read full text pdf - journal of biological chemistry a gift of prof. h. clevers (university of...
TRANSCRIPT

Ets Gene PEA3 Cooperates with �-Catenin-Lef-1 and c-Jun inRegulation of Osteopontin Transcription*
Received for publication, October 9, 2003, and in revised form, February 15, 2004Published, JBC Papers in Press, February 27, 2004, DOI 10.1074/jbc.M311131200
Mohamed El-Tanani‡§, Angela Platt-Higgins¶, Philip S. Rudland¶,and Frederick Charles Campbell‡
From the ‡Department of Surgery, Cancer Research Centre, Queen’s University of Belfast, Grosvenor Road,Belfast BT12 6BJ and the ¶Cancer and Polio Research Laboratories, School of Biological Sciences,University of Liverpool, Liverpool L69 7ZB, United Kingdom
Osteopontin (OPN) is a multifunctional protein impli-cated in mammary development, neoplastic change, andmetastasis. OPN is a target gene for �-catenin-T cellfactor signaling, which is commonly disturbed duringmammary oncogenesis, but the understanding of OPNregulation is incomplete. Data base-assisted bioinfor-matic analysis of the OPN promoter region has revealedthe presence of T cell factor-, Ets-, and AP-1-bindingmotifs. Here we report that �-catenin, Lef-1, Ets tran-scription factors, and the AP-1 protein c-Jun eachweakly enhanced luciferase expression from a OPN pro-moter-luciferase reporter construct, transiently trans-fected into a rat mammary cell line. OPN promoter re-sponsiveness to �-catenin and Lef-1, however, wasconsiderably enhanced by Ets transcription factors in-cluding Ets-1, Ets-2, ERM, and particularly PEA3. PEA3also enhanced promoter responsiveness to the AP-1 pro-tein c-Jun. Co-transfection of cells with �-catenin, Lef-1,PEA3, and c-Jun in combination increased luciferaseexpression by up to 280-fold and induced expression ofendogenous rat OPN. In six human breast cell lines,those that highly expressed OPN also expressed PEA3and Ets-1. Moreover, there was a significant associationof immunocytochemical staining for OPN and one of�-catenin, Ets-1, Ets-2, PEA3, or c-Jun, in the 29 humanbreast carcinomas tested. This study shows that �-cate-nin/Lef-1, Ets, and AP-1 transcription factors can coop-erate in a rat mammary cell line in stimulating tran-scription of OPN and that their independent presence isassociated with that of OPN in a group of human breastcancers. These results suggest that the presence of thesetranscription factors in human breast cancer is respon-sible in part for the overexpression of OPN that, in turn,is implicated in mammary neoplastic progression andmetastasis.
Osteopontin (OPN)1 is an acidic hydrophilic glycophospho-protein that binds to cell surface integrins and may activate
growth factor receptors (1). OPN transcription may be acti-vated by the Ras oncogene (2) and plays a key role in neoplastictransformation, metastasis (3), and cancer progression (4).OPN is usually absent or expressed at a low level in normaltissues but is up-regulated in certain preneoplastic and neo-plastic epithelia (4–7), including that of the breast (8). Trans-fection of an expression vector for OPN induces malignanttransformation and induction of metastasis in a benign ratmammary epithelial cell line (9), whereas transfection of OPNantisense cDNA inhibits these processes in a cell line alreadyoverexpressing osteopontin (10, 11). These results suggest thatOPN overexpression may represent a key molecular event intumor progression and metastasis, particularly that of thebreast. Unlike many proto-oncogenes activated by a gain offunction mutation, OPN is not typically mutated during step-wise tumorigenesis (3). Instead various responsive elements inits promoter regulate OPN expression for its diverse physiolog-ical roles (12–14), and it is presumably these elements thatallow the overexpression of OPN in certain cancers.
�-Catenin is a component of the Wnt signal transductionpathway, implicated inter alia in initiation and progression ofbreast cancer (15, 16). �-Catenin translocates to the nucleus,where it binds to the Tcf/Lef family to initiate transcription ofresponsive genes (17–19). �-Catenin and OPN overexpressionmay coincide (20), and activated �-catenin may induce OPNexpression in migrating cells (21). OPN may be a transcrip-tional target of the �-catenin-Tcf complex (22). Lef/Tcf factorsare sequence-specific DNA-binding proteins that have a singlehigh mobility group domain, located in the middle or near theirC terminus (23). The ability of Lef-1 to regulate transcriptionmay involve its association with different DNA-binding pro-teins (24–26). The OPN promoter also contains inter alia, AP-1(27), and Ets transcription factor-binding domains (12). Mu-tagenesis of AP-1 or Ets sites separately impede OPN tran-scription in several cell lines and reporter-promoter systems,suggesting their involvement at this promoter (27). In a database-assisted bioinformatics analysis, we identified three Lef-1/Tcf-binding sites (CAAAG) in addition to one AP-1-bindingdomain (TGAGTCA) and three Ets-binding motifs (AGGAAR)within the rat OPN promoter. Although Ets proteins bind toDNA recognition sites bearing a 5�-AGGA(A/T)-3� central core,flanking sequences influence their binding specificity (28).AP-1 and Ets proteins participate in transcriptional regulationof metastasis-associated matrix metalloproteinases (29) andsynergize with �-catenin-Lef-1 in regulation of tumor associ-ated matrilysin (30). Activation of transcription factors AP-1and members of the Ets family may therefore represent keyevents in cell transformation (31). To test the hypothesis that�-catenin-Lef-1 synergizes with AP-1 and Ets in regulation ofOPN expression, we have transfected Rama 37 rat mammary
* This work was supported by grants from Action Cancer (NorthernIreland), the Royal Victoria Hospital Research Fund (Belfast), theNorth West Research Fund, and the Cancer and Polio Research FundLtd. The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
§ To whom correspondence should be addressed. Tel.: 44-28-90-632528; Fax: 44-28-321811; E-mail: [email protected].
1 The abbreviations used are: OPN, osteopontin; Tcf, T cell factor;Lef-1, lymphoid enhancer factor; AP-1, activation protein 1; MAb,monoclonal antibody; Rama, rat mammary; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 20, Issue of May 14, pp. 20794–20806, 2004© 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org20794
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

epithelial cells with an OPN promoter-luciferase reporter con-struct and cDNA expression vectors for �-catenin, Lef-1, theAP-1 protein c-Jun, and for the Ets family of transcriptionfactors, alone and in combination. PEA3 (polyomavirus en-hancer activator protein 3), an Ets family member, renderedthe OPN promoter strongly responsive to transactivation by�-catenin/Lef-1 or c-Jun. Ets family members PEA3 and Ets-1were overexpressed in human breast cancer cell lines thathighly expressed OPN. Moreover, Ets family members as wellas c-Jun and �-catenin were highly expressed in the primarybreast tumors that highly expressed OPN. We conclude that�-catenin, AP-1, and Ets pathways can synergize to provoketranscriptional up-regulation of OPN expression in a culturedrat mammary cell line. The co-incident expression of �-catenin,AP-1, Ets family members, and OPN in breast cancer cell linesand primary tumors suggest that they may be implicated in thedysregulation of OPN in breast cancer.
EXPERIMENTAL PROCEDURES
Plasmids and Oligonucleotides—Expression vectors for human Ets-1(32), Ets-2 (33), and mouse PEA3 (34) within the expression vectorspcDNA3, pCMV5, and pSG5, respectively, were gifts of Dr. Ejii Hara(Paterson Institute for Cancer Research, Manchester, UK), with thepermission of Prof. J. M. Leiden (Harvard University), Dr. C. A. Hauser(Scripps Research Institute), and Dr. Y. de Launoit (Pasteur Institute,Lille, France). Expression vectors for human c-Jun (35) and ERM (30)within the expression vector pcDNA3 were purchased from ATCC (Ma-nassas, VA). The expression vectors for human Lef-1 (36) in pcDNA3were a gift of Prof. H. Clevers (University of Utrecht, Utrecht, Holland),and a stable mutant form of mutant �-catenin that lacks the N-terminaldomain (37) in vector pCI-neo (Promega, Madison, WI) was a gift ofProf. B. Vogelstein (Johns Hopkins Oncology Center, Baltimore, MD).The synthetic 20-mer double-stranded oligonucleotides containing thewild type Lef-1-binding sites with sequences GGG TTA CAA AGA GTCCTG G and AGA CGA TTC AAA GAC GTT A or a mutant Lef-1-bindingsite with the sequence AGA TCG ACC GCG GTA CGT TA were pro-duced on an automated DNA synthesizer by Invitrogen. A 2.3-kilobasepair fragment of the 6-kilobase pair rat OPN promoter (38) was ampli-fied by PCR and then coupled to a firefly luciferase reporter construct,as described previously (22, 39).
Mutagenesis of the Osteopontin Promoter—The OPN promoter fireflyluciferase reporter constructs with mutated Lef-1-, AP-1-, and Ets-binding sites were generated with the QuikChange site-directed mu-tagenesis kit (Stratagene, La Jolla, CA). A mutant of one Lef-1-bindingsite was generated at position �1954 to �1960 (Mut �1960) using CACTCA GGG GTC CGC GGT AGT CCT GGA AGG GTC and GAC CCTTCC AGG ACT ACC GCG GAC CCC TGA GTG forward and reverseprimers, respectively. The start of the 2285-bp promoter fragment isdefined as �1, the TATA box is in positions �28 to �23, and the startof the primary transcript from the OPN gene is at �1. A double mutantin two Lef-1-binding sites was generated at positions �1457 to �1463 inaddition to the mutated site above (Mut �1960/�1463), using GATGGC TAG TAC CCG CGG TGT TTG ACT TAA TTC and GGA TTA AGTCAA ACC CCG CGG GTA CTA GCC ATC as forward and reverseprimers, respectively. A triple mutant in three Lef-1-binding sites wasgenerated at positions �1115 to �1121 in addition to the two mutatedsites above (Mut �1960/�1463/�1121) using CTT AAA GAT CGA CCGCGG TAC CTT ACA AAT C and GAT TTG TAA CGT ACC GCG GTCGAT CTT TAA G as forward and reverse primers, respectively. Amutation of the single AP-1-binding site was generated at positions�1872 to �1866 (Mut �1872) using TAT ACC TCC ATA ATT CGT GTCGAG TCG TTC CTG TGG GCT CAG GG and CCC TGA GCC CAC AGGAAC GAC TCG ACA CGA ATT ATG GAG GTA TA as forward andreverse primers, respectively. For the Ets-binding sites, a mutation wasgenerated at position �2198 to �2194 (Mut �2198), using forward andreverse primers GTC AGT GTA TGA AGC AGT CAG TCC TGT CGAand TCG ACA GGA CTG ACT GCT TCA TAC ACT CAC, respectively.A mutation of the second Ets-binding site was separately generated atposition �1361 to �1357 (Mut �1361) using forward and reverse prim-ers CAG GTA ATT GAA GAA AGG AAG TAA TTG CAG and CTG CAATTA CTT CCT TTC TTC AAT TAC CTG, respectively. A mutation of thethird Ets-binding site was separately generated at position �1215 to�1211 (mut-1215) using forward and reverse primers AAC CTT ATATAT TCT ATG AAA TAA AAC TCA and TGA GTT TTA TTT CAT AGAATA TAT AAG GTT, respectively.
Cell Lines and Transient Transfections—Rama 37, ZR-75, T47-D,MDA-MB-231, Huma 7, Huma 109, and Huma 123 cell lines werecultured in Dulbecco’s modified Eagle’s medium, 10% (v/v) fetal calfserum, 100 �g/ml penicillin, 100 �g/ml streptomycin (Invitrogen). Thecells were harvested and seeded in multiwell plates at 2.5 � 105 cells/3.5-cm-diameter well in 1 ml of serum-free medium. After 24 h, the cellswere co-transfected with the predetermined optimal amounts of thefollowing, where indicated: 200 ng of Ets-1, Ets-2, ERM, and PEA3 intheir respective expression vectors; 300 ng of Lef-1 and �-catenin ex-pression vectors; and 1 �g of wild type or mutant OPN promoter fireflyluciferase reporter constructs. The assays for the firefly luciferase-linked promoter also contained a control expression vector of 5 ng ofpRL Renilla luciferase (Promega). The concentration of pRL Renillaluciferase was reduced to avoid interference with the firefly luciferaseassay, in accordance with the manufacturer’s instructions. The cellswere incubated for a further 24 h and harvested in 300 �l of ReporterLysis Buffer (Promega), and firefly luciferase and control Renilla lucif-erase were simultaneously assayed, as described in the dual luciferasereporter assay system (Promega) and reported previously (40). Fireflyluciferase activity was normalized to Renilla luciferase activity. Maxi-mum activity was reached by 48 h, and the results at 72 h were similar.
Gel Shift Assays—Gel shift assays were performed as described pre-viously (22) by incubating 0.5 ng of 32P-labeled double-stranded 20-meroligonucleotides containing one wild type Lef-1-binding site (specificactivity �5 � 108 cpm/�g), either GGG GTT ACA AAG AGT CCT GGand AGA TCG ATT CAA AGA CGT TA or one mutated Lef-1-bindingsite, AGA TCG ACC GCG GTA CGT TA (specific activity, �5 � 108
cpm/�g) with 8 �l of protein extract from a reticulocyte cell-free tran-scription-translation protein synthesizing lysate for 1 h at 20 °C in 30 �lof 0.03 M KCl, 1 mM Na2HPO4, 0.01 M HEPES (pH 7.9), 0.25 mM
dithiothreitol, 10% (w/v) glycerol, 1.4 �g of poly(dI-dC) (AmershamBiosciences), 1 �g of single-stranded salmon sperm DNA, 0.01% (w/v)SDS for 40 min at 0 °C. The samples were electrophoresed throughnondenaturing 4% (w/v) polyacrylamide gels, which were dried, exposedto Fuji X-Omat film for 18 h with an intensifying screen, and processedfor autoradiography. The reticulocyte lysate was incubated with theexpression vectors for Lef-1 or with buffer alone in a cell-free protein-synthesizing transcription-translation system to generate the relevantprotein, as described previously (40).
Northern Blotting—Complementary DNA probes were generated bydigestion of OPN, Ets-1, or PEA3 expression vectors with EcoRI/KpnI,Hind III/NotI, or EcoRI restriction enzymes, respectively. Fifty ng ofeach purified cDNA probe was radioactively labeled using a randomprimed DNA labeling kit (Roche Applied Science) to a specific activity of5 � 108 dpm/�g. Total cellular RNA was isolated from the relevant celllines using the RNeasy kit (Qiagen), and 10 �g of total RNA was run ona 1% (w/v) agarose denaturing formaldehyde gel. Nucleic acids wereblotted on to Hybond N� membrane (Amersham Biosciences) by capil-lary transfer in 10� SSC buffer (1� SSC in 0.15 M NaCl plus 0.015 M
sodium citrate). The probe was hybridized to a blot in UltraHyb buffer(Amersham Biosciences), which was washed to high stringency. Thewashed membrane was exposed to x-ray film (Fuji) for 3 days at �70 °C,using an intensifying screen. The resultant autoradiographs were an-alyzed by densitometry using a CS-9000 dual wavelength Flying Spotscanner (Shimadzu, Tokyo, Japan). The OPN, Ets-1, and PEA3 densi-tometer readings were corrected for differences in the amounts of RNAloaded in each lane by normalizing them relative to the GAPDH mRNAreading for each lane (41, 42).
Immunocytochemical Analysis—Formalin-fixed, paraffin-embeddedspecimens from 29 patients with breast cancer were randomly selectedfrom the group described previously (43), cut serially into 4-�m sec-tions, and mounted on three aminopropyltriethoxy silane-coated slides.After staining of representative sections with hematoxylin and eosin,all of the specimens proved to be invasive breast carcinoma of no specialtype. For subsequent immunocytochemical staining for �-catenin, PEA,and c-Jun, the sections were subjected to antigen retrieval by immers-ing them in 0.01 M sodium citrate (pH 6.0) and heating in a domesticmicrowave oven at 850 watts for 15 min. The slides were allowed to coolfor a further 15 min in citrate buffer (44). The sections for the use of allantibodies were then incubated with 0.05% (w/v) H2O2 in methanol toinhibit endogenous peroxidase activity (45).
Mouse MAb MBIII B10 (46) raised against rat OPN was purchasedfrom the Developmental Studies Hybridoma Bank, Department of Bi-ological Sciences, University of Iowa (Iowa City, IA). It recognizedhuman, bovine, and rat OPN in Western blots and immunocytochem-istry, as described previously (45). MAbs to �-catenin (E5, sc-7963) andTcf-4 (6H5–3, T5817) were purchased from Santa Cruz Biotechnology(Santa Cruz, CA) and Sigma, respectively and have been characterized
Ets, �-Catenin, and c-Jun Regulate Osteopontin 20795
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

previously (22, 39). Rabbit polyclonal antibodies to Ets-1 (C-20, sc-350),Ets-2 (C-20, sc-351), c-Jun (9162), and MAb to PEA3 (sc-113) werepurchased from Santa Cruz Biotechnology for three items and fromNew England Biolabs for c-Jun. All of these antibodies recognized thecorrect size antigens on Western blots of SDS-polyacrylamide gels.Blocking antigens for OPN, �-catenin, Tcf-4, Ets-1, Ets-2, PEA3, andc-Jun, respectively, were human recombinant OPN (cc-1074), blockingpeptide to �-catenin (sc-1496P), blocking peptide to Tcf-4 (sc-8631P),blocking peptide to Ets-1 (sc-350P), blocking peptide to Ets-2 (sc-351P),general blocking peptide for Ets family (sc-112P), and reticulocyte cell-free transcription translation protein synthesizing lysate primed withexpression vector for PEA3 as described earlier under “ExperimentalProcedures” (40), and c-Jun fusion protein (6093). These items werepurchased from Chemicon Europe (Chandlers Ford, UK) for OPN; fromSanta Cruz Biotechnology Inc. (AutogenBioclear, Calne, Swindon, UK)for �-catenin, Tcf-4, Ets-1, Ets-2, and Ets family (PEA3); and from NewEngland Biolabs (Hitchen, UK) for c-Jun.
Indirect immunocytochemistry was carried out using slightly differ-ent procedures of detection for different primary antibodies with acommercially available antibody biotin complex containing horseradishperoxidase, as described previously (45). The antibodies to �-catenin,Tcf-4, Ets-1, Ets-2, PEA3, c-Jun, and OPN were diluted to 1:20, 1:100,1:500, 1:400, 1:10, 1:50, and 1:30, respectively. The sections were incu-bated at room temperature for 3 h for antibodies to �-catenin, Ets-1,Ets-2, and Tcf-4; for 16 h overnight at room temperature for anti-OPN;and 16 h overnight at 4 °C for antibodies to PEA3 and c-Jun. The boundantibodies were detected with a StreptABComplex horseradish per-oxidase Duet Mouse/Rabbit kit (Dako Ltd., Cambridge, UK) for anti-bodies to �-catenin, Ets-1, Ets-2, PEA3, and c-Jun and with a DAKOEnVisionTM � System (Dako Ltd.) (47) for anti-Tcf-4 or with 1:200diluted biotinylated sheep anti-mouse Ig (Amersham Biosciences) for1 h followed by a commercially available ABComplex made according to
the manufacturer’s instructions for antibodies to OPN (Dako Ltd.). Thebound complexes were visualized with 3,31-diaminobenzidine (Merck)and 0.003% (v/v) H2O2. The cellular nuclei were lightly counterstainedblue with Mayers’ hemalum and the sections mounted in DPX (MerckLtd.). For blocking purposes, the primary antibodies were incubatedwith 100–200 �g/ml of blocking antigens or a 50% mixture of reticulo-cyte cell-free protein-synthesizing lysate for 3 h at 37 °C in phosphatebuffered saline (pH 7.4), 0.5% (v/v) bovine serum albumin and storedovernight at 4 °C prior to use. The slides from the 29 specimens werestained separately for all seven antibodies and analyzed independentlyby two observers using light microscopy. The specimens were recordedas staining positively for each antibody if �5% of the carcinoma cells/field were well stained, and the remaining specimens were classified asnegatively stained; the results from two sections of each specimen with10 fields/section at �200 magnification were recorded, as describedpreviously (8). Increasing the concentration of antibodies 5-fold gaveidentical results in sections selected at random. Retrieval of antigens(44) increased the level of staining for �-catenin, PEA3, and c-Jun butnot for the other antibodies used. However, it failed to alter the ob-served subcellular distribution of any of the antigens under study.Photographs were recorded on a Reichert Polyvar microscope fittedwith a Wratten 44 blue green filter (45). Testing for significance of theassociation of the staining for OPN and for each transcription factor inturn was undertaken using Fisher’s Exact test; the two-sided values ofp were given.
RESULTS
Assay of Lef-1 Protein Binding to Tcf Motifs within the OPNPromoter—The rat OPN promoter bears three consensus Tcf-binding sites (5�-(A/T)(A/T)CAAAG-3�) between nucleotides�1960 and �1954 (TACAAAG), between nucleotides �1463
FIG. 1. A, structure of the rat osteopontin promoter. Note the positions of the three consensus Tcf-binding sites, three Ets-binding sites, and oneAP-1-binding site relative to the transcription start site (GenBankTM accession number AF017274). B, electrophoretic mobility shift assay for Lef-1binding to oligonucleotides containing Tcf-binding sites. The synthetic double-strand 20-mer oligonucleotides containing the wild type first orsecond Tcf-binding sites (Wt�1960 and Wt�1463) were incubated with protein-synthesizing lysates programmed with expression vectors for Lef-1or for Lef-1 and �-catenin (�-cat). The oligonucleotide with wild type Tcf site �1960 was incubated with unprogrammed lysate (Lysate). Also shownis a 20-mer oligonucleotide containing a mutated Tcf site (mut �1960) co-incubated with the same amount of lysate programmed by expressionvector for Lef-1.
Ets, �-Catenin, and c-Jun Regulate Osteopontin20796
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

and �1457 (AACAAAG), and between nucleotides �1121 and�1115 (TTCAAAG). Nucleotide �1 fragment is at the start ofthe 2285-bp OPN promoter. The negative numbers representnucleotides increasing from this point, read in a 3� direction.The primary transcript from the gene starts at nucleotide �1(Fig. 1A). Lef-1 affinity for these binding sites with or without�-catenin was investigated in an electromobility shift assay.Lef-1 and �-catenin proteins synthesized by a reticulocyte cell-free transcription-translation-coupled protein synthesizingsystem (“Experimental Procedures”) were incubated with a20-mer oligonucleotide containing the sequence TACAAAG orAACAAAG. These oligonucleotides have the same sequence asthe first two Tcf-binding sites present in the OPN promoter(Fig. 1B). A mutant 20-mer oligonucleotide was also generatedfrom the second Tcf-binding site bearing mutations known todiminish Tcf binding (CCGCGGT). The addition of Lef-1 pro-tein lysate produced a slower running band than that caused bythe radioactive oligonucleotide alone for either of the oligonu-cleotides containing the Tcf-binding sites. This result is con-sistent with the formation of a DNA-protein complex. Lef-1protein lysates failed to produce a similar slower running bandwith the oligonucleotide containing the mutant Tcf-bindingsite, even at the highest protein lysate concentrations tested(Fig. 1B). The addition of �-catenin lysates produced a furtherslower running band for either of the oligonucleotides contain-ing the Tcf-binding sites, the presence of which was dependenton the mixtures containing Lef-1 lysates (data not shown).These results are consistent with the formation of a ternarycomplex between the two proteins and either of the two 20-meroligonucleotides containing the Tcf-binding site.
AP-1 and Ets Transactivation of the OPN Promoter—TheOPN promoter has a canonical AP-1 site (5�-TGAGTCA-3�)located between nucleotides �1872 and �1866 and candidateEts-binding sites (5�-AGGA(A/T)-3�) located from nucleotides�2198 to �2194, �1361 to �1357, and �1215 to �1211 (Fig.
1A). To investigate the possibility of AP-1-mediated OPN trans-activation, we co-transfected the Rama 37 cells with the 2.3-kilobase pair OPN promoter-luciferase reporter construct(OPN-Luc) and an expression vector for c-Jun, which whensynthesized is capable of dimerizing to initiate AP-1 transcrip-tion factor activity (see “Experimental Procedures”). Luciferasereporter activity was enhanced by �6-fold by the addition ofc-Jun lysates (Fig. 2). Previously determined optimal stimulat-ing doses of mammalian expression vectors for Ets-1, Ets-2,PEA3, and its subfamily gene, ERM (200 ng/reaction) wereco-transfected with OPN-Luc separately and in combinationwith the expression vector for c-Jun. Ets-1 and Ets-2 lysatesenhanced OPN-promoter luciferase-reporter activity by ap-proximately 5-fold and also further enhanced c-Jun-mediatedactivity approximately 2-fold (Fig. 2). The lysates containingPEA3 or ERM appeared more potent and enhanced OPN pro-moter-reporter activity by 7–8-fold, and they synergized withlysates containing c-Jun to enhance this activity by a further5–6-fold. These latter combinations therefore caused a 38–44-fold increase above the activity of the OPN promoter-reporteralone (Fig. 2). These findings indicate a certain degree of pref-erence of the PEA3 and ERM subclass of the Ets transcriptionfactor family for the OPN promoter either alone or in combi-nation with the AP-1 co-activator c-Jun.
Co-activation of �-Catenin-Lef-1-mediated OPN PromoterActivity by c-Jun or PEA3—Rama 37 cells were co-transfectedwith the expression vectors for c-Jun and for Lef-1 and/or�-catenin together with the OPN promoter reporter construct(OPN-Luc). Lysates containing �-catenin or Lef-1 were rela-tively weak individual or combined inducers of OPN promoterreporter activity. The combination of lysates increased the pro-moter activity by 9-fold (Fig. 3A). Co-transfection of the expres-sion vector for c-Jun had only a relatively weak enhancingeffect on �-catenin/Lef-1-mediated OPN promoter activity ofapproximately 1.3-fold, increasing the overall stimulation to
FIG. 2. Effect of Ets transcription factors on the osteopontin promoter with and without c-Jun. A total of 1.5 � 105 Rama 37 cells wereco-transfected with the rat OPN-promoter luciferase-reporter construct and protein-synthesizing lysates primed by expression vectors for variousEts transcription factors, as indicated. The output of the OPN promoter was measured by the activity of the firefly luciferase reporter gene at 24 h(“Experimental Procedures”). The data are presented as fold induction of the OPN promoter-reporter activity relative to that for co-transfection ofthe promoter constructs with empty expression vectors. The values obtained were normalized by co-transfection of the same cells with constitu-tively active simian virus 40-driven Renilla luciferase. The data bars represent the means of experiments repeated a minimum of three times, andeach transfection was performed in triplicate. The error bars represent the standard errors.
Ets, �-Catenin, and c-Jun Regulate Osteopontin 20797
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12-fold (Fig. 3). Co-expression of PEA3 with either Lef-1 or�-catenin increased the OPN promoter reporter activity byapproximately 8-fold, whereas expression of PEA3, Lef-1, and�-catenin increased the overall activity by 18-fold, indicatingthat PEA3, �-catenin, and Lef-1 were strong co-activators ofthe OPN promoter reporter construct. Moreover, co-expressionof PEA3, c-Jun, and either LEF-1 or �-catenin enhanced theOPN promoter reporter activity by 140- and 180-fold overall,respectively (Fig. 3A). This stimulation of OPN promoter re-porter activity was enhanced still further with lysates contain-ing all four protein factors to 280-fold overall. These resultsshow synergistic co-activator functions of PEA3, c-Jun with�-catenin, and Lef-1 on OPN promoter reporter activity.
Activation of Endogenous OPN Expression by PEA3, c-Jun,�-Catenin, and Lef-1—Rama 37 cells that typically express OPNmRNA at a low level were transiently transfected with expres-sion vectors for PEA3, c-Jun, �-catenin, and Lef-1, individuallyand in combination at the same optimal concentrations and for
the same time period used in the OPN promoter-reporter assays.Stepwise increases in endogenous OPN mRNA were observedafter transfection with combined expression vectors for PEA3/c-Jun with Lef-1 and with Lef-1/�-catenin (Fig. 3B). These in-creases corresponded to 2.5-, 5.2-, and 7.2-fold, respectively, overthat produced endogenously after normalization for the expres-sion of a constitutively expressed GAPDH mRNA.
c-Jun and �-Catenin-Lef-1 Act Independently to Synergizewith PEA3—One possible explanation for synergy of �-cateninwith PEA3 on the activity of the OPN promoter could involveup-regulation of expression of c-Jun by �-catenin (30). To ad-dress this possibility, we constructed an OPN promoter-lucif-erase reporter vector containing inactivating point mutationsin the AP-1 site (mAP1-OPN-Luc). Mutant and wild type OPN-luciferase reporters were co-transfected with combination ofPEA3, c-Jun, Lef-1, and �-catenin expression vectors intoRama 37 cells. Mutation of the AP-1 site did not alter basalOPN promoter reporter activity over that of the wild type
FIG. 3. Effects of PEA3 and c-Junon �-catenin-Lef-1 stimulation of OPN promoter reporter activity and endogenous geneexpression. A, activation of the OPN promoter by PEA3, c-Jun, �-catenin (�-cat), and Lef-1. The OPN-Luc construct was co-transfected withcombinations of expression vectors for PEA3, c-Jun, Lef-1, and �-catenin into 1 � 105 Rama 37 cells. The output of the OPN promoter was measuredby the activity of the firefly luciferase reporter gene after 24 h (“Experimental Procedures”). The data are presented as fold induction of the OPNpromoter reporter relative to co-transfection of the reporter with empty expression vectors. The values were normalized to those obtained with theSV40-Renilla luciferase internal. The data bars represent the means of four experiments, each performed in triplicate. The error bars representthe standard errors. B, induction of endogenous OPN mRNA by transfection of transcription factors. Rama 37 cells were transiently transfectedwith the expression vectors indicated, total RNA was harvested from cells 24 h later, and the Northern blots were performed for OPN and forGAPDH. The autoradiograph image of the blot is shown, and the positions of OPN mRNA at 1.4 kb and GAPDH mRNA at 1.2 kb are shown.
Ets, �-Catenin, and c-Jun Regulate Osteopontin20798
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

promoter (Fig. 4). Expression vectors for PEA3, alone or incombination with those for Lef-1 and �-catenin, activated boththe mutant (mAP1-OPN-Luc) and wild type OPN-luciferasepromoter-reporter constructs to similar degrees (Fig. 4). How-ever, no enhancement of the mutant reporter was observed bythe expression vector for c-Jun, although c-Jun enhanced lucif-erase activity from the wild type OPN-Luc reporter, with PEA3and Lef-1 alone or PEA3 with Lef-1 and �-catenin, by 17- and14-fold, respectively (Fig. 4). Thus the effects of c-Jun uponmAP1-OPN-Luc activation were not additive to those of expres-sion vectors for PEA3, alone or in combination with Lef-1 and�-catenin (Fig. 4). These results indicate that Lef-1/�-catenin
synergy with PEA3 on OPN promoter reporter transactivationis independent of the expression of c-Jun.
�-Catenin-Lef-1 Synergize with PEA3/c-Jun through Tcf-binding Motifs, in the OPN Promoter—To investigate Tcf-bind-ing motifs as potential �-catenin-Lef-1-responsive elementswithin the OPN promoter, we generated OPN-promoter lucif-erase-reporter constructs mutated at one Tcf recognition site(single mutation at position �1960 [mTcf �1960-OPN-Luc]), attwo sites (double mutation at positions �1960/�1463, [mTcf�1960/�1463-OPN-Luc]), and at three sites (triple mutation atpositions �1960/�1463/�1121, mTcf �1960/�1463/�1121-OPN-Luc). Luciferase activity from any of the three mutated
FIG. 4. Effects of transcription fac-tors on mutation of the AP-1 site ofthe OPN promoter reporter. Mutationof the AP-1 site (mAP) nominally affectsonly c-Jun transactivation of the OPNpromoter. A 2-bp inactivating mutation ofthe AP-1 site (TGAGTCA to CGAGTCG)was introduced into the OPN-promoter-luciferase-reporter to create mAP1 of theOPN promoter-luciferase reporter. Themutant promoter was co-transfected intoRama 37 cells with combinations of thePEA3, c-Jun, Lef-1, and �-catenin (�-cat)expression vectors as indicated, in paral-lel experiments with the wild type pro-moter-reporter construct. The data arepresented as fold induction of the OPNpromoter relative to co-transfection of thepromoter reporter constructs with emptyexpression vectors. The values were nor-malized to those obtained by co-transfec-tion with simian virus 40-driven Renillaluciferase. The data bars represent themeans of experiments repeated a mini-mum of three times, each transfectionperformed in triplicate. The error barsrepresent the standard errors.
FIG. 5. Effects of transcription fac-tors on Tcf site mutations of the OPNpromoter-reporter. Inactivating pointmutations were introduced into one, two,or three of the Tcf-binding sites of theOPN promoter-Luc reporter construct tocreate Mut-OPN (�1960 Tcf), Mut-OPN(�1960/�1463 Tcf), and Mut-OPN(�1960/�1463/�1115 Tcf) promoter re-porters, respectively. The wild type andmutant promoter-reporters were co-transfected into Rama 37 cells in parallelwith combinations of the expression vec-tors for PEA3, c-Jun, Lef-1, and �-catenin(�-cat). The output of the OPN promoterwas measured by the activity of the fireflyluciferase reporter gene after 24 h (“Ex-perimental Procedures”). The data arepresented as fold induction of the OPNpromoter relative to co-transfection of thereporter with empty expression vectors.The values were normalized to those ob-tained with the SV40-Renilla luciferaseinternal control. The data bars representthe means of three experiments, each per-formed in triplicate. The error bars repre-sent the standard errors.
Ets, �-Catenin, and c-Jun Regulate Osteopontin 20799
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from
OPN promoter reporter constructs was unaffected by expres-sion vectors for PEA3 or c-Jun, alone or in combination (Fig. 5).However, the Lef-1 and Lef-1/�-catenin expression vector en-hancement of PEA3/c-Jun-mediated OPN promoter-reporteractivity was reduced by mutation of the Tcf-binding sites. Sin-gle mutation at position �1960 was associated with a decline ofLef-1 and Lef-1/�-catenin enhancement PEA3/c-Jun-mediatedOPN promoter-reporter activity to 48% for Lef-1 alone (65-foldenhancement for mTcf �1960 versus 136-fold for wild type) and46% for Lef-1/�-catenin (120-fold enhancement for mTcf �1960versus 260-fold for wild type). The corresponding values for theTcf double mutant versus wild type were 42% for Lef-1 (57-foldenhancement for mTcf �1960/�1463 versus 136-fold for wildtype) and 29% for Lef-1/�-catenin (75-fold enhancement formTcf �1960/�1463 versus 260-fold for wild type). Mutation ofall three Tcf-binding sites in the OPN promoter effectivelyabolished Lef-1 and Lef-1/�-catenin enhancement of PEA3/c-Jun-mediated OPN promoter-reporter activity (Fig. 5). Theseresults indicate that �-catenin and LEF-1 cooperate independ-ently with c-Jun/PEA3 to transactivate the OPN promoter.
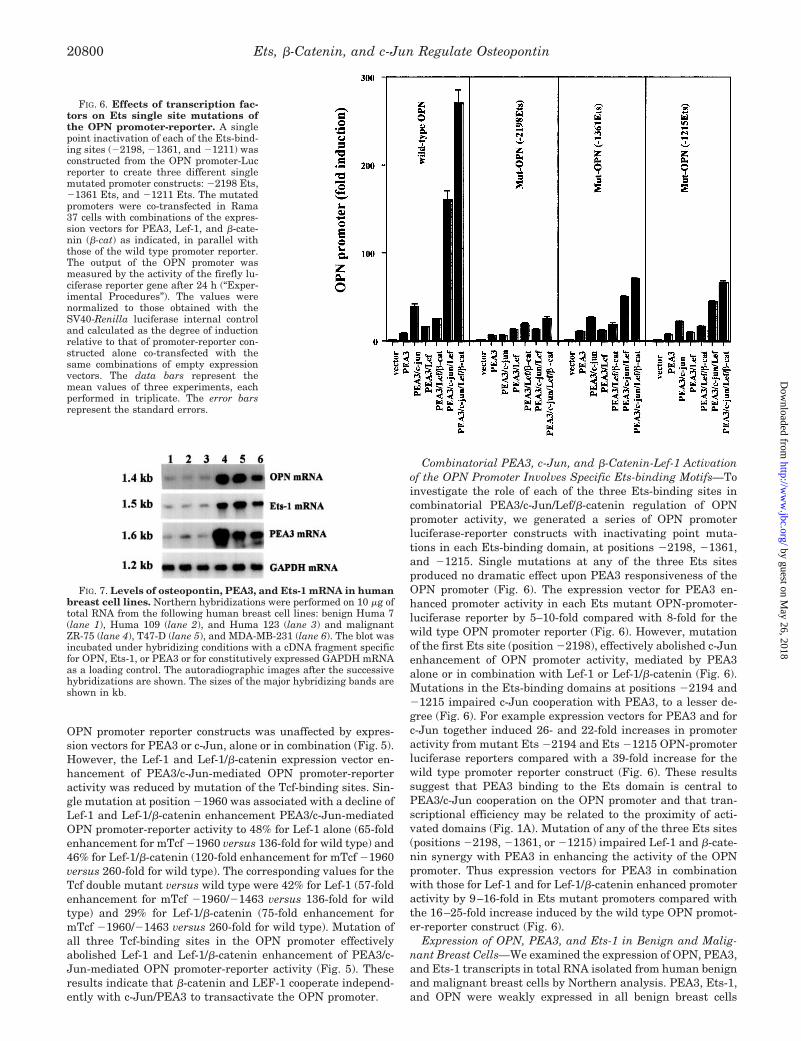
Combinatorial PEA3, c-Jun, and �-Catenin-Lef-1 Activationof the OPN Promoter Involves Specific Ets-binding Motifs—Toinvestigate the role of each of the three Ets-binding sites incombinatorial PEA3/c-Jun/Lef/�-catenin regulation of OPNpromoter activity, we generated a series of OPN promoterluciferase-reporter constructs with inactivating point muta-tions in each Ets-binding domain, at positions �2198, �1361,and �1215. Single mutations at any of the three Ets sitesproduced no dramatic effect upon PEA3 responsiveness of theOPN promoter (Fig. 6). The expression vector for PEA3 en-hanced promoter activity in each Ets mutant OPN-promoter-luciferase reporter by 5–10-fold compared with 8-fold for thewild type OPN promoter reporter (Fig. 6). However, mutationof the first Ets site (position �2198), effectively abolished c-Junenhancement of OPN promoter activity, mediated by PEA3alone or in combination with Lef-1 or Lef-1/�-catenin (Fig. 6).Mutations in the Ets-binding domains at positions �2194 and�1215 impaired c-Jun cooperation with PEA3, to a lesser de-gree (Fig. 6). For example expression vectors for PEA3 and forc-Jun together induced 26- and 22-fold increases in promoteractivity from mutant Ets �2194 and Ets �1215 OPN-promoterluciferase reporters compared with a 39-fold increase for thewild type promoter reporter construct (Fig. 6). These resultssuggest that PEA3 binding to the Ets domain is central toPEA3/c-Jun cooperation on the OPN promoter and that tran-scriptional efficiency may be related to the proximity of acti-vated domains (Fig. 1A). Mutation of any of the three Ets sites(positions �2198, �1361, or �1215) impaired Lef-1 and �-cate-nin synergy with PEA3 in enhancing the activity of the OPNpromoter. Thus expression vectors for PEA3 in combinationwith those for Lef-1 and for Lef-1/�-catenin enhanced promoteractivity by 9–16-fold in Ets mutant promoters compared withthe 16–25-fold increase induced by the wild type OPN promot-er-reporter construct (Fig. 6).
Expression of OPN, PEA3, and Ets-1 in Benign and Malig-nant Breast Cells—We examined the expression of OPN, PEA3,and Ets-1 transcripts in total RNA isolated from human benignand malignant breast cells by Northern analysis. PEA3, Ets-1,and OPN were weakly expressed in all benign breast cells
FIG. 6. Effects of transcription fac-tors on Ets single site mutations ofthe OPN promoter-reporter. A singlepoint inactivation of each of the Ets-bind-ing sites (�2198, �1361, and �1211) wasconstructed from the OPN promoter-Lucreporter to create three different singlemutated promoter constructs: �2198 Ets,�1361 Ets, and �1211 Ets. The mutatedpromoters were co-transfected in Rama37 cells with combinations of the expres-sion vectors for PEA3, Lef-1, and �-cate-nin (�-cat) as indicated, in parallel withthose of the wild type promoter reporter.The output of the OPN promoter wasmeasured by the activity of the firefly lu-ciferase reporter gene after 24 h (“Exper-imental Procedures”). The values werenormalized to those obtained with theSV40-Renilla luciferase internal controland calculated as the degree of inductionrelative to that of promoter-reporter con-structed alone co-transfected with thesame combinations of empty expressionvectors. The data bars represent themean values of three experiments, eachperformed in triplicate. The error barsrepresent the standard errors.
FIG. 7. Levels of osteopontin, PEA3, and Ets-1 mRNA in humanbreast cell lines. Northern hybridizations were performed on 10 �g oftotal RNA from the following human breast cell lines: benign Huma 7(lane 1), Huma 109 (lane 2), and Huma 123 (lane 3) and malignantZR-75 (lane 4), T47-D (lane 5), and MDA-MB-231 (lane 6). The blot wasincubated under hybridizing conditions with a cDNA fragment specificfor OPN, Ets-1, or PEA3 or for constitutively expressed GAPDH mRNAas a loading control. The autoradiographic images after the successivehybridizations are shown. The sizes of the major hybridizing bands areshown in kb.
Ets, �-Catenin, and c-Jun Regulate Osteopontin20800
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

tested (Huma 7, Huma 109, and Huma 123). Conversely, co-incident high expression of OPN, PEA3, and Ets-1 was ob-served in malignant breast cells (ZR-75, T47-D, and MDA-MB231) (Fig. 7).
Immunocytochemical Staining of Human Breast Cancers forTranscription Factors and OPN—Nearly adjacent histologicalsections from specimens from 29 patients with primary inva-sive breast carcinoma were immunocytochemically stained forOPN and for different transcription factors. All of the antibod-ies used against OPN and the transcription factors stainedhistological sections from some of these breast carcinomas(Figs. 8, A, C, and E, and 9, A, C, E, and G). The specificity ofthe antibodies used against OPN (Fig. 8, A and B), �-catenin(Fig. 8, C and D), Tcf-4 (Fig. 8, E and F), Ets-1, (Fig. 9, A andB), Ets-2 (Fig. 9, C and D), PEA3 (Fig. 9, E and F), and c-Jun(Fig. 9, G and H) was tested by their prior incubation with therequisite antigen described under “Experimental Procedures.”Immunocytochemical staining was suppressed completely bythis treatment (Figs. 8, B, D, and F, and 9, B, D, F, and H) butnot with antibodies incubated with noncognate antigens (notshown), demonstrating that the antibodies were specific. Stain-ing of the carcinoma cells for OPN and �-catenin was predom-
inantly cytoplasmic, although both membranous and nuclearstaining was observed for the latter but not the former antigen(Fig. 10, A and B). Specific staining for OPN was also observedin the cytoplasm of reactive stromal cells/lymphocytes as pre-viously reported (8). Staining for Ets-1, Ets-2, PEA3, and c-Junvaried from nuclear to cytoplasmic depending both on the iden-tity of the transcription factor and the specimen (Fig. 10, C–F).Staining for Tcf-4 was mainly nuclear (not shown). The sam-ples were considered positively stained if more than 5% of thecarcinoma cells were stained strongly by the requisite antibody(“Experimental Procedures”). The percentage of positivelystained samples for �-catenin, Tcf-4, Ets-1, Ets-2, PEA3, c-Jun,and OPN were �71, 35, 78, 78, 63, 78, and 66%, respectively.Staining for OPN was significantly correlated with that for�-catenin, Ets-1, Ets-2, PEA3, and c-Jun (p � 0.001) but notwith that for Tcf-4 (p � 0.36) when cross-tabulated and as-sessed using Fisher’s Exact test (“Experimental Procedures”)(Table I). Similarly there was a significant correlation for sam-ples staining positively for any one of the transcription factorsin comparison with another (p � 0.01), except only a borderlinecorrelation of staining for �-catenin and c-Jun (p � 0.06). Nosignificant correlation between staining for Tcf-4 and any fac-
FIG. 8. Specificity of immunocytochemical staining of human breast cancers for OPN, �-catenin, and Tcf-4. A and B, sections fromthe same area of a tumor specimen incubated with either MAb to OPN (A) or MAb preabsorbed with OPN (B). C and D, sections from a similararea of a tumor specimen incubated with either MAb to �-catenin (C) or MAb preabsorbed with �-catenin blocking peptide (D). E and F, sectionsfrom a similar area of a specimen incubated with either MAb to Tcf-4 (E) or MAb preabsorbed with Tcf-4 blocking peptide (F). Positiveimmunocytochemical staining in A, C, and E is abolished in B, D, and F. Magnification, �230; bar, 50 �m.
Ets, �-Catenin, and c-Jun Regulate Osteopontin 20801
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

tor (p � 0.19) was observed. Within the same specimens, im-munocytochemical staining for �-catenin and OPN; for Ets-1,Ets-2, and OPN; and for PEA3, c-Jun, and OPN (not shown)occurred in similar areas, often in the same cells. This was notthe case for staining for Tcf-4 and OPN (not shown). Whenstaining for four or more of �-catenin, Ets-1, Ets-2, PEA3, andc-Jun was positive in the same specimen, that specimen wasalso positive for OPN. When staining for three or less of thesame transcription factors occurred in the same specimen, thatspecimen remained unstained for OPN (Table II).
DISCUSSION
A number of mitogenic molecules including growth factors(48), oncogenes (49), and tumor promoters (10) induce the ex-
pression of OPN, which has a key role in metastasis (9) andprognosis of breast cancer (45). The understanding of regula-tory mechanisms for transcription of OPN is incomplete. Stud-ies on the mouse or human OPN promoter has revealed ahighly modular structure (50) involving four regions, as fol-lows: (i) A nuclear receptor and a Ras-responsive region en-coded by the OPN promoter, nucleotide sequence �1000 to�550 (2); peroxisome proliferator-activated receptor � (51), vi-tamin D receptor (13), and estrogen receptor-related receptor-�(52, 53) all bind to elements in this region. (ii) A regulatoryregion relevant to osteoblast differentiation that maps to theOPN promoter region nucleotides �220 to �110 and is regu-lated by Polyoma enhancer-binding protein, 2�A/core-binding
FIG. 9. Specificity of immunocytochemical staining of human breast cancers for Ets-1, Ets-2, PEA3, and c-Jun. A and B, sections fromthe same area of a specimen incubated with either rabbit polyclonal antibody to Ets-1 (A) or antibody preincubated with blocking peptide to Ets-1(B). C and D, sections from a similar area of a specimen incubated with either rabbit polyclonal antibody to Ets-2 (C) or antibody preincubated withblocking peptide to Ets-2 (D). E and F, sections from the same area of a specimen incubated with either MAb to PEA3 (E) or MAb preincubatedwith reticulocyte protein-synthesizing lysate primed with an expression vector for PEA3 (F). Preincubation with the general blocking peptide forthe Ets family gave the same results. G and H, sections from a similar area of a specimen incubated with either rabbit antibody to c-Jun (G) orantibody preincubated with c-Jun fusion protein (H). Positive immunocytochemical staining in A, C, E, and G is abolished in B, D, F, and H.Magnification, �230; bar, 50 �m.
Ets, �-Catenin, and c-Jun Regulate Osteopontin20802
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

factor, and Ets-1 (12), and bone morphogenetic protein simu-lated through an interaction with a homeodomain transcrip-tion factor, Hox-8, and Smad1 (BMP2-regulated Hoxc8:Smad1)(54). (iii) A basal promoter region that maps to nucleotides�107 to �78 relative to the start site. (iv) Enhancer element�799 to �864 that supports Sox2:Oct4-regulated expression(55). Cooperation between factors interacting at multiple sitesmay be implicated in context-specific OPN gene expression.
We have previously shown the responsiveness of the rat OPNpromoter to Tcf components of the �-catenin signaling pathway(22). AP-1 and the Ets transcription family influence OPNexpression (2, 12, 14, 27) and have oncogenic potential in mam-mary epithelium (56–58). In a data base-assisted bioinformat-ics analysis of the rat OPN promoter, we have found threeTcf-binding sites at nucleotide positions �1960 to �1954,�1463 to �1457, and �1121 to �1115 and three Ets-bindingdomains at nucleotide positions �2198 to �2194, �1361 to�1357, and �1215 to �1211, and an AP-1 domain was con-firmed at position �1872 to �1866. We have previously shownbinding of Tcf-1 and Tcf-4 to an oligonucleotide representingthe Tcf-binding site at nucleotide positions �1121 to �1115(40). Here, we report assembly of Lef-1: DNA complexes and
co-localization of �-catenin with Lef-1 and the two other Tcf-binding domains at nucleotide positions �1960 and �1463.Mutagenesis of the Tcf domain at nucleotide �1960 effectivelyblocked its binding to Lef-1.
To assess AP-1 and/or Ets regulation of OPN transcription,we performed co-transfection experiments of a wild type OPNpromoter-luciferase reporter construct with expression vectorsfor c-Jun, Ets-1, Ets-2, PEA3, and/or the Ets subfamily gene,ERM alone, or in the combinations outlined under “Experimen-tal Procedures.” Individually, each of the Ets factors enhancedluciferase activity of the OPN promoter reporter construct andall synergized with c-Jun. PEA3 and ERM showed the largestincrease in activity within the Ets family for the OPN pro-moter, whether alone or in combination with c-Jun.
Functional synergy between PEA3, the AP-1 activator pro-tein c-Jun, Lef-1, and �-catenin for promoter activity was in-vestigated with the wild type OPN promoter-linked luciferasereporter. We found that PEA3 had a modest co-activator func-tion with �-catenin-Lef-1. However, co-expression of c-Jun withPEA3 and either Lef-1 or �-catenin enhanced luciferase expres-sion from the promoter by 140- and 180-fold, respectively,whereas a large 280-fold stimulation of promoter activity was
FIG. 10. Immunocytochemical staining of human breast cancers for OPN and for different transcription factors at highermagnification. A and B, sections from the same specimen of a tumor incubated with MAb to OPN (A) or �-catenin (B). Cytoplasmic staining isindicated by a thick arrow in both A and B, whereas membranous and nuclear staining in B are indicated by thin and curved arrows, respectively.C and D, sections from the same specimen of a second tumor incubated with polyclonal antibody to Ets-1 (C) or polyclonal antibody to Ets-2 (D),showing cytoplasmic (thick arrow) and nuclear (curved arrow) staining. E and F, sections from the same specimen of a third tumor incubated withMAb to PEA3 (E) or polyclonal antibody to c-Jun (F), showing predominantly nuclear staining (curved arrows) with the occasional cell containingstaining in the cytoplasm (thick arrow). Magnification, �580; bar, 20 �m.
Ets, �-Catenin, and c-Jun Regulate Osteopontin 20803
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

observed by co-transfection of PEA3, c-Jun, �-catenin, andLef-1. Luciferase reporter systems provide useful informationconcerning transcriptional regulation of a gene of interest, al-though their quantitative output may be influenced by trans-fection efficiency, translation, and activity of the reporter pro-tein, as well as transcription from the promoter. Confirmatoryassays of endogenous gene transcription can lend weight tofindings. In this study, we show that the increases in the levels
of endogenous OPN mRNA triggered by the various combina-tions of expression vectors reflect the increases in OPN promot-er-reporter activity. However, the overall combination of ex-pression vectors for PEA3, c-Jun, �-catenin, and Lef-1 thatinduced a maximum increase in OPN promoter-reporter activ-ity of 280-fold induced only a maximum increase in endogenousOPN mRNA of 7.2-fold. This difference could be a reflection ofdifferences in and/or stability of OPN mRNA.
To investigate the specific role of AP-1 factors in this collab-orative regulatory network, the activity of the OPN promoterluciferase reporter construct bearing an inactivating mutationat the AP-1 site (mAP-OPN-Luc) was tested. Mutational in-activation of the AP-1 site had no effects of transactivation onthe OPN promoter by PEA3 alone or in combination withLef-1 or Lef-1 and �-catenin. These results do not supportfunctional overlap between AP-1, PEA3, or �-catenin/Lef-1 onthe activity of the OPN promoter. However, this mutationabolished synergy of c-Jun with PEA3 alone, with PEA3/Lef-1, and with PEA3/Lef-1/�-catenin on the activity of theOPN promoter. AP-1, Ets, and Tcf transcription factors mayshare some common co-regulator proteins (59, 60). Conceiv-ably, protein binding to an intact AP-1 site may enable re-cruitment of co-activators for Ets- and Tcf-mediated OPNpromoter activity.
The influence of identified Tcf-binding sites on OPN promot-er-reporter transactivation was next investigated. Mutationsat the defined Tcf-binding sites had no effect upon OPN pro-moter-reporter responsiveness to PEA3 or c-Jun, alone or incombination. However, Lef-1 synergy with these transcriptionfactors for transactivation of the OPN promoter-reporter wasaffected to a variable degree. Mutation at all three Tcf-bindingsites effectively abolished Lef-1 synergy on promoter activity,in co-transfection experiments. Of necessity, mutations at po-sition �1960 and �1463 invoked major changes to the spatialorganization between the single AP-1 site and the next adja-cent Tcf-binding motif. The distance between these bindingdomains increased from �100 bp in the wild type OPN pro-moter to �400 and 700 bp in the single �1960 and double�1960/�1463 mutants, respectively. Conversely, the maxi-mum distance between any single Tcf- and Ets-binding domainremained relatively constant in the single and double mutants,for either transcription factor-binding site (Fig. 1A). However,the three pairs of Tcf- and Ets-binding sites lay within 240-bpproximity in the wild type promoter. Single �1960 and double�1960/�1463 mutants of the Tcf-1-binding site had only twoand one pair separated by this distance or less, respectively.PEA3/Lef-1 synergy on the activity of the OPN promoter, al-though diminished, remained detectable in �1960 single anddouble �1960/�1463 mutants. Conversely, Lef-1 synergy withc-Jun/PEA3 and c-Jun/PEA3/�-catenin on promoter activitywas inhibited to a greater degree in single and double Tcf-binding site mutants of the OPN promoter. Differential effectsof these mutations upon Tcf/Ets and Tcf/AP-1 synergy on theactivity of the OPN promoter may be related to the positionalrelationships between Tcf, AP-1, and Ets-binding domains ofthis promoter. These studies have also shown that �-cateninsynergy with PEA3/c-Jun/Lef-1 on the activity of this promoteris mediated through the identified Tcf sites and not throughany cryptic Tcf domains, as has been reported for other pro-moters (61).
Unlike many other families of transcription factors, the Etsfamily does not appear to associate as homo- or heterodimers(62) and by themselves may display only weak transactivationproperties. Instead, the Ets family may form complexes withunrelated factors to initiate or enhance transcription (63). In-teractions between Ets factors and AP-1 proteins have variable
TABLE IAssociation of immunocytochemical staining of breast carcinomas for
OPN with that for individual transcription factorsHistological sections from samples of invasive ductal carcinomas of
the breast were immunocytochemically stained for �-catenin, Tcf-4,Ets-1, Ets-2, PEA3, c-Jun, and OPN as described under ‘‘ExperimentalProcedures.’’ The samples where the percentage of stained carcinomacells was �5% are recorded as positive, and the rest are recorded asnegative. The statistical significance of correlated staining betweenpaired samples was obtained from Fisher’s Exact test (two-sided value),where a probability of p � 0.05 was considered significant. (‘‘Experi-mental Procedures’’).
Transcription factor OPN-negative OPN-positive Statisticalsignificance
% %
�-Catenin � 7 (78) 1 (5)�-Catenin � 2 (22) 18 (95) �0.001Tcf-4 � 6 (86) 11 (58)Tcf-4 � 1 (14) 8 (42) 0.36Ets-1 � 6 (75) 0 (0)Ets-1 � 2 (25) 19 (100) �0.001Ets-2 � 6 (75) 0 (0)Ets-2 � 2 (25) 19 (100) �0.001PEA3 � 8 (100) 2 (11)PEA3 � 0 (0) 17 (89) �0.001c-Jun � 6 (75) 0 (0)c-Jun � 2 (25) 19 (100) �0.001
TABLE IIAssociation of immunocytochemical staining for multiple transcription
factors with that for OPN in breast carcinomasImmunocytochemical staining of breast carcinomas for �-catenin,
Tcf-4, Ets-1, Ets-2, PEA3, c-Jun, and OPN as described under ‘‘Exper-imental Procedures.’’ Those sections where the percentage of stainedcarcinoma cells was �5% are recorded as positive (�), and the remain-der are recorded as negative (�) or not determined (ND).
Patient numberImmunostaining
�-Catenin Tcf-4 Ets-1 Ets-2 PEA3 c-Jun OPN
1 � � � � � � �2 � � � � � � �3 � � � � � � �4 � � � � � � �5 � � � � � � �6 � � � � � � �7 � � � � � � �8 � � � � � � �9 � � � � � � �
10 � � � � � � �11 � � � � � � �12 � � � � � � �13 � � � � � � �14 � � ND ND ND ND �15 � � � � � � �16 � � � � � � �17 � � ND ND ND ND �18 � � � � � � �19 � � � � � � �20 � � � � � � �21 � ND � � � � �22 � � � � � � �23 ND ND � � � � �24 � � � � � � �25 � � � � � � �26 � � � � � � �27 � � � � � � �28 � � � � � � �29 � ND � � � � �
Ets, �-Catenin, and c-Jun Regulate Osteopontin20804
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

effects upon transcription (64). In the present study, muta-tional analysis was carried out to investigate the role of eachEts motif in PEA3-mediated transactivation of the OPN pro-moter, alone or in combination with other factors. PEA3 com-bined with c-Jun stimulated the activity of the wild type OPNpromoter reporter by an additional 5.5-fold, over that withPEA3 alone (Fig. 2). No single mutation at any Ets site abol-ished OPN transactivation by PEA3 alone or in combinationwith other factors. Mutation at the �2198 Ets-binding site,which was closest to the AP-1 site, had the greatest inhibitoryeffect upon PEA3-, PEA3/c-Jun-, or PEA3/c-Jun/Lef-1-medi-ated OPN transactivation. For example, the combination ofPEA3 and c-Jun transactivated the OPN promoter-reporter ofeach of the single Ets-binding site mutants �2198, �1361, and�1215 by approximately 0-, 2.5-, and 3-fold, respectively, overthat with PEA3 alone and by approximately 0-, 4.5-, and 5-fold,respectively, over that with PEA3 and Lef-1, in comparisonwith stimulation of the wild type promoter (Fig. 6). Thus theEts-binding site closest to the AP-1-binding domain appearedto exert the greatest effect on the activity of the OPN pro-moter, by c-Jun or c-Jun/Lef-1 transactivation. One of the mostintriguing findings was that mutation at any one of the Ets-binding sites was associated with a substantial inhibition ofcooperative PEA3/c-Jun/Lef-1 or PEA3/c-Jun/Lef-1/�-catenintransactivation of the OPN promoter reporter construct. Thusfull occupancy of all Ets sites within the OPN promoter is prob-ably essential for maximal synergy between these cis-actingregulatory elements, in activation of the OPN promoter.
In addition to osteopontin, other genes are co-regulated byvarious combinations of PEA3, �-catenin/Tcf, and AP-1 tran-scription factors. Cross-coupling of different signaling path-ways through these cis-acting regulatory domains influencesexpression of TWIST transcription factor (65), which inhibitsmammary cell differentiation and expression of matrilysin, amember of the matrix metalloproteinase family (matrix metal-loproteinase 7) (30). Co-incident expression of OPN and matrixmetalloproteinase has been reported in conditions of tissueinjury or wound repair (66) and tumorigenesis (67), whereasmatrix metalloproteinase 7 may be co-expressed with OPN in aremodeling tissue, like the postpartum uterus (68, 69). OPN isproteolytically cleaved by matrilysin with a resulting increaseof its activity in adhesion and migration (70). Taken together,these data suggest that PEA3-, �-catenin/Tcf-, and AP-1 tran-scription factor-co-regulated target genes may be implicated inkey events of mammary tumorigenesis, including suppressionof differentiation (65), neoplastic transformation (49), and cellmigration across tissue barriers (67). Further work is requiredto identify other PEA3, �-catenin/Tcf, and AP-1 combinatorialtarget genes implicated in mammary oncogenesis.
To test the biological significance of our findings that definedtranscription factors can stimulate the OPN promoter in tran-sient transfection assays in a rat mammary model cell line,their co-expression with OPN has been assessed using North-ern blotting to detect their mRNAs in human breast cell linesand by immunocytochemical techniques to detect their proteinsin specimens from primary breast cancers. Prior incubation ofantibodies to OPN and to each of the transcription factors withthe requisite antigen completely abolished any immunocyto-chemical staining, confirming that such staining was specificfor the antigen in question. Those cell lines showing high levelsof OPN mRNA also showed high levels of mRNA for Ets-1 andPEA3. The high expressing cell lines were isolated from malig-nant sources, in agreement with the idea that OPN is associ-ated with tumor progression and metastasis (4). The associa-tion of immunocytochemical staining for OPN separately withthat for �-catenin, Ets-1, Ets-2, PEA3, and c-Jun, but not with
that for Tcf-4 in the human breast cancers, is consistent withthe finding for the OPN reporter in transient transfection as-says of the rat mammary cell line. Lef-1, the transcriptionfactor homologous to Tcf-4, showed little stimulation of theactivity of the OPN promoter reporter construct without �-cate-nin, whereas the Ets and AP-1 classes of transcription factorsdid show stimulation. Moreover, the same areas of the tumorspecimens showed coordinated elevation of staining for OPNand for staining for �-catenin and the other two classes oftranscription factors. In the group of tumors studied, albeitsmall, no specimen showed immunocytochemical staining forOPN above the threshold when staining for one of the threeclasses of transcription factors �-catenin/Tcf, Ets, and AP-1was designated as negative. Thus it is possible that the syner-gistic overexpression of �-catenin/Tcf, Ets, and AP-1 classes oftranscription factors are responsible, in part, for the overpro-duction of OPN in human breast cancers. These three classes oftranscription factors alone, however, cannot specify overpro-duction of OPN in the human tumors, because they are oftenelevated in some reactive stromal cells, probably myofibro-blasts, which fail to express immunoreactive OPN.2 Thus otherregulators of OPN expression are also probably required tostimulate OPN production, e.g. the activated steroid hormonereceptors for vitamin D3 (71) and estradiol (40). AP-1, Etsproteins, and �-catenin/Lef-1 synergize in the up-regulationof the matrix metalloproteinase matrilysin (72), another pro-tein associated with tumor invasion and metastasis. Theoverexpression of transcription factors AP-1 (73) and mem-bers of the Ets (58) (73–75) family as well as �-catenin/Tcf(76) may reflect selection in some tumors for those cellscapable of mounting a coordinated response to increase thelevels of proteins important for successful development of thecancer.
REFERENCES
1. Tuck, A. B., Hota, C., Wilson, S. M., and Chambers, A. F. (2003) Oncogene 22,1198–1205
2. Guo, X., Zhang, Y. P., Mitchell, D. A., Denhardt, D. T., and Chambers, A. F.(1995) Mol. Cell Biol. 15, 476–487
3. Weber, G. F. (2001) Biochim. Biophys. Acta 1552, 61–854. Agrawal, D., Chen, T., Irby, R., Quackenbush, J., Chambers, A. F., Szabo, M.,
Cantor, A., Coppola, D., and Yeatman, T. J. (2002) J. Natl. Cancer Inst. 94,513–521
5. Senger, D. R., and Perruzzi, C. A. (1985) Cancer Res. 45, 5818–58236. Senger, D. R., Perruzzi, C. A., Gracey, C. F., Papadopoulos, A., and Tenen,
D. G. (1988) Cancer Res. 48, 5770–57747. Devoll, R. E., Li, W., Woods, K. V., Pinero, G. J., Butler, W. T., Farach-Carson,
M. C., and Happonen, R. P. (1999) J. Oral Pathol. Med. 28, 97–1018. Rudland, P. S., Platt-Higgins, A., El-Tanani, M., De Silva Rudland, S.,
Barraclough, R., Winstanley, J. H., Howitt, R., and West, C. R. (2002)Cancer Res. 62, 3417–3427
9. Oates, A. J., Barraclough, R., and Rudland, P. S. (1996) Oncogene 13, 97–10410. Su, L., Mukherjee, A. B., and Mukherjee, B. B. (1995) Oncogene 10, 2163–216911. Behrend, E. I., Craig, A. M., Wilson, S. M., Denhardt, D. T., and Chambers,
A. F. (1994) Cancer Res. 54, 832–83712. Sato, M., Morii, E., Komori, T., Kawahata, H., Sugimoto, M., Terai, K.,
Shimizu, H., Yasui, T., Ogihara, H., Yasui, N., Ochi, T., Kitamura, Y., Ito,Y., and Nomura, S. (1998) Oncogene 17, 1517–1525
13. Noda, M., Vogel, R. L., Craig, A. M., Prahl, J., DeLuca, H. F., and Denhardt,D. T. (1990) Proc. Natl. Acad. Sci. U. S. A. 87, 9995–9999
14. Bidder, M., Shao, J. S., Charlton-Kachigian, N., Loewy, A. P., Semenkovich,C. F., and Towler, D. A. (2002) J. Biol. Chem. 277, 44485–44496
15. Lin, S. Y., Xia, W., Wang, J. C., Kwong, K. Y., Spohn, B., Wen, Y., Pestell, R. G.,and Hung, M. C. (2000) Proc. Natl. Acad. Sci. U. S. A. 97, 4262–4266
16. Miyoshi, K., Rosner, A., Nozawa, M., Byrd, C., Morgan, F., Landesman-Bollag,E., Xu, X., Seldin, D. C., Schmidt, E. V., Taketo, M. M., Robinson, G. W.,Cardiff, R. D., and Hennighausen, L. (2002) Oncogene 21, 5548–5556
17. Korinek, V., Barker, N., Morin, P. J., van Wichen, D., de Weger, R., Kinzler,K. W., Vogelstein, B., and Clevers, H. (1997) Science 275, 1784–1787
18. Morin, P. J., Sparks, A. B., Korinek, V., Barker, N., Clevers, H., Vogelstein, B.,and Kinzler, K. W. (1997) Science 275, 1787–1790
19. Rubinfeld, B., Robbins, P., El-Gamil, M., Albert, I., Porfiri, E., and Polakis, P.(1997) Science 275, 1790–1792
20. Nagai, S., Hashimoto, S., Yamashita, T., Toyoda, N., Satoh, T., Suzuki, T., andMatsushima, K. (2001) Int. Immunol. 13, 367–376
2 M. El-Tanani, A. Platt-Higgins, P. S. Rudland, and F. C. Campbell,unpublished results.
Ets, �-Catenin, and c-Jun Regulate Osteopontin 20805
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21. Muller, T., Bain, G., Wang, X., and Papkoff, J. (2002) Exp. Cell Res. 280,119–133
22. El-Tanani, M., Barraclough, R., Wilkinson, M. C., and Rudland, P. S. (2001)Cancer Res. 61, 5619–5629
23. Giese, K., Pagel, J., and Grosschedl, R. (1997) Proc. Natl. Acad. Sci. U. S. A.94, 12845–12850
24. Hsu, S. C., Galceran, J., and Grosschedl, R. (1998) Mol. Cell Biol. 18,4807–4818
25. Carlsson, P., Waterman, M. L., and Jones, K. A. (1993) Genes Dev. 7,2418–2430
26. Giese, K., and Grosschedl, R. (1993) EMBO J. 12, 4667–467627. Kim, H. J., Lee, M. H., Shin, H. I., Choi, J. Y., and Ryoo, H. M. (2002) J. Cell
Biochem. 87, 93–10228. Oikawa, T., and Yamada, T. (2003) Gene (Amst.) 303, 11–3429. Himelstein, B. P., Lee, E. J., Sato, H., Seiki, M., and Muschel, R. J. (1997)
Oncogene 14, 1995–199830. Crawford, H. C., Fingleton, B., Gustavson, M. D., Kurpios, N., Wagenaar,
R. A., Hassell, J. A., and Matrisian, L. M. (2001) Mol. Cell Biol. 21,1370–1383
31. Wasylyk, B., Wasylyk, C., Flores, P., Begue, A., Leprince, D., and Stehelin, D.(1990) Nature 346, 191–193
32. Walunas, T. L., Wang, B., Wang, C. R., and Leiden, J. M. (2000) J. Immunol.164, 2857–2860
33. Foos, G., and Hauser, C. A. (2000) Oncogene 19, 5507–551634. de Launoit, Y., Baert, J. L., Chotteau, A., Monte, D., Defossez, P. A., Coutte, L.,
Pelczar, H., and Leenders, F. (1997) Biochem. Mol. Med. 61, 127–13535. Ryder, K., and Nathans, D. (1988) Proc. Natl. Acad. Sci. U. S. A. 85,
8464–846736. Milatovich, A., Travis, A., Grosschedl, R., and Francke, U. (1991) Genomics 11,
1040–104837. Barth, A. I., Pollack, A. L., Altschuler, Y., Mostov, K. E., and Nelson, W. J.
(1997) J. Cell Biol. 136, 693–70638. Ridall, A. L., Daane, E. L., Dickinson, D. P., and Butler, W. T. (1995) Ann. N. Y.
Acad. Sci. 760, 59–6639. El-Tanani, M. K., Barraclough, R., Wilkinson, M. C., and Rudland, P. S. (2001)
Oncogene 20, 1793–179740. El-Tanani, M., Fernig, D. G., Barraclough, R., Green, C., and Rudland, P.
(2001) J. Biol. Chem. 276, 41675–4168241. El-Tanani, M. K., and Green, C. D. (1997) J. Steroid Biochem. Mol. Biol. 60,
269–27642. El-Tanani, M. K., and Green, C. D. (1996) Mol. Cell Endocrinol. 121, 29–3543. Anandappa, S. Y., Winstanley, J. H., Leinster, S., Green, B., Rudland, P. S.,
and Barraclough, R. (1994) Br. J. Cancer 69, 772–77644. Platt-Higgins, A. M., Renshaw, C. A., West, C. R., Winstanley, J. H., De Silva
Rudland, S., Barraclough, R., and Rudland, P. S. (2000) Int. J. Cancer 89,198–208
45. Rudland, P. S., Platt-Higgins, A., Renshaw, C., West, C. R., Winstanley, J. H.,Robertson, L., and Barraclough, R. (2000) Cancer Res. 60, 1595–1603
46. Gorski, J. P., Griffin, D., Dudley, G., Stanford, C., Thomas, R., Huang, C., Lai,E., Karr, B., and Solursh, M. (1990) J. Biol. Chem. 265, 14956–14963
47. Sabattini, E., Bisgaard, K., Ascani, S., Poggi, S., Piccioli, M., Ceccarelli, C.,Pieri, F., Fraternali-Orcioni, G., and Pileri, S. A. (1998) J. Clin. Pathol. 51,506–511
48. Li, G., Oparil, S., Kelpke, S. S., Chen, Y. F., and Thompson, J. A. (2002)Circulation 106, 854–859
49. Wu, Y., Denhardt, D. T., and Rittling, S. R. (2000) Br. J. Cancer. 83, 156–16350. Denhardt, D. T., Noda, M., O’Regan, A. W., Pavlin, D., and Berman, J. S.
(2001) J. Clin. Invest. 107, 1055–106151. Oyama, Y., Akuzawa, N., Nagai, R., and Kurabayashi, M. (2002) Circ. Res. 90,
348–35552. Bonnelye, E., Vanacker, J. M., Dittmar, T., Begue, A., Desbiens, X., Denhardt,
D. T., Aubin, J. E., Laudet, V., and Fournier, B. (1997) Mol. Endocrinol. 11,905–916
53. Vanacker, J. M., Pettersson, K., Gustafsson, J. A., and Laudet, V. (1999)EMBO J. 18, 4270–4279
54. Yang, X., Ji, X., Shi, X., and Cao, X. (2000) J. Biol. Chem. 275, 1065–107255. Botquin, V., Hess, H., Fuhrmann, G., Anastassiadis, C., Gross, M. K., Vriend,
G., and Scholer, H. R. (1998) Genes Dev. 12, 2073–209056. de Launoit, Y., Chotteau-Lelievre, A., Beaudoin, C., Coutte, L., Netzer, S.,
Brenner, C., Huvent, I., and Baert, J. L. (2000) Adv. Exp. Med. Biol. 480,107–116
57. Baert, J. L., Monte, D., Musgrove, E. A., Albagli, O., Sutherland, R. L., and deLaunoit, Y. (1997) Int. J. Cancer 70, 590–597
58. Span, P. N., Manders, P., Heuvel, J. J., Thomas, C. M., Bosch, R. R., Beex,L. V., and Sweep, C. G. (2002) Oncogene 21, 8506–8509
59. Yang, C., Shapiro, L. H., Rivera, M., Kumar, A., and Brindle, P. K. (1998) Mol.Cell Biol. 18, 2218–2229
60. Sun, Y., Kolligs, F. T., Hottiger, M. O., Mosavin, R., Fearon, E. R., and Nabel,G. J. (2000) Proc. Natl. Acad. Sci. U. S. A. 97, 12613–12618
61. Tetsu, O., and McCormick, F. (1999) Nature 398, 422–42662. Roussel, M. F., Davis, J. N., Cleveland, J. L., Ghysdael, J., and Hiebert, S. W.
(1994) Oncogene 9, 405–41563. Boyd, K. E., and Farnham, P. J. (1999) Proc. Soc. Exp. Biol. Med. 222, 9–2864. Buttice, G., Duterque-Coquillaud, M., Basuyaux, J. P., Carrere, S., Kurkinen,
M., and Stehelin, D. (1996) Oncogene 13, 2297–230665. Howe, L. R., Watanabe, O., Leonard, J., and Brown, A. M. (2003) Cancer Res.
63, 1906–191366. Yamagiwa, H., Tokunaga, K., Hayami, T., Hatano, H., Uchida, M., Endo, N.,
and Takahashi, H. E. (1999) Bone 25, 197–20367. Philip, S., Bulbule, A., and Kundu, G. C. (2001) J. Biol. Chem. 276,
44926–4493568. Kao, L. C., Tulac, S., Lobo, S., Imani, B., Yang, J. P., Germeyer, A., Osteen, K.,
Taylor, R. N., Lessey, B. A., and Giudice, L. C. (2002) Endocrinology 143,2119–2138
69. Wilson, C. L., Heppner, K. J., Rudolph, L. A., and Matrisian, L. M. (1995) Mol.Biol. Cell 6, 851–869
70. Agnihotri, R., Crawford, H. C., Haro, H., Matrisian, L. M., Havrda, M. C., andLiaw, L. (2001) J. Biol. Chem. 276, 28261–28267
71. Koszewski, N. J., Reinhardt, T. A., and Horst, R. L. (1996) J. Steroid Biochem.Mol. Biol. 59, 377–388
72. Crawford, H. C., Fingleton, B. M., Rudolph-Owen, L. A., Goss, K. J., Rubinfeld,B., Polakis, P., and Matrisian, L. M. (1999) Oncogene 18, 2883–2891
73. Sato, T., and Miwa, A. (2002) Apmis 110, 347–35374. Nakayama, T., Ito, M., Ohtsuru, A., Naito, S., and Sekine, I. (2001) Mod.
Pathol. 14, 415–42275. Kinoshita, J., Kitamura, K., Tanaka, S., Sugimachi, K., Ishida, M., and Saeki,
H. (2002) Surgery 131, S222–22576. Hugh, T. J., Dillon, S. A., O’Dowd, G., Getty, B., Pignatelli, M., Poston, G. J.,
and Kinsella, A. R. (1999) Int. J. Cancer 82, 504–511
Ets, �-Catenin, and c-Jun Regulate Osteopontin20806
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

CampbellMohamed El-Tanani, Angela Platt-Higgins, Philip S. Rudland and Frederick Charles
Osteopontin Transcription-Catenin-Lef-1 and c-Jun in Regulation ofβEts Gene PEA3 Cooperates with
doi: 10.1074/jbc.M311131200 originally published online February 27, 20042004, 279:20794-20806.J. Biol. Chem.
10.1074/jbc.M311131200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/279/20/20794.full.html#ref-list-1
This article cites 76 references, 34 of which can be accessed free at
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from