1997). Progress towardthe structure and therapeutic use of the hairpinribozyme. Antisense Nucleic Drug Dev. 7, 403-411.
Earnshaw, D. J. & Gait, M. J. (1998). Hairpin ribozymecleavage catalyzed by aminoglycoside antibioticsand the polyamine spermine in the absence ofmetal ions. Nucl. Acids Res. 26, 5551-5561.
Earnshaw, D. J., Masquida, B., MuÈ ller, S., Sigurdsson,S. T., Eckstein, F., Westhof, E. & Gait, M. J. (1997).Inter-domain cross-linking and molecular modellingof the hairpin ribozyme. J. Mol. Biol. 247, 1-16.
Esteban, J. A., Banerjee, A. R. & Burke, J. M. (1997). Kin-etic mechanism of the hairpin ribozyme. Identi®-cation and characterization of two nonexchangeableconformations. J. Biol. Chem. 272, 13629-13639.
Evans, C. H. (1990). The Biochemistry of Lanthanides, pp.85-204, Plenum Press, London.
Faulhammer, D. & Famulok, M. (1997). Characterizationand divalent metal-ion dependence of in vitroselected deoxyribozymes which cleave DNA/RNAchimeric oligonucleotides. J. Mol. Biol. 269, 188-202.
Feig, A. L. & Uhlenbeck, O. C. (1999). The role of metalions in RNA biochemistry. In RNA World(Gesteland, R. F., Cech, T. R. & Atkins, J. F., eds),2nd edit., pp. 287-319, Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
Feig, A. L., Scott, W. G. & Uhlenbeck, O. C. (1998).Inhibition of the hammerhead ribozyme cleavagereaction by site-speci®c binding of Tb. Science, 279,81-84.
Feig, A. L., Panek, M., Horrocks, W. D. & Uhlenbeck,O. C. (1999). Probing the binding of Tb(III) andEu(III) to the hammerhead ribozyme using lumines-cence spectroscopy. Chem. Biol. 6, 801-810.
Frey, M. W., Frey, S. T., Horrocks, W. D., Kaboord, B. F.& Benkovic, S. J. (1996). Elucidation of the metal-binding properties of the Klenow fragment ofEscherichia coli polymerase I and bacteriophage T4DNA polymerase by lanthanide(III) luminescencespectroscopy. Chem. Biol. 3, 393-403.
Gast, F. U., Kempe, D., Spieker, R. L. & Sanger, H. L.(1996). Secondary structure probing of potato spin-dle tuber viroid (PSTVd) and sequence comparisonwith other small pathogenic RNA replicons pro-vides evidence for central non-canonical base-pairs,large A-rich loops, and a terminal branch. J. Mol.Biol. 262, 652-670.
Gersanovski, D., Colson, P., Houssier, C. & Fredericq, E.(1985). Terbium(3�) as a probe of nucleic acidsstructure. Does it alter the DNA conformation insolution?. Biochim. Biophys. Acta, 824, 313-323.
Geyer, C. R. & Sen, D. (1997). Evidence for the metal-cofactor independence of an RNA phosphodiester-cleaving DNA enzyme. Chem. Biol. 4, 579-593.
Gross, D. S. & Simpkins, H. J. (1981). Evidence for two-site binding in the terbium(III)-nucleic acid inter-action. J. Biol. Chem. 256, 9593-9598.
Hampel, A. & Cowan, J. A. (1997). A unique mechanismfor RNA catalysis: the role of metal cofactors inhairpin ribozyme cleavage. Chem. Biol. 4, 513-517.

554 Cation Binding in the Hairpin Ribozyme
Hampel, A. & Tritz, R. (1989). RNA catalytic propertiesof the minimum (ÿ)sTRSV sequence. Biochemistry,28, 4929-4933.
Hampel, K. J., Walter, N. G. & Burke, J. M. (1998). Thesolvent-protected core of the hairpin ribozyme-sub-strate complex. Biochemistry, 37, 14672-14682.
Horrocks, W. D. (1993). Luminescence spectroscopy.Methods Enzymol. 226, 495-538.
Horrocks, W. D., Schmidt, G. F., Sudnick, D. R., Kittrell,C. & Bernheim, R. A. (1977). Laser-induced lantha-nide ion luminescence lifetime measurements bydirect excitation of metal ion levels. A new class ofstructural probe for calcium-binding proteins andnucleic acids. J. Am. Chem. Soc. 99, 2378-2380.
Jack, A., Ladner, J. E., Rhodes, D., Brown, R. S. & Klug,A. (1977). A crystallographic study of metal-bindingto yeast phenylalanine transfer RNA. J. Mol. Biol.111, 315-328.
Jayasena, V. K. & Gold, L. (1997). In vitro selection ofself-cleaving RNAs with a low pH optimum. Proc.Natl Acad. Sci. USA, 94, 10612-10617.
Kieft, J. S. & Tinoco, I. (1997). Solution structure of ametal-binding site in the major groove of RNAcomplexed with cobalt(III) hexammine. Structure, 5,713-721.
Kim, S. H., Shin, W. C. & Warrant, R. W. (1985). Heavymetal ion-nucleic acid interaction. Methods Enzymol.114, 156-167.
Laing, L. G., Gluick, T. C. & Draper, D. E. (1994). Stabil-ization of RNA structure by Mg ions. Speci®c andnon-speci®c effects. J. Mol. Biol. 237, 577-587.
Leontis, N. B. & Westhof, E. (1998). A common motiforganizes the structure of multi-helix loops in 16 Sand 23 S ribosomal RNAs. J. Mol. Biol. 283, 571-583.
Lott, W. B., Pontius, B. W. & von Hippel, P. H. (1998).A two-metal ion mechanism operates in the ham-merhead ribozyme-mediated cleavage of an RNAsubstrate. Proc. Natl Acad. Sci. USA, 95, 542-547.
Matsumura, K. & Komiyama, M. (1997). Enormouslyfast hydrolysis by lanthanide(III) ions under physio-logical conditions: eminent candidates for noveltools of biotechnology. J. Biochem. 122, 387-394.
McFail-Isom, L., Shui, X. & Williams, L. D. (1998). Diva-lent cations stabilize unstacked conformations ofDNA and RNA by interacting with base p systems.Biochemistry, 37, 17105-17111.
Milligan, J. F. & Uhlenbeck, O. C. (1989). Synthesis ofsmall RNAs using T7 RNA polymerase. MethodsEnzymol. 180, 51-62.
Morrow, J. R. (1996). Hydrolytic cleavage of RNA cata-lyzed by metal ion complexes. Methods Ions Biol.Syst. 33, 561-592.
Murchie, A. I., Thomson, J. B., Walter, F. & Lilley, D. M.(1998). Folding of the hairpin ribozyme in its natu-ral conformation achieves close physical proximityof the loops. Mol. Cell, 1, 873-881.
Murray, J. B., Seyhan, A. A., Walter, N. G., Burke, J. M.& Scott, W. G. (1998). The hammerhead, hairpinand VS ribozymes are catalytically pro®cient inmonovalent cations alone. Chem. Biol. 5, 587-595.
Narlikar, G. J. & Herschlag, D. (1997). Mechanisticaspects of enzymatic catalysis: lessons from com-parison of RNA and protein enzymes. Annu. Rev.Biochem. 66, 19-59.
Nesbitt, S., Hegg, L. A. & Fedor, M. J. (1997). An unu-sual pH-independent and metal-ion-independentmechanism for hairpin ribozyme catalysis. Chem.Biol. 4, 619-630.
Nesbitt, S. M., Erlacher, H. A. & Fedor, M. J. (1999). Theinternal equilibrium of the hairpin ribozyme:temperature, ion and pH effects. J. Mol. Biol. 286,1009-1024.
Pan, T. & Uhlenbeck, O. C. (1992). A small metalloribo-zyme with a two-step mechanism. Nature, 358, 560-563.
Pan, T., Long, D. M. & Uhlenbeck, O. C. (1993). Divalentmetal ions in RNA folding and catalysis. In RNAWorld (Gesteland, R. F., Cech, T. R. & Atkins, J. F.,eds), pp. 271-302, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
Pinard, R., Heckman, J. E. & Burke, J. M. (1999a).Alignment of the two domains of the hairpin ribo-zyme-substrate complex de®ned by interdomainphotoaf®nity crosslinking. J. Mol. Biol. 287, 239-251.
Pinard, R., Lambert, D., Walter, N. G., Heckman, J. E.,Major, F. & Burke, J. M. (1999b). Structural basis forthe guanosine requirement of the hairpin ribozyme.Biochemistry, 38, 16035-16039.
Pyle, A. M. (1993). Ribozymes: a distinct class of metal-loenzymes. Science, 261, 709-714.
Pyle, A. M. (1996). Role of metal ions in ribozymes.Metal Ions Biol. Syst. 32, 479-520.
Sigurdsson, S. T., Thomson, J. B. & Eckstein, F. (1998).Small ribozymes. In RNA Structure and Function(Simons, R. W. & Grunberg-Manago, M., eds), pp.339-375, Cold Spring Harbor Press, Cold SpringHarbor, NY.
Smith, D. (1995). Magnesium as the catalytic center ofRNA enzymes. In The Biological Chemistry of Mag-nesium (Cowan, J. A., ed.), pp. 111-136, VCH, NewYork.
Smith, D., Burgin, A. B., Haas, E. S. & Pace, N. R.(1992). In¯uence of metal ions on the ribonuclease Preaction. Distinguishing substrate binding from cat-alysis. J. Biol. Chem. 267, 2429-2436.
Stein, C. A. & Cohen, J. A. (1988). Oligodeoxynucleo-tides as inhibitors of gene expression: a review.Cancer Res. 48, 2659-2668.
Steitz, T. A. & Steitz, J. A. (1993). A general two-metal-ion mechanism for catalytic RNA. Proc. Natl Acad.Sci. USA, 90, 6498-6502.
Suga, H., Cowan, J. A. & Szostak, J. W. (1998). Unusualmetal ion catalysis in an acyl-transferase ribozyme.Biochemistry, 37, 10118-10125.
Tajmir-Riahi, H. A., Ahmad, R. & Naoui, M. J. (1993).Interaction of calf-thymus DNA with trivalent La,Eu, and Tb ions. Metal ion binding, DNA conden-sation and structural features. J. Biomol. Struct.Dynam. 10, 865-877.
Thomson, J. B. & Lilley, D. M. (1999). The in¯uence ofjunction conformation on RNA cleavage by thehairpin ribozyme in its natural junction form. RNA,5, 180-187.
Topal, M. D. & Fresco, J. R. (1980). Fluorescence of ter-bium ion-nucleic acid complexes: a sensitive speci®cprobe for unpaired residues in nucleic acids. Bio-chemistry, 19, 5531-5537.
Tung, C. S., Harvey, S. C. & McCammon, J. A. (1984).Large-amplitude bending motions in phenylalaninetransfer RNA. Biopolymers, 23, 2173-2193.
Walter, N. G. & Burke, J. M. (1997). Real-time monitor-ing of hairpin ribozyme kinetics through base-speci®c quenching of ¯uorescein-labeled substrates.RNA, 3, 392-404.
Walter, N. G. & Burke, J. M. (1998). The hairpin ribo-zyme: structure, assembly and catalysis. Curr. Opin.Chem. Biol. 2, 24-30.

Cation Binding in the Hairpin Ribozyme 555
Walter, N. G., Hampel, K. J., Brown, K. M. & Burke,J. M. (1998). Tertiary structure formation in the hair-pin ribozyme monitored by ¯uorescence resonanceenergy transfer. EMBO J. 17, 2378-2391.
Walter, N. G., Burke, J. M. & Millar, D. P. (1999).Stability of hairpin ribozyme tertiary structure isgoverned by the interdomain junction. NatureStruct. Biol. 6, 544-549.
Wedekind, J. E. & McKay, D. B. (1998). Crystallographicstructures of the hammerhead ribozyme: relation-ship to ribozyme folding and catalysis. Annu. Rev.Biophys. Biomol. Struct. 27, 475-502.
Welch, P. J., Hampel, A., Barber, J., Wong-Staal, F. &Yu, M. (1996). Inhibition of HIV replication by thehairpin ribozyme. Nucl. Acids Mol. Biol. 10, 315-327.
Wenzel, T. J. & Collette, L. M. (1988). Lanthanide ionsas luminescent chromophores for the liquid chroma-
tographic detection of polynucleotides and nucleicacids. J. Chromatog. 436, 299-307.
Wincott, F., DiRenzo, A., Shaffer, C., Grimm, S., Tracz,D., Workman, C., Sweedler, D., Gonzalez, C.,Scaringe, S. & Usman, N. (1995). Synthesis,deprotection, analysis and puri®cation of RNAand ribozymes. Nucl. Acids Res. 23, 2677-2684.
Yarus, M. (1993). How many catalytic RNAs? Ions andthe Cheshire cat conjecture. FASEB J. 7, 31-39.
Yonuschot, G., Helman, D., Mushrush, G., Woude, G. V.& Robey, G. (1978). Terbium as a solid-state probefor RNA. Bioinorg. Chem. 8, 405-418.
Young, K. J., Gill, F. & Grasby, J. A. (1997). Metal ionsplay a passive role in the hairpin ribozymecatalysed reaction. Nucl. Acids Res. 25, 3760-3766.
Edited by J. Doudna
(Received 12 January 2000; received in revised form 14 March 2000; accepted 14 March 2000)