public health risk of giardia and cryptosporidium posed by ... · public health risk of giardia and...
TRANSCRIPT

Public health risk of Giardia and Cryptosporidium posed by reintroduction of
beavers into Scotland
Background
Following publication of ‘Scottish Beaver Trial Independent Public Health Monitoring 2009-2014 Report and
Recommendations’ (Mackie, 2014), two pieces of complementary work were undertaken in parallel to assess
the potential contribution of reintroduced beavers in Scotland to the public health burden of disease
attributed to Giardia spp. and Cryptosporidium spp. parasites. The first, a risk assessment,
addressing the question ‘What is the likelihood that re-introduced beavers will have a significant
impact on the contamination of drinking water supplies with Cryptosporidium parvum and Giardia
lamblia?’ (Appendix 1), was conducted by Scottish Government’s Centre of Expertise on Animal
Disease Outbreaks (EPIC). This reviewed evidence from data and publications across the world, as
well as evidence from the beaver trial and SNH’s Tayside beaver reports, and used this to assess the
likely additional contribution of beavers to the risk associated with exposure to these parasites in
Scotland. The second, ‘What is the likelihood that beavers will be an important source of
contamination of drinking water supplies with Cryptosporidium parvum and Giardia intestinalis?’
(Appendix 2), was prepared by Health Protection Scotland (HPS), Scottish Parasite Diagnostic
Reference Laboratory (SPDL) and Drinking Water Quality Regulator for Scotland (DWQR). This
reviewed the diagnostics, surveillance and epidemiology of these infections in people in Scotland.
The summary conclusions and recommendations of the two reports are as follows:
Conclusions
Human infection with Cryptosporidium and Giardia spp. results in time-limited diarrhoeal
disease in otherwise healthy humans. However, in immunosuppressed individuals, these
parasites may have life-threatening consequences.
Beavers may carry Giardia and Cryptosporidium and may contaminate raw water sources.
They are thus likely to form part of the reservoir community for both parasites.
Other sources of infection are likely to result in a greater numbers of oocysts/cysts shed into
the environment (such as humans, livestock and other existing wildlife) and will pose a more
significant risk to water contamination than beavers.
Therefore, the reintroduction of beavers is likely to present a very small additional risk as the
number of beavers excreting these organisms will be likely to be small relative to the large
number of wild, domestic and livestock animals and humans which contribute to existing
public health risk.

Further, based on the current epidemiology and implemented control measures, the main
potential route of additional risk of infection with Cryptosporidium or Giardia is via the
contamination of drinking water supplies; however:
o The majority of the population is served by public water supplies which have
effective barriers and monitoring in place such that the introduction of beavers is
unlikely to pose an additional public health risk via this route
o A smaller proportion of the population which is served by, or exposed to, private
water supplies
o Owners of private water supplies are advised, encouraged and supported by Scottish
Government and local authority initiatives to ensure adequate treatment processes
are in place to prevent microbiological contamination
o HPS and the SPDRL work together to provide robust surveillance of human cases of
these infections and recent trends show downward trends for both organisms
o Increased monitoring and epidemiological assessment should continue including
molecular studies (human, animal and environmental) to further characterise likely
sources of acquisition
o There is no evidence from Norway that the presence of beavers has adversely
affected the number of human cases of Cryptosporidium or Giardia
o The public is provided with advice about how to avoid exposure to infection when
enjoying the countryside
Recommendations
As a precaution, and to provide further assurance, the following are recommended:
Enhanced surveillance by HPS and SPDRL of all cryptosporidiosis and giardiasis human cases
within Scotland for a pre-determined period e.g. twelve months.
Continued promotion of best practice in relation to public and private water supplies and
the public’s interaction with the countryside.
Plans to re-introduce beavers in PWS catchments should be discussed with local authority
Environmental Health teams to ensure that risk is fully evaluated.
Any proposed re-introduction of beavers should be discussed in detail with Scottish Water to
allow appropriate risk assessments to be undertaken and water safety plans to be modified
if required.

Appendix 1
Public health risk of Giardia and Cryptosporidium posed by re-introduced beavers into Scotland.
RISK QUESTION: What is the likelihood that re-introduced beavers will have a significant impact on the contamination of drinking water supplies with Cryptosporidium parvum and Giardia lamblia?
1. SUMMARY OF OVERALL RISK AND RECOMMENDED ACTION This risk assessment was compiled according to terms of reference provided by the Scottish Government regarding time of delivery, title of veterinary risk assessment (VRA) and level of detail required. EPIC scientists have created a generic framework suitable for VRAs; collated and updated existing information on risks. This document may require updating as new information becomes available or legislation develops, or if more in-depth assessment is necessary. DEFINITIONS OF RISK LEVEL (Defra, 2011; OIE, 2004) Negligible: So rare that it does not merit consideration Very low: Very rare but cannot be excluded Low: Rare but could occur Medium: Occurs regularly High: Occurs very often Very High: Events occur almost certainly RISK ESTIMATION: The likelihood of beavers as an important source of contamination of Cryptosporidium and Giardia spp. to water supplies is VERY LOW to LOW (HIGH UNCERTAINTY) in the context of other sources of contamination (such as humans, livestock, other wildlife and domestic animals).
2.MAIN MESSAGES
A. Likely sources of water contamination with Cryptosporidium spp. and Giardia spp. A1. Likelihood that beavers are a source of water contamination: The likelihood of beavers as a source of contamination is VERY LOW TO LOW (HIGH UNCERTAINTY) but contingent on the source country of any reintroduced beavers, the presence of infection and adequacy of testing to detect infection and excretion of oocysts prior to reintroduction. Once beavers are reintroduced to Scottish waters, there is LOW – MEDIUM likelihood (HIGH UNCERTAINTY) that beavers will become infected with these parasites already in Scottish waters. There is strong evidence that beavers are hosts for these parasites. However, the evidence for amplification is HIGHLY UNCERTAIN as few studies have investigated this. A2. Likelihood that wildlife is a source of raw untreated water contamination is MEDIUM to HIGH with HIGH UNCERTAINTY. Infection status is inferred because of the known

susceptibility of certain wildlife species in the area, and the presence of parasites in the water supply. A3. Likelihood that livestock is a source of contamination of raw untreated water supplies is HIGH, LOW-MEDIUM UNCERTAINTY. There is strong evidence to support the view that infection with Giardia and Cryptosporidium may be endemic on some farms; this increases the likelihood that it will become a source of environmental and water contamination. A4. Likelihood that domestic pets are a source of contamination of raw untreated water supplies is NEGLIGIBLE. There is MEDIUM-HIGH UNCERTAINTY about the role of domestic pets as a source of outbreaks through direct or indirect transmission. A5. Likelihood that humans are a source of contamination of raw untreated water supplies is MEDIUM to HIGH. Uncertainty is LOW as there is evidence that control of sewage is an important component of reducing exposure to water-borne parasites.
B Transmission pathway B1. Likelihood of consumption of contaminated water supplies (raw untreated and treated drinking water). Likelihood of contamination of raw water supplies is HIGH but effective mitigation is in place (treatment and operational procedures) to ensure that risk of contamination is minimized in public drinking water supplies. Therefore, the likelihood of consumption of contaminated, treated drinking water is VERY LOW to LOW (VERY LOW UNCERTAINTY). The likelihood of consuming contaminated raw water is LOW (VERY LOW UNCERTAINTY). Sources of contamination are most likely to be livestock, particularly young animals, such as lambs and calves. Although aquatic animals play a role as a reservoir of Cryptosporidium and Giardia spp., it is not clear whether they are primary or secondary host species that are being infected through contact via human contamination routes. Beavers, as one type of wildlife species which is also susceptible to Cryptosporidium and Giardia spp. infection will also make a contribution to the risk of contamination. However, relative to other sources, this is likely to be minor (although uncertainty is high). The significance of the contribution will depend on the likelihood of infection and the proximity of beavers to important water sources for public water supplies.
3. LEGISLATION, DEFINITIONS AND ASSUMPTIONS European beavers have been extinct in Scotland for around 400 years. In 2008, Scottish Government granted a licence for a scientifically monitored trial reintroduction of European beaver (Castor fiber) into Knapdale, Scotland for a five year period. There is the potential to extend this reintroduction to establish a population of beavers in Scotland. However, there are questions over the extent to which beavers pose a threat to the quality and safety of water for human consumption. The purpose of this risk assessment is to inform any future decision over the public health risks of water-borne parasites (such as Giardia spp. and Cryptosporidium spp.) posed by further beaver reintroductions into Scotland.
4. HAZARD IDENTIFICATION Both Giardia spp. and Cryptosporidium spp. are parasitic protozoa that can cause infection to humans through contaminated drinking water. Cryptosporidium Hazard: Disease in humans is predominantly caused by the species Cryptosporidium hominis and Cryptosporidium parvum. C. parvum and C. hominis are under established surveillance in the United Kingdom, and so data on Cryptosporidium infections in the European Surveillance System represents the total Cryptosporidium infections reported in the United Kingdom. Health Protection Scotland has received approximately 450-700 reports of laboratory confirmed cryptosporidiosis cases every year since 2005 (Pollock, Alexander, Robertson-Kellie and Anderson, 2015). Transmission Oocysts are released in the faeces and are immediately infectious. There are numerous direct and indirect (through fomites on food or in water) transmission routes. Individuals become infected through ingesting food, soil, or water contaminated with oocysts or by person-to-person, or animal-person, transmission. Animal reservoirs include livestock such as cattle and sheep, domestic pets (including cats and dogs) and wildlife species. C. parvum infects both humans and domesticated animals, particularly cattle, and is epidemiologically associated with zoonotic transmission both via direct contact with infected cattle and indirect transmission through drinking water (Goh et al., 2005; Hunter and Thompson, 2005). C. hominis is mostly associated with human sources. Survival in the environment Cryptosporidium can survive outside the body in severe environmental conditions for several months in moist conditions. It can survive water chlorination but it can be physically removed by filtration or inactivated by UV treatment. Inadequate water filtration or UV irradiation can expose a person to risk to infection (Pollock, Alexander, Robertson-Kellie and Anderson, 2015; Pollock et al. 2008). Detection Detection methods for Cryptosporidium and Giardia are similar and can be performed at the same time. Routine diagnostic methods are based on microscopy (usually of faecal samples) and are not capable of parasite identification to species level. Detection methods for the water industry require very large sample volumes and specialized sampling and analysis to detect the very low numbers of protozoan cysts likely to be present. Samples positive for Cryptosporidium can be typed to species, genotype or higher resolution for epidemiological purposes, although this is not routinely done. Newer immunological and PCR-based detection methods are more sensitive, faster and easier to perform than microscopy and offer the possibility of differentiation of species and genotypes. Detection depends on sufficient cysts being present for microscopy, immunological, or molecular methods (rapid microbiology website). Public Health Consequences Drinking water is a rare but recognized risk factor for human illness (Pollock, Alexander, Robertson-Kellie and Anderson, 2015; Pollock et al. 2008). Symptoms of cryptosporidiosis are primarily profuse, watery diarrhoea, often accompanied by bloating, abdominal pain and nausea or vomiting. Clinical signs appear

after 2-10 days (mean 7). Illness can last 2-3 weeks, but it is normally self-limiting. Individuals who are immunocompromised may develop severe chronic diarrhoea or atypical gastrointestinal infection, which may prove fatal (Hunter and Nichols, 2002). Giardia Hazard: Human infection with Giardia is commonly with Giardia duodenalis (also known as G. lamblia and G. intestinalis). Transmission: As for Cryptosporidium, infectious cysts are released in the faeces. Individuals become infected through ingesting contaminated food, soil, water or by person-to-person transmission. The situation is less clear cut for Giardia, compared to Cryptosporidium, but the evidence does not, in general, support zoonotic transmission as a major risk for human infections (Hunter and Thompson, 2005). Most cases in Scotland are associated with foreign travel (Pollock, Alexander, Robertson-Kellie and Anderson, 2015). Survival in the environment: Giardia cysts can survive for extended periods of time (weeks to months) in the environment. A major reservoir of the parasite is contaminated surface water. Environmental dispersal can lead to the contamination of drinking water and food (Cacciò et al., 2005). Outbreaks in other parts of the world are linked to inadequate treatment of drinking water, and infants and children are at particular risk of infection. Detection: Giardia is likely to be underreported but has the potential to cause large outbreaks (Pollock, Alexander, Robertson-Kellie and Anderson, 2015). Molecular technologies are important in genotying Giardia (and are more sensitive than microscopy). Public health consequences: Infection can be asymptomatic. However, it can also cause acute or chronic disease (including belching, nausea and weight loss). In Scotland, the majority of cases are infected with assemblage A, which is associated with milder infections.
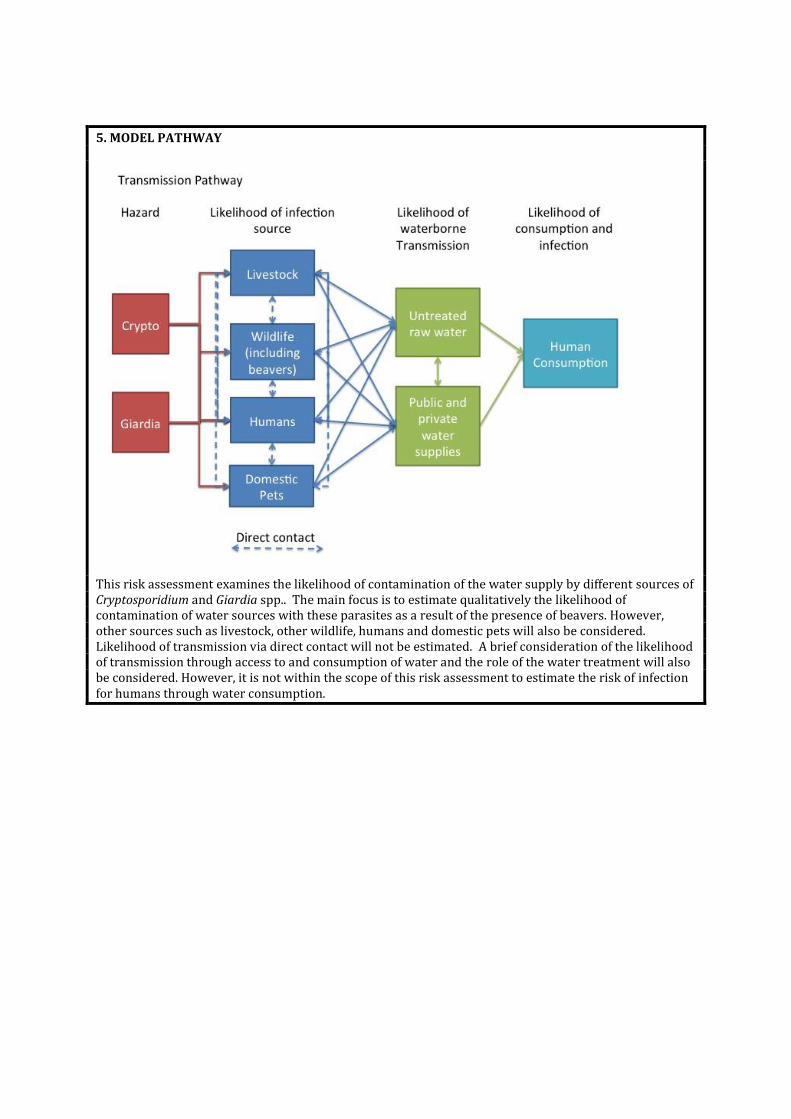
5. MODEL PATHWAY
This risk assessment examines the likelihood of contamination of the water supply by different sources of Cryptosporidium and Giardia spp.. The main focus is to estimate qualitatively the likelihood of contamination of water sources with these parasites as a result of the presence of beavers. However, other sources such as livestock, other wildlife, humans and domestic pets will also be considered. Likelihood of transmission via direct contact will not be estimated. A brief consideration of the likelihood of transmission through access to and consumption of water and the role of the water treatment will also be considered. However, it is not within the scope of this risk assessment to estimate the risk of infection for humans through water consumption.
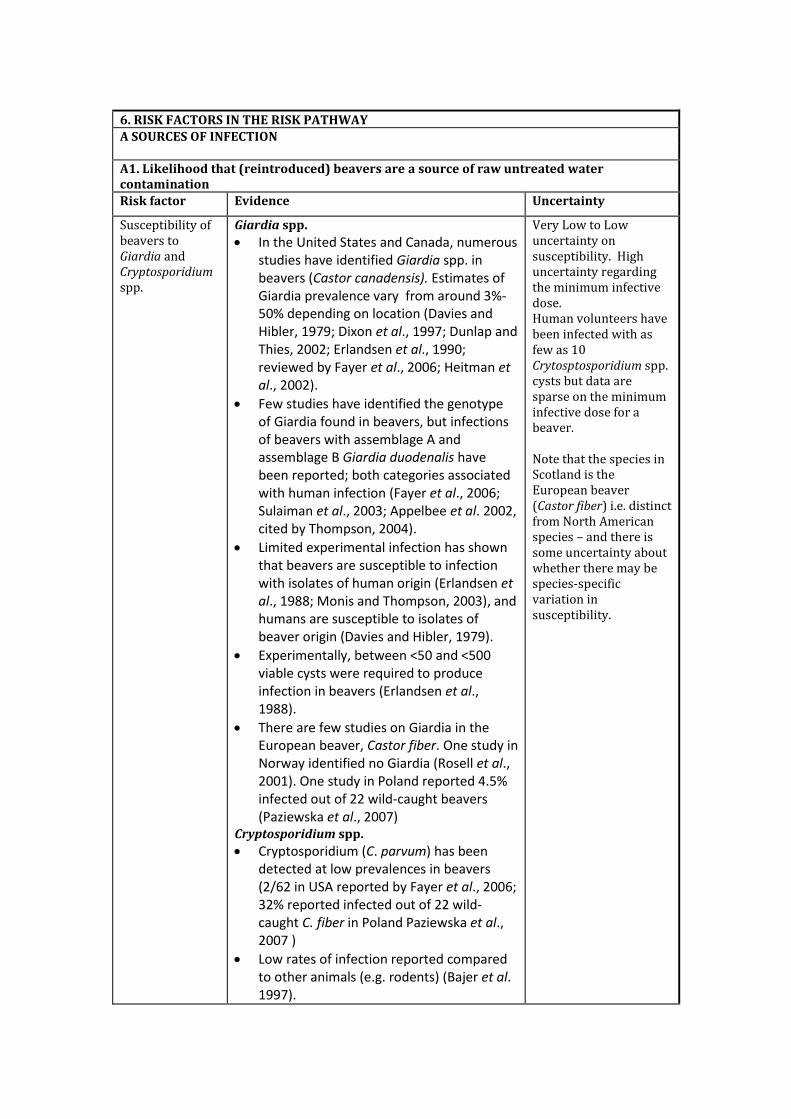
6. RISK FACTORS IN THE RISK PATHWAY
A SOURCES OF INFECTION
A1. Likelihood that (reintroduced) beavers are a source of raw untreated water contamination
Risk factor Evidence Uncertainty
Susceptibility of beavers to Giardia and Cryptosporidium spp.
Giardia spp.
In the United States and Canada, numerous studies have identified Giardia spp. in beavers (Castor canadensis). Estimates of Giardia prevalence vary from around 3%-50% depending on location (Davies and Hibler, 1979; Dixon et al., 1997; Dunlap and Thies, 2002; Erlandsen et al., 1990; reviewed by Fayer et al., 2006; Heitman et al., 2002).
Few studies have identified the genotype of Giardia found in beavers, but infections of beavers with assemblage A and assemblage B Giardia duodenalis have been reported; both categories associated with human infection (Fayer et al., 2006; Sulaiman et al., 2003; Appelbee et al. 2002, cited by Thompson, 2004).
Limited experimental infection has shown that beavers are susceptible to infection with isolates of human origin (Erlandsen et al., 1988; Monis and Thompson, 2003), and humans are susceptible to isolates of beaver origin (Davies and Hibler, 1979).
Experimentally, between <50 and <500 viable cysts were required to produce infection in beavers (Erlandsen et al., 1988).
There are few studies on Giardia in the European beaver, Castor fiber. One study in Norway identified no Giardia (Rosell et al., 2001). One study in Poland reported 4.5% infected out of 22 wild-caught beavers (Paziewska et al., 2007)
Cryptosporidium spp.
Cryptosporidium (C. parvum) has been detected at low prevalences in beavers (2/62 in USA reported by Fayer et al., 2006; 32% reported infected out of 22 wild-caught C. fiber in Poland Paziewska et al., 2007 )
Low rates of infection reported compared to other animals (e.g. rodents) (Bajer et al. 1997).
Very Low to Low uncertainty on susceptibility. High uncertainty regarding the minimum infective dose. Human volunteers have been infected with as few as 10 Crytosptosporidium spp. cysts but data are sparse on the minimum infective dose for a beaver. Note that the species in Scotland is the European beaver (Castor fiber) i.e. distinct from North American species – and there is some uncertainty about whether there may be species-specific variation in susceptibility.
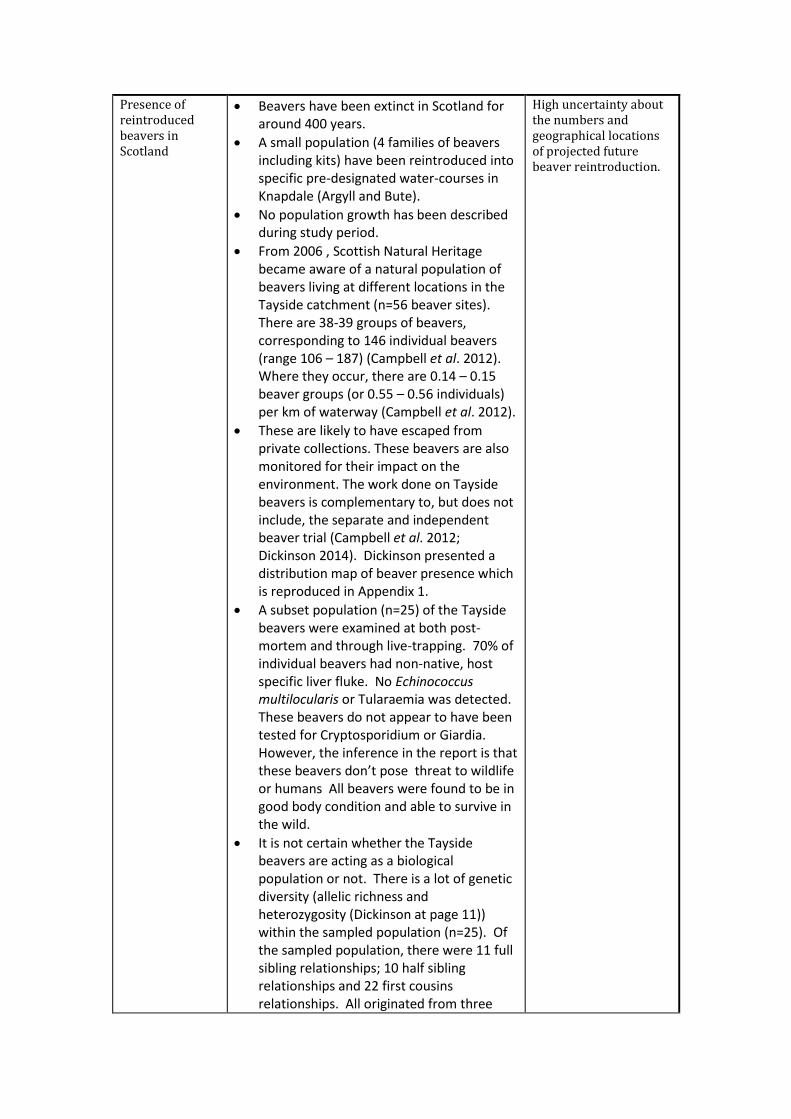
Presence of reintroduced beavers in Scotland
Beavers have been extinct in Scotland for around 400 years.
A small population (4 families of beavers including kits) have been reintroduced into specific pre-designated water-courses in Knapdale (Argyll and Bute).
No population growth has been described during study period.
From 2006 , Scottish Natural Heritage became aware of a natural population of beavers living at different locations in the Tayside catchment (n=56 beaver sites). There are 38-39 groups of beavers, corresponding to 146 individual beavers (range 106 – 187) (Campbell et al. 2012). Where they occur, there are 0.14 – 0.15 beaver groups (or 0.55 – 0.56 individuals) per km of waterway (Campbell et al. 2012).
These are likely to have escaped from private collections. These beavers are also monitored for their impact on the environment. The work done on Tayside beavers is complementary to, but does not include, the separate and independent beaver trial (Campbell et al. 2012; Dickinson 2014). Dickinson presented a distribution map of beaver presence which is reproduced in Appendix 1.
A subset population (n=25) of the Tayside beavers were examined at both post-mortem and through live-trapping. 70% of individual beavers had non-native, host specific liver fluke. No Echinococcus multilocularis or Tularaemia was detected. These beavers do not appear to have been tested for Cryptosporidium or Giardia. However, the inference in the report is that these beavers don’t pose threat to wildlife or humans All beavers were found to be in good body condition and able to survive in the wild.
It is not certain whether the Tayside beavers are acting as a biological population or not. There is a lot of genetic diversity (allelic richness and heterozygosity (Dickinson at page 11)) within the sampled population (n=25). Of the sampled population, there were 11 full sibling relationships; 10 half sibling relationships and 22 first cousins relationships. All originated from three
High uncertainty about the numbers and geographical locations of projected future beaver reintroduction.
distinct lineages of Castor fiber from Germany (Dickinson at page 11).
The Tayside report assesses environmental impact of beaver presence. This is outwith the scope of this risk assessment.
Population structure
Beavers live in extended family units, most commonly composed of a mated adult pair and their offspring from the previous 2 3 breeding seasons (Busher et al., 2009; Crawford, 2007; Novak, 1977; Svendsen, 1980). Svendsen (1989) has reported that beaver pairs remained together for an average of 2.5 years.
Dispersal of juveniles and population structure may be influenced by landscape characteristics such as the connectivity and spatial distribution of aquatic habitats (Crawford, 2007).
Low –Medium uncertainty This information is based on North American research. The assumption is that the population structure of the European beaver is likely to be broadly similar to the North American species.
Effect on the environment
Beavers can shape wetland habitats through dam and lodge construction, and food acquisition. These activities have been shown to significantly modify wetlands; altering stream flow, water chemistry, sediment load, vertebrate and invertebrate species composition and are thought to increase species (Naiman et al., 1988; Nummi and Kuuluvainen, 2013; Wright and Jones, 2002).
Beavers prefer deep pools of water around lodges and will attempt to dam free-flowing water to create these pools (Havens 2006, cited by Crawford, 2007).
Intensive dam repair may facilitate the downstream movement of oocysts. Mud from the bottom of ponds is used to plaster the upstream face and top of the dam – and if this contains broken faecal pellets, Giardia cysts may wash over the top of the dam (Monzingo and Hibler, 1987).
Dam building and the impact of beavers in the environment may also affect water turbidity which may subsequently impact upon the effectiveness of filtration mechanisms for oocyst removal (pers. comm. B. Wells). The magnitude of the impact would be likely to depend on the proximity of beavers to intake mechanisms for human drinking water supplies, population size and extent of beaver activity.
Low- Medium uncertainty The assumption is that behaviour of the European beaver is likely to be broadly similar to the North American species.

In Tayside, 3 beaver groups had built dams. In these groups, seven dams were maintained. The dam-building activities of two of these groups were leading to localized issues, necessitating frequent dam-removal by the landowners (Campbell et al. 2012).
The Tayside beaver report indicates that dam-building may have been responsible for erosion of the banks and changes in water flows. In addition, 9 sites (out of 56) reported issues with associated flooding from impeded drainage. Flooding may increase the risk of exposure to parasites if these are already present in the water supply. The use of drainage pipes to mitigate this effect appears to have been successful in at least one instance where the pipe was correctly installed.
Age Some studies have found a greater prevalence of Giardia spp. in juvenile (subadults or kits) beavers than adults (Erlandsen et al., 1990; Fayer et al., 2006; Frost et al., 1980). It is unknown whether adults become free of infection or develop latent infection.
Kits appear to become infected prior to, or just after, leaving the winter lodge. This may suggest beaver-to-beaver transmission or water-borne transmission (Monzingo and Hibler, 1987).
Female beavers ingest kits’ excrement for 3 days after birth (Novak 1998, cited by Dunlap and Thies, 2002). This may be a source of reinfection.
Other studies found no differences in prevalence with age (Dunlap and Thies, 2002).
Low- Medium uncertainty Age determination may be a source of uncertainty. Some ageing is done by weight. Small numbers of beavers have been sampled in these studies.
Habitat and behaviour
Susceptibility may be attributable to the semi-aquatic habitat, burrowing behavior and coprophagy (Dunlap and Thies, 2002)
Season, habitat and stress may play a role in beaver infection. This may result in varying rates of infection depending on access to food, changes in diet, parturition.
Beavers exhibit coprophagy (Beuch, 1984), which may be an important feature of Giardia epidemiology. Beavers ingest their own faeces directly from the rectum and thus can be continually re-exposed to infection (Dunlap and Thies, 2002;
Medium uncertainty Studies sampling beavers have varied according to location (almost all have taken place in the United States) and numbers of beavers sampled. The largest studies have sampled fewer than 200 beavers. The assumption is that behaviour of the European beaver is likely to be broadly
Monzingo and Hibler, 1987).
Parturition and post-parturition behaviour may influence frequency of coprophagy.
similar to the North American species.
Infection status of reintroduced beavers in Knapdale
1. Prevalence of disease in country of origin of reintroduced beavers
All beavers arriving from Norway were quarantined in accordance with statutory rabies precautions. All animals were extensively screened for health and for zoonoses and rodent pathogens (Goodman et al., 2012).
Rosell (Rosell et al., 2001) in Telemark County, Norway, examined 241 beavers for Giardia, detecting no cysts in any sample. In the same study, the faeces of 182 beavers (103 live- trapped and 79 shot) were examined using in-vitro diagnosis and found no Cryptosporidium oocysts in any sample.
There is evidence of Giardia and Cryptosporidium in Norway at low prevalences in the human population (Giardia (179 human cases/16368 confirmed cases in the EU/EEA) and Cryptosporidium (4/9581 confirmed human case in the EU/EEA)) (ECDC).
Disease status of animals prior to release
All animals tested negative for Cryptosporidium and Giardia, as well as Salmonella and Campylobacter (Goodman et al., 2012).
Disease status of animals during and after reintroduction:
Beavers were trapped and subjected to veterinary examination throughout the study period. Tests included screening blood and faecal samples for Cryptosporidium and Giardia. All tests were negative, with the exception of a decomposed beaver kit recovered from Loch Linne, which tested positive for Cryptosporidium (Mackie, 2014 at 3.1.2 at p6)
High uncertainty No data to answer any questions about the effectiveness of quarantine or screening. Speciation of Cryptosporidium found in samples from beavers has not been adequately performed, so it remains in doubt whether they carry species of the organism capable of zoonotic infection.
Subsequent infection of naïve beavers (reintroduced or progeny) through exposure to Cryptosporidium and Giardia already in
A study of the microbiological quality of watercourses and private water supplies in the study area between 2001 and 2003 identified Cryptosporidium and Giardia prior to beaver re-introduction (Morrison, 2004). Escherichia coli and faecal streptococci were also detected.
This suggests the presence of wild and
High uncertainty No control samples were taken from catchments where beavers were not present. Beavers were not sampled for
environment domestic animals (e.g. deer, foxes, sheep, etc.) in the immediate area of the lochs or burns. These burns are supplied from different catchment areas (Mackie, 2014).
No Giardia, Salmonella, Cryptosporidium or faecal streptococci were isolated from drinking water supplies. However, there was evidence of faecal contamination of water supplies (presence of E. coli- and coliform bacteria). This is not uncommon for untreated supplies.
It is possible that uninfected beavers may become infected with Cryptosporidium or Giardia spp. already in the water courses in Scotland.
Cryptosporidium or Giardia after the reintroduction period so there is no evidence as to whether infection took place during the trial. The study was inconclusive as to whether beavers represented an additional factor in the presence or absence of Cryptosporidium or Giardia in any particular water system in the study area over the period of the trial
Likelihood of amplification and shedding of Cryptosporidium and Giardia in reintroduced beavers
There is some speculation that beavers may serve as an amplification host for Giardia but this evidence is limited (Monzingo and Hibler 1987; Thompson 2004).
High uncertainty over amplification of numbers of parasites by beavers or other wildlife.
Detection of infection in reintroduced beavers
Adequate sampling of faeces and water samples must be undertaken in order to assess this appropriately.
Beavers don’t appear to be physically compromised by Giardia infection. However, the epidemiology of infection is reasonably unknown.
Microscopy alone will not be sufficient to determine whether Giardia or Cryptosporidium spp. originated from beavers or from the environment.
Medium uncertainty Many historical surveys for Giardia spp. use detection methods that rely on detection of cysts rather thanactive trophozoites (e.g. Davies and Hibler 1979; Frost et al. 1980, Beuch 1984; Monzingo and Hibler 1987). This may result in large numbers of false negatives (Sensitivity 76% - Davies and Hibler 1979)
Environmental contamination
See Hazard Identification (section 4)
Depends on parasite species, precipitation, temperature, season, habitat and water flow.
One study in the USA identified the greatest number of Giardia-infected beaver from marshes in the spring and summer, ponds during autumn and winter, and creeks during summer and autumn (Dunlap and Thies, 2002). The authors speculated that this could be due to the suspension behaviour of cysts in the water. In dry hot conditions when water flow is slow, cysts
High uncertainty No substantive evidence of environmental contamination in a Scottish or international context. Kirner et al (1978) did not describe the examination of other mammals likely to be present in the watersheds. The Scottish study conducted in

concentrate in shallower muddier waters; as rainfall increases, water flow and speed increases and cysts are flushed downstream into ponds. If these settle, they will concentrate as the pond freezes in winter.
If beavers are active in reservoirs, the risk of contamination of the water supply is likely to increase with increased proximity to intakes for human consumption.
In winter, if temperatures are consistently below freezing, beavers are likely to be confined to their lodge pond. There are no data about the prevalence of Giardia in beavers at this time and streams are not likely to be flowing to disperse the cysts. The extreme conditions necessary for this to occur are a very unlikely occurrence in Scotland.
The source of Giardia infection in beavers in a number of studies was of human origin (Fayer et al., 2006; Sulaiman et al., 2003; Appelbee et al. 2002, cited by Thompson, 2004).
There is limited evidence of beavers as the primary source of contamination for water sources. It is thought more likely that they become infected with contamination from human or domestic animal origin, potentially acting as an amplifier (Thompson, 2004). One study that traced an outbreak to a single beaver which was infected with Giardia failed to look for infection in other wildlife sources (Kirner et al., 1978).
In Scotland, the beaver trial did not produce evidence of further contamination of water supplies from beavers. Giardia and Cryptosporidium were detected in watercourses, but at rates consistent with similar samples taken prior to the introduction of beavers. There was no association between the presence or absence of Giardia and Cryptosporidium in watercourses and beaver populations in the same catchments. In the catchment with the greatest number of beavers, no Cryptosporidium or Giardia oocysts were detected during the study period, even though these had been detected prior to beaver release.
conjunction with the release of beavers was inconclusive as to whether beavers present an additional risk for the presence of Cryptosporidium or Giardia. No control samples were taken from catchments in which beavers were not present; relying instead on the pre-release study for control data. Characterization of Giardia DNA of animal origin is urgently required, particularly from aquatic animals, given the abundance of water-borne cysts and the scarcity of water-borne outbreaks (Cacciò et al. 2005).
A1. Likelihood that beavers are a source of raw untreated water contamination: The likelihood of beavers as a source of contamination is VERY LOW TO LOW (HIGH UNCERTAINTY) but contingent on the source country of any reintroduced beavers, the presence of infection and adequacy of testing to detect infection and excretion and speciation of oocysts prior to reintroduction. Once beavers are reintroduced to Scottish waters, there is LOW – MEDIUM likelihood (HIGH UNCERTAINTY) that beavers will become infected with these parasites already in Scottish waters. There is strong evidence that beavers are hosts for these parasites. However, the evidence for amplification is HIGHLY UNCERTAIN as few studies have investigated this.
A2. Likelihood that other wildlife are a source of raw untreated water contamination
Risk factor Evidence Uncertainty
Presence of wildlife
European otter (Lutra lutra) and American mink (Neovison vison) have been recorded in the beaver trial area during the study period (Morrison, 2004).
Deer, fox, red squirrel, pine marten and a range of small mammals such as rodents and shrews are also commonly present in Argyll’s forests (Mackie, 2014).
Low uncertainty Wildlife detected depends on sampling strategy.
Susceptibility of wildlife to Giardia and Cryptosporidium spp.
Giardia spp.
Animals such as nutria, deer, rodents and birds are also commonly infected with Giardia in North America (Dunlap and Thies, 2002; Heitman et al., 2002; Monzingo and Hibler, 1987)
Specific information on what genotype of Giardia wildlife species carry is limited. Recent studies have, however, confirmed that white-tailed deer in the wild can harbour infections with zoonotic genotypes of G. duodenalis (assemblage A) (Trout et al., 2003).
Although wildlife are susceptible to infection with zoonotic genotypes of G. duodenalis, the limited evidence collected under natural, pristine conditions suggests that wildlife harbour their own genotypes/species of Giardia (Thompson, 2004).
Cryptosporidium spp.
Cryptosporidium has been reported to infect over 150 species of animals (Xiao et al., 2004) and over 60 genotypes (Chalmers et al. 2013), (some with a limited host range), C. parvum, a zoonotic pathogen, is the species reported most often and with the widest range of hosts (Fayer et al., 2006).
Small wild mammals shed large numbers of oocysts per gramme faeces (Sturdee et al., 2003).
Medium-high uncertainty over current prevalence. Low uncertainty over susceptibility. No wildlife species in Scotland were concurrently sampled for Cryptosporidium or Giardia spp. during the beaver reintroduction trial.
Age Age may play a role in the susceptibility of wildlife to parasite infection.
High uncertainty as there is little evidence here..
Habitat and behaviour
Any wildlife that rely on aquatic or semi-aquatic environments for their habitat will be at risk of infection or contaminating the aquatic environment with parasites.
Burrowing or copraphagic behaviour may increase this risk (see A1).
Season, habitat and stress may play a role in infection rates and may depend on access to food, changes in diet and/or parturition.
Medium uncertainty Further evidence about impact on the epidemiology of these parasites is required.
Environmental contamination
See Hazard Identification (section 4)
Depends on parasite species, precipitation, temperature, season, habitat and water flow.
The presence of Cryptosporidium oocysts (detected in a pre-beaver-release study) in surface water burns was thought to signify the presence of wild and domestic animals (e.g. deer, foxes, sheep, etc.) in the immediate area of the lochs or burns. These burns are supplied from different catchment areas (Mackie, 2014, p. 7).
The suspension of (oo)cysts in water will play a role in the degree of contamination.
The significance of non-human hosts as sources of water contamination with Giardia has remained an unresolved issue, as has the role of zoonotic transmission in the epidemiology of human Giardia infection (Cacciò et al., 2005; Thompson, 2004).
There is little evidence that aquatic mammals have been the originating contaminating source in water-borne outbreaks. Rather, these animals are more likely to have become infected from water contaminated with faecal matter of human origin (Thompson, 2004).
High uncertainty due to sparse empirical evidence.
A2. Likelihood that wildlife is a source of raw untreated water contamination is MEDIUM to HIGH with HIGH UNCERTAINTY. Infection status is inferred because of the known susceptibility of certain wildlife species in the area, and the presence of parasites in the water supply. However, no sampling plan is in place to determine prevalence of infection in wildlife. A3. Likelihood that livestock are a source of raw untreated water contamination
Risk factor Evidence Uncertainty
Presence, number and density of livestock
A high density of farm animals (especially cattle), will increase the likelihood of infection with Cryptosporidium and Giardia spp. and therefore increase the potential for high excretion rates of oocysts and contamination of the environment.
In addition, farms may provide a habitat for wild mammals that are sources of oocysts (Sturdee et al. 1999; Chilvers et al. 1998) and may act as reservoirs and vectors of infection (Sturdee et al. 2003).
Very low uncertainty Information is available on the presence and location of farm livestock in Britain based on movement records, agricultural census and other data sources.

Susceptibility of livestock to Giardia and Cryptosporidium spp.
Giardia spp.
Giardia infections are widely reported in North America and Europe in dairy and beef cattle often at high farm and herd prevalences (for example Dixon et al., 2011; Gillhuber et al., 2013), and particularly in calves . Cysts appear in faeces of calves at around 4 weeks of age, and peak between 4 and 12 weeks, at around 105 to 106 cysts per gramme of faeces (reviewed by Thompson, 2004).
Sheep and goats are also reported to be infected but generally at lower prevalences than cattle (Geurden et al., 2008; Ryan et al., 2005; Santín et al., 2007). There is little information on the prevalence of Giardia in pigs, but one study reports high prevalence in weaners (Maddox-Hyttel et al., 2006).
The importance of livestock in carrying the zoonotic forms of Giardia duodenalis is unclear. The zoonotic genotypes A and B have been found in cattle, sheep, goats and pigs, but are less common than host adapted genotype E in several studies (Budu-Amoako et al., 2012; O’Handley et al., 2000; Sprong et al., 2009; Van Keulen et al., 2002) including in UK (Minetti et al., 2013).
Cryptosporidium spp.
Farm livestock, and cattle in particular, have been implicated as major sources of oocysts contaminating surface waters (Wells et al. 2015; Brook et al. 2008; Brook et al. 2009; Sturdee et al., 2003).
Surveys in the UK and abroad have identified farms positive for Cryptosporidium (Sturdee et al. 2003; Lindsay et al., 2000; Wade et al., 2000; Enemark et al., 2002)
In a longitudinal UK (England) study, Cryptosporidium was endemic in livestock and small wild mammals (Sturdee et al., 2003).
Seasonally combined data for adult livestock, young livestock and small wild mammals showed all three categories tended to have the greatest Cryptosporidium prevalences in the autumn (Smith et al. 2005; Sturdee et al., 2003).
Prevalence of Cryptosporidium vary according to species. In one study, bull beef, 3.6%; dairy cows, 3.5%; ewes, 6.4%; horses, 8.9%; calves (home bred), 52%; calves (bought-in) 23.2%; lambs, 12.9%; small wild mammals (rodents)
Low uncertainty about the role of livestock in transmission pathway for Cryptosporidium and Giardia parasites. The Cryptosporidium data from this study come from Warwickshire England from a well-maintained estate with good biosecurity and animal health. These data may represent, the baseline, the lowest possible levels to be expected, for Cryptosporidium infection and oocyst production on a lowland farm in the United Kingdom. Cryptosporidium data from animal disease surveillance are passive, and thus based on convenience sampling. There may be some bias according to submissions to different surveillance centres. Multiple reports may come from a single animal but this cannot be determined from the data. At this stage, no data are available on Giardia.

living in and around farm buildings, 32.8%; small wild mammals (mainly rodents) living in areas of pasture, 29.9% (Sturdee et al., 2003).
Animals from populations with the greatest prevalences of Cryptosporidium also tend to shed the greatest numbers of oocysts (ranging
from 1.4 × 103 for bull beef to 1.1 × 105 for calves).
Under experimental conditions, calves shed very high numbers of Cryptosporidium oocysts (Blewett, 1989; Henriksen, 1989; Current, 1985; Anderson, 1981).
For Cryptosporidium, peak shedding occurs in cattle at 14 days (reviewed by Thompson et al., 2005).
Viability of oocysts once outside the animal may vary (Bukhari and Smith 1997; Sturdee et al. 2003).
Scottish surveillance data on Cryptosporidium are collected by SAC consulting surveillance centres. These represent numerator data (number of reports by animal species and type of sample – such as carcase, blood or faeces). These are passively submitted samples (and may be biased towards different centres) and are not speciated.
Between 2010-2014, 1449 reports in which Cryptosporidium spp. was identified, approximately 94% (1360 reports) were from bovine, and 6% from ovine samples. There was 1 avian case, 1 case in a deer and 1 in a goat). Note that it is not apparent from the surveillance data whether every report is from a unique animal.
The following table describes the distribution of Cryptosporidium reports in bovine animals by health authority in Scotland.
Health Authority Number %
AA 157 11.54
BR 54 3.97
DG 378 27.79
FF 32 2.35
FV 42 3.09
GC 29 2.13
GR 224 16.47
HG 186 13.68
LN 57 4.19
LO 61 4.49
OR 29 2.13
SH 6 0.44
TY 101 7.43
WI 4 0.29
Grand Total 1360 100
Environmental
contamination See Hazard Identification (section 4)
Depends on parasite species, precipitation, temperature, season, habitat and water flow.
The viability of oocysts may affect environmental impact (Sturdee et al., 2003).
In a UK study, a stream draining the area surrounding a farm (in Warwickshire) was positive for oocysts on the majority of occasions tested during the course of an 18-month study (Bodley-Tickell et al., 2002) indicating widespread contamination of the local environment
Early reports of human cryptosporidiosis drew attention to zoonotic transmission (contact with infected young cattle or sheep and consumption of livestock- contaminated drinking water) (Current et al. 1983; Department of the Environment 1995 cited by Cacciò et al. 2005)
There was a decline (81%) in the reported incidence of human C. parvum cases in the UK, coinciding with a foot and mouth disease outbreak. During the spring of 2001, strict restrictions on access to the countryside were imposed, which reduced human contact with livestock, feral animals and their faeces (Hunter et al., 2003).
Concurrent decrease in giardiasis was not detected during this period, suggesting that this parasite has significantly different reservoirs of infection and/or routes of transmission, at least in the UK (Smerdon et al. 2003 cited by Cacciò et al. 2005).
In one Scottish study of sporadic human cryptosporidiosis, C. parvum was the aetiological agent in 84% of 67 cases, supporting livestock faecal pollution of water sources as the leading cause of sporadic cryptosporidiosis (Goh et al., 2004).
There is anthroponotic transmission of C. parvum. Some subtypes of C. parvum (GP60 IIc) have never been identified in any other host than humans, so finding C. parvum in a human does not imply zoonotic transmission, although it is possible.
Approximately half the samples that are speciated in Scotland are 50:50 parvum:
Low uncertainty for
Cryptosporidium spp.
Medium/high
uncertainty for
Giardia spp. – the
role of livestock in
zoonotic waterborne
transmission is
unknown.
Although this
evidence comes from
a study from a
different area of the
UK, it may still be
informative; the
farm high standards
of animal husbandry
and land
management.
Despite this,
Cryptosporidium
infection is endemic
and wild mammal
populations were
persistently infected
acting as reservoirs
of the parasite.
hominis. The only samples likely to be submitted for speciation are those deemed to be part of an outbreak. Samples from what is likely to be a sporadic case are unlikely be speciated (pers. comm. Hotchkiss).
The observed spring peak in human C. parvum infection is thought to be associated with an increase in contact with livestock faecal material during the calving/lambing season (Pollock, Alexander, Robertson-Kellie and Anderson, 2015).
Human isolates of C. parvum often have the same subtypes as animal sources supporting the hypothesis that ruminants are a source of infection (Deshpande et al. 2015; Pollock, Alexander, Robertson-Kellie and Anderson, 2015).
Although cattle have been repeatedly implicated as sources of water-borne cryptosporidiosis outbreaks, genotyping the contaminating isolate(s) has often implicated human effluent as the source, as with the notorious Milwaukee outbreak (Zhou et al. 2003, cited by Cacciò et al. 2005)
The role of livestock in contributing to human Giardia infection is less clear. Given the high rate of production of oocysts in calves, it is thought that even a few calves infected could present a significant contribution to water contamination (Thompson 2004)
The clinical presentation of Giardia infection in ruminants is inconsistent; it is often subclinical, but may be associated with diarrhoea and ill thrift in calves (reviewed by Thompson, 2004).
A3.Likelihood that livestock is a source of contamination of raw untreated water supplies
is HIGH, LOW-MEDIUM UNCERTAINTY. There is strong evidence to support the view that
infection with Giardia and Cryptosporidium spp. may be endemic on some farms; this increases
the likelihood that livestock will become a source of environmental and water contamination.
A4. Likelihood that domestic pets are a source of water contamination
Risk factor Evidence Uncertainty
Presence High frequency of domestic pets in Scotland – particularly cats and dogs in urban and rural settings.
Medium to high
uncertainty over
domestic pet
distribution.
Need specific
evidence to
substantiate if
numbers are
required.
Susceptibility of
domestic pets to
Giardia and
Cryptosporidium
spp.
Giardia spp.
G. duodenalis has been identified as the most common enteric parasite of domestic dogs and cats in Australia, and is common in pets in other countries (reviewed by Thompson, 2004)
Domestic dogs are susceptible to infection with both host-adapted (G. duodenalis assemblage C or G. canis) and zoonotic Giardia genotypes (Cacciò et al., 2005), with assemblage A and B identified in dogs and cats (Van Keulen et al., 2002). One study in Germany found 60% of Giardia identified in dogs to be assemblage A (Leonhard et al., 2007).
Although Giardia is common in dogs and cats, it is rarely associated with clinical disease in these animals. Parasites may be present at subclinical concentrations and cyst excretion may be intermittent or cyclical (Leib and Zajec, 1999; McGlade et al., 2003).
Treatment of Giardia- infected dogs and cats is usually recommended whether or not they are clinically ill, because of the perceived potential for zoonotic transmission (Thompson, 2004).
The recent development of vaccines for the treatment and prevention of Giardia infections in dogs and cats, and their apparent ability to reduce the duration of shedding of cysts may provide an alternative to drugs for reducing carrier rates in pets and subsequent environmental contamination (Thompson, 2004).
Dogs from multi-dog households were more commonly infected with Giardia than dogs in single-dog households, emphasizing the potential ease with which Giardia can be spread to in-contact animals and therefore presumably to humans (Bugg et al., 1999).
Cryptosporidium spp.
Although dogs and cats often carry Cryptosporidium, they are most commonly infected with the host adapted strains C. canis and C. felis (Abe et al.,
High uncertainty
This requires further
molecular
characterization of
oocysts from
infected animals in
endemic areas
before the
assumption that
domestic animals
don’t contribute to
human cases can be
verified (Cacciò et
al. 2005).
The previous
uncertainty may not
be relevant to the
question posed by
the risk assessment
that is concerned
with contamination
of water supplies.
2003; reviewed by Monis and Thompson, 2003).
Age Data required High uncertainty.
Requires data.
Habitat and
behaviour Close direct contact between people and
domestic pets is likely to be the primary source of transmission.
If clinical giardiasis in domestic pets is reported, it is usually associated with kennel or cattery situations, where the effects of over-crowding may cause stress and exacerbate the effects of an infection (Robertson et al., 2000).
High uncertainty.
Requires data.
Environmental
contamination Rural domestic pets on farms may contribute
to the burden of environmental contamination posed by livestock although this has not been quantified and is likely to be a relatively minor contribution.
Recent molecular epidemiological studies demonstrate zoonotic transmission in an endemic focus in situations in which humans and dogs live closely together (Traub et al., 2004).
Dogs and cats are susceptible to infection with zoonotic genotypes of Giardia but the chances of a contamination event from a dog or cat leading to a water-borne outbreak in humans would seem unlikely (Thompson, 2004).
There have been few reports of human cases of Cryptosporidium or Giardia linked to pet ownership.
In 1998, in Sydney, Australia, dead dogs found near a water supply, were incriminated as a possible source of contamination with Giardia, but there was no evidence of infection in the dogs (Thompson 2004)
High uncertainty due
to lack of data
A4. Likelihood that domestic pets are a source of contamination of raw untreated water
supply is NEGLIGIBLE. There is MEDIUM-HIGH UNCERTAINTY about the role of domestic pets
as a source of outbreaks through direct or indirect transmission.
A5. Likelihood that humans are a source of raw untreated water contamination
Risk factor Evidence Uncertainty
Susceptibility of
humans to
Giardia and
Cryptosporidium
Most common infections in humans are by Cryptosporidium parvum and C. hominis.
Giardia duodenalis genotypes A and B have so far been associated with human
Low uncertainty
about role of
humans in onward
spp. infections.
The infective dose is low (1-10 oocysts) for both parasites (Cacciò et al., 2005).
Oocysts are immediately infectious when excreted in faeces, and can be transmitted by person-to-person contact (Cacciò et al., 2005).
Giardia spp.
Swallowing water while swimming, drinking treated tap water, contact with recreational raw water and eating lettuce have all been identified as risk factors for human infection (Stuart et al., 2003).
Giardia cysts probably survive less well in raw water than do C. parvum oocysts, although cysts are usually detected more frequently and with greater abundance in raw and drinking water. The association with eating lettuce (and other fruit and vegetables which receive minimal heat treatment before consumption) also highlights the role of contaminated wastewater, uncomposted sewage sludge or manure used as fertilizer, in addition to direct contamination of produce by wildlife, or transmission vectors such as refuse-feeding birds and non biting filth flies (Stuart et al., 2003).
In 2012, the UK reported 4,138 cases of Giardia out of a possible 16, 369 confirmed cases in the EU (25%) (ECDC Giardia data), incidence rate 5.39 per 100 000 cases, Age Standardized Rate (ASR) 5.67
Cryptosporidium spp.
C. hominis and C. parvum have been identified as the major causes of human cryptosporidiosis but their prevalences vary in different regions of the world (Cacciò et al., 2005).
C. parvum causes more human infections in Europe (57%, on average), especially in the UK.
The infective dose of Cryptosporidium is very small and about two to ten oocysts are estimated to be sufficient to cause the disease.
Humans may have symptomatic and asymptomatic infection.
In 2012, the UK reported 6,533 cases (68% of all cases reported in the EU) (ECDC
transmission.
Molecular assays
must be applicable
to clinical (human
and non-human
hosts) and
environmental
(including food)
samples, particularly
for species and
genotype identity
and source, and
disease tracking.
Specific
requirements for
developing effective
molecular
identification and
genotyping systems
in environmental
samples include
increased
discrimination,
specificity and
sensitivity, which
can be exploited to
improve
understanding of the
epidemiology of
infection, disease
and outbreak
investigations.
Spatial data on
human infections
will be recorded
according to the
residential location
of the individual
rather than
necessarily
reflecting where
infection occurs
(Pollock, Alexander,
Robertson-Kellie and
Cryptosporidium data). All cases were confirmed positive. Incidence rate 10.37 per 100 000 cases, Age Standardized Rate (ASR) of 9.97.
The provisional incidence rate for Cryptosporidium in Scotland for 2013 is 8.07 per 100 000 population ( Health Protection Scotland, data as at 20 June 2014)
Anderson, 2015).
Thus, any infections
acquired by people
living in urban areas
during access to the
countryside may not
be accurately
located.
Age Children and young adults are more likely than older people to become infected and act as infection sources (Rose et al. 1991; Robertson et al. 2002; Roy et al. 2004; Hunter et al. 2004; Cacciò et al. 2005)
Low uncertainty
Habitat and
behaviour High population densities increase
transmission rates.
Rural versus urban populations increase likelihood of infection due to close contact with farm animals, roles in intensive livestock husbandry, birthing and feeding of susceptible calves and lambs (Learmonth et al. 2004, Xiao et al. 2004, cited by Cacciò et al. 2005). This is especially true for C. parvum. In Scotland, relatively large numbers of C. parvum isolates are observed in Grampian, Fife and Ayrshire/Arran compared to urban areas (Glasgow and Lothian) (Pollock, Alexander, Robertson-Kellie and Anderson, 2015; see also Pollock et al. 2010).
Close contact (e.g. during recreational bathing or consumption of contaminated water) (Cacciò et al., 2005). Swimming in raw water or public swimming pools were positively associated with infection in Australian and USA studies (Robertson et al 2002; Roy et al 2004).
Travelling abroad and contact with a diarrhoeic individuals (Hunter et al. 2004)
Season: In UK, a bimodal seasonal pattern of disease has been described: one peak during the spring and the second during late summer to early autumn (McLauchlin et al. 2000). The spring peak is almost exclusively due to C. parvum, whereas both C. parvum and C. hominis occur in the late summer to early autumn peak (Cacciò et al. 2005).
Low uncertainty
Likelihood of
environmental The control of human waste is important in
protecting aquatic wildlife from infection with
Very low uncertainty
contamination Giardia (Thompson 2004) and Cryptosporidium spp.
Suk et al. (27) reported that surface water in recreational areas with high human usage had significantly greater rates of Giardia cysts in the water (44.9%) than did water from areas with low recreational use (17.2%).
“The media-popularized term "beaver fever," used in regard to the role of these animals as a source of cysts in water-borne giardiasis, may actually be a misnomer, since human usage of the watersheds may have led to the contamination of the drinking water” (Erlandsen et al. 1988; 1990)
The source of Giardia infection in beavers is likely to be of human origin (Dixon et al., 2002; Monzingo and Hibler, 1987; Rickard et al., 1999).
Evidence that greater numbers of C. hominis than C. parvum oocysts are excreted by infected humans (McLauchlin et al. 1999; Xiao et al. 2001 cited by Cacciò et al. 2005). This may prove important when estimating the impact of human sewage as a means by which oocysts are transmitted into the environment.
Water is likely to become contaminated if recreational access is possible or if water is contaminated with sewage.
A5. Likelihood that humans are a source of contamination of raw untreated water supplies
is MEDIUM to HIGH. Uncertainty is LOW as there is evidence that control of sewage is an
important component of reducing exposure to water-borne parasites.
B. TRANSMISION PATHWAYS
B1. Likelihood of consumption of contaminated raw untreated water supplies and treated
drinking water
Risk factor Evidence Uncertainty
Level of existing raw
untreated water
pollution
Water pollution will be affected by location, water speed and sources of pollution such as sewage outlets.
Water speed has an impact on water pollution. Cysts of Giardia spp. settle rapidly in slow-moving waters (Monzingo and Hibler, 1987).
Cryptosporidiosis has a bi-modal seasonality. C. parvum predominates April to June; C. hominis prevails between August and November (Pollock et al. 2010; Deshpande et al. 2015 cited by Pollock,
Medium uncertainty
Data are available on
the geographical
location of public
water reservoirs,
private water
sources and sewage
outlets and
treatment plants
(Scottish Water and
Alexander, Robertson-Kellie and Anderson, 2015). This spring peak appears to be associated with contamination by animal faecal material during the calving/lambing season (Pollock, Alexander, Robertson-Kellie and Anderson, 2015).
Cryptosporidium oocysts can be removed or inactivated by some water treatment processes. Inadequate filtration can expose persons to risk of infection (Pollock et al. 2008).
Low concentrations of Cryptosporidium oocysts have already been detected in 65-97% of surface water supplies supplying raw water to treatment works in Scotland (Pollock, Alexander, Robertson-Kellie and Anderson, 2015). This is mitigated by effective water treatment of public drinking water supplies.
DWQR).
Access to
contaminated raw
untreated water
Likelihood of access to the countryside. For example, the forestry area in North Knapdale (where beavers have been reintroduced) is popular with fishermen, walkers, cyclists and horse-riders, with public access promoted by Forestry Commission Scotland. There is limited recreational use of freshwater lochs, mainly restricted to the use of boats for angling, which is controlled through local clubs (Mackie, 2014)
There is a high incidence of giardiasis among outdoor recreationalists in the USA. However, the evidence for an association between drinking backcountry water and acquiring giardiasis was minimal. Other potential risk behaviours were suggested and the importance of hand-washing was identified (Welch, 2004).
High Uncertainty
Uncertainty about
human behaviour
with respect to
appropriate access
to and consumption
of untreated water
from sources.
Private versus public
water supplies Private water supplies in Scotland account
for approximately 3% of the water supply (Pollock, Alexander, Robertson-Kellie and Anderson, 2015) and the majority of these are concentrated in rural areas. Thus, exposure to private water supplies occurs for owners and users of these supplies and their visitors, and also visitors to holiday accommodation.
Water quality in private water supplies is variable. Outbreaks of zoonotic pathogens
High uncertainty
Data on
Cryptosporidium
from private water
supplies are poor
because it is difficult
practically, costly,
and not a legislative
requirement to test
have been associated with private water supply consumption because these are most susceptible to faecal contamination, and are the most likely to have insufficient treatment systems (Pollock, Alexander, Robertson-Kellie and Anderson, 2015).
Users of private water supplies have reported greater rates of C. parvum cases (Pollock et al. 2010). This has also been described in Ireland (Pollock, Alexander, Robertson-Kellie and Anderson, 2015). This is likely to be due to difficulties in detection outlined above.
Private supplies derived from surface waters and those which are influenced by surface water will be vulnerable to contamination from Cryptosporidium or Giardia, particularly in agricultural catchments, prone to contamination by the faeces of animals, if there is a rapid route for recharge to reach the raw water intake and if there are inadequate treatment barriers. Filtration using filters designed for oocysts removal, and adequately designed UV systems, can remove Cryptosporidium, although advice should always be sought from local authority Environmental Health teams.
A small number of dwelling-houses utilize surface waters arising in the Scottish beaver trial area as sources for private water supplies. The pre-release study
found no Cryptosporidium orGiardia in samples taken from those private water supplies when sampled at the kitchen tap (Morrison, 2004)
for the agent
routinely, although
legislation requires
samples to be taken
if there is considered
to be a risk.
Large samples of
water are required
to minimize the
likelihood of false
negatives.
Treatment of
contaminated raw
water
See Hazard Identification in Section 4.
Disinfection alone may be insufficient to reduce contamination with Giardia spp. (Monzingo and Hibler 1987).
Ineffective filtration is an important cause of outbreaks and cases of Giardia in some parts of the world (Monzingo and Hibler 1987)
Properly-operated conventional water treatment processes designed for removal of particulate material are usually very effective in removing Cryptosporidium oocysts from water (Bouchier Report, page 12, cited by Mackie Report 2014 at 2.6.2;
Very low – low
uncertainty about
the treatment of
public drinking
water and mitigation
processes in place to
prevent human
infection.

Pollock et al. 2010).
Effective methods include coagulation, followed by dissolved air flotation or sedimentation (although some treatment works have direct filtration), followed by rapid gravity or pressure filtration ; membrane filtration can also be very effective (Pollock, Alexander, Robertson-Kellie and Anderson, 2015).
In Scotland, infrastructure upgrades to water treatment have been strongly associated with general decreasing trend in Cryptosporidium reports over time (Pollock, Alexander, Robertson-Kellie and Anderson, 2015).
Consumption of
contaminated raw
untreated water or
drinking water
supplies
The consumption of drinking water other than metropolitan mains, or other very carefully designed filtered supplies may represent a risk for giardiasis (Hoque et al. 2002; Jakubowski and Graun, 2002).
In Scotland the public supply serves many rural areas and these should meet the equivalently high standards.
The majority of water-borne giardiasis outbreaks in humans have occurred in unfiltered surface or groundwater systems affected by surface run-off or sewage discharges (Jakubowski and Graun, 2002).
Irrigation waters used for food crops that are traditionally consumed raw may also represent a high risk as a source of Giardia (Thurston- Enriquez et al., 2002). Environmental contamination of such water systems and supplies may result from human, agricultural and wildlife sources (Heitman et al., 2002).
High uncertainty
about human
behaviour and
knowledge about
risks and
consumption
preferences.
B1.Likelihood of consumption of contaminated water supplies (raw and treated drinking
water).
Likelihood of contamination of the raw untreated water supplies is HIGH but effective treatment
and mitigation procedures and risk assessment and sampling are in place to ensure that public
drinking water is safe to drink. Therefore, the likelihood of consumption of contaminated
drinking water is VERY LOW to LOW (VERY LOW UNCERTAINTY).
The likelihood of consuming contaminated raw water is LOW (VERY LOW UNCERTAINTY) This is
because raw water may be untreated, human behaviour and knowledge about the risks of Giardia
and Cryptosporidium spp. is uncertain and there are a number of uncontrolled sources of
infection. The most likely source of contamination is livestock. Although aquatic animals play a
role as a reservoir of Cryptosporidium and Giardia spp., it is not clear whether they are primary or

secondary host species that are being infected through contact via same routes as humans.
Beavers, as one type of wildlife species which is also susceptible to Cryptosporidium and Giardia
spp. infection will also make a contribution to the risk of contamination. However, relative to
other sources, this is likely to be minor (although uncertainty is high). The significance of the
contribution will depend on the likelihood of infection and the proximity of beavers to important
water sources for public water supplies.
7. CONSEQUENCE ASSESSMENT Disease consequences: Beavers may carry Giardia and Cryptosporidium and may contaminate raw water sources. They are thus likely to form part of the reservoir community for both parasites. However, it is important to note that the risk of water contamination that is attributable to beavers (as a source of Cryptosporidium and Giardia spp..) remains unquantified. Other sources of infection are likely to result in a greater numbers of oocysts shed into the environment (such as humans, livestock and other existing wildlife) and will be likely to pose a more significant risk to water contamination than beavers. Therefore, it is unlikely that the reintroduction of beavers will have a significant impact on the public health risk posed by these two pathogens. The low-very low risk posed by beavers must be viewed in the context of the positive influence that beaver presence has on river and stream ecosystems, reducing erosion, trapping sediment, reducing fluctuations in run-off and nutrient cycling (Monzingo and Hibler 1987). Economic consequences: Unknown and outwith scope of the risk assessment. Public health consequences: Human infection with Cryptosporidium and Giardia spp. results in time-limited diarrhoeal disease in otherwise healthy humans. However, in immunosuppressed individuals, these parasites may have life-threatening consequences. The public should be advised to take appropriate precautions when drinking raw untreated water from wild water sources or to avoid doing this. Appropriate water treatment and testing of drinking water should continue to be rigorously implemented by Scottish water (public water supplies) and local authorities (private water supplies).
8. SOURCES OF EXPERT ADVICE
9. AUTHORS
First draft compiled by: Lisa Boden Date: 22 January 2015
Revised by: Lisa Boden, Harriet Auty Date: 25 February 2015
Reviewed within EPIC by: Emily Hotchkiss Date: 3 March 2015
Externally reviewed by: Date:
10.REFERENCES
Abe, N.N., Kimata, I., Iseki, M., 2003. Identification of genotypes of Giardia intestinalis isolates from dogs in Japan by direct sequencing of the PCR amplified glutamate dehydrogenase gene. J. Vet. Med. Sci. 61, 29–33. Anderson, B.C., 1981. Patterns of shedding of Cryptosporidial oocysts in Idaho Calves. J. Am. Vet. Med. Assoc. 178, 982–984

Bajer, A., Bednarska, M., Sinski, E. 1997. Wildlife Rodents from Different Habitats as a Reservoir for Cryptosporidium parvum. Acta Parasitol. 42, 192-194. Beuch, R.R., 1984. Ontogeny and Diurnal Cycle of Fecal Reingestion in the North American Beaver (Castor canadensis). J. Mammal. 65, 347–350. Blewett, D.A., 1989. Quantitative techniques in Cryptosporidium research. In: Angus, K.W., Blewett, D.A. (Eds.), Proceedings of the First International Workshop on Cryptosporidiosis, Edinburgh, Scotland, p. 85. Bodley-Tickell, A.T., Kitchen, S.E., Sturdee, A.P., 2002. Occurrence of Cryptosporidium in agricultural surface waters during an annual farming cycle in lowland UK. Water Res. 36, 1880–6. Budu-Amoako, E., Greenwood, S.J., Dixon, B.R., Barkema, H.W., McClure, J.T., 2012. Occurrence of Cryptosporidium and Giardia on beef farms and water sources within the vicinity of the farms on Prince Edward Island, Canada. Vet. Parasitol. 184, 1–9. Brook, E.J., Hart, C.A., French, N.P., Christley, R.M. 2009. Molecular epidemiology of Cryptosporidium subtypes in cattle in England. The Veterinary Journal. 179(3), 378-382. doi:10.1016/j.tvjl.2007.10.023 Brook, E., Hart, C.A., French, N.P., Christley, R.M. 2008. Prevalence and risk factors for Cryptosporidium spp. infection in young calves. Veterinary Parasitology. 152(1-2), 46-52. Bukhari, Z., Smith, H.V., 1997. Cryptosporidium parvum: oocyst excretion and viability patterns in experimentally infected lambs. Epidemiol. Infect. 119, 105–108. Bugg, R.J., Robertson, I., Elliot, A.D., Thompson, R.C.A., 1999. Gastro intestinal parasites of urban dogs in Perth, Western Australis. Vet. J. 157. Busher, P.E., Warner, R.J., Jenkins, S.H., 2009. Population density, colony composition, and local movements in two Sierra Nevadan beaver populations. J. Mammal. 64, 314–318. Cacciò, S.M., Thompson, R.C.A., McLauchlin, J., Smith, H. V, 2005. Unravelling Cryptosporidium and Giardia epidemiology. Trends Parasitol. 21, 430–7. Campbell, R.D., Harrington, A., Ross, A. and Harrington, L. 2012. Distribution, population assessment and activities of beavers in Tayside. Scottish Natural Heritage Commissioned Report No. - SNH use only, available at http://www.snh.org.uk/pdfs/publications/commissioned_reports/540.pdf (accessed 19 February 2015). Chalmers R.M., Katzer F. 2013. Looking for Cryptosporidium: the application of advances in detection and diagnosis. Trends Parasitol. 29, 237-251 Chilvers, B.L., Cowan, P.E., Waddington, D.C., Kelly, P.J., Brown, T.J., 1998. The prevalence of infection of Giardia spp. and Cryptosporidium spp. in wild animals on farmland, southeastern North Island, New Zealand. Int. J. Environ. Health 8, 59–64. Crawford, J.C., 2007. Mating , kinship , and population structure in Illinois beaver populations. Keep. Davies, R.B., Hibler, C.P., 1979. Animal reservoirs and cross species transmission of Giardia, in: Jakubowski, W., Hoff, J.C. (Eds.), Waterborne Transmission of Giardiasis. U.S. Environmental Protection Agency, Cincinnati, Ohio, pp. 104–126. Current, W.L., 1985.Cryptosporidiosis. J. Am. Vet. Med. Assoc. 187, 1334–1338. Defra, 2011. Qualitative assessment of the risk of introduction of Equine Infectious Anaemia (EIA) into Great Britain from an EIA endemic area through temporary movement of UK origin horses (Roberts H. and Paterson A.). Veterinary Science Team, 17 Smith Square, London. Deshpande, A.P., Jones, B.L., Connelly, L. et al. 2015. Molecular characterization of Cryptosporidium parvum isolates from human cryptosporidiosis cases in Scotland. Parasitol. 142, 318-325.

Dickinson, H. 2014. Tayside Beaver Study Group Final Report. Dixon, B., Parrington, L., Cook, A., Pintar, K., Pollari, F., Kelton, D., Farber, J., 2011. The potential for zoonotic transmission of Giardia duodenalis and Cryptosporidium spp. from beef and dairy cattle in Ontario, Canada. Vet. Parasitol. 175, 20–6. Dixon, B.R., Parenteau, M., Martineau, C., Fournier, J., 1997. A comparison of conventional microscopy, immunofluorescence microscopy and flow cytometry in the detection of Giardia lamblia cysts in beaver fecal samples. J. Immunol. Methods 202, 27–33. Dunlap, B.G., Thies, M.L., 2002. Giardia in Beaver (Castor canadensis ) and Nutria (Myocastor coypus ) From East Texas. J. Parasitol. 88, 1254–1258. Erlandsen, S.L., Sherlock, L. a., Bemrick, W.J., Ghobrial, H., Jakubowski, W., 1990. Prevalence of Giardia spp. in beaver and muskrat populations in Northeastern States and Minnesota: Detection of intestinal trophozoites at necropsy provides greater sensitivity than detection of cysts in fecal samples. Appl. Environ. Microbiol. 56, 31–36. ECDC Cryptosporidium data available at http://www.ecdc.europa.eu/en/publications/_layouts/xlviewer.aspx?id=/en/publications/Report%20Assets/AER-FWD-2014/Number-and-rates-of-confirmed-cryptosporidiosis-reported-cases-EUEEA-2008-2012.xlsx&DefaultItemOpen=1 (accessed 15 January 2015). Enemark, H.L., Ahrens, P., Lowery, C.J., Thamsborg, S.M., Enemark, J.M.D., Bille-Hansen, V., Lind, P., 2002 Cryptosporidium andersoni from a Danish cattle herd: identification and preliminary characterisation. Vet. Parasitol. 107, 37–49. Erlandsen, S.L., Sherlock, L.A., Januschka, M., Schupp, D.G., Schaefer, F.W., Jakubowski, W., Bemrick, W.J., 1988. Cross-Species Transmission of Giardia spp .: Inoculation of Beavers and Muskrats with Cysts of Human , Beaver , Mouse , and Muskrat Origin. Appl. Environ. Microbiol. 54, 2777–2785. Fayer, R., Santin, M., Trout, J.M., DeStefano, S., Koenen, K., Kaur, T., 2006. Prevalence of Microsporidia , Cryptosporidium spp ., and Giardia spp . in Beavers ( Castor canadensis ) in Massachusetts. J. Zoo Wildl. Med. 37, 492–497. Frost, F., Plan, B., Liechty, B., 1980. Giardia prevalence in commercially trapped mammals. J. Environ. Health 42, 245–249. Geurden, T., Thomas, P., Casaert, S., Vercruysse, J., Claerebout, E., 2008. Prevalence and molecular characterisation of Cryptosporidium and Giardia in lambs and goat kids in Belgium. Vet. Parasitol. 155, 142–5. Gillhuber, J., Pallant, L., Ash, A., Thompson, R.C.A., Pfister, K., Scheuerle, M.C., 2013. Molecular identification of zoonotic and livestock-specific Giardia-species in faecal samples of calves in Southern Germany. Parasit. Vectors 6, 346. Goh, S., Reacher, M., Casemore, D.P., Verlander, N.Q., Chalmers, R., Knowles, M., Williams, J., Osborn, K., Richards, S., 2004. Sporadic cryptosporidiosis, North Cumbria, England, 1996-2000. Emerg. Infect. Dis. 10, 1007–1015. Goh, S., Reacher, M., Casemore, D.P., Verlander, N.Q., Charlett, A., Chalmers, R.M., Knowles, M., Pennington, A., Williams, J., Osborn, K., Richards, S., 2005. Sporadic cryptosporidiosis decline after membrane filtration of public water supplies, England, 1996-2002. Emerg. Infect. Dis. 11, 251–259. Goodman, G., Girling, S., Pizzi, R., Meredith, a., Rosell, F., Campbell-Palmer, R., 2012. Establishment of a Health Surveillance Program for Reintroduction of the Eurasian Beaver (Castor Fiber) into Scotland. J. Wildl. Dis. 48, 971–978.

Heitman, T.L., Frederick, L.M., Viste, J.R., Guselle, N.J., Morgan, U.M., Thompson, R.C. Olson, M.E., 2002. Prevalence of Giardia and Cryptosporidium and characterization of Cryptosporidium spp. isolated from wildlife, human, and agricultural sources in the North Saskatchewan River Basin in Alberta, Canada. Can. J. Microbiol. 48, 530–541. Henriksen, S.A., 1989. Epidemiology of cryptosporidiosis in calves. In: Angus, K.W., Blewett, D.A. (Eds.), Proceedings of the First International Workshop on Cryptosporidiosis, p. 79. Hunter, P.R., Chalmers, R.M., Syed, Q., Hughes, L.S., Woodhouse, S., Swift, L., 2003. Foot and mouth disease and cryptosporidiosis: possible interaction between two emerging infectious diseases. Emerg. Infect. Dis. 9, 109–12. Hunter, P.R. et al. 2004. Health sequelae of human cryptosporidiosisin immunocompetent patients.Clin. Infect. Dis. 39, 504–510 Hunter, P.R., Nichols, G., 2002. Epidemiology and Clinical Features of Cryptosporidium Infection in Immunocompromised Patients Epidemiology and Clinical Features of Cryptosporidium Infection in Immunocompromised Patients. Clin. Microbiol. Rev. 15, 145–154. Hunter, P.R., Thompson, R.C.A., 2005. The zoonotic transmission of Giardia and Cryptosporidium. Int. J. Parasitol. 35, 1181–90. Kirner, J.C., Littler, J.D., Angelo, L.A., 1978. A waterborne outbreak of Giardiasis in Camas, Wash. J. Am. Water Works Assoc. 70, 35–40. Learmonth, J.J. et al., 2004. Genetic characterization and trans- 65 mission cycles of Cryptosporidium species isolated from humans in New Zealand. Appl. Environ. Microbiol. 70, 3973–3978. Leib, M.S., Zajec, A.M., 1999. Giardiasis in dogs and cats. Vet. Med. 9, 793–802. Leonhard, S., Pfister, K., Beelitz, P., Wielinga, C., Thompson, R.C.A., 2007. The molecular characterisation of Giardia from dogs in Southern Germany. Vet. Parasitol. 150, 33–38. Lindsay, D.S., Upton, S.J., Owens, D.S., Morgan, U.M., Mead, J.R., Blagburn, B.L., 2000. Cryptosporidium andersoni. sp. (Apicomplexa: Cryptosporiidae) from cattle, Bostaurus. J. Eukaryot. Microbiol. 47, 91–95 Mackie, P., 2014. Scottish Beaver Trial Independent Public Health Monitoring 2009-2014 Report and Recommendations, Planning and Regulatory Services Argyll and Bute Council. Maddox-Hyttel, C., Langkjaer, R.B., Enemark, H.L., Vigre, H., 2006. Cryptosporidium and Giardia in different age groups of Danish cattle and pigs--occurrence and management associated risk factors. Vet. Parasitol. 141, 48–59. McGlade, T.R., Robertson, I.D., Elliot, A.D., Thompson, R.C.A., 2003. High prevalence of Giardia detected in cats by PCR. Vet. Parasitol. 110, 197–203. Minetti, C., Taweenan, W., Hogg, R., Featherstone, C., Randle, N., Latham, S.M., Wastling, J.M., 2013. Occurrence and Diversity of Giardia duodenalis Assemblages in Livestock in the UK. Transbound. Emerg. Dis. 61. Monis, P.T., Thompson, R.C.A., 2003. Cryptosporidium and Giardia-zoonoses: Fact or fiction? Infect. Genet. Evol. 3, 233–244. Monzingo, D.L., Hibler, C.P., 1987. Prevalence of Giardia spp. in a beaver colony and the resulting environmental contamination. J. Wildl. Dis. 23, 576–585. Morrison, A., 2004. Trial re-introduction of the European beaver to Knapdale: public health monitoring 2001–3. Scottish Natural Heritage Commissioned Report No. 077 (ROAME No. F02AC327).

Naiman, R.J., Johnston, C. a, Kelley, J.C., 1988. Alteration of North American Streams by Beaver: The structure and dynamics of streams are changing as beaver recolonize their historic habitat. Bioscience 38, 753–762. Novak, M., 1977. Determining the average size and composition of beaver families. J. Wildl. Manage. 41, 751–754. Nummi, P., Kuuluvainen, T., 2013. Forest disturbance by an ecosystem engineer: Beaver in boreal forest landscapes. Boreal Environ. Res. 18, 13–24. O’Handley, R.M., Olsen, A.M., Fraser, D., Adams, P., Thompson, R.C.A., 2000. Prevalence and genotypic characterisation of Giardia in dairy calves from Western Australia and Western Canada. Vet. Parasitol. 90, 193–200. OIE, 2004. Handbook on Import Risk Analysis for Animals and Animals products - Volume I. Introduction and qualitative risk analysis. Paziewska, A., Bednarska, M., Niewęgłowski, H., Karbowiak, G., Bajer, A., 2007. Distribution of Cryptosporidium and Giardia spp. in selected species of protected and game mammals From North-Eastern Poland. Ann. Agric. Environ. Med. 14, 265–270. Pollock, K.G.J., Young, D., Smith, H.V., Ramsay, C.N. 2008. Cryptosporidiosis and filtration of water from Loch Lomond, Scotland. Emerg. Infect. Dis. 14, 115-120. Pollock, K.G.L., Ternent, H., Mellor, D.J. et al. (2010) Spatial and Temporal Epidemiology of Sporadic Cryptosporidiosis in Scotland. Zoonoses Pub Health. 57, 487-492. Pollock, K.G.J., Alexander, C., Robertson-Kellie, C. and Anderson, E., (2015). What is the likelihood that beavers will be an important source of contamination of the water supply with Cryptosporidium parvum and Giardia intestinalis? Available from K Pollock on request. Private Water Supplies Technical Manual, available at http://www.privatewatersupplies.gov.uk/private_water/files/Full%20Doc.pdf (accessed 25 February 2015). Robertson, I.D., Irwin, P.J., Lymbery, A.J., Thompson, R.C.A., 2000. The role of companion animals in the emergence of parasitic zoonoses. Int. J. Parasitol. 30, 1369–1377. Robertson L.J and Gjerde B. 2001. Occurrence of Cryptosporidium oocysts and Giardia cysts in raw waters in Norway. Scand J Public Health. 29:200 DOI:10.1177/14034948010290030901 Available online at http://sjp.sagepub.com/content/29/3/200 Robertson, B. et al. 2002.Case-control studies of sporadic crypto- sporidiosis in Melbourne and Adelaide, Australia. Epidemiol. Infect.128, 419–431. Rose, J.B., Haas, C. and Regli, S. 1991. Risk Assessment and Control of Water-borne Giardiasis. Am J Public Health. 81, 709-713. Rosell, F., Rosef, O., Parker, H., 2001. Investigations of waterborne pathogens in Eurasian beaver (Castor fiber) from Telemark County, southeast Norway. Acta Vet. Scand. 42, 479–482. Roy, S.L. et al. 2004. Risk factors for sporadic cryptosporidiosis among immunocompetent persons in the United States from1999–2001. J. Clin. Microbiol. 42, 2944–2951. Ryan, U.M., Bath, C., Robertson, I., Read, C., Elliot, A., McInnes, L., Traub, R., Besier, B., 2005. Sheep may not be an important zoonotic reservoir for Cryptosporidium and Giardia parasites. Appl. Environ. Microbiol. 71, 4992–7.

Santín, M., Trout, J.M., Fayer, R., 2007. Prevalence and molecular characterization of Cryptosporidium and Giardia species and genotypes in sheep in Maryland. Vet. Parasitol. 146, 17–24. Smerdon, W.J. et al. (2003) Foot and mouth disease in livestock and reduced cryptosporidiosis in humans, England and Wales.Emerg. Infect. Dis. 9, 22–28 Smith, H.V., Nichols, R. A. B. ,Mallon, M., Macleod, A., Tait, A. Reilly, W.J., Browning, L.M., Gray, D., Reid, S.W.J., Wastling, J.M., 2005. Natural Cryptosporidium hominis infections in Scottish cattle. Veterinary Record 156, 710-711 doi:10.1136/vr.156.22.710 Sprong, H., Cacciò, S.M., Van Der Giessen, J.W.B., 2009. Identification of zoonotic genotypes of Giardia duodenalis. PLoS Negl. Trop. Dis. 3, 1–12. Stuart, J.M., Orr, H.J., Warburton, F.G., Jeyakanth, S., Pugh, C., Morris, I., Sarangi, J., Nichols, G., 2003. Risk factors for sporadic giardiasis: a case control study in south western England. Emerg. Infect. Dis. 9, 229–233. Sturdee, A.P., Bodley-Tickell, A.T., Archer, A., Chalmers, R.M., 2003. Long-term study of Cryptosporidium prevalence on a lowland farm in the United Kingdom. Vet. Parasitol. 116, 97–113. Sulaiman, I.M., Fayer, R., Bern, C., Gilman, R.H., Trout, J.M., Schantz, P.M., Das, P., Lal, A. a., Xiao, L., 2003. Triosephosphate Isomerase Gene Characterization and Potential Zoonotic Transmission of Giardia duodenalis. Emerg. Infect. Dis. 9, 1444–1452. Svendsen, G.E., 1980. Population Parameters and Colony Composition of Beaver ( Castor canadensis ) in Southeast Ohio. Am. Midl. Nat. 104, 47–56. Svendsen, G.E., 1989. Pair formation, duration of pair-bonds, and mate replacement in a population of beavers (Castor canadensis). Can. J. Zool. 67, 336–340. Thompson, R.C.A., 2004. The zoonotic significance and molecular epidemiology of Giardia and giardiasis. Vet. Parasitol. 126, 15–35. Thompson, R.C.A., Olsen, A.M., Zhu, G., Enomoto, S., Abrahamsen, M.S., Hijjawi, N.S., 2005. Cryptosporidium and cryptosporidiosis. Adv. Parasitol. 59, 77–158. Traub, R.J., Monis, P.T., Robertson, I.D., Irwin, P., Mencke, N., Thompson, R.C.A., 2004. Epidemiological and molecular evidence supports the zoonotic transmission of Giardia among humans and dogs living in the same community. Parasitology 128, 258–262. Trout, J.M., Santin, M., Fayer, R., 2003. Identification of assemblage A Giardia in white-tailed deer. J. Parasitol. 89, 1254–5. Van Keulen, H., Macechko, P.T., Wade, S., Schaaf, S., Wallis, P.M., Erlandsen, S.L., 2002. Presence of human Giardia in domestic, farm and wild animals, and environmental samples suggests a zoonotic potential for giardiasis. Vet. Parasitol. 108, 97–107. Wade, S.E., Mohammed, H.O., Schaaf, S.L., 2000. Prevalence of sp., Cryptosporidium parvum and Cryptosporidium muris (C. andersoni) in 109 dairy herds in five counties of southeastern New York. Vet. Parasitol. 93, 1–11. Welch, T., 2004. Evidence-based medicine in the wilderness: The safety of back country water. Wilderness Environ. Med. 15, 235–237. Wells, B., Shaw, H., Hotchkiss, E. Gilray, J., Ayton, R., Katzer F., Wells, A., Innes, E. 2015. Prevalence, species identification and genotyping Cryptosporidium from livestock and deer in a catchment in the Cairngorms with a history of a contaminated public water supply. Parasit Vectors. 4 (8(1)), 66.

Wright, J.P., Jones, C., 2002. An Ecosystem Engineer, the Beaver, Increases Species Richness at the Landscape Scale. Oecologia 132, 96–101. Xiao, L., Fayer, R., Ryan, U., Upton, S.J., 2004. Cryptosporidium taxonomy: recent advances and implications for public health. Clin. Microbiol. Rev. 17, 72–97. Zhou, L. et al. (2003) Molecular surveillance of Cryptosporidium spp. in raw wastewater in Milwaukee: implications for understanding 43 outbreak occurrence and transmission dynamics. J. Clin. Microbiol.41, 5254–5257
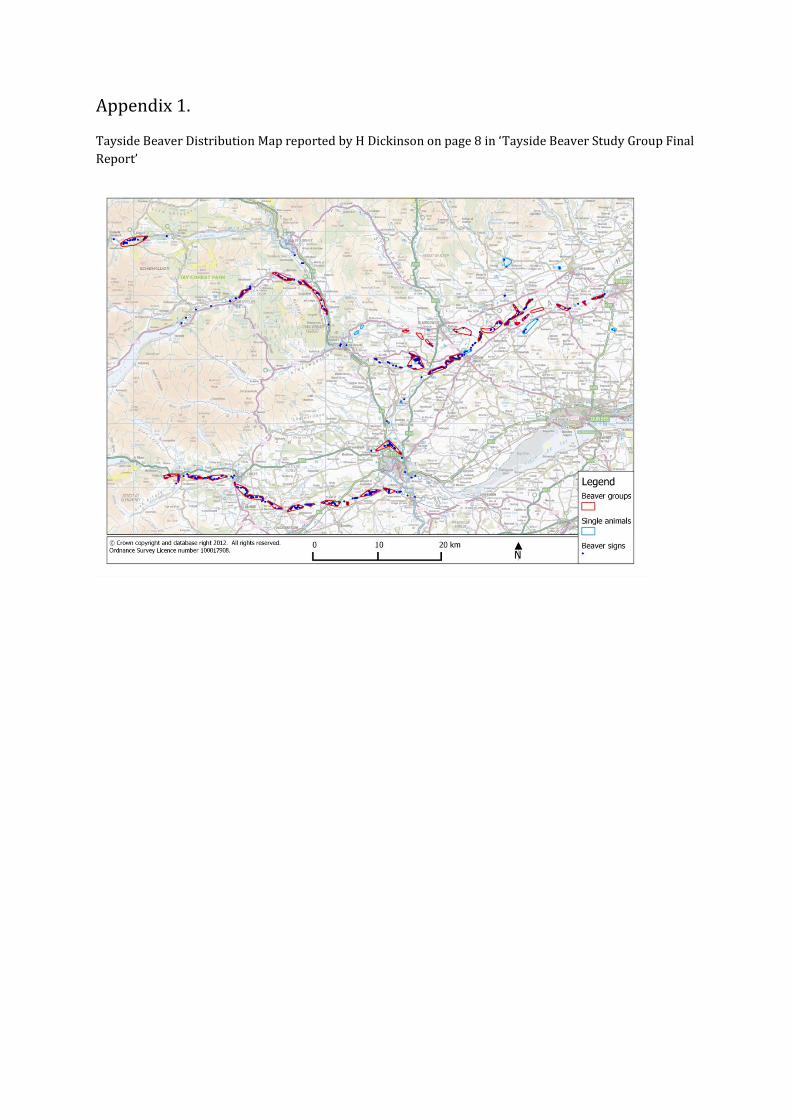
Appendix 1.
Tayside Beaver Distribution Map reported by H Dickinson on page 8 in ‘Tayside Beaver Study Group Final
Report’

Appendix 2
What is the likelihood that beavers will be an important source of contamination of
drinking water supplies with Cryptosporidium parvum and Giardia intestinalis?
Pollock KGJ 1, Alexander C
2, Robertson-Kellie C
3, Anderson E
1
1 Health Protection Scotland, Meridian Court, Glasgow,
2 Scottish Parasite Diagnostic Reference Laboratory, Lister Building, Glasgow,
3 Drinking Water Quality Regulator for Scotland, Edinburgh.
INTRODUCTION
In 2009/10, 16 Norwegian Eurasian beavers (Castor fiber) were reintroduced to
Knapdale in Argyll, Scotland as part of a 5-year reintroduction trial (Scottish Beaver Trial). A
veterinary health surveillance programme was commenced and beaver faecal and blood
samples longitudinally obtained to assess presence of micro-organisms, which may have an
impact on public health (Gidona et al., 2012). Although no protozoan pathogens such as
Cryptosporidium spp. and Giardia intestinalis were identified, detection and sampling
methodologies for protozoans were arguably sub-optimal.
HPS and partners (University of Glasgow, School of Veterinary Medicine; Drinking
Water Quality Regulator for Scotland) were asked to supplement this work with a qualitative
risk assessment of the additional public health risk posed by the re-introduction of beavers to
Scotland, if any, in relation to Cryptosporidium and Giardia infections. A review of
published literature on the risk to public health posed by beavers in similar settings has been
undertaken. This paper provides an overview of the human epidemiology of these infections
supplementing the literature review, and placing it in the context of related extant public
health measures in Scotland.
Diagnostic methods to detect Cryptosporidium spp. and Giardia intestinalis
The standardised methods adopted by clinical microbiology laboratories throughout
Scotland to detect Cryptosporidium oocysts and Giardia cysts from human faeces are based
on the Standard for Microbiology Investigations (SIM Staining Procedures, TP 39). This

describes the combined use of stains and microscopy to detect parasitic structures. Although
microscopy is inexpensive, it requires significant expertise, hence laboratories are
increasingly turning to commercial kits which are deemed to be more sensitive in situations
where microscopy expertise is lacking. The kits are ELISA / capillary-based and detect
parasite-specific antigens which eliminates the requirement for microscopy expertise
(Alexander et al, 2013).
However, neither microscopy nor antigen detection can be used to speciate
Cryptosporidium which is necessary when determining if the oocysts represent species which
have the potential to cause disease in humans and animals. Oocysts of pathogenic and non-
pathogenic species are similar in size, therefore isolating oocysts from environmental samples
does not necessarily mean they will impact adversely on human health. Similarly, antigen-
based kits may be a useful screening tool but they do not differentiate the species.
The advent of molecular tools allows Cryptosporidium DNA, extracted from oocysts,
to be amplified and speciated (Hadfield et al. 2011). In addition to speciation, further in-
depth information is possible using molecular amplification assays which target specific
regions of parasite DNA e.g. GP60 gene (for Cryptosporidium; Glaberman et al. 2002) and
the β-Giardin gene (for Giardia; Caccio et al, 2002) to determine subtypes and assemblages,
respectively. This information is essential, in addition to speciation, for source tracing during
outbreaks.
Surveillance of human cases of Cryptosporidium spp. and Giardia intestinalis
HPS is the national surveillance centre in Scotland for communicable and infectious
diseases. Reports of identifications of pathogens are sent to HPS from all the laboratories
within NHS Boards in Scotland by a system of voluntary reporting. This system relies upon
the goodwill of the laboratories and it is assumed that there is close collaboration and a high
degree of co-operation between all involved, including both the laboratories and the public
health teams in Scotland. Surveillance of Cryptosporidium spp. and Giardia intestinalis in
Scotland is at the genus level, although outbreak cases of Cryptosporidium are speciated.

Cryptosporidium
Clinical features
Characteristic signs of Cryptosporidium infection are profuse, watery diarrhoea, often
accompanied by bloating, abdominal pain, and nausea or vomiting. Illness is typically self-
limiting but can last for 2–3 weeks; studies suggest an association with long-term health
sequelae, such as post-infection irritable bowel syndrome in a proportion of individuals
(Thabane et al., 2009).
Epidemiology
Recent trends
Cryptosporidiosis, the most common parasitic gastrointestinal disease in the Scottish
population, is caused mainly by two protozoan pathogens, Cryptosporidium
parvum (C. parvum) and Cryptosporidium hominis (C. hominis) which account for over 90%
of human cryptosporidiosis worldwide (Putignani and Menichella, 2010). Each year since
2005, Health Protection Scotland has received reports of between 450–700 laboratory-
confirmed cases of cryptosporidiosis (9–14 cases/100,000 population/year) (figure 1).
Seasonality
Seasonality is usually markedly biphasic, peaking in spring and early autumn.
The epidemiology of cryptosporidiosis in Scotland tends to exhibit a bi-modal seasonality,
where C. parvum predominates from April to June, and C. hominis is more prevalent from
August to November (Pollock et al., 2010; Deshpande et al., 2015). The Spring peak usually
corresponds to an increase of infectious
faecal material during the calving/lambing season. Isolates from human cases within and out-
with the UK have the same GP60 subtypes as animal sources supporting the hypothesis that
ruminants are the main sources of infection (Deshpande et al., 2015).
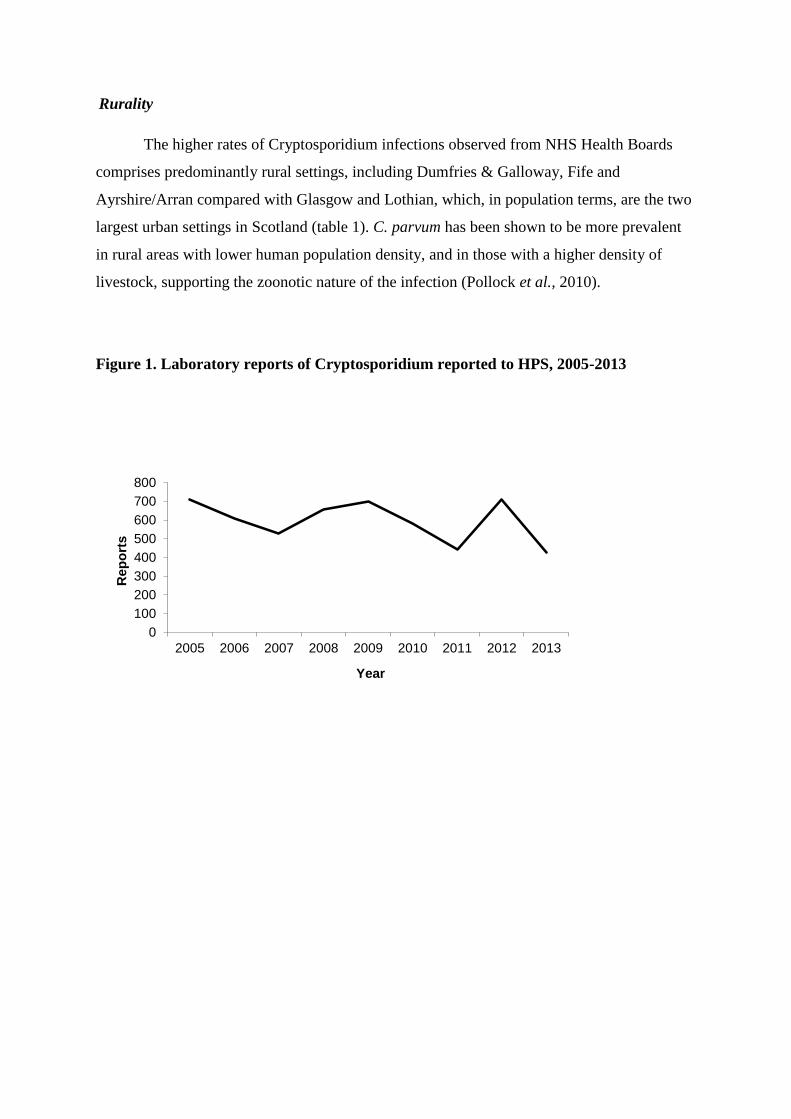
Rurality
The higher rates of Cryptosporidium infections observed from NHS Health Boards
comprises predominantly rural settings, including Dumfries & Galloway, Fife and
Ayrshire/Arran compared with Glasgow and Lothian, which, in population terms, are the two
largest urban settings in Scotland (table 1). C. parvum has been shown to be more prevalent
in rural areas with lower human population density, and in those with a higher density of
livestock, supporting the zoonotic nature of the infection (Pollock et al., 2010).
Figure 1. Laboratory reports of Cryptosporidium reported to HPS, 2005-2013
0
100
200
300
400
500
600
700
800
2005 2006 2007 2008 2009 2010 2011 2012 2013
Rep
ort
s
Year
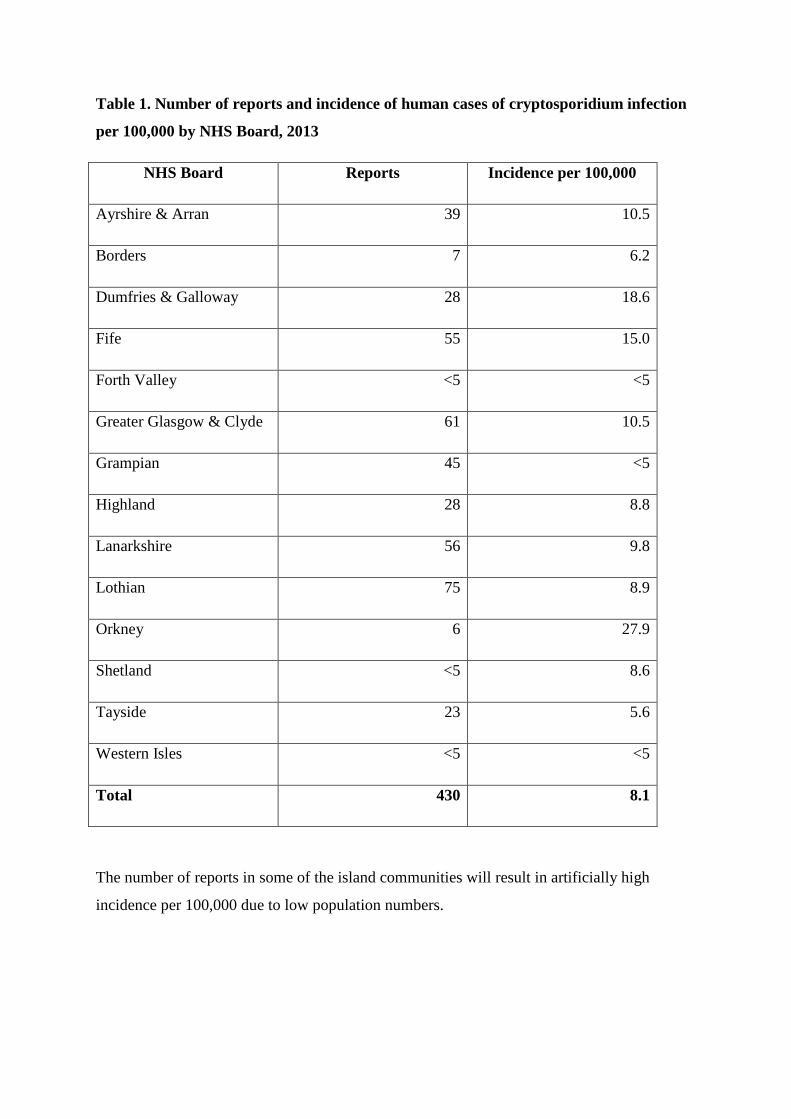
Table 1. Number of reports and incidence of human cases of cryptosporidium infection
per 100,000 by NHS Board, 2013
NHS Board Reports Incidence per 100,000
Ayrshire & Arran 39 10.5
Borders 7 6.2
Dumfries & Galloway 28 18.6
Fife 55 15.0
Forth Valley <5 <5
Greater Glasgow & Clyde 61 10.5
Grampian 45 <5
Highland 28 8.8
Lanarkshire 56 9.8
Lothian 75 8.9
Orkney 6 27.9
Shetland <5 8.6
Tayside 23 5.6
Western Isles <5 <5
Total 430 8.1
The number of reports in some of the island communities will result in artificially high
incidence per 100,000 due to low population numbers.

Risk factors for infection
The main risk factors for Cryptosporidium infection in Scotland include : exposure to
contaminated water (either via contaminated drinking water or exposure related to outdoor
recreational activities), direct contact with animals including during lambing, or via visits to
petting farms, foreign travel, and direct person to person spread. Cases and outbreaks related
to failure of treatment of swimming pool water also occur.
Exposure to contaminated water
Drinking water contaminated with Cryptosporidium oocysts from species known to
induce human disease is a recognized risk factor for human illness (Pollock et al., 2008). The
sources of drinking water such as rivers or reservoirs can be contaminated by faecal material
from a variety of sources, including livestock, feral animals, farming activities such as slurry
spreading or human sewage. Oocysts can remain infectious in the environment for prolonged
periods of time and can withstand freezing. There have been historic issues with drinking
water supplies as oocysts are resistant to chlorination, the process used to disinfect all public
drinking water supplies and many private water supplies. To prevent human exposure,
oocysts must be physically removed from water supplies by filtration. Additionally, UV
disinfection is commonly practised on many private water supplies and can inactivate
oocysts. However, deficiencies in these processes can expose persons to risk of infection
from viable oocysts (Ibid.).
Where drinking-water filtration has been enhanced on public drinking water supplies
to reduce oocyst counts, the incidence of reported clinical Cryptosporidium infection has
been reduced (Ibid.). However, reported rates of infection are subject to variation, depending
on factors such as local laboratory testing criteria, and exposure source attribution depends on
the quality of risk-factor exposure data. Therefore, assessing trends in clinical infection rates
might not be sufficiently sensitive for detecting changes in single-exposure risks. Variations
in other risk factors (e.g. foreign travel, direct animal contact) can also obscure an effect
associated with reduced exposure to oocysts in drinking water. Assessment of the effects of
changes in environmental oocyst exposure would ideally be based on measuring population-
level indicators, rather than relying on reported (self-selected) cases of laboratory-confirmed
cryptosporidiosis (Ramsay et al., 2014).

Public drinking water supplies
To minimise the risk for waterborne Cryptosporidium infection from drinking-water
supplies, the water industry identified the need for effective barrier water treatment systems.
In Scotland, considerable investment in treatment processes known to remove oocysts from
water, such as coagulation and filtration, and membrane filtration, along with optimisation of
existing water treatment processes and risk-based sampling of supplies has significantly
reduced the concentration of Cryptosporidium oocysts in treated water (Pollock et al., 2010).
Cryptosporidium is monitored by Scottish Water in the raw water of some water
treatment works and in the treated water of all works. The 2013 Annual Report from the
Drinking Water Quality Regulator for Scotland (DWQR) reported that 17% of treatment
works had one or more samples of treated water containing oocysts; this compared with 29%
in 2012 and 34% in 2011. All detections are investigated by Scottish Water and reported to
Health Boards, the Drinking Water Quality Regulator and local authorities. These
infrastructure upgrades are strongly associated with the general decreasing trend in
Cryptosporidium reports over time (656 cases in 2009 to 430 cases in 2013) (Pollock et al.,
2008; Pollock et al., 2014). The Scottish total of 430 lab reports in 2013 is the lowest number
of reports since meaningful surveillance commenced in 1988.
The most effective method of protecting water supplies from Cryptosporidium is to
prevent the contamination of source waters; however, appropriate, effectively operated and
maintained treatment of public drinking water supplies minimises Cryptosporidium oocyst
loads and concomitantly reduces risk of waterborne infection. It is unlikely that the re-
introduction of beavers to areas to catchment areas supplying the public drinking water
supply should have a significant impact on contamination of the water supply over and above
the faecal load excreted by livestock and feral animals. However, it is recommended that any
proposed re-introduction of beavers should be discussed in detail with Scottish Water to
allow appropriate risk assessments to be undertaken and water safety plans to be modified if
required.
Private water supplies
Within the rural areas of Scotland, there are over 19,811 reported private water
supplies compared to less than 240 in urban areas. Treatment of PWS is highly variable

ranging from no treatment to carefully-maintained UV-filtered treatment. Areas with a
greater number of private water supplies to human inhabitants have reported a greater number
of C. parvum cases (Pollock et al. 2010). Similarly, in Ireland, an increased prevalence of C.
parvum in rural, lowly populated areas has been described (Zintl et al., 2009). The
distribution of C. parvum sub-types from human isolates by Health Board (Table 2)
highlights the molecular potential to monitor changes in aetiology associated with potential
introduction of a beaver population.
Table 2. Distribution of C. parvum subtypes from human isolates across Scotland, 2012-
2013.
Approximately 3% of the Scottish population (170,000) uses a PWS for drinking
water. Inadequately treated PWS may pose a threat to public health (Pollock et al., 2010;
Kilchoman E. coli O157 Outbreak Control Team report) and outbreaks of zoonotic pathogens
such as E. coli O157 have been associated with PWS consumption. Non-means tested
Scottish Government grants are available for upgrading private water supplies e.g.
installation of UV filters but many supplies ( a reported 81%) have no treatment; this is
thought to be because residents in such areas either presume immunological experience to
pathogens such as Cryptosporidium, or are unwilling to install treatment, or simply do not
maintain existing treatment systems.

Lay evaluations of zoonotic risk reflect intuitive and experience-based estimates of
the risk rather than probabilistic estimates (Strachan et al., 2011). Because of the lack of
treatment, and lack of management of many PWS, it is PWS which are most susceptible to
faecal contamination, and transient consumers of these who are most at risk of infection from
C. parvum especially if the water supply is inadequately treated or managed. The eventual
prevalence of beavers shedding cryptosporidium will likely present a small additional risk to
human health relative to the risk associated with excretion by the large numbers of various
domestic and wild animals, livestock and human sources which contribute to the current
endemnicity. Further, while re-introduction of beavers to catchments, which serve PWS, may
augment the risk of infection with Cryptosporidium, the number of potentially exposed
individuals represents a small proportion of the total population, and all owners of PWS are
encouraged and incentivised to ensure effective treatment is in place on their supplies. As a
precaution, however, it is recommended that any plans to re-introduce beavers and in PWS
catchments should be discussed with local authority Environmental Health teams to ensure
that the local risk is fully evaluated.

Giardia
Giardia intestinalis (synonyms: G. lamblia, G. duodenalis) is a flagellated protozoan
parasite, the species complex of which comprises a series of largely host-adapted
assemblages.
Clinical features
Infection with Giardia can be asymptomatic but it usually manifests as an acute or
chronic disease. Symptoms include chronic diarrhoea, bloating, belching (historically referred
to as the ‘purple burps’), nausea and significant weight loss. Furthermore, long-term sequelae
associated with this infection can be debilitating (Wensaas et al., 2012) including irritable
bowel syndrome and chronic fatigue.
Epidemiology
Recent trends
Between 1988 and 1998, the number of laboratory-confirmed reports of human cases
of Giardia infection in Scotland fluctuated between 300-400 (Pollock et al., 2005). However,
from 2007, the annual number of Giardia reports to HPS has gradually declined to less than
200 (figure 2).
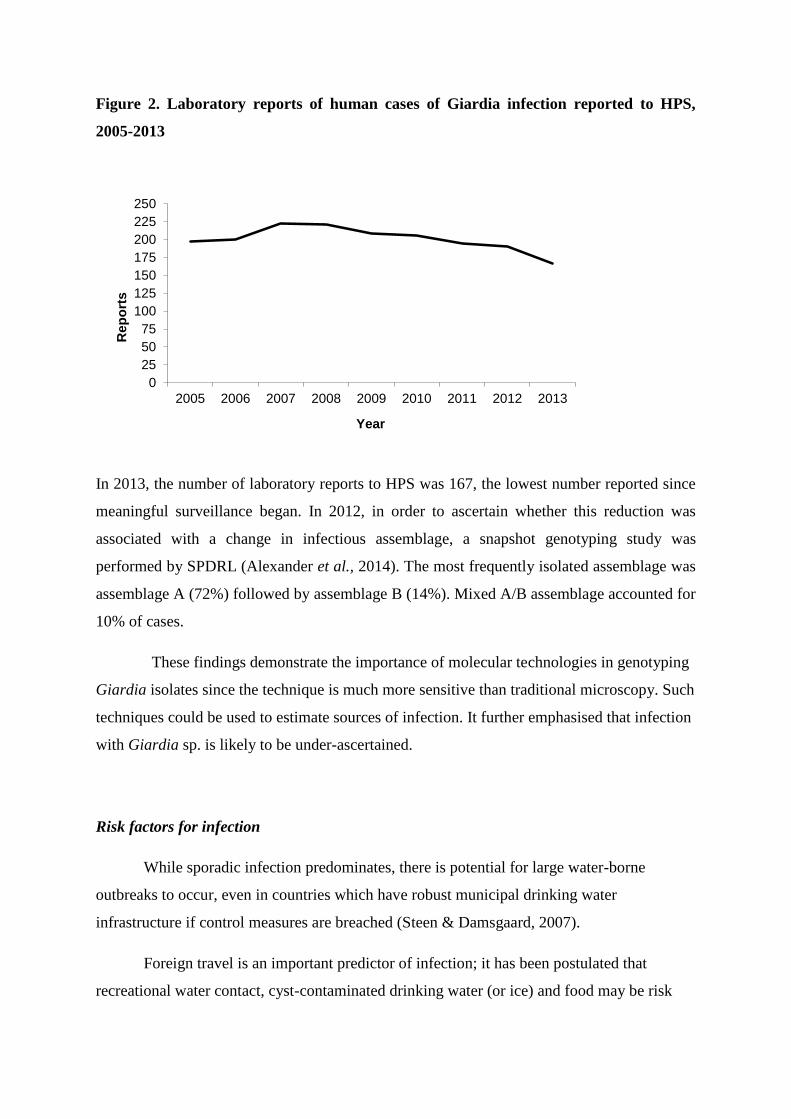
Figure 2. Laboratory reports of human cases of Giardia infection reported to HPS,
2005-2013
In 2013, the number of laboratory reports to HPS was 167, the lowest number reported since
meaningful surveillance began. In 2012, in order to ascertain whether this reduction was
associated with a change in infectious assemblage, a snapshot genotyping study was
performed by SPDRL (Alexander et al., 2014). The most frequently isolated assemblage was
assemblage A (72%) followed by assemblage B (14%). Mixed A/B assemblage accounted for
10% of cases.
These findings demonstrate the importance of molecular technologies in genotyping
Giardia isolates since the technique is much more sensitive than traditional microscopy. Such
techniques could be used to estimate sources of infection. It further emphasised that infection
with Giardia sp. is likely to be under-ascertained.
Risk factors for infection
While sporadic infection predominates, there is potential for large water-borne
outbreaks to occur, even in countries which have robust municipal drinking water
infrastructure if control measures are breached (Steen & Damsgaard, 2007).
Foreign travel is an important predictor of infection; it has been postulated that
recreational water contact, cyst-contaminated drinking water (or ice) and food may be risk
0
25
50
75
100
125
150
175
200
225
250
2005 2006 2007 2008 2009 2010 2011 2012 2013
Rep
ort
s
Year

factors for acquisition of infection abroad (Pollock et al., 2005; Breathnach et al., 2010).
While the sub-Indian continent has historically been associated with infection, travellers
returning from Africa and Europe may also be affected.
In Scotland, the majority of Giardia cases are infected with assemblage A, which is
possibly associated with milder infections. This may partly account for the apparent decrease
in Giardia cases in Scotland, since those infected may be less likely to consult their general
practitioner. At odds with this finding is that 1 in 5 is admitted to hospital for further
treatment
Given that most reported cases of Giardia sp. in Scotland are due to foreign travel, it
unlikely that re-introduction of beavers to Scotland will have a significant impact on the
incidence of this pathogen.
Epidemiology of human cases of cryptosporidiosis and giardiasis in European countries
where beavers are endemic
Castor fiber is endemic in both Norway and Sweden. A brief literature review of
scientific publications through PubMed from 2010, indicate that the epidemiology of human
cases of both cryptosporidiosis and giardiasis in these countries is similar to that in Scotland.
Multi-locus genotyping of human Giardia isolates in Sweden suggests limited zoonotic
transmission (Lebbad et al., 2011). The pattern of outbreaks of cryptosporidiosis in Sweden
are comparable to that historically observed in Scotland i.e. large outbreaks through
inadequately treated municipal drinking water (Widerstrom et al., 2014), foodborne
outbreaks and family clusters (Insulander et al., 2013). There is no explicit mention of the
role of beavers as a source of transmission in these studies.
In Norway, since 2006, investigators (primarily from the Dept of Food Safety
in Oslo) assessed the zoonotic potential of Cryptosporidium from sheep, goats, red foxes,
deer, reindeer, suckling pigs, dogs and calves. However, there have been no published
studies appraising the zoonotic potential of these protozoal pathogens from beavers.
Outbreaks of cryptosporidiosis in Norway mirror that of Scotland with waterborne
transmission via poorly-managed treatment systems, foodborne transmission, and zoonotic
transmission from ruminants (Lange et al., 2014).

Investigators from the aforementioned Dept of Food Safety in Oslo also assessed the
zoonotic potential of Giardia from a number of hosts such as musk rats, calves and dogs but
not from beavers. The epidemiology of giardiasis in Norway appears similar to Scotland i.e.
sporadic infection predominates; however, a large water-borne outbreak of giardiasis did
occur in 2004, through municipal drinking water which had been contaminated with human
sewage (Steen & Damsgaard, 2007). There is no explicit mention of the role of beavers as a
source of transmission in these studies.
Conclusion
The above epidemiological and microbiological information is provided to
contextualise the literature search apropos the potential additional risk to human public health
relating to infections with Cryptosporidium and Giardia posed by the re-introduction of
beavers to Scotland. In conclusion:
The re-introduction is likely to present a very small additional risk as the number of
beavers excreting these organisms will be likely to be small relative to the large number
of wild, domestic and livestock animals and humans which contribute to existing public
health risk.
Further, based on the current epidemiology and implemented control measures,
the main potential route of additional risk of infection with Cryptosporidium or Giardia
is via the contamination of drinking water supplies; however:
The majority of the population is served by public water supplies which have
effective barriers and monitoring in place such that the introduction of beavers is
unlikely to pose an additional public health risk via this route
A smaller proportion of the population which is served by, or exposed to, private
water supplies
Owners of private water supplies are advised, encouraged and supported by Scottish
Government and local authority initiatives to ensure adequate treatment processes are
in place to prevent microbiological contamination

HPS and the SPDRL work together to provide robust surveillance of human cases of
these infections and recent trends show downward trends for both organisms
Increased monitoring and epidemiological assessment should continue including
molecular studies (human, animal and environmental) to further characterise likely
sources of acquisition
There is no evidence from Norway that the presence of beavers has adversely affected
the number of human cases of Cryptosporidium or Giardia
The public is provided with advice about how to avoid exposure to infection when
enjoying the countryside
Recommendations
As a precaution, and to provide further assurance, the following are recommended:
Enhanced surveillance by HPS and SPDRL of all cryptosporidiosis and giardiasis
human cases within Scotland for a pre-determined period e.g. twelve months.
Continued promotion of best practice in relation to public and private water supplies
and the public’s interaction with the countryside.
Plans to re-introduce beavers in PWS catchments should be discussed with local
authority Environmental Health teams to ensure that risk is fully evaluated.
Any proposed re-introduction of beavers should be discussed in detail with Scottish
Water to allow appropriate risk assessments to be undertaken and water safety plans
to be modified if required.

References
Alexander, C., Jones, B., Inverarity, D., Pollock, K.G. (2014) Genotyping of Giardia isolates
in Scotland: a descriptive epidemiological study. Epidemiol Infect, 142, 1636-1639.
Alexander CL, Niebel M, Jones B. The rapid detection of Cryptosporidium and Giardia
species in clinical stools using the Quik Chek immunoassay. Parasitol Int 2013
Dec;62(6):552-3. doi: 10.1016/j.parint.2013.08.008. Epub 2013 Aug 24.
Breathnach, A.S., McHugh, T.D., Butcher, P.D. (2010) Prevalence and clinical correlations
of genetic subtypes of Giardia lamblia in an urban setting. Epidemiol Infect, 138 (10), 1459-
1467.
-Giardin gene and
development of a polymerase chain reaction-restriction fragment length polymorphism assay
to genotype Giardia duodenalis cysts from human faecal samples. Int J Parasitol 2002; 32:
1023-1030.
Daly, E.R., Roy S.J., Blaney D.D. et al. (2010) Outbreak of giardiasis associated with a
community drinking-water source. Epidemiol Infect, 138 (4), 491-500.
Deshpande, A.P., Jones, B.L., Connelly, L. et al. (2015) Molecular characterization of
Cryptosporidium parvum isolates from human cryptosporidiosis cases in Scotland. Parasitol,
142, 318-325.
Glaberman, S., Moore, J. E., Lowery, C. J., Chalmers, R. M., Sulaiman, I., Elwin, K.,
Rooney, P. J., Millar, B. C., Dooley, J. S., Lal, A. A. and Xiao, L. (2002). Three drinking-
water-associated cryptosporidiosis outbreaks, Northern Ireland. Emerg Infect Dis 8, 631–633.
Goodman, G., Girling, S., Pizzi, R. et al. (2012) Establishment of a health surveillance
program for reintroduction of the Eurasian beaver (Castor fiber) into Scotland. J Wild Dis, 48
(4), 971-978.
Hadfield, S. J., Robinson, G., Elwin, K. and Chalmers, R.M. (2011).

Detection and differentiation of Cryptosporidium spp. in human clinical
samples by use of real-time PCR. J Clin Microbiol 49,
918–924.
Hanevik, K., Dizdar, V., Langeland, N., Hausken, T. (2009) Development of functional
gastrointestinal disorders after Giardia lamblia infection. BMC Gastroenterol, 9, 27.
Insulander, M., Silverlås, C., Lebbad M. et al. (2013) Molecular epidemiology and clinical
manifestations of human cryptosporidiosis in Sweden. Epidemiol Infect. 141(5): 1009-20.
Lange, H., Johansen, O.H., Vold, L. et al. (2014) Second outbreak of infection with a rare
Cryptosporidium parvum genotype in schoolchildren associated with contact with lambs/goat
kids at a holiday farm in Norway. Epidemiol Infect. 142(10): 2105-13.
Lebbad, M., Petersson, I., Karlsson, L. et al. (2011) Multilocus genotyping of human Giardia
isolates suggests limited zoonotic transmission and association between assemblage B and
flatulence in children. PLoS Negl Trop Dis. 5(8) :e1262.
Morch, K., Hanevik, K., Robertson, L.J., Strand, E.A., Langeland, N. (2008) Treatment-
ladder and genetic characterisation of parasites in refractory giardiasis after an outbreak in
Norway. J Infect, 56 (4), 268-273.
Pollock, K.G.J., Smith, H.V., Young, D., Ramsay, C.N., Reilly, W.J. (2005) Giardia
surveillance in Scotland, 1988-2003. Eur J Clin Microbiol Infect Dis, 24 (8), 571-573.
Pollock, K.G.J., Young, D., Smith, H.V., Ramsay, C.N. (2008) Cryptosporidiosis
and filtration of water from Loch Lomond, Scotland. Emerg Infect Dis, 14, 115–20.
Pollock, K.G.J., Ternent, H., Mellor, D.J. et al. (2010) Spatial and Temporal Epidemiology of
Sporadic Human Cryptosporidiosis in Scotland. Zoonoses Pub Health, 57, 487-492.
Putignani, L. and Menichella, D. (2010) Global distribution, public health and clinical impact
of the protozoan pathogen Cryptosporidium. Interdis Perspect Infect Dis, 1–39.

Ramsay CN, Wagner AP, Robertson C, Smith HV, Pollock KG. (2014)
Effects of drinking-water filtration on Cryptosporidium seroepidemiology, Scotland.
Emerg Infect Dis. 20(1): 70-6.
Robertson, L., Gjerde, B., Hansen, E.F., Stachurska-Hagen, T. (2009) A water contamination
incident in Oslo, Norway during October 2007; a basis for discussion of boil-water notices
and the potential for post-treatment contamination of drinking water supplies. J Water
Health, 7 (1), 55-66.
Standards for Microbiology Investigations; Staining Procedures TP 39.
http://www.hpa.org.uk/ProductsServices/MicrobiologyPathology/UK
StandardsForMicrobiologyInvestigations/TermsOfUseForSMIs/AccessTo
UKSMIs/SMITestProcedures/
Steen, K., Damsgaard, E. (2007) [The Giardia epidemic in 2004 and out-of-hours service in
Bergen]. Tidsskr Nor Laegeforen, 127 (2), 187-189.
Strachan, N.J.C., Hunter, C.J., Jones, C.D.R. et al. (2011) The relationship between lay and
technical views of Escherichia coli O157 risk. Philos Trans R Soc Lond B Biol Sci, 366,
1999-2009.
Thabane, M., Marshall, J.K. (2009) Post-infectious irritable bowel syndrome. World J
Gastroenterol, 15, 3591–6.
Widerström, M., Schönning, C., Lilja, M., et al. (2014) Large outbreak of Cryptosporidium
hominis infection transmitted through the public water supply, Sweden. Emerg Infect Dis.
20(4): 581-9