protein folding, processing and degradation 2 protein folding protein in native state is not static...
TRANSCRIPT

Protein Folding, Processing Protein Folding, Processing and Degradationand Degradation

22
Protein FoldingProtein Folding
• Protein in native state is not staticProtein in native state is not static– 2° structural elements and domains 2° structural elements and domains
movemove
• Function of proteins often dependent Function of proteins often dependent on large conformational changes, on large conformational changes, triggered by ligand bindingtriggered by ligand binding

33
Energy of FoldingEnergy of Folding
• Difference in energy (free energy) Difference in energy (free energy) between folded (native) and unfolded between folded (native) and unfolded (denatured) state is small, 5-15 (denatured) state is small, 5-15 kcal/mol.kcal/mol.
• Two major contributions to energy Two major contributions to energy difference between unfolded and difference between unfolded and folded state, enthalpy and entropy folded state, enthalpy and entropy

44
Free energy of foldingFree energy of folding• Enthalpy (H) is increased upon folding Enthalpy (H) is increased upon folding
– Noncovalent interactions are maximizedNoncovalent interactions are maximized
•Stronger and more frequent in native stateStronger and more frequent in native state– Enthalpy difference can reach several hundred Enthalpy difference can reach several hundred
kcal/molkcal/mol
• Entropy is decreased upon folding Entropy is decreased upon folding – Folding causes one main conformation=highly Folding causes one main conformation=highly
ordered structureordered structure– Entropy difference can reach several hundred Entropy difference can reach several hundred
kcal/molkcal/mol

55
Free energy of foldingFree energy of folding• Enthalpy and entropy differences balance Enthalpy and entropy differences balance
each other, and each other, and G is a small positive G is a small positive number.number.
• Small Small G is necessary because too large a G is necessary because too large a free energy change would mean a very free energy change would mean a very stable protein, one that would never stable protein, one that would never changechange
• However, structural flexibility is important However, structural flexibility is important to protein function, and proteins need to to protein function, and proteins need to be degradedbe degraded

66
Protein foldingProtein folding• For any given protein, there is one For any given protein, there is one
conformation that has significantly lower conformation that has significantly lower free energy than any other statefree energy than any other state
• Achieved through kinetic pathway of Achieved through kinetic pathway of unstable intermediates (not all unstable intermediates (not all intermediates are sampled)intermediates are sampled)
• Assisted by chaperones and protein Assisted by chaperones and protein disulfide isomerases so intermediates are disulfide isomerases so intermediates are not trapped in a local low energy statenot trapped in a local low energy state

77
Pathways to foldingPathways to folding
• There are multiple folding pathways There are multiple folding pathways for some proteins, but a single main for some proteins, but a single main pathway for other proteinspathway for other proteins

88
Transition State ModelTransition State Model
• Free energy barrier separates the D Free energy barrier separates the D (denatured) state from the N (native) state(denatured) state from the N (native) state
• Folding pathway can vary between proteinsFolding pathway can vary between proteins– Can be a single energy barrier with one pathway Can be a single energy barrier with one pathway
or or – A single folding pathway that has sequential A single folding pathway that has sequential
transitional states that have limited flexibilities transitional states that have limited flexibilities along the pathway oralong the pathway or
– Can have multiple transition states with similar Can have multiple transition states with similar energy values and a variety of pathways to get energy values and a variety of pathways to get to the final folded stateto the final folded state

99Fig. 6-39, p.189

1010Fig. 6-40, p.190

1111
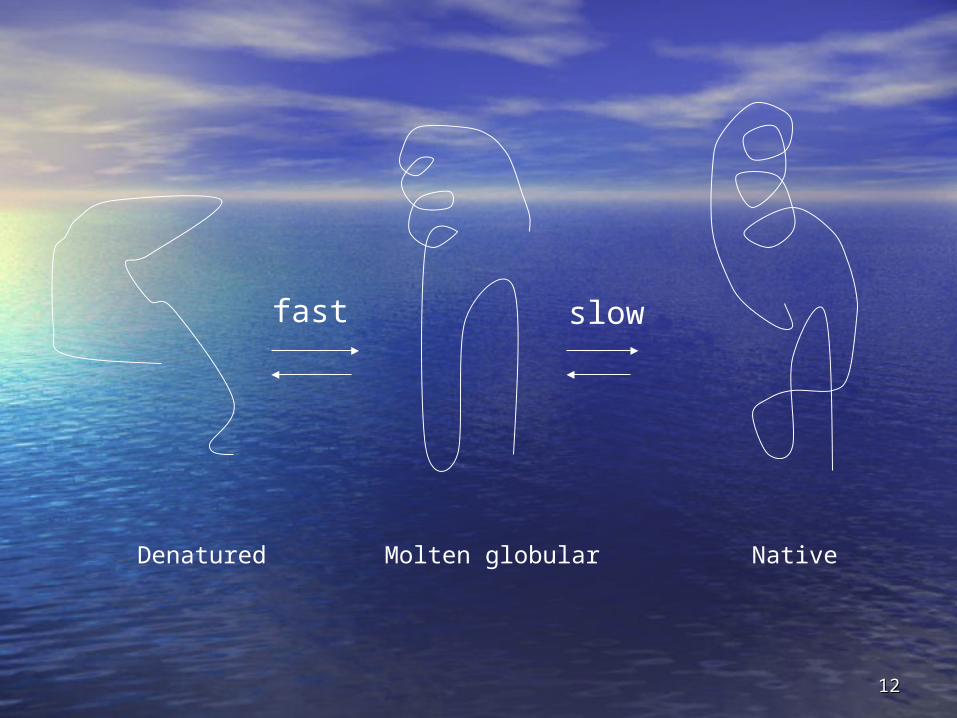
Pathways to folded statePathways to folded state
• Molten globularMolten globular is state in between is state in between unfolded and final folded stateunfolded and final folded state– Can be multiple molten globular states for each Can be multiple molten globular states for each
proteinprotein
• Has helices and sheets, which may be Has helices and sheets, which may be positioned correctlypositioned correctly
• Unfolded loops, less compaction, different Unfolded loops, less compaction, different conformations of surface structuresconformations of surface structures
1212
fast slow
Denatured Molten globular Native

1313Fig. 6-41, p.191

1414
Disulfide bond formationDisulfide bond formation
• Cellular environment is Cellular environment is reducingreducing• Cysteines must be oxidized to form S-Cysteines must be oxidized to form S-
S bondsS bonds• Bacteria use Disulfide bridge-forming Bacteria use Disulfide bridge-forming
protein, protein, Dsb,Dsb, for oxidation in for oxidation in periplasmperiplasm
• In eucaryotes, protein disulfide In eucaryotes, protein disulfide isomerase, isomerase, PDIPDI,, used for oxidation in used for oxidation in the endoplasmic reticulum (ER)the endoplasmic reticulum (ER)

1515
DsbDsb
• Crystal structure shows a thioredoxin-like domain Crystal structure shows a thioredoxin-like domain
• Mechanism of thioredoxin is based on reversible Mechanism of thioredoxin is based on reversible oxidation of two cysteine thiol groups to a oxidation of two cysteine thiol groups to a disulfide--redox-active disulfide bridge is Cys-X-X-disulfide--redox-active disulfide bridge is Cys-X-X-Cys (X any amino acid)Cys (X any amino acid)
• Oxidized Dsb is less stable than reduced form--Oxidized Dsb is less stable than reduced form--– S-S is very reactive, therefore a strong oxidizing agentS-S is very reactive, therefore a strong oxidizing agent

1616
Isomerization of prolinesIsomerization of prolines
• Most peptide bonds are Most peptide bonds are transtrans (100x more stable (100x more stable than than ciscis))
• When second residue is proline, When second residue is proline, transtrans form is only form is only 4x more stable4x more stable
• In native proteins, In native proteins, ciscis-proline peptides are -proline peptides are stabilized by tertiary structure but in unfolded stabilized by tertiary structure but in unfolded state there is an equilibrium between state there is an equilibrium between ciscis and and transtrans isomers isomers
• Cis-transCis-trans isomerization of proline peptides is the isomerization of proline peptides is the rate-limiting step in folding for some proteinsrate-limiting step in folding for some proteins

1717
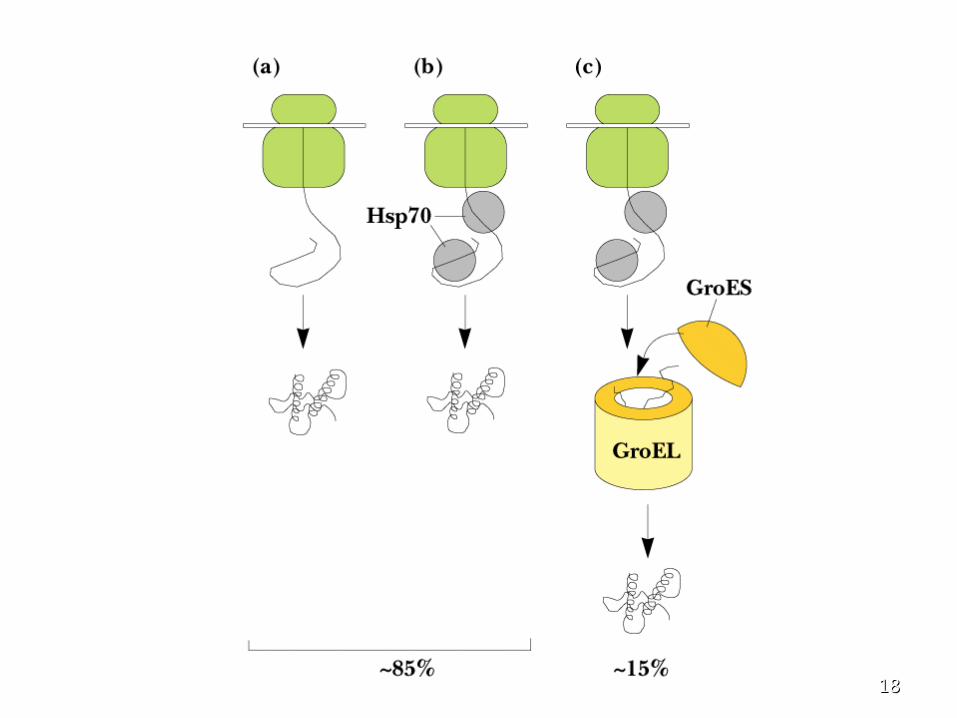
Protein FoldingProtein Folding • Proteins are assisted in folding by Proteins are assisted in folding by
molecular chaperonesmolecular chaperones
• Hsp60Hsp60 ( (chaperonins)chaperonins) Hsp70 and Hsp90Hsp70 and Hsp90 are are three main classes three main classes
• Hsp70 recognizes exposed, unfolded Hsp70 recognizes exposed, unfolded regions of new protein chains - especially regions of new protein chains - especially hydrophobic regions hydrophobic regions
• It binds to these regions, protecting them It binds to these regions, protecting them until productive folding reactions can occuruntil productive folding reactions can occur
1818

1919Fig. 31-2b, p. 1026

2020
The GroES-GroEL ComplexThe GroES-GroEL Complex • The principal chaperonin in The principal chaperonin in E. coliE. coli• GroEL forms two stacked 7-membered rings of GroEL forms two stacked 7-membered rings of
60 kD subunits; GroES is a dome on the top60 kD subunits; GroES is a dome on the top• Nascent protein apparently binds reversibly Nascent protein apparently binds reversibly
many times to the walls of the donut structure, many times to the walls of the donut structure, each time driven by each time driven by ATP hydrolysisATP hydrolysis, eventually , eventually adopting its folded structure, then being adopting its folded structure, then being released from the GroES-GroEL complex released from the GroES-GroEL complex
• RhodaneseRhodanese (as one example) requires hydrolysis (as one example) requires hydrolysis of of 130 ATP130 ATP to reach fully folded state to reach fully folded state

2121Fig. 31-3a, p. 1027

2222Fig. 31-3b, p. 1027

2323Fig. 31-3c, p. 1027

2424
Eucaryotic Hsp90 Eucaryotic Hsp90 ChaperonesChaperones
• Hsp 90 is 1-2% of total cytosolic Hsp 90 is 1-2% of total cytosolic proteinprotein
• Signal transduction molecules such Signal transduction molecules such as tyrosine kinase receptors, steroid as tyrosine kinase receptors, steroid hormone receptors, non-receptor hormone receptors, non-receptor tyrosine kinases are tyrosine kinases are clients clients for Hsp90for Hsp90

2525
Fate of nascent proteinsFate of nascent proteins
• Association of nascent polypeptide Association of nascent polypeptide with various chaperone systems with various chaperone systems commits them to folding pathwayscommits them to folding pathways
• If proteins fail to fold, they are If proteins fail to fold, they are recognized and targeted for recognized and targeted for degradationdegradation

2626
Protein processingProtein processing
• Post-translational modificationsPost-translational modifications are are covalent modifications that alter covalent modifications that alter functionfunction
• Hundreds of amino acid variations Hundreds of amino acid variations have been describedhave been described
• Methylation, phosphorylation, lipidMethylation, phosphorylation, lipid and and carbohydrate additioncarbohydrate addition are a few are a few
• Human proteins number around Human proteins number around 300,000 from 30,000 genes300,000 from 30,000 genes

2727
Proteolytic cleavageProteolytic cleavage
• Why remove amino acids?Why remove amino acids?– Create diversityCreate diversity
•Met-amidopeptidaseMet-amidopeptidase removes Met from removes Met from peptide so that not all amino termini are Metpeptide so that not all amino termini are Met
– Serves as activation mechanismServes as activation mechanism•Metabolically active enzymes are Metabolically active enzymes are pro-pro-
proteinsproteins or or zymogens zymogens such as digestive such as digestive enzymesenzymes
– Involves targeting of proteins to proper Involves targeting of proteins to proper destinationsdestinations

2828
Protein TranslocationProtein TranslocationCommon features in all organismsCommon features in all organisms
• Proteins are made as Proteins are made as preproteinspreproteins containing containing domains that act as sorting signals domains that act as sorting signals
• Signal recognition particles (SRPs)Signal recognition particles (SRPs) recognize the recognize the sorting signals as they emerge from the ribosome sorting signals as they emerge from the ribosome
• Membranes involved in protein translocation have Membranes involved in protein translocation have specific specific signal receptorssignal receptors
• TransloconsTranslocons (selectively permeable protein- (selectively permeable protein-conducting channels) catalyze the movement of the conducting channels) catalyze the movement of the proteins across the membrane with metabolic proteins across the membrane with metabolic energy (ATP, GTP, ion gradients) essential energy (ATP, GTP, ion gradients) essential – Stop-transfer signalsStop-transfer signals allow diffusion of protein into allow diffusion of protein into
membranemembrane
• Preproteins bind to chaperones to stay loosely foldedPreproteins bind to chaperones to stay loosely folded

2929
Prokaryotic Protein Prokaryotic Protein TranslocationTranslocationAn essential process for membrane proteins An essential process for membrane proteins
and secretory proteinsand secretory proteins
• Such proteins are synthesized with a Such proteins are synthesized with a leader leader peptidepeptide, or , or signal sequencesignal sequence of about 16-26 of about 16-26 amino acids amino acids
• The signal sequence has a basic N-terminus, The signal sequence has a basic N-terminus, a central domain of 7-13 hydrophobic a central domain of 7-13 hydrophobic residues, and a nonhelical C-terminus residues, and a nonhelical C-terminus
• The signal sequence The signal sequence directs the newly directs the newly synthesized protein to its proper destinationsynthesized protein to its proper destination

3030
Prokaryotic Protein Prokaryotic Protein TransportTransport
All non-cytoplasmic proteins must be All non-cytoplasmic proteins must be translocated translocated
• The leader peptide retards the folding of the The leader peptide retards the folding of the protein so that molecular chaperone protein so that molecular chaperone proteins can interact with it and direct its proteins can interact with it and direct its folding folding
• The leader peptide also The leader peptide also provides recognition provides recognition signalssignals for the translocation machinery for the translocation machinery
• A A leader peptidaseleader peptidase removes the leader removes the leader sequence when folding and targeting are sequence when folding and targeting are assured assured

3131
Eukaryotic Protein SortingEukaryotic Protein Sorting
Eukaryotic cells contain many membrane-Eukaryotic cells contain many membrane-bounded compartmentsbounded compartments
• Most (but not all) targeting sequences are N-Most (but not all) targeting sequences are N-terminal, terminal, cleaveable presequences cleaveable presequences
• Charge distribution, polarity and secondary Charge distribution, polarity and secondary structurestructure of the signal sequence, rather than a of the signal sequence, rather than a particular sequence, appears to target to particular sequence, appears to target to particular organelles and membranes particular organelles and membranes
• Synthesis of Synthesis of secretory and membrane proteinssecretory and membrane proteins is coupled to translocation across ER is coupled to translocation across ER membranemembrane

3232
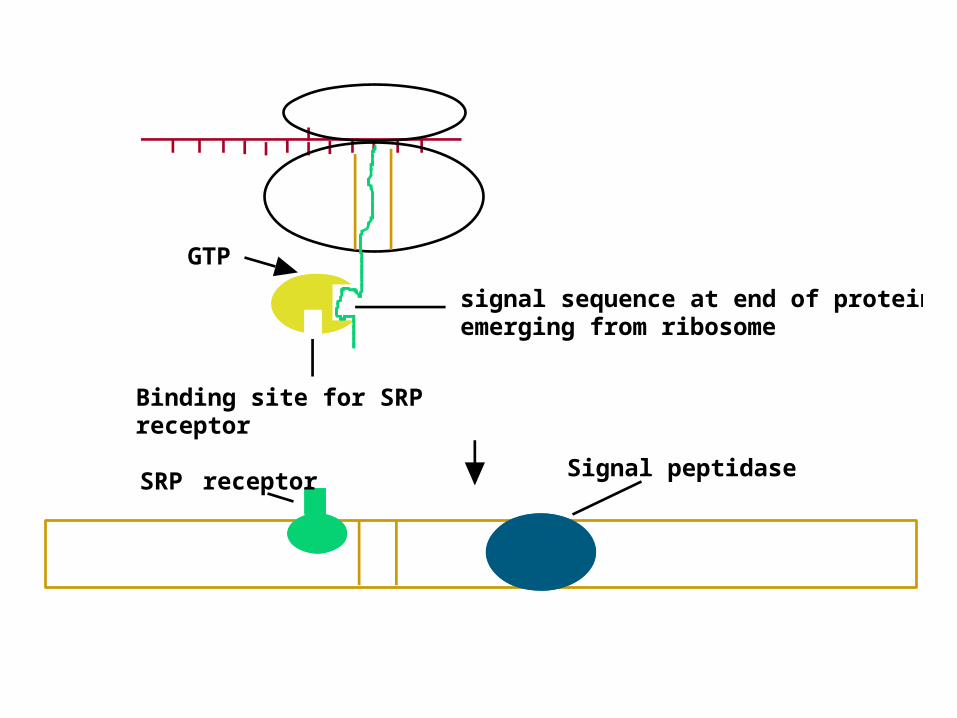
Events at the ER MembraneEvents at the ER Membrane • As the signal sequence emerges from the As the signal sequence emerges from the
ribosome, a ribosome, a signal recognition particle (SRP)signal recognition particle (SRP) finds it and escorts it to the ER membrane finds it and escorts it to the ER membrane
• There it docks with a There it docks with a docking protein or docking protein or signal receptor (SR)signal receptor (SR) - see Figure 31.5 - see Figure 31.5
• SRP dissociates in a SRP dissociates in a GTP-dependent processGTP-dependent process
• Protein synthesis resumes and protein Protein synthesis resumes and protein passes into ER or into ER membrane; signal passes into ER or into ER membrane; signal is cleaved is cleaved

mRNA
encoded signal sequence

signal sequence at end of protein emerging from ribosome
Signal recognition particle (SRP)
signal sequence at end of protein emerging from ribosome
Binding site for SRP receptor
Signal peptidaseSRP receptor
GTP

Signal peptidaseGDP + Pi
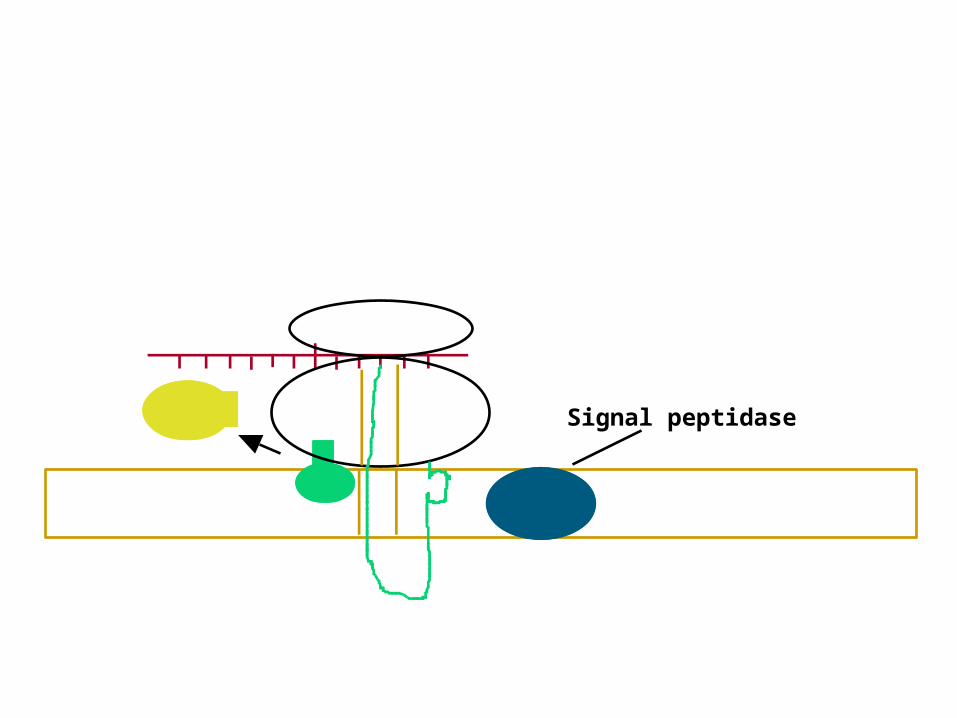
Signal peptidase

Signal peptidase cleaves signal peptide
Ribosome dissociates

Completed polypeptide chain

4040

4141
Signal Recognition ParticleSignal Recognition Particle
• Complex of six polypepides and one Complex of six polypepides and one 300-base RNA molecule300-base RNA molecule
• Signal peptide recognition depends Signal peptide recognition depends on one protein of 54,000 daltons-on one protein of 54,000 daltons-SRP54SRP54
• RNA (7S) is probably structuralRNA (7S) is probably structural
• Removal of RNA disrupts SRPRemoval of RNA disrupts SRP

4242
Mitochondrial protein importMitochondrial protein import
• Mitochondria have two membranes, Mitochondria have two membranes, and two spaces in between the and two spaces in between the membranesmembranes
• Signal sequences are N-terminal, Signal sequences are N-terminal, positively charged regions of 10-70 aapositively charged regions of 10-70 aa
• Form amphipathic Form amphipathic -helices, positive -helices, positive on one side and uncharged, on one side and uncharged, hydrophobic on the other (Fig. 31.6)hydrophobic on the other (Fig. 31.6)

4343
Mitochondrial protein import Mitochondrial protein import (Fig 31.7)(Fig 31.7)• Binding of preprotein to Binding of preprotein to TOM (mitochondrial TOM (mitochondrial
outer membrane translocon)outer membrane translocon)• For outer membrane proteins, transfer to For outer membrane proteins, transfer to
SAM (sorting and assembly complex)SAM (sorting and assembly complex)• Inner membrane proteins traverse TOM and Inner membrane proteins traverse TOM and
are taken up by are taken up by TIM22 (inner mitochondrial TIM22 (inner mitochondrial membrane translocon)membrane translocon)
• Matrix proteins are taken up byMatrix proteins are taken up by TIM23 TIM23 and and transferred across the inner membrane into transferred across the inner membrane into the matrixthe matrix

4444
Protein DegradationProtein Degradation
• Some protein degradation pathways are Some protein degradation pathways are nonspecific - randomly cleaved proteins nonspecific - randomly cleaved proteins seem to be rapidly degraded seem to be rapidly degraded
• However, there is also However, there is also a selective, ATP-a selective, ATP-dependent pathway for degradationdependent pathway for degradation - the - the ubiquitin-mediated pathway ubiquitin-mediated pathway
• Ubiquitin is a highly-conserved, 76 Ubiquitin is a highly-conserved, 76 residue (8.5 kD) protein found widely in residue (8.5 kD) protein found widely in eukaryotes eukaryotes
• Proteins are Proteins are committed to degradationcommitted to degradation by by conjugation with ubiquitinconjugation with ubiquitin

4545
Ubiquitin and DegradationUbiquitin and Degradation Three proteins involved: EThree proteins involved: E11, E, E22 and E and E33
• EE11 is the is the ubiquitin-activating enzymeubiquitin-activating enzyme - it forms - it forms a thioester bond with C-terminal Gly of a thioester bond with C-terminal Gly of ubiquitin ubiquitin
• Ubiquitin is then transferred to a Cys-thiol of Ubiquitin is then transferred to a Cys-thiol of EE22, the , the ubiquitin-carrier proteinubiquitin-carrier protein
• Ligase (ELigase (E33)) selects proteins for degradation. selects proteins for degradation. the Ethe E22-S~ubiquitin complex transfers ubiquitin -S~ubiquitin complex transfers ubiquitin to these selected proteins to these selected proteins
• More than one ubiquitin may be attached to a More than one ubiquitin may be attached to a protein targetprotein target

4646Fig. 31-8, p. 1034
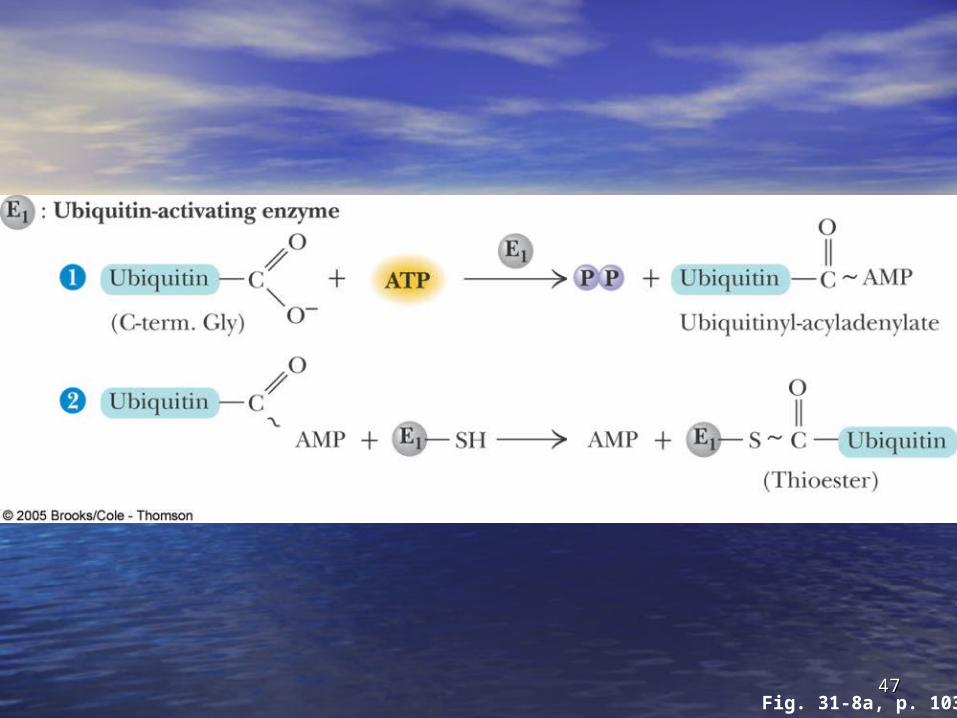
4747Fig. 31-8a, p. 1034

4848Fig. 31-8b, p. 1034

4949Fig. 31-8c, p. 1034

5050Fig. 31-9, p. 1034

5151Fig. 31-10a, p. 1035

5252