purple - journal of bacteriologyjb.asm.org/content/81/5/688.full.pdf · turned brown when sprayed...
TRANSCRIPT

PATHWAYS FOR BIOSYNTHESIS OF A BACTERIALCAPSULAR POLYSACCHARIDE1
I. CHARACTERIZATION OF THE ORGANISM AND POLYSACCHARIDE
W. HERMAN TAYLOR2 AND ELLIOT JUNI
Department of Microbiology, Enmory University, Atlanta, Geolgia
Received for ptiblication October 24, 1960
There has been no demonstration of netsynthesis of carbohydrate from fats in animaltissues (Weinman, Strisower, and Chaikoff,1957). Many microorganisms, however, are ableto grow with acetate and other fatty acids assole carbon sources and must therefore convertthese compounds to cellular carbohydrate.
This investigation was primarily concernedwith the conversion of two-carbon compoundsinto bacterial polysaccharide capsules. Bacteriawhich grow and produce capsules in a mineralsalts medium supplemented with single carbonsources were isolated from soil. A study hasbeen made of capsule synthesis by one of theseisolates, a gram-negative coccus. This paperreports the characterization of the organism anda study of the composition of the polysaccharidecapsule which it elaborates. A preliminary reportof this work has been presented (Taylor andJuni, 1958).
MATERIALS AND METHODS
The bacterium was isolated from soil on amineral salts medium (S-2) (Monod and Woll-man, 1947) containing 0.5% meso-2,3-butanediolas the sole carbon source. The organism, desig-nated BD-4, was maintained on nutrient agar.
Cell suspensions were spread on the surfacesof mineral agar plates containing 0.5% of thecarbon source and incubated at 30 C for 18 to24 hr. The cells were harvested using a rubberscraper.
Cells and capsules were visualized by spreadingan India ink suspension of organisms on a slide,drying, and then staining with crystal violet
I This work was supported by grant E-1545from the National Institute of Allergy and In-fectious Diseases, U. S. Public Health Service.
2 Present address: Biology Division, Oak RidgeNational Laboratory, Oak Ridge, Tennessee.
(Duguid, 1951). The cells appeared purple Nithcolorless capsules surrounded by a black back-ground. Capsule diameter was estimated inthese stained preparations by using a calibratedocular micrometer and measuring the capsulediameters of 10 cells. The values reported arethe average diameters obtained.
Total counts were made in a Petroff-Hausercounting chamber.
Optical rotation was measured in a 2 dmpolarimeter tube (inside diameter, 4 mm) wvitha Rudolph model no. 70 polarimeter using asodium arc light source.
Melting points were determined in a Thieletube filled with mineral oil and are given un-corrected.Rhamnose and fucose 2, 5-dichlorophenyl-
hydrazones were prepared by the method ofMandl and Neuberg (1952).
Paper chromatograms of sugars were prel)aredon Whatman no. 1 filter paper and developedby descending chromatography at room temper-ature. The following solvent systems were used:n-butanol-ethanol-water (40:10:50), n-butanol-pyridine-water (40:40:20), and aqueous phenol(39.5 ml of water dissolved in 100 g of phenol).The ratios given are volume to volume. Thesugar spots on the developed chromatogramswere located with phloroglucinol reagent (Boren-freund and Dische, 1957), p-anisidine reagent(Hough, Jones, and Wadman, 1950), and am-moniacal silver nitrate reagent (Partridge andWestall, 1948). Amino sugar was tested for with0.2% ninhydrin in 95% ethanol.
Polysaccharide and sugar concentrations weremeasured by the anthrone reagent (Lewin, 1956),methyl pentose by the cysteine-H2SO4 test(Dische and Shettles, 1948), hexose by thecysteine-H2S04 test (Dische, Shettles, and
688
on May 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
BIOSYNTHESIS OF A CAPSULAR POLYSACCHARIDE. I
Osnos, 1949), and glucose by the glucose oxidasereagent (Worthington Biochemical Corporation).
Polysaccharide was prepared from encapsu-lated cells by the following procedures:
Procedure 1.
1) A thick suspension of cells was agitated inan ice-cooled Waring Blendor for 30-min.
2) Cells were centrifuged from the suspension,leaving a viscous, opalescent supernatant.
3) The supernatant was poured into threevolumes of cold acetone and the fibrous pre-cipitate was collected on a stirring rod.
Procedure 2.
1) Cells were suspended in an equal volume of30% KOH and heated at 100 C for 20 min.
2) The resulting digest was poured into severalvolumes of cold acetone or 95% ethanol and thefibrous precipitate was collected.
3) The precipitate was dissolved in water andreprecipitated with five volumes of 95% ethanol.
4) The precipitate was dissolved in water andprotein was removed by a series of 25 Sevagextractions (Sevag, Lackman, and Smolens,1938).
5) The polysaccharide was precipitated withfive volumes of 95%0 ethanol, redissolved inwater, and dialyzed for 12 hr against cold runningwater.
6) The dialyzed polysaccharide was thendried by lyophilization.
Polysaccharide samples were hydrolyzed bydissolving them in 1 N H2SO4 and heating at120 C for 140 min in an autoclave. The smallamount of suspended solid material was re-moved by filtration. The filtrate was neutralizedto pH 7 with solid Ba(OH)2 and the precipitatedBaSO4 was removed by filtration. The filtratewas decolorized with a pinch of acid-washedNorit A (Pfanstiehl Laboratories, Inc.) andfiltered. This colorless filtrate was used forpaper chromatographic analysis. Paper chroma-tograms of some preparations revealed a con-siderable amount of ninhydrin-positive materialwhich was removed by passing the sugar solutionthrough a column containing Duolite C-3(Chemical Process Company) in the hydrogenion cycle and Duolite A-4 in the hydroxyl ioncycle (Sakami, 1955).Rhamnose and glucose in water eluates from
ion exchange columns were separated on a
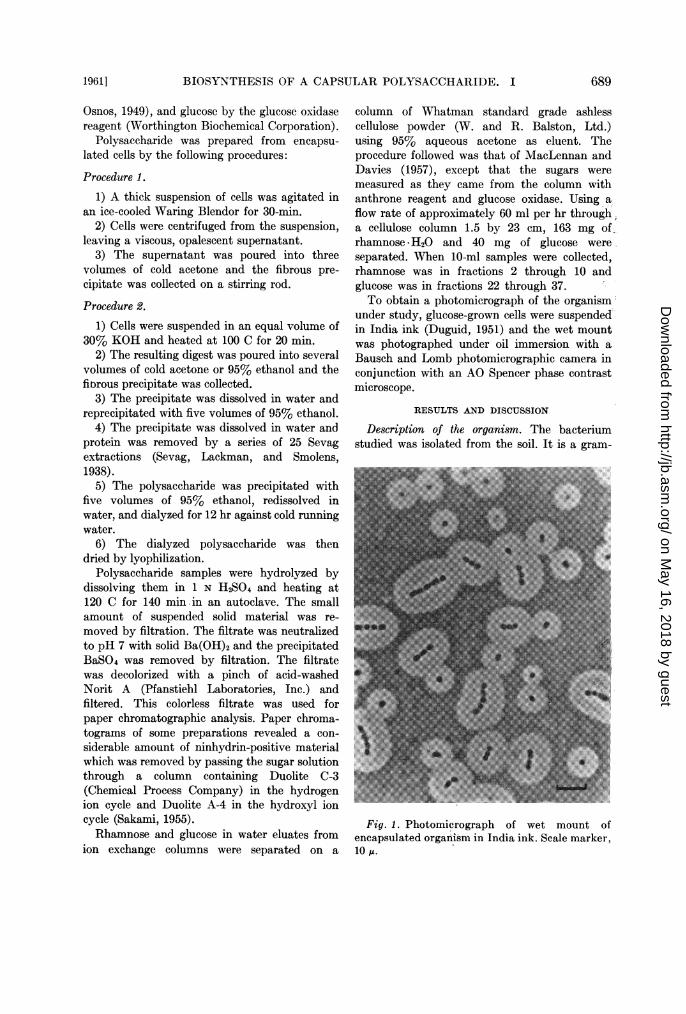
column of Whatman standard grade ashlesscellulose powder (W. and R. Balston, Ltd.)using 95% aqueous acetone as eluent. Theprocedure followed was that of MacLennan andDavies (1957), except that the sugars weremeasured as they came from the column withanthrone reagent and glucose oxidase. Using aflow rate of approximately 60 ml per hr through,a cellulose column 1.5 by 23 cm, 163 mg ofrhamnose * H20 and 40 mg of glucose wereseparated. When 10-ml samples were collected,rhamnose was in fractions 2 through 10 andglucose was in fractions 22 through 37.To obtain a photomicrograph of the organism
under study, glucose-grown cells were suspendedin India ink (Duguid, 1951) and the wet mountwas photographed under oil immersion with aBausch and Lomb photomicrographic camera inconjunction with an AO Spencer phase contrastmicroscope.
RESULTS AND DISCUSSION
Description of the organism. The bacteriumstudied was isolated from the soil. It is a gram-
Fig. 1. Photomicrograph of wet mount ofencapsulated organism in India ink. Scale marker,10 ,u.
19611 689
on May 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TAYLOR AND JUNI
TABLE 1Growth substrates and capsule production
Substrate Avg CapsuleDiameter
Is
Ethanol ........................... 3.3Acetate ............................ 1.6D, L-Lactate ...................... 2.02-Ketoglutarate .................... 1.4D, L-Malate ........... ............. 3.1Succinate .......................... 3.1D, L-1, 2-Propanediola ........ ....... 3.3Meso-2,3-butanediol ............... 3.4D (+)-Glucose ...................... 3.1D (+)-Gluconate .................. 2.2
Growth was tested by inoculating a half-strength mineral-agar plate with a suspension ofthe organism. The carbon source was added togive a final concentration of 0.5%. The followingcompounds were also tested but would not supportgrowth: D(-)-fructose, D(+)-galactose, L(-)-sorbose, L(-)- and D(+)-xylose, D(+)-mannitol,D(+)-sorbitol, D(-)-ribose, D(-)- and L(+)-arabinose, lactose, maltose, sucrose, starch,trehelose, L(+)-rhamnose, L(-)-fucose, D(+)-glucuronate, and methanol.
a Growth occurred only when 0.25% yeastextract was added.
negative, nonmotile, aerobic coccus whichgrows well on a mineral salts medium with singlecarbon sources. Fig. 1 is a photomicrograph of awet mount of the organism suspended in Indiaink (Duguid, 1951). The cells, which appeareither singly, in pairs, or in short chains, arespherical when growing on semisolid media,but are short rods when growing in liquid media.Colonies of the organism are round, opaque,moist, and convex. According to the system ofclassification in Bergey's Manual of Determi-native Bacteriology (Breed, Murray, and Smith,1957), the organism could be classified in thegenus Neisseria. The organism closely resemblesthe one isolated and described by Lemoigne,Girard, and Jacobelli (1952) which was namedNeisseria uwinogradskyi.A quellung reaction was observed with
meningococcus antiserum. The cells clumped inpneumococcus type II antiserum and exhibiteda slight quellung reaction.
Acid is produced on glucose, but no acid orgas is produced from lactose, sucrose, mannitol,
-J
w
z
7--J
wco
-J
r,c00CA
"I)
m
I-
Cs
TIME - HOURS
Fig. 2. Growth and polysaccharide production.A 2.8-liter Fernbach flask containing 500 ml ofS-2 (half-strength) medium and 0.77% ethanolwas inoculated with 15 ml of a 12-hr culture andplaced on a 30 C shaker. Total counts and anthronetests (expressed as glucose) were made at the timesindicated.
maltose, or starch. The oxidase test is negative,no pigment is produced on Loeffler's medium,and no hemolysis is observed on blood agar. Thecatalase test is positive.
Growth and polysaccharide production. Table 1contains a list of substrates tested for ability tosupport growth and capsule production. Of thesimple sugars, disaccharides, and sugar deriv-atives tested, only glucose and gluconate sup-ported growth.
Fig. 2 shows the growth curve and corres-ponding polysaccharide production for thisorganism. After the cell number had reached amaximum, polysaccharide production almostdoubled in the next 14 hr.
Identification of neutral sugar components of thepolysaccharide. Paper chromatography of ahydrolyzate of polysaccharide from glucose-grown cells gave the results listed in Table 2.Sugar A had the same RF as glucose and bothturned brown when sprayed with phloroglucinolor p-anisidine. Sugar B had the same RF as
690 [Vol-. 81
on May 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

BIOSYNTHESIS OF A CAPSULAR POLYSACCHARIDE. I
TABLE 2Paper chromatography of sugars in a hydrolyzate of
polysaccharide from glucose-grown cells
Solvent System
Sugar Butanol- Acetone- Phenoethanol- wActoer waterolwater
wae wtr
RF RF RF
A .............. .. 0.30 0.19 0.41Glucose ............. 0.30 0.19 0.40B ................... 0.48 0.39 0.62Rhamnose ............ 0.47 0.39 0.62
rhamnose, and both spots turned yellow whensprayed with phloroglucinol or p-anisidine. Cap-sular polysaccharide is composed of the samesugars regardless of the carbon source used forgrowth. The same components were present inthe polysaccharide from cells grown on succinate,ethanol, and glucose. The two common methylpentoses, L-rhamnose and L-fucose, are easilydifferentiated from one another by paper chroma-tography.
Identification of the methyl pentose component.The 2,5-dichlorophenylhydrazone of L-rhamnosewas prepared and compared with the corres-ponding derivative of methyl pentose from thepolysaccharide. The melting point of the de-rivative of isolated methyl pentose was 175 to178 C, whereas the melting point of knownL-rhamnose derivative was 176 to 178 C. Amixture of the known and unknown derivativesmelted at 175 to 177 C. The reported meltingpoint for the 2, 5-dichlorophenylhydrazone ofrhamnose is 171 C, whereas the melting pointof the fucose derivative is reported to be 205 C(Mandl and Neuberg, 1952).The cysteine-sulfuric acid test, when carried
out as described by Dische and Shettles (1948),is specific for methyl pentose and gives a colorwith a characteristic absorption spectrum. Ascan be seen in Table 3, the methyl pentosesample reacted in both the anthrone test andthe cysteine-sulfuric acid test, but gave noreaction in the glucose oxidase test. The ab-sorption spectrum of the color produced by theunknown methyl pentose in the cysteine-sulfuricacid test was identical with the absorptionspectrum of the color produced by a rhamnosestandard.The specific rotation of the methyl pentose
TABLE 3Analysis of hexose and methyl pentose separated
from a hydrolyzate of polysaccharide on acellulose column
Total Total Methyl Total GlucoseFractions Anthrone Pentose in in GlucosePooled Value Cysteine-H2S04 Oxidase TestTest
mg mg mg
2-10 163a 154 022-37 40b 0 38
a A rhamnose-H20 standard was used to calcu-late this value.
b A glucose standard was used to calculate thisvalue.
TABLE 4Analysis of a solution of purified polysaccharide
Klett-SummersonUnitsa in
Anthrone Test
1 Ml of polysaccharide solutionb 104Contribution due to 36.5,ug of rham-
nosec............ 75.8Contribution due to 10.3 j,g of glu-
cosed........... 19.6Total units as rhamnose plus glu-
cose. 95.4
aThese units are optical density X 500 as meas-ured in a 16-mm pyrex tube and read in a Klett-Summerson colorimeter with a 540 m,u filter.
bPolysaccharide, 78,g of dry weight per ml.c Determined by the cysteine-H2SO4 test for
methyl pentose (Dische and Shettles, 1948) per-formed directly on the polysaccharide solution.
d Calculated from the ratio of glucose to rham-nose found in a hydrolyzed sample of the poly-saccharide.
isolated from polysaccharide was found to be[a]D + 8.6 (c = 0.358, in H20). The value forthe specific rotation of L-rhamnose is reported tobe [a]D + 8.32 (Hudson and Yanovsky, 1917).
In view of these data, the methyl pentosecomponent of the polysaccharide appears to beL-rhamnose.
Identification of the hexose component. Inaddition to chromatographic identification (Table2), the hexose component was identified asD-glucose by its reactivity with glucose oxidase.As seen in Table 3, the isolated hexose com-
1961 ] 691
on May 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TAYLOR ANI) JUNI
TABLE 5Paper chromatograph?y of a hydrolyzate of purified polysaccharide
Amt ofHydrolyzate Solvent Spray Resul tApplied'
pg
310 Butanol-pyridine-water Ninhydrin No amino sug,ar1,400 Butanol-pyridine-water p-Anisidine Very light spot of uronic
acid-not more than 10
300 Butanol-acetic acid-water Naphthoresoreiinol No uronic acid spot300 Butanol-acetic acid-water Ammoniacal silver nitrate No sugar alcohol spot
a The amount of hydrolyzate applied is listed as micrograms of rhamnose in the hydrolyzate asmeasured by the cysteine-H2SO4 test.
ponent reacted equally in the glucose oxidaseand anthrone tests. The glucose oxidase enzymehas been shown to be specific for D-glucose(Keilin and Hartree, 1948, 1952).
Analysis of a purified preparation of poly-saccharide. To determine in greater detail thechemical composition of the polysaccharide itwas necessary to obtain a relatively pure prepa-ration. This was accomplished by use of pro-cedure 2, which includes a series of Sevag extrac-tions. Table 4 contains the analysis of a solutionof purified polysaccharide by use of the anthrone,glucose oxidase, and cysteine-sulfuric acid tests,demonstrating that 91.8% of the anthrone colorgiven by the solid can be attributed to thecontent of rhamnose and glucose; however, only60% of the dry weight of the polysaceharidepreparation is rhamnose and glucose. Controlexperiments demonstrated that glucose andrhamnose mixtures reacted in an additive mannerin the anthrone test. It seemed likely that theremay have been some major component of thepolysaceharide which had not been detected. Aseries of paper chromatograms were preparedto search for components other than rhamnoseand glucose. The results of these experiments aregiven in Table 5. It can be seen that no aminosugars or sugar alcohols could be detected. Ifuronic acid was present, it was only a minorcomponent. No pentose component has beendetected in hydrolyzates by use of any sprayreagent. It is important to note that no majorspots other than rhamnose and glucose weredetected using the ammoniacal silver nitratespray reagent. This spray reagent will detectnonreducing compounds such as sugar alcohols.It is concluded that D-glucose and L-rhamnose
are the only major organic components of thepolysaccharide. The lpurified preparation mayhave contained water in a bound form or someinorganic salts, which would not have beendetected.
SUMMARY
A gram-negative coccus, which forms largepolysaccharide cal)sules during growth on amineral salts medium containing single carbonsources, was isolated from the soil. The capsularpolysaceharide produced by this organism wasisolated, purified, and hydrolyzed. The com-ponent sugars of the polysaceharide were identi-fied as D-glucose and L-rhamnose. No aminosugar, sugar alcohol, uronic acid, or pentosecomponent could be detected.
REFERENCES
BORENFREUND, E., AND Z. DISCHE 1957 A new
spray for spotting of sugars on paper chroma-tograms. Arch. Biochem. Biophys., 67,239-240.
BREED, R. S., E. G. D. MURRAY, AND N. R. SMITH1957 Bergey's manual of determinative bac-teriology, 7th ed. The Williams & WilkinsCo., Baltimore.
DISCHE, Z., AND L. B. SHETTLES 1948 A specificcolor reaction of methylpentoses and a spec-trophotometric micromethod for their deter-mination. J. Biol. Chem., 175, 595-603.
DISCHE, Z., L. B. SHETTLES, AND M. OSNOS 1949New color reactions of hexoses and spectro-photometric micromethods for their determi-nation. Arch. Biochem., 22, 169-184.
DUGUID, J. P. 1951 The demonstration ofbacterial capsules and slime. J. Pathol.Bacteriol., 63, 673-685.
692 [VOL. 81
on May 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

BIOSYNTHESIS OF A CAPSULAR POLYSACCHARIDE. I
HOUGH, L., J. K. N. JONES, AND W. H. WADMAN1950 Quantitative analysis of mixtures ofsugars by the method of partition chroma-tography. V. Improved methods for theseparation and detection of the sugars andtheir methylated derivatives on the paperchromatogram. J. Chem. Soc., 1950, 1702-1706.
HUDSON, C. S., AND E. YANOVSKY 1917 Indirectmeasurements of the rotary powers of somea- and ,8-forms of the sugars by means ofsolubility experiments. J. Am. Chem. Soc.,39, 1013-1038.
KEILIN, D., AND E. F. HARTREE 1948 Propertiesof glucose oxidase (notation). Biochem. J.,42, 221-229.
KEILIN, D., AND E. F. HARTREE 1952 Specificityof glucose oxidase (notation). Biochem. J.,50, 331-341.
LEMOIGNE, M., H. GIRARD, AND G. JACOBELLI1952 Bacterie du soil utilisant facilement le2-3 butanediol. Ann. inst. Pasteur, 82,389-398.
LEWIN, R. A. 1956 Extracellular polysaccha-rides of green algae. Cani. J. Microbiol., 2,665-672.
MAcLENNAN, A. P., AND D. A. L. DAVIES 1957The isolation of D-glycero-D-galactoheptoseand other sugar components from the specificpolysaccharide of Chromobacterium violaceum(BN). Biochem. J., 66, 562-567.
Y 1] I I., AND C. NEUBERG 1952 Characteriza-tion of some sugars of interest to biochem-istry. Arch. Biochem. Biophys., 35, 326-334.
MONOD, J., AND E. WOLLMAN 1947 L'inhibitionde la croissance de l'adaptation enzymatiqueches les bacteries infectees par le bacterio-phage. Ann. inst. Pasteur, 73, 937-956.
PARTRIDGE, S. M., AND R. G. WESTALL 1948Filter-paper partition chromatography ofsugars. I. General description and applicationto the qualitative analysis of sugars in applejuice, egg white and foetal blood of sheep.Biochem. J., 42, 238-250.
SAKAMI, W. (Editor) 1955 Handbook of isotopetracer methods, p. 163. Western ReserveUniversity, Cleveland.
SEVAG, M. G., D. B. LACKMAN, AND J. SMOLENS1938 The isolation of the components ofstreptococcal nucleoproteins in serologicallyactive form. J. Biol. Chem., 124,425-436.
TAYLOR, W. H., AND E. JUNI 1958 Synthesis ofL-rhamnose from ethanol by an aerobicbacterium. Bacteriol. Proc., 1958, 125.
WEINMAN, E. O., E. H. STRISOWER, AND I. L.CHAIKOFF 1957 Conversion of fatty acidsto carbohydrate: application of isotopes tothis problem and the role of the Krebs cycleas a synthetic pathway. Physiol. Rev., 37,252-272.
69319611
on May 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from