prodomains – adaptors – oligomerization: the pursuit of caspase activation in apoptosis
TRANSCRIPT

TIBS 24 – JANUARY 1999
10968–0004/99/$ – See front matter © 1999, Elsevier Science. All rights reserved. PII: S0968-0004(98)01332-2
The mammalian homologues of theCaenorhabditis elegans death proteaseCED-3, now known as caspases, are keycomponents of the apoptotic machin-ery1–3. They are normally present as pre-cursor molecules in healthy cells and,upon receiving a death signal, undergoproteolytic cleavage to produce two sub-units that constitute the active enzyme.The crystal structure of fully processedcaspases indicates that the active enzymeis a heterotetramer that comprises twoheterodimers derived from two precursormolecules1–3. Several earlier studies sug-gested that some caspases are able toautoprocess. In vitro data suggest thatonce initial upstream caspases are acti-vated, they can mediate the processing ofdownstream caspases1–3. However, untilrecently, the exact mechanisms of cas-pase activation in apoptosis had remainedillusive. Several recent reports now indi-cate that the activation of a number of caspases occurs through oligomerization.
Caspase prodomainsOn the basis of their primary structures,
caspases can be grouped into two classes:those that possess long N-terminal pro-domains (class I caspases); and thosethat either possess short prodomains orlack prodomains (class II caspases). Theprodomains in class I caspases were initially thought to be negative regu-latory domains, but these regions arenow known to contain protein–protein-interaction domains that help recruitclass I caspase precursors to specificdeath complexes. Activation of class IIcaspases, such as caspase-3, caspase-6and caspase-7, appears to require pro-teolytic processing by class I caspases1–3.Thus, class II caspases are the down-stream caspases that mediate proteolysisof a number of cellular proteins in a cellthat is destined to die. Activation ofclass I caspases is of fundamental impor-tance in cell death commitment. Here, wereview recent developments in our under-standing of the mechanisms that medi-ate the activation of class I caspases.
The first direct evidence for direct re-cruitment of a class I caspase to a death
complex came when two groups ob-served that caspase-8 associates withthe Fas death-inducing signalling com-plex by binding to the adaptor FADD4,5.Subsequent studies identified an adap-tor molecule, RAIDD, that specificallyinteracts with procaspase-2 (Ref. 6).RAIDD also interacts with RIP, a serine/threonine kinase that is recruited to tumour necrosis factor (TNF) receptor 1(TNFR-1) and mediates TNF-induced NF-kB activation7. Another recently clonedRIP-like kinase, CARDIAK, specifically interacts with caspase-1 (Ref. 8). In allcases, the interaction with the adaptormolecules and procaspases is mediatedby specific domains that are present inthe caspase prodomains.
Caspase recruitmentHomotypic interactions between vari-
ous components of the death complexesare crucial for apoptotic signalling. Threetypes of protein–protein-interaction do-mains are present in apoptotic molecules7:death domains (DDs); death-effector do-mains (DEDs); and caspase-recruitmentdomains (CARDs). DDs are common inupstream components of the apoptoticpathways, such as death receptors (e.g.CD95, TNFR-1 and DR3) and moleculesthat are recruited to these receptors (e.g.FADD, TRADD and RIP), whereas DEDsand CARDs are responsible for recruitingclass I caspases to specific death com-plexes through adaptor molecules7. DD-mediated association between receptorsand adaptors is usually a consequenceof ligand-dependent activation of deathreceptors. The overall structure of DDs,DEDs and CARDs is very similar (all con-sist of six a-helices); however, there aredistinct differences between the threedomains. For instance, certain mu-tations in the Fas DD inhibit protein–protein interactions; corresponding mu-tations have no effect on the FADD DED.In addition, a hydrophobic region in theFADD DED that is crucial for binding tothe caspase-8 DED is absent from theFas DD9,10. Residues that are required forthe apoptotic activity of DEDs and DDsare not conserved in CARDs, which
suggests that these three domains use a different set of residues to define their binding specificity and function11.Electrostatic interactions appear to me-diate homotypic interactions in DDs andCARDs, whereas the interactions betweenDEDs probably involve hydrophobic regions9–11.
CARDs are present in CED-3, caspase-1, caspase-2, caspase-9 and the adaptormolecules CED-4, CARDIAK, RAIDD andAPAF-18,11,12 (Table 1). The NMR structureof the RAIDD CARD indicates that this do-main consists of six antiparallel amphi-pathic a-helices, which are closely packedaround a hydrophobic core in a fashionsimilar to the structure of the Fas DD(Ref. 11). However, the orientation of twohelices in the RAIDD CARD differs signifi-cantly from the corresponding helices inthe Fas DD. The surface of the RAIDDCARD contains a basic region formed byhelices 1, 3 and 4, and an acidic regionformed by helices 2, 5 and 6. Modellingof the caspase-2 CARD indicates thatthis domain has a remarkably similarstructure and surface polarity; the inter-action between RAIDD and caspase-2might, therefore, be mediated by electro-static bonding between the two CARDs.Chou et al.11 propose a similar mode ofprotein–protein interaction for theCARDs in APAF-1 and caspase-9, and forthe CARDs in CED-4 and CED-3. Thesestructural studies have provided signifi-cant insight into how the recruitment ofprocaspase molecules can be mediatedthrough homotypic interactions betweenspecific domains.
Oligomerization of adaptors andprocaspases
The concept of caspase activationthrough oligomerization is not new. In1995, Gu et al.13 proposed that oligomer-ization is required for autoprocessing ofthe caspase-1 zymogen. Later studies,using two-hybrid assays, found that procaspase-1 dimerization in yeast ismediated by the prodomain14. Similar interactions are evident in caspase-2(Ref. 15). The dimerization of the caspase precursors occurs prior to processing of
FRONTLINES
Prodomains – adaptors –oligomerization: the pursuit ofcaspase activation in apoptosis
Table 1. Caspases, adaptors and thedomains that mediate their interactions
Caspase Adaptor Type of interaction
CED-3 CED-4 CARD–CARDCaspase-1 CARDIAK? CARD–CARDCaspase-2 RAIDD CARD–CARDCaspase-8 FADD DED–DEDCaspase-9 APAF-1 CARD–CARDCaspase-10 FADD DED–DED

FRONTLINES TIBS 24 – JANUARY 1999
2
the caspase molecules. Therefore, prodomain-dependent dimerization islikely to be necessary for processing andactivation to occur. These studies alsoshowed that activation is dependent onthe autocatalytic activities of the cas-pase precursors, and that removal ofthe prodomain abolishes their process-ing in Saccharomyces cerevisiae15.
The homodimerization of caspase pre-cursors is distinct from heterodimer-ization between a procaspase and anadaptor molecule. However, it is notclear whether both sets of interactionsare mediated exclusively by CARDs orwhether other regions within the pro-domains of caspase-1 and caspase-2 arerequired for homodimerization of pre-cursor molecules. Given that both cas-pases can homodimerize without anyapparent need for other molecules, andthat dimerization is required and suffi-cient for processing to occur, the functionof the adaptors CARDIAK and RAIDD isprobably simply to recruit procaspase-1and procaspase-2 to a death-signallingcomplex. Caspase-8 and caspase-10 con-tain two DEDs in their prodomain regions(Table 1). At least one of the DEDs inthese caspases interacts with the DED inFADD, recruiting the procaspase to acti-vated death receptors, such as Fas (Ref. 7),whereas the other DED might be requiredfor homodimerization. Several recentpapers show that when prodomains ofsome caspases are replaced by proteindomains that can be induced to dimerizeby artificial means, forced oligomerizationis sufficient to mediate procaspase acti-vation16–20. In another study, our groupshowed that fusion of the caspase-2prodomain to procaspase-3, a class IIcaspase that is unable to autoprocess, issufficient to mediate autocatalytic acti-vation of the chimeric molecule in trans-fected cells21. The activation of chimericcaspase-3 is a result of dimerizationthrough the caspase-2 prodomain21.
The activation of caspase-9 requiresAPAF-1, dATP and cytochrome c, which isreleased from the mitochondria in cellscommitted to apoptosis. Binding ofcytochrome c and dATP to APAF-1 pre-sumably causes conformational changesin the molecule that expose the N-terminalCARD; this enables APAF-1 to interactwith the CARD domain in procaspase-9(Ref. 19). APAF-1 can also form oligomersthrough its CED-4-like domains19. Adap-tor oligomerization and recruitment ofcaspase-9 through CARD mediates auto-catalytic processing of caspase-9. CED-4also oligomerizes – a process necessaryfor the activation of proCED-3 (Ref. 20).
Interestingly, the regions of APAF-1 andCED-4 that are required for oligomer-ization are distinct from N-terminalCARDs in these proteins.
When overexpressed, the DEDs ofFADD and caspase-8 form intracellularfilaments, presumably through oligo-merization, which are capable of recruit-ing procaspases22. The caspase-2 precur-sor and the prodomain of caspase-2 alsoform filamentous and dot-like structures,mostly within the nucleus of the trans-fected cells, possibly as a result of CARD-mediated oligomerization23. Althoughformation of these higher-order structuresmight simply be due to overexpressionof the protein, these experiments demon-strate that, under conditions that pro-mote localized high concentrations ofDEDs or CARDs, oligomerization canoccur efficiently. All these results sug-gest that the primary role of recruitmentthrough adaptors is to bring procaspasemolecules into close proximity. This presumably generates a high localizedconcentration of procaspase moleculesthat is sufficient to induce their auto-proteolytic processing, which generatesactive caspase.
How does oligomerization mediate caspaseactivation?
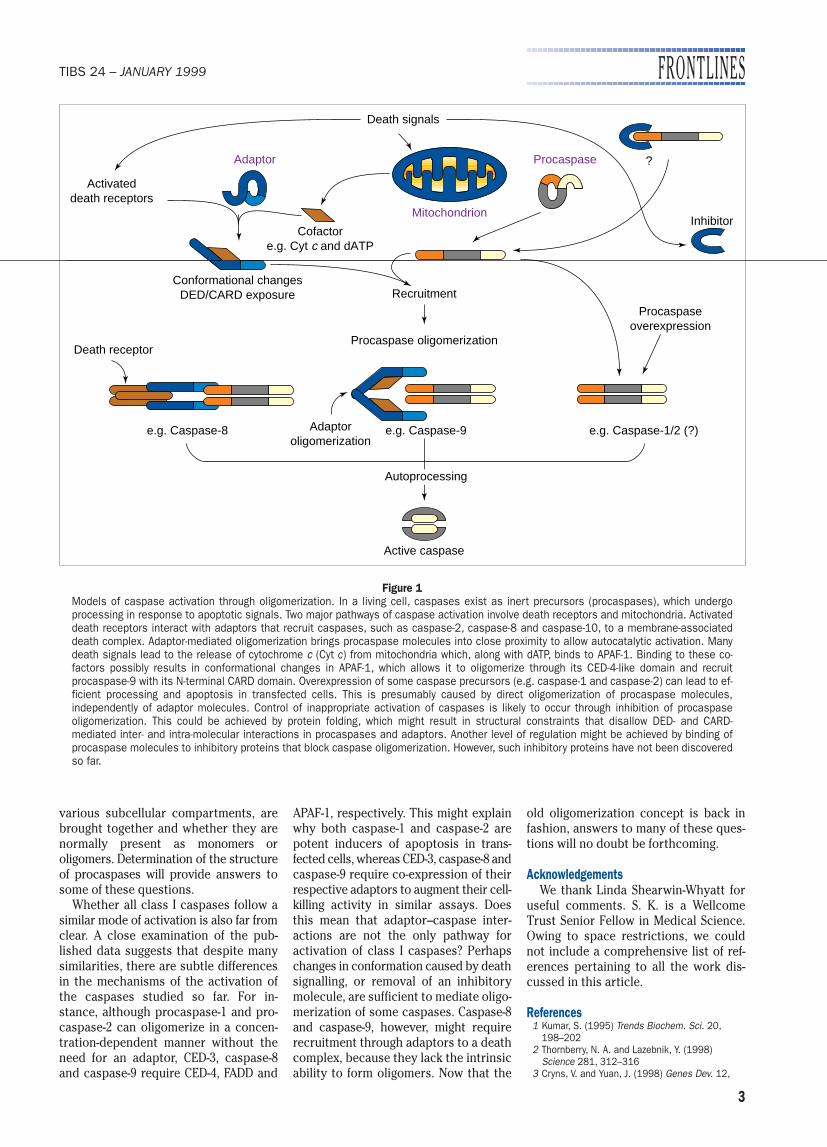
The events that lead to activation ofclass I caspases appear to follow the se-quence outlined in Fig. 1: (1) confor-mational changes caused by upstreamsignalling events allow recruitment ofadaptor molecules to a death complex;(2) this results in additional changesthat allow recruitment of caspasesthrough specific domains; (3) once inclose proximity, procaspase moleculesform transient oligomers, which allowsinter- or intra-molecular catalysis andactivation. The exact mechanism of auto-catalytic processing of procaspases isnot fully understood but, clearly, for autocatalytic processing to occur, eitherthe caspase precursors carry some intrinsic enzyme activity, or proximity-induced dimerization of precursors allows refolding, which generates astructure similar to that of an active,processed enzyme. The caspase-8 zymo-gen has low levels of enzymatic activ-ity16; however, it is not clear whether asingle precursor molecule can acquire a catalytically active conformation.
How is caspase activation regulated? Activation of the initial caspase in an
apoptotic pathway is the priming eventin cell death. As such, it is likely to be atightly regulated process. Given that
caspase activation is achieved througholigomerization of the precursor mol-ecules, the regulation of oligomerizationmust be important. Caspase-8-like mol-ecules, such as viral and cellular FLIPs,that contain DEDs but lack the catalyticfunction inhibit caspase-8 activation byblocking caspase-8 recruitment to deathcomplexes2,3. Another inhibitor of apop-tosis, ARC, contains a CARD domainand, thus, might function by blockingthe interaction between specific CARD-containing molecules24. APAF-1 requiresbinding of cytochrome c and dATP beforeit can interact with procaspase-9. A trun-cated APAF-1 protein that lacks the C-terminal region, which comprises severalWD40 repeats, can activate procaspase-9independently of dATP and cytochrome c.This suggests that WD40 repeats exert anegative regulatory effect, possibly bymasking the CARD and CED-4-like do-mains, which are required for APAF-1–caspase-9 heterodimerization and APAF-1–APAF-1 homodimerization, respectively19.As mentioned earlier, conformationalchanges induced by cytochrome c anddATP probably induce refolding of themolecule, which exposes the CARD andenables it to interact with procaspase-9.The binding of CED-9 to CED-4 blocksoligomerization of CED-4 molecules, whichsuggests that CED-9 and its mammaliancounterparts, the anti-apoptotic BCL2members, inhibit apoptosis primarily by modulating oligomerization-mediatedcaspase activation20. Another level ofregulation might be provided by directbinding of inhibitory molecules to pro-caspases themselves, which wouldblock inappropriate dimerization andactivation (Fig. 1). Caspase activation isa complex phenomenon, requiring mul-tiple protein–protein interactions andpossibly resulting in conformationalchanges in various proteins. Evolutionof this multifaceted activation mecha-nism possibly allowed tighter control ofapoptotic initiation through blocking ofinappropriate oligomerization of class Icaspases.
What next? There is now little doubt that protein–
protein interactions and oligomerizationplay a vital role in the activation of class Icaspases; however, several key questionsremain. For example, we do not knowwhether, in a healthy cell, procaspasesare usually bound to adaptors, or whetherthey are only recruited by adaptors inresponse to a death signal. We also donot know how procaspase molecules,which might normally be dispersed in
TIBS 24 – JANUARY 1999
3
various subcellular compartments, arebrought together and whether they arenormally present as monomers oroligomers. Determination of the structureof procaspases will provide answers tosome of these questions.
Whether all class I caspases follow asimilar mode of activation is also far fromclear. A close examination of the pub-lished data suggests that despite manysimilarities, there are subtle differencesin the mechanisms of the activation ofthe caspases studied so far. For in-stance, although procaspase-1 and pro-caspase-2 can oligomerize in a concen-tration-dependent manner without theneed for an adaptor, CED-3, caspase-8and caspase-9 require CED-4, FADD and
APAF-1, respectively. This might explainwhy both caspase-1 and caspase-2 arepotent inducers of apoptosis in trans-fected cells, whereas CED-3, caspase-8 andcaspase-9 require co-expression of theirrespective adaptors to augment their cell-killing activity in similar assays. Doesthis mean that adaptor–caspase inter-actions are not the only pathway for activation of class I caspases? Perhapschanges in conformation caused by deathsignalling, or removal of an inhibitorymolecule, are sufficient to mediate oligo-merization of some caspases. Caspase-8and caspase-9, however, might requirerecruitment through adaptors to a deathcomplex, because they lack the intrinsicability to form oligomers. Now that the
old oligomerization concept is back infashion, answers to many of these ques-tions will no doubt be forthcoming.
AcknowledgementsWe thank Linda Shearwin-Whyatt for
useful comments. S. K. is a WellcomeTrust Senior Fellow in Medical Science.Owing to space restrictions, we couldnot include a comprehensive list of ref-erences pertaining to all the work dis-cussed in this article.
References1 Kumar, S. (1995) Trends Biochem. Sci. 20,
198–2022 Thornberry, N. A. and Lazebnik, Y. (1998)
Science 281, 312–3163 Cryns, V. and Yuan, J. (1998) Genes Dev. 12,
FRONTLINESDeath signals
Death receptor
e.g. Caspase-8 e.g. Caspase-9
Active caspase
Adaptoroligomerization
e.g. Caspase-1/2 (?)
Procaspaseoverexpression
Mitochondrion
Procaspase
Cofactore.g. Cyt c and dATP
Activateddeath receptors
Conformational changesDED/CARD exposure Recruitment
Procaspase oligomerization
?
Inhibitor
Adaptor
Autoprocessing
Figure 1Models of caspase activation through oligomerization. In a living cell, caspases exist as inert precursors (procaspases), which undergoprocessing in response to apoptotic signals. Two major pathways of caspase activation involve death receptors and mitochondria. Activateddeath receptors interact with adaptors that recruit caspases, such as caspase-2, caspase-8 and caspase-10, to a membrane-associateddeath complex. Adaptor-mediated oligomerization brings procaspase molecules into close proximity to allow autocatalytic activation. Manydeath signals lead to the release of cytochrome c (Cyt c) from mitochondria which, along with dATP, binds to APAF-1. Binding to these co-factors possibly results in conformational changes in APAF-1, which allows it to oligomerize through its CED-4-like domain and recruit procaspase-9 with its N-terminal CARD domain. Overexpression of some caspase precursors (e.g. caspase-1 and caspase-2) can lead to ef-ficient processing and apoptosis in transfected cells. This is presumably caused by direct oligomerization of procaspase molecules,independently of adaptor molecules. Control of inappropriate activation of caspases is likely to occur through inhibition of procaspase oligomerization. This could be achieved by protein folding, which might result in structural constraints that disallow DED- and CARD-mediated inter- and intra-molecular interactions in procaspases and adaptors. Another level of regulation might be achieved by binding ofprocaspase molecules to inhibitory proteins that block caspase oligomerization. However, such inhibitory proteins have not been discoveredso far.

FRONTLINES TIBS 24 – JANUARY 1999
4 0968–0004/99/$ – See front matter © 1999, Elsevier Science. All rights reserved. PII: S0968-0004(98)01339-5
1551–1570 4 Muzio, M. et al. (1996) Cell 85, 817–8275 Boldin, M. P., Goncharov, T. M., Goltsev, Y. V. and
Wallach, D. (1996) Cell 85, 803–8156 Duan, H. and Dixit, V. M. (1997) Nature 385,
86–897 Ashkenazi, A. and Dixit, V. M. (1998) Science
281, 305–3088 Thome, M. et al. (1998) Curr. Biol. 16, 885–8889 Huang, B. et al. (1996) Nature 384, 638–641
10 Eberstadt, M. et al. (1998) Nature 392, 941–94511 Chou, J. J., Matsuo, H., Duan, H. and Wagner, G.
(1998) Cell 94, 171–18012 Hoffman, K., Bucher, P. and Tschopp, J. (1997)
Trends Biochem. Sci. 22, 155–156 13 Gu, Y. et al. (1995) EMBO J. 14, 1923–193114 Van Criekinge, W. et al. (1996) J. Biol. Chem.
271, 27245–2724815 Butt, A. J., Harvey, N. L., Parasivam, G. and
Kumar, S. (1998) J. Biol. Chem. 273,6763–6768
16 Muzio, M. et al. (1998) J. Biol. Chem. 273,2926–2930
17 Yang, X., Chang, H. Y. and Baltimore, D. (1998)Mol. Cell 1, 319–325
18 MacCorkle, R. A., Freeman, K. W. and Spencer, D. M. (1998) Proc. Natl. Acad. Sci.U. S. A. 95, 3655–3660
19 Srinivasula, S. M., Ahmad, M., Fernandes-Alnemri, T. and Alnemri, E. S. (1998) Mol. Cell1, 949–957
20 Yang, X., Chang, H. Y. and Baltimore, D. (1998)Science 281, 355–357
21 Colussi, P. A., Harvey, N. L., Shearwin-Whyatt, L. M.
and Kumar, S. (1998) J. Biol. Chem. 273,26566–26570
22 Siegel, R. M. et al. (1998) J. Cell Biol. 141,1243–1253
23 Colussi, P. A., Harvey, N. L. and Kumar, S.(1998) J. Biol. Chem. 273, 24535–24542
24 Koseki, T., Inohara, N., Chen, S. and Nunez, G.(1998) Proc. Natl. Acad. Sci. U. S. A. 95,5156–5160
SHARAD KUMAR AND PAUL A. COLUSSI
Hanson Centre for Cancer Research, Instituteof Medical and Veterinary Science, Frome Rd,Adelaide, Australia.Email: [email protected]
To understand the mechanism of con-densation of transcriptionally inactivechromatin we must identify the detailsof interactions between its components.The fundamental structural unit of thecondensed 300-Å fibre is the chromato-some (see Box 1); this contains a histoneoctamer and one molecule of linker his-tone, which are associated with 168 bpof DNA. The chromatosomes are sep-arated by varying lengths of linker DNA,and the folding of the fibre is facilitatedby linker histone H1 and its variants,which include H5. The linker histonesthemselves contain basic N- and C-terminal tails, which flank a centralglobular domain. This domain is suffi-cient for chromatosome formation butits position in the particle has remainedcontroversial1. One recent model2 pro-poses that the globular domain bridgestwo superhelical turns between one ter-minus and the dyad (see Box 1), while asecond places the domain on the insideof one turn away from the dyad3,4. Newevidence5 indicates that, at least in bulkchromatin, the first model is correct.
Location and orientation of the globulardomain of histone H5 in the nucleosome
An early model of the 300-Å fibre pro-posed that it has a solenoidal structurein which a helical array of nucleosomesis linked by histone H1 and that theglobular domain of this histone binds tothe dyad region located on the interiorof the solenoid6. In this model, adjacent
nucleosomes in the helix are connectedby a single linker DNA. Since this pro-posal, various alternative structures thatdiffer in the path of the linker DNA (e.g.the zig-zag model) or in the location oflinker histone on the chromatosomehave been proposed.
Ramakrishnan and co-workers7 experi-mentally confirmed the interior locationof H1 by neutron diffraction, but evidencefor its binding site in the chromatosomeremained elusive1. The three-dimensionalstructures of the globular domains ofboth H1 and H5 (GH1 and GH5, respec-tively) showed that the globular domainis a three-helix bundle that shares struc-tural homology with helix-turn-helixDNA-binding proteins. These structuresalso revealed the existence of a second-ary putative DNA-binding site, which isseparated from the recognition helix by25 Å8,9 – an observation that is consistentwith the demonstration that this do-main can bind two DNA duplexes simul-taneously10,11. This structure suggesteda model in which the globular domainbridges two adjacent DNA gyres (seeBox 1 and Fig. 1).
Both these predictions have now beenconfirmed. Goytisolo et al.12 have demon-strated that mutations in the secondarybinding site abolish the ability of GH5 tobind two duplexes and to form chro-matosomes. In addition, the groups ofMuyldermans and Ramakrishnan, incollaboration with our group5, havemapped the binding site of the globular
domain of linker histone H5 on mixed-sequence chicken chromatosomes by conjugating a protein–DNA cross-linking reagent to specific Ser→Cys sub-stitutions. These experiments showthat helix III binds in the major grooveof the first helical turn of the chromato-somal DNA (i.e. close to the end) whilethe secondary DNA-binding site on theopposite face of the globular domain ofH5 contacts the chromatosomal DNAclose to its midpoint. By exploiting theability of some Ser→Cys mutants toself-dimerize, we5 inferred that helix Iand helix II of the globular domain of H5face the solvent and the nucleosome,respectively. In bulk chromatin, theglobular domain of the linker histonethus forms a bridge between one termi-nus of chromatosomal DNA and the mid-point. This position of GH5 on the chro-matosome would place its C-terminuson the outside of the particle, betweenone end of chromatosomal DNA and thecentral gyre, and its N-terminus on theinner surface of the entering DNA. Theorientation of the globular domain rela-tive to the surface of the octamer is alsoconsistent with the protection of lysineresidues in helix II from reductivemethylation in chromatin containing H5(Ref. 13).
Both the original model for GH5 pos-itioning proposed by Allan et al.14 andour more recent variant5 argue that GH5binds at, or close to, the chromatosomedyad (Fig. 1b,c). However, a second radi-cally different model was recently pro-posed by Wolffe and Hayes and theircolleagues3,4. This model is based oncrosslinking3, site-directed DNA cleav-age4 and micrococcal-nuclease (an en-zyme that cuts linker DNA) mapping15
on a chromatosome formed on Xenopusborealis somatic 5S rDNA, and proposesthat the globular domain of linker histone
The location of the linker histoneon the nucleosome