radiotherapy (eagle & foley, 1956; pet- - europe pubmed central
TRANSCRIPT

Br. J. Cancer (1981) 44, 775
DO CELL LINES IN VITRO REFLECT THE PROPERTIES OF THETUMOURS OF ORIGIN? A STUDY OF LINES DERIVED FROM
HUMAN MELANOMA XENOGRAFTS
K. M. TVEIT AND A. PIHL
From. Norsk Hydro's Institute for Cancer Research and The Norwegian Cancer Society,Montebello, Oslo 3, Norway
Received 28 AIay 1981 Accepte(1 2 September 1981
Summary.-The characteristics of 7 human melanoma cell lines were comparedwith those of the xenografts from which they were established. The ultrastructure,melanin content, isozyme pattern and chromosome numbers of the cell lines wereclosely similar to those of the corresponding xenografts. The different cell lines gaverise to colonies in soft agar of size and morphology similar to the parent xenografts,and the plating efficiencies were clearly correlated. However, no correlation wasfound between the growth rates in vivo and either the doubling times and saturationdensities in monolayer cultures or the plating efficiencies in soft agar. Moreover, oneof the cell lines lost its tumorigenic ability upon establishment in culture. Thus,although the properties of the cell lines by and large reflected those of the parentxenografts, important inconsistencies were seen. The data emphasize that extra-polations from continuous cell lines in vitro to tumour cells in vivo are not neces-sarily valid.A high content of cellular fibronectin was correlated with a compact colony
morphology in soft agar and rapid attachment and spreading on plastic. The growthrates and cellular morphology of the cell lines were strongly influenced by TPA,DMSO, retinoic acid and theophylline, but not by a-melanocyte-stimulating hormone.A murine cell line established from one of the xenografts grew in soft agar and
produced sarcomas in mice. The malignant murine cells had arisen by transforma-tion of murine stromal cells during the first subcultures in vitro, possibly caused by afactor produced by the human melanoma cells.
Human tumour cell lines growing con-tinuously in tissue culture have beenwidely used as models in studies of variousaspects of tumour biology and chemo- andradiotherapy (Eagle & Foley, 1956; Pet-tersen et al., 1974; Fogh, 1975; Drewinkoet al., 1976). Such cell lines possess severaladvantages from an experimental pointof view; the cells can be grown in largequantities, and can be stored and studiedrepeatedly under defined experimentalconditions. However, continuous cell linesalso possess obvious limitations as modelsof tumours in situ. Factors which mayinfluence the biological behaviour of amalignant tumour in a patient, such asthe immunological response, the vascular-
53
ization of the tumour and the presence ofstromal cells cannot be studied in cellscultivated in vitro. Moreover, the possi-bility must be considered that during theestablishment of the permanent cell lineschanges may occur in their biologicalproperties. Few detailed comparisons havebeen made between such cell lines andtheir tumours of origin, and it is not clearwhether the cell lines indeed reflect thecharacteristics of the malignant cell popu-lations in vivo.Whether or not continuous cell lines
reflect the properties of the tumour cellsin situ is difficult to study using patients'tumours directly. We have therefore usedhuman tumours serially transplanted in

K. M. TVEIT AND A. PIHL
athymic (nude) mice. We have establishedcontinuous cell lines from the xenograftsand studied to what extent they reflectthe properties of the parent tumours. Wehave compared the morphology, chromo-some constitution, isozyme patterns,growth rates and colony formation in softagar of the cell lines with the same para-meters in the xenografts.
MATERIALS AND METHODS
Xenografting in athymic mice.-The originof the xenografts and the procedure forheterotransplantation in athymic (nude) micehas previously been described (Fodstad et al.,1980). Cells grown in monolayer cultureswere harvested by scraping, and 5 x 106-107cells were inoculated s.c. into athymic miceof BALB/c origin.
Histology, electron microscopy, chromosomeand isozyme analyses.-The methods usedare described in previous papers (Tveit et al.,1980a,b).
In vitro cultivation.-Xenografts measuring8-12 mm in diameter were minced andseeded into culture flasks as previouslydescribed (Tveit et al., 1980b). RPMI 1640medium with 25mM HEPES (Gibco Biocult,Glasgow) supplemented with 15% foetal calfserum (Gibco Biocult), 100 iu/ml penicillin and100 ,tg/ml streptomycin, was routinely used.In certain experiments, where the melanincontent in cells was measured, Dulbecco'smodified medium (Gibco Biocult) was used.Subculturing was carried out twice a week,using a 0.05% trypsin/0 02% EDTA solution.The cell lines were tested for mycoplasma asdescribed below, and were all negative.Growth curves of cultured cells were
obtained by seeding 2 x 105 cells into 25cm2culture flasks and counting every second day.Medium was changed on Days 4, 6 and 8.Treatment of cells with 10-7M 12-0-tetra-decanoyl phorbol-13-acetate (TPA), 1.5%dimethyl sulphoxide (DMSO), 10-5M all-transretinoic acid and 1mM theophylline was per-formed as previously described (Tveit et al.,1980b). Treatment with o-melanocyte-stimu-lating hormone (a-MSH) was carried out byadding 2 x 10-7M ax-MSH (Sigma ChemicalCo., St Louis, U.S.A.) to 25cm2 flasks with2 x 105 cells. The cultured cells were re-fedwith medium supplemented with x-MSH onDays 4, 6 and 8. The melanin content was
measured by the method described byWhittaker (1963). The rapidity of attachmentand spreading on a plastic surface wasexamined after seeding out trypsinized cellsinto wells of culture plates immediately,after 4 h or 24 h incubation in medium withor without serum.
Cultivation of tumour cells in soft agar wasperformed as described by Courtenay & Mills(1978). Single-cell suspensions, both fromsolid tumours growing in athymic mice andfrom cells growing in monolayer cultures,were prepared by trypsin/EDTA treatment.After washing in serum-containing medium,an appropriate number of cells was seeded outin soft agar cultures in triplicate. Colonies ofmore than 50 cells were scored after 2 weeks'incubation. In order to establish tumour celllines free from normal cells the agar withcolonies was pipetted vigorously and trans-ferred to culture flasks. When cells andcolonies had settled, serum-containing mediumwas added and further cultivation was per-formed in monolayer cultures.
Staining with Hoechst 33258.-This fluores-cence staining technique was used both foridentifying human and murine cells (Moseret al., 1975) and for mycoplasma screening.Cells grown on coverslips were fixed inmethanol: glacial acetic acid = 3:1, treatedwith fluorochrome Hoechst 33258 (CalbiochemAG, Lucerne, Switzerland) at a concentrationof 1 ,tg/ml for 2-3 min, washed in phosphatebuffered saline (PBS), mounted in glycerol/PBS and examined in a Zeiss fluorescenceinvertoscope.
Immunofluorescence.-Cells growing expo-nentially on coverslips were fixed in 3%paraformaldehyde and washed in PBS. Fordetermination of intracellular fibronectin thecells were subsequently treated for 20 minwith Nonidet P40 detergent according toLaurila et al. (1978). The detergent treatmentwas omitted when surface-associated fibro-nectin was determined. The cells were washedx 5 and treated for 30 min with rabbit anti-human-fibronectin serum (a gift from Dr A.Vaheri, Helsinki, Finland), diluted 1:100,followed by washing and further incubationfor 30 min with swine anti-rabbit serum IgG,conjugated with fluorescein-isothiocyanate(Daco-Immunoglobulins, Copenhagen, Den-mark), diluted 1:100. All incubations wereat room temperature. The cells were washed,mounted and examined in the fluorescencemicroscope.
776

XENOGRAFT-DERIVED HUMAN MELANOMA CELL LINES
Examination of cellular proteins.-Lacto-peroxidase-catalysed iodination (1251) of sur-face proteins was performed on exponentiallygrowing cells in 100mm Petri dishes (106cells/dish). Each dish was incubated with5mM glucose (Sigma Chemical Co.), 1 ulactoperoxidase (Sigma Chemical Co.) 06 uglucose oxidase (Worthington BiochemicalCorp., New Jersey, U.S.A.) and 0-2 mCi Na1251 (The Radiochemical Centre, Amersham,Bucks) in 4 ml PBS for 20 min at roomtemperature. The dishes were washed oncewith 015M Nal in 10mM Na phosphate(pH 7.4) and in PBS x 3. One ml samplebuffer (3% SDS and 1% 2-mercaptoethanolin 01M Tris-HCl, pH 6 8) was added to thePetri dish and the cells were scraped off.
Slab-gel electrophoresis used a 4.5% stack-ing gel (4.5% acrylamide and 0-1% SDS inTris-HCl, pH 6.8) and a 7% separation gel(7% acrylamide and 0.1% SDS in Tris-HCl,pH 8 8). The gels were dried, and auto-radiography was performed at -70°C usingKodak X-Omat Film.
RESULTS
Establishment of continuous cell linesIn this study we have established con-
tinuous cell lines from human melanomaxenografts by two different procedures:by cultivation directly in monolayers orfrom colonies in soft agar. Interestingly,the 4 xenografts showed differences intheir behaviour.When tumour fragments and dispersed
cells were seeded out in monolayers, cul-tures from xenograft GE were usuallyovergrown by normal murine cells in the2nd-4th subculture. However, in oneinstance, when the melanoma cells grewmore rapidly than usual in the primaryculture, the murine cells disappeared anda continuous melanoma cell line, GEM,developed. The notation for the xenograftsand the derived cell lines is apparent fromTable I. Xenograft EF immediately gaverise to the cell line EFM, which has beendescribed previously (Tveit et al., 1980b).This line grew rapidly right from the start,and no contamination with murine cellswas seen. In contrast, primary culturesfrom xenograft EE were regularly over-
TABLE I.-Chromosome numbers in humanmelanoma xenografts and in cell linesestablished from them
Cell line establishedin vitro
Xeno-graftGE
EF
Directly
GEM-6tGEM
EFM-6tEFM
EE
VNVNM-8tVNM
Chromosomenumber*
A
Via softagar Modal
434746
GEMS-6t 46GEMS 43
585454
EFMS-6t 53EFMS 52
57EEMS-5t 55
827980
VNMS-6t 74VNMS 74
Range29-5827-7925-7622-7922-5737-7425-9318-10331-8726-8929-8426-8955-10360-10152-9839-10241-97
* 50 or 100 metaphases analysed.t Number of the subculture studied. The number
was omitted when the cell lines had been subeulturedmore than 70 times.
grown by murine fibroblasts, and no cellline could be established.The behaviour of xenograft VN was
particularly interesting. When fragmentsand single cells from this xenograft weregrown in culture flasks, in most cases twocell populations were found growing to-gether. The two populations were identi-fied as human tumour cells and abnormalmurine cells by chromosome and isozymeanalyses, as well as by Hoechst staining.Both populations were usually present toabout the 30th subculture. Thereafter, in2 experiments a human cell line emerged,one of which, VNM, was further studied.Usually the abnormal murine cells tookover and gave rise to continuous murinecell lines with a transformed phenotype.This cell line will be discussed below.When colonies formed in soft agar were
further grown in monolayers, the xeno-grafts gave rise to cell lines (denoted byadding S to the name), which were per-manent, except EEMS, which could not besubcultured for more than 6 months.
777

K. M. TVEIT AND A. PIHL
FIG. 1. Histological sections of human melanoma xerlografts. H. & E. x 400. (a) EF, (b) VN, (c) GE.FIG. 2. Photomicrographs of cells in tissue culture. Phase contrast. x 160. (a) EFM, (b) VNM, (C) GEM.FIG. 3.-Colonies in soft agar (after 2 weeks' incubation). x 160. (a) EMF, (b) VNM, (C) GEM.
Characterization of melanoma cell lines andcomparison with the xenografts of originMorphology and pigmentation.-Three
of the melanoma xenografts were closelysimilar in histology, whereas in xenograftVN the cells contained less cytoplasmand had nuclei of a more uniform size(Fig. 1). The derived melanoma cell linesall showed individual and characteristiccytological appearance and growth pat-
tern (Fig. 2). The EFM and EFMS cellswere triangular or bipolar, grew in irregularpatterns and piled up when grown to highdensity. The VNM cells were bipolar,often arranged in a regular parallel fashionand a variable number of living cellsfloated free in the medium. Interestingly,the VNMS cells, established by a differentmethod from the same xenografted mela-noma, showed a more triangular, cuboidal
778

XENOGRAFT-DERIVED HUMAN MELANOMA CELL LINES
and irregular appearance. In the remainingcell lines (G8EM, GEMS and EEMS)cuboidal cells were most prominent.The xenograft EF and the derived cell
lines EFM and EFMS showed pigmenta-tion, detectable by microscopic inspectionof stained histological sections or mono-
layer cultures, and electron microscopyrevealed melanosomes. In none of theother xenografts or cell lines was thereevidence of melanin. Prolonged treatmentof the permanent cell lines EFM, VNMSand GEMS with 2 x 10-7M o-MSH hadno effect on melanin production, growthrate or cellular morphology, in agreementwith the finding that; most human mela-noma lines so far studied did not respondto this hormone (Fuller & Meyskens,1979; Lotan & Lotan, 1980). However,when treated with TPA, DMSO, retinoicacid and theophylline, our cells changedmorphology and developed elongated den-(Iritic processes. No melanin appeared inthe melanin-free cell lines AVNMS and(XEMS, though we have previously shownincreased production of melanin in the cellline EFM when treated with these agents(Tveit et al., 1980b).
Chromnosome and isozyme studies. Themelanoma xenografts showed dissimilarchromosome numbers (Table I). Thus,xenograft GE was near-diploid, xeno-
grafts EF and EE were hyperdiploid andxenograft VN was hypotetraploid. Thein vitro cell lines had chromosome numberssimilar to the xenografts from whichthey were derived (Table I) though therewas a tendency for a slight downwarddrift for 3 xenografts. In consecutive sub-cultures the chromosome numbers re-
mained stable, except in the case of theGEMS line, where a slight downward driftcontinued.
Also, when the lactate dehydrogenase(LDH) isozyme pattern was studied,individual differences were found betweenthe xenografts. Thus, in three of thexenografts (EF, GE and EE) Bands 3 and4 were the most heavily stained, indicatingnearly equal amounts of A and B poly-peptide chains (not shown). In contrast,
in xenograft VN, Bands 4 and 5 were themost heavily stained, corresponding tohigher amounts of A than B chains. Againthe cell lines in vitro showed similarpatterns to the xenografts from whichthey were derived.
Growth characteristics.-Growth curvesof 5 cell lines in vitro, established from 3of the xenografts, are shown in Fig. 4.It is seen that the cell line EFM showedthe highest growth rate, and that in thetwo cases where two cell lines were ob-tained from the same xenograft thesegrew at essentially the same rate. InTable II the doubling times in culture ofall cell lines are given, together with thetumour volume-doubling times (Td) pre-viously observed in vivo during exponen-tial growth (Fodstad et al., 1980). It canbe seen that the growth rates of the celllines are not correlated with those of thexenografts. Thus, the most fast-growingcell line in vitro, EFM (doubling time 18 h),which also showed the highest saturationdensity, was established from a xenograftwith an intermediate growth rate in vivo(Td 6 3 days). Three more slow growing celllines'originated from two xenografts whichhad considerably highergrowth rates in vivo
Ln~ ~~Dy in ulur
50xA
-~~~~~~~ ~~0
line 0tvir.Smos:O F;C
20 -
10 -
--
o 2 A
2 4 6 8 10
Days in cultureFiG,. 4.-Growth curves for 5 melanoma cell
lines in vitro. Symbols: 0, EFM; 0~,VNM; U, VNMIS; A, GEMS; A, GEMI.
779

K. AM. TVEIT ANI) A. PIHL
TABLE II.-Growth characteristics of human melanoma xenografts in vivo and of thecorresponding cell lines in vitro*
Doubling timI
XenograftEF
VN
GE
EE
Cell line
EFM-6EFMEFMS-6EFMS
VNAM-8VNMVNAIS-6VNMS
GEM-6GEMGEMS-6GEMS
EES-5
In vivot(days)
6-3 (577-70)
3-3 (2'9-3.4)
9-5 (9 0-9-8)
3 0 (2 9-3.0)
SatutrationIt vitro (lenlsity
(h1) (cells X 10-4/Cm2)
32 (29-36)18 (17-21)42 (38-46)25 (23-27)
37 (36-38)30 (26-32)33 (31-35)29 (26-32)
42 (42)39 (36-42)39 (36-42)31 (28-36)
42 (42)
12 (10-14)
11 (11)
3.5 (3.0-4 0)
5-5 (466-64)
3-2 (2-5-4-0)
4-6 (42-5.0)
5-0 (3-5-6 5)
PEin soft agarl
12 (5-21)27 (18-35)53 (35-78)28 (20-36)47 (35-65)0 7 (0 5-1-5)1-0 (0-5-1-5)5-0 (3-5-6.5)I- t(0.6-1(6)3 2 (2.0-4 8)0-5 (0.2-1 0)0 9 (0-3-1-2)1-1 (0-5-1-5)1-0 (0-4-1-5)2-0 (0 9-2 5)
10 (5-20)12 (6-15)
* Averages of 2-5 experiments (ranges in parentheses).t Tumour volume-doubling time (volume = n/6 x (mean (liameter):3).t Number of colonies (> 50 cells)/Number of cells platedI x 100.
than EF. Interestingly, it was found thatthe amount of cellular fibronectin wasinversely related to the growth rate invivo. Treatment of the cell lines EFM,VNMS and GEMS with the differentiatingagents TPA, DMSO, retinoic acid andtheophylline always decreased the growthrates (Table III), though individual dif-ferences in response were observed.The plating efficiencies (PE) of the dif-
ferent xenografts and the derived celllines differed widely (Table II). The PEof the cell lines were clearly correlatedwith and consistently higher than thePE of the corresponding xenografts. Inthe cell lines the PE in soft agar wasgenerally highest in the lines showing lowdoubling times, whereas in the xeno-grafts there was no correlation betweenPE and doubling times.
Colony morphology.-The morphologyof the colonies in soft agar was charac-teristic for each particular xenograft, andthis morphology was maintained in thein vitro cell lines. Thus, the xenograft VNand the human cell lines established fromit gave rise to colonies of very looselyattached cells (Fig. 3), while the xenograft
GE and the cell lines derived froml- itgave compact small colonies (Fig. 3).Xenograft EF and the corresponding celllines EFM and EFMS gave rise to coloniiesof an intermediate morphology (Fig. 3).The results indicate that the cohesionbetween the cells differed in soft-agarcolonies from different melanomas.To study in more detail the adhesive
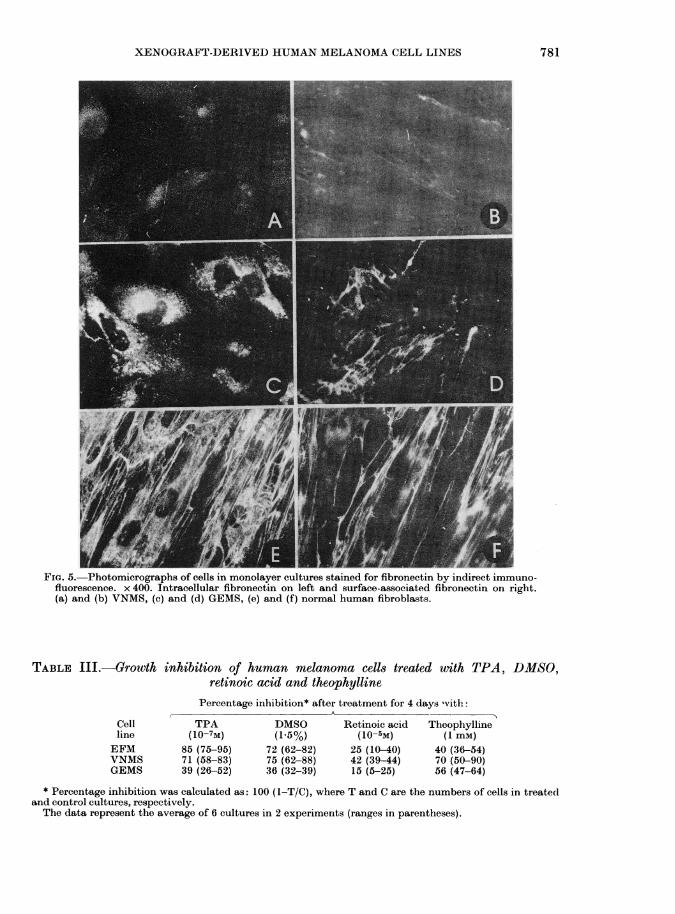
properties of the cells, their attachment toplastic surface and their protein contentwere examined. The rate of attachment toand spreading on plastic surface werestudied in 3 cell lines showing differentcolony morphology. The GEMS cell line,which had the most compact small col-onies, attached and spread out as fast asnormal human fibroblasts, whereas theEFM cells, and particularly the VNMScells, which formed colonies of looselyattached cells, needed more time for bothattachment and spreading. All 3 cell linescontained surface-associated and intra-cellular fibronectin, as revealed by immu-nofluorescence. The rapidly spreading cellline GEMS had about as much fibronectinas human fibroblasts, while EFM cells, andespecially the slow-spreading VNMS cells,
780
XENOGRAFT-DERIVED HUMAN MELANOMA CELL LINES
Lwf!
FIG. 5. Photomicrographs of cells in monolayer cultures stained for fibronectin by indirect immuno-fluorescence. x 400. Intracellular fibronectin on left and surface-associated fibronectin on right.(a) and (b) VNMS, (c) and (d) GEMS, (e) and (f) normal human fibroblasts.
TABLE III.-Growth inhibition of human melanoma cell8 treated with TPA, DMSO,retinoic acid and theophylline
Percentage inhibition* after treatment for 4 days 'vith
Cell TPA DMSO Retinoic acid Theophyllineline (10-7M) (1.5%) (10-5M) (1 mM)EFM 85 (75-95) 72 (62-82) 25 (10-40) 40 (36-54)VNMS 71 (58-83) 75 (62-88) 42 (39-44) 70 (50-90)GEMS 39 (26-52) 36 (32-39) 15 (5-25) 56 (47-64)
* Percentage inhibition was calculated as: 100 (1-T/C), where T and C are the numbers of cells in treatedand control cultures, respectively.The data represent the average of 6 cultures in 2 experiments (ranges in parentheses).
781
7K. M1. TVEiTrr ANI) A. PIHL
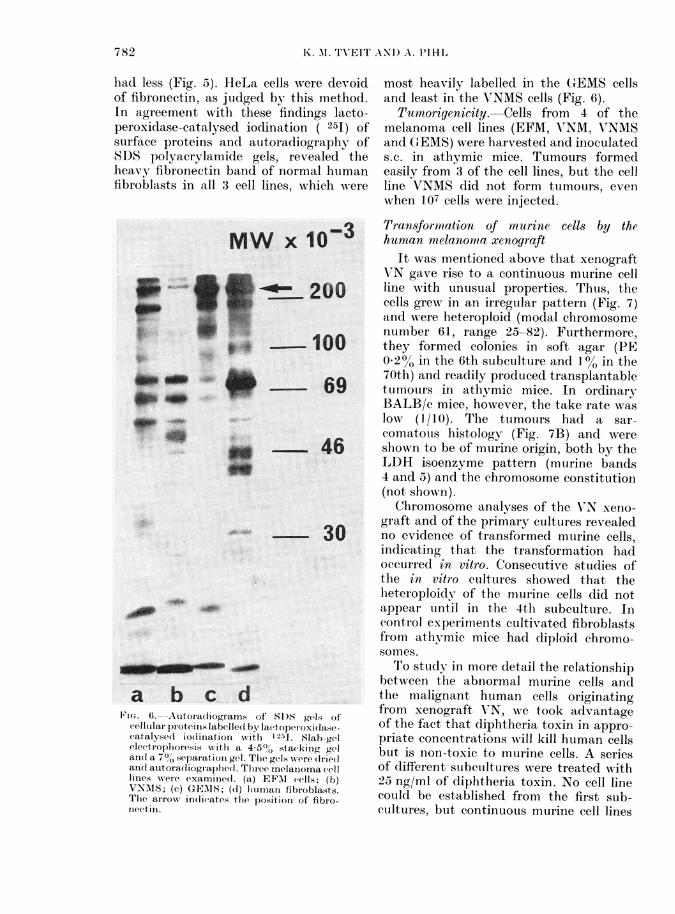
had less (Fig. 5). HeLa cells were devoidof fibronectin, as judged by this method.In agreement with these findings lacto-peroxidase-catalysed ioclination ( 251) ofsurface proteins and autoradiography ofkSDS polyacrylamide gels, revealed theheavy fibronectin band of normal humanfibroblasts in all 3 cell lines, which wAere
....aFIG. 6.-Autoradiograms of SD)S gels of
cellutlai proteiis labellel byr lacttopetroxidasecatalysed io(liniation xwith 12-41. Slab-gelelectroplioresis with a 4-5% stacking gelan(l a 700 separation gel. The gelss were (Irie(lanlel autoradliogiaphed. Tlhree melanioma celllinies were examined. (a) EFAI cells; (b)VNMIS; (c) GEMS; (d) hliumatn fibiroblasts.The aIIow indicates tihe position of fibro-nectin.
most heavily labelled in the G-EMS cellsand least in the V7NMS cells (Fig. 6).
Tumoriyenicity. Cells from 4 of themelanoma cell lines (EFM, AVNM, A'NMSand GEMS) were harvested and inoculateds.c. in athymic mice. Tumours formedeasily from 3 of the cell lines, but the cellline VNMS did not form tumours, evenwhen 107 cells were injected.
Transformation of murine cells by thehuman melanomta xenograft
It was mentioned above that xenograftVN gave rise to a continuous murine cellline with unusual properties. Thus, thecells grew in an irregular pattern (Fig. 7)and were heteroploid (modal chromosomenumber 61, range 25-82). Furthermore,they formed colonies in soft agar (PE0.2% in the 6th subculture and 1% in the70th) and readily produced transplantabletumours in athymic mice. In ordinaryBALB/c mice, however, the take rate waslow (I/10). The tumours had a sar-comatous histology (Fig. 7B) and wereshown to be of murine origin, both by theLDH isoenzyme pattern (murine bands4 and 5) and the chromosome constitution(not shoxvn).Chromosome analyses of the VrN xeno-
graft and of the primary cultures revealedno evidence of transformed murine cells,indicating that the transformation hadoccurred in vitro. Consecutive studies ofthe in vitr-o cutltures showed that theheteroploidv of the murine cells did notcappear until in the 4th subculture. Incontrol experiments cultivated fibroblastsfrom athymic mice had diploid chromo-somes.To study in more detail the relationship
between the abnormnal murine cells andthe malignant human cells originatingfrom xenograft VN, we took advantageof the fact that diphtheria toxin in appro-priate concentrations will kill human cellsbut is non-toxic to murine cells. A seriesof different subcultures were treated with25 ng/ml of diphtheria toxin. No cell linecould be established from the first sub-cultures, but continuous murine cell lines
782

XENOGRAFT-DERIVED HUMAN MELANOMA CELL LINES
Fim. 7.-Photomicrographs of the malignant murine cell line established from xenograft VN (a) Cellsin monolayer culture. Phase contrast x 160. (b) Histological section of tumour formed in athymicmice. H. & E. x 400.
did emerge in the 4th and later subcul-tures. These murine cells showed the samemalignant characteristics as the spon-taneous murine cell lines. It thus appearsthat the transformation of the murinecells occurred during the early subcultures
and that somehow the presence of thehuman cells was necessary for the trans-formation.
DISCUSSIONContinuous human cell lines have been
established from a variety of malignancies,
783

K. M. TVEIT AND A. PIHL
including primary and metastatic malig-nant melanomas (Romsdahl & Hsu, 1972;Gerner et al., 1975; G-iovanella et al., 1976).It is difficult to tell whether these cell linesreflect the properties of the tumour cellsin vivo by direct comparison with thepatients' tumours. However, evidence isaccumulating that human xenograftsgrown in immune-deprived animals largelyreflect the properties of the parenttumours, though kinetic differences havebeen observed (Steel & Peckham, 1980).Since such serially heterotransplantedtumours represent a bank of easily avail-able tumour tissue and permit repeatedstudies of tumour cells from the samepatient, they greatly facilitate comparisonsbetween the properties of established invitro cell lines and the tumour cells oforigin.The present results show that, as expec-
ted, the melanoma cell lines, like the xeno-grafts from which they were derived, havedistinct individual properties. The com-parisons of the cell lines with the corre-sponding xenografts demonstrate that theproperties of the cell lines largely reflectthose of the tumours of origin. Thus, theultrastructural picture and the melaninpigmentation were the same in vitro andin vivo, in agreement with the findings ofFoa & Aubert (1977). Also the isozymepatterns of the xenografts were retainedin the cultured cell lines. The coloniesformed in soft agar from the xenograftsand from the derived cell lines were closelysimilar in respect of size and compactness.Likewise, the chromosome constitutionwas, by and large, retained on cultivation,though a slight downward drift in chromo-some number was found during theestablishment of some of the cell lines.However, some of the in vivo character-
istics were not retained. Importantly, thegrowth rates and the saturation densitiesof the cell lines in monolayer cultures, aswell as the PE in soft agar, showed nocorrelation with the early growth rates ofthe melanomas in vivo. The reason for thisdiscrepancy is not clear. One possibility isthat the in vitro conditions influence the
growth of tumour cells from differenttumours to varying extents. Unexpec-tedly, one of the cell lines, VNMS, estab-lished via soft agar, did not form tumourswhen injected back into athymic mice.Apparently, the genetic information neces-sary for tumorigenicity had been lost orsuppressed during the establishment inculture. It is noteworthy that in this cellline the chemosensitivity did not reflectthat of the parent xenograft, in contrastto the situation with the other cell lines(Tveit et al., 1981). The data emphasizethat extrapolations from continuous celllines in tissue culture to the parent tumourcells in vivo are not always valid.
It is a general view that fibronectinplays a central role in cell adhesion (Hynes,1979; Kleinman et al., 1981). The presentfinding that the amounts of fibronectincorrelated with the compactness of thecolonies formed in soft agar and therapidity of attachment and spreading on aplastic surface, indicates that fibronectinmay be responsible for the differentdegrees of compactness seen in soft-agar colonies. It has been speculatedwhether the amount of fibronectin on thesurface of tumour cells correlates to anyin vivo characteristics; e.g. stage of disease,growth rate or metastatic properties(Lloyd et al., 1979). Our study shows thatin the 3 tumour cell lines studied theamount of fibronectin indeed correlatedwith the growth rates in vivo, as the slow-est growing xenograft (GE) gave rise toa cell line in vitro with high amounts offibronectin, whilst the xenograft with theshortest doubling time (VN) gave rise tomelanoma cells with least fibronectin.A difficulty in establishing cell lines
from human tumours is that most fre-quently stromal fibroblasts grow alongwith the malignant cell population andtend to overgrow the culture. To eliminatethese normal cells several methods havebeen used, such as differential centrifuga-tion, differential trypsinization, physicalremoval of colonies and the use of specialculture media (Fogh, 1975). Here we haveintroduced an additional procedure for the
784

XENOGRAFT-DERIVED HUMAN MELANOMA CELL LINES 785
elimination of fibroblasts. We have takenadvantage of the fact that, in solidtumours, only malginant cells can formcolonies in semi-solid medium. Thus, wehave cultivated in monolayers coloniesformed in soft agar. This procedure forestablishing cell lines free of normal cellscan also be applied directly to patients'biopsies, and we have obtained severalpure malignant cell populations frompatients' melanomas.We have reported (Tveit et al., 1980a)
tha-t whnen a hurnan embryonal carcinomaxenograft was grown in tissue culture, atransformed murine cell line appeared.This abnormal heteroploid murine cell linegrew in soft agar, but failed to formtumours in athymic mice. In contrast, themurine cell line which appeared in thepresent study upon cultivation of themelanoma xenograft VN, was clearlymalignant as it gave sarcomas on injectioninto mice. Goldenberg and Pavia havereported that when human xenografts of3 different histological types were grownin culture, malignant murine cells con-sistently emerged, capable of formingsarcomas when injected into nude mice(Pavia & Goldenberg, 1979; Goldenberg &Pavia, 1981). On this basis they suggestedthat such transformation may be a generalphenomenon when xenografts are culturedin vitro. Our results do not support thisview, as only one of the 4 melanoma xeno-grafts studied gave rise to transformedmurine cells.
It is apparent from this work that themalignant transformation of murine cellstook place during the first subcultures. Itis unlikely that spontaneous transforma-tion can account for the transformedmurine cells here observed, since they werefound in all experiments involving xeno-grafts VN and never when other melanomaxenografts or skin fibroblasts from nudemice were grown in vitro. Furthermore,when the human cells were killed withdiphtheria toxin, it was concluded thatthe presence of human cells during the first4 subcultures was necessary for the estab-lishment of the malignant murine cell
lines. The data suggest that the malignanttransformation of murine stromal cells iscaused by a transforming factor producedby the human melanoma cells during thefirst weeks of in vitro cultivation. This wassupported by the finding that mediumfrom the permanent melanoma cell lineVNM, established from the same xeno-graft, consistently stimulated the growthof nude mice fibroblasts in monolayercultures, far more than did medium fromcontrol cells. Moreover, in two instancesmurine fibroblasts grown in this condi-tioned medium formed colonies in softagar (PE 0.2%). However, further workis clearly needed to elucidate the mechan-ism of this transformation.
The authors are indebted to Drs 0. Fodstad andS. Olsnes for stimulating discussions, to Dr A.Sundan for help with the electrophoreses and toL. Ness for skilful technical assistance.
REFERENCESCOURTENAY, V. D. & MILLS, J. (1978) An in vitro
colony assay for human tumours grown in immune-suppressed mice and treated in vivo with cytotoxicagents. Br. J. Cancer, 37, 261.
DREWINKO, B., Loo, T. L., BROWN, B., GOTTLIEB,J. A. & FREIREICH, E. J. (1976) Combinationchemotherapy in vitro with Adriamycin: Observa-tions of additive, antagonistic, and synergisticeffects when used in two-drug combinations oncultured human lymphoma cells. Cancer Biochem.Biophys., 1, 187.
EAGLE, H. & FOLEY, G. E. (1956) The cytotoxicaction of carcinolytic agents in tissue culture.Am. J. Med., 21, 739.
FOA, C. & AUBERT, C. (1977) Ultrastructural com-parison between cultured and tumor cells ofhuman malignant melanoma. Cancer Res., 37,3957.
FODSTAD, 0., AASS, N. & PIHL, A. (1980) Assessmentof tumour growth and of response to chemo-therapy of human melanomas in athymic, nudemice. Br. J. Cancer, 41, Suppl. IV, 146.
FoGH, J. (1975) Human Tumor Cells in vitro. NewYork: Plenum Press.
FULLER, B. B. & MEYSKENS, F. L. (1979) Endocrineresponsiveness of normal and malignant humanmelanocytes in culture. Proc. Am. Assoc. CancerRes., 20, 93.
GERNER, R. E., KITAMURA, H. & MOORE, G. E.(1975) Studies of tumor cell lines derived frompatients with malignant melanoma. Oncology, 31,31.
GIOVANELLA, B. C., STEHLIN, J. S., SANTAMARIA, C.& 6 others (1976) Human neoplastic and normalcells in tissue culture. I. Cell lines derived frommalignant melanomas and normal melanocytes.J. Natl Cancer Inst., 56, 1131.
GOLDENBERG, D. M. & PAVIA, R. A. (1981) Malig-

786 K. M. TVEIT AND A. PIHL
nant potential of murine stromal cells after trans-plantation of human tumors into nude mice.Science, 212, 65.
HYNES, R. 0. (1979) Proteins and glycoproteins. InSurfaces of Normal and Malignant Cells. Ed.Hynes. New York: John Wiley & Sons. p. 103.
KLEINMAN, H. K., KLEBE, R. J. & MARTIN, G. R.(1981) Role of collagenous matrices in the adhesionand growth of cells. J. Cell Biol., 88, 473.
LAURILA, P., VIRTANEN, I., WARTIOVAARA, J. &STENMAN, S. (1978) Fluorescent antibodies andlectins stain intracellular structures in fixed cellstreated with nonionic detergent. J. Histochem.Cytochem., 26, 251.
LLOYD, K. O., TRAVASSOS, L. R., TAKAHASHI, T. &OLD, L. J. (1979) Cell surface glycoproteins ofhuman tumor cell lines: Unusual characteristicsof malignant melanoma. J. Natl Cancer Inst., 63,623.
LOTAN, R. & LOTAN, D. (1980) Stimulation ofmelano-genesis in a human melanoma cell line byretinoids. Cancer Res., 40, 3345.
MOSER, F. G., DORMAN, B. P. & RUDDLE, F. H.(1975) Mouse-human heterokarvon analysis witha 33258 Hoechst-Giemsa techinique. J. Cell Biol.,66, 676.
PAVIA, R. A. & GOLDENBERG, D. M. (1979) Malig-nant transformation of murine fibroblasts propa-
gated from a human cervical squamous cancerxenografted in nude mice. In Vitro, 15, 227.
PETTERSEN, E. O., OFTEBRO, R. & BRUSTAD, T.(1974) Survival after X-irradiation of extremelyhypoxic human cells cultured in vitro. Int. J.Radiat. Biol., 26, 305.
ROMSDAHL, M. M. & Hsu, T. C. (1972) Establish-ment and characterization of human malignantmelanoma cell lines grown in vitro. In Pigmenta-tion. Its Genesis and Biologic Control. Ed. Riley.New York: Appleton-Century-Crofts. p. 461.
STEEL, G. G. & PECKHAM, M. J. (1980) Humantumour xenografts: A critical appraisal. Br. J.Cancer, 41, Suppl. IV, 133.
TVEIT, K. M., FODSTAD, 0., BROGGER, A. & OLSNES,S. (1 980a) Human embryonal carcinoma grownin athymic mice and in vitro. Cancer Res., 40, 949.
TVEIT, K. M., FODSTAD, 0., JOHANNESSEN, J. V. &OLSNES, S. (1980b) A human melanoma cell lineestablished from xenograft in athymic mice.Br. J. Cancer, 41, 724.
TVEIT, K. M., FODSTAD, 0. & PIHL, A. (1981) Theusefulness of human tumour cell lines in the studyof chemosensitivity. A study of malignant mela-nomas. Int. J. Cancer, 28, 403.
WHITTAKER, J. R. (1963) Changes in melanogenesisduring the dedifferentiation of chick retinal pig-ment cells in cell culture. Dev. Biol., 8, 99.