prostatic acid phosphatase as a regulator of endo/exocytosis and
TRANSCRIPT

Prostatic Acid Phosphatase as a Regulator of Endo/Exocytosis and Lysosomal Degradation
MEDICUMDEPARTMENT OF CLINICAL CHEMISTRY AND HAEMATOLOGYFACULTY OF BIOLOGICAL AND ENVIRONMENTAL SCIENCESDOCTORAL PROGRAMME IN INTEGRATIVE LIFE SCIENCEUNIVERSITY OF HELSINKI
CÉSAR L. ARAUJO
dissertationes scholae doctoralis ad sanitatem investigandam universitatis helsinkiensis 7/2016
7/2016Helsinki 2016 ISSN 2342-3161 ISBN 978-951-51-1849-3
CÉ
SA
R L
. AR
AU
JO P
rostatic Acid
Ph
osph
atase as a Regu
lator of En
do/E
xocytosis and
Lysosom
al Degrad
ation
Recent Publications in this Series
94/2015 Jani SaksiStroke-Associated Carotid Atherosclerosis - a Role for FABP4 in Atheroma Vulnerability and Cardiometabolic Risk95/2015 Otto HemminkiCancer Immunotherapy with a Gene Modified Serotype 3 Oncolytic Adenovirus96/2015 Jukka KoskelaIndividual Trajectories in Asthma and COPD: A Longitudinal Perspective to Obstructive Lung Disease97/2015 Miira KlemettiTrends In Obstetric and Perinatal Outcomes of Women with Type 1 Diabetes During 1988-2011 – A Finnish Population-Based Observational Study98/2015 Ileana B. QuinteroPhysiological Functions of Prostatic Acid Phosphatase99/2015 Minna MatikainenLaunching New Products in the Finnish Pharmaceutical Industry: A Relationship Approach100/2015 Päivi Ylä-AnttilaPhagophore Membrane Connections and RAB24 in Autophagy101/2015 Kaisa KyöstiläIdentification of Novel Mutations and Molecular Pathways for Canine Neurodegeneration and Chondrodysplasia102/2015 Emmi JoensuuEpigenetic Alterations in Sporadic and Familial Cancers103/2015 Elina ReponenPreoperative Risk-Assessment Methods, Short-Term Outcome, and Patient Satisfaction in Elective Cranial Neurosurgery104/2015 Riina KandolinCardiac Sarcoidosis in Giant Cell Myocarditis in Finland106/2015 Karmen KappPolyphenolic and Essential Oil Composition of Mentha and Their Antimicrobial Effect107/2015 Dina PopovaNeurophysiological mechanisms of Plasticity Induced in Adult Brain1/2016 Pauliina SaurusRegulation of Podocyte Apoptosis in Diabetic Kidney Disease – Role of SHIP2, PDK1 and CDK22/2016 Sanna ToivonenDerivation of Hepatocyte Like Cells from Human Pluripotent Stem Cells3/2016 Marjaana PeltolaAMIGO-Kv2.1 Potassium Channel Complex: Identification and Association with Schizophrenia-Related Phenotypes4/2016 Niko-Petteri NykänenCellular Physiology and Cell-to-Cell Propagation of Tau in Neurodegeneration: The Impact of Late-Onset Alzheimer’s Disease Susceptibility Genes5/2016 Liisa KorkaloHidden Hunger in Adolescent Mozambican Girls: Dietary Assessment, Micronutrient Status, and Associations between Dietary Diversity and Selected Biomarkers6/2016 Teija OjalaLactobacillus crispatus and Propionibacterium freudenreichii: A Genomic and Transcriptomic View

Prostatic acid phosphatase as a regulator of endo/exocytosis and
lysosomal degradation
César L. Araujo
Department of Clinical Chemistry and Haematology, Medicum
Department of Biosciences, Division of Biochemistry and Biotechnology, Faculty of Biological and Environmental Sciences
FinPharma Doctoral Program Drug Discovery Section (FPDP-D) Integrative Life Science Doctoral Program
University of Helsinki
Finland
Academic Dissertation
To be publicly discussed with the permission of Faculty of Biological and Environmental Sciences, University of Helsinki, in
Lecture hall 2, Haartman Institute, on Friday 29th of January 2016 at 12 o’clock noon.
Helsinki 2016

Supervisor
Professor Pirkko Vihko, M.D., Ph.D. Department of Clinical Chemistry and Haematology University of Helsinki
Thesis Advisory Committee Päivi Lakkisto, M.D., Ph.D., Adj. Prof.
Department of Clinical Chemistry and Haematology University of Helsinki
Hannu Koistinen, D.Sc., Adj. Prof.
Department of Clinical Chemistry and Haematology University of Helsinki
Pre-examiners
Hannu Koistinen, D.Sc., Adj. Prof. Department of Clinical Chemistry and Haematology University of Helsinki
Sakari Kellokumpu, Ph.D., Adj. Prof.
Department of Biochemistry University of Oulu
Opponent Professor Jorma Palvimo, Ph.D.
Institute of Biomedicine University of Eastern Finland
Custos Professor Kari Keinänen, Ph.D.
Department of Biosciences, Division of Biochemistry and Biotechnology University of Helsinki
Cover picture was rendered by wordle.net web service. Published in the series Dissertationes Scholae Doctoralis Ad Sanitatem Investigandam Universitatis Helsinkiensis ISBN 978-951-51-1849-3 (Paperback) ISBN 978-951-51-1850-9 (PDF) ISSN 2342-3161 (print) ISSN 2342-317X (online) http://ethesis.helsinki.fi Hansaprint Oy Vantaa 2016

“Faith and reason are like two wings on which the human spirit rises to the contemplation of truth; and God has placed in the human heart a desire to know the truth—in a word, to know himself—so that, by knowing and loving God, men and
women may also come to the fullness of truth about themselves“
(St. John Paul II, Fides et Ratio)
To my family…


TABLE OF CONTENTS
LIST OF ORIGINAL PUBLICATIONS............................................................................................. I
ABBREVIATIONS ...................................................................................................................... II
ABSTRACT ............................................................................................................................... IV
1 REVIEW OF THE LITERATURE ................................................................................................ 1
1.1 Prostatic acid phosphatase and prostate cancer .................................................................... 1 1.2 Structure of prostatic acid phosphatase ................................................................................. 2 1.3 Prostatic acid phosphatase as a 5’-ectonucleotidase and a thiamine
monophosphatase .................................................................................................................... 4 1.4 Homologous histidine acid phosphatases found in other organisms .................................... 4
1.4.1 Histidine acid phosphatase of Francisella tularensis (FtHAP) ............................................... 4 1.4.2 Membrane-bound acid phosphatase of Trypanosoma brucei (TbMBAP) ............................ 5
1.5 Lysosomal biogenesis and function ......................................................................................... 6 1.6 Function of the tyrosine-based lysosomal targeting motif Yxxφ ........................................... 7 1.7 The SNARE complex and the mechanism of exocytosis ......................................................... 8 1.8 Disturbed endocytosis as a hallmark of cancer....................................................................... 9 1.9 Mouse submandibular salivary glands as a model of exocrine organ ................................. 11
2 AIMS OF THIS STUDY .......................................................................................................... 14
3 MATERIALS AND METHODS................................................................................................ 15
3.1 Ethics statement ..................................................................................................................... 15 3.2 Mice ........................................................................................................................................ 15 3.3 Stable transfected LNCaP cells ............................................................................................... 15 3.4 Bioinformatics determination of TMPAP sequence elements ............................................. 15 3.5 Prediction of nucleotide sequence for rat TMPAP variant ................................................... 16 3.6 Microarray analyses ............................................................................................................... 16 3.7 miRNA sequencing analyses of mouse SMG ......................................................................... 17 3.8 TMP and AMP histochemistry of mouse SMG ...................................................................... 17 3.9 Measurement of total salivation volume under sympathetic and parasympathetic
stimulation .............................................................................................................................. 18 3.10 Determination of total protein concentration and α-amylase activity in mouse
saliva ....................................................................................................................................... 18 3.11 Assesment of GCT and acinar cell proliferation and apoptosis ............................................ 18 3.12 Determination of cell areas.................................................................................................... 19 3.13 Cell-cyle analysis of LNCaP cells by flow cytometry .............................................................. 19
4 RESULTS .............................................................................................................................. 21
4.1 Alternative splicing of Acpp gene produces a novel PAP transmembrane isoform ............ 21 4.2 The novel TMPAP isoform is a type-I transmembrane protein with a Yxxφ motif .............. 22

4.3 TMPAP is widely expressed and the ratio between PAP isoforms changes in prostate cancer ...................................................................................................................... 24
4.4 PAP localizes to the plasma membrane and to subcellular compartments of the endosomal-lysosomal and exosomal pathway ..................................................................... 25
4.5 PAP activity accounts for 50% of the total acid phosphatase activity in wild-type saliva ....................................................................................................................................... 27
4.6 Male PAP-/- mice secrete more stimulated saliva than wild-type mice ............................... 27 4.7 TMP activity is specific for GCT cells in mouse SMG ............................................................ 29 4.8 The SMGs of male PAP-/- mice exhibit dysregulation of gene and miRNA
expression .............................................................................................................................. 29 4.9 The SMGs of male PAP-/- exhibit an increased cell proliferation rate and intense
inflammatory changes ........................................................................................................... 29 4.10 LNCaP cells overexpressing TMPAP grow at slow rate and accumulate in the G1
phase ...................................................................................................................................... 30 4.11 LNCaP-TMPAP cells show downregulation of transferrin receptor and reduced
transferrin endocytosis .......................................................................................................... 31 4.12 LNCaP-TMPAP and LNCaP-SPAP cells show increased HRP uptake and cell size ................ 31 4.13 LNCaP cells overexpressing TMPAP show dysregulated gene expression .......................... 31
5 DISCUSSION ........................................................................................................................ 33
5.1 TMPAP traffics to the lysosomes by means of its Yxxφ motif .............................................. 33 5.2 TMPAP endocytosis regulates cell cycle progression in LNCaP cells ................................... 34 5.3 PAP serves specific function in male mouse SMG and saliva ............................................... 35
6 CONCLUSIONS AND FUTURE PERSPECTIVES ....................................................................... 38
7 ACKNOWLEDGEMENTS ....................................................................................................... 39
8 BIBLIOGRAPHY .................................................................................................................... 41

i
LIST OF ORIGINAL PUBLICATIONS‡
This thesis is based on the following original publications that are referred in the main text by their Roman numerals.
I. Prostatic Acid Phosphatase is Not a Prostate Specific Target. Quintero, I. B.§, Araujo, C. L.§, Pulkka, A. E., Wirkkala, R. S., Herrala, A. M., Eskelinen, E. L., Jokitalo, E., Hellström, P. A., Tuominen, H. J., Hirvikoski, P. P., Vihko, P. T. Cancer Res. 67 (14): 6549-6554 (2007). (IF 9.0).
II. Prostatic Acid Phosphatase (PAP) is the Main Acid Phosphatase with 5'-Ectonucleotidase Activity in the Male Mouse Saliva and Regulates Salivation. Araujo, C. L., Quintero, I. B., Kipar, A., Herrala, A. M., Pulkka, A. E., Saarinen, L., Hautaniemi, S., and Vihko, P. American Journal of Physiology. Cell Physiology 306: C1017-27 (2014). (IF 4.0). Included in APSselect July 2014 and nominated for article of the year 2014.
III. Transmembrane prostatic acid phosphatase (TMPAP) delays cells in G1 phase of the cell cycle. Araujo, C. L., Quintero, I. B., Ovaska, K., Herrala, A. M., Hautaniemi, S., Vihko, P. The Prostate 76: 151-162 (2016). (IF 3.6).
‡ The original publications are reproduced in this book with the permission of the
copyright holders. § Authors with equal contribution.

ii
ABBREVIATIONS
BMP Bis-(monoacylglycero) phosphate CCP Clathrin-coated pit CCV Clathrin-coated vesicle CDR Circular dorsal ruffles cPAP Cellular prostatic acid phosphatase DEG Differentially expressed gene DRG Dorsal-root ganglia ECM Extracellular matrix EGF Epidermal growth factor EGFR Epidermal growth factor receptor ER Endoplasmic reticulum ESCRT Endosomal sorting complex required for transport FC Fold-change FtHAP Francisella tularensis histidine acid phosphatase GCT Granular convoluted tubule GFP Green fluorescent protein GPCR G-protein coupled receptor HRP Horse raddish peroxidase IEM Immuno-electron microscopy ILV Intraluminal vesicle IRG Interferon regulated genes LAP Lysosomal acid phosphatase LNCaP Lymph node carcinoma of the prostate miRNA Micro RNA MVE Multivesicular endosome NSF N-ethylmaleimide-sensitive factor PAP Prostatic acid phosphatase PAR1 Protease-activated receptor 1 PM Plasma membrane p-NPP p-Nitrophenyl phosphate PSA Prostate specific antigen QC Quality control RTK Receptor tyrosine kinase SMG Submandibular gland SNAP Soluble NSF attachment protein

iii
SNARE SNAP receptor SPAP Secreted prostatic acid phosphatase TbMBAP Trypanosoma brucei membrane-bound acid phosphatase TF Transferrin TGN Trans-Golgi network TMP Thiamine mono-phosphate TMPAP Transmembrane prostatic acid phosphatase

iv
ABSTRACT Prostatic acid phosphatase (PAP) was discovered during the mid-1930s, but the
molecular mechanisms in which this protein is involved remain poorly understood. This enzyme was originally described as a highly-expressed protein in the human prostate that was secreted to the seminal fluid. It has always been associated to prostate cancer, since high levels of acid phosphatase activity were found in the sera of patients with metastatic disease. Therefore, for 40 years, research was focused on the improvement of biochemical assays in an effort to find specific substrates for clinical application. However, in the 1980s PSA (prostate specific antigen) superseded PAP as a biomarker for early detection of the disease and became the preferred marker for diagnosis. Later, in the mid-1990s with the advent of new molecular techniques such as cloning and high-scale protein purification it was possible to obtain high-quality crystals for 3D-structural determinations of PAP. In addition, new concepts emerged concerning the physiological role of the enzyme, with renewed speculation regarding the molecular mechanisms in which this protein could be involved. The fact that the serum levels of the enzyme are increased in prostate cancer patients with metastatic disease rendered this enzyme an attractive target for immunotherapies against advanced prostate cancer. However, this hypothesis has thus far neglected the existence of any potential isoforms that could be expressed in other organs and tissues.
Therefore, as part of this thesis, the molecular mechanisms where PAP is involved will be investigated. For this purpose, two biological tools were employed: a PAP-knockout mouse model and stable virus-transfected LNCaP cell lines. A novel transmembrane type-I isoform of PAP (TMPAP) was first characterized as a product of alternative splicing of the same gene (ACPP) that encodes for the well-known secretory isoform (SPAP). TMPAP is distributed throughout mouse tissues, including prostate, lung, kidney, endometrium, salivary glands, and dorsal-root ganglia. The enzyme comprises an N-terminal domain containing the catalytic active site, a transmembrane helical domain, and a short C-terminal cytosolic domain that carries a tyrosine-based motif (Yxxφ) that targets the enzyme to the endosomal-lysosomal/exosomal pathway. This was confirmed by co-localization studies that revealed that PAP localizes to the plasma membrane as well as to the intracellular membranes of vesicles, lysosomes, and intraluminal vesicles of the multivesicular endosomes. Microarray experiments were performed on mouse tissues to study the differential gene expression profile between wild-type and PAP-knockout mice. The differential gene expression that was observed between the prostates of wild-type and PAP-knockout mice suggested that PAP is involved in secretory mechanisms, as many genes related to this process appeared dysregulated. Moreover, the results obtained from two-hybrid system experiments suggested that snapin (a SNARE-associated protein) was a likely candidate protein that could interact with TMPAP. This interaction was recently proved by co-

v
localization and florescence resonance energy transfer (FRET) studies. The PAP-knockout mice developed prostate adenocarcinoma and showed dysregulation of genes related to vesicular traffic. Consequently, this investigation was focused on murine submandibulary glands (SMG) as a model of an exocrine organ. The expression of PAP in SMG was found to be even higher than in mouse prostate. In addition to microarrays and miRNA analyses, physiological and biochemical determinations help to demonstrate that there is an increased salivation volume in PAP-knockout mice upon stimulation with secretagogue drugs. This supports the hypothesis that PAP is involved in the regulation of secretory and exocytic processes. PAP was found to account for 50% of the total acid phosphatase activity in male mouse saliva and it is expressed by the granular convoluted tubular cells of the male SMG but not by the acinar cells. Unlike prostate gland, however, the mouse SMG does not develop signs of hyperplasia or adenocarcinoma in spite of an observed increased acinar cell proliferation. This discrepancy was explained by studying the degree of lymphocyte infiltration, the dysregulation of miRNAs, and the differentially expressed genes in microarray data. In SMGs of PAP-knockout mice, the innate immune system was shown to be responsive and able to remove proliferating acinar cells, which may explain the absence of adenocarcinoma. In addition, the upregulation of anti-inflammatory molecules may prevent the extension of tissue damage. Finally, we compared the effect of the overexpression of SPAP and TMPAP in LNCaP cells with empty-vector cells. As a result, the TMPAP-LNCaP cells exhibited slower growth than SPAP-LNCaP or empty-vector cells. Cells overexpressing either SPAP or TMPAP isoform showed increased 2D-projection area and increased HRP-uptake when compared with empty-vector cells. These two observations suggested an increased vesicular traffic in endo/exocytic pathways to maintain cell membrane homeostasis. Thus, vesicles loaded with TMPAP are most likely sorted to lysosomes by means of its Yxxφ motif. Consequently, there is an increased degradation of cargo molecules—such as receptor tyrosine kinases expressed on the cell surface—that could explain the observed slow growth of LNCaP cells that overexpress TMPAP.
The molecular mechanisms identified in this study will definitely contribute to a better understanding of the physiological role of PAP in diseases and to a critical re-evaluation of existing immunotherapies. The knowledge of the molecular determinants responsible for the presence of TMPAP in the endo/exocytic pathway can also be exploited for the future development of radio-imaging and drug delivery protocols.


1
1 REVIEW OF THE LITERATURE
1.1 Prostatic acid phosphatase and prostate cancer
Prostatic acid phosphatase (PAP) is an enzyme highly expressed by the human prostate gland (Kutscher & Wolbergs 1935). It has been known in its soluble form since mid-1930s and it was associated to prostate cancer since high levels of the enzymatic activity are detected in serum of patients with metastasizing disease (Gutman & Gutman 1938). Nevertheless the molecular mechanisms behind this association remained poorly understood. One hypothesis postulates that the presence of PAP in serum is due to leakage from normal tissue due to the damage provoked by the cancerous cells. In addition, PAP activity was also detected in focal areas of bone metastasis (Bruce et al. 1980). Therefore, efforts have been focused in the development of clinical assays to determine the levels of PAP activity in serum towards the diagnosis of prostate cancer stage or recurrence. Since then the clinical laboratory methods evolved from colorimetric assays to radioimmunoassays (RIA) (Vihko 1978a; Griffiths 1983) with improved sensitivity and specificity. However, during the 1980s, the clinical use of PAP as biomarker was superseded by prostate specific antigen (PSA) that enabled the diagnostic of the disease in early stages (Stamey et al. 1987; Catalona et al. 1991). But in the last decade PSA determinations turned controversial due to the risk of over diagnosis (Draisma et al. 2003) that could lead to unnecessary treatment of patients that would never develop the disease. During the 1990s, a hypothesis emerged regarding the existence of a cytosolic cellular form (cPAP) of the enzyme. According to this hypothesis, cPAP has tumor-suppressor activity in prostate cancer cells by direct dephosphorylation of ErbB2 (an EGFR family member) (Lin & Clinton 1988; Lin et al. 1992). However, this hypothetical enzyme has never been cloned. The accumulation of results that could be better explained by a membrane associated form sparked the interest in the search for a potential alternative isoform (Vihko 1979; Kontturi et al. 1988). The alternative cDNA transcript sequence (GenBank: BC007460.1) (Strausberg et al. 2002) found in biological databases and encoding for a histidine-acid phosphatase with high degree of sequence identity to the already known soluble form of PAP, finally led to focus into the characterization of this new transcript and its product. Furthermore, a signal peptide sequence of 32 amino acids long present in the precursor of ACPP gene product that is absent in the mature form of the protein, confines the enzyme to the ER-Golgi-PM secretory route. The expression of a construct carrying a deletion of the signal peptide from hPAP sequence failed to lead to detectable levels of protein product (Hurt et al. 2012). Finally, PAP-deficient mice slowly develop prostate adenocarcinoma in a way that resembles the stages of the human disease (Quintero et al. 2013).

Review of the Literature
2
1.2 Structure of prostatic acid phosphatase
PAP is a histidine acid phosphatase characterized by the general signature motif RHGXRXP (Van Etten 1982). Other members of the family include the lysosomal acid phosphatase, the lysophosphatidic acid phosphatase, and the testicular acid phosphatase. According to the folding of the protein, PAP belongs to the phosphoglycerate mutase-like superfamily (Schneider et al. 1993). During the 1980s and 1990s, a lot of effort was focused in structural studies and determination of the enzymatic mechanism (Van Etten 1982; Buchwald et al. 1984; Vihko et al. 1993; Lindqvist et al. 1993; Schneider et al. 1993; Lindqvist et al. 1994; Porvari et al. 1994). The mass-production of recombinant protein in insect cells yielded suitable amounts of the enzyme for these crystallographic studies (Vihko et al. 1993). The active enzyme is a dimer of molecular weight ca. 100 kDa (Ostrowski & Rybarska 1965) that shows positive cooperativity upon substrate binding (Luchter-Wasylewska 2001). The active site of each subunit lies in the cleft between an α/β domain and an α-helical domain.
The reaction pocket is formed by arginine residues (Arg11 and Arg15 that belongs to the histidine acid phosphatase signature motif RHGXRXP and Arg79) (Schneider et al. 1993). These positive-charged residues provide a stabilizing environment that contributes to the positioning of the negative-charged phosphate group of the substrate. The mechanism of hydrolysis is a two-step mechanism with a first step represented by a concerted SN2 mechanism that involves the histidine residue His12 as the nucleophile to form a phosphohistidine intermediate and a second rate-limiting step where the aspartic acid residue (Asp258)—as a general acid-base catalyst—assists in the release of the alcohol molecule (Porvari et al. 1994; Sharma et al. 2008; Sharma & Juffer 2009) (Figure 1). The presence of aliphatic alcohols in the reaction media may enhance the phosphohydrolase activity of the enzyme by acting as a phosphate acceptor in a transphosphorylation reaction (Buchwald et al. 1984; Valcour et al. 1989).
The active site of PAP is big enough to accommodate different kinds of substrates as suggested by in vitro experiments (Schneider et al. 1993). The list of substrates that PAP dephosphorylates includes β-glycerophosphate, phosphorylcholine, lysophosphatidic acid, phosphoaminoacids, and nucleotides (Serrano et al. 1976; Vihko 1978b; Dziembor-Gryszkiewicz et al. 1978; Li et al. 1984; Tanaka et al. 2004). The exact nature of the physiological substrates remains elusive, however. Only the direct dephophorylation of extracellular 5’-AMP by PAP has been demonstrated to be associated to the physiological mechanisms of pain sensing (Zylka et al. 2008). Recently, it was shown that TMPAP is the same enzyme known as thiamine monophosphatase (TMPase) (a.k.a. fluoride-resistant acid phosphatase, FRAP) (Zylka et al. 2008). Strikingly, thiamine monophosphate (TMP) is a very specific substrate of PAP as already observed in other studies about

Review of the Literature
3
fluoride-resistant acid phosphatase (Knyihar-Csillik et al. 1986). During the structural characterization of rat PAP, the residues located at the entrance of the active site (Tyr123 and Arg127) were suggested to play a role in substrate specificity (Lindqvist et al. 1993; Porvari et al. 1994). These residues are not conserved in homologous lysosomal acid phosphatase (LAP) where they are replaced by lysine and glycine residues respectively (Lindqvist et al. 1993). Vanha-Perttula et al. observed that PAP and LAP exhibit differences in sensitivity to inhibition by L-(+)-tartrate ion (Vanha-Perttula et al. 1972). When the sensitivity to inhibition by L-(+)-tartrate was compared between rPAPTyr123Lys, Arg127Gly mutant and hLAP, the results indicated that both enzymes behave similarly toward concentrations of inhibitor greater than 10 mM. However, the enzymatic activity against the
substrates such as p-nitrophenylphosphate, phosphocholine, or phosphocreatine was not greatly affected. This suggested that Tyr123 and Arg127 are not the only determinants for specificity in PAP (Porvari et al. 1994).
Figure 1. Schematic representation of PAP mechanism based on Lindqvist et al. 1994.

Review of the Literature
4
1.3 Prostatic acid phosphatase as a 5’-ectonucleotidase and a thiamine monophosphatase
The 5’-ectonucleotidase activity of PAP has been known for many years (Matsuda & Ogoshi 1966; Vihko 1978b; Randerath et al. 1989), but its role in purinergic signaling has been surprisingly overlooked. On the other hand, during the 1980s there was an effort to identify the hitherto unidentified enzyme called thiamine monophosphatase (TMPase, a.k.a fluoride-resistant acid phosphatase, FRAP) that was known as a marker for nociceptive neurons in dorsal root ganglia (DRG). Due to this property, it was thought that TMPase could have a role in nociception. Nevertheless, the gene encoding TMPase was never identified. During their studies, Dodd and co-workers reached the conclusion that TMPase could be the same secreted PAP enzyme—the only known isoform of PAP at the time—after biochemical identification of partially purified TMPase from rat DRG (Dodd et al. 1983). However, later immunostaining studies using anti-PAP antibodies failed to identify PAP in the same locations where TMPase staining was known to be positive, and therefore made it impossible to assert that TMPase was indeed the secreted PAP (Dodd et al. 1983; Silverman & Kruger 1988). In 2008, Zylka et al. based on our findings, finally solved the open question and revealed that TMPase was the novel transmembrane isoform of PAP (TMPAP) (Quintero et al. 2007; Zylka et al. 2008; Taylor-Blake & Zylka 2010). Based on these two fundamental findings, subsequent rigorous studies finally established the role of the 5’-ectonucleotidase activity of PAP in nociceptive circuits (Zylka et al. 2008; Sowa et al. 2010; Hurt & Zylka 2012).
1.4 Homologous histidine acid phosphatases found in other organisms
1.4.1 Histidine acid phosphatase of Francisella tularensis (FtHAP)
The bacterial pathogen Francisella tularensis, which is the infectious agent responsible for the disease tularemia, expresses a histidine acid phosphatase known as FtHAP. Its closest structural homolog is hPAP in spite of sharing only 27% amino acid sequence identity (Singh et al. 2009). The three-dimensional structure of FtHAP was determined in complex with inorganic phosphate ion (Pi), L-(+)-tartrate ion, and 3’-AMP as substrate (Singh et al. 2009). The 3’-AMP substrate was selected from in vitro experiments for which FtHAP exhibited the highest normalized activity. Interestingly, the authors developed a trapping mutant FtHAPAsp181Ala which locks the substrate 3’-AMP in the active site. This is a well-established approach used to trap physiological substrates for protein-tyrosine phosphatases (PTPs) (Flint et al. 1997). In spite of the structural homology between both enzymes, the active site of FtHAP seems to be more compact than the active

Review of the Literature
5
site of hPAP. This difference in size accounts for the increased specificity of FtHAP against 3’-AMP. Similar to PAP, the FtHAP structure is organized as a core α/β domain and a α-helical domain that caps the core domain (Singh et al. 2009). However, the most prominent difference between the folding of both enzymes resides in the different position of the first α-helix of the cap domain. This difference can be explained by means of the disulphide bond between Cys129 and Cys340 observed in hPAP structure that locks the α-helix away the entrance of the active site (Singh et al. 2009). In addition, in FtHAP the Tyr135 residue—located at the beginning of the first α-helix of the cap domain—and the Phe23 residue form a π-stacking interaction with the adenine ring of the substrate (Singh et al. 2009). These elements that are missing in hPAP are considered fundamental to the specificity of FtHAP towards 3’-AMP (Figure 2).
1.4.2 Membrane-bound acid phosphatase of Trypanosoma brucei (TbMBAP)
Trypanosoma brucei is the parasite responsible for the African trypanosomiasis, known as sleeping sickness in humans. This microorganism expresses two genes encoding transmembrane histidine acid phosphatases, TbMBAP1 and TbMBAP2, which are 58% identical and show 40% and 41% amino
Figure 2. (A) Molecular overlay between hPAP (PDB: 2hpa, blue) and FtHAP (PDB: 3it1, orange). (B) Magnification view showing the active site sphere (translucent light-blue) with L-(+)-tartrateion inside from 2hpa structure along with the FtHAP Tyr135 and Phe23 residues (light-blue), andthe cap domains for hPAP (yellow) and FtHAP (purple). The cap domain in hPAP is locked inposition by a disulphide bridge (arrow). Based on (Singh et al. 2009).

Review of the Literature
6
acid identity to the well-characterized membrane bound acid phosphatase from Leshmania Mexicana (LmxMBAP), respectively (Engstler et al. 2005). Characterization studies revealed that 87±5% of the tartrate-sensitive acid phosphatase activity from T. brucei detergent extract corresponds to TbMBAP1. This protein shares 24% amino acid identity with human TMPAP (Engstler et al. 2005). Unlike TMPAP, the C-terminus of TbMBAP1 does not contain a tyrosine-based lysosomal targeting motif and the genes encoding the AP2 complex cannot be found in the trypanosome genome database. Therefore, this enzyme is absent in lysosomes, even in conditions of protein overexpression. The enzyme is present in recycling endosomes and it has been shown to be essential for the mainteinance of endo/exocytosis when expressed in the mammalian stage of the trypanosome (Engstler et al. 2005).
1.5 Lysosomal biogenesis and function
Lysosomes are membrane-enclosed cellular organelles at the intersection of exocytic and endocytic pathways that concentrate acid hydrolases in charge of degrading macromolecules. Therefore, two main cellular metabolic functions are attributed to lysosomes: the recycling of building blocks of material and the downregulation of cell signaling, both processes contributing to maintain cellular homeostasis (Luzio et al. 2007). In addition, lysosomal-related organelles (LROs) such as melanosomes, lytic granules, MHC class II compartments and platelet-dense granules, are specialized forms of cell-type specific lysosomes that are involved in specialized protective functions (Dell'Angelica et al. 2000). Thereby, lysosomes were also observed to play a role in other fundamental processes such as autophagy, secretion, cell death, pathogen destruction—concomitant with antigen presentation—and plasma membrane-wound healing (Wileman 2013; Blank et al. 2014; Andrews et al. 2014; Mrschtik & Ryan 2015). Lysosomes are characterized by an acidic lumen maintained at pH of 4.6-5.0 by proton-vacuolar pumping ATPases (Mellman et al. 1986). Soluble lysosomal enzymes are synthesized in the rough endoplasmic reticulum along with other secretory proteins. The lysosomal enzymes are tagged with a mannose 6-phosphate group to differentiate them from the secretory proteins. Thereafter, the tagged molecules are packed in clathrin-coated vesicles and sorted from TGN to the endosomes by a receptor-mediated mechanism (M6PR) (von Figura & Hasilik 1986). Once the complex reach an endosome, it dissociates in the acidic environment and the receptor is recycled, whereas the ligand is delivered to the lysosomes. However, other lysosomal membrane associated proteins, such as lysosomal acid phosphatase (LAP), do not require a receptor-based trafficking mechanism and they can reach the lysosomes through either a direct or an indirect pathway that involves the plasma membrane and the endocytic route (Luzio et al. 2007).

Review of the Literature
7
1.6 Function of the tyrosine-based lysosomal targeting motif Yxxφ
All cells interact with their environment by the export and import of substances through the plasma membrane via the mechanisms of endo/exocytosis. In particular, the main route of entry into the cell for many soluble molecules and membrane bound proteins is clathrin-mediated endocytosis (Sorkin & Puthenveedu 2013). Endocytosis is a fundamental mechanism, not only to import extracellular substances, but to maintain cell-membrane homeostasis and regulate cell-surface receptor signalling by internalization and sorting to degradation or recycling pathways.
The process of formation of clathrin-coated vesicles (CCVs) has been extensively studied during the last decades. Clathrin is a cytosolic soluble protein that is attached to membrane lipids by heterotetrameric adaptor proteins (AP) that bind to both clathrin and phosphatidylinositol-4,5-diphosphate [PtdIns(4,5)P2], a lipid that is enriched in the inner leaflet of the plasma membrane (Sorkin & Puthenveedu 2013). The process continues with the recruiting of accessory proteins (Slepnev & De Camilli 2000) and the polymerization of clathrin to form clathrin-coated pits (CCPs) on the intracellular side of the plasma membrane. Clathrin is a heterohexamer formed by light and heavy chains that adopts a conformation of triskelion that has the ability to form a three-dimensional lattice leading to the formation of buds and vesicles (Sorkin & Puthenveedu 2013). The heavy chains serve for structural purposes whereas the light chains provide regulatory functions. While the heavy chains are essential for the formation of vesicles (Huang et al. 2004), the light chains are not indispensable for the general process of clathrin-mediated endocytosis and it is thought that they play a role in the endocytosis of particular cargo molecules (Ferreira et al. 2012).
The interaction between specific sorting sequences present in cargo molecules and adaptor proteins that also bind to clathrin concentrates cargo molecules in CCPs. Some proteins require the binding of the ligand to promote the binding to the adaptor protein (regulated endocytosis) whereas other proteins bind to the adaptor proteins irrespective of the ligand (constitutive endocytosis). For instance, in the case of receptor tyrosine kinases, the binding of the ligand produces a conformational change with a subsequent auto-phosphorylation of cytosolic tyrosine residues that promotes the binding of adaptor and accessory proteins, such as AP2, EPS15, and epsin, among others (Traub 2009). A typical example of constitutive endocytosis is showed by the transferrin receptor (TfR) where only the

Review of the Literature
8
presence of the tyrosine-based motif (Yxxφ) is sufficient to recruit the adaptor protein (Lampe et al. 2014).
At present, the tyrosine-based endosomal-lysosomal targeting motif (Yxxφ) is one of the more studied sorting motifs (Sorkin & Puthenveedu 2013) found in many integral membrane proteins, such as lysosome-associated membrane glycoproteins (Lamp), lysosomal hydrolases (e.g. LAP), and nutrient receptors (e.g. TfR). The Yxxφ motifs bind to the μ-subunits of AP complexes (AP-1, AP-2, AP-3, AP-4 and AP-5). The different AP complexes participate in distinct intracellular routes. For instance, while the AP-1 complex is found associated to CCVs formed at TGN as part of the biosynthetic pathway, the AP-2 complex is the main adaptor protein complex found associated to CCVs that form at the plasma membrane. The AP-3 complex participates in the transport of cargo proteins in the endo-lysosomal pathway, whereas the AP-4 complex takes part in the sorting of proteins to the basolateral surface in polarized cells. The AP-5 complex has recently been described to be part of late endosome trafficking (Agrawal et al. 2013).
In spite of the advances in the identification of molecular components that take part in the clathrin-mediated endocytic process, yet unanswered questions remain to be solved. For many of these components, still unidentified post-translational modifications can add a layer of modulation to the dynamics of the process as reviewed elsewhere (Bonifacino & Traub 2003). For instance, the phosphorylation of the tyrosine residue of the Yxxφ motif in the T-cell co-stimulatory receptor (CTLA4) upon activation blocks the interaction with the μ2 subunit of the AP2 complex and inhibits the internalization of the receptor. In addition, the phosphorylated tyrosine residue recruits molecules for further signaling events. However, the phosphorylation of a serine residue at position Y-1 of the tyrosine-based motif in AQP4 enhances the interaction with μ3A subunit and lysosomal targeting of the protein (Madrid et al. 2001).
1.7 The SNARE complex and the mechanism of exocytosis
Cellular compartmentalization is a fundamental characteristic of all eukaryotic cells. By keeping molecular components such as enzymes separated by lipid membranes, cell dynamics can be controlled in a spatial-temporal fashion. Compartmentalization also precludes the enzymatic activity to occur on incorrect substrates. The communication between different compartments is carried out by the trafficking of vesicles conveying the cargo molecules. This mechanism implies that the vesicles must fuse their membrane with the plasma membrane or the membrane of the target compartment (e.g. another vesicle or organelle) in order to deliver their cargo. This membrane fusion process is driven by the SNARE (soluble

Review of the Literature
9
N-ethylmaleimide sensitive factor attachment receptor) complex (Sollner et al. 1993). SNARE proteins were identified in the late 1980s as part of the synapses in the brain (Trimble et al. 1988; Oyler et al. 1989; Inoue et al. 1992; Bennett et al. 1992). The SNARE complex is formed when some components attached to the target membrane (e.g. syntaxin and SNAP25 in neuronal cells or the ubiquitously expressed SNAP23 in non-neuronal cells) bind to the vesicle associated membrane proteins (e.g. synaptobrevin). Even when the formation of the SNARE complex is sufficient for membrane fusion (Hu et al. 2003), other proteins can participate in the modulation of the process. Two important modulators are synaptotagmins that are calcium-sensor proteins (Geppert et al. 1994) and snapin which is a SNARE-associated protein that facilitates the fusion event by enhancing the interaction between SNAP25 and synaptotagmin in neuronal cells and between SNAP23 and syntaxin in non-neuronal cells (Buxton et al. 2003).
Several mechanisms have been described to explain the dynamics of regulated exocytosis. More precisely, in neuronal cells the presence of syanptotagmin (the Ca+2-sensor protein) complexed with SNAP25 triggers the fusion event of the docked vesicle. When Ca+2 ions bind to the synaptotagmin calcium-binding domain it promotes a conformational change that brings the vesicle membrane closer to the plasma membrane (Vrljic et al. 2010). In addition, the association between SNAP25 and synaptotagmin is enhanced by PKA-dependent phosphorylation of snapin (Chheda et al. 2001). However, some integral membrane proteins, e.g. EBAG9, bind snapin resulting in inhibition of the PKA-dependent phosphorylation of snapin that results in a slowdown of the exocytic process (Ruder et al. 2005).
1.8 Disturbed endocytosis as a hallmark of cancer
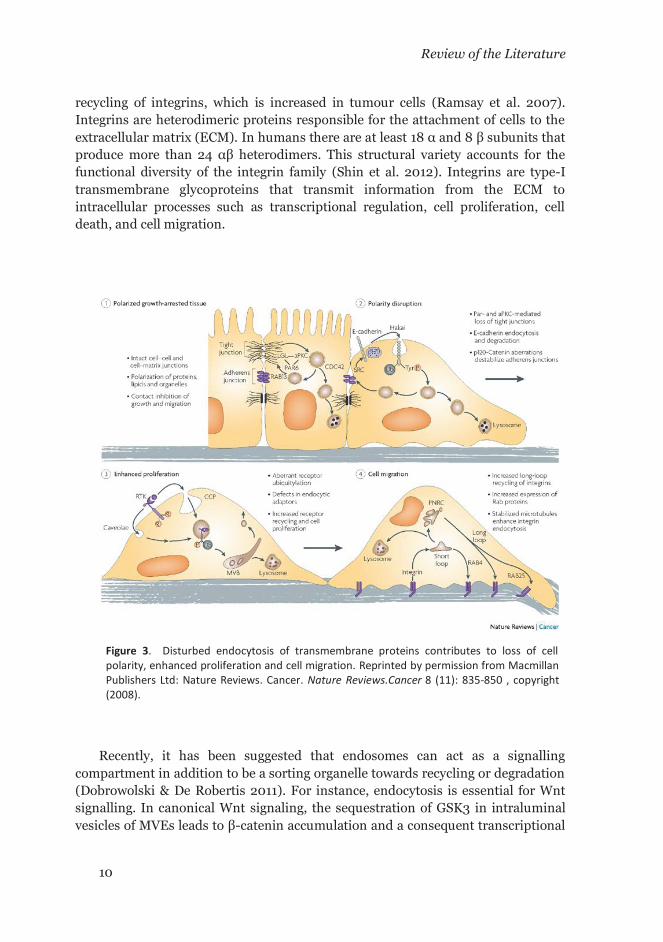
In cancerous cells, normal physiological processes are hijacked to gain survival advantage compared to their normal counterparts. Endocytosis is an important mechanism for internalization, recycling or degradation of key mitogens such as RTKs. Therefore, a dysregulation on the endocytic mechanism is expected to impact in the dynamics of RTKs downregulation and other membrane molecules. Hence it will result in a sustained propagation of intracellular proliferation signals (Figure 3). The changes in the overall appearance and dynamics of the plasma membrane suggest that dysregulated endocytosis is a hallmark of cancer (Mellman & Yarden 2013). At present, different endocytic mechanisms have been described. These include clathrin-mediated endocytosis (CME), caveolae-mediated endocytosis (CavME), clathrin- and caveolae-independent endocytosis, micropinocytosis, and circular dorsal ruffles (CDR) (Mosesson et al. 2008). All these mechanisms converge into lysosomes through several endosomal compartments. However, recycling mechanisms exist that allow internalized proteins to return back to the plasma membrane. One such example involves the
Review of the Literature
10
recycling of integrins, which is increased in tumour cells (Ramsay et al. 2007). Integrins are heterodimeric proteins responsible for the attachment of cells to the extracellular matrix (ECM). In humans there are at least 18 α and 8 β subunits that produce more than 24 αβ heterodimers. This structural variety accounts for the functional diversity of the integrin family (Shin et al. 2012). Integrins are type-I transmembrane glycoproteins that transmit information from the ECM to intracellular processes such as transcriptional regulation, cell proliferation, cell death, and cell migration.
Recently, it has been suggested that endosomes can act as a signalling compartment in addition to be a sorting organelle towards recycling or degradation (Dobrowolski & De Robertis 2011). For instance, endocytosis is essential for Wnt signalling. In canonical Wnt signaling, the sequestration of GSK3 in intraluminal vesicles of MVEs leads to β-catenin accumulation and a consequent transcriptional
Figure 3. Disturbed endocytosis of transmembrane proteins contributes to loss of cell polarity, enhanced proliferation and cell migration. Reprinted by permission from Macmillan Publishers Ltd: Nature Reviews. Cancer. Nature Reviews.Cancer 8 (11): 835-850 , copyright (2008).

Review of the Literature
11
regulation of target genes (Taelman et al. 2010). On the other hand, in the non-canonical Wnt signalling pathway, the frizzled receptor is internalized upon binding of Wnt and recruits Dishivelled2 (DVL2) which interacts with the μ2-subunit of AP2 complex and leads to activation of small GTPases Rho and Rac (Yu et al. 2007).
In particular, the endocytosis of EGFR has been extensively studied as a model of receptor—mediated endocytosis and different endocytic pathways were described for this particular RTK (Mosesson et al. 2008). After internalization, the EGFR can recycle back to the plasma membrane upon dissociation of the ligand in early endosomes (EE)—from where it can continue signaling (Dobrowolski & De Robertis 2011)—or it can follow a degradative pathway to lysosomes that requires the sorting of EGFR to the intraluminal vesicles (ILV) of MVE. This sorting mechanism is mediated by mono-ubiquitination of the receptor at the cell surface (Katzmann et al. 2002). The mono-ubiquitinated receptor interacts with the endosomal sorting complexes required for transport (ESCRTs) that along with other accessory proteins transfer the cargo to ILVs for subsequent degradation into lysosomes. However, under derailed endocytosis, the EGFR translocates to the nucleus where it can exert transcriptional regulation of gene expression. The aberrant nuclear localization of EGFR is observed in several epithelial carcinomas and determines a poor prognostic indicator (Bitler et al. 2010; De Angelis Campos et al. 2011). In addition, not all the proteins sorted into ILVs are mono-ubiquitinated. For instance, the PAR1 receptor, a G protein-coupled receptor for thrombin, is sorted to the lysosomes independently of ubiquitination. Instead, PAR1 is sorted to ILVs by binding to the adaptor protein ALIX and the ESCRT-III complex. The binding between PAR1 is favored by the adaptor protein AP3 which binds to the tyrosine-based cytosolic motif (YxxL) of the receptor (Dores et al. 2012).
1.9 Mouse submandibular salivary glands as a model of exocrine organ
In mammals, saliva is a fundamental biological body fluid that in addition to digestive enzymes contains other bioactive compounds (e.g. mucosal glycoproteins, antimicrobial components, immunoglobulins, growth factors, small peptides, and hormones) (Barka 1980). In this way, saliva plays an important role in keeping oral and digestive tract homeostasis. In animals other than humans it also contributes to wound healing through licking. In Mus musculus, the saliva also contains pheromones that mediates mate recognition when it is spread on pelage and the surroundings after grooming (Talley et al. 2001; Laukaitis et al. 2005).

Review of the Literature
12
In mammals the saliva is produced by a set of specialized major glands: the parotid, the submandibular and the sublingual glands which are innervated by the sympathetic and parasympathetic nervous systems. Additionally, the oral mucosa presents other minor glands that are disseminated throughout the oral cavity. The activation of adrenergic and cholinergic receptors associated to the sympathetic and parasympathetic nerves after stimulation produces the discharge of saliva from the glands. When both types of nerves are stimulated simultaneously, the amount of saliva discharged by the glands is greater than the corresponding individual stimulus (Proctor 1998). However, in the absence of any nerve stimulation small amounts of saliva are constitutively secreted (Proctor 1998).
Upon stimulation the secretion is mainly produced in the acini constituted by the acinar cells which are the canonical secretory units of the gland. The fluid is collected by the secretory ducts, where its composition is further changed by mechanisms of active sodium reabsorption, addition of potassium ions and bicarbonate buffering which provides a final pH ranging from 7.6 to 10.0 in C57BL/6J mice (Ma et al. 1999; Ruiz-Ederra et al. 2009). This basic pH range is essential to prevent the formation of dental caries. The final composition of saliva depends on the stimulatory input signal as revealed by the fact that the activation of α-adrenergic receptors and muscarinic receptors are responsible for the electrolyte and fluid secretion with a modest protein secretion, while the β-adrenergic stimulation produces protein secretion (Baum & Wellner 1999). In rodents, other peripheral input signals contribute to the stimulation of saliva secretion. These signals include chewing, olfaction, tasting, drinking, grooming, and heat exposure (Matsuo 1999; Proctor & Carpenter 2007).
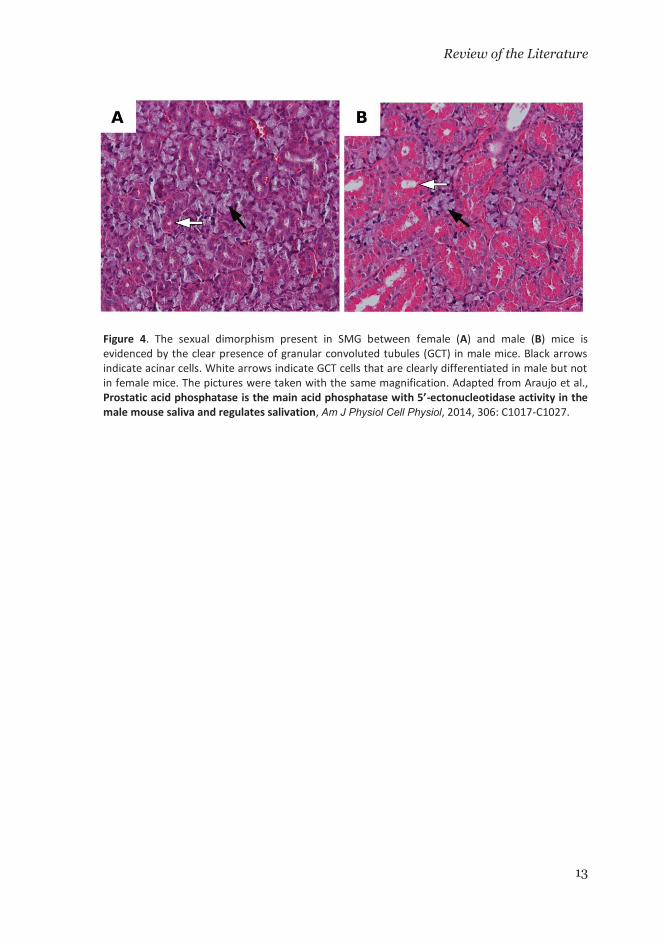
The submandibular gland (SMG) of the mice exhibits sexual dimorphism, a characteristic that is not present in humans (Junqueira & Fajer 1949; Treuting & Dintzs 2012). This sexual dimorphism is manifested by the development of the granular convoluted tubules (GCT) in male mice (Figure 4). In addition to the acini, the GCTs are exocrine structures that are fully developed after 4 weeks when they conform the ca. 19% of the gland volume in males, but only ca. 8% in female mice (Jayasinghe et al. 1990). The GCTs cells synthesize a variety of the biologically active peptides that are finally present in saliva. These bioactive molecules include nerve growth factor (NGF), epidermal growth factor (EGF), renin, erythropoietin, and kallikreins (Mori et al. 1992; Gresik 1994). In mice, the saliva secretion from SMG has proven to be more effective in wound healing through licking than the saliva from minor, sublingual and parotid glands (Bodner et al. 1991).
Review of the Literature
13
Figure 4. The sexual dimorphism present in SMG between female (A) and male (B) mice isevidenced by the clear presence of granular convoluted tubules (GCT) in male mice. Black arrows indicate acinar cells. White arrows indicate GCT cells that are clearly differentiated in male but not in female mice. The pictures were taken with the same magnification. Adapted from Araujo et al.,Prostatic acid phosphatase is the main acid phosphatase with 5’-ectonucleotidase activity in the male mouse saliva and regulates salivation, Am J Physiol Cell Physiol, 2014, 306: C1017-C1027.

14
2 AIMS OF THIS STUDY The principal focus of this doctoral investigation was to elucidate the molecular
mechanisms in which the prostatic acid phosphatase (PAP) is involved. PAP belongs to the histidine-acid-phosphatase family and, for more than 70 years, only one secreted soluble isoform (SPAP) was known.
The main goals of this study were:
1. To characterize a novel transcript of PAP enzyme.
2. To study the effects of PAP-deficiency in a mouse model at functional and gene expression levels.
3. To study the effect of the overexpression of PAP isoforms in LNCaP cells.

15
3 MATERIALS AND METHODS
3.1 Ethics statement
The animal experimentation protocols were approved by the Animal Experimentation Committee of the National Animal Experiment Board of Finland (ELLA). The work was undertaken under the project license numbers ESLH-2009-07019/Ym-23.
3.2 Mice
PAP-/- mice were generated by removing exon 3 (PAPΔ3/Δ3) of the prostatic acid phosphatase gene (Acpp), completely abolishing the expression of SPAP and TMPAP gene products (Vihko et al. 2005). PAP-/- mice have been backcrossed to the C57BL/6J strain (Harlan Laboratories) for 16 generations. They were analysed alongside age-matched C57BL/6J wild-type mice as controls. For tissue sampling, mice were killed by cervical dislocation and the SMG were immediately dissected unless otherwise specified.
3.3 Stable transfected LNCaP cells
Stably transfected LNCaP cells overexpressing TMPAP (LNCaP-TMPAP) and empty vector (LNCaP-pMX) were generated as described elsewhere (Quintero et al. 2013). The same procedure and protocol was used to generate the virus-stable transfected LNCaP cells overexpressing SPAP (LNCaP-SPAP). The transfected LNCaP cells were grown and maintained in complete RPMI-1640 [RPMI-1640 supplemented with 10% FCS, 2mM glutamine, and 100 U/ml penicillin, and 100 μg/ml streptomycin (Sigma)]. Geneticin G418 (200μg/ml) was used for selection of transformed cells. Original cell line LNCaP clone FGC was obtained from ATCC cell bank. After transfection, each cell line was stored as stock in liquid nitrogen. The cryopreserved suspensions were resuscitated after 4 months. All the cell lines were routinely tested for +PSA (prostate specific antigen) and +PAP secretion.
3.4 Bioinformatics determination of TMPAP sequence elements
Sequence alignment amongst PAP and LAP sequences of rat, mouse and human species was done with ClustalW (Thompson et al. 1994). ESPript software (Gouet et al. 1999) was used for figures. PHD tool via PredictProtein server (Rost &

Materials and Methods
16
Liu 2003) was used to obtain secondary structure elements for ESPript software. Prediction of transmembrane segment was done with TMHMM 2.0 server (Kahsay et al. 2005) and signal peptide was predicted with SignalP server (Bendtsen et al. 2004).
3.5 Prediction of nucleotide sequence for rat TMPAP variant
The nucleotide sequence for rat TMPAP variant was searched in a region of 146307 bp from NCBI reference sequence NW_047801.1 by pattern matching implemented in a Perl script using a regular expression (G...{117}(TGA|TAG|TAA))—to
match the region coding the C-terminus of the mouse variant—after the portion of the sequence that matches the first methionine and the following four amino acids (ATGAGAGCTGTCCCT). Matching sequences that contain an earlier stop codon or gaps by matching the regular expression pattern ^(G..)(...)*(TGA|TAG|TAA|NNN)(...)*(TGA|TAG|TAA)$
were disregarded. The obtained segments were translated to protein code and compared with the C-terminus of the mouse sequence.
3.6 Microarray analyses
Total RNA was isolated from SMG of two-month-old male PAP-/- and wild-type mice (8 per group), cut into four equal pieces and stored in RNA stabilization reagent (RNAlater, Qiagen). Each tissue fragment was lysed with Lysing Matrix D ceramic beads in LT buffer (Qiagen) with 2-mercaptoethanol (Sigma), using a FastPrep FP120 homogenizer (Thermo Savant). The four lysates were pooled and total RNA isolated with the RNeasy Midi kit (Qiagen) according to the manufacturer’s instructions. The microarray experiments were performed at the Functional Genomics Unit (FuGU), Biomedicum Helsinki by Two-Color Microarray-Based Gene Expression Analysis. Labeling and hybridization were performed according to the manufacturer´s instructions. Combined experimental and reference samples were mixed with blocking agent, fragmentation, and hybridization buffer, and hybridized to Agilent Mouse Genome Microarray 4x44K in dye-swap configuration, following the manufacturer’s instructions. Microarrays were washed and scanned with an Agilent Scanner, using protocols provided by the manufacturer. Feature Extraction software was used for image analysis. All control and saturated spots from the microarray data were excluded. The data were then subjected to linear normalization to allow comparison between arrays. Differentially expressed genes (DEGs) between PAP-/- and wild-type replicate samples were calculated using the median fold change (FC) value of the replicate pairs. DEGs with Benjamini and Hochberg FDR corrected p-value cut-off 0.05 were selected for further analyses (Ovaska et al. 2010; Development Core Team 2014).

Materials and Methods
17
Ontological analyses were performed using GoMiner software (Zeeberg et al. 2003), and ontological groups with “p-value changed” less than 0.05 were considered as significant. Genes were considered significantly downregulated or upregulated with FC cut-off 1.5. Network analyses were made with GeneMANIA software (Warde-Farley et al. 2010). The INTERFEROME database (Rusinova et al. 2013) was used to search for interferon regulated genes in DEGs list. The database was queried with parameters for in vivo experiments in Mus musculus with FC cut-off 1.5.
3.7 miRNA sequencing analyses of mouse SMG
Total RNA was extracted from frozen 2-month-old male mouse SMG samples (4 animals per genotype) stored in RNAlater, using the miRNeasy kit (Qiagen) following the manufacturer’s protocol and stored at -70°C until further processing. Then, 1.0 μg of RNA was used for preparation of smallRNA libraries according to the user´s guide of the TruSeq Small RNA kit (Illumina). Library quality control was evaluated with an Agilent Bioanalyzer (Agilent Technologies). Caliper LabChipXT (PerkinElmer) was used for enrichment of miRNA containing 147 bp ± 5% fraction from the smallRNA library amplification products. This fraction contains mature microRNAs generated from approximately 22 nt RNA fragments. The Illumina HiSeq platform was used for 50 bp PE sequencing of miRNA enriched libraries. The miRseq data were analyzed by the Institute for Molecular Medicine Finland (FIMM) miRseq analysis pipeline with the main inputs being metric plots and expression data. The FIMM miRseq analysis pipeline is a modified version of the miRNA analysis and expression profiling pipeline, E-miR, originally developed by Buermans et al. 2010. The Bowtie (Langmead et al. 2009) mature transcript per million (tpm) table was filtered for significantly (p-value < 0.05) dysregulated miRNAs by carrying out a t-test comparison between PAP-/- and wild-type Bowtie tpm. Fold-changes were calculated from the ratio between PAP-/- and wild-type mean values.
3.8 TMP and AMP histochemistry of mouse SMG
Wild-type and PAP-/- 2-month-old mice (3 per group) were anesthetized with Mebunat Vet (60 mg/ml, Orion Pharma) and intracardially perfused with phosphate buffer pH 7.4 (PB), followed by 4% PFA (paraformaldehyde) prepared in PB. SMG were post-fixed for 2 h in 4% PFA, then cryoprotected in 30% sucrose in PB. SMG were embedded in OCTTM (Tissue-Tek, Sakura). From these, 30 μm cryosections were prepared. Samples were assayed for PAP 5’-ectonucleotidase activity with TMP, AMP, ADP, and ATP substrates (Sigma-Aldrich) according to Knyihar-Csillik’s modified protocol (Knyihar-Csillik et al. 1986). Briefly,

Materials and Methods
18
cryosections were dried for 15 min at 37°C, washed twice with 0.04 M Trizma-Maleate buffer (TMB) pH 5.6 and once with 8% (w/v) sucrose in TMB. The slides were then incubated for 2 h at 37°C in a solution containing 0.25% (w/v) substrate (TMP, AMP, ADP or ATP), 8% (w/v) sucrose, and 0.08% (w/v) Pb(NO3)2 in TMB. Subsequently, slides were washed for 1 min in 2% acetic acid, followed by 3 washes with TMB and development for 10 sec in 1% Na2S in TMB. Sections were then washed in H2O, dehydrated in an alcohol gradient and xylene, and mounted with Pertex (HistoLab Products AB).
3.9 Measurement of total salivation volume under sympathetic and parasympathetic stimulation
Two-month-old wild-type (17 males, 9 females) and PAP-/- (15 males, 8 females) mice were weighed and then anaesthetized by intraperitoneal injection of a mixture of 10 mg/kg xylazin (Rompun® Vet 20 mg/ml, Bayer) and 36 mg/kg ketamine (Ketalar 50 mg/ml, Pfizer Oy) in sterile isotonic saline solution. After anesthetization, animals were intraperitoneally injected with a cocktail of secretagogues containing 2 mg/kg of (-)-isoproterenol hydrochloride (Sigma-Aldrich) and 0.5 mg/kg of pilocarpine hydrochloride (99% titration, Sigma-Aldrich). Five min after injection, the total saliva was collected in Eppendorf tubes on ice for 10 min, using a peristaltic pump at constant speed. The volume was determined with a micropipette. Samples were stored at -20°C for further analyses. The animals were assayed in small groups (4 to 5 animals) at the same time of the day to avoid differences in the salivation properties due to circadian rhythm.
3.10 Determination of total protein concentration and α-amylase activity in mouse saliva
Total protein concentrations were measured in the saliva samples collected from stimulated mice with the Pierce BCA Protein Assay kit (Thermo) following the manufacturer’s instructions. The enzymatic assay to evaluate the α-amylase activity was performed, following a modified Sigma A3176-SSSTAR01 protocol adjusted to microscale.
3.11 Assesment of GCT and acinar cell proliferation and apoptosis
SMG of male PAP-/- and wild-type mice, aged 3, 6, and 12 months (4 mice per group) were fixed in formalin and the specimens were embedded in paraffin. To assess the degree of cell proliferation and apoptosis, sections (5 μm) were prepared,

Materials and Methods
19
deparaffinized, rehydrated and stained. To detect proliferating cells, a rabbit polyclonal anti-Ki67 antibody (AB15580, Abcam) and the Vectastain Elite ABC Kit (Vector Laboratories) with hematoxylin counterstaining were used. To detect apoptotic cells, terminal deoxynucleotidyl transferase–mediated dUTP nick end labeling (TUNEL) staining was conducted, using the FragEL DNA Fragmentation Detection Kit (Calbiochem) following the manufacturer’s instructions, and hematoxylin counterstaining. Ki67- and TUNEL-positive cells were quantified as follows: the sections were photographed with a Labovert FS microscope (magnification 40x), 10 random fields were captured for each section from non-inflammatory areas. In each field, the total number of acinar and GCT cells, as well as the number of Ki67- and TUNEL-positive acinar cells, were counted. The acinar and GCT cells were discriminated based on their morphological differences that were readily identifiable under experimental conditions, without need for additional staining. The ratio of positive cells to the total amount of cells per sample was compared between PAP-/- and wild-type groups for each marker, using two-sample t-test for equality of proportions with continuity correction for each age group, as implemented in the R statistical package version 2.10.1.
3.12 Determination of cell areas
One frame from the IncuCyte videos taken from the plates containing 2500 or 5000 cells/cm2 was extracted at 48h and the cell areas were measured using CorelDraw X6 software. The images were kept at the same dimensions for comparison. Polyline contours were manually drawn around cells avoiding round cells or cells clearly in division. The area value was then taken from the ‘area’ attribute of the polyline object in square-inches (in.2) and converted to μm2 using the scale bar on the pictures. The differences in mean values were compared for statistical significance by Mann-Whitney test.
3.13 Cell-cyle analysis of LNCaP cells by flow cytometry
LNCaP-TMPAP, -SPAP or -pMX cells were seeded at a density of 15000 cells/cm2. Cells were then incubated for 24, 48, 72 or 96 h and a pulse with BrdU (final concentration 15 μM) for 60 min was given to the cells prior to trypsinization. Cells were washed with PBS and fixed with 70% ethanol. For the flow cytometry stain, cells were first washed with PBS, permeabilized in 0.1% Triton X-100 in PBS, and their DNA was denatured with 1 M HCl for 30 min. After DNA denaturation, the cells were washed twice with 1% BSA in PBS and incubated 30 min antiBrdU-FITC antibody (BD347583). The cells were washed again with 1% BSA in PBS and finally re-suspended in 5 μg/ml propidium iodine (Sigma P4864) solution in PBS. Samples were assessed in a BD AccuriC6 flow cytometer. Flow cytometry data files

Materials and Methods
20
in FCS format were exported to text file format using FCSTrans v1.3 conversion utility (Qian et al. 2012) and the resulting text files were imported into TOPCAT v4.1 software (Taylor 2005; Taylor 2006; Taylor 2009) for cell cycle analysis. The differences in mean values were compared for statistical significance by two-tailed Student’s t-test.

21
4 RESULTS
4.1 Alternative splicing of Acpp gene produces a novel PAP transmembrane isoform
A high degree of homology was found between the cDNA sequence for the mouse TMPAP variant and the PCR products obtained from total RNA derived from human, mouse and rat tissues. The rat cDNA (GenBank accession no. DQ826426) exhibits 91% and 81% of identities with reported cDNA sequences of mouse (NM_207668) and human (BC007460) PAP, respectively (Roiko et al. 1990; Strausberg et al. 2002). The analysis of the exon-intron junctions suggests that PAP variants originate from alternative splicing in rat, mouse and human. The position of the splicing of the 10th intron is similar at the end of the 10th exon. This splicing leads to the secreted variant mRNA in rat and mouse, and to the transmembrane variant in human. In mouse and rat, the splicing of the 10th intron, the 11th exon, and the 11th intron yields the transmembrane variant. In human, the
secreted variant mRNA originates as a consequence of the absence of intron 10 and hence the open reading frame continues over the splicing site until the stop codon (Figure 5). The long isoform of the rat PAP was absent from biological databases; therefore a custom search was made of the rat genome. The nucleotide sequence
Figure 5. Schematic representation of alternative splicing in Acpp gene for human, mouse and rat. Vertical red lines represent stop codons.

Results
22
encoding the long isoform in the rat genome was found by using regular expression matching. Just one out of the 212 amino acid sequences encoded by the matched regular expression pattern exhibited 100% amino acid identity with the C-terminal part of mouse transmembrane isoform.
4.2 The novel TMPAP isoform is a type-I transmembrane protein with a Yxxφ motif
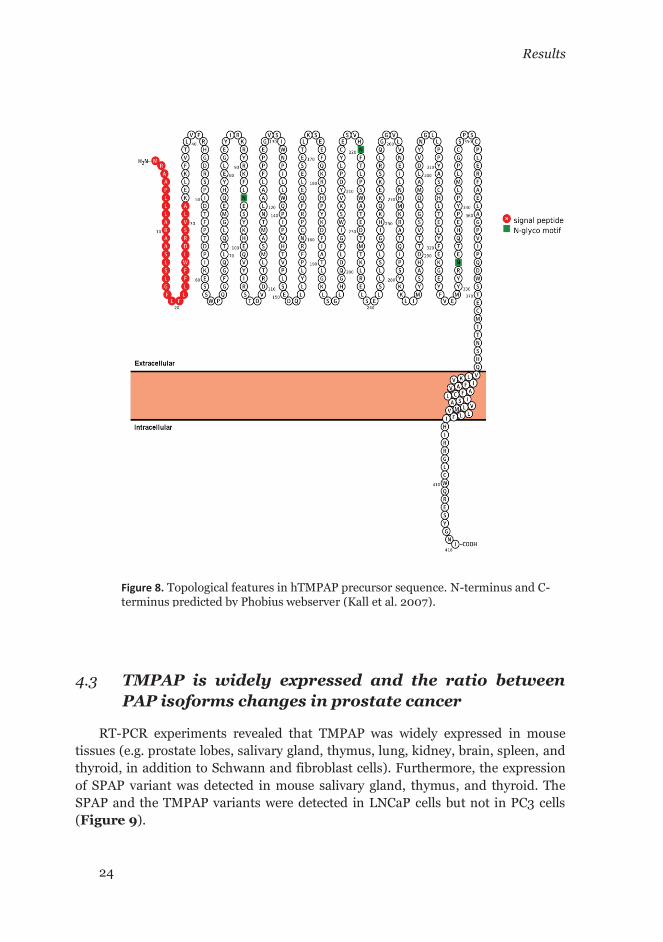
Sequence analysis of TMPAP revealed the presence of a transmembrane segment at the C-terminus followed by a short cytosolic tail. The arrangement between the first 32 amino acids (signal peptide) and the transmembrane segment defines the topology of the TMPAP isoform as a transmembrane type-I protein. Hence, the N-terminal domain carries the catalytically active site and the C-terminus comprises a short cytosolic tail with a tyrosine-based motif (Yxxφ) at the end (Figure 7 and Figure 8). The Figure 6 shows the comparison of the C-terminus sequence between PAP and lysosomal acid phosphatase LAP. The Figure 7B shows phospho-serine/threonine and phospho-tyrosine motifs predicted in the short cytosolic tail of TMPAP.
Figure 6. Comparison of C-terminal sequence of hPAP and hLAP toward protease activity. The different fragments generated by proteases acting on hLAP are showed in different colors.

Results
23
Figure 7. (A) Alignment of PAP isoform sequences from human, mouse and rat. Only the N- andC-terminus of TMPAP and SPAP isoforms are shown. Red arrowheads indicate potential phosphorylation sites of phosphobinding motifs detailed in B. Adapted by permission from theAmerican Association for Cancer Research: Quintero et al., Prostatic Acid Phosphatase Is Not aProstate Specific Target, Cancer Research, 2007, 67: (14), 6549-54. (B) Serine/threonine andtyrosine motifs recognized by kinases, phosphatases and binding proteins (unpublished).
Results
24
4.3 TMPAP is widely expressed and the ratio between PAP isoforms changes in prostate cancer
RT-PCR experiments revealed that TMPAP was widely expressed in mouse tissues (e.g. prostate lobes, salivary gland, thymus, lung, kidney, brain, spleen, and thyroid, in addition to Schwann and fibroblast cells). Furthermore, the expression of SPAP variant was detected in mouse salivary gland, thymus, and thyroid. The SPAP and the TMPAP variants were detected in LNCaP cells but not in PC3 cells (Figure 9).
Figure 8. Topological features in hTMPAP precursor sequence. N-terminus and C-terminus predicted by Phobius webserver (Kall et al. 2007).

Results
25
When the relative expression of both isoforms in human prostate tissue was studied, both PAP variants were detected in human benign prostatic hyperplasia (BPH) and well-differentiated prostate cancer specimens (Figure 9). Evaluation of the relative abundance of PAP variants in those specimens by qRT-PCR showed the average level of TMPAP transcript variant to be 0.5 ± 0.26 in BPH and 0.45 ± 0.34 in prostate cancer, while the values corresponding to the SPAP transcript variant were 0.60 ± 0.3 and 0.31 ± 0.33, respectively. The expression of both variants was observed in all the specimens, but the expression of the secreted variant was significantly downregulated in prostate cancer (P < 0.05).
4.4 PAP localizes to the plasma membrane and to subcellular compartments of the endosomal-lysosomal and exosomal pathway
The subcellular localization of PAP was studied by immunofluorescence and immunoelectron microscopy in cell and tissue samples. The results indicated that, in tissue specimens, PAP localizes to vesicles located both at the basal and apical cytoplasm, to the lumen of the glands, and to bis-(monoacylglycero) phosphate (BMP) containing vesicles (Figure 11A). At higher resolution, IEM results showed
that PAP localizes to the limiting and internal membranes of apical, electron-luscent vesicles, as well as to the internal membranes of multivesicular endosomes. In agreement with the observations made by immunofluorescence labeling, PAP co-localized with BMP in the same structures (Figure 11C). PAP was also observed in membranous structures in the lumen of the gland (Figure 11B, left). The results in cells showed a similar distribution of PAP in LNCaP cells where PAP was observed in endosome-like vesicles and lysosomes (Figure 12A). In the mouse
Figure 9. Expression of TM-PAP in different mouse cells and tissues, human prostate cancer cells, and benign prostatic hyperplasia (BPH) and prostate cancer (PC) patient samples. Reprinted by permission from the American Association for Cancer Research: Quintero et al., Prostatic Acid Phosphatase Is Not a Prostate Specific Target, Cancer Research, 2007, 67: (14), 6549-54.

Results
26
Schwann cells, PAP was observed in the plasma membrane segments and filopodia-like structures (Figure 12B) where as in the skeletal muscle fibers PAP expression was present in the sarcolemma (Figure 10A). Furthermore, the expression distribution of a TMPAP-GFP construct was analyzed by confocal microscopy in PC3 cells—a prostate cancer cell line that do not express endogenous PAP. The TMPAP-GFP was observed in the plasma membrane and in intracellular vesicles (Figure 10B). In empty-vector transfected cells, the signal was evenly distributed throughout the cell. In addition, PAP was observed to co-localize with both lysosomal-associated membrane proteins (LAMP1 and LAMP2), as well as with flotillin-1 [a protein commonly associated to lipid rafts, lysosomes, maturing phagosomes, early endosomes and exosomes (Otto & Nichols 2011)] in plasma membrane and intracellular vesicles (Figure 10, C and D).
Human LAP precursor is processed in lysosomes by a sequence of two proteolytic events resulting in the trapping of soluble LAP into the lumen of the lysosomes (Gottschalk et al. 1989). A comparison between hLAP and hTMPAP amino acid sequences reveal no amino acid identities at the protease cleavage sites or their neighboring residues (Figure 6).
Figure 10. (A) TM-PAP localization in sarcolemma of human skeletal muscle detected by immuno-histochemistry. (B) Localization of TM-PAP-GFP in plasma membrane and vesicles of PC-3 cells after transfection. (C and D) Co-localization of PAP with different cell markers in LNCaP cells: PAP co-localized with flotillin-1 in small vesicles and in the plasma membrane (C), PAP co-localization with lysosomal associated membrane protein-2 (LAMP-2; D). White arrows, co-localization sites (yellow). Reprinted by permission from the American Association for Cancer Research: Quintero et al., Prostatic Acid Phosphatase Is Not a Prostate Specific Target, Cancer Research, 2007, 67: (14), 6549-54.

Results
27
4.5 PAP activity accounts for 50% of the total acid phosphatase activity in wild-type saliva
When PAP activity was measured in male C57BL/6J saliva, as the tartrate-sensitive fraction of the total acid phosphatase activity, the total acid phosphatase activity against p-NPP was reduced ca. 50% in the presence of tartrate ion (Article II: Figure 1A). This effect remains undetected in saliva samples of PAP-/- male mice or wild-type female mice.
4.6 Male PAP-/- mice secrete more stimulated saliva than wild-type mice
Figure 11. PAP and BMP co-localization in human prostate cancer tissue. (A) PAP was localized in vesicles both in the basal and apical cytoplasm, and strong labeling was observed in the lumen of the glands. PAP showed an almost complete co-localization with BMP (green, PAP; red, BMP; blue, nuclei; yellow, co-localization). (B) In addition, PAP labeling was observed in the membranous structures in the lumen. PAP (10-nm gold) co-localized with BMP (5-nm gold; arrowheads). Some BMP labeling was also observed in the endosomes (right, arrowheads). (C) PAP localized in the limiting and internal membranes of apical, electron-lucent vesicles (black arrow). PAP was also observed in the lumen of multivesicular endosomes. AJ, adherent junction; L, lumen. Adapted by permission from the American Association for Cancer Research: Quintero et al., Prostatic Acid Phosphatase Is Not a Prostate Specific Target, Cancer Research, 2007, 67: (14), 6549-54.

Results
28
The presence of TMPAP in endosomal-lysosomal/exosomal pathway, and the observed interaction of TMPAP with snapin, led to the hypothesis that TMPAP is involved in endo/exocytic processes. Therefore, to indirectly test this, the total salivation volume (normalized by body weight) was measured in anesthesized male and female mice upon stimulation with secretagogues. The results indicated a small but statistically significant difference between controls and PAP-/- male mice with PAP-/- mice secreting a higher volume of saliva (P = 0.03, n = 15) (Article II: Figure 1B). However, no modification of other measured parameters was observed, such as; total protein concentration or α-amylase activity (Article II: Figure 1, C and D). A non-statistically significant increase was seen in the salivation volume of female PAP-/- mice compared to wild-type counterparts. No differences in total protein concentration nor α-amylase activity were evident between genotypes either. In addition, no statistically significant differences in body weight were observed between wild-type and PAP-/- genotype groups of each sex.
Figure 12. PAP localization in LNCaP and SW10. (A) In LNCaP cells, PAP was detected in caveosome-like structures (f100 nm; left), small endosome-like vesicles (f65 nm; middle), and lysosomes (right). Left, gold labeling was enhanced by silver just enough to make the nanoparticles visible (arrows), to avoid any masking of the structures by the precipitate; middle and right, a stronger signal was obtained by longer enhancement period. (B) PAP in isolated mouse Schwann cells. PAP was localized in the plasma membrane domains and filopodias. Reprinted by permission from the American Association for Cancer Research: Quintero et al., Prostatic Acid Phosphatase Is Not a Prostate Specific Target, Cancer Research, 2007, 67: (14), 6549-54.

Results
29
4.7 TMP activity is specific for GCT cells in mouse SMG
In order to compare the levels of acid phosphatase activity in SMG cryosections between wild-type and PAP-/- male mice, TMP, AMP, ADP, and ATP were used as substrates. TMP was confirmed as a specific substrate for PAP since staining was only observed in GCT cells of SMG wild-type samples (Article II: Figure 3, A and C), but it was completely absent in PAP-/- specimens (Article II: Figure 3, B, and D). However, 5’-ectonucleotidase activity against AMP was observed in both GCT and acinar cells of wild-type samples (Article II: Figure 3, E and G), and only remains in acinar cells of PAP-/- specimens (Article II: Figure 3, F and H). No activity was observed against ADP nor ATP substrates in any of the specimens under experimental conditions.
4.8 The SMGs of male PAP-/- mice exhibit dysregulation of gene and miRNA expression
To determine the molecular changes associated to the observed functional and morphological changes in SMG of PAP-/- mice, an initial microarray screening was carried out comparing the SMG of 2-month-old male PAP-/- and wild-type mice. The ontological analyses of significant DEGs showed an enrichment of upregulated genes associated with cell proliferation (e.g. Mki67, Aurkb and Birc5) along with immune response processes (e.g. Irf7, Cxcl9, Ccl3, Ccl4, Ccl8 and Fpr2) in PAP-/- mice (Article II: Figure 4 and Table 1). These findings were also confirmed by other methods. At first, qRT-PCR analyses revealed changes in expression of Irf7 and Cxcl9 in good agreement with microarray data (FC = 4.9, P = 0.001, n = 7 and FC = 2.5, P = 0.002, n = 7, respectively). Moreover, a query of the INTERFEROME database (Rusinova et al. 2013) with the list of DEGs showed that over 40% of the genes correspond to interferon regulated genes (IRGs). A co-localization network of DEGs also shows a clear separation into two clusters defined by positive regulation of cell cycle processes and innate immune response (Article II: Figure 4).
Subsequently, differential miRNA expression in the SMG of PAP-/- and wild-type male mice was studied using a next-generation sequencing platform. The results indicated that miR-146a is the most significantly upregulated miRNA (FC = +1.84).
4.9 The SMGs of male PAP-/- exhibit an increased cell proliferation rate and intense inflammatory changes
To corroborate the observed upregulation of genes associated with cell proliferation in the microarray data, the proliferative and apoptotic status of acinar

Results
30
and GCT cells in SMG of the mice was evaluated. For this purpose, in situ staining was performed with an antibody against Ki67—a marker of proliferation that shows upregulated transcriptional level in the microarray. The results indicated a significant increased proliferation of acinar cells in 6- and 12-months-old PAP-/- mice (P < 0.001, n = 4) compared with wild-type mice. Similarly, the apoptotic status of the cells was determined by counting the number of positive cells in TUNEL-assays. No differences were observed regarding the degree of apoptotic cell death between any of the groups at any age (Article II: Figures 5, A and B). A similar analysis was conducted to evaluate the proliferative and apoptotic status of GCT cells, demonstrating a significantly higher proliferation rate in the GCT cells of 6-months-old PAP-/- mice (P < 0.001, n = 4). However, the significance disappeared at the age of 12-months (Article II: Figure 5C). Moreover, at this age, the ratio between the number of acinar cells and the total amount of acinar and GCT cells was significantly decreased (P < 0.001, n = 8) in PAP-/- mice, but remained constant in wild-type animals (Article II: Figure 6). In addition, in order to corroborate the observed upregulation of immune response genes in SMG of PAP-/- mice, the degree and type of inflammatory infiltration in the SMG of the 3-, 6- and 12-months-old wild-type and PAP-/- male-mice were compared. A significantly higher degree of lymphocyte infiltration was observed in PAP-/- mice at the age of 3 months (average T cell score PAP-/-: 2.0, wild-type: 0.75, P = 0.02, n = 4). At the age of 6-months, the average T-cell score was still higher but not statistically significant (average T cell score PAP-/-: 4.25, wild-type: 3.0, P = 0.06, n = 4). At 12-months of age, the lymphocyte infiltration was similar in both intensity and distribution between genotypes (average T cell score PAP-/-: 4.75, wild-type: 4.5), which represented a further increase in the degree of inflammation in wild-type mice, but an almost stagnant degree in the PAP-/- mice. The results are summarized in Article II: Figures 5, 6 and 8.
4.10 LNCaP cells overexpressing TMPAP grow at slow rate and accumulate in the G1 phase
To further investigate the molecular mechanisms associated with cell cycle progression, flow cytometry was employed to analyze LNCaP cells that overexpress SPAP, TMPAP, or pMX-empty vector. The results indicated that at 48 h after plating, a significantly higher proportion of LNCaP-TMPAP cells (73.7%, P < 0.001) accumulated at G1 phase of the cell cycle when compared with LNCaP-pMX (61.3%, P < 0.001) and LNCaP-SPAP (57.6%, P < 0.01) cells (Article III: Figure 1).

Results
31
4.11 LNCaP-TMPAP cells show downregulation of transferrin receptor and reduced transferrin endocytosis
In order to corroborate if the observed downregulation of transferrin receptor gene expression (TFRC-1.66) is also manifested at the functional level, labeled-transferrin (TF) uptake experiments were performed. LNCaP-TMPAP cells exhibited a clear reduction of TF endocytosis when compared with LNCaP-SPAP cells. This was demonstrated by a significant reduction of TF bound (B) to the plasma membrane of LNCaP-TMPAP cells compared to LNCaP-SPAP (P = 0.006, n = 15) or LNCaP-pMX (P = 0.04, n = 15) at +4°C. After the cells were incubated at +37°C for 30 min, the amount of TF (B+E) remained significantly lower in LNCaP-TMPAP cells compared with LNCaP-SPAP (P = 2.2×10-5, n1 = 15, n2 = 12, respectively) or LNCaP-pMX (P = 0.03, n = 15). The results are summarized in (Article III: Figure 2 and Table 1).
4.12 LNCaP-TMPAP and LNCaP-SPAP cells show increased HRP uptake and cell size
The overexpression of SPAP or TMPAP is hypothesized to lead to an increase in endo/exocytic processes that would manifest as differences in cell area as a consequence of the insertion and removal of vesicular membranous material. The cell surface area approximately measured from a 2D-projection area is in good agreement with this hypothesis. As expected, both LNCaP-SPAP (828±80 μm2) and LNCaP-TMPAP (767±97 μm2) cells are significantly bigger in size compared with empty-vector transfected cells (589±20 μm2) (P=7.7×10-10, n1=46, n2=175 and P=3.6×10-3, n1=66, n2=175, respectively) (Article III: Figure 4).
4.13 LNCaP cells overexpressing TMPAP show dysregulated gene expression
Differences in cell growth were evident between LNCaP-TMPAP cells and LNCaP-pMX cells, but not between LNCaP-SPAP cells and LNCaP-pMX cells. Therefore, microarray analysis was conducted in order to identify dysregulated genes that could explain the observations. The ontological analyses of DEGs revealed that the overrepresented ontological groups related to cell cycle included a large group of downregulated histone genes from families H1, H2A and H2B (Article III: Supl. tables 1 and 2). Other DEGs observed in microarray data were the transferrin receptor TFRC(-1.66) and the adenosine receptors ADORA1(-1.47), ADORA2A(-1.91), ADORA2B(-2.05) and ADORA3(-1.59). The microarray data were

Results
32
validated by qRT-PCR on selected genes and the results showed good agreement with the observed expression values in microarrays: H2AFJ (FC=-8.17, P=0.002), CDK6 (FC=-4.45, P=0.001), ADORA2A(FC=-1.84, P=0.02), IGFBP3 (FC=+4.22, P=0.0003) and TFRC (FC=-1.47, P=0.02).

33
5 DISCUSSION Prostatic acid phosphatase (PAP) has been known for 70 years in its soluble
secreted form as an enzyme that is highly expressed in human prostate. This led to the conception that it is a prostate-specific enzyme up to the point that it could be considered a prostate-specific antigen suitable for the development of immunotherapies against prostate cancer (Kantoff et al. 2010; Cheever & Higano 2011). The existence of an extracellular soluble isoform of PAP led some researchers to postulate the hypothesis of the existence of a cytosolic isoform (cPAP) derived from the soluble form and capable of dephosphorylating the ErbB2 receptor (Meng & Lin 1998; Veeramani et al. 2005; Chuang et al. 2010). However, during the course of this investigation, a novel transmembrane type-I isoform of PAP (TMPAP) was identified that localizes to the plasma membrane and to the endosomal-lysosomal pathway and consequently it fails to match the characteristics of cPAP, which was never sequenced. Therefore, animal and cell models were characterized here in order to establish a functional molecular framework to discuss the molecular context in which TMPAP should be considered in future research.
5.1 TMPAP traffics to the lysosomes by means of its Yxxφ motif
Soluble lysosomal hydrolases are delivered to the lysosomes by a direct route from the TGN and involves the M6PR pathway (von Figura & Hasilik 1986). However, membrane-associated hydrolases such as LAP are delivered to the lysosomes through an indirect pathway that involves the plasma membrane and the clathrin-dependent endocytic route (Braun et al. 1989). In the case of LAP, it recycles through the plasma membrane and the lysosomes several times until it gets trapped into the lumen of the lysosomes by a proteolytic cleavage (Gottschalk et al. 1989). The topological arrangement of signal peptide and transmembrane segment of TMPAP precursor resemble those of the lysosomal acid phosphatase (LAP) and lysosomal associated-membrane glycoproteins (LAMPs). The presence of the Yxxφ motif is sufficient for lysosomal localization of proteins when additional constraints are met. These constraints include the distance between the tyrosine residue and the TM segment (Rohrer et al. 1996), the localization of the tyrosine-based motif at the C-terminal end of the sequence (Bonifacino & Traub 2003), and the nature of the bulky hydrophobic residue (Gough et al. 1999). In TMPAP, all of these additional constraints are satisfied. Other endocytic cargoes with a tyrosine-based motif that are not destined to lysosomes—e.g. transferrin receptor—carry an inner Yxxφ motif (Bonifacino & Traub 2003).

Discussion
34
The cellular localization studies confirmed that TMPAP-GFP is present in the plasma membrane and intraluminal vesicles of MVE (Article I), as well as the exosomes (Quintero et al. 2013), whereas the signal was evenly distributed throughout the cell in the empty-vector transfected cells. In addition, TMPAP-GFP co-localized with LAMP2 protein which is a typical marker for lysosomes (Article I). An illustration of the proposed mechanism for the intracellular trafficking of TMPAP is depicted in Figure 13.
5.2 TMPAP endocytosis regulates cell cycle progression in LNCaP cells
The overexpression of TMPAP or SPAP was studied in LNCaP cells in order to discriminate between the effects on endo/exocytosis produced by each isoform. Consequently, the overexpression of TMPAP but not SPAP produces an increased endo/exocytosis with a concomitant delay at the G1/S phase of the cell cycle. Changes at the transcriptional level were observed, in line with observations. LNCaP-TMPAP cells show differences in cell cycle progression that are independent of their shared 5’-ectonucleotidase activity; both PAP isoforms carry
Figure 13. A proposed trafficking mechanism for TMPAP. Ado: adenosine, TGN: trans-Golgi network, P: phosphate group, AP-2: adaptor protein complex 2, AC: adenylate cyclase, G-prot: trimeric G-protein, PKA: protein kinase A, AMP: adenosine monophosphate, cAMP: cyclic adenosine monophosphate, VDCC: Voltage-dependent calcium channel. Green dotted-lines represent clathrin-coat. Adapted from Quintero, IB et al. Transmembrane Prostatic Acid Phosphatase (TMPAP) Interacts with Snapin and Deficient Mice Develop Prostate Adenocarcinoma. PLoS One, 2013, 8(9):e73072.

Discussion
35
an identical extracellular N-terminus. Overall, the observed differences can be explained by the transmembrane type-I nature of TMPAP and the presence of a tyrosine-based motif at the C-terminus (Article III).
The observed changes at the gene-expression level in LNCaP-TMPAP cells are compatible with a delay in G1/S cell-cycle phase and with a dysregulated endo/exocytic process. The downregulation of histones and CDK6 can explain the retarded cell cycle. In addition, the extracellular adenosine pathway appears to be affected by the downregulation of adenosine receptors (ADORA1, ADORA2A, ADORA2B, and ADORA3) and adenosine kinase (ADK). These changes can be explained after considering that the increased expression of TMPAP results in an increased extracellular 5’-ectonucleotidase activity (Article III).
In the proposed mechanistic model the endocytic vesicles loaded with TMPAP are sorted to lysosomes by means of the Yxxφ motif. Consequently other cargo molecules carried by TMPAP-loaded vesicles are targeted for degradation. This can serve as an alternate mechanism of degradation for those cell surface receptors (such as RTKs or GPCR) that fail to follow the canonical monoubiquitination pathway. Moreover, the presence of TMPAP in the exosomes of LNCaP cells (Quintero et al. 2013) can be another alternative cellular mechanism to downregulate cell surface signaling receptors (Article III).
5.3 PAP serves specific function in male mouse SMG and saliva
The balance between exo- and endocytosis controls the cell membrane homeostasis and other cellular processes such as cell division. We also found that TMPAP interacts with snapin that is a SNARE-associated protein and PAP-/- mice developed prostate adenocarcinoma (Quintero et al. 2013). In addition, snapin is a component of BLOC1—a complex required for the biogenesis of specialized organelles of the endosomal-lysosomal pathway. Therefore, by considering that TMPAP is in the endosomal-lysosomal/exosomal pathway we decided to study the SMGs of PAP-/- and wild-type mice as a model of exocrine organ. In mice, PAP accounts for 50% of the total acid phosphatase activity in male saliva whereas female saliva lacks PAP activity (Article II). In male wild-type mice, PAP activity exclusively localizes to the GCT cells. Moreover, under the reported experimental conditions, PAP is the only 5’-ectonucleotidase expressed in GCT cells (Article II) whereas there are other 5’-ectonucleotidases present in acinar cells such as Cd73 (Nt5e) (Lavoie et al. 2011). Another parameter to consider is the pH of the saliva. According to the literature, the pH of the C57BL/6J male-mouse saliva lies in the range 8.0 to 11.0 (Ma et al. 1999; Ruiz-Ederra et al. 2009). In mammals, the high pH of saliva is required to prevent the formation of dental caries (Catalan et al.
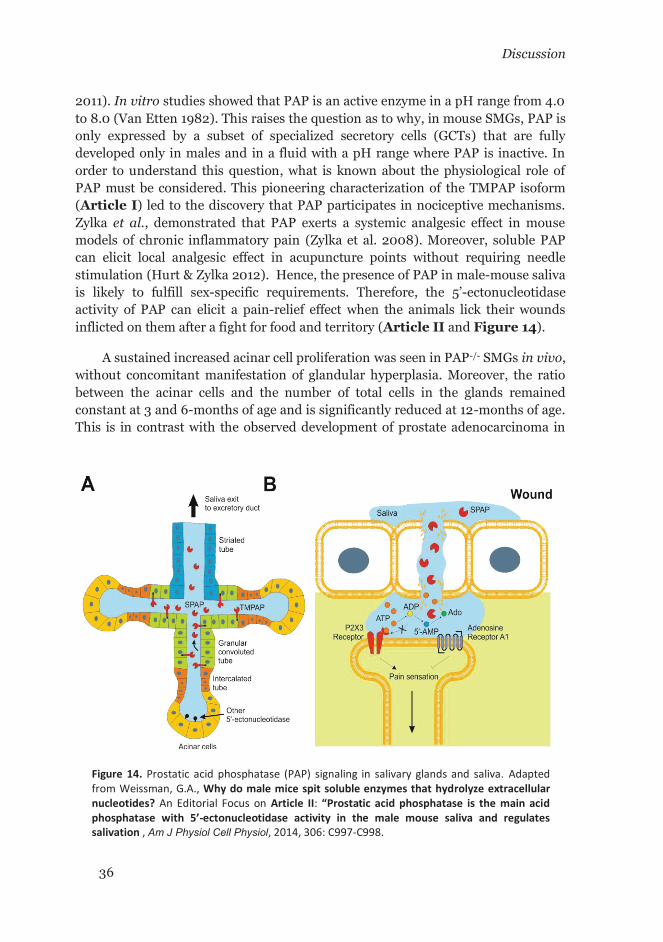
Discussion
36
2011). In vitro studies showed that PAP is an active enzyme in a pH range from 4.0 to 8.0 (Van Etten 1982). This raises the question as to why, in mouse SMGs, PAP is only expressed by a subset of specialized secretory cells (GCTs) that are fully developed only in males and in a fluid with a pH range where PAP is inactive. In order to understand this question, what is known about the physiological role of PAP must be considered. This pioneering characterization of the TMPAP isoform (Article I) led to the discovery that PAP participates in nociceptive mechanisms. Zylka et al., demonstrated that PAP exerts a systemic analgesic effect in mouse models of chronic inflammatory pain (Zylka et al. 2008). Moreover, soluble PAP can elicit local analgesic effect in acupuncture points without requiring needle stimulation (Hurt & Zylka 2012). Hence, the presence of PAP in male-mouse saliva is likely to fulfill sex-specific requirements. Therefore, the 5’-ectonucleotidase activity of PAP can elicit a pain-relief effect when the animals lick their wounds inflicted on them after a fight for food and territory (Article II and Figure 14).
A sustained increased acinar cell proliferation was seen in PAP-/- SMGs in vivo, without concomitant manifestation of glandular hyperplasia. Moreover, the ratio between the acinar cells and the number of total cells in the glands remained constant at 3 and 6-months of age and is significantly reduced at 12-months of age. This is in contrast with the observed development of prostate adenocarcinoma in
Figure 14. Prostatic acid phosphatase (PAP) signaling in salivary glands and saliva. Adapted from Weissman, G.A., Why do male mice spit soluble enzymes that hydrolyze extracellular nucleotides? An Editorial Focus on Article II: “Prostatic acid phosphatase is the main acid phosphatase with 5’-ectonucleotidase activity in the male mouse saliva and regulates salivation , Am J Physiol Cell Physiol, 2014, 306: C997-C998.

Discussion
37
PAP-/- mice (Quintero et al. 2013). To understand this phenomenon at the molecular level, microarray and microRNA experiments were performed on SMG tissue from 2-month-old wild-type and PAP-/- mice. They demonstrated an upregulation of genes related to cell proliferation and immune system response. In addition, miR-146a—an important regulator of inflammation—was upregulated. Thus, a histological evaluation of the glands was made. The results indicated significantly increased lymphocyte infiltration in 3-month-old mice that reached a plateau from 6 to 12-months of age. The increased acinar cell proliferation at 2-month-of-age occurs concomitantly with the upregulation of inflammatory and immune system genes. Consequently, the immune reaction is likely responsible for keeping the proliferative acinar cells under control. In addition, the upregulation of mir-146a and pro-resolving inflammatory genes, such as Irf7 and Fpr2, may be responsible for avoiding extensive tissue damage that otherwise should occur as a consequence of the increased T-lymphocyte infiltration as observed in mouse models of autoimmune diseases (e.g. Sjögren’s syndrome) (Varin et al. 2012).

Discussion
38
6 CONCLUSIONS AND FUTURE PERSPECTIVES This work reveals new fundamental clues regarding the molecular pathways
where PAP is involved. During the characterization of the novel TMPAP isoform its localization in the endosomal/lysosomal and exosomal route was predicted. These predictions were experimentally confirmed and led to the formulation of a mechanistic working model that opens new avenues of research. For instance, it led to the final characterization of TMPase—a hitherto uncharacterized enzyme involved in mechanisms of nociception—and placed PAP into the realm of 5’-ectonucleotidases and pain signaling. In addition, it was observed that a virally-driven overexpression of TMPAP in the rostral ventrolateral medulla oblongata led to a significant reduction in arterial blood pressure in rats—an effect attributed to the 5’-ectonucleotidase activity (Marina et al. 2015). By studying the mouse SMG as a model of exocrine organ the presence of PAP in male mouse saliva was concluded to be required to support sex-specific requirements related to the antinociceptive effect elicited by the 5’-ectonucleotidase activity on wounds. The lack of PAP expression in male mice SMG also fails to compromise either the innate immune response against highly proliferative cells or the pro-resolving anti-inflammatory processes responsible to keep tissue homeostasis. This is an important difference with the observed development of adenocarcinoma in the prostate—an immunologically ignored organ (Bruno et al. 2012). Furthermore, TMPAP, but not SPAP, regulates LNCaP cell proliferation and this led to the conclusion that this effect is associated to the presence of TMPAP in the endocytic pathway. Therefore, future retrospective studies will bring explanations of many observations that could not be explained by considering only the secreted isoform of PAP. For instance, the radio-imaging detection of metastatic lesions observed by Vihko et al. about 30 years ago, which can be a valuable tool for diagnosis (Kontturi et al. 1988). In addition, the presence of TMPAP in the endosomal-lysosomal/exosomal pathway could be exploited by drug delivery protocols. Moreover, PAP can become a fundamental therapeutic tool for the management of neuropathic pain, which is a common co-morbidity of many diseases, including metastatic prostate cancer.

39
7 ACKNOWLEDGEMENTS This study was carried out at the Research Center for Molecular Endocrinology,
Faculty of Medicine, University of Oulu; the Department of Biosciences, Faculty of Biological and Environmental Sciences, University of Helsinki; and the Department of Clinical Chemistry and Haematology, Faculty of Medicine, University of Helsinki.
I wish to express my sincere gratitude to Prof. Pirkko Vihko for the opportunity to carry out this thesis work in her research group. I greatly appreciate her support, guidance and dedication through all these years.
I also thank the pre-examiners Adj. Prof. Hannu Koistinen and Adj. Prof. Sakari Kellokumpu for their careful review accompanied by very valuable comments and a constructive criticism aimed to improve the content of this thesis. I also wish to thank my thesis committee members Adj. Prof. Hannu Koistinen and Adj. Prof. Päivi Lakkisto for their support and always constructive comments. I express my gratitude also to Prof. Tom Böhling and Prof. Ulf-Håkan Stenman for their kind support during the writing of this thesis book.
I wish to acknowledge to all my co-authors and coworkers that through their talented contribution bring this work to a conclusion: Ileana Quintero, Annakaisa Herrala, Anitta Pulkka, Riikka Wirkkala, Eeva-Liisa Eskelinen, Eija Jokitalo, Pekka Hellström, Hannu Tuominen, Pasi Hirvikoski, Anja Kipar, Sampsa Hautaniemi, Kristian Ovaska, and Lilli Saarinen.
I also wish to express my sincere gratitude to Prof. Mika Scheinin and Dr. Eeva Valve, the director and coordinator of the FPDP-D (former Drug Discovery Graduate School) respectively, for the support and the enriching atmosphere during the annual meetings organized on wonderful places across Finland.
I would like to express also my special gratitude and affection to Annakaisa Herrala for her unconditional support inside and outside the lab, for allow me and my family to be part of her family, and especially for her friendship that made Finland a shinier and warmer place for me.
I thank Nick Domansky for his friendship, his unconditional help, and his good sense of humor, and for being always present in some of the most important moments of my personal life in Finland.
I wish to thank my former and present colleagues at the Department of Biosciences and the Department of Clinical Chemistry and Haematology for their support and contribution.

40
I wish to thank my former colleagues in Oulu Anitta Pulkka and Riita Kurkela for their valuable support and contribution and more important to let me share with them wonderful moments while visiting different places in Finland that make me feel part of a big family. I also want to thank Prof. Andre Juffer, Serena Donini, Satyan Sharma, Outi Lampela and Niko Pursiainen from the Biocomputing Group at the University of Oulu for sharing their knowledge and expertise during the early stages of this work.
I cordially thank the guys at Redmond for making our working life more challenging after every update of their software products.
I also warmly thank my friends from Oulu Eija and Topi, Milly and Manuel, María and Omar, for their unconditional support, to make me feel part of their families and the wonderful moments we shared and still do when someone travels to break the distance. I also thank Alberto and María and their extraordinary family for the shared moments and especially for their dedication to translate to me the homily during the services at the Catholic Parish of Holy Family of Nazareth of Oulu. I wish also to thank Hernán for his friendship, his sense of humor, the sharing of his political views and the Pisco Sour.
I feel extremely grateful to my family, my parents Hugo and Rosa, and my sister, Alejandra that always supported and believed in me. Thanks for all their efforts, the encouragement and the values they transmitted to me and for taught me how to keep the faith always in spite of any adverse circumstance.
Finally, all my love and gratitude to Ileana the woman that let me share beside her this wonderful journey through the life. Thanks for the support, the comprehension, the patience and the unconditional love. And to my son Santiago, thanks for filling my days with joy, for not let me forget what the important things on life are and at the same time let me forget about the distance from my homeland with every smile.
This thesis work has received the financial support from CIMO, FPDP-D, K. Albin Johanssons Foundation, Academy of Finland, The Finnish Cancer Foundation, Biocenter Oulu, Magnus Ehrnrooth Foundation, Kliinisen Kemian Tutkimussäätiö, University of Helsinki, HUS-Evo, Sigrid Jussélius Foundation, Laboratoriolääketieteen Edistämissäätiö, The Finnish Society of Sciences and Letters, and Ida Montin Foundation.
César L. Araujo
Helsinki, 2016

41
8 BIBLIOGRAPHY
Agrawal, T., Schu, P., and Medigeshi, G. R. 2013. "Adaptor Protein Complexes-1 and 3 are Involved at Distinct Stages of Flavivirus Life-Cycle." Scientific Reports 3: 1813. doi:10.1038/srep01813 [doi].
Andrews, N. W., Almeida, P. E., and Corrotte, M. 2014. "Damage Control: Cellular Mechanisms of Plasma Membrane Repair." Trends in Cell Biology 24 (12): 734-742. doi:10.1016/j.tcb.2014.07.008 [doi].
Barka, T. 1980. "Biologically Active Polypeptides in Submandibular Glands." The Journal of Histochemistry and Cytochemistry : Official Journal of the Histochemistry Society 28 (8): 836-859.
Baum, B. J. and R. B. Wellner. 1999. "Receptors in Salivary Glands." In Neural Mechanisms of Salivary Gland Secretion, edited by J. R. Garret, J. Ekström and L. C. Anderson. Vol. 11, 44: Karger.
Bendtsen, J. D., Nielsen, H., von Heijne, G., and Brunak, S. 2004. "Improved Prediction of Signal Peptides: SignalP 3.0." Journal of Molecular Biology 340 (4): 783-795. doi:10.1016/j.jmb.2004.05.028 [doi].
Bennett, M. K., Calakos, N., and Scheller, R. H. 1992. "Syntaxin: A Synaptic Protein Implicated in Docking of Synaptic Vesicles at Presynaptic Active Zones." Science (New York, N.Y.) 257 (5067): 255-259.
Bitler, B. G., Goverdhan, A., and Schroeder, J. A. 2010. "MUC1 Regulates Nuclear Localization and Function of the Epidermal Growth Factor Receptor." Journal of Cell Science 123 (Pt 10): 1716-1723. doi:10.1242/jcs.062661 [doi].
Blank, U., Madera-Salcedo, I. K., Danelli, L., Claver, J., Tiwari, N., Sanchez-Miranda, E., Vazquez-Victorio, G., Ramirez-Valadez, K. A., Macias-Silva, M., and Gonzalez-Espinosa, C. 2014. "Vesicular Trafficking and Signaling for Cytokine and Chemokine Secretion in Mast Cells." Frontiers in Immunology 5: 453. doi:10.3389/fimmu.2014.00453 [doi].
Bodner, L., Knyszynski, A., Adler-Kunin, S., and Danon, D. 1991. "The Effect of Selective Desalivation on Wound Healing in Mice." Experimental Gerontology 26 (4): 357-363.
Bonifacino, J. S. and Traub, L. M. 2003. "Signals for Sorting of Transmembrane Proteins to Endosomes and Lysosomes." Annual Review of Biochemistry 72: 395-447. doi:10.1146/annurev.biochem.72.121801.161800.

Bibliography
42
Braun, M., Waheed, A., and von Figura, K. 1989. "Lysosomal Acid Phosphatase is Transported to Lysosomes Via the Cell Surface." The EMBO Journal 8 (12): 3633-3640.
Bruce, A. W., Mahan, D. E., and Beleville, W. D. 1980. "Radioimmunoassay of Bone Marrow Prostatic Acid Phosphatase." The Prostate 1 (4): 457-463.
Bruno, T. C., Rothwell, C., Grosso, J. F., Getnet, D., Yen, H. R., Durham, N. M., Netto, G., Pardoll, D. M., and Drake, C. G. 2012. "Anti-Tumor Effects of Endogenous Prostate Cancer-Specific CD8 T Cells in a Murine TCR Transgenic Model." The Prostate 72 (5): 514-522. doi:10.1002/pros.21453 [doi].
Buchwald, S. L., Saini, M. S., Knowles, J. R., and Van Etten, R. L. 1984. "Stereochemical Course of Phospho Group Transfer by Human Prostatic Acid Phosphatase." The Journal of Biological Chemistry 259 (4): 2208-2213.
Buermans, H. P., Ariyurek, Y., van Ommen, G., den Dunnen, J. T., and 't Hoen, P. A. 2010. "New Methods for Next Generation Sequencing Based microRNA Expression Profiling." BMC Genomics 11: 716-2164-11-716. doi:10.1186/1471-2164-11-716; 10.1186/1471-2164-11-716.
Buxton, P., Zhang, X. M., Walsh, B., Sriratana, A., Schenberg, I., Manickam, E., and Rowe, T. 2003. "Identification and Characterization of Snapin as a Ubiquitously Expressed SNARE-Binding Protein that Interacts with SNAP23 in Non-Neuronal Cells." The Biochemical Journal 375 (Pt 2): 433-440. doi:10.1042/BJ20030427.
Catalan, M. A., Scott-Anne, K., Klein, M. I., Koo, H., Bowen, W. H., and Melvin, J. E. 2011. "Elevated Incidence of Dental Caries in a Mouse Model of Cystic Fibrosis." PloS One 6 (1): e16549. doi:10.1371/journal.pone.0016549 [doi].
Catalona, W. J., Smith, D. S., Ratliff, T. L., Dodds, K. M., Coplen, D. E., Yuan, J. J., Petros, J. A., and Andriole, G. L. 1991. "Measurement of Prostate-Specific Antigen in Serum as a Screening Test for Prostate Cancer." The New England Journal of Medicine 324 (17): 1156-1161. doi:10.1056/NEJM199104253241702 [doi].
Cheever, M. A. and Higano, C. S. 2011. "PROVENGE (Sipuleucel-T) in Prostate Cancer: The First FDA-Approved Therapeutic Cancer Vaccine." Clinical Cancer Research : An Official Journal of the American Association for Cancer Research 17 (11): 3520-3526. doi:10.1158/1078-0432.CCR-10-3126.
Chheda, M. G., Ashery, U., Thakur, P., Rettig, J., and Sheng, Z. H. 2001. "Phosphorylation of Snapin by PKA Modulates its Interaction with the SNARE Complex." Nature Cell Biology 3 (4): 331-338. doi:10.1038/35070000.

Bibliography
43
Chuang, T. D., Chen, S. J., Lin, F. F., Veeramani, S., Kumar, S., Batra, S. K., Tu, Y., and Lin, M. F. 2010. "Human Prostatic Acid Phosphatase, an Authentic Tyrosine Phosphatase, Dephosphorylates ErbB-2 and Regulates Prostate Cancer Cell Growth." The Journal of Biological Chemistry 285 (31): 23598-23606. doi:10.1074/jbc.M109.098301 [doi].
De Angelis Campos, A. C., Rodrigues, M. A., de Andrade, C., de Goes, A. M., Nathanson, M. H., and Gomes, D. A. 2011. "Epidermal Growth Factor Receptors Destined for the Nucleus are Internalized Via a Clathrin-Dependent Pathway." Biochemical and Biophysical Research Communications 412 (2): 341-346. doi:10.1016/j.bbrc.2011.07.100 [doi].
Dell'Angelica, E. C., Mullins, C., Caplan, S., and Bonifacino, J. S. 2000. "Lysosome-Related Organelles." FASEB Journal : Official Publication of the Federation of American Societies for Experimental Biology 14 (10): 1265-1278.
Development Core Team, R. 2014. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. http://www.R-project.org.
Dobrowolski, R. and De Robertis, E. M. 2011. "Endocytic Control of Growth Factor Signalling: Multivesicular Bodies as Signalling Organelles." Nature Reviews.Molecular Cell Biology 13 (1): 53-60. doi:10.1038/nrm3244 [doi].
Dodd, J., Jahr, C. E., Hamilton, P. N., Heath, M. J., Matthew, W. D., and Jessell, T. M. 1983. "Cytochemical and Physiological Properties of Sensory and Dorsal Horn Neurons that Transmit Cutaneous Sensation." Cold Spring Harbor Symposia on Quantitative Biology 48 Pt 2: 685-695.
Dores, M. R., Paing, M. M., Lin, H., Montagne, W. A., Marchese, A., and Trejo, J. 2012. "AP-3 Regulates PAR1 Ubiquitin-Independent MVB/Lysosomal Sorting Via an ALIX-Mediated Pathway." Molecular Biology of the Cell 23 (18): 3612-3623. doi:10.1091/mbc.E12-03-0251 [doi].
Draisma, G., Boer, R., Otto, S. J., van der Cruijsen, I. W., Damhuis, R. A., Schroder, F. H., and de Koning, H. J. 2003. "Lead Times and Overdetection due to Prostate-Specific Antigen Screening: Estimates from the European Randomized Study of Screening for Prostate Cancer." Journal of the National Cancer Institute 95 (12): 868-878.
Dziembor-Gryszkiewicz, E., Fikus, M., Kazimierczuk, Z., and Ostrowski, W. 1978. "Activity of Human Prostatic Acid Phosphatase Toward Purine 5'-Phosphonucleosides." Bulletin De L'Academie Polonaise Des Sciences.Serie Des Sciences Biologiques 26 (12): 815-821.
Engstler, M., Weise, F., Bopp, K., Grunfelder, C. G., Gunzel, M., Heddergott, N., and Overath, P. 2005. "The Membrane-Bound Histidine Acid Phosphatase

Bibliography
44
TbMBAP1 is Essential for Endocytosis and Membrane Recycling in Trypanosoma Brucei." Journal of Cell Science 118 (Pt 10): 2105-2118. doi:jcs.02327 [pii].
Ferreira, F., Foley, M., Cooke, A., Cunningham, M., Smith, G., Woolley, R., Henderson, G., Kelly, E., Mundell, S., and Smythe, E. 2012. "Endocytosis of G Protein-Coupled Receptors is Regulated by Clathrin Light Chain Phosphorylation." Current Biology : CB 22 (15): 1361-1370. doi:10.1016/j.cub.2012.05.034 [doi].
Flint, A. J., Tiganis, T., Barford, D., and Tonks, N. K. 1997. "Development of "Substrate-Trapping" Mutants to Identify Physiological Substrates of Protein Tyrosine Phosphatases." Proceedings of the National Academy of Sciences of the United States of America 94 (5): 1680-1685.
Geppert, M., Goda, Y., Hammer, R. E., Li, C., Rosahl, T. W., Stevens, C. F., and Sudhof, T. C. 1994. "Synaptotagmin I: A Major Ca2+ Sensor for Transmitter Release at a Central Synapse." Cell 79 (4): 717-727. doi:0092-8674(94)90556-8 [pii].
Gottschalk, S., Waheed, A., Schmidt, B., Laidler, P., and von Figura, K. 1989. "Sequential Processing of Lysosomal Acid Phosphatase by a Cytoplasmic Thiol Proteinase and a Lysosomal Aspartyl Proteinase." The EMBO Journal 8 (11): 3215-3219.
Gouet, P., Courcelle, E., Stuart, D. I., and Metoz, F. 1999. "ESPript: Analysis of Multiple Sequence Alignments in PostScript." Bioinformatics (Oxford, England) 15 (4): 305-308. doi:btc028 [pii].
Gough, N. R., Zweifel, M. E., Martinez-Augustin, O., Aguilar, R. C., Bonifacino, J. S., and Fambrough, D. M. 1999. "Utilization of the Indirect Lysosome Targeting Pathway by Lysosome-Associated Membrane Proteins (LAMPs) is Influenced Largely by the C-Terminal Residue of their GYXXphi Targeting Signals." Journal of Cell Science 112 ( Pt 23) (Pt 23): 4257-4269.
Gresik, E. W. 1994. "The Granular Convoluted Tubule (GCT) Cell of Rodent Submandibular Glands." Microscopy Research and Technique 27 (1): 1-24. doi:10.1002/jemt.1070270102 [doi].
Griffiths, J. C. 1983. "The Laboratory Diagnosis of Prostatic Adenocarcinoma." Critical Reviews in Clinical Laboratory Sciences 19 (3): 187-204. doi:10.3109/10408368309165763 [doi].
Gutman, A. B. and Gutman, E. B. 1938. "An " Acid " Phosphatase Occurring in the Serum of Patients with Metastasizing Carcinoma of the Prostate Gland." The Journal of Clinical Investigation 17 (4): 473-478. doi:10.1172/JCI100974.

Bibliography
45
Hu, C., Ahmed, M., Melia, T. J., Sollner, T. H., Mayer, T., and Rothman, J. E. 2003. "Fusion of Cells by Flipped SNAREs." Science (New York, N.Y.) 300 (5626): 1745-1749. doi:10.1126/science.1084909 [doi].
Huang, F., Khvorova, A., Marshall, W., and Sorkin, A. 2004. "Analysis of Clathrin-Mediated Endocytosis of Epidermal Growth Factor Receptor by RNA Interference." The Journal of Biological Chemistry 279 (16): 16657-16661. doi:10.1074/jbc.C400046200 [doi].
Hurt, J. K., Fitzpatrick, B. J., Norris-Drouin, J., and Zylka, M. J. 2012. "Secretion and N-Linked Glycosylation are Required for Prostatic Acid Phosphatase Catalytic and Antinociceptive Activity." PloS One 7 (2): e32741. doi:10.1371/journal.pone.0032741 [doi].
Hurt, J. K. and Zylka, M. J. 2012. "PAPupuncture has Localized and Long-Lasting Antinociceptive Effects in Mouse Models of Acute and Chronic Pain." Molecular Pain 8: 28-8069-8-28. doi:10.1186/1744-8069-8-28; 10.1186/1744-8069-8-28.
Inoue, A., Obata, K., and Akagawa, K. 1992. "Cloning and Sequence Analysis of cDNA for a Neuronal Cell Membrane Antigen, HPC-1." The Journal of Biological Chemistry 267 (15): 10613-10619.
Jayasinghe, N. R., Cope, G. H., and Jacob, S. 1990. "Morphometric Studies on the Development and Sexual Dimorphism of the Submandibular Gland of the Mouse." Journal of Anatomy 172: 115-127.
Junqueira, L. C. and Fajer, A. 1949. "Biochemical and Histochemical Observations on the Sexual Dimorphism of Mice Submaxillary Glands." Journal of Cellular Physiology 34 (1): 129-58, incl 6 pl.
Kahsay, R. Y., Gao, G., and Liao, L. 2005. "An Improved Hidden Markov Model for Transmembrane Protein Detection and Topology Prediction and its Applications to Complete Genomes." Bioinformatics (Oxford, England) 21 (9): 1853-1858. doi:bti303 [pii].
Kantoff, P. W., Higano, C. S., Shore, N. D., Berger, E. R., Small, E. J., Penson, D. F., Redfern, C. H., et al. 2010. "Sipuleucel-T Immunotherapy for Castration-Resistant Prostate Cancer." The New England Journal of Medicine 363 (5): 411-422. doi:10.1056/NEJMoa1001294.
Katzmann, D. J., Odorizzi, G., and Emr, S. D. 2002. "Receptor Downregulation and Multivesicular-Body Sorting." Nature Reviews.Molecular Cell Biology 3 (12): 893-905. doi:10.1038/nrm973 [doi].
Knyihar-Csillik, E., Bezzegh, A., Boti, S., and Csillik, B. 1986. "Thiamine Monophosphatase: A Genuine Marker for Transganglionic Regulation of

Bibliography
46
Primary Sensory Neurons." The Journal of Histochemistry and Cytochemistry : Official Journal of the Histochemistry Society 34 (3): 363-371.
Kontturi, M., Lukkarinen, O., Vihko, P., and Vihko, R. 1988. "Radioimaging of Lymph Node Involvement in Prostatic Carcinoma." Scandinavian Journal of Urology and Nephrology.Supplementum 110: 113-117.
Kutscher, W. and Wolbergs, H. 1935. "Prostataphosphatase." Z. Physiol. Chem. 236: 237.
Lampe, M., Pierre, F., Al-Sabah, S., Krasel, C., and Merrifield, C. J. 2014. "Dual Single-Scission Event Analysis of Constitutive Transferrin Receptor (TfR) Endocytosis and Ligand-Triggered Beta2-Adrenergic Receptor (beta2AR) Or Mu-Opioid Receptor (MOR) Endocytosis." Molecular Biology of the Cell 25 (19): 3070-3080. doi:10.1091/mbc.E14-06-1112 [doi].
Langmead, B., Trapnell, C., Pop, M., and Salzberg, S. L. 2009. "Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome." Genome Biology 10 (3): R25-2009-10-3-r25. Epub 2009 Mar 4. doi:10.1186/gb-2009-10-3-r25; 10.1186/gb-2009-10-3-r25.
Laukaitis, C. M., Dlouhy, S. R., Emes, R. D., Ponting, C. P., and Karn, R. C. 2005. "Diverse Spatial, Temporal, and Sexual Expression of Recently Duplicated Androgen-Binding Protein Genes in Mus Musculus." BMC Evolutionary Biology 5: 40. doi:10.1186/1471-2148-5-40.
Lavoie, E. G., Gulbransen, B. D., Martin-Satue, M., Aliagas, E., Sharkey, K. A., and Sevigny, J. 2011. "Ectonucleotidases in the Digestive System: Focus on NTPDase3 Localization." American Journal of Physiology.Gastrointestinal and Liver Physiology 300 (4): G608-20. doi:10.1152/ajpgi.00207.2010; 10.1152/ajpgi.00207.2010.
Li, H. C., Chernoff, J., Chen, L. B., and Kirschonbaum, A. 1984. "A Phosphotyrosyl-Protein Phosphatase Activity Associated with Acid Phosphatase from Human Prostate Gland." European Journal of Biochemistry / FEBS 138 (1): 45-51.
Lin, M. F. and Clinton, G. M. 1988. "The Epidermal Growth Factor Receptor from Prostate Cells is Dephosphorylated by a Prostate-Specific Phosphotyrosyl Phosphatase." Molecular and Cellular Biology 8 (12): 5477-5485.
Lin, M. F., DaVolio, J., and Garcia-Arenas, R. 1992. "Expression of Human Prostatic Acid Phosphatase Activity and the Growth of Prostate Carcinoma Cells." Cancer Research 52 (17): 4600-4607.
Lindqvist, Y., Schneider, G., and Vihko, P. 1994. "Crystal Structures of Rat Acid Phosphatase Complexed with the Transition-State Analogs Vanadate and

Bibliography
47
Molybdate. Implications for the Reaction Mechanism." European Journal of Biochemistry / FEBS 221 (1): 139-142.
Lindqvist, Y., Schneider, G., and Vihko, P. 1993. "Three-Dimensional Structure of Rat Acid Phosphatase in Complex with L(+)-Tartrate." The Journal of Biological Chemistry 268 (28): 20744-20746.
Luchter-Wasylewska, E. 2001. "Cooperative Kinetics of Human Prostatic Acid Phosphatase." Biochimica Et Biophysica Acta 1548 (2): 257-264.
Luzio, J. P., Pryor, P. R., and Bright, N. A. 2007. "Lysosomes: Fusion and Function." Nature Reviews.Molecular Cell Biology 8 (8): 622-632. doi:nrm2217 [pii].
Ma, T., Song, Y., Gillespie, A., Carlson, E. J., Epstein, C. J., and Verkman, A. S. 1999. "Defective Secretion of Saliva in Transgenic Mice Lacking Aquaporin-5 Water Channels." The Journal of Biological Chemistry 274 (29): 20071-20074.
Madrid, R., Le Maout, S., Barrault, M. B., Janvier, K., Benichou, S., and Merot, J. 2001. "Polarized Trafficking and Surface Expression of the AQP4 Water Channel are Coordinated by Serial and Regulated Interactions with Different Clathrin-Adaptor Complexes." The EMBO Journal 20 (24): 7008-7021. doi:10.1093/emboj/20.24.7008 [doi].
Marina, N., Ang, R., Machhada, A., Kasymov, V., Karagiannis, A., Hosford, P. S., Mosienko, V., et al. 2015. "Brainstem Hypoxia Contributes to the Development of Hypertension in the Spontaneously Hypertensive Rat." Hypertension 65 (4): 775-783. doi:10.1161/HYPERTENSIONAHA.114.04683 [doi].
Matsuda, M. and Ogoshi, H. 1966. "Specificity of DNase I. Estimation of Nucleosides Present at the 5'-Phosphate Terminus of a Limit Digest of DNA by DNase I." Journal of Biochemistry 59 (3): 230-235.
Matsuo, R. 1999. "Central Connections for Salivary Innervations and Efferent Impulse Formation." In Neural Mechanisms of Salivary Gland Secretion, edited by J. R. Garret, J. Ekström and L. C. Anderson. Vol. 11, 26: Karger.
Mellman, I., Fuchs, R., and Helenius, A. 1986. "Acidification of the Endocytic and Exocytic Pathways." Annual Review of Biochemistry 55: 663-700. doi:10.1146/annurev.bi.55.070186.003311 [doi].
Mellman, I. and Yarden, Y. 2013. "Endocytosis and Cancer." Cold Spring Harbor Perspectives in Biology 5 (12): a016949. doi:10.1101/cshperspect.a016949 [doi].

Bibliography
48
Meng, T. C. and Lin, M. F. 1998. "Tyrosine Phosphorylation of C-ErbB-2 is Regulated by the Cellular Form of Prostatic Acid Phosphatase in Human Prostate Cancer Cells." The Journal of Biological Chemistry 273 (34): 22096-22104.
Mori, M., Takai, Y., and Kunnikata, M. 1992. "Biologically Active Peptides in the Submandibular Salivary Gland - Role of the Granular Convoluted Tubule." Acta Histochem Cytochem 25: 325-341.
Mosesson, Y., Mills, G. B., and Yarden, Y. 2008. "Derailed Endocytosis: An Emerging Feature of Cancer." Nature Reviews.Cancer 8 (11): 835-850. doi:10.1038/nrc2521 [doi].
Mrschtik, M. and Ryan, K. M. 2015. "Lysosomal Proteins in Cell Death and Autophagy." The FEBS Journal 282 (10): 1858-1870. doi:10.1111/febs.13253 [doi].
Ostrowski, W. and Rybarska, J. 1965. "Studies on Human Prostatic Acid Phosphomonoesterase. further Purification and Molecular Weight of the Enzyme." Biochimica Et Biophysica Acta 105 (1): 196-198.
Otto, G. P. and Nichols, B. J. 2011. "The Roles of Flotillin Microdomains--Endocytosis and Beyond." Journal of Cell Science 124 (Pt 23): 3933-3940. doi:10.1242/jcs.092015 [doi].
Ovaska, K., Laakso, M., Haapa-Paananen, S., Louhimo, R., Chen, P., Aittomaki, V., Valo, E., et al. 2010. "Large-Scale Data Integration Framework Provides a Comprehensive View on Glioblastoma Multiforme." Genome Medicine 2 (9): 65. doi:10.1186/gm186; 10.1186/gm186.
Oyler, G. A., Higgins, G. A., Hart, R. A., Battenberg, E., Billingsley, M., Bloom, F. E., and Wilson, M. C. 1989. "The Identification of a Novel Synaptosomal-Associated Protein, SNAP-25, Differentially Expressed by Neuronal Subpopulations." The Journal of Cell Biology 109 (6 Pt 1): 3039-3052.
Porvari, K. S., Herrala, A. M., Kurkela, R. M., Taavitsainen, P. A., Lindqvist, Y., Schneider, G., and Vihko, P. T. 1994. "Site-Directed Mutagenesis of Prostatic Acid Phosphatase. Catalytically Important Aspartic Acid 258, Substrate Specificity, and Oligomerization." The Journal of Biological Chemistry 269 (36): 22642-22646.
Proctor, G. B. 1998. "Secretory Protein Synthesis and Constitutive (Vesicular) Secretion by Salivary Glands." In Glandurlar Mechanisms of Salivary Secretion, edited by J. R. Garret, J. Ekström and L. C. Anderson. Vol. 10, 73: Karger.

Bibliography
49
Proctor, G. B. and Carpenter, G. H. 2007. "Regulation of Salivary Gland Function by Autonomic Nerves." Autonomic Neuroscience : Basic & Clinical 133 (1): 3-18. doi:10.1016/j.autneu.2006.10.006.
Qian, Y., Liu, Y., Campbell, J., Thomson, E., Kong, Y. M., and Scheuermann, R. H. 2012. "FCSTrans: An Open Source Software System for FCS File Conversion and Data Transformation." Cytometry.Part A : The Journal of the International Society for Analytical Cytology 81 (5): 353-356. doi:10.1002/cyto.a.22037 [doi].
Quintero, I. B., Araujo, C. L., Pulkka, A. E., Wirkkala, R. S., Herrala, A. M., Eskelinen, E. L., Jokitalo, E., et al. 2007. "Prostatic Acid Phosphatase is Not a Prostate Specific Target." Cancer Research 67 (14): 6549-6554. doi:10.1158/0008-5472.CAN-07-1651.
Quintero, I. B., Herrala, A. M., Araujo, C. L., Pulkka, A. E., Hautaniemi, S., Ovaska, K., Pryazhnikov, E., et al. 2013. "Transmembrane Prostatic Acid Phosphatase (TMPAP) Interacts with Snapin and Deficient Mice Develop Prostate Adenocarcinoma." PloS One 8 (9): e73072. doi:10.1371/journal.pone.0073072; 10.1371/journal.pone.0073072.
Ramsay, A. G., Keppler, M. D., Jazayeri, M., Thomas, G. J., Parsons, M., Violette, S., Weinreb, P., Hart, I. R., and Marshall, J. F. 2007. "HS1-Associated Protein X-1 Regulates Carcinoma Cell Migration and Invasion Via Clathrin-Mediated Endocytosis of Integrin Alphavbeta6." Cancer Research 67 (11): 5275-5284. doi:67/11/5275 [pii].
Randerath, K., Randerath, E., Danna, T. F., van Golen, L., and Putman, K. L. 1989. "A New Sensitive 32P-Postlabeling Assay Based on the Specific Enzymatic Conversion of Bulky DNA Lesions to Radiolabeled Dinucleotides and Nucleoside 5'-Monophosphates." Carcinogenesis 10 (7): 1231-1239.
Rohrer, J., Schweizer, A., Russell, D., and Kornfeld, S. 1996. "The Targeting of Lamp1 to Lysosomes is Dependent on the Spacing of its Cytoplasmic Tail Tyrosine Sorting Motif Relative to the Membrane." The Journal of Cell Biology 132 (4): 565-576.
Roiko, K., Janne, O. A., and Vihko, P. 1990. "Primary Structure of Rat Secretory Acid Phosphatase and Comparison to Other Acid Phosphatases." Gene 89 (2): 223-229.
Rost, B. and Liu, J. 2003. "The PredictProtein Server." Nucleic Acids Research 31 (13): 3300-3304.
Ruder, C., Reimer, T., Delgado-Martinez, I., Hermosilla, R., Engelsberg, A., Nehring, R., Dorken, B., and Rehm, A. 2005. "EBAG9 Adds a New Layer of Control on Large Dense-Core Vesicle Exocytosis Via Interaction with Snapin."

Bibliography
50
Molecular Biology of the Cell 16 (3): 1245-1257. doi:10.1091/mbc.E04-09-0817.
Ruiz-Ederra, J., Levin, M. H., and Verkman, A. S. 2009. "In Situ Fluorescence Measurement of Tear Film [Na+], [K+], [Cl-], and pH in Mice shows Marked Hypertonicity in Aquaporin-5 Deficiency." Investigative Ophthalmology & Visual Science 50 (5): 2132-2138. doi:10.1167/iovs.08-3033; 10.1167/iovs.08-3033.
Rusinova, I., Forster, S., Yu, S., Kannan, A., Masse, M., Cumming, H., Chapman, R., and Hertzog, P. J. 2013. "Interferome V2.0: An Updated Database of Annotated Interferon-Regulated Genes." Nucleic Acids Research 41 (Database issue): D1040-6. doi:10.1093/nar/gks1215; 10.1093/nar/gks1215.
Schneider, G., Lindqvist, Y., and Vihko, P. 1993. "Three-Dimensional Structure of Rat Acid Phosphatase." The EMBO Journal 12 (7): 2609-2615.
Serrano, J. A., Shannon, W. A.,Jr, Sternberger, N. J., Wasserkrug, H. L., Serrano, A. A., and Seligman, A. M. 1976. "The Cytochemical Demonstration of Prostatic Acid Phosphatase using a New Substrate, Phosphorylcholine." The Journal of Histochemistry and Cytochemistry : Official Journal of the Histochemistry Society 24 (10): 1046-1056.
Sharma, S. and Juffer, A. H. 2009. "Hydrolysis of Phosphohistidine in Water and in Prostatic Acid Phosphatase." Chemical Communications (Cambridge, England) (42):6385-7. doi (42): 6385-6387. doi:10.1039/b910451h [doi].
Sharma, S., Rauk, A., and Juffer, A. H. 2008. "A DFT Study on the Formation of a Phosphohistidine Intermediate in Prostatic Acid Phosphatase." Journal of the American Chemical Society 130 (30): 9708-9716. doi:10.1021/ja710047a [doi].
Shin, S., Wolgamott, L., and Yoon, S. O. 2012. "Integrin Trafficking and Tumor Progression." International Journal of Cell Biology 2012: 516789. doi:10.1155/2012/516789 [doi].
Silverman, J. D. and Kruger, L. 1988. "Acid Phosphatase as a Selective Marker for a Class of Small Sensory Ganglion Cells in several Mammals: Spinal Cord Distribution, Histochemical Properties, and Relation to Fluoride-Resistant Acid Phosphatase (FRAP) of Rodents." Somatosensory Research 5 (3): 219-246.
Singh, H., Felts, R. L., Schuermann, J. P., Reilly, T. J., and Tanner, J. J. 2009. "Crystal Structures of the Histidine Acid Phosphatase from Francisella Tularensis Provide Insight into Substrate Recognition." Journal of Molecular Biology 394 (5): 893-904. doi:10.1016/j.jmb.2009.10.009.

Bibliography
51
Slepnev, V. I. and De Camilli, P. 2000. "Accessory Factors in Clathrin-Dependent Synaptic Vesicle Endocytosis." Nature Reviews.Neuroscience 1 (3): 161-172. doi:10.1038/35044540 [doi].
Sollner, T., Whiteheart, S. W., Brunner, M., Erdjument-Bromage, H., Geromanos, S., Tempst, P., and Rothman, J. E. 1993. "SNAP Receptors Implicated in Vesicle Targeting and Fusion." Nature 362 (6418): 318-324. doi:10.1038/362318a0 [doi].
Sorkin, A. and M. A. Puthenveedu. 2013. "Clathrin-Mediated Endocytosis." In Vesicle Trafficking in Cancer, edited by Y. Yasef and G. Tarcic. New York: Springer.
Sowa, N. A., Taylor-Blake, B., and Zylka, M. J. 2010. "Ecto-5'-Nucleotidase (CD73) Inhibits Nociception by Hydrolyzing AMP to Adenosine in Nociceptive Circuits." The Journal of Neuroscience : The Official Journal of the Society for Neuroscience 30 (6): 2235-2244. doi:10.1523/JNEUROSCI.5324-09.2010; 10.1523/JNEUROSCI.5324-09.2010.
Stamey, T. A., Yang, N., Hay, A. R., McNeal, J. E., Freiha, F. S., and Redwine, E. 1987. "Prostate-Specific Antigen as a Serum Marker for Adenocarcinoma of the Prostate." The New England Journal of Medicine 317 (15): 909-916. doi:10.1056/NEJM198710083171501.
Strausberg, R. L., Feingold, E. A., Grouse, L. H., Derge, J. G., Klausner, R. D., Collins, F. S., Wagner, L., et al. 2002. "Generation and Initial Analysis of More than 15,000 Full-Length Human and Mouse cDNA Sequences." Proceedings of the National Academy of Sciences of the United States of America 99 (26): 16899-16903. doi:10.1073/pnas.242603899 [doi].
Taelman, V. F., Dobrowolski, R., Plouhinec, J. L., Fuentealba, L. C., Vorwald, P. P., Gumper, I., Sabatini, D. D., and De Robertis, E. M. 2010. "Wnt Signaling Requires Sequestration of Glycogen Synthase Kinase 3 Inside Multivesicular Endosomes." Cell 143 (7): 1136-1148. doi:10.1016/j.cell.2010.11.034 [doi].
Talley, H. M., Laukaitis, C. M., and Karn, R. C. 2001. "Female Preference for Male Saliva: Implications for Sexual Isolation of Mus Musculus Subspecies." Evolution; International Journal of Organic Evolution 55 (3): 631-634.
Tanaka, M., Kishi, Y., Takanezawa, Y., Kakehi, Y., Aoki, J., and Arai, H. 2004. "Prostatic Acid Phosphatase Degrades Lysophosphatidic Acid in Seminal Plasma." FEBS Letters 571 (1-3): 197-204. doi:10.1016/j.febslet.2004.06.083.
Taylor, M. B. 2006. "STILTS - A Package for Command-Line Processing of Tabular Data." Astronomical Data Analysis Software and Systems XV ASP Conference Series 351.

Bibliography
52
Taylor, M. B. 2009. "STILTS Plotting Tools." Astronomical Data Analysis Software and Systems XVIII ASP Conference Series 411: 510.
Taylor, M. B. 2005. "TOPCAT & STIL: Starlink Table/VOTable Processing Software.".
Taylor-Blake, B. and Zylka, M. J. 2010. "Prostatic Acid Phosphatase is Expressed in Peptidergic and Nonpeptidergic Nociceptive Neurons of Mice and Rats." PloS One 5 (1): e8674. doi:10.1371/journal.pone.0008674 [doi].
Thompson, J. D., Higgins, D. G., and Gibson, T. J. 1994. "CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice." Nucleic Acids Research 22 (22): 4673-4680.
Traub, L. M. 2009. "Tickets to Ride: Selecting Cargo for Clathrin-Regulated Internalization." Nature Reviews.Molecular Cell Biology 10 (9): 583-596. doi:10.1038/nrm2751 [doi].
Treuting, P. M. and S. Dintzs. 2012. "Salivary Glands." In Comparative Anatomy and Histology. A Mouse and Human Atlas., edited by P. M. Treuting, S. Dintzs, D. Liggitt and C. W. Frevert, 111-120: Elsevier.
Trimble, W. S., Cowan, D. M., and Scheller, R. H. 1988. "VAMP-1: A Synaptic Vesicle-Associated Integral Membrane Protein." Proceedings of the National Academy of Sciences of the United States of America 85 (12): 4538-4542.
Valcour, A. A., Bowers, G. N.,Jr, and McComb, R. B. 1989. "Evaluation of a Kinetic Method for Prostatic Acid Phosphatase with use of Self-Indicating Substrate, 2,6-Dichloro-4-Nitrophenyl Phosphate." Clinical Chemistry 35 (6): 939-945.
Van Etten, R. L. 1982. "Human Prostatic Acid Phosphatase: A Histidine Phosphatase." Annals of the New York Academy of Sciences 390: 27-51.
Vanha-Perttula, T., Niemi, R., and Helminen, H. J. 1972. "Separate Lysosomal and Secretory Acid Phosphatases in the Rat Ventral Prostate." Investigative Urology 9 (4): 345-352.
Varin, M. M., Guerrier, T., Devauchelle-Pensec, V., Jamin, C., Youinou, P., and Pers, J. O. 2012. "In Sjogren's Syndrome, B Lymphocytes Induce Epithelial Cells of Salivary Glands into Apoptosis through Protein Kinase C Delta Activation." Autoimmunity Reviews 11 (4): 252-258. doi:10.1016/j.autrev.2011.10.005; 10.1016/j.autrev.2011.10.005.
Veeramani, S., Yuan, T. C., Chen, S. J., Lin, F. F., Petersen, J. E., Shaheduzzaman, S., Srivastava, S., MacDonald, R. G., and Lin, M. F. 2005. "Cellular Prostatic Acid Phosphatase: A Protein Tyrosine Phosphatase Involved in Androgen-

Bibliography
53
Independent Proliferation of Prostate Cancer." Endocrine-Related Cancer 12 (4): 805-822. doi:12/4/805 [pii].
Vihko, P. 1978a. Human Prostatic Acid Phosphatase and its Radioimmunoassay Acta Universitatis Ouluensis: Medica.
Vihko, P. T., Quintero,I., Rönkö,A. E., Herrala,A., Jäntti,P., Porvari,K., Lindqvist,Y., Kaija,H., Pulkka,A., Vuoristo,J., Sormunen,R., Soini,Y., Halmekytö,M., Jänne,J., Luokkala,T., and Kurkela,R. 2005. "Prostatic Acid Phosphatase (PAP) is PI(3)P-Phosphatase and its Inactivation Leads to Change of Cell Polarity and Invasive Prostate Cancer." Abstract. Proceedings of the AACR Abstract 5239, : 96th Annual Meeting. Anaheim, CA, USA.
Vihko, P. 1978b. "Characterization of the Principal Human Prostatic Acid Phosphatase Isoenzyme, Purified by Affinity Chromatography and Isoelectric Focusing. Part II." Clinical Chemistry 24 (10): 1783-1787.
Vihko, P. 1979. "Human Prostatic Acid Phosphatases: Purification of a Minor Enzyme and Comparisons of the Enzymes." Investigative Urology 16 (5): 349-352.
Vihko, P., Kurkela, R., Porvari, K., Herrala, A., Lindfors, A., Lindqvist, Y., and Schneider, G. 1993. "Rat Acid Phosphatase: Overexpression of Active, Secreted Enzyme by Recombinant Baculovirus-Infected Insect Cells, Molecular Properties, and Crystallization." Proceedings of the National Academy of Sciences of the United States of America 90 (3): 799-803.
von Figura, K. and Hasilik, A. 1986. "Lysosomal Enzymes and their Receptors." Annual Review of Biochemistry 55: 167-193. doi:10.1146/annurev.bi.55.070186.001123 [doi].
Vrljic, M., Strop, P., Ernst, J. A., Sutton, R. B., Chu, S., and Brunger, A. T. 2010. "Molecular Mechanism of the Synaptotagmin-SNARE Interaction in Ca2+-Triggered Vesicle Fusion." Nature Structural & Molecular Biology 17 (3): 325-331. doi:10.1038/nsmb.1764 [doi].
Warde-Farley, D., Donaldson, S. L., Comes, O., Zuberi, K., Badrawi, R., Chao, P., Franz, M., et al. 2010. "The GeneMANIA Prediction Server: Biological Network Integration for Gene Prioritization and Predicting Gene Function." Nucleic Acids Research 38 (Web Server issue): W214-20. doi:10.1093/nar/gkq537; 10.1093/nar/gkq537.
Wileman, T. 2013. "Autophagy as a Defence Against Intracellular Pathogens." Essays in Biochemistry 55: 153-163. doi:10.1042/bse0550153 [doi].
Yu, A., Rual, J. F., Tamai, K., Harada, Y., Vidal, M., He, X., and Kirchhausen, T. 2007. "Association of Dishevelled with the Clathrin AP-2 Adaptor is Required

Bibliography
54
for Frizzled Endocytosis and Planar Cell Polarity Signaling." Developmental Cell 12 (1): 129-141. doi:S1534-5807(06)00467-9 [pii].
Zeeberg, B. R., Feng, W., Wang, G., Wang, M. D., Fojo, A. T., Sunshine, M., Narasimhan, S., et al. 2003. "GoMiner: A Resource for Biological Interpretation of Genomic and Proteomic Data." Genome Biology 4 (4): R28.
Zylka, M. J., Sowa, N. A., Taylor-Blake, B., Twomey, M. A., Herrala, A., Voikar, V., and Vihko, P. 2008. "Prostatic Acid Phosphatase is an Ectonucleotidase and Suppresses Pain by Generating Adenosine." Neuron 60 (1): 111-122. doi:10.1016/j.neuron.2008.08.024.

Prostatic Acid Phosphatase as a Regulator of Endo/Exocytosis and Lysosomal Degradation
MEDICUMDEPARTMENT OF CLINICAL CHEMISTRY AND HAEMATOLOGYFACULTY OF BIOLOGICAL AND ENVIRONMENTAL SCIENCESDOCTORAL PROGRAMME IN INTEGRATIVE LIFE SCIENCEUNIVERSITY OF HELSINKI
CÉSAR L. ARAUJO
dissertationes scholae doctoralis ad sanitatem investigandam universitatis helsinkiensis 7/2016
7/2016Helsinki 2016 ISSN 2342-3161 ISBN 978-951-51-1849-3
CÉ
SA
R L
. AR
AU
JO P
rostatic Acid
Ph
osph
atase as a Regu
lator of En
do/E
xocytosis and
Lysosom
al Degrad
ation
Recent Publications in this Series
94/2015 Jani SaksiStroke-Associated Carotid Atherosclerosis - a Role for FABP4 in Atheroma Vulnerability and Cardiometabolic Risk95/2015 Otto HemminkiCancer Immunotherapy with a Gene Modified Serotype 3 Oncolytic Adenovirus96/2015 Jukka KoskelaIndividual Trajectories in Asthma and COPD: A Longitudinal Perspective to Obstructive Lung Disease97/2015 Miira KlemettiTrends In Obstetric and Perinatal Outcomes of Women with Type 1 Diabetes During 1988-2011 – A Finnish Population-Based Observational Study98/2015 Ileana B. QuinteroPhysiological Functions of Prostatic Acid Phosphatase99/2015 Minna MatikainenLaunching New Products in the Finnish Pharmaceutical Industry: A Relationship Approach100/2015 Päivi Ylä-AnttilaPhagophore Membrane Connections and RAB24 in Autophagy101/2015 Kaisa KyöstiläIdentification of Novel Mutations and Molecular Pathways for Canine Neurodegeneration and Chondrodysplasia102/2015 Emmi JoensuuEpigenetic Alterations in Sporadic and Familial Cancers103/2015 Elina ReponenPreoperative Risk-Assessment Methods, Short-Term Outcome, and Patient Satisfaction in Elective Cranial Neurosurgery104/2015 Riina KandolinCardiac Sarcoidosis in Giant Cell Myocarditis in Finland106/2015 Karmen KappPolyphenolic and Essential Oil Composition of Mentha and Their Antimicrobial Effect107/2015 Dina PopovaNeurophysiological mechanisms of Plasticity Induced in Adult Brain1/2016 Pauliina SaurusRegulation of Podocyte Apoptosis in Diabetic Kidney Disease – Role of SHIP2, PDK1 and CDK22/2016 Sanna ToivonenDerivation of Hepatocyte Like Cells from Human Pluripotent Stem Cells3/2016 Marjaana PeltolaAMIGO-Kv2.1 Potassium Channel Complex: Identification and Association with Schizophrenia-Related Phenotypes4/2016 Niko-Petteri NykänenCellular Physiology and Cell-to-Cell Propagation of Tau in Neurodegeneration: The Impact of Late-Onset Alzheimer’s Disease Susceptibility Genes5/2016 Liisa KorkaloHidden Hunger in Adolescent Mozambican Girls: Dietary Assessment, Micronutrient Status, and Associations between Dietary Diversity and Selected Biomarkers6/2016 Teija OjalaLactobacillus crispatus and Propionibacterium freudenreichii: A Genomic and Transcriptomic View