psycho-social phenotype and brain n...
TRANSCRIPT

PSYCHO-SOCIAL PHENOTYPE AND BRAIN N-SULFATED HEPARAN
SULFATES IN MECP2 MUTANT MICE
A DISSERTATION SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAI‘I AT MĀNOA IN PARTIAL FULFILLMENT OF THE
REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
IN
PSYCHOLOGY
MAY 2012
By
Brandon L. Pearson
Dissertation Committee:
Robert Blanchard, Chairperson D. Caroline Blanchard
Adrian Dunn Lorey Takahashi Timothy Tricas

ACKNOWLEDGEMENTS
Special thanks to my mother, Theresa Forshee for encouraging my curiosity and to my
wife, Daria Ebneter for her outstanding support and her incredible ability to keep me grounded.
Jaclyn Bettis, Erwin Defensor, Dr. Ksenia Meyza, Sam O’Hanlon, Larry Oasay, Dr.
Roger Pobbe, Amy Vasconcellos and Lace Yamamoto provided assistance with behavioral
studies. Ted Murphy constructed behavioral arenas. Michael Corley, Ashley Jensen, Dr.
Frederic Mercier and Dr. Roger Pobbe assisted with immunohistochemistry. Tina Carvalho of
the Biological Electron Microscope Facility (BEMF) at the University of Hawaii provided critical
confocal training and support. Funded by grant NIH MH081845-01 to Dr. Robert J. Blanchard.
i

ABSTRACT
A variety of inbred and mutant strains of mice exhibit impairments in social behaviors
and restricted, repetitive behaviors; these constitute face validity for autism models. However,
researchers rarely evaluate the psychological state eliciting the impaired social investigation and
there remain no reliable biomarkers for autism. A series of studies was conducted to assess
social behavior in methyl CpG binding protein 2 (MeCP2308/Y) mutant mice, which were
subsequently shown to be hypersocial. Analyses of psychostimulant-based behavioral
responses were assessed in the hypersocial MeCP2 mutant mice and in hypo-social BTBR
T+tf/J mice relative to their respective controls. This body of work suggests that dysregulated
reward processing in candidate mutant mice, and specifically, MeCP2 hypomorphs, may
underlie inflexible and abnormal social behaviors. N-sulfated heparan sulfate alterations in the
brain may underlie brain developmental and behavioral abnormalities in autism. To ascertain
whether MeCP2 mutation influences heparan sulfate proteoglycan systems, fluorescence
immunohistochemistry was performed on wild-type and mutant mouse brains throughout the
neurogenic lateral ventricle subventricular zone and it was shown that, counter to the prediction,
MeCP2308/Y mutants display normal levels of heparan sulfate and laminin extracellular matrix
markers in the neurogenic subventricular zone. Perhaps other brain regions or alternative
markers should be included in future analyses; yet, the overall influence of adult neurogenesis
and other brain plasticity processes in autism-relevant behavior deficits has not been fully
characterized. This body of work utilizes multiple levels of analyses in an attempt to elucidate
neurobiological and psychological contributions to abnormal and autism-relevant behavioral
disturbances.
ii

Table of Contents
CHAPTER 1: INTRODUCTION 1
1.1 MeCP2308/Y SOCIABILITY AND STEREOTYPY PHENOTYPE 2
1.2 INTRODUCTION 2
1.3 MATERIALS AND METHODS 4
1.3.1 Experimental Subjects 5
1.3.2 Experimental Design 5
1.3.3 Visible Burrow System 5
1.3.4 Three-Chamber Social Approach 6
1.3.5 Autogrooming 7
1.3.6 Repetitive Novel Object Contact Task/ Locomotion 8
1.3.7 Social Proximity 9
1.3.8 Urinary Scent Marking 9
1.3.9 Resident Intruder 10
1.3.10 Statistical Analyses 10
1.4 RESULTS 11
1.4.1 Body Weight 11
1.4.2 Visible Burrow System 11
1.4.3 Three-Chamber Social Approach 12
1.4.4 Autogrooming 12
1.4.5 Repetitive Novel Object Contact Task/ Locomotion 13
1.4.6 Social Proximity 13
1.4.7 Urinary Scent Marking 13
1.4.8 Resident Intruder 13
1.5 DISCUSSION 14
iii

CHAPTER 2: EMOTIONAL CHARACTERISTICS OF MeCP2308/Y MUTANTS 25
2.1 INTRODUCTION 25
2.2 MATERIALS AND METHODS 26
2.2.1 Experimental Subjects 26
2.2.2 Experimental Design 26
2.2.3 Elevated Plus-Maze 27
2.2.4 Elevated Zero-Maze 27
2.2.5 Mouse Defense Test Battery 28
2.2.6 Female Urine Sniffing Test 29
2.3 RESULTS 29
2.3.1 Elevated Plus Maze 29
2.3.2 Elevated Zero Maze 30
2.3.3 Mouse Defense Test Battery 30
2.3.4 Female Urine Sniffing Test 30
2.4 DISCUSSION 30
CHAPTER 3: REWARD SYSTEMS IN MOUSE MODELS OF AUTISM 35
3.1 INTRODUCTION 36
3.2 MATERIALS AND METHODS 37
3.2.1 Experimental Subjects 37
3.2.2 Social Conditioned Place Preference 39
3.2.3 Social Interactions 39
3.2.4 Autogrooming 39
3.2.5 Statistical Analysis 40
3.3 RESULTS 40
iv

3.3.1 Social Conditioned Place Preference 40
3.3.2 Social Interactions 41
3.3.3 Autogrooming 41
3.4 DISCUSSION 42
3.5 PSYCOSTIMULANT REACTIVITY 48
3.5.1 Reward Tasks in MeCP2308/Y Mutants 50
3.5.2 Psychostimulant Response in Low-Social Mice 50
3.6 DISCUSSION 53
CHAPTER 4: N-SULFATED HEPARAN SULFATE ABUNDANCE AND DISTRIBUTION IN
MeCP2308/Y MICE 54
4.1 INTRODUCTION 54
4.2 MATERIALS AND METHODS 55
4.2.1 Experimental Subjects 55
4.2.2 Dual-Channel Indirect Fluorescent Immunohistochemistry 55
4.3 RESULTS 56
4.3.1 Laminin 56
4.3.2 Heparan Sulfate 56
4.3.3 Heparan/Laminin Ratio 56
4.4 DISCUSSION 57
CHAPTER 5: CONCLUSIONS AND FUTURE DIRECTIONS 61
5.1 NEUROBIOLOGY OF SOCIABILITY 61
5.2 THE ASSOCIATIONS OF NEURODEVELOPMENT AND NEUROPLASTICITY
WITH BEHAVIOR 62
v

REFERENCES 67
vi

LIST OF TABLES
Table 1. Review of MeCP2 Mutant Rodent Studies Assessing Social Behavior 64
Table 2. ANOVA Results for MeCP2308/Y in the Visible Burrow System 65
Table 3. Comparison of MeCP2308/Y Mutant and Wild-type Mice in the Mouse Defense
Test Battery 66
vii

LIST OF FIGURES
Figure 1. MeCP2308/Y Mutant Mice Show Increased Affiliation in the Visible Burrow
System 18
Figure 2. Three-Chamber Sociability in MeCP2308/Y Mutant and Wild-Type
Littermates 19
Figure 3. Subtle Alterations in Patterns of Autogrooming in MeCP2308/Y Mutant Mice 20
Figure 4. No Difference in MeCP2 Genotype-Related Performance in the Repetitive
Novel Object Contact Task 21
Figure 5. Patterns of Social Interaction in the Social Proximity Task 22
Figure 6. Elevated Scent Marking in MeCP2308/Y Mutant Mice 23
Figure 7. Comparison of Agonistic Behavior of MeCP2308/Y Mutant and Wild-Type Mice in
the Resident-Intruder Task 24
Figure 8. Elevated Plus-Maze (EPM) testing of MeCP2308/Y and Wild-Type
Littermates 33
Figure 9. Elevated Zero-Maze (EZM) Testing of MeCP2308/Y and Wild-Type
Littermates 35
Figure 10. Social Conditioned Place Preference Testing in B6 and BTBR Mice 45
Figure 11. Scan Sampling of Social Conditioning Sessions in B6 and BTBR Mice 46
Figure 12. Assessment of Autogrooming Behavior Before and After Social Conditioning
During Exploration 47
Figure 13. Cocaine-Induced Conditioned Place Preference Testing in B6 and BTBR
Mice 49
Figure 14. Locomotor Response to Repeated Cocaine Administration in MeCP2308/Y
Mutant and Wild-Type Mice 51
Figure 15. Acute Locomotor Response to IP Cocaine in B6 and BTBR Mice 52
viii

ix
Figure 16. Representative Confocal Fluorescent Immunohistochemistry Micrographs of
Mouse Brain Lateral Ventricle Immunolabeled for Laminin and Heparan Sulfate 59
Figure 17. Immunohistochemical Analyses of Brain Extracellular Matrix Markers Laminin
and Heparan Sulfate in MeCP2308/Y Wild-Type and Mutant Mice 60

1
CHAPTER 1: INTRODUCTION
Autism spectrum disorders (ASD) are a group of pervasive developmental
abnormalities characterized by three clusters of symptoms: deficits in reciprocal social
behaviors, impaired communication, and restricted interests and repetitive behaviors
(APA 2000). Though overall, ASD is highly heritable, the disorder is characterized by a
pronounced heterogeneity (Bill & Geschwind 2009). Less than 20% of ASD diagnoses
stem from known chromosomal or allelic disruptions (Benvenuto et al, 2009). Therefore,
most cases of ASD are idiopathic, with no specific endophenotypes or biomarkers to
aide in diagnosis or development of improved therapeutics. The diverse and complicated
pathophysiology of ASD potentially leads to an over-reliance on behavioral symptoms to
identify cases of non-syndromal neurodevelopmental disorders. A persistent goal of
clinicians and pre-clinical scientists will be to identify physiological abnormalities that are
present in ASD.
ASD disorders such as Rett syndrome and Fragile X mental retardation are
associated with known genetic mutations (MECP2 and FMR1, respectively), while others
such as Angelman syndrome, 15 q11-15 duplication syndrome and Williams syndrome
result from chromosome region deletions, duplications, or inversions across multiple
genes (i.e. Benvenuto et al, 2009). Since mice almost universally possess homologues
to these loci, knockout strategies have been performed and have yielded substantial
progress in the comprehension of these diseases. Much of the work presented here has
utilized such mutant mice.
In order for rodent models to be informative, they must exhibit reliable
approximations of the human condition. What’s more, the behavioral variables of the
rodent testing must be appropriate to the symptomatology of the disease. A variety of
rodent models have been developed which, on the surface, display many autism-like
characteristics (Moy & Nadler 2008). However, in order to make progress in revealing
consistent neurobiological markers for autism in rodent models, the systems underlying
the core deficits in ASD must be realized.
The following set of studies describes an innovative approach to assess explicit
components of social behavior in mice. These assessments encompass analyses of
emotional contributions to variation in affiliative social behavior with particular emphasis
on motivation. These approaches are based on the notion that social deficits are
maintained by alteration in social motivation, an impairment directly implicated in ASD

2
(Dawson et al, 1998; Dawson et al, 2005; Schultz 2005). Impaired social motivation can
encompass a lack of reinforcement for social interactions or a lack of reward for attention
to social stimuli (Scott-Van Zeeland et al, 2010). With regards to pre-clinical models and
experimental psychopharmacology, analyses of social reward mechanisms might be a
particularly suitable approach. In order to test for social motivation abnormalities, studies
were performed to compare mice with known social deficits to those with normal social
behavior, and to MeCP2308/Y mutant mice which were revealed to possess clear
enhancement of social behavior. Although this work could, and should be expanded in a
variety of directions, the final study assessed what is proving to be a potential biomarker
of altered brain function in animal models of ASD.
Despite vast advances in autism research, ASD remains a syndrome
characterized by behavioral impairments; no consistent diagnostic or prognostic
biomarkers have been identified. Identification is hindered by the fact that autism is not a
singular construct, but rather, a collection of neurodevelopmental abnormalities that
manifest as social impairments on cognitive and behavioral restrictions (Walsh et al,
2011). A potential improvement to pre-clinical approaches to the comprehension of ASD
will be to reveal the basic brain substrates of relevant symptoms in rodent models.
1.1 MeCP2308/Y Sociability and Stereotypy Phenotype
1.2 INTRODUCTION
Mutations of the MECP2 gene underlie the majority of cases of Rett syndrome
(Amir et al, 1999; Bienvenu et al, 2000). Investigations are revealing the complex
functions of this gene in intra- and inter-generational epigenetic regulation in non-
disease states, and in somatic and psychiatric disorders. The gene’s protein product,
methyl-CpG binding protein 2 (MeCP2), plays a prominent role in gene regulation due to
its recognition of promoter-specific methylated CpG sites, and its subsequent
interactions with co-repressors (Chahrour et al, 2008; Jørgensen & Bird, 2002; Lewis et
al, 1992; Samaco et al, 2009), as well as its influences on alternative splicing of mRNA
(Young et al, 2005) and microRNA (Wu et al, 2010). Females with MECP2 mutations
display a wide range of symptom severity, due to patterns of X-chromosome inactivation
(XCI), which in-turn leads to mosaicism (Braunschweig et al, 2004) and subsequent
complications in the clinical characterization as well as the generation of informative
mouse models. Mutations of MECP2 were previously considered embryonic lethal for
hemizygous males, but recently, MECP2 mutations have been noted in human males

3
and are associated with mental retardation, movement abnormalities, and other related
symptoms (Clayton-Smith et al, 2000; Dotti et al, 2002; Kleefstra et al, 2002; Orrico et al,
2000). Evidence is accumulating that partial loss-of function mutations of MECP2 may
contribute to neurodevelopmental and social disorders (Samaco et al, 2008).
Rett syndrome is characterized by normal early development, followed by a
pronounced regression at about 6-18 months in age with loss of verbal functions,
epilepsy, gastro-intestinal problems, stereotypies, and severe musculo-skeletal
deformations (Hagberg et al, 1983). Rett syndrome is classified as a pervasive
developmental disorder and is often considered an Autism Spectrum Disorder (ASD)
due to impairments in social communication and presence of restricted-repetitive
behavior (APA, 2000). Although Rett syndrome is comparatively rare (approximately
1:10,000-15,000, Hagberg et al, 1985) compared to ASD (1:110, CDC 2009) there has
been an upsurge of pre-clinical studies of single-gene and other ASD candidate mouse
models under the ultimate impetus to reveal the neural genetics and molecular signature
of developmental disorders affecting social behavior. These mouse models may play an
important role in elucidating some of the biology underlying ASD.
A variety of mutant MeCP2 mice have been engineered to study the
pathophysiology resulting from mutations in this gene (Ricceri et al, 2008). Newer cell
and brain region-specific null mice are proving valuable in the characterization of
pathways responsible for the phenotypic alterations associate with normal and perturbed
MeCP2 expression. Global, whole gene null MeCP2 male mice exhibit severe symptoms
such as seizures and reduced survivability (Chen et al, 2001; Guy et al, 2001; Pelka et
al, 2006). Truncated and conditional knockouts show a range of symptom severity, age
of onset, and mortality (see Calfa et al, 2011 for an exhaustive review).
Behavioral investigations have revealed an apparent decrease in social
interactions indexed by impaired nest building and social avoidance of an unfamiliar
stimulus mouse behind a partition within the home cage in MeCP2308/Y truncated protein
mutants (Moretti et al, 2005). Gemelli et al, (2006) demonstrated that conditional
postnatal knockout MeCP2 mice spent less time interacting with an unfamiliar mouse
located behind a wire mesh enclosure in an unfamiliar arena, relative to wild-type
littermates. Amygdala-specific siRNA knockdown of MeCP2 did not influence three-
chamber social approach behaviors but impaired juvenile play behaviors in male rats
(Kurian et al, 2008). With respect to social communication, MeCP2308/Y mutant mouse

4
pups show impaired ultrasonic vocalization calls upon separation from their mother (De
Filippis et al, 2010). These reductions in social behaviors in MeCP2 mutants generally
have not been attributable to impaired social recognition abilities (Ricceri et al, 2008).
In contrast with these findings, a number of recent studies suggest that MeCP2
mutations in rodent models may be associated with increased sociability. Three-
chamber social approach behavior was shown to be enhanced rather than decreased in
the Mecp21lox mutant (Schaevitz et al, 2010). Similarly, MeCP2 conditional knockout
mice (Sim1 –cre BAC transgenics) and their controls (Mecp2flox/+), who also show
reduced MeCP2 expression, both show increased sociability as indexed by more
proportional time in proximity to familiar and unfamiliar stimulus mice behind a wire-
mesh enclosure (Fyffe et al, 2008; Samaco et al, 2008, 2009). A recent report has
indicated that GABA-ergic neuron MeCP2 mutants show enhanced social interactions in
the partition test and in a modified three-chamber task (Chao et al, 2010). Mecp2llooxx//YY
hypomorphs have similar social approach compared to that of wild-types, but show an
apparent augmentation of social-novelty preference (Kerr et al, 2008). Table 1 provides
an overview of studies revealing alterations in social behavior in targeted mutations of
MeCP2 in rodents. Studies of MeCP2 mutant mice with no social behavior analyses
were not included. These studies indicate that, at a minimum, MeCP2 may be an
important gene for the investigation of mechanisms underlying social behaviors.
The purpose of the current study was to perform analyses of social behavior with
attempts to differentiate pro-social from aggressive or sexual motivations in social
interactions. A battery of behavioral tests in male MeCP2 wild-type and their hemizygous
mutant littermates was performed. The MeCP2308/Y mouse exhibits the same Rett
syndrome-like abnormalities as the whole gene knockouts, but with improved
background strain survivability and a later age of symptom onset (Shahbazian et al,
2002). This permitted the assessment of adult mice prior to the onset of severe sleep,
neurological, and musculo-skeletal deficits. The tests used were designed to reveal
subtle and overt distinctions in social interactions and repetitive behavior, which have
proven to be relevant variables for mouse models of neurodevelopmental disorders
(Arakawa et al, 2007; Defensor et al, 2011; Moy et al, 2004; Pearson et al, 2010, 2011;
Pobbe et al, 2010).
1.3 MATERIALS AND METHODS

5
1.3.1 Experimental Subjects
Experimental animals were bred from C57BL/6J-backcrossed stock obtained
from The Jackson Laboratory (B6.129S-Mecp2tm1Hzo/J, stock # 005439) and bred from
heterozygous mutant dams and hemizygous father sibling pairs. Stimulus mice used for
social behavior tests were adult CD-1 mice bred in-house from stock obtained from
Charles River Labs, and C57BL/6J (B6) mice bred from stock obtained from The
Jackson Laboratory. Mutant mouse genotype was determined according to the PCR
parameters obtained from The Jackson Laboratory with purified DNA collected from tail
biopsy after weaning at post-natal day 25. Since MeCP2 is an X-linked gene, only wild-
type (y/+) and hemizygous (y/-) males were obtained and compared in behavioral
testing. Mice were housed with up to five same-sex littermates under a 12-h light/dark
schedule with lights on at 0600h. Mice had ad libitum access to tap water and standard
laboratory rodent diet. Mice were 10-13 weeks old at the beginning of behavioral studies.
All procedures were performed according to protocols approved by the University of
Hawaii Laboratory Animal Service Institutional Animal Care and Use Committee.
1.3.2 Experimental Design
In total, nine mice per genotype were used as subjects. Mice were subjected to a
series of behavioral tests in a sequence intended to prioritize both the accumulating
effects of multiple testing and the influence of housing and experience on the measures.
All mice were weighed prior to the first behavioral test, and weighed again after testing
for urinary scent marking to determine baseline weight differences and weight gain
trajectories. One mutant animal showing degenerative features, severe weight loss and
seizures and was euthanized after the scent marking test; its data were excluded from
scent marking scores. No other animals showed complications during testing. One
mutant Visible Burrow System (VBS) colony (n=3) was not included in the analyses due
to a recorder error. Animals were moved to the behavioral testing room at least 30
minutes prior to testing and all arenas and equipment were cleaned with 70% ethanol
and dried completely between mice. Unless otherwise noted, testing was performed
under ambient fluorescent lighting between 0900 and 1800 hours. Temperature (22±1 ̊C)
and humidity (50-70%) were controlled in the experimental room.
1.3.3 Visible Burrow System (VBS)

6
Three days prior to behavioral testing, mice were anesthetized with an IP
injection of 12.5mg/ml of Avertin (2,2,2-Tribromoethanol, Sigma-Aldrich, St. Louis, MO)
at a dose of 336 mg/kg. Once mice were unresponsive, peroxide based hair bleach was
applied to the dorsal coat of each mouse so they were distinguishable in videotape
analysis. Mice were allowed to recover two days prior to being placed in the VBS
according to procedures outlined previously (Pobbe et al, 2010). Briefly, each colony
was housed in a rectangular, galvanized metal bin, 86 × 61 × 26 cm (H). The colony
arena consisted of three chambers, each 12 × 7 × 6 cm (H), which were positioned
behind a barrier wall extending across a short width (61 cm) of the bin, 30 cm from the
end wall. This wall separated an open surface area (30 × 61 × 26cm [H]) from the
chambers in the other compartment. These chambers were connected to the wall via
clear Plexiglas tubes 5 cm in diameter. Two of the three chambers, each connected to
the surface area via a Z shaped tube, were connected to each other via a straight clear
Plexiglas tube. The third chamber was connected only to the surface via a straight tube.
The animals could pass freely between each chamber and the surface area, or between
the two connected chambers, by these tubes. The experimental room was maintained
on a 12-h light/dark cycle (lights on at 0600h), being illuminated by fluorescent lamps
during the light period and by infrared light during the dark phase. An assistant who was
blind to genotype scored VBS videotapes by sub-sampling 30 second intervals every 10
minutes for the first 4 hours of the dark and light phases. The observer manually
recorded all occurrences of Approach Front, Approach Rear, Flight, Chase/Follow,
Vigorous behavior, Autogroom, Allogroom, and Huddle. Approach [to the] Front/Back of
another animal was defined in terms of a line bisecting the approached mouse,
perpendicular to the long axis of its body. Flight was defined as rapid locomotion away
from an approaching animal; Chase/Follow was rapid locomotion toward another animal,
or a slow approach toward an animal that was moving away. Vigorous behavior was
high-intensity chase, bite, or attack. Autogrooming was lick or rub self while
Allogrooming is lick or rub with paws of another animal. Huddle was characterized by a
mouse lying in contact with another animal for more than 10 seconds of the 30-second
time sample. Data were summarized separately for the dark and light phases, and the
mean frequency of each behavior for each genotype was compared across the three
days of testing.
1.3.4 Three-Chamber Social Approach

7
Twenty-four hours after removal from the VBS, mice were tested for social
approach behavior in the three chamber apparatus, which was constructed according to
published studies (Moy et al, 2004). Initially, mice are placed into the center of the
divided 41 x 70 x 28 cm (H) apparatus, which contained two empty, inverted wire cups
(Galaxy Pencil/Utility Cup, Spectrum Diversified Designs, Inc., Streetsboro, OH). Empty
glass jars of the same diameter were placed on top of the base of the wire cup to
prevent movement of the enclosures, or escape by stimulus mice. For the habituation
phase, the sliding doors were elevated and the mouse was permitted ten minutes to
explore the three chambers. At the end of the ten minute habituation session, mice were
placed back into the middle of the apparatus, the sliding doors were lowered, and an
unfamiliar adult (57-83 day old) male CD-1 mouse was placed into one of two outer wire
cups, and the doors were again lifted and the mouse was permitted to explore the entire
apparatus for ten minutes; this constituted the sociability phase. The time spent in each
compartment during both sessions was collected in real-time with two stopwatches by a
single observer who was blind to the genotype of the subject. During both the
habituation and sociability phases, cameras were mounted in front of both outer
compartments and connected to a DVD recorder. The frequency and duration of Rear,
Autogroom, Contact (with the stimulus cup), Sniff, Stretch-Attend, Quick-Withdraw, and
Nose-to-Nose were scored off-line using Noldus Observer software (Noldus Information
Technology, Wageningen, The Netherlands) for each of the two outer compartments
during the sociability phase for each subject.
1.3.5 Autogrooming
Detailed grooming analyses were performed as previously described (Pearson et
al, 2011). Mice were placed in a 14 x 7 x 30 (H) cm Plexiglas chamber for 20 minutes
under normal fluorescent lighting. An aluminum lid that permitted air circulation, but
prevented escape, was placed over the top. Two digital cameras were used to collect
videotapes from the front and side aspect so that the mouse’s grooming behavior was
always visible. Recorded DVDs were scored using Noldus software for the frequency
and duration of Paw Licking, Head Washing, Body Grooming, Leg Licking, and
Tail/Genital grooming. In addition to the collection of frequency and duration of body
directed grooming, the following variables were determined according to Kalueff et al,
(2007). A Bout was defined as at least one episode of any category of grooming, or an
uninterrupted sequence of grooming types. Bouts were divided by at least 6 seconds of

8
inactivity or by an activity other than grooming. An Interrupted Bout was defined as a
grooming bout that is interrupted by less than 6 seconds; the proportion of bouts that
were interrupted was calculated as (interrupted bouts/total bouts)*100. Transitions were
transfers between regional grooming subtypes. Incorrect transitions were transfers,
which did not follow the cephalo-caudal progression (0-No Grooming, 1-Paw Licking, 2-
Head Wash, 3-Body Groom, 4-Leg Licking, 5-Tail/Genital). Proportion of incorrect
transitions was calculated as (incorrect transitions/total transitions)*100.
1.3.6 Repetitive Novel Object Contact Task/Locomotion
On the day following the autogrooming analyses, mice were relocated to the
behavior room at least 30 minutes before habituation and testing for the repetitive novel
object contact task (Pearson et al, 2011). The habituation session consisted of placing a
mouse in a clean standard mouse cage containing 1 cm of bedding. No lid, water bottle,
or food hopper was present. A micro-isolator lid was modified by removing the filter
element and frame, and thick gauge wire bisecting both horizontal planes of the lid were
added which divided the overhead image into four equal sized compartments. The
number of transitions between quadrants and amount of time spent within each quadrant
was scored for a 10-minute habituation session under normal fluorescent illumination
using Noldus Observer software. This permitted assessment of any baseline differences
in motor activation or exploration tendencies of the genotypes.
On the following day, and at the same time of day as the prior habituation
session, mice were placed into the same apparatus which contained four small plastic
objects (a multicolored 3 cm long arrangement of Lego blocks, a green 4 cm long jacks
piece, a 1.5 cm3 multicolored die, and a 3.5 cm long white and red bowling pin) arranged
2 cm from the four corners. Video was collected for ten minutes and all instances of
investigation of the objects, and the order in which they were investigated, were scored
offline. Investigation was defined as clear facial or vibrissae contact with or burying of
the novel objects; merely passing or pausing by an object was insufficient for
investigation. The occurrence of repetitive contact with three and four toys and the
frequency of times that the mice buried each object were counted. Total frequency of
contact with each of the four toys, and the total number of burying episodes were also
calculated. In order to determine if there was a strain effect on the tendency to display
preferences for particular toys, the frequencies of contact with each object were ranked

9
in decreasing order from maximum to minimum preference (contact) values for each
subject, and the frequencies were averaged by genotype and compared.
1.3.7 Social Proximity
Social proximity testing was conducted in a clear rectangular chamber (7 ×14 ×
30 cm H) constructed of acrylic plastic according to previously reported parameters
(Defensor et al, 2011). The dimensions of this arena were identical to that used for
analyses of individual autogrooming behavior. For testing, the subject mouse and an
unfamiliar 5-7 month old male B6 mouse were placed into the chamber simultaneously
and an aluminum lid was placed over the top to prevent escape. Video from two
cameras providing front and side views was transferred to a video merge processor,
which combined both channels into a single side-by-side output. The availability of both
views aided in the discrimination of behaviors by reducing occlusion of one animal from
view by the other. The output from the video processor displaying both the front and side
view was transmitted to a DVD recorder for storage and subsequent analysis. The
frequencies of the following behaviors were manually quantified by an observer blind to
the subject’s genotype:
Nose-to-Nose- subject’s nose tip and/or vibrissae contact the nose tip
and/or vibrissae of the other mouse.
Nose-to-Anus- subject’s nose or vibrissae contacts the base of the tail or
anogenital region of the other mouse.
Crawl Over- subject’s forelimbs cross the midline of the dorsal surface of
the other mouse.
Crawl Under- subject’s head goes under the ventral surface of the other
mouse to a depth of at least the ears of the subject animal crossing the
midline of the other mouse’s body.
Upright- subject displays a reared posture oriented towards the other
mouse with head and/or vibrissae contact.
1.3.8 Urinary Scent Marking
Baseline scent marking in a divided arena was characterized on the first day of
testing, after seven days of single housing. Previous studies have demonstrated that
single housing is critical to establishing motivation to engage in detectable scent marking

10
(Arakawa et al, 2009). The scent-marking arena was an inverted rat cage with a steel
mesh divider wall installed to bisect the arena. This apparatus was placed on top of a 30
x 45 cm section of drawing paper, and the subject mouse was placed on one side for a
20-minute baseline session. Twenty-four hours later, the mouse was tested for urinary
scent marking to an unfamiliar CD-1 (91-103 days old) mouse stimulus placed
immediately before in the opposite half of the arena. At the end of the baseline and
social marking sessions the mice were removed and the number of fecal boli counted.
The placement of the compartments was marked and the paper was allowed to dry
overnight. The paper was then fixed and stained with a 6% solution of ninhydrin (Fisher)
in methanol and dried. To quantify the amount of urinary scent marking, a 1x1 cm
printed transparency grid was placed over the paper and the number of squares
containing a stained mark was counted manually by an assistant blind to the genotype of
the subject.
1.3.9 Resident Intruder
An unfamiliar, ten month old intruder B6 mouse was introduced into a resident
MeCP2 mutant or wild-type home cage, and the interactions between those animals
were recorded for five minutes. The latency to and duration of agonistic attack by the
resident was scored. Additionally, the frequency and duration of the following behaviors
was quantified: Face Sniff, Body Sniff, Anogenital Sniff, Tail Sniff, Vigorous Allogroom,
and Following. Mice analyzed in this test, having already been assessed in all of the
previous behavioral tasks were then euthanized by CO2 inhalation.
1.3.10 Statistical Analyses
Two-way repeated-measures analyses of variance (ANOVA) were performed to
compare body weights. Genotype was the between-subjects factor and the day of weight
measurement was the within-subjects repeated-measures factor. Two-way repeated-
measures ANOVAs were performed for data obtained from the VBS, with genotype as a
between-subjects factor, and day as the repeated-measures, within-subjects factor.
Separate analyses were performed across means from the dark and light phases.
Similarly, for the Three-Chamber Social Approach test, two-way repeated-measures
ANOVAs were performed for the duration of time spent within the two outer stimulus
chambers, and the frequency and duration values for each behavioral category except
that stimulus compartment (empty cup vs stimulus mouse) was the within-subjects
factor. Two-way repeated measures ANOVAs were applied to assess any differences in

11
the amount of time spent in each of the four compartments during the habituation phase
of the repetitive novel object investigation task and to compare the number of scent
marks during both scent marking conditions. For all ANOVA tests Bonferroni post-hoc
analyses were conducted to reveal any significant effects of genotype across the within-
subjects factor when significant main effects or interactions resulted. Independent t-tests
were performed to compare average frequencies and durations of each behavior
category in the autogrooming analysis, the social proximity task, the resident-intruder
test, and the number of lines crossed during the habituation phase of the repetitive novel
object contact task. Similarly, for the Three-Chamber Social Approach Task, the mean
frequency and duration of Stretch-Attend, Quick Withdraw, and Nose-to-Nose for each
genotype was compared with unpaired t-tests. When assumptions were violated for
these comparisons (non-normal distributions or unequal variances), nonparametric
Mann-Whitney U tests were performed instead. Statistica (v.6) and GraphPad PRISM
(v.4) software programs were used for statistical analyses and figures.
1.4 RESULTS
1.4.1 Body Weight
All mice were weighed before anesthetization for fur bleaching and on the
second day of scent marking. Mean ± 1 S.E.M. body weights before the VBS are as
follows: wild-type mice weighed 23.2 ± 0.6 g whereas mutant mice weighed 26.0 ± 0.7 g.
After the second scent marking session, wild-type mice weighed 22.7 ± 0.5 g and
hemizygous mice weighed 23.6 ± 1.0 g. A two-way repeated measures ANOVA revealed
no significant main effect of Genotype [F(1,16)=3.635, p=0.075], but a significant main
effect for Day [F(1,16)=16.21, p=0.001] and a significant Genotype x Day interaction
[F(1,16)=6.892, p=0.018, data not shown]. Post-hoc tests failed to indicate any
significant differences between genotypes within each day of weight measurement.
Hemizygous mice weighed slightly more at the beginning of experimentation, but no
significant weight differences were noted towards the end of behavioral assessments.
1.4.2 Visible Burrow System
Table 2 provides statistics for two-way repeated measures ANOVA for VBS
colonies for each behavior during light and dark phases. Figure 1 displays Mean ± 1
S.E.M frequencies of all behaviors in the VBS across the dark (upper panel) and light
(lower panel) phases for hemizygous and wild-type MeCP2 mice. Significant genotype

12
differences were of particular note. During the light phase, mutant mice engaged in
higher rates of frontal-oriented approaches and they also made fewer rear-oriented
approaches during the dark light cycle. Hemizygous mice had lower frequencies of
autogrooming during the light phase, with significantly more allogrooming in the dark
phase. Finally, hemizygous males showed increased rates of huddling in the dark phase.
1.4.3 Three-Chamber Social Approach
Mice displayed no significant side preferences in the habituation phase [WT,
t=1.529, p=0.146; KO, t=-0.910, p=0.376, data not shown]. Figure 2a displays the
amount of time spent in each compartment of the three-chamber social approach arena.
Duration of time spent in the center compartment is displayed for reference but was not
included in analyses. During the sociability phase, two-way ANOVA revealed a
significant main effect for Stimulus [F(1,32)=22.44, p<0.0001] indicating that both
genotypes spent a greater amount of time in the chamber associated with the stimulus
mouse compared to that containing the empty cup. The main effect for Genotype [F(1,32)=
0.003, p=0.959] and the Genotype x Stimulus interaction [F(1,32)=3.376, p=0.076] were
not statistically significant. Bonferroni post-hoc comparisons revealed no significant
difference between time in the social and non-social chambers for the wild-type mice
[t=2.051, p>0.05], but a significant increase in time spent in the social chamber for
hemizygous mice [t=4.649, p<0.001] (Figure 2a). Figures 2b&c present the frequency
and duration of behaviors scored during the sociability phase of the three-chamber task.
Significant main effects for Genotype for frequency and duration of contact [frequency:
F(1,16)=6.52, p=0.021; duration: F(1,16)=6.768, p=0.019] were found. Bonferroni post-hoc
tests indicated a significant genotype difference in the duration of contact with the empty
cup (t=2.626, p<0.05); mutant males displayed a reduced duration. There was also a
significant Stimulus main effect for the frequency and duration of sniffing [frequency:
F(1,16)=69.32, p<0.0001; duration: F(1,16)=60.05, p<0.0001]. No other statistically
significant main differences were found.
1.4.4 Autogrooming
Hemizygous mice showed a significantly increased frequency of tail/genital
grooming [t(16)=-2.7, p=0.016] and increased duration of paw licking [t(16)=-4.074,
p=0.0009] relative to wild-type littermates (Figures 3 a&b). No statistically significant
differences in the frequency or duration of other autogrooming sub-types were found.
Similarly, no significant differences in the number of bouts, interrupted bouts, proportion

13
of interrupted bouts, transitions between grooming stages, incorrect transitions, and
proportion of incorrect transitions were found (Figure 3c).
1.4.5 Repetitive Novel Object Contact Task/Locomotion
During the habituation phase, no significant differences were noted in the number
of lines crossed [t(16)=-0.598, p=0.558, data not shown] nor in the duration of time spent
within each quadrant [F(1,16)=0.827, p=0.377, data not shown]. Wild-type and hemizygous
mice investigated unfamiliar novel objects at a similar rate [F(1,64)=0, p=1] (Figure 4a).
Similarly, when proportional preferences for each toy were ranked and averaged for
each genotype, no significant difference in object preferences was found [F(1,32)=0.803,
p=0.377] (Figure 4b). Finally, when the number of identical three- and four-object
sequences was compared between genotypes, no differences were observed [F(1,64)=4E-
12, p=1] (Figure 4c). Taken together, MeCP2 mutants and wild-type mice are
indistinguishable in cognitive, object- investigation-based measures of stereotypy or
restricted interest.
1.4.6 Social Proximity
Figures 5 a&b display the frequency and duration of social behaviors of MeCP2
wild-type and mutant mice to unfamiliar B6 mice. Mann-Whitney U-tests indicated a
significant increase in the frequency of Crawl Under behavior in hemizygous mutants
[p=0.006]. They also showed significant elevations in the duration of Crawl-Over
[p=0.026] and Crawl-Under [p=0.010] behavior relative to wild-type littermates. Finally,
MeCP2 hemizygous males showed an increased duration of Nose-to-Face behavior
[p=0.040]. No other statistically-significant effects were noted in the social proximity
chamber between hemizygous and wild-type MeCP2 mice.
1.4.7 Urinary Scent Marking
Figure 6 presents the average number of 1 x 1cm squares containing scent
marks in the baseline and social scent marking conditions. Two-way repeated measures
ANOVA revealed a significant main effect of Genotype [F(1,32)=6.15, p=0.0195]. Neither
the main effect for scent marking Condition [F(1,32)=0.024, p=0.878] nor the Genotype x
Condition interaction were statistically significant [F(1,32)=0.280, p=0.601]. No significant
post-hoc differences were noted between Genotypes in either Condition. Therefore,
mutant MeCP2 mice show elevated scent marking across social and non-social
contexts.
1.4.8 Resident Intruder

14
No attack behavior was noted in any of the MeCP2 wild-type or mutant mice;
therefore, no attack variables are reported. Resident male MeCP2 hemizygous mutants
demonstrated lower average durations of following behavior than wild-type littermates
[t(15)=2.273, p=0.038]. No other significant differences were noted (Figure 7a,b).
1.5 DISCUSSION
Mutations of the MECP2 gene in humans and laboratory animals are associated
with a variety of neurological and behavioral alterations. However, the specific role of
this gene and the MeCP2 protein in normal variation in behavior and in disease states
remains to be elucidated. A major goal of this set of studies was to unravel the specific
types of abnormalities in social behaviors and stereotypies in male mice with targeted
mutations in Mecp2. Since existing reports are inconsistent in the directional influence of
mutations of this gene on social behavior, we performed a battery of tasks designed to
reveal subtle differences in social motivation under diverse social contexts. In the VBS, a
laboratory-based semi-natural environment in which subjects are free to establish their
own time budgets, MeCP2308/Y mutant males preferentially approach one another from
the front rather than the back, a preference which appears to be associated with pro-
social motivation (Arakawa et al, 2007). Mutant MeCP2308/Y mice also spent a greater
amount of time huddling during the dark phase, which indicates that they prefer contact
and spend less time alone compared to wild-type littermates. In the three-chamber
apparatus, whereas both genotypes spent more time in the side containing an unfamiliar
stimulus mouse, this effect was greater for the mutant mice. These results resemble
those noted in Mecp21lox and Mecp2flox mutants (Fyffe et al, 2008; Samaco et al, 2008,
2009; Schaevitz et al, 2010) albeit with some possible differences in interpretation in
light of other results in these studies (see below). Kerr et al, (2008) noted enhanced
social novelty preference for Mecp2lox/y mutants. However, a limitation of the Kerr et al,
(2008) study is that it did not counterbalance the social novelty phase, possibly
confounding social and object novelty (Pearson et al, 2010).
Mutant MeCP2308/Y mice have been reported to be hypoactive in the dark phase
and hyperactive in the light (De Filippis et al, 2010; Moretti et al, 2005). However, in the
present study, huddling behavior in mutants was comparable to that of wild-type mice in
the light phase, when mutants tend to be hyperactive, while allogrooming was elevated
in the dark phase, the period when mutants tend to be hypoactive. Moreover, we found

15
no genotype differences in locomotor activity in the habituation phase of the repetitive
novel object contact task which was assessed in the light phase. Similarly, an overall
decrease in contact with the stimulus cups in the three-chamber task was noted in
mutants. It is currently unclear why this might have occurred. However, confounding
distinctions in activity patterns seem an insufficient explanation, because mutants tend to
be hyperactive in the period when tested. Thus, the affiliative inclination of MeCP2
mutants does not appear to reflect hypoactivity in the dark phase and hyperactivity in the
light.
The social proximity task was designed to differentiate close-quarter differences
in social interactions (Defensor et al, 2011). In the current study, MeCP2308/Y mice
showed elevated frequencies and/or durations of crawl-under, crawl-over behaviors, and
nose-to-face investigation. Notably, low-social BTBR T+tf/J (BTBR) mice also displayed
increases in crawl-over and -under behaviors, which were interpreted as attempts to
avoid the other pair-member in a situation in which physical distance from that animal
could not be achieved; a view that was supported by findings that diazepam significantly
reduced this behavior in BTBRs (Defensor et al, 2011). In the present, highly social,
MeCP2308/Y mutants, these crawl-under/over results are consonant with their enhanced
social approach, investigation, and huddling in the VBS. An intriguing possibility is that
enhanced crawl under/over may nonetheless reflect a subtle social deficiency since
control mice (C57BL/6J) in the Defensor et al, (2011) study tended to orient to the pair-
partner mouse and move around it, rather than crawling under or over it, as might be
more appropriate for an inanimate object.
These data, including VBS indices, the three-chamber task, social proximity
findings, and urinary scent marking suggest that this specific partial loss of function
mutation of the MeCP2 gene in male mice produces an up-regulation of affiliative social
behavior. An additional goal of this set of studies was to understand further the
motivational components of these enhanced behaviors. Many commonly utilized social
behavior tasks (e.g. three chamber) do not inherently discriminate aggressive
motivations in approach and proximity variables. This is a particular issue for MeCP2
mutants because brain region and cell-specific MeCP2 mutants (Fyffe et al, 2008;
Samaco et al, 2009) were more aggressive in a resident-intruder situation, and also
spent more time in social proximity in the partition test, suggesting an aggressive
motivation in such approaches to the stimulus mouse. The present findings that the 308

16
mutant mice show higher levels of allogrooming are consonant with this interpretation, in
that intense bouts of allogrooming are often interpreted as agonistic behaviors reflective
of dominance motivations (“rough grooming”; Litvin et al, 2007; Long et al, 1972).
Similarly, increased scent marking for MeCP2308/Y in this study suggests an interpretation
in terms of enhanced aggressive or dominance motivations. This interpretation is not
supported by findings that MeCP2308/Y mice failed to show attack in the resident-intruder
task. However, overt attack may not be the most sensitive measure of dominance or
competitive motives, and an overall evaluation of the present data suggests that the
contribution of such motivation to social approach for the 308 mutant cannot currently be
dismissed.
Much of the early work with MeCP2308/Y mutant mice indicated that wild-type mice
seem to avoid contact with the mutant mice in the tube test and resident intruder task
(Shahbazian et al, 2002). An additional aspect of social interaction, relevant to
interpretation in terms of autism-like behaviors, is reciprocity. One possibility is that it is
the lack of reciprocation on the part of the recipient (stimulus) mouse that drives this
enhanced sociability in the mutants: Another is that the MeCP2308/Y mutants are deficient
in some behavioral, or perhaps physiological, characteristics that elicit sociality in WT
mice. In this context, the above-mentioned enhanced crawl under/over behaviors may
suggest such a social deficiency. These possibilities have yet to be tested, but suggest
some additional and potentially important aspects to the complex set of events that
constitute sociality in mammals. However, the present study revealed no differences in
the frequency of directed investigation towards mutant vs wild-type residents; and a
slight augmentation of duration of olfactory investigation including a significant increase
in duration of anogenital sniffing of mutants by intruder wild-types (data not shown) in the
resident intruder paradigm. The original 308/Y mutants in the Shahbazian et al, (2002)
study were on a mixed background, while those in the current study were on a C57BL/6J
background which may, in part, account of this difference.
Mouse models of ASD commonly display patterns of restricted repetitive
behaviors (Moy et al, 2008; Pearson et al, 2011). Published studies have characterized
hindlimb clasping when lifted by the tail, and forelimb stereotypies in MeCP2 mutants,
and it is thought that these might represent face validity in models of Rett syndrome
(Chen et al, 2001; Gemelli et al, 2006; Guy et al, 2001; Moretti et al, 2005; Shahbazian
et al, 2002). We found no disturbances in patterns of object investigation, and only

17
minor alterations in forepaw- and anogenital-directed grooming behaviors. The latter
results may extend previous discoveries of altered stereotypy in MeCP2 mutants by
noting that qualitative and region-specific differences in grooming may exist, but the
cephalo-caudal patterning appears to be normal.
Taken together, the hypersocial phenotype of MeCP2308/Y mutants demonstrates
one of the many conceptual issues arising from models of pervasive developmental
disorders- that autism-relevant features may occur without actual reductions in affiliative
social behavior. Mouse models of symptom- or domain-specific alterations in
neurodevelopmental disorders may be as informative as those that are generated in an
attempt to create an entire syndrome. To that end, a future goal might be to clarify the
distinct effects of the variety of region and allele targeted mutants of MeCP2 on specific
forms of social behavior, and the downstream targets of MeCP2 protein that influence
behavior, particularly throughout the developmental span. These results contribute to a
growing body of research that suggests MeCP2 is critical in the bi-directional fine-tuning
of behavioral responses, possibly up to, and including complex social interactions.

Figure 1. MeCP2308/Y Mutant Mice Show Increased Affiliation in the Visible Burrow System. Figures 1a and b display the frequencies of each of the eight behaviors scored during 24 scans across the first four hours of the dark (upper panel) and light phases (lower panel). Horizontal lines indicate a significant main effect of Day, and vertical lines indicate a significant Genotype main effect. y/+ denotes wild-type and y/- are hemizygous mutant mice. * p<0.05; ** p<0.01, *** p<0.0001. Symbols above individual means indicate a significant post-hoc effect: # p<0.05; $ p<0.001. Wild-type (y/+) N=9, mutant (y/-) N=6.
18
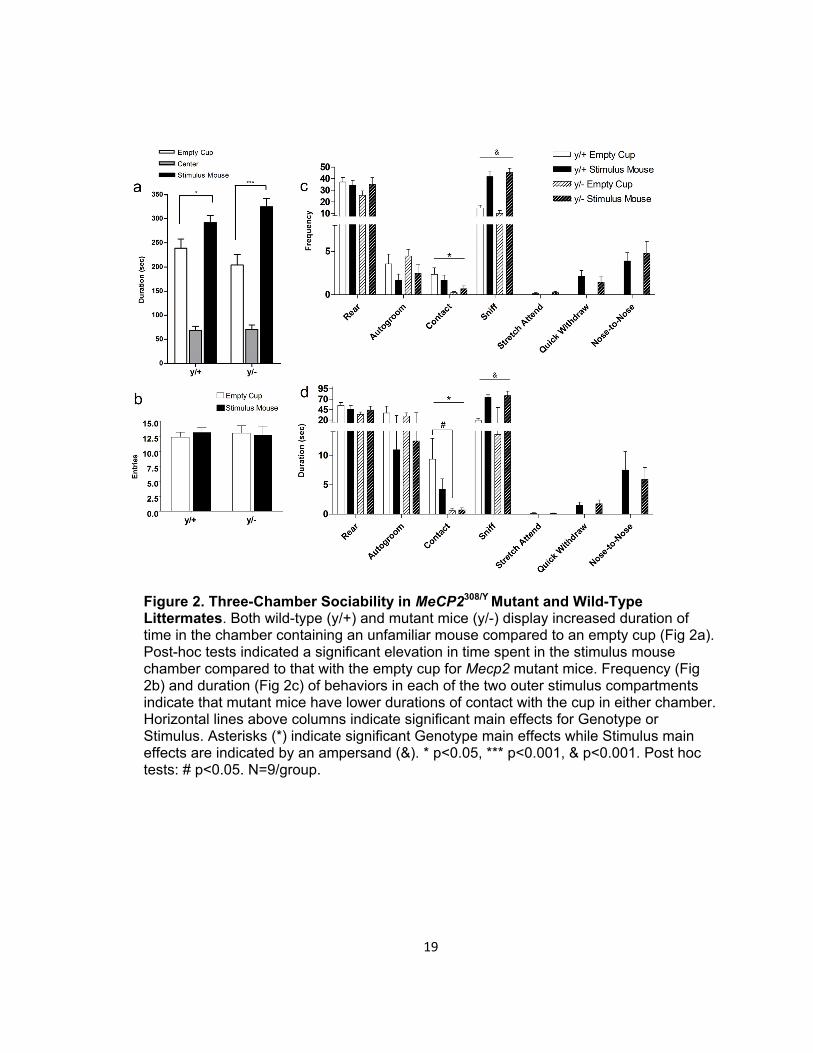
Figure 2. Three-Chamber Sociability in MeCP2308/Y Mutant and Wild-Type Littermates. Both wild-type (y/+) and mutant mice (y/-) display increased duration of time in the chamber containing an unfamiliar mouse compared to an empty cup (Fig 2a). Post-hoc tests indicated a significant elevation in time spent in the stimulus mouse chamber compared to that with the empty cup for Mecp2 mutant mice. Frequency (Fig 2b) and duration (Fig 2c) of behaviors in each of the two outer stimulus compartments indicate that mutant mice have lower durations of contact with the cup in either chamber. Horizontal lines above columns indicate significant main effects for Genotype or Stimulus. Asterisks (*) indicate significant Genotype main effects while Stimulus main effects are indicated by an ampersand (&). * p<0.05, *** p<0.001, & p<0.001. Post hoc tests: # p<0.05. N=9/group.
19

Figure 3. Subtle Alterations in Patterns of Autogrooming in MeCP2308/Y Mutant Mice. Frequency and duration of body site-specific patterns of grooming (Fig 3a,b). Mecp2 mutant mice (y/-) show increased frequency of tail/genital grooming and an increased duration of paw licking compared to wild-type littermate controls (y/+). No significant differences were found in syntactical parameters of grooming (Fig 3c). *p<0.05; **p<0.01. N=9/group.
20
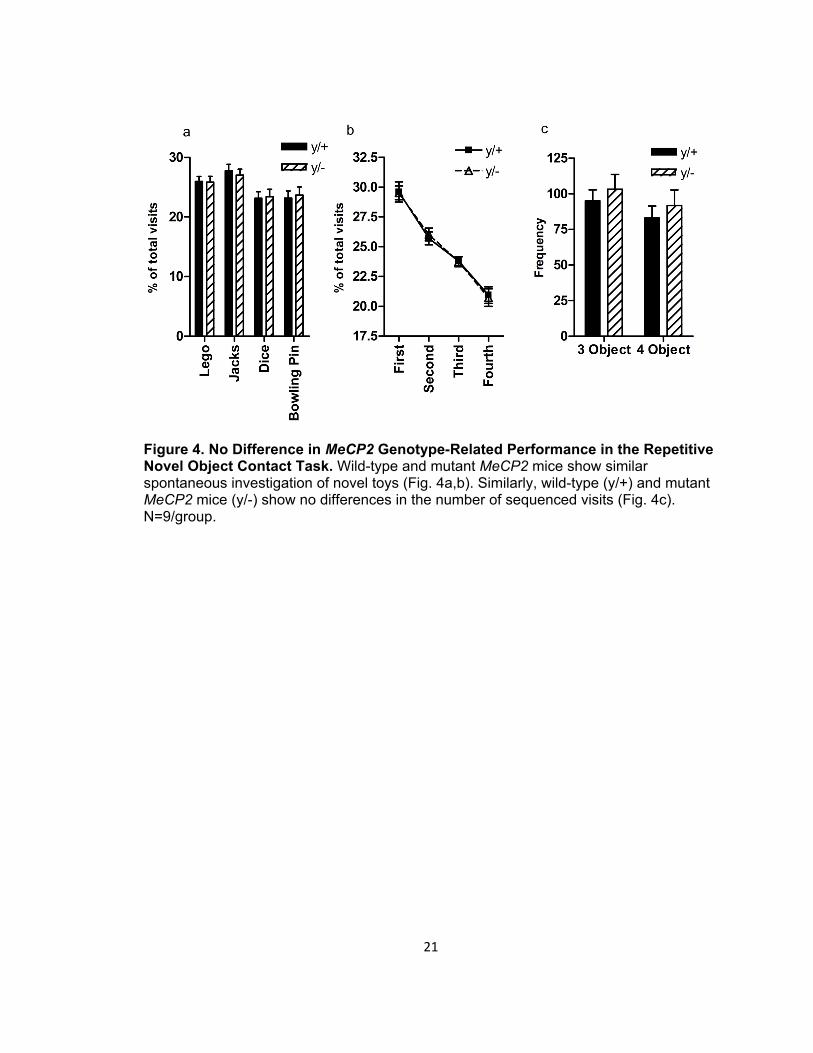
Figure 4. No Difference in MeCP2 Genotype-Related Performance in the Repetitive Novel Object Contact Task. Wild-type and mutant MeCP2 mice show similar spontaneous investigation of novel toys (Fig. 4a,b). Similarly, wild-type (y/+) and mutant MeCP2 mice (y/-) show no differences in the number of sequenced visits (Fig. 4c). N=9/group.
21

Figure 5. Patterns of Social Interaction in the Social Proximity Task. MeCP2 mutant mice (y/-) show increased frequency of crawl under (Fig 5a). Additionally, they show higher duration of nose-to-face interactions, as well as increased durations of crawl over and crawl under behavior (Fig 5b) compared to wild-type littermates (y/+). *p<0.05; **p<0.01. N=9/group.
22

Figure 6. Elevated Scent Marking in MeCP2308/Y Mutant Mice. Mutant mice (y/-) scent mark more to an empty chamber and when an unfamiliar adult CD-1 mouse is behind a divider compared to wild-type littermates (y/+). Wild-type (y/+) N=9, mutant (y/-) N=8. Genotype main effect * p<0.05.
23

Figure 7. Comparison of Agonistic Behavior of MeCP2308/Y Mutant and Wild-Type Mice in the Resident-Intruder Task. Mecp2 mutant mice (y/+) show similar frequencies of behavior to an unfamiliar intruder compared to their wild-type littermates (y/+)(Fig. 7a). Mutant mice show decrease duration of following behavior (Fig. 7b). *p<0.05. Wild-type (y/+) N=9, mutant (y/-) N=8.
24

25
CHAPTER 2: EMOTIONAL CHARACTERISTICS OF MECP2308/Y MUTANTS
2.1 INTRODUCTION
The protein methyl-CpG binding protein 2 (MeCP2) is transcribed from the X-
linked gene MECP2. MeCP2 is intricately involved in the regulation of gene expression
via its actions on methylated DNA and microRNA as well as influences on alternative
splicing (Chahrour et al, 2008; Jørgensen & Bird 2002; Lewis et al, 1992; Samaco et al,
2009; Wu et al, 2010; Young et al, 2005). Not only does MECP2 mutation underlie most
cases of Rett syndrome (Amir et al. 1999; Bienvenu et al. 2000), it has been implicated
in non-syndromal autism (Beyer et al, 2002; Carney et al, 2003; Nagarajan et al, 2006)
and other psychiatric disorders (Hammer et al, 2002; Van Esch et al, 2005). Emerging
literature also supports a central role of MECP2 in addiction processes because, in
rodents, psychostimulant drugs induce MeCP2 expression (Cassel et al, 2006), while
targeted mutation of MeCP2 alters amphetamine and cocaine sensitivity (Deng et al,
2010; Im et al, 2010). This has led to the hypothesis that fine tuning of MeCP2 protein
level in the brain is critical in normal brain functioning (Kaufmann et al, 2005) early in
brain development as well as in the adult (McGraw et al, 2011).
Shahbazian et al, (2002) created a transgenic mouse (MeCP2308/Y) that is
missing approximately one-third of the C-terminal region of the MeCP2 gene. This
partial-loss of function mutation is highly clinically relevant (e.g., Meloni et al, 2000; Wan
et al, 1999) and furthermore, permits the assessment of male mutants and mice that
reach adulthood prior to debilitating epilepsy and musculoskeletal abnormalities.
Constitutive MeCP2 knockout, as well as truncated and cell and brain-region specific
knockout mice show pronounced alterations in anxiety-related, cognitive and social
behaviors (see Calfa et al, 2011 for an exhaustive review). Interestingly, work from our
lab (Chapter 1 and Pearson et al, 2012) has noted a hypersocial phenotype in
MeCP2308/Y mice. More specifically, MeCP2308/Y mutants displayed an augmentation of
social approach and investigation with an absence of aggression. These results
complement findings from Schaevitz and colleagues (2010) noting increased three-
chamber sociability in Mecp21lox null mutants.
A variety of phenomena may contribute to the social characteristics of a given
mouse strain or engineered mutant. Cognitive or sensorimotor disturbances can
influence behaviors of interest and alterations in emotional reactivity influence the

26
behaviors expressed in a given paradigm (Crawley 2007a). It is entirely conceivable that
much of the social phenotype of a mouse (such as that of the MeCP2308/Y mutant mouse)
may reflect changes in emotional, defensive or cognitive disruptions. Previous reports
have indicated that MeCP2308/Y mice show several cognitive deficits such as spatial and
contextual fear memory (Moretti et al, 2006). Despite motor dexterity abnormalities
(Shahbazian et al, 2002; Pelka et al, 2006), sensory systems appear intact in MeCP2
mutants (Ricceri et al, 2008). Increases in anxiety and stress responsivity have been
found in MeCP2308/Y mice (McGill et al, 2006; Shabazian et al, 2002).
To replicate and extend previous reports, male MeCP2308/Y mutants and their
wild-type littermates were tested in the elevated plus-maze and the elevated zero-maze.
To determine whether the mutants show altered reactions to imminent threats, (fear) the
same animals were tested in the mouse defense test battery (MDTB) which is a series of
tests under which a threatening, predatory stimulus is presented under varying threat
distance and availability of escape. These tests were performed to assess potential
emotional phenotype characteristics of the MeCP2308/Y truncated mutant. We predicted
that the hyper-social phenotype of MeCP2 mutants would be accompanied by overt
decreases in anxiety and fear.
2.2 MATERIALS AND METHODS
2.2.1 Experimental Subjects
Experimental animals were bred from C57BL/6J-backcrossed stock obtained
from Jackson Laboratory (B6.129S-Mecp2tm1Hzo/J, stock # 005439) and bred from
heterozygous mutant dams and hemizygous father sibling pairs. Subject mouse
genotype was determined according to the PCR parameters from The Jackson
Laboratory with purified DNA collected from tail biopsy after weaning at post-natal day
25. Since MeCP2 is an X-linked gene, only wild-type (y/+: WT) and hemizygous (y/-: KO)
males were obtained and compared in behavioral testing. Stimulus mice of the CD-1
outbred strain were acquired from Charles River and bred in-house. Mice were housed
with up to five same-sex littermates under a 12-h light/dark schedule with lights on at
0600h. Mice had ad libitum access to tap water and lab rodent diet. All procedures were
performed according to protocols approved by the University of Hawaii Laboratory
Animal Service (LAS) Institutional Animal Care and Use Committee.
2.2.2 Experimental Design

27
Seventeen each of WT and MeCP2 KO mice were assigned to two experimental
cohorts. Eight of each were assessed in the elevated plus-maze, the female urine
sniffing test, and the cocaine-induced behavioral sensitization test. A second cohort of
nine of each genotype were tested in the elevated zero-maze and the mouse defense
test battery.
2.2.3 Elevated Plus-Maze
The test apparatus is based on that described by Lister (1987) and comprised of
two open arms (30 × 7 × 2 cm) and two closed arms (30 × 7 × 20) that extend from a
common central platform (5 × 5 cm). The apparatus was constructed from wood and
Plexiglas and was raised to a height of 40 cm above floor level. To prevent mice falling
off, a rim of Plexiglas (0.25 cm high) surrounded the perimeter of the open arms. One
ceiling-mounted video camera was used to record mouse behavior during the test and
the experimental room was illuminated with standard fluorescent lamps. The mean
intensity of luminosity on the open arms of the EPM was 55 lux. Testing was initiated by
placing the subject on the central platform of the maze, facing one of the open arms.
Test sessions lasted five minutes and, between subjects, the maze was thoroughly
cleaned with 70% ethanol and dry paper towels. The results were expressed as
frequency of entries into the closed and open arms, as well as percentage of time spent
in open arms with respect to total time in either closed or open arms. Ethological
measures comprised frequency scores for stretched attend postures (mouse stretches
forward and maintains a “flat-back” posture), head dips (mouse protrudes its head over
the edge of an open arm to a length equal to or exceeding the caudal aspect of the
pinnae).
2.2.4 Elevated Zero-Maze
The test apparatus consisted of an acrylic circular platform (6 cm in width)
composed of open and closed segments, with the latter being surrounded by 20 cm high
acrylic walls. The platform was raised to a height of 50 cm above floor level and the
diameter of the maze was 65 cm. To prevent mice falling off, a rim of Plexiglas (0.25 cm
high) surrounded the perimeter of the open segments. One ceiling-mounted video
camera was used to record the behavior during the test and the experimental room was
illuminated with standard fluorescent lamps. The mean intensity of luminosity on the
open segments of the zero-maze was 70 lux. Testing was initiated by placing the subject
in the center of the open arm. Test sessions lasted five minutes. The results were

28
expressed as mean entries into the closed and open sections as well as mean
percentage of time spent in open segments over total time spent in both open and
closed segments. Ethological measures comprised frequency scores for stretched
attend postures (mouse stretches forward and maintains a “flat-back” posture), head
dips (mouse protrudes its head over the edge of an open arm to a length greater than or
exceeding the caudal aspect of the pinnae). Videotapes collected for the elevated plus
and elevated zero mazes were scored by an observer blind to genotype using
commercially-available annotation software (Observer, Noldus Information Technology,
The Netherlands)
2.2.5 Mouse Defense Test Battery
Ten minutes before the test session began, an adult Sprague–Dawley rat
(average weight of 450 g) to be used as the predator stimulus was deeply anesthetized
with sodium pentobarbital (80 mg/kg, i.p.) in order to minimize its discomfort. The Mouse
Defense Test Battery (MDTB) consists of the following subtests:
1) Pre-test: subjects were placed into the MDTB apparatus for a 3-min
familiarization period during which total line crossings and wall rears were
recorded. Predator avoidance test: avoidance and escape distances were
measured when a predator stimulus (a hand-held rat) was brought up to the
subject at a speed of approximately 0.5 meters per second (m/s). Approach
was terminated when contact with the subject was made or if the subject ran
away from the approaching rat. This was repeated five times.
2) Chase/flight test: the anesthetized rat was brought up to the subject at a
speed of approximately 2.0 m/sec. Chase was initiated only when the subject
was standstill with its head oriented toward the rat, and completed when the
subject had traveled a distance of 14.4m (three laps in the runway). The time
spent by the mouse to travel this distance was recorded. Mean flight speed
(in m/s) was calculated based on the obtained latency to complete the three
laps. In addition, the number of stops (pauses in locomotion), orientations
(subject orients its head toward the oncoming rat) and reversals (subject
turned and ran in the rat direction) were also recorded.
3) Straight alley test: the runway was converted into a straight alley, 80 cm long,
by the closing of a door at one end and the placement of a removable barrier
at the other. The rat is placed at one end while the mouse begins the test at

29
the other. During a 30 sec period, the number of approach-withdrawals
(subject moves toward the rat more than 20 cm and then returns), as well as
voluntary contacts with the rat stimulus were recorded. Other measures
scored included immobility time (freezing) and the frequency of defensive
uprights.
4) Forced contact test: in this situation, the straight alley length was reduced to
40 cm. The rat was brought up by the experimenter in five sudden contacts
directed toward the subject. For each such contact, the number of
vocalizations, defensive uprights, jump attacks, jump escapes and bites were
recorded. This procedure was repeated three times.
5) Post-test: upon completion of the forced contact test, the alley doors were
opened allowing the subject free locomotion around the oval runway. The
subject exploratory activity was recorded for another 3 min period during
which the experimenter and the rat stimulus were out of sight. Line crossings
and wall rears were recorded.
2.2.6 Female Urine Sniffing Test
Reward sensitivity in MeCP2 mutants might underlie their behavioral phenotype.
To test this possibility, MeCP2 wild-type and knock out (KO) mice, previously analyzed
for social behavior and stereotypy, were tested in a female urine sniffing test of hedonia
according to procedures outlined in Malkesman et al, (2010). Fresh urine was collected
from adult female CD-1 mice and used as a point-source stimulus. Individually-housed
MeCP2 wild-type and mutant mice were initially exposed to a sterile foam-tipped surgical
swab containing 20 µL distilled water for a five minute session. Immediately following
this baseline session, they were permitted access to a new swab containing 20 μL fresh
female urine for an additional five minutes. Urine was collected for females primed to
soiled male mouse bedding for four consecutive days to ensure estrous and/or
proestrous status (Dalal et al, 2001), and pooled such that all samples were uniform.
The average duration of time sniffing the swab, which was defined as the subject
suspending all other activities while maintaining proximity of the nose to the tip of the
swab at a distance ≤ 2 cm, was analyzed for both sessions and compared by genotype.
2.3 RESULTS
2.3.1 Elevated Plus-Maze

30
No significant differences were noted in the frequency of entries into the areas of
the plus-maze arena (Figure 8a). Wild-type and knockout mice showed similar rates of
stretch-attend and head dip postures during the five minute test (Figure 8b). Finally,
there were no significant differences in the duration of time spent in the open, center and
closed arms of the elevated plus-maze as well in the proportion of time spent in the open
arm relative to the other arms (Figure 8c).
2.3.2 Elevated Zero-Maze
MeCP2 wild-type and mutant mice demonstrated similar frequencies of entries
into the open and closed portions of the elevated zero-maze (Figure 9a). Though the two
genotypes displayed similar rates of Stretch Attend postures, mutant mice showed a
significant elevation in the frequency of Head Dips (t(16)=-3.55, p=0.003, Figure 9b).
When the mean duration of time spent in the open and closed compartments was
compared for the wild-type and mutant mice, the results demonstrated no significant
differences. Furthermore, the proportion of time spent in the open arms was comparable
for the two genotypes (Figure 9c).
2.3.3 Mouse Defense Test Battery
Table 3 displays the mean and standard error for the MDTB parameters, and
independent t-test and Mann-Whitney U-test values. In brief, MeCP2 mutants showed an
enhanced avoidance distance of the predatory stimulus in the predator avoidance test.
Mutants showed enhanced flight speed, fewer stops, and more reversals in the
chase/flight component as well as fewer approaches and contacts; they also showed
increased freezing in the straight alley test. When the mice were exposed to forced
contact with the rat stimulus, mutants performed jump escape behaviors more than wild-
type mice. When mice were exposed to the post-test, when given free access to the
entire oval arena with an absence of the predatory stimulus, mutants performed more
rearing (exploratory) behaviors.
2.3.4 Female Urine Sniffing Test
Two-way ANOVA indicated no significant main effect of Genotype
(F(1,14)=0.004, p=0.952) or of the Stimulus (F(1,14)=1.522, p=0.238). The Genotype x
Stimulus interaction also failed to reach statistical significance (F(1,14)=0.291, p=0.598).
2.4 DISCUSSION

31
The purpose of these studies was to determine whether alterations in
fundamental affective and approach/withdrawal systems might underlie the unusual
social phenotype of MeCP2 mutant mice. More specifically, the prediction was tested
that enhanced affiliative sociability might be secondary to a reduction in anxiety or fear-
which would render novel stimuli or contexts less threatening. The overall hypothesis
that an augmentation of social affiliation might be the consequence of a general
enhancement of sensitivity to reinforcers was also tested. We noted normal anxiety in
the elevated plus- and zero-mazes but a complex enhancement of fear and defensive
responses in the mouse defense test battery. The latter finding was the opposite of the
predicted changes. Taken together these results suggest that the hyper-social
phenotype of MeCP2308/Y mutant mice cannot be attributed to a general effect of the
protein’s dysfunction on failures of the emotional systems showing appropriate risk
assessment and defensive responses to novel, potentially threatening social systems or
contexts. Likewise, the social phenotype cannot be understood solely in terms of overall
enhanced sensitivity to rewarding olfactory stimuli. Further testing will be necessary to
assess which sensory or emotional systems drive MeCP2308/Y hyper-sociability.
Previous studies of MeCP2 mutants have demonstrated increased anxiety
(Gemelli et al, 2006); however, others have demonstrated normal and decreased
anxiety, depending on the type of mutation (Guy et al, 2001; Pelka et al, 2006). Indeed,
our reports are contrary to others demonstrating elevated anxiety in the MeCP2308/Y
mutant (McGill et al, 2006; Shahbazian et al, 2002). A potential explanation of this
contradiction is that earlier reports utilize mutants on a hybrid background, while the
current study used mice fully backcrossed onto a C57BL/6J background (The Jackson
Laboratory Strain # 005439). Another consideration is that previous reports may
conclude that MeCP2 mutants possess an anxiety phenotype but this may be based on
open field test parameters, for instance, in the absence of light-dark box impairments
(e.g. Fyffe et al, 2008) or investigations of the plus or zero mazes. This conclusion would
be problematic given locomotor disturbances in many MeCP2 mutants. Altogether, the
influence of MeCP2 on anxiety may be complex, but our work does not support an
anxious phenotype in adult MeCP2308/Y mutants.
One limitation of our results is that the exploration parameters collected in the
elevated zero-maze might be affected by a ceiling effect. Subject mice were placed into
the open zone facing a closed portion and many of both genotypes inhibited exploration.

32
Therefore, proportion open area time approached one hundred percent; this could have,
to some degree masked the ability to demonstrate a difference. On the other hand, both
wild-type and mutant mice showed this lack of exploration around the perimeter of the
zero-maze and thus spent very little time in the protected areas. Although protocols for
elevated zero-maze often indicate placement of the subject into the closed area first,
others do not (e.g. Bader et al, 2011). In the future, researchers should consider placing
MeCP2 mutants and controls into the closed zone when using the elevated zero-maze to
reveal any abnormal anxiety-like behaviors in MeCP2 mutants. Alternatively, the light-
dark box may be a more appropriate measure of anxiety-like behavior in future analyses.
However, given the absence of anxiety-like behavior in the elevated plus maze, coupled
with the normal exploration noted in the mouse defense test battery prior to the
predatory threat stimulus presentation, it does not appear the MeCP2308/Y mice (on a
pure C57BL/6J background) display robust alterations in anxiety.
To explore the possibility that this mutation causes other affective disturbances,
MeCP2308/Y mice were tested for fear and panic-like parameters in the MDTB. Altogether,
mutant mice were substantially more defensive, as evidenced by augmented avoidance
and escape behaviors, as well as increases in active defensive responses to the
threatening predatory stimulus. These data suggest that not only are fear and defense
intact in the hypomorphs, it surpasses that of their normal littermates. Although fear
conditioning has been assessed in MeCP2308/Y mice, it has been shown to be normal, in
a context conditioning paradigm (but not in a cue-conditioning test) to footshock (Gemelli
et al, 2006; Shahbazian et al, 2002). Since fear conditioning and unconditioned fear
possess unique properties (Rosen et al, 2004), further research should analyze the
implications of altered unconditioned fear in MeCP2 mutants.

Figure 8. Elevated Plus-Maze (EPM) testing of MeCP2308/Y and Wild-Type Littermates. MeCP2 mutant (Y/-) and wild-type (Y/+) mice show similar frequencies of entries into the three areas of the EPM (a). They also display comparable durations and proportions of time in the open and closed arms (c). Rates of risk assessment and exploration behaviors were similar as well (b). N=8/group.
33
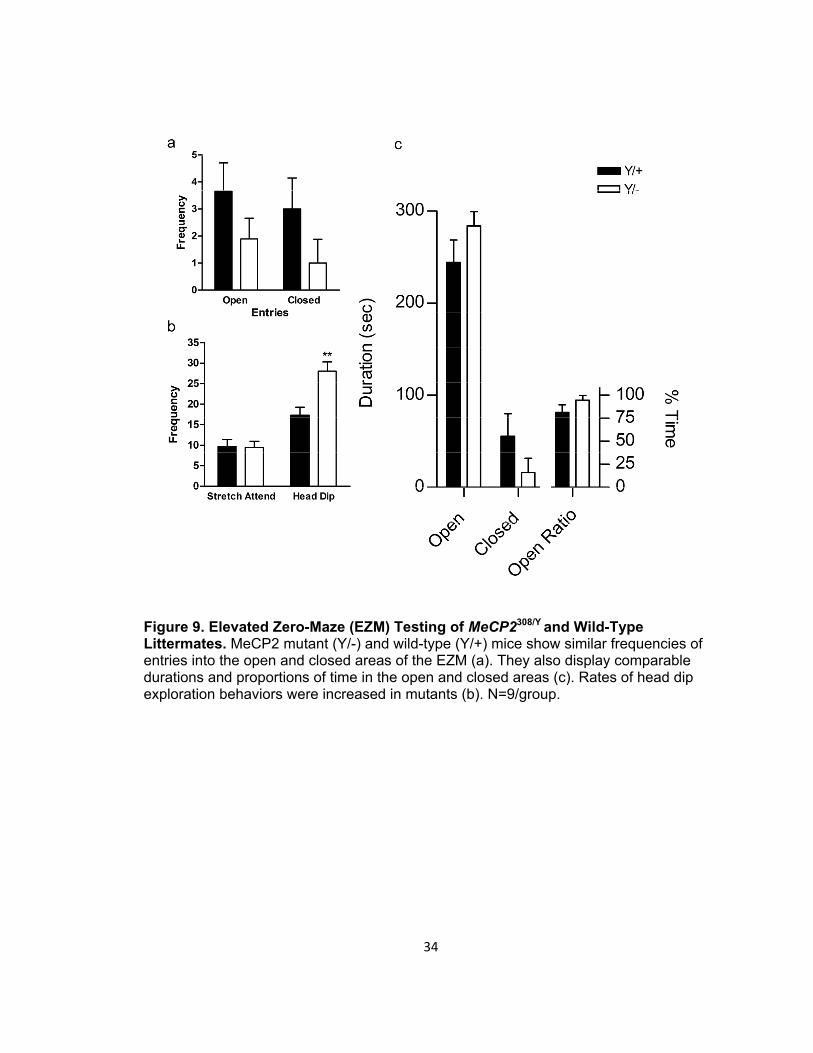
Figure 9. Elevated Zero-Maze (EZM) Testing of MeCP2308/Y and Wild-Type Littermates. MeCP2 mutant (Y/-) and wild-type (Y/+) mice show similar frequencies of entries into the open and closed areas of the EZM (a). They also display comparable durations and proportions of time in the open and closed areas (c). Rates of head dip exploration behaviors were increased in mutants (b). N=9/group.
34

35
CHAPTER 3: REWARD SYSTEMS IN MOUSE MODELS OF AUTISM
Basic models of behavioral regulation (see Lang et al, 1998) tend to posit that
emotional systems regulate learning, experience and behavioral output which are, in turn
driven by internal motivational states. This simple model may be useful in attempts to
characterize central processes regulating psychiatric-disorder-relevant behavioral
displays in rodents. One potentially understudied system in murine models of autism is
motivation (Crawley 2007b). Since social motivation deficits have been implicated in
autism (Dawson 1998; Dawson et al, 2005; Waterhouse et al, 1996), pre-clinical
researchers should consider distinctions in sensitivity to rewarding stimuli in mice
showing variability in social approach and affiliation.
Much research in behavioral neuroscience has been concerned with the classical
and instrumental underpinnings of approach, avoidance and consummatory behaviors.
As such, many explanations of a given behavior tend to focus on reinforcement and
contingency. Although social interactions are a motivated behavior (Ikemoto & Panksepp
1999), very few murine models of autism have been assessed for aberrant brain reward
systems in conditions of disordered social behavior. In particular, neurodevelopmental
disorders with impaired sociability may result from an overall lack of motivation to
engage unfamiliar persons. This notion is supported by the fact that many individuals
with ASD show limited affiliation, communication and bonding with family members but
show a virtual absense of appropriate affiliation with others (Kanner 1943). A simple
approach to comprehend the mechanisms of these traits is to measure the hedonic
value of social reinforcers within rodent models. Any disturbances in brain reward
circuitry might contribute to, or reflect impairments in appropriate assignment of value to
exogenous social signals and potentially reveal a biomarker or target for therapeutics.
An aim complementary to analyses of emotional parameters is general assessment of
motivation and reward sensitivity in animal models of autism. Therefore the prediction
was made that since MeCP2308/Y mutants are hypersocial, they would show increased
sensitivity to a chemical reinforcer, and this altered corollary might underlie components
of their social phenotype.
To begin realizing the role of reward and hedonia in sociability, low-social BTBR
T+tf/J (BTBR) mice and normally-social C57Bl/6J (B6) mice were tested in a novel social
conditioned place preference task. More specifically, to verify that social interactions are

36
reinforcing in mice, a normal and socially-impaired strain were studied for positive
incentive evaluation-type behavior, to social interactions. Mice were tested in a hybrid
apparatus that was prepared by adding uniform black and white stripes to the outer two
compartments of two three chamber arenas- in doing so, allowing the distinct vertical
and horizontal stripes that are unique to the outer compartments to act as conditioned
stimuli in an a Pavolivan task. Rather than using a chemical reinforcer, one of the
unique contexts was paired with an unfamiliar conspecific during repeated days of
conditioning to assess the positive incentive value of the interaction. As outlined below-
while B6 mice showed a place preference for the side containing the CS associated with
social interactions, BTBR mice did not. This suggests that BTBR mice do not find the
interactions reinforcing, as predicted by the social motivation hypothesis.
3.1 INTRODUCTION
Autism spectrum disorder (ASD) is a highly heritable, but varied set of
neurodevelopmental abnormalities characterized by deficits in reciprocal social
interactions, communication, and restricted or repetitive behaviors (American
Psychological Association [APA], 2000). Increasingly, researchers are applying modern
molecular techniques and genetic association in order to determine the underlying
pathology of ASD (Betancur 2011; DiCicco-Bloom et al, 2006). Rodent models of ASD
have proven informative in the recapitulation of the syndrome, in revealing physiological
abnormalities and in pre-clinical drug screening (Silverman et al, 2010a).
Since ASD is conceptualized as a social disorder, a major research emphasis is
on revealing brain systems that regulate social behavior, and in particular, those that are
disturbed by environmental or genetic processes (Insel & Fernald 2004). Social behavior
encompasses a broad accumulation of constructs (emotion, and sexual and aggressive
motivation) and is modulated by a variety of stimuli and sensory modalities (olfactory,
visual and tactile); therefore, some specificity is required in the interpretation of
behavioral variables in social behavior tests (Insel & Fernald, 2004). One way to
approximate the positive incentive (pro-social or affiliative) motivation of social
interaction in rodents is through tests which measure value of social interactions via
conditioning. Relative motivation for social interaction has been tested with operant
lever-pressing for social contact (Evans et al, 1994), through conditioned place
preferences (CPP) to interactions with littermates (Panksepp et al, 2007; Panksepp &

37
Lahvis 2007) and via exposure to familiar and unfamiliar conspecifics (Calcagnetti &
Schechter 1992; Thiel et al, 2008; Trezza et al, 2009). In fact, the degree of social-
conditioned place preference in rodents corresponds directly to variability in social
motivation secondary to extent of previous social isolation (Trezza et al., 2009). In rats,
social interactions are sufficiently reinforcing in their ability to reverse cocaine CPP (Fritz
et al, 2011); however, restricting tactile contact with an unconditioned social stimulus
markedly attenuates or blocks social CPP (Kai et al, 2011). The degree of conditioned
place preference can therefore be considered an index of the positive incentive value
(Bardo & Bevins 2000) of social access and provides an outstanding measure of social
cognition in rodents.
Social motivation measures may provide complementary measures in animal
models of ASD (Crawley, 2007). In fact, attenuated conditioned place preference to
conspecifics has been demonstrated in fmr1 mutant mice, a model for Fragile X mental
retardation disorder (Pacey et al, 2011). In terms of translation, social motivation testing
may be critical in revealing brain substrates of impaired affiliative social behavior in ASD
(Dawson 2005; Waterhouse et al, 1996) and perhaps the development of such
impairments through altered assignment of reward to social stimuli (Dawson et al, 2005).
In the current study, a social reinforcement paradigm was developed to analyze
social conditioned place preference in BTBR T+tf/J (BTBR) mice. We present a
modification of the three-chamber social apparatus for the analysis of place preference
stemming from context-specific access to a novel juvenile conspecific. By applying this
social conditioning task to compare juvenile, inbred C57BL/6J (B6) mice to the socially-
impaired BTBR strain, we were able to demonstrate differential place preferences to a
context associated with social interactions.
3.2 MATERIALS AND METHODS
3.2.1 Experimental Subjects
C57BL/6J (B6) and BTBR T+tf/J (BTBR) mice were bred from stock obtained
from Jackson Laboratories (Bar Harbor, ME). Subject mice were weaned on postnatal
day 21-25 and housed with up to seven same-sex littermates. An equal number of male
and female mice from each strain were tested beginning at 25 - 30 days of age, at least
24 hours after being weaned. All mice were individually identified with numbered ear
tags.

38
3.2.2 Social Conditioned Place Preference
Three-chamber arenas (Nadler et al, 2004) were modified so that the two outer
compartments included alternating 3.8 cm black and white stripes on the walls; one
chamber contained vertical stripes while the other contained horizontal stripes (Figure
10a). The center compartment was comprised of black Plexiglas walls. Two separate
arenas were used simultaneously during testing- each of which had the opposite pattern
of stripes to eliminate confounding influences of baseline stripe orientation or side
preference. On the first day of testing (Day 0), all subjects were placed in the middle
compartment and the doors were lifted to permit full access to all three compartments.
Mice could freely explore the entire arena for twenty minutes during which time an
observer manually recorded the duration of time in each of the two striped outer
compartments with hand-held stopwatches. This session served as habituation and
permitted determination of baseline side preferences for each subject. Mice were then
randomly assigned to either 5 or 10 days of social conditioning, creating four groups: B6
5 day, B6 10 day, BTBR 5 day, and BTBR 10 day. In total, twenty mice from each strain
were used, representing an equal sex ratio, with ten each per condition. On the day
following baseline testing, and for 5 or 10 consecutive days, conditioning was performed.
Mice were randomly assigned to receive contact with a conspecific (unconditioned
stimulus) in either the horizontally- or vertically- striped (conditioned stimulus)
compartment. During conditioning, doors were left closed and the mice were placed into
the non-social chamber and permitted to explore alone for 10 minutes, then the mouse
was immediately removed and placed into the opposite compartment which contained
an unfamiliar age-matched male or female B6 mouse which had been placed in the
“social” chamber immediately prior to transferring the subject mouse. The subject mouse
and stimulus mouse were then permitted 10 additional minutes to explore or interact.
The side in which the subject mouse was initially placed was alternated daily; therefore,
the order in which they received the social and non-social sessions was
counterbalanced daily. An unfamiliar group of six naïve male and female mice (n=3/sex)
served as stimulus animals. As a result, mice conditioned for 10 days were exposed to
mice they were in contact with previously for the last four out of the ten days, while 5 day
conditioned mice were exposed to unfamiliar mice daily. Subjects of both sexes were
conditioned to stimuli of both sexes. On the three consecutive days following the last
conditioning session, mice were tested daily for place preference during a free

39
exploration session for 20 minutes, exactly as they were tested during the baseline
period.
3.2.3 Social Interactions
The degree of social conditioned place preference may be directly influenced by
the quality or quantity of interactions during conditioning. Mouse strains avoiding social
contact might show impaired social conditioning secondary to inadequate social contact
during conditioning. To account for this possibility, videotapes of conditioning sessions
were sub-sampled for the presence of social proximity. Proximity was defined as
maintaining a body distance (excluding tail) of less than or equal to 2 cm at any point in
a ten second scan collected every 60 seconds for each session per day. Therefore, the
presence/absence of proximity was sub-sampled for ten seconds at each of the nine
minutes (1-10, the first minute was excluded) for five or ten days, yielding at total of 45,
and 90 scans for the two conditions, respectively.
3.2.4 Autogrooming
BTBR mice show consistent enhancements of self-grooming rates compared to
B6 control mice (McFarlane et al, 2008). BTBR mice may avoid social contact in order to
perform high rates of such body care activities. This hypothesis is predicated on the
possibiliy that vigilance in a social context and self-grooming might be state behaviors at
odds with one another. Therefore, during baseline and the first testing session, the time
spent self-grooming was scored and calculated as a percentage time [grooming duration
(in seconds)/chamber duration x 100]; the calculated values on the first test day were
then subtracted from the baseline value to obtain a change in percent time spent
grooming in each of the three compartments during testing. I predicted that BTBR mice,
but not B6 mice would spend less proportional time grooming on the side they associate
with social isolation.
All sessions were videotaped from above with a digital camcorder connected to a
DVD recorder. Side preference testing was performed in real time by two observers;
social proximity and self-grooming were scored offline from DVDs by an assistant blind
to the strain and condition of the mouse. Testing arenas were sanitized with a 70%
ethanol solution and dried between subjects. Conditioning and testing was performed
between 0800 and 1300 hours under standard fluorescent lighting. Experiments and
husbandry were performed according to protocols approved by the University of Hawaii
Laboratory Animal Service (LAS) Animal Care and Use Committee.

40
3.2.5 Statistical Analysis
For three consecutive test days after conditioning, percent preference for the side
to which the subject mouse was socially conditioned was calculated [time in social
compartment/ (time in nonsocial compartment + time in social compartment) x 100] and
for graphical and statistical purposes, the baseline preference (calculated in the same
manner) for the social side was subtracted. Therefore, values represent the change in
percent from baseline, in the preference for the social conditioned chamber. A two-way
repeated measures analysis of variance (ANOVA) was performed with Strain and
Conditioning Days as the two factors and Test Day as the repeated measures variable.
To determine strain differences independent of testing day, a two-way ANOVA was
performed utilizing normalized social compartment preference scores collapsed across
all three days of testing. Similarly, two-way repeated measures ANOVAs were utilized to
compare mean percentages of scan observations during conditioning in which the
mouse was in proximity to the stimulus between the two strains within the five- and ten-
day conditioning groups. A series of one-way ANOVAs were performed to compare
mean durations of grooming during the habituation and first testing day as well as the
difference of the two, to compare proportion of time spent grooming in each of the three
chambers. In cases where ANOVA analyses reveal significant main effects or
interactions, Bonferroni post-hoc comparisons were performed to compare strain means.
3.3 RESULTS
3.3.1 Social Conditioned Place Preference
Two- way repeated measures ANOVA revealed a significant main effect of Strain
(F(1,36)=4.196, p=0.048), indicating that BTBR mice showed decreased percent
preference for the socially-conditioned side compared to B6 mice. The main effect for
the number of Conditioning Days (F(1,36)=1.765, p=0.192) failed to reach statistical
significance. Strain x Conditioning Days (F(1,36)=0.099, p=0.755), and Strain x
Conditioning Days x Test Day (F(2,72)=0.295, p=0.746) interactions were not statistically
significant. When mean chamber preferences were collapsed across the three days,
two-way ANOVA revealed a significant effect of strain (F(1,116)=9.914, p=0.003) but the
interaction between Strain and number of Conditioning Days failed to reach statistical
significance (F(1,116)=0.216, p=0.643). Interestingly, with the mean values collapsed
across all three days of testing, the Conditioning Days main effect bordered statistical

41
significant (F(1,116)=3.867, p=0.052) suggesting that the amount of conditioning might
influence the strength of place preference formation. Figure 10b presents percent
preference (-baseline) for the socially-conditioned side across three days of testing in B6
and BTBR 5- and 10-day conditioned mice. Figure 10c displays the average preference
for the social compartment across all three days. Positive values indicate preference for
the compartment associated with the social interactions while negative values indicate
an aversion or decrease in preference for the social compartment, relative to baseline.
3.3.2 Social Interactions
For mice conditioned for five days, there was no significant main effects of Strain
or Day, or a Strain x Day interaction (F(1,72)= 0.212, p=0.651; F(4,72)= 0.880, p=0.481;
F(4,72)=1.502, p=0.211, respectively). Similarly, there were no significant main effects of
Strain or Day, or Strain x Day interaction for mice conditioned for ten days to social
interactions (F(1,162)= 4.148, p=0.057; F(9,162)= 0.850, p=0.572; F(9,162)=, 0.391, p=0.938
respectively). These analyses indicate that although there was a slight decrease in the
percent time BTBR mice were in proximity during conditioning; this effect was not
statistically significant and perhaps also was not behaviorally relevant with regards to
opportunity to form social reward-related contingencies. BTBR mice showed similar
proportions of time in proximity during conditioning. Figure 11 presents mean
percentages of scan observations of social proximity for five (a) and ten (b) day
conditioned mice.
3.3.3 Autogrooming
Figure 12 presents the mean percent time self-grooming in each of the three
chambers of the conditioning apparatus during the baseline session (a) as well as at the
first day of testing (b) and the difference between the two (c), which represents the mean
change in percent time grooming after social conditioning. Although significant
differences in time spent grooming in each of the three chambers was noted, most of the
variance contributing the difference was between the outer two conditioning chambers
and the center. If BTBR mice were particularly avoidant of social interactions because of
their pre-occupation with self-grooming, we would expect decreased percent time
grooming in the social compartment after conditioning and this was not noted. In fact,
BTBR mice spent as much time self-grooming in the social chamber as they did the non-
social chamber (Figure 12b,c). This suggests that BTBR mice are not avoiding social

42
contact or failing to be reinforced to the presence of a conspecific in order to groom
themselves in a context associated with isolation.
3.4 DISCUSSION
Utilizing the commonly-utilized three-chamber apparatus for sociability testing,
we have validated a novel approach to analyze direct social motivation deficits by
demonstrating an absence of place-preference elicited by an animate social incentive in
BTBR mice- a strain known to exhibit face and construct validity for autism. Though our
results revealed that the number of conditioning days did not contribute a statistically
significant influence on the variance, the means (Figure 10b) strongly indicate that more
days of conditioning elicit a stronger place preference; an effect noted previously (Thiel
et al., 2008).
Subject mouse behavior throughout testing was assessed to determine if the
amount of voluntary social contact during conditioning was related to differences in
social conditioned place preference. We noted that both strains, independent of social
phenotype, showed similar rates of social proximity during conditioning. Therefore, the
absence of conditioned place preference to social interactions in BTBR mice cannot be
solely explained by avoidance of social interactions during conditioning sessions.
Since BTBR mice groom themselves at abnormally high levels and at highly
stereotyped patterns (McFarlane et al, 2008; Pearson et al, 2011), it was predicted that
BTBR mice might avoid social stimuli in order to perform bouts of self-grooming, which is
a time when mice cannot engage in high rates of vigilance. Our results demonstrate that
BTBR mice spent a similar proportion of time grooming in the area associated with social
interactions after conditioning when compared to the non-social conditioned chamber.
Furthermore, grooming patterns were somewhat incongruent across chambers during
baseline sessions, particularly for BTBR mice. To rule out baseline side and/or grooming
preferences, the difference between the baseline and first test session were calculated
and compared. Even when eliminating the baseline rates in grooming preferences,
BTBR mice were not preferentially grooming in the non-social compartment and avoiding
self-grooming while inside of the social compartment. These results suggest that social
deficits in BTBR mice are not secondary to avoidance of social stimuli because of an
urge to engage in self-grooming.

43
An inherent issue in learning-based assays of motivation is potential cognitive
disturbances in the candidate model. BTBR mice have been shown to have a deficit in
footshock-based fear conditioning (MacPherson et al, 2008). However, MacPherson et
al. (2008) demonstrated intact object recognition abilities in BTBR mice. Furthermore,
Silverman et al, (2010b) noted that BTBR mice have a higher nociception threshold,
suggesting that the aforementioned fear conditioning impairment (MacPherson et al,
2008) may have something to do with sensory, rather than cognitive disparities. Newer
work by Amodeo and colleagues (2012) indicates that BTBR mice have impairments in
reversal learning, an impairment that is highly relevant to restricted interests and
insistence on sameness deficits in ASD. Acquisition and maintenance of a rewarded
contingency are required for CPP, so the perseveration deficits noted by Amodeo et al,
(2012) in the BTBR mice would presumably have little or no effect. Ultimately, the
responses to non-social positive reinforcers (psychostimulants or sweetened solutions)
should be examined to determine whether learning and/or memory deficits, which are
independent of social incentive, influence conditioning performance in the BTBR strain.
An additional limitation is that one cannot rule out aggression and sex as
unconditioned stimulus in these tasks since rodents will operantly respond to contexts
associated with agonistic and sexual interactions (Bell et al, 2010; Martínez et al, 1995;
Mathews et al, 2005). Another consideration is whether subjects should be individually
housed of subjects prior to conditioning; this manipulation has been shown to increase
social play in juvenile rats (Trezza et al., 2009) and may increase the positive incentive
value of the unconditioned stimulus (Douglas et al, 2004). Single housing of male
subjects might cause increased drive for agonistic interactions. Alternatively, revealing
strain or genotype distinctions in social conditioned place preference without social
isolation in the home cage may be preferable. Other important parameters are whether
the stimulus animals are siblings or unfamiliar, and the age of both. The study presented
herein utilized juvenile mice; a design which may have effectively diminished aggressive
or sexual motivating factors in the conditioned response.
By utilizing place-preference conditioning testing, we have modified the standard
three-chamber sociability test for the use as a social reinforcement assay. Our results
strongly suggest that BTBR mice lack social motivation. We propose social conditioning
may be an alternative and/or complementary test, to social approach paradigms for
social impairments in rodent models of ASD. This technique may aide in determining

44
neurobiological system impairments and novel therapeutics in animal models of ASD
where social motivation is impaired.
Figure 10. Social Conditioned Place Preference Testing in B6 and BTBR Mice. (a) Social conditioned place preference arena. (b) While B6 mice increased the proportion of time spent on the side associated with social interactions, BTBR mice showed a significantly decreased conditioned place preference to repeated social interactions over three consecutive days of testing. (c) When preference was collapsed across three days of testing, B6 mice showed a clear percent preference for the social conditioned side, relative to baseline (subtracted) and to BTBR mice. N=10/group.
45

Figure 11. Scan Sampling of Social Conditioning Sessions in B6 and BTBR Mice. No significant differences were noted in the percent of scan observations wherein the B6 or BTBR mice were in proximity to the stimulus mouse for those conditioned for five days (a) or for ten days (b). N=10/group.
46

Figure 12. Assessment of Autogrooming Behavior Before and After Social Conditioning During Exploration. Mean percentages of time spent self-grooming during baseline (a), test day one (b), and the difference between baseline and test day as a measure in change in self-grooming preferences for mice before and after social conditioning. Values represent Mean ± 1 S.E.M. Horizontal lines denote significant main effects: * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001. Bonferroni pairwise post-hoc comparisons: # p ≤ 0.05, λ p ≤ 0.01, p ≤ 0.001, Ψ p ≤ 0.0001. N=10/group.
47

48
3.5 PSYCHOSTIMULANT REACTIVITY
A potential confound to the social conditioning results would be the presence of
any cognitive impairments in BTBR mice, particularly in conditioning; this would
influence the ability of the mice to generate contingency, independent of social
motivation. Previous reports have demonstrated fear conditioning impairments (to
footshock) in the strain, relative to B6 mice (MacPherson et al, 2008). An obvious
challenge to the interpretation that BTBR mice are impaired in learning or memory is that
BTBR mice possess a higher nociception threshold (Silverman et al, 2010b). As such,
conditioning deficits should be assessed to non-painful reinforcers as well.
To ascertain whether SCPP impairments in BTBR mice stem from lack of social
reinforcement or a cognitive deficit, a cocaine-induced conditioned place preference was
performed in the same arenas. Figure 13 illustrates the change, relative to baseline, in
preference for the side associated with cocaine administration (4 days each of
alternating saline and cocaine conditioning). Two-way repeated measures analysis of
variance (ANOVA) revealed a significant strain effect. However, neither strain showed a
reliable increase in preference for the side associated with the chemical reinforcer. In
fact, BTBR mice showed an apparent aversion to the conditioning side, which is virtually
meaningless since it was independent of treatment. If forced to speculate, one could
argue that BTBR mice are more avoidant of the side the mouse was last placed into
after an injection, relative to B6 mice. Given the small sample size, and the degree of
variance, it would be difficult to assert that this is demonstrative of intact active
avoidance conditioning. Differential psychostimulant conditioning should be investigated
in these both the BTBR and MeCP2 strains in the future to fully realize their
characteristics specific to non-social positive associations.

Figure 13. Cocaine-Induced Conditioned Place Preference Testing in B6 and BTBR Mice. Percent preference for the CS-US paired side, relative to pre-conditioning baseline for three consecutive days in B6 and BTBR mice treated with saline or cocaine. Positive numbers represent increased preference for the US-paired side. Both strains failed to show a reliable conditioned place preference to cocaine administration. N=5/group.
49

50
3.5.1 Reward Tasks MeCP2308/Y Mutants
MeCP2 mutant and wild-type mice were tested for acute and chronic locomotor
responses to cocaine, a potent, dopamine (DA) indirect agonist, in a cocaine-induced
behavioral sensitization experiment. Mutant mice showed a decreased acute response
to cocaine, with neither genotype differing in behavioral sensitization to the drug (Figure
14). These data complement emerging work showing the pervasive and bi-directional
influence of MeCP2 on psychostimulant response and the psychostimulant influence on
MeCP2 expression. This test was performed to test the hypothesis that the hypersocial
phenotype of the mutant mice might result from enhanced reward sensitivity. The
obtained results are opposite to the prediction, and may indicate that the lack of MeCP2
protein may result in decreased reward sensitivity, driving an enhanced appetitive drive
to social engagement. A multitude of tests will need to be performed to assess this
possibility.
3.5.2 Psychostimulant Response in Low-Social Mice
A logical extension of the psychostimulant results in MeCP2 KO mice is to
determine whether the low-social BTBR mouse strain would show the opposite -
enhancement of acute locomotor response to cocaine. Within the CIPP experiment
mentioned previously, locomotor responses during the 20 minute conditioning sessions
to saline and cocaine, were analyzed from videotapes. BTBR mice showed a slight, but
consistent attenuation of locomotor response to cocaine, relative to saline, than did B6
mice (Figure 15). These data (reduced acute psychostimulant responses) are similar to
aforementioned data in the cocaine-induced behavioral sensitization task for MeCP2
mutants. Attenuated acute response to a psychostimulant drug may reflect DA
abnormalities in both MeCP2 mutant and BTBR mice despite opposing social
phenotypes.

* *
Figure 14. Locomotor Response to Repeated Cocaine Administration in MeCP2308/Y
Mutant and Wild-Type Mice. Upon daily, repeated injection of cocaine (10mg/kg) in an open field arena, MeCP2 mutant mice showed significantly decreased distance on the first day, relative to wild-type mice (a). Similarly, they showed significantly reduced total distance traveled (b) and velocity (c) on day 1. N=9/group.
51

Figure 15. Acute Locomotor Response to IP Cocaine in B6 and BTBR Mice. BTBR mice show an attenuated locomotor response to cocaine (10mg/kg) relative to saline injection, compared to B6 mice when accounting for the locomotor behavior in the previous day of saline injection. N=5/group.
52

53
3.6 DISCUSSION
Despite widespread analysis of the role of the mesolimbic reward neurocircuitry
in specific forms of social behaviors such as pair bonding and maternal behavior (e.g.
Champagne et al, 2004; Liu & Wang 2003), there has been very little consideration of
the role of this system in disturbances in other forms of social affiliation. The specific role
or function of reward processing in the brain of animal models of autism as well as in
human neurodevelopmental syndromes has not been sufficiently studied.
The neurobiology of behavior, but more specifically complex social behavioral
phenotypes are being considered in light of emerging evidence of dynamic changes in
genome regulation and neural plasticity (Hofmann 2003). These processes have vast
implications for normal and abnormal functioning, particularly with regards to
development, cognition and coping in humans and model organisms alike. Furthermore,
neural and behavioral plasticity phenomena, themselves, may prove to be critical and
effective targets for novel therapeutics (e.g. Manji et al, 2003).
The candidate mouse strains assessed here (BTBR inbred line and MeCP2
hypomorphs vs. wild-type inbred controls) show opposite, but nonetheless aberrant
behavioral displays relevant to autism. In light of emerging work demonstrating
pronounced disruptions of normal genetic and cellular plasticity cascades in these
candidates (Asaka et al, 2006; Deng et al, 2010; Martinowich et al, 2003; Stephenson et
al, 2011), it seems relatively evident that neural plasticity might be directly involved in
their syndrome-related behaviors. Pre-clinical research should aim to characterize the
specific mechanisms (ie. neurogenesis, synaptogenesis, epigenetics etc.) and their
interactions with known or predicted risk factors that culminate in pathology and
neurodevelopmental syndromes.

54
CHAPTER 4: N-SULFATED HEPARAN SULFATE ABUNDANCE AND DISTRIBUTION
IN MECP2308/Y MICE
4.1 INTRODUCTION
A major goal in research into autism and other related neurodevelopmental
disorders is to identify biomarkers that may aide in detection and treatment of these
troubling abnormalities (Walsh et al, 2011). Since MeCP2 protein regulates transcription
of a variety of genes, and MeCP2308/Y mutants have alterations in brain development and
neuroplasticity, a logical hypothesis would be that MeCP2308/Y mutants show alterations
in brain systems which regulate dynamic cyto-architectural and neurotrophic processes.
Emerging research has demonstrated that the subependymal layer of the lateral
ventricle (LV) contains small fractal structures (fractones) immunoreactive for laminin
(LAM) and n-sulfated heparan sulfate (HS) proteoglycans (Mercier et al, 2002).
Research suggests these constituents are crucial for growth factor binding and cognate
receptor signaling- processes which directly regulate cell proliferation and neurite growth
(Kerever et al, 2007). Their abundant presence in the sub-ependymal layer of the lateral
ventricle subventricular zone suggests a direct role in neurogenesis since this region and
the dentate gyrus are the two main adult neurogenic zones (Ming & Song 2005). More
specifically, n-sulfated HS sequesters a variety of growth and guidance factors that are
critical to normal stochastic processes in the adult brain (Yamaguchi 2001). Any
disturbances in the distribution or function of these HS proteoglycans, at any point in the
life-span, but particularly early in development, might severely disrupt brain circuit
formation and function.
Laminins are glycoproteins that are important constituents of the basement
membrane extracellular matrix (Timpl et al, 1979) and are expressed by endothelial and
epithelial cells (Foidart et al, 1980). As the most abundant glycoprotein constituent of
the basement membrane, laminin has central roles in cell-to-cell interactions as well as
is diverse signaling functions (Martin & Timpl 1987). Laminin is the first detectable
glycoprotein present in the basement membrane during embryogenesis, and binds
directly to heparan sulfate proteoglycans (Martin & Timpl 1987).
BTBR mice, an outstanding behavioral model of all three symptom clusters of
autism, show vast reductions in the amount of HS in the subventricular zone of the LVs
compared to B6 mice (Blanchard et al, 2012, Meyza et al, 2012). Furthermore, BTBR

55
mice show corpus callosum agenesis and a reduced hippocampal commissure
(Wahlsten et al, 2003). Although this work is ongoing, it suggests a potential mechanism
which might underlie aberrant brain development and the display of abnormalities in
social and repetitive behaviors reminiscent of autism. A major goal is to determine the
extent to which proteoglycan disturbances might affect normal functioning in models of
autism and other pervasive neurodevelopmental disorders. The following experiment
was designed to determine if n-sulfated heparan sulfate disturbances are characteristic
of MeCP2 mutant mice.
4.2 MATERIALS AND METHODS
4.2.1 Experimental Subjects
Adult (pnd 56-88) male MeCP2 WT and KO mice (n=4/genotype) were
euthanized and whole brains were removed and flash frozen in isopentane cooled to -
80ºC. Brain sections (10 µM) were collected and mounted on poly l-lysine (Sigma 1/5
dilution) coated glass slides and stored at -20ºC.
4.2.2 Dual-Channel Indirect Fluorescent Immunohistochemistry
Prior to immunolabeling, sections were fixed in acetone (-20ºC) for two minutes.
Sections were hybridized in wells created on the slide with a hydrophobic barrier pen
(Vector Labs). Primary antibodies directed at laminin (Sigma: rabbit IgG 1/1000) and HS
(Seikagaku Corp: 10E4: mouse IgM 1/400) were incubated for 120 minutes. Secondary
antibodies (Invitrogen: AlexaFluor 546 goat anti-rabbit IgG and AlexaFluor 488 goat anti-
mouse IgM) recognized the laminin and HS primaries, respectively and were incubated
for 40 minutes. After several rinses, slides were mounted and coverslipped with a
mounting medium containing DAPI nuclear marker (Vector Labs).
Microscope images (20X) were collected on an Olympus Fluoview 1000 laser
scanning confocal microscope utilizing a multi-argon laser for DAPI quantification as well
as red and green HeNe lasers for the laminin and heparan sulfate immunoreactivity (ir),
respectively, at the University of Hawaii Biological Electron Microscope Facility. High
resolution (1024 x 1024 pixel) images of the lateral wall of the right lateral ventricle were
combined as a montage in Photoshop and imported into ImageJ software (NIH) as 8 bit
RGB images. The images were divided into stacks which yielded grayscale images of
each of the three channels (Figure 16). The polygon selection tool was used to trace the
entire dorsal-ventral extent of the subventricular zone of the lateral aspect of the lateral

56
ventricle (caudate side) at each of five sections per subject (Bregma +1.0; +0.5; 0.0; -
0.5; and -1.0. Area, integrated density and mean gray value were collected for the
experimental zone selected as well as for three background areas which were used to
calculate corrected total fluorescence [Integrated density – (area of selected zone x
mean fluorescence of background images)] according to procedures previously
published (e.g. Burgess et al, 2010). Mean corrected total fluorescence for each
genotype at each region for the laminin- and heparan sulfate-immunoreactivity (LAM-ir,
HS-ir, respectively) as well as the heparan sulfate/laminin ratio (HS/LAM-ir) was
compared with Two-way analyses of variance (ANOVA) with Strain and AP Point as the
between- and within-subjects factors, respectively.
4.3 RESULTS
4.3.1 Laminin
Figure 17a displays the mean level of laminin fluorescence (mean signal intensity
in arbitrary units) across the five anterio-posterior sub-ventricular walls. No significant
difference was noted in the mean LAM-ir across AP Points (F(4,24)=1.150, p=0.357).
Likewise, there was no significant main effect of Genotype across the AP extent
(F(1,24)=0.165, p=0.699). Finally, the Genotype x AP Point interaction failed to reach
statistical significance (F(4,24)=1.120, p=0.370).
4.3.2 Heparan Sulfate
Figure 17b displays the mean level of heparan sulfate fluorescence across the
five anterio-posterior sub-ventricular zones. No significant difference was noted in the
mean HS-ir across AP Points (F(4,24)=1.976, p=0.130). Likewise, there was no significant
main effect of Genotype across the AP extent (F(1,24)=0.162, p=0.702). Finally, the
Genotype x AP Point interaction failed to reach statistical significance (F(4,24)=1.800,
p=0.162).
4.3.3 Heparan/Laminin Ratio
Figure 17c displays the mean level of heparan sulfate:laminin fluorescence
across the five anterio-posterior sub-ventricular walls. No significant difference was
noted in the mean HS/LAM-ir across AP Points (F(4,24)=425, p=0.789). Likewise, there
was no significant main effect of Genotype across the AP extent (F(1,24)=0.212, p=0.661).
Finally, the Genotype x AP Point interaction failed to reach statistical significance
(F(4,24)=1.138, p=0.363).

57
4.4 DISCUSSION
Despite widespread behavioral anomalies in male mice with a truncated form of
the MeCP2 gene, the subventricular zone heparan sulfate/ laminin system appears to be
relatively intact in MeCP2308/Y mutant mice. These data are in stark contrast to those
obtained when comparing normal B6 mice to BTBR mice. BTBR mice show pronounced
reductions of both heparan and laminin in the lateral ventricle subventricular zone
(Meyza et al, 2012). It is thought that this distruption might be secondary to or causal to
the brain anatomical abnormalities such as callosal agenesis and collapsed anterior
lateral ventricles of this autism candidate strain (Meyza et al, 2012). In the current study,
the same coordinates were assessed for similar disturbances in wild-type and
MeCP2308/Y mutants, but no differences emerged.
Since heparan sulfates are large multi-unit and varying proteoglycan molecules,
they are not under transcriptional control, per se. Rather, a variety of enzymes modify
the structure of the polysaccharide side chains of the complex molecules largely through
sulfation patterns, which in turn affects its affinity for growth factors (Kerever et al, 2007;
Turnbull et al, 2001). On the contrary, these regulatory enzymes are under
transcriptional/translational regulation. MeCP2 mutation directly affects the abundance of
their transcripts in microarray analyses (Chahrour et al, 2008). Furthermore, though not
to any extent like that of BTBR mice, MeCP2 mutant mice show alterations in brain
growth factors (Nag et al, 2008) as well as brain morphological alterations suggesting
perturbed cellular development (Stearns et al, 2007). Ultimately, no differences in the
LAM-ir were expected, as the basement membrane signal should be relatively stable.
The main rationale for including it in these assays is to provide a unit of the measured
amount of basement membrane to determine how much of it, and to what extent, it is
immune-positive for heparan sulfate; thus the HS/LAM variable.
Since MeCP2 protein regulates expression of methyl-transferases and other
such heparan sulfate proteoglycan enzymes, this strongly suggests that MeCP2308/Y
truncated mutants might have altered abundance and distribution of n-sulfate heparan
sulfate in the brain subventricular zones. We did not note such an effect. The case that
N-sulfated regions on heparan sulfate proteoglycans are not disrupted with this particular
mutation, but other enriched domains are abnormal (ie. O-sulfated zones), is a distinct
possibility that has not been assessed here. Furthermore, alterations in N-sulfated

58
heparan sulfates may occur in the brain of MeCP2308/Y mutants but in areas outside of
the anterio-posterior boundaries (Bregma +1.0 to -1.0) and/or in areas outside of the
subventricular zone. Alterations might also exist in other developmental times
(embryonically, and early postnatal periods) and these were not noted with the analyses
performed in adult animals. Altogether, the absence of significant findings here does not
preclude a role of heparan sulfate proteoglycans in the abnormal behavioral and
physiological phenotype of this candidate mutant, but it does exemplify the possibility
that the HS system does not underlie altered neurodevelopmental trajectories in animal
models of autism or in those clinically affected with ASD.

DAPI HS MERGELAM
DAPI LAM HS MERGE
a
hgfe
b c d
Figure 16. Representative Confocal Fluorescent Immunohistochemistry Micrographs of Mouse Brain Lateral Ventricle Immunolabeled for Laminin and Heparan Sulfate. Ventral lateral ventricle revealed by DAPI nuclear marker (a) as well as laminin (b)and heparan sulfate (c) and the three channels merged (d, e). The three channels were separated into grayscale images (e-g) and the subventricular zone was manually traced (dotted yellow lines). ImageJ intensity analyses were performed in the selected regions to quantify the fluorescence immunoreactivity in the laminin (f) and heparan sulfate (g) channels.
59

a
b
c
Figure 17. Immunohistochemical Analyses of Brain Extracellular Matrix Markers Laminin and Heparan Sulfate in MeCP2308/Y Wild-Type and Mutant Mice. Laminin immunoreactivity (LAM-ir) throughout the antero-posterior extent of analysis was comparable (a). Likewise, no difference in the heparan sulfate immunoreactivity (HS-ir) was evident (b). The ratio of HS to LAM was also similar across the coordinates for wild-type (WT) and mutant (KO) mice (c). N=4/group.
60

61
CHAPTER 5: CONCLUSIONS AND FUTURE DIRECTIONS
Utilizing an engineered mouse mutant with alterations in social behavior, and
comparing its emotional and motivation characteristics in reference to normal siblings
and an inbred strain with known alterations in social behaviors, strides have been made
in characterizing the role of emotion, motivation and brain subventricular development
systems in mouse models of autism. Although the MeCP2308/Y mutant showed an
accentuation of social behavior rather than a deficit, this specific phenotype might prove
exceptionally useful in determining the brain mechanisms of altered social behavior. This
possibility is supported by the fact that the various types of engineered MeCP2
mutations can cause opposing and varied influences on social behaviors (Table 1).
Other findings of note in this mutant candidate were subtle changes in autogrooming
patterns that might show face validity to the motor stereotypies characteristic of Rett
syndrome (Hagberg et al, 1983). Furthermore, the influences of the MeCP2308/Y mutation
on fear-related anxiety responses and reactions to cocaine are notable. Despite these
advances, these results raise more questions than are answered. Altogether, the search
for novel comprehension of the pathobiology and treatment of autism-spectrum
disorders will require continued advances in basic and translational research into the
fundamental mechanisms of social and repetitive behavior symptoms.
5.1 NEUROBIOLOGY OF SOCIABILITY
Three major theoretical constructs are essential to our comprehension of the role
of the nervous system in social behaviors, but more specifically in the understanding of
the brain in social cognition (Adolphs 2001). More specifically, when assessing the
mechanisms of autism-relevant social disturbances, key questions must be regarded
with respect to what the deficits might reflect; the first is how the organism - more
specifically, the nervous system receives and processes exogenous social information
(Young et al, 2002). Disturbances in social information processing might contribute
greatly to the development of social impairments in neurodevelopmental disorders and
this influence must be considered in rodent models of ASD (Crawley 2004).
A second and perhaps equally important set of processes are the influences of
the gathered social information on emotional processes and motivation (Adolphs 2001).
Take, for example, the immense body of literature examining the differential activation of

62
the amygdala, a set of limbic nuclei principally involved in emotional processing, in
response to facial expressions and other social signals in ASD (Baron-Cohen 2000). The
notion that emotion- principally, the evolutionarily-preserved basic emotion systems,
might be directly involved in decisions about when and how to engage social stimuli
(which are potentially threatening or rewarding) are quite comprehensible. Similarly, the
motivational and emotional systems can thus integrate and differentially regulate the
motoric output systems which underlie behavior (Adolphs et al, 2001). This regulatory
model suggests how the organism is afforded additional opportunities for social learning
and reward (through feed-forward experience) which shape the cognitive and emotional
experience of a social entity.
The work illustrated here has been concerned mainly with assessing the
emotional and motivational contributions to pro-social or affiliative forms of social
behavior in male mice - forms of affiliation that are not secondary to reproduction. The
theoretical framework should apply also to other forms of social behavior (parenting,
mating and aggression) but the emphasis here is to analyze behaviors in a manner that
informs the neurobiology of social avoidance in ASD.
5.2 THE ASSOCIATIONS OF NEURODEVELOPMENT AND
NEUROPLASTICITY WITH BEHAVIOR
Discoveries surrounding the dynamic and experience-driven changes in the brain
and genome are fast revolutionizing biological and psychological sciences. Yet,
researchers have only scratched the surface in the revelation of relationships between
genes, experience and cellular plasticity (neuron/neurite/synapse proliferation and
development) and behavioral phenotypes. Crucial studies will be those that reveal direct
relationships between dynamic changes within the brain and forms of behavior that are
disturbed in autism. What’s more, the developmental conditions and exogenous
perturbations that regulate these brain changes will need to be understood in greater
detail. Based upon the data presented here and the expanding literature of the role of
MeCP2, there is a substantial amount of support that engineered mutations of the
MeCP2 gene can reveal mechanisms of the relationships between neural plasticity, DNA
methylation, histone modification and behaviors relevant to psychological and behavioral
impairments throughout development and within adulthood. This notion is best illustrated
by studies demonstrating MeCP2 is critical for normal neuroplastic responses to

63
psychostimulant drugs of abuse (Deng et al, 2010) and those showing that post-mortem
tissue collected from those affected with Rett syndrome have decrements in
hippocampal (CA1 pyramidal cell) spine density and structure, suggesting synaptic
plasticity abnormalities (Chapleau et al, 2009). Altogether, impairments in neural activity-
dependent phosphorylation of MeCP2 have been shown to regulate function and
development of the nervous system through its gene-regulatory influences stemming
from chromatin influences (Cohen et al, 2011). How MeCP2 is involved in social
cognition will require intensive assessment, but this suggests that the impairments in
response to drugs of abuse might also underlie abnormalities in the socio-emotional
phenotype of MeCP2 hypomorphs.
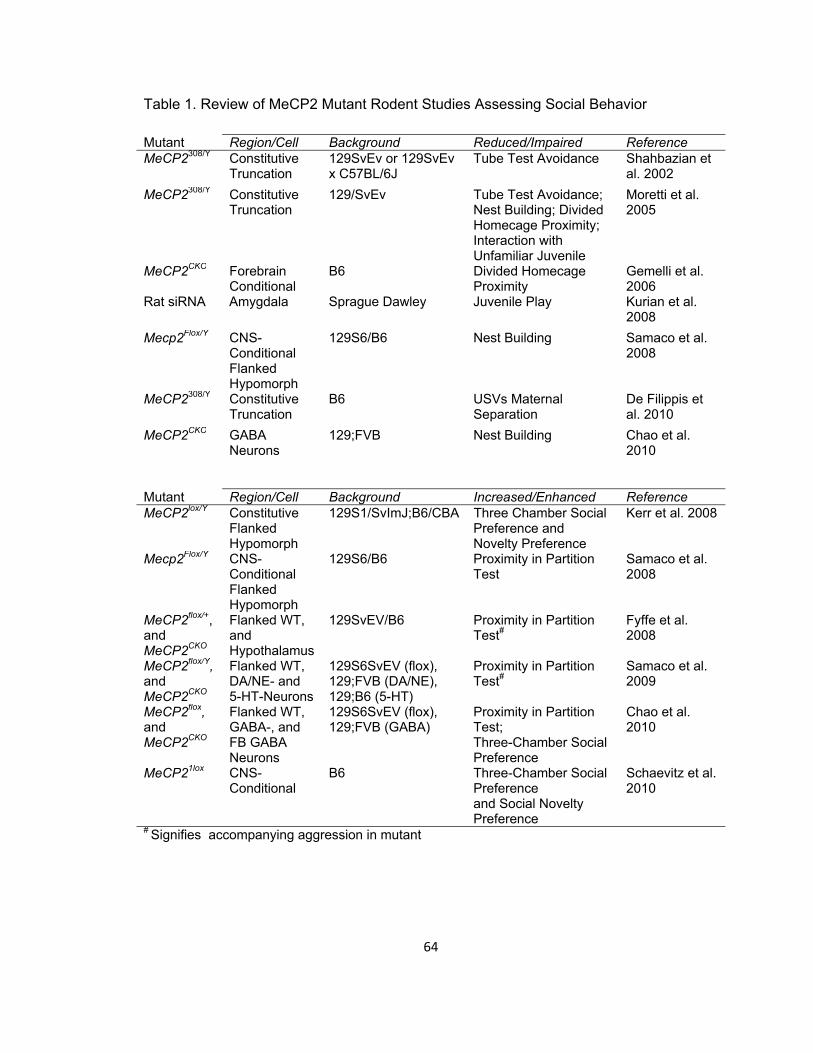
64
Table 1. Review of MeCP2 Mutant Rodent Studies Assessing Social Behavior
Mutant Region/Cell Background Reduced/Impaired Reference MeCP2308/Y Constitutive
Truncation 129SvEv or 129SvEv x C57BL/6J
Tube Test Avoidance Shahbazian et al. 2002
MeCP2308/Y Constitutive Truncation
129/SvEv Tube Test Avoidance; Nest Building; Divided Homecage Proximity; Interaction with Unfamiliar Juvenile
Moretti et al. 2005
MeCP2CKO Forebrain Conditional
B6 Divided Homecage Proximity
Gemelli et al. 2006
Rat siRNA Amygdala Sprague Dawley Juvenile Play Kurian et al. 2008
Mecp2Flox/Y CNS-Conditional Flanked Hypomorph
129S6/B6 Nest Building Samaco et al. 2008
MeCP2308/Y Constitutive Truncation
B6 USVs Maternal Separation
De Filippis et al. 2010
MeCP2CKO GABA Neurons
129;FVB Nest Building Chao et al. 2010
Mutant Region/Cell Background Increased/Enhanced Reference MeCP2lox/Y Constitutive
Flanked Hypomorph
129S1/SvImJ;B6/CBA Three Chamber Social Preference and Novelty Preference
Kerr et al. 2008
Mecp2Flox/Y CNS-Conditional Flanked Hypomorph
129S6/B6 Proximity in Partition Test
Samaco et al. 2008
MeCP2flox/+, and MeCP2CKO
Flanked WT, and Hypothalamus
129SvEV/B6 Proximity in Partition Test#
Fyffe et al. 2008
MeCP2flox/Y, and MeCP2CKO
Flanked WT, DA/NE- and 5-HT-Neurons
129S6SvEV (flox), 129;FVB (DA/NE), 129;B6 (5-HT)
Proximity in Partition Test#
Samaco et al. 2009
MeCP2flox, and MeCP2CKO
Flanked WT, GABA-, and FB GABA Neurons
129S6SvEV (flox), 129;FVB (GABA)
Proximity in Partition Test; Three-Chamber Social Preference
Chao et al. 2010
MeCP21lox CNS-Conditional
B6 Three-Chamber Social Preference and Social Novelty Preference
Schaevitz et al. 2010
# Signifies accompanying aggression in mutant
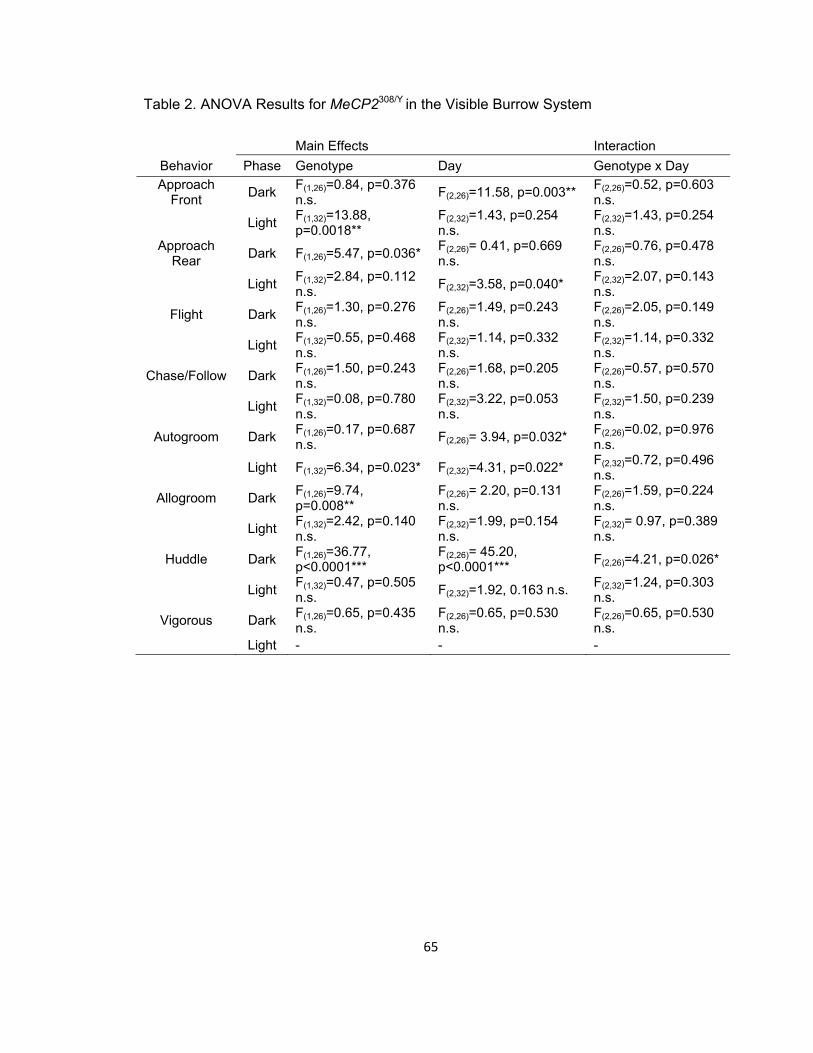
Table 2. ANOVA Results for MeCP2308/Y in the Visible Burrow System
Main Effects Interaction Behavior Phase Genotype Day Genotype x Day Approach
Front Dark F(1,26)=0.84, p=0.376 n.s. F(2,26)=11.58, p=0.003** F(2,26)=0.52, p=0.603
n.s.
Light F(1,32)=13.88, p=0.0018**
F(2,32)=1.43, p=0.254 n.s.
F(2,32)=1.43, p=0.254 n.s.
Approach Rear Dark F(1,26)=5.47, p=0.036* F(2,26)= 0.41, p=0.669
n.s. F(2,26)=0.76, p=0.478 n.s.
Light F(1,32)=2.84, p=0.112 n.s. F(2,32)=3.58, p=0.040* F(2,32)=2.07, p=0.143
n.s.
Flight Dark F(1,26)=1.30, p=0.276 n.s.
F(2,26)=1.49, p=0.243 n.s.
F(2,26)=2.05, p=0.149 n.s.
Light F(1,32)=0.55, p=0.468 n.s.
F(2,32)=1.14, p=0.332 n.s.
F(2,32)=1.14, p=0.332 n.s.
Chase/Follow Dark F(1,26)=1.50, p=0.243 n.s.
F(2,26)=1.68, p=0.205 n.s.
F(2,26)=0.57, p=0.570 n.s.
Light F(1,32)=0.08, p=0.780 n.s.
F(2,32)=3.22, p=0.053 n.s.
F(2,32)=1.50, p=0.239 n.s.
Autogroom Dark F(1,26)=0.17, p=0.687 n.s. F(2,26)= 3.94, p=0.032* F(2,26)=0.02, p=0.976
n.s.
Light F(1,32)=6.34, p=0.023* F(2,32)=4.31, p=0.022* F(2,32)=0.72, p=0.496 n.s.
Allogroom Dark F(1,26)=9.74, p=0.008**
F(2,26)= 2.20, p=0.131 n.s.
F(2,26)=1.59, p=0.224 n.s.
Light F(1,32)=2.42, p=0.140 n.s.
F(2,32)=1.99, p=0.154 n.s.
F(2,32)= 0.97, p=0.389 n.s.
Huddle Dark F(1,26)=36.77, p<0.0001***
F(2,26)= 45.20, p<0.0001*** F(2,26)=4.21, p=0.026*
Light F(1,32)=0.47, p=0.505 n.s. F(2,32)=1.92, 0.163 n.s. F(2,32)=1.24, p=0.303
n.s.
Vigorous Dark F(1,26)=0.65, p=0.435 n.s.
F(2,26)=0.65, p=0.530 n.s.
F(2,26)=0.65, p=0.530 n.s.
Light - - -
65

66
Table 3. Comparison of MeCP2308/Y Mutant and Wild-type mice in the Mouse Defense
Test Battery
Y/+ Y/-
Pre-test activity Mean ± 1 S.E.M. Result
Line crossings 108.56 ± 9.48 99.11 ± 9.31 t(16)= 0.71, p=0.487
Rears 6.33 ± 1.34 6.00 ± 1.40 t(16)= 0.17, p=0.866
Predator avoidance test
Avoidance distance (cm) 36.67 ± 14.14 141.11 ± 34.90 U=14.5, p=0.021 Escape distance (cm) 431.11 ± 39.03 400.00 ± 44.72 t(16)=0.52, p=0.607
Chase/flight test
Flight speed (m/sec) 0.315 ± 0.03 0.512 ± 0.04 t(16)= -4.37, p<0.001 Stops 11.90 ± 1.24 7.67 ± 1.03 t(16)= 2.62, p=0.018 Jump Escapes 0.00 ± 0.00 0.33 ± 0.17 t(16)= -2.00, p=0.063
Reversals 1.22 ± 0.52 0.00 ± 0.00 t(16)= 2.35, p=0.032 Straight alley test
Approaches/withdrawals 1.33 ± 0.29 0.22 ± 0.15 t(16)= 3.43, p=0.003 Contacts 0.89 ± 0.26 0.00 ± 0.00 t(16)= 3.41, p=0.004 Freezing (sec) 0.00 ± 0.00 1.78 ± 0.83 t(16)= -2.14, p=0.048 Uprights 3.89 ± 0.61 2.56 ± 0.82 t(16)= 1.31, p=0.210
Forced contact test
Vocalizations 1.11 ± 0.75 0.11 ± 0.11 U=26.5, p=0.115
Uprights 5.56 ± 1.44 2.33 ± 0.62 U=19.5, p=0.061
Jump attacks 0.56 ± 0.24 0.33 ± 0.30 t(16)= -1.08, p=0.297
Jump escapes 2.33 ± 0.60 6.56 ± 0.94 t(16)= -3.77, p=0.002 Bites 0.00 ± 0.00 0.22 ± 0.22 t(16)= -1.00, p=0.332
Post-test
Line crossings 108.22 ± 6.41 117.56 ± 10.04 t(16)= -0.78, p=0.445
Rears 14.44 ± 0.90 20.22 ± 1.67 t(16)= -3.05, p=0.008
n=9/ group.

67
REFERENCES
Adolphs R (2001). The neurobiology of social cognition. Curr Opin Neurobiol 11: 231-
239.
Amir RE, Van den Veyver IB, Wan M, Tran CQ, Francke U, Zoghbi HY (1999) Rett
syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding
protein 2. Nat Genet 23(2):185–188.
American Psychiatric Association (2000) Diagnostic criteria for 299.00 autistic disorder.
In: Diagnostic and statistical manual of mental disorders, 4th, text revision, DSM-IV-TR
ed.
Amodeo DA, Jones JH, Sweeney JA, Ragozzino ME (2012). Differences in BTBR T+tf/J
and C57BL/6J mice on probabilistic reversal learning and stereotyped behaviors. Behav
Brain Res 227(1): 64-72.
Arakawa H, Blanchard DC, Blanchard RJ (2007) Colony formation of C57BL/6J mice in
visible burrow system: identification of eusocial behaviors in a background strain for
genetic animal models of autism. Behav Brain Res 176(1):27-39.
Arakawa H, Arakawa K, Blanchard DC, Blanchard RJ (2009) Social features of scent-
donor mice modulate scent marking of C57BL/6J recipient males. Behav Brain Res
205(1):138-145.
Asaka Y, Jugloff DGM, Zhang L, Eubanks JH, Fitzsimonds RM (2006). Hippocampal
synaptic plasticity is impaired in the Mecp2-null mouse model of Rett syndrome.
Neurobiol Disease 21(1): 217-227.
Bader PL, Faizi M, Kim LH, Owen SF, Tadross MR, Alfa RW, Bett GCL, Tsien RW,
Rasmusson RL, Shamloo M (2011). Mouse model of Timothy syndrome recapitulates
triad of autistic traits. Proc Natl Acad Sci USA 108(37): 15432-15437.
Bardo MT, Bevins RA (2000). Conditioned place preference: what does it add to our
preclinical understanding of drug reward? Psychopharmacology 153(1): 31-43.

68
Baron-Cohen S, Ring HA, Bullmore ET, Wheelwright S, Ashwin C, Williams SCR (2000).
The amygdala theory of autism. Neurosci Biobehav Rev 24(3): 355-364.
Bell MR, Meerts SH, Sisk CL (2010). Male Syrian hamsters demonstrate a conditioned
place preference for sexual behavior and female chemosensory stimuli. Horm Behav
58(3), 410-414.
Benvenuto A, Moavero R, Alessandrelli R, Manzi B, Curatolo P (2009) Syndromic
autism: causes and pathogenic pathways. World J Pediatr 5(3): 169-176.
Betancur C (2011). Etiological heterogeneity in autism spectrum disorders: more than
100 genetic and genomic disorders and still counting. Brain Res 1380: 42-77.
Beyer KS, Blasi F, Bacchelli E, Klauck SM, Maestrini E, Poustka A. Mutation analysis of
the coding sequence of the MECP2 gene in infantile autism (2002). Hum Genet 111:
305–309.
Bienvenu T, Carrie A, de Roux N, Vinet MC, Jonveaux P, Couvert P, Villard L,
Arzimanoglou A, Beldjord C, Fontes M, Tardieu M, Chelly J (2000) MECP2 mutations
account for most cases of typical forms of Rett syndrome. Hum Mol Genet 9(9):1377–
1384.
Bill BR, Geschwind DH (2009). Genetic advances in autism: heterogeneity and
convergence on shared pathways. Curr Opin Genet Dev 19(3): 271-278.
Blanchard DC, Defensor EB, Meyza KZ, Pobbe RL, Pearson BL, Bolivar VJ, Blanchard
RJ (2012). BTBR T+tf/J mice: autism-relevant behaviors and reduced fractone-
associated heparan sulfates. Neurosci Biobehav Rev 36(1):285-296.
Braunschweig D, Simcox T, Samaco RC, LaSalle JM (2004). X-Chromosome
inactivation ratios affect wild-type MeCP2 expression within mosaic Rett syndrome and
Mecp2−/+ mouse brain. Hum Mol Genet 13(12): 1275-1286.

69
Burgess A, Vigneron S, Brioudes E, Labbé J-C, Lorca T, Castro A (2010). Loss of
human Greatwall results in G2 arrest and multiple mitotic defects due to deregulation of
the cyclin B-Cdc2/PP2A balance. Proc Natl Acad Sci USA 107: 12564–12569.
Calcagnetti DJ, Schechter MD (1992). Place conditioning reveals the rewarding aspect
of social interaction in juvenile rats. Physiol Behav 51(4), 667-672.
Calfa G, Percy AK, Pozzo-Miller L (2011) Experimental models of Rett syndrome based
on Mecp2 dysfunction. Exp Biol Med 236(1):3-19.
Carney RM, Wolpert CM, Ravan SA, Shahbazian M, Ashley-Koch A, Cuccaro ML,
Vance JM, Pericak-Vance MA (2003). Identification of MeCP2 mutations in a series of
females with autistic disorder. Pediatr Neurol 28: 205–211.
Cassel S, Carouge D, Gensburger C, Anglard P, Burgun C, Dietrich JB, Zwiller J (2006).
Fluoxetine and cocaine induce the epigenetic factors MeCP2 and MDB1 in adult rat
brain. Mol Pharmacol 70(2): 487-492.
Centers for Disease Control and Prevention (2009). Prevalence of Autism Spectrum
Disorders - Autism and Developmental Disabilities Monitoring Network, United States,
2006. 58(SS10): 1-20.
Chahrour M, Jung SY, Shaw C, Zhou X, Wong ST, Qin J, Zoghbi HY (2008). MeCP2, a
key contributor to neurological disease, activates and represses transcription. Science
320(5880): 1224-1229.
Champagne FA, Chretien P, Stevenson CW, Zhang TY, Gratton A, Meaney MJ (2004).
Variations in nucleus accumbens dopamine associated with individual differences in
maternal behavior in the rat. J Neurosci 24(17): 4113-4123.
Chapleau CA, Calfa GD, Lane MC, Albertson AJ, Larimore JL, Kudo S, Armstrong DL,
Percy AK, Pozzo-Miller L (2009). Dendritic spine pathologies in hippocampal pyramidal
neurons from Rett syndrome brain and after expression of Rett-associated MECP2
mutations. Neurobiol Dis 35: 219-233.

70
Chao HT, Chen H, Samaco RC, Xue M, Chahrour M, Yoo J, Neul JL, Gong S, Lu HC,
Heintz N, Ekker M, Rubenstein JL, Noebels JL, Rosenmund C, Zoghbi HY (2010).
Dysfunction in GABA signalling mediates autism-like stereotypies and Rett syndrome
phenotypes. Nature 468(7321): 263-269.
Chen RZ, Akbarian S, Tudor M, Jaenisch R (2001). Deficiency of methyl-CpG binding
protein-2 in CNS neurons results in a Rett-like phenotype in mice. Nat Genet 27(3): 327-
331.
Clayton-Smith J, Watson P, Ramsden S, Black GC (2000). Somatic mutation in MECP2
as a non-fatal neurodevelopmental disorder in males. Lancet 356(9232): 830-832.
Cohen S, Gabel HW, Hemberg M, Hutchinson AN, Sadacca LA, Ebert DH, Harmin DA,
Greenberg RS, Verdine VK, Zhou Z, Wetsel WC, West AE, Greenberg ME (2011).
Genome-wide activity-dependent MeCP2 phosophorylation regulated nervous system
development and function. Neuron 72(1): 72-85.
Crawley JN (2004). Designing mouse behavioral tasks relevant to autistic-like behaviors.
Ment Retard Dev Disabil Res Rev 10: 248-258.
Crawley JN (2007a) What’s wrong with my mouse?: behavioral phenotyping of
transgenic and knockout mice. John Wiley & Sons, Inc: Hoboken, New Jersey.
Crawley, J.N. (2007b). Mouse behavioral assays relevant to the symptoms of autism.
Brain Pathol 17(4), 448-459.
Dalal SJ, Estep JS, Valentin-Bon IE, Jerse AE (2001). Standardization of the Whitten
effect to induce susceptibility to Neisseria gonorrhoeae in female mice. Contemp Top
Lab Anim Sci 40(2): 13-17.
Dawson G, Meltzoff AN, Osterling J, Rinaldi J, Brown E (1998). Children with autism fail
to orient to naturally occurring social stimuli. J Autism Dev Disord 28(6): 479-485

71
Dawson G, Webb SJ, McPartland J (2005). Understanding the nature of face processing
impairment in autism: insights from behavioral and electrophysiological studies. Dev
Neuropsychol 27: 403–424.
De Filippis B, Ricceri L, Laviola G (2010). Early postnatal behavioral changes in the
Mecp2-308 truncation mouse model of Rett syndrome. Genes Brain Behav 9(2): 213-
223.
Defensor EB, Pearson BL, Pobbe RL, Bolivar VJ, Blanchard DC, Blanchard RJ (2011). A
novel social proximity test suggests patterns of social avoidance and gaze aversion-like
behavior in BTBR T+ tf/J mice. Behav Brain Res 217(2): 302-308.
Deng JV, Rodriguiz RM, Hutchinson AN, Kim IH, Wetsel WC, West AE (2010). MeCP2
in the nucleus accumbens contributes to neural and behavioral responses to
psychostimulants. Nat Neurosci 13(9): 1128-1136.
DiCicco-Bloom E, Lord C, Zwaigenbaum L, Courchesne E, Dager SR, Schmitz C,
Schultz RT, Crawley J, & Young LJ (2006). The developmental neurobiology of autism
spectrum disorder. J Neurosci 26(26): 6897-6906.
Dotti MT, Orrico A, De Stefano N, Battisti C, Sicurelli F, Severi S, Lam CW, Galli L,
Sorrentino V, Federico A (2002). A Rett syndrome MECP2 mutation that causes mental
retardation in men. Neurology 58(2):226-230.
Douglas LA, Varlinskaya EI, Spear LP (2004). Rewarding properties of social
interactions in adolescent and adult male and female rats: impact of social versus isolate
housing of subjects and partners. Dev Psychobiol 45(3): 153-162.
Evans, MJ, Duvel A, Funk ML, Lehman B, Sparrow J, Watson NT, Neuringer A (1994).
Social reinforcement of operant behavior in rats: a methodological note. J Exp Anal
Behav 62: 149-156.
Foidart JM, Bere EW Jr, Yaar M, Rennard SI, Gullino M, Martin GR, Katz SI (1980).
Distribution and immunoelectron microscopic localization of laminin, a noncollagenous
basement membrane glycoprotein. Lab Invest 42(3): 336-342.

72
Fritz M, El Rawas R, Salti A, Klement S, Mardo MT, Kemmler G, Dechant G, Saria A,
Zernig G (2011). Reversal of cocaine-conditioned place preference and
mesocorticolimbic Zif268 expression by social interaction in rats. Addict Biol 16(2): 273-
284.
Fyffe SL, Neul JL, Samaco RC, Chao HT, Ben-Shachar S, Moretti P, McGill BE,
Goulding EH, Sullivan E, Tecott LH, Zoghbi HY (2008). Deletion of Mecp2 in Sim1-
expressing neurons reveals a critical role for MeCP2 in feeding behavior, aggression,
and the response to stress. Neuron 59(6): 947-958.
Gemelli T, Berton O, Nelson ED, Perrotti LI, Jaenisch R, Monteggia LM (2006).
Postnatal loss of methyl-CpG binding protein 2 in the forebrain is sufficient to mediate
behavioral aspects of Rett syndrome in mice. Biol Psychiatry 59(5): 468-476.
Guy J, Hendrich B, Holmes M, Martin JE, Bird A (2001). A mouse Mecp2-null mutation
causes neurological symptoms that mimic Rett syndrome. Nat Genet 27(3): 322-326.
Hagberg B, Aicardi J, Dias K, Ramos O (1983). A progressive syndrome of autism,
dementia, ataxia, and loss of purposeful hand use in girls: Rett’s syndrome: report of 35
cases. Ann Neurol 14(4): 471–479.
Hagberg B (1985). Rett's syndrome: prevalence and impact on progressive severe
mental retardation in girls. Acta Paediatr Scand 74(3): 405–408.
Hammer S, Dorrani N, Dragich J, Kudo S, Schanen C (2002). The phenotypic
consequences of MECP2 mutations extend beyond Rett syndrome. Ment Retard Dev
Disabil Res Rev 8: 94-98.
Hofmann HA (2003). Functional genomics of neural and behavioral plasticity. J
Neurobiol 54(1): 272-282.
Ikemoto S, Panksepp J (1999). The role of nucleus accumbens dopamine in motivated
behavior: a unifying interpretation with special reference to reward-seeking. Brain Res
Rev 31(1): 6-41.

73
Im HI, Hollander JA, Bali P, Kenny PJ (2010). MeCP2 controls BDNF expression and
cocaine intake through homeostatic interactions with microRNA-212. Nat Neurosci 13(9):
1120-1127.
Insel TR, Fernald RD (2004). How the brain processes social information: searching for
the social brain. Annu Rev Neurosci 27: 697-722.
Jørgensen HF, Bird A (2002). MeCP2 and other methyl-CpG binding proteins. Ment
Retard Dev Disabil Res Rev 8(2): 87-93.
Kai K, Sabine K, Vincent E, Michael JM, Alois S, Gerald Z (2011). Conditioned place
preference for social interaction in rats: contribution of sensory components. Front
Behav Neurosci 5: 80.
Kalueff AV, Aldridge JW, LaPorte JL, Murphy DL, Tuohimaa P (2007). Analyzing
grooming microstructure in neurobehavioral experiments. Nat Protoc 2(10): 2538-2544.
Kanner L (1943). Autistic disturbances of affective contact. Nerv Child 2: 217-250.
Kaufmann WE, Johnston MV, Blue ME (2005). MeCP2 expression and function during
brain development: implication for Rett syndrome’s pathogenesis and clinical evolution.
Brain Dev 27: S77-S87.
Kerever A, Schnack J, Vellinga D, Ichikawa N, Moon C, Arikawa-Hirasawa E, Efird JT,
Mercier F (2007). Novel extracellular matrix structures in the neural stem cell niche
capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu.
Stem Cells 25(9): 2146-2157.
Kerr B, Alvarez-Saavedra M, Sáez MA, Saona A, Young JI (2008). Defective body-
weight regulation, motor control and abnormal social interactions in Mecp2 hypomorphic
mice. Hum Molec Genet 17(12): 1707-1717.

74
Kleefstra T, Yntema HG, Oudakker AR, Romein T, Sistermans E, Nillessen W, van
Bokhoven H, de Vries BB, Hamel BC (2002). De novo MECP2 frameshift mutation in a
boy with moderate mental retardation, obesity, and gynaecomastia. Clin Genet 61(5):
359-362.
Kurian JR, Bychowski ME, Forbes-Lorman RM, Auger CJ, Auger AP (2008). Mecp2
organizes juvenile social behavior in a sex-specific manner. J Neurosci 28(28): 7137-
7142.
Lang PJ, Bradley MM, Cuthbert BN (1998). Emotion, motivation, and anxiety: brain
mechanisms and psychophysiology. Biol Psychiatry 44(12): 1248-1263.
Lewis JD, Meehan RR, Henzel WJ, Maurer-Fogy I, Jeppesen P, Klein F, Bird A (1992).
Purification, sequence, and cellular localization of a novel chromosomal protein that
binds to methylated DNA. Cell 69(6): 905-914.
Lister RG (1987). The use of a plus-maze to measure anxiety in the mouse.
Psychopharmacology 92(2): 180-185.
Litvin Y, Blanchard DC, Pentkowski NS, Blanchard RJ (2007). A pinch or a lesion: a
reconceptualization of biting consequences in mice. Aggress Behav 33(6): 545-551.
Liu Y, Wang ZX (2003). Nucleus accumbens oxytocin and dopamine interact to regulate
pair bind formation in female prairie voles. Neuroscience 121(3): 537-544.
Long SY (1972). Hair-nibbling and whisker-trimming as indicators of social hierarchy in
mice. Anim Behav 20(1): 10–12.
MacPherson P, McGaffigan R, Wahlsten D, Nguyen PV (2008). Impaired fear memory,
altered object memory and modified synaptic plasticity in split-brain mice. Brain Res
1210: 179-188.
Malkesman O, Scattoni ML, Paredes D, Tragon T, Pearson B, Shaltiel G, Chen G,
Crawley JN, Manji HK (2010). The female urine sniffing test: a novel approach for
assessing reward-seeking behavior in rodents. Biol Psychiatry 67(9): 864-871.

75
Manji HK, Quiroz JA, Sporn J, Payne JL, Denicoff K, Gray NA, Zarate CA Jr, Charney
DS (2003). Enhancing neuronal plasticity and cellular resilience to develop novel,
improved therapeutics for Difficult-to-Treat depression. Biol Psychiatry 53(8): 707-742.
Martin GR, Timpl R (1987). Laminin and other basement membrane components. Ann
Rev Cell Biol 3: 57-85.
Martínez M, Guillén-Salazar F, Salvador A., Simón VM (1995). Successful intermale
aggression and conditioned place preference in mice. Physiol Behav 58(2): 323-328.
Martinowich K, Hattori D, Wu H, Fouse S, He F, Hu Y, Fan G, Sun YE (2003). DNA
methylation-related chromatin remodeling in activity-dependent Bdnf gene regulation.
Science 302(5646): 890-893.
Mathews JT, Abdelbaky P, Pfaff D (2005). Social and sexual motivation in the mouse.
Behav Neurosci 119(6): 1628-1639.
McFarlane HG, Kusek GK, Yang M, Phoenix JL, Bolivar VJ, Crawley JN (2008). Autism-
like phenotypes in BTBR T+tf/J mice. Genes Brain Behav 7(2): 152-163.
McGill BE, Bundle SF, Yaylaoglu MB, Carson JP, Thaller C, Zoghbi HY (2006).
Enhanced anxiety and stress-induced corticosterone release are associated with
increased Crh expression in a mouse model of Rett syndrome. Proc Natl Acad Sci USA
103(48): 18267-18272.
McGraw CM, Samaco RS, Zoghbi HY (2011). Adult neural function requires MeCP2.
Science 333(6039): 186.
Meloni I, Bruttini M, Longo I, Mari F, Rizzolio F, D'Adamo P, Denvriendt K, Fryns JP,
Toniolo D, Renieri A (2000). A mutation in the rett syndrome gene, MECP2, causes X-
linked mental retardation and progressive spasticity in males. Am J Hum Genet 67
(2000): 982–985.

76
Mercier F, Kitasako JT, Hatton GI (2002). Anatomy of the brain neurogenic zones
revisited: fractones and the fibroblast/macrophage network. J Comp Neurol 451(2):170-
188.
Meyza KZ, Blanchard DC, Pearson BL, Pobbe RL, Blanchard RJ (2012). Fractone-
associated N-sulfated heparan sulfate shows reduced quantity in BTBR T+tf/J mice: A
strong model of autism. Behav Brain Res 228(2): 247-253.
Ming G, Song H (2005). Adult neurogenesis in the mammalian central nervous system.
Annu Rev Neurosci 28: 223-250.
Moretti P, Bouwknecht JA, Teague R, Paylor R, Zoghbi HY (2005). Abnormalities of
social interactions and home-cage behavior in a mouse model of Rett syndrome. Hum
Mol Genet 14(2):205-220.
Moy SS, Nadler JJ, Perez A, Barbaro RP, Johns JM, Magnuson TR, Piven J, Crawley
JN (2004). Sociability and preference for social novelty in five inbred strains: an
approach to assess autistic-like behavior in mice. Genes Brain Behav 3(5): 287-302.
Moy SS, Nadler JJ (2008). Advances in behavioral genetics: mouse models of autism.
Mol Psychiatry 13: 4-26.
Moy SS, Nadler JJ, Poe MD, Nonneman RJ, Young NB, Koller BH, Crawley JN, Duncan
GE, Bodfish JW (2008). Development of a mouse test for repetitive, restricted behaviors:
relevance to autism. Behav Brain Res 188(1):178-194.
Nadler JJ, Moy SS, Dold G, Trang D, Simmons N, Perez A, Young NB, Barbaro RP,
Piven J, Magnuson TR, Crawley JN (2004). Automated apparatus for rapid quantitation
of autism-like social deficits in mice. Genes Brain Behav 3: 303–314.
Nag N, Mellott TJ, Berger-Sweeney JE (2008). Effects of postnatal dietary choline
supplementation on motor regional brain volume and growth factor expression in a
mouse model of Rett syndrome. Brain Res 1237: 101-109.

77
Nagarajan RP, Hogart AR, Gwye Y, Martin MR, LaSalle JM (2006). Reduced MeCP2
expression is frequent in autism frontal cortex and correlates with aberrant MECP2
promoter methylation. Epigenetics 1(4): e1-11.
Orrico A, Lam C, Galli L, Dotti MT, Hayek G, Tong SF, Poon PM, Zappella M, Federico
A, Sorrentino V (2000). MECP2 mutation in male patients with non-specific X-linked
mental retardation. FEBS Lett 481(3): 285-288.
Pacey LKK, Doss L, Cifelli C, van der Kooy D, Heximer SP, Hampson DR (2011).
Genetic deletion of regulator of G-protein signaling 4 (RGS4) rescues a subset of fragile
X phenotypes in the FMR1 knockout mouse. Mol Cell Neurosci 46: 563-572.
Panksepp JB, Jochman KA, Kim JU, Koy JJ, Wilson ED, Chen Q, Wilson CR, Lahvis GP
(2007). Affiliative behavior, ultrasonic communication and social reward are influenced
by genetic variation in juvenile mice. PLoS One 2: e351.
Panksepp JB, Lahvis GP (2007). Social reward among juvenile mice. Genes Brain
Behav 6(7): 661-671.
Pearson BL, Defensor EB, Blanchard DC, Blanchard RJ (2010). C57BL/6J mice fail to
display preference for social novelty in the three-chamber apparatus. Behav Brain Res
213(2):189-194.
Pearson BL, Defensor EB, Pobbe RLH, Yamamoto L, Bolivar VJ, Blanchard DC,
Blanchard RJ (In Press) MeCP2 mutation in male mice promotes affiliative social
behavior. Behav Genet 42(2): 299-312.
Pearson BL, Pobbe RL, Defensor EB, Oasay L, Bolivar VJ, Blanchard DC, Blanchard RJ
(2011) Motor and cognitive stereotypies in the BTBR T+tf/J mouse model of autism.
Genes Brain Behav 10(2): 228-235.

78
Pelka GJ, Watson CM, Radziewic T, Hayward M, Lahooti H, Christodoulou J, Tam PPL
(2006). Mecp2 deficiency is associated with learning and cognitive deficits and altered
gene activity in the hippocampal region of mice. Brain 129(4): 887–898.
Pobbe RL, Pearson BL, Defensor EB, Bolivar VJ, Blanchard DC, Blanchard RJ (2010).
Expression of social behaviors of C57BL/6J versus BTBR inbred mouse strains in the
visible burrow system. Behav Brain Res 214(2): 443-449.
Ricceri L, De Filippis B, Laviola G (2008). Mouse models of Rett syndrome: from
behavioural phenotyping to preclinical evaluation of new therapeutic approaches. Behav
Pharmacol 19(5-6): 501-517.
Rosen JB (2004). The neurobiology of conditioned and unconditioned fear: a
neurobehavioral system analysis of the amygdale. Behav Cogn Neurosci Rev 3(1): 23-
41.
Samaco RC, Fryer JD, Ren J, Fyffe S, Chao HT, Sun Y, Greer JJ, Zoghbi HY, Neul JL
(2008). A partial loss of function allele of Methyl-CpG-binding protein 2 predicts a human
neurodevelopmental syndrome. Hum Mol Genet 17(12): 1718-1727.
Samaco RC, Mandel-Brehm C, Chao HT, Ward CS, Fyffe-Maricich SL, Ren J, Hyland K,
Thaller C, Maricich SM, Humphreys P, Greer JJ, Percy A, Glaze DG, Zoghbi HY, Neul
JL (2009). Loss of MeCP2 in aminergic neurons causes cell-autonomous defects in
neurotransmitter synthesis and specific behavioral abnormalities. Proc Natl Acad Sci
USA 106(51): 21966-21971.
Schaevitz LR, Moriuchi JM, Nag N, Mellot TJ, Berger-Sweeney J (2010). Cognitive and
social functions and growth factors in a mouse model of Rett syndrome. Physiol Behav
100(3): 255-263.
Schultz RT (2005). Developmental deficits in social perception in autism: the role of the
amygdala and fusiform face area. Int J Dev Neurosci 23: 125-141.

79
Scott-Van Zeeland AA, Dapretto M, Ghahremani DG, Poldrack RA, Bookheimer SY
(2010) Reward processing in autism. Autism Res 3(2): 53-67.
Shahbazian M, Young J, Yuva-Paylor L, Spencer C, Antalffy B, Noebels J, Armstrong D,
Paylor R, Zoghbi H (2002). Mice with truncated MeCP2 recapitulate many Rett
syndrome features and display hyperacetylation of histone H3. Neuron 35(2): 243–254.
Silverman JL, Yang M, Lord C, Crawley JN (2010a). Behavioral phenotyping assays for
mouse models of autism. Nat Rev Neurosci 11(7): 490-502.
Silverman JL, Yang M, Turner SM, Katz AM, Bell DB, Koenig JI, Crawley JN (2010b).
Low stress reactivity and neuroendocrine factors in the BTBR T+tf/J mouse model of
autism. Neuroscience 171(4): 1197-1208.
Stearns NA, Schaevitz LR, Bowling H, Nag N, Berger UV, Berger-Sweeney J (2007).
Behavioral and anatomical abnormalities in Mecp2 mutant mice: a model for Rett
syndrome. Neuroscience 146(3): 907-921.
Stephenson DT, O’Neill SM, Narayan S, Tiwari A, Arnold E, Samaroo HD, Du F, Ring
RH, Campbell B, Pletcher M, Vaidya VA, Morton D (2011). Histopathalogical
characterization of the BTBR mouse model of autistic-like behavior reveals selective
changes in neurodevelopmental proteins and adult hippocampal neurogenesis. Mol
Autism 2:7.
Thiel KJ, Okun AC, Neisewander JL (2008). Social reward-conditioned place preference:
a model revealing an interaction between cocaine and social context reward in rats.
Drug Alcohol Depend 96(3): 202-212.
Timpl R, Rohde H, Robey PG, Rennard SI, Foidart J, Martin GR (1979). Laminin-a
glycoprotein from basement membranes. J Biol Chem 254(19): 9933-9937.
Trezza V, Damsteegt R, Vanderschuren LJ (2009). Conditioned place preference
induced by social play behavior: parametrics, extinction, reinstatement and disruption by
methylphenidate. Eur Neuropsychopharmacol 19(9): 659-669.

80
Turnbull J, Powell A, Guimond S (2001). Heparan sulfate: decoding a dynamic
multifunctional cell regulator. Trends Cell Biol 11(2): 75-82.
Van Esch H, Bauters M, Ignatius J, Jansen M, Raynaud M, Hollanders K, Lugtenberg D,
Bienvenu T, Jensen LR, Gecz J, Moraine C, Marynen P, Fryns JP, Froyen G (2005).
Duplication of the MECP2 region is a frequent cause of severe mental retardation and
progressive neurological symptoms in males. Am J Hum Genet. 77(3): 442–453.
Wahlsten D, Metten P, Crabbe JC (2003). Survey of 21 inbred mouse strains in two
laboratories reveals that BTBR T/+ tf/tf has severely reduced hippocampal commissure
and absent corpus callosum. Brain Res 971(1): 47-54.
Walsh P, Elsabbagh M, Bolton P, Singh I (2011). In search of biomarkers for autism:
scientific, social and ethical challenges. Nat Rev Neurosci 12: 603-612.
Wan M, Lee SS, Zhang X, Houwink-Manville I, Song HR, Amir RE, Budden S, Naidu S,
Pereira JL, Lo IF, Zoghbi HY, Schanen NC, Francke U (1999). Rett syndrome and
beyond: recurrent spontaneous and familial MECP2 mutations at CpG hotspots. Am J
Hum Genet 65(6): 1520-1529.
Waterhouse L, Fein D, Modahl C (1996). Neurofunctional mechanisms in autism.
Psychol Rev 103: 457-489.
Wu H, Tao J, Chen PJ, Shahab A, Ge W, Hart RP, Ruan X, Ruan Y, Sun YE (2010).
Genome-wide analysis reveals methyl-CpG-binding protein 2-dependent regulation of
microRNAs in a mouse model of Rett syndrome. Proc Natl Acad Sci USA 107(42):
18161-18166.
Yamaguchi Y (2001). Heparan sulfate proteoglycans in the nervous system: their diverse
roles in neurogenesis, axon guidance, and synaptogenesis. Semin Cell Dev Biol 12(2):
99-106.

81
Young JI, Hong EP, Castle JC, Crespo-Barreto J, Bowman AB, Rose MF, Kang D,
Richman R, Johnson JM, Berget S, Zoghbi HY (2005). Regulation of RNA splicing by the
methylation-dependent transcriptional repressor methyl-CpG binding protein 2. Proc Natl
Acad Sci USA 102(49): 17551-17558.