regulation of stiffness by skeletomotor reflexesphysio.northwestern.edu/docs/houk/houk79.pdf ·...
TRANSCRIPT

Anr~ Rcv. Physio£ 1979. 41:99-114Copyright © 1979 by Annual Reviews ln~ All rights reserved
REGULATION OF STIFFNESSBY SKELETOMOTOR REFLEXES
~1208
James C Houk
Department of Physiology, Northwestern University Medicaland Dental Schools, Chicago, Illinois 60611
Since the early 1950s when Merton (44) first introduced his "follow-upservo hypothesis" of movement control, the dominant opinion has held thatstretch and unloading reflexes function to control the length of a muscle inopposition to changes in mechanical load, thus providing load compensa-tion. In this article I review recent evidence indicating that this notion iswrong, or at least incomplete, and proceed to develop an alternative idea(47) that neither muscle length nor force are regulated as individual vari-ables, but that a property called stiffness (the ratio of force change to lengthchange) is maintained relatively constant by skeletomotor reflexes. Thisleads to a discussion of the actions of descending motor commands on thesegmental system, from which I conclude that these may be much lessversatile than was formerly believed.
The reader is referred to another review (55) and to Matthews’ mono-graph (42) for additional information.
AUTOGENETIC REFLEXES AND THE MOTORSERVO
Stretch and unloading reflexes are mediated by combined actions of severalautogenetic neural pathways. "Autogenetic" means that the stimulus ex-cites receptors located in the same muscle that is the target of the reflexresponse. The most important of these muscle receptors are the primary andsecondary endings in muscle spindles, sensitive to length change, and theGolgi tendon organs, sensitive to contractile force. The autogenetie circuitsappear to function as servoregulatory loops that convey continuouslygraded amounts of excitation and inhibition to the large (alpha) skeletomo-
990066-4278/79/0301-0099501.00
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.
1~ HOUK
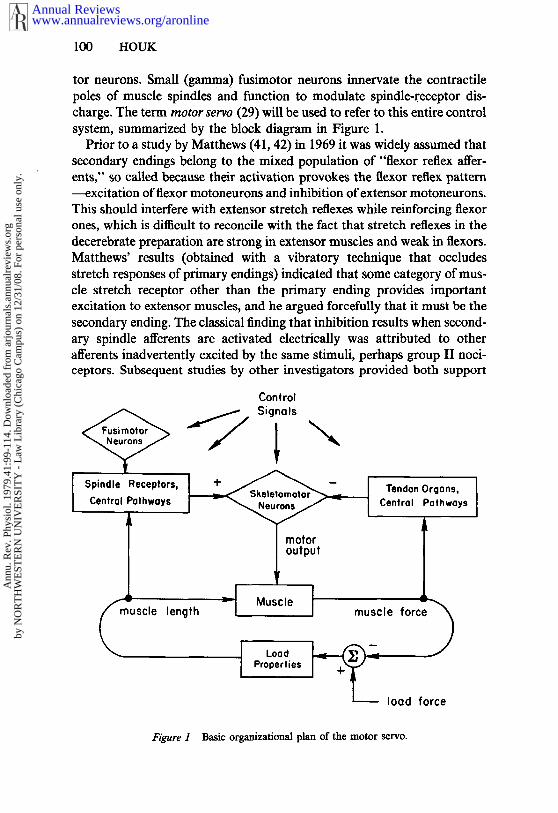
tor neurons. Small (gamma) fusimotor neurons innervate the contractilepoles of muscle spindles and function to modulate spindle-receptor dis-charge. The term motor servo (29) will be used to refer to this entire controlsystem, summarized by the block diagram in Figure 1.
Prior to a study by Matthews (41, 42) in 1969 it was widely assumed thatsecondary endings belong to the mixed population of "flexor reflex after-ents," so called because their activation provokes the flexor reflex pattern---excitation of flexor motoneurons and inhibition of extensor motoneurons.This should interfere with extensor stretch reflexes while reinforcing flexorones, which is difficult to reconcile with the fact that stretch reflexes in thedecerebrate preparation are strong in extensor muscles and weak in flexors.Matthews’ results (obtained with a vibratory technique that occludesstretch responses of primary endings) indicated that some category of mus-cle stretch receptor other than the primary ending provides importantexcitation to extensor muscles, and he argued forcefully that it must be thesecondary ending. The classical finding that inhibition results when second-ary spindle afferents are activated electrically was attributed to otherafferents inadvertently excited by the same stimuli, perhaps group II noci-ceptors. Subsequent studies by other investigators provided both support
ControlSignals
Spindle Receptors,
Central Pathways
~’~-u scle length
Ioutput
Muscle
LoadProperties
Tendon Organs,Central Pathways
muscle forc~ ~
J~load force
Figure 1 Basic organizational plan of the motor servo.
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

SKELETOMOTOR REGULATION OF STIFFNESS 101
for and opposition to Matthew’s conclusion. The reader may consult tworecent studies (34, 54) that support excitation by secondaries and alsosummarize other studies. The conclusion that the discharge of both primaryand secondary endings contributes excitation now seems firmly established,in the opinion of this author. However, the classical inhibitory actionsevoked by electrical stimulation, and in some cases by stretch, requirefurther explanation (cf 52).
The pathways from primary and secondary endings are treated com-monly in Figure 1, since both receptors are sensitive to muscle length andboth provoke retiex excitation. However, primary endings show an addi-tional sensitivity to the dynamic phase of length change, called dynamicresponsiveness, and they also show a much-enhanced sensitivity to smallchanges in muscle length (42). Recent studies indicate that both of thesespecial response features of primary endings derive from a common nonlin-ear property, which is a decreasing sensitivity to length change that occurswhenever the muscle is stretched beyond a certain point (25). Prior to thisstudy it usually had been assumed that the dynamic response is a velocityresponse that provides velocity feedback. In fact, the dependence onvelocity is both weak and highly nonlinear (53), and the role of the dynamiccomponent of feedback must be reevaluated (cf 30, 51).
Two features omitted from the block diagram of the motor servo mayhave to be added to it in the future. One concerns the motor fibers that sendbranches to both the main muscle and to spindles, referred to asskeletofusimotor (or beta) fibers. There is now increasing evidence of theimportance of skeletofusimotor innervation in mammals as well as lowervertebrates (24). The other omission concerns feedback to fusimotor neu-rons, the existence of which has been demonstrated in several laboratories(4, 10, 17, 32). What remains unclear at present is the specific modality andactual importance of these projections. The incorporation of both of thesefeatures into theories of muscle control has been considered elsewhere (28).Neither modification alters the theoretical conclusion drawn in the nextsection.
STRUCTURAL ANALYSIS OF THE MOTOR SERVO
The motor servo comprises three closed circuits (Figure 1), two neuralfeedback pathways, and one circuit representing the mechanical interactionbetween a muscle and its load. One of the feedback pathways, that fromspindle receptors, conveys information concerning muscle length, and itfollows that this loop will act to keep muscle length constant. The otherfeedback pathway, that from tendon organs, conveys information concern-ing muscle force, and it acts to keep force constant.
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

102 HOUK
In general, it is physically impossible to maintain both muscle length andforce constant when external loads vary; in this situation the action of thetwo feedback loops will oppose each other. For example, an increased loadforce will lengthen the muscle and cause muscular force to increase as themuscle is stretched out on its length-tension curve. The increased length willlead to excitation of motoneurons, whereas the increased force will lead toinhibition. It follows that the net regulatory action conveyed by skeletomo-tor output will depend on some relationship between force change andlength change and on the strength of the feedback from muscle spindles andtendon organs. A simple mathematical derivation (47) demonstrates thatthe change in skeletomotor output, the error signal of the motor servo,Should be proportional to the difference between a regulated stiffness andthe actual stiffness provided by the mechanical properties of the muscle,where stiffness has the units of force change divided by length change. Theregulated stiffness is determined by the ratio of the gain of length to forcefeedback.
It follows that the combination of spindle receptor and tendon organfeedback will tend to maintain the stiffness of the neuromuscular apparatusat some regulated level. If this level is high, due to a high gain of lengthfeedback and a low gain of force feedback, one could simply forget aboutforce feedback and treat muscle lcagth as the regulated variable of thesystem. However, if the regulated level of stiffness is intermediate in value,i.e. not appreciably different from the average stiffness arising from musclemechanical properties in the absence of reflex actions, one would concludethat stiffness, or its inverse, compliance, is the regulated property of themotor servo.
EVIDENCE FAVORING STIFFNESS, RATHERTHAN MUSCLE LENGTH, AS THE REGULATEDPROPERTY OF THE MOTOR SERVO
For the past several decades most investigators have assumed, either explic-itly or implicitly, that the regulated property of the stretch reflex is musclelength, or one of its derivatives such as velocity. The contemporary notionthat the stretch reflex is a mechanism for load compensation derives directlyfrom this assumption, since effective length (or velocity) regulation wouldprevent load changes from appreciably affecting movements or postures.
Recently several authors have reported that the load compensatingcapabilities of the stretch reflex system are rather modest. In particular,Vallbo (58) has shown, with a calculation based on spindle responses human subjects during voluntarily graded contractions, that the stiffness ofthe motor servo is rather low. Bizzi, Dev, Morasso & Polit (7) compared
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

SKELETOMOTOR REGULATION OF STIFFNESS 103
load compensation before and after dorsal rhizotomy and found only rela-tively small differences. Rack (50) has emphasized that muscle mechanicalproperties can contribute an appreciable amount of stiffness without theassistance of a reflex, and it has been shown that this muscular stiffness maynot differ substantially from the stiffness of an intact deeerebrate stretchreflex, at least in some situations (22, 49).
If the assumption that muscle length is the regulated property of themotor servo were correct, this evidence would indicate that the system israther ineffective and indeed of minor importance in normal motor control.Another interpretation is that the motor servo is actually designed to regu-late stiffness. All of the findings just discussed are compatible with the ideaof stiffness regulation, but they provide no direct test of it. Two recentstudies, one in the decerebrate cat (47) and the other in normal humans (8),were designed specifically to test the hypothesis of stiffness regulation andto assess its efficacy.
The rationale for these experiments is that the inherent mechanical stiff-ness of a muscle varies, depending on the amplitude and the direction oflength change (33, 47). Thus, if the function of the motor servo is to regulatestiffness, reflex action should vary to maintain stiffness approximately con-stant. In the animal experiments reflex action was assessed mechanically,as the difference between a net change in reflex force and the force changeattributable to the purely mechanical response of the muscle. In the humanexperiments reflex action was assessed more indirectly, by the changes inelectromyographic (EMG) activity. In both cases, it was found that reflexaction is greater during muscle stretch than it is during release, and thisdifference effectively compensates for an opposite asymmetry in the me-chanical response of the unregulated muscle. In the cat experiments thedependence of stiffness on the amplitude of length change was also studied.Muscular stiffness was shown to vary widely and in some instances actuallyto assume negative values. Although the stiffness of the motor servo was notabsolutely constant, the variations were small in comparison with those ofthe unregulated muscle, which indicated that the regulation of stiffness wasquite effective.
Although the results obtained with human subjects were qualitativelysimilar to those obtained in the decerebrate cat, there were two potentiallyimportant differences. Changes in the state of a decerebrate preparationwere sometimes accompanied by alterations in reflex stiffness, even whenit was assessed at the same initial length and force (46). These changes mustbe attributed to extrinsic neural signals that alter gain in the motoservopathways, and it is notable that such changes could not be demonstratedin the experiments with normal human subjects. The second difference wasthat the human reflex actions in response to unloading were often in a
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

1~ HOUK
direction that assisted rather than opposed length change. This observationprovides strong evidence against the hypothesis for length regulation, sug-gests a higher gain of tendon organ feedback in intact man than in decere.brate eat (8), and indicates that the regulated stiffness may be morecompliant than the inherent stiffness of the muscle under some circum-stances.
The results just described test the efficacy of stiffness regulation when theamplitude and direction of length change are varied. Another test of thetheory would be to explore differences in initial force, since muscular stiff-ness is known to increase in approximate proportion with initial force (47)and effective servo action would be expected to diminish this dependence.
Matthews (39, 40) studied the relation between muscle force and lengthunder a wide range of conditions by applying stretch at a low and constantvelocity to the soleus muscle of the decerebrate cat. He showed that thestiffness (the slope of the stretch reflex curves) was low near the thresholdof the reflex, increased as the stretch progressed, and then tended to reacha constant value as indicated by a linear relationship between muscle forceand length. The dependence of stiffness on initial force has also been as-sessed from incremental responses to length change, with similar results(31). The evidence from both of these studies suggests that the stiffness the motor servo remains relatively constant whenever the initial force ex-ceeds approximately 15% of its maximal value, and decreases progressivelywith lower forces. The latter authors suggested that the more appreciablevariation at low initial forces might be accentuated by the presence of a"recruitment nonlinearity," which was postulated to arise from the recruit-ment of motor units in the order small to large [the size principle (26)]. Thisfactor would compound with the dependence of muscular stiffness on initialforce mentioned earlier. The fact that the stiffness of the motor servodepends so little on initial force (in the range 15-100% of maximum) good evidence in support of stiffness regulation.
The relation of stiffness to initial force appears to be similar in humansto that demonstrated in the decerebrate eat, although most of the availablemeasurements were made in the range of low initial forces where stiffnessvaries most (5, 15, 37).
SIMPLIFIED MODELS OF THE MOTOR SERVO
I now consider how these results relate to more general problems of motorcontrol. Here it is useful to have a summary model of the motor servo thataccounts for its net responses to mechanical disturbances and to controlsignals that may be sent to it from other regions of the central nervoussystem.
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.
SKELETOMOTOR REGULATION OF STIFFNESS 105
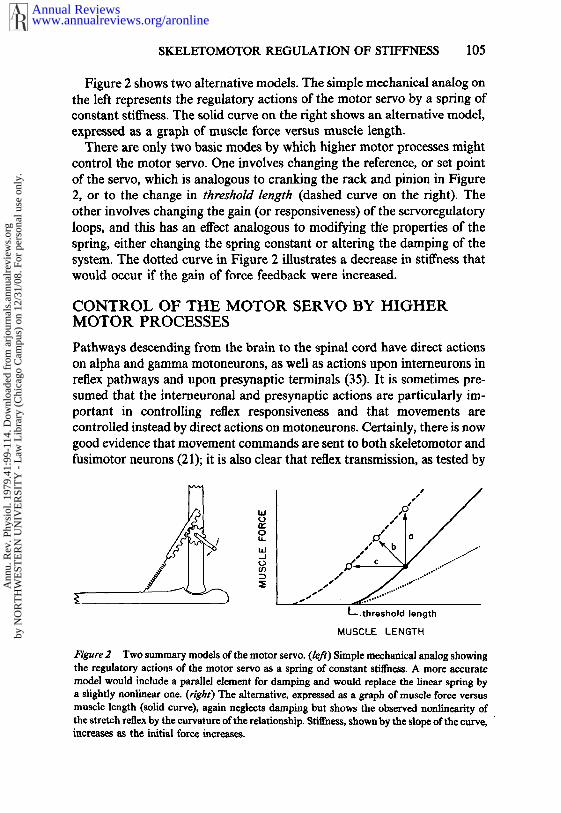
Figure 2 shows two alternative models. The simple mechanical analog onthe left represents the regulatory actions of the motor servo by a spring ofconstant stiffness. The solid curve on the right shows an alternative model,expressed as a graph of muscle force versus muscle length.
There are only two basic modes by which higher motor processes mightcontrol the motor servo. One involves changing the reference, or set pointof the servo, which is analogous to cranking the rack and pinion in Figure2, or to the change in threshoM length (dashed curve on the righ0. Theother involves changing the gain (or responsiveness) of the servoregulatoryloops, and this has an effect analogous to modifying tlle properties of thespring, either changing the spring constant or altering the damping of thesystem. The dotted curve in Figure 2 illustrates a decrease in stiffness thatwould occur if the gain of force feedback were increased.
CONTROL OF THE MOTOR SERVO BY HIGHERMOTOR PROCESSES
Pathways descending from the brain to the spinal cord have direct actionson alpha and gamma motoncurons, as well as actions upon intcrncurons inreflex pathways and upon presynaptic terminals (35). It is sometimes prc-sumcd that thc intcrncuronal and prcsynaptic actions arc particularly im-portant in controlling reflex responsiveness and that movcments arecontrolled instead by direct actions on motoneurons. Certainly, there is nowgood evidence that movement commands are sent to both skclctomotor andfusimotor neurons (2 I); it is also clcar that rcflcx transmission, as tcstcd
L-,threshold length
MUSCLE LENGTH
t~’gure 2 Two summary modds of the motor scrvo. (left) Simple mechanical analog showingthe regulatory actions of the motor servo as a spring of constant stiffness. A more accuratemodel would include a parallel element for damping and would replace the linear spring bya slightly nonlinear one. (right) The alternative, expressed as a graph of muscle force versusmuscle length (solid curve), again neglects damping but shows the observed nonllncarity the stretch reflex by the curvature of the relationship. Stiffness, shown by the slope of the curve,increases as the initial force increases.
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

106 HOUK
electrophysiological techniques, can be altered by descending activity.However, changes in reflex transmission only demonstrate the anatomicalconvergence of descending and reflex pathways; the normal actions of aconvergent circuit might be ones that change the reference of the motorservo by adding to an ongoing input from the periphery (a change inthreshold length), rather than ones that alter the gain of the ,reflex pathway(a change in stiffness). This question can only be answered by,studies thatemploy natural stimuli and as much as possibIe preserve normal activitystates of the spinal cord. The decerebrate cat preparation, during ongoingreflex activity, approximates these requirements, but the best tests are madein intact animal and human subjects.
Studies in decerebrate animals in which electrical stimulation mimicsmotor commands show that these sources of excitatory and inhibitory inputsimply bias the threshold of the stretch reflex (the alteration in thresholdlength shown by the dashed curve in Figure 2) without appreciably alteringits slope (16, 40). In terms of the analog of the motor servo (Figure 2), theseactions crank the rack and pinion without altering stiffness. Of course, thereis no guarantee that the electrical stimuli used in these studies are actuallyanalogous to normal motor commands.
In experiments with humari subjects it is easy to provoke authentic motorcommands by asking the subject to move to a new position, but it is moredit~cult to ascertain that responses to mechanical disturbances applied atthat new position are, in fact, purely reflexive. Asatryan and Fel’dman (5)introduced the procedure of instructing their subjects not to intervenevoluntarily as a means for obtaining responses to mechanical disturbancesthat were presumed to be reflexive. Fairly conclusive evidence that this canbe accomplished has been given recently by Crago, Houk & Hasan (8).
Asatryan and Fel’dman (5) showed that when the subject is~asked move to a new initial position the relations between elbow moment andangle, described in an earlier section, are shifted along the abscissa of thetorque versus angl~ graph with little additional change in the relation.Fel’dman (15) also demonstrated by mechanical analysis that this shift wasaccounted for by assuming that the thresholds of the stretch reflexes of themuscles acting about the elbow were biased in the manner discussed above.The effects of altered loads on head movements described recently by Bizzi,Polit & Morasso (6) are also well .accounted for on this basis. Thus, theavailable evidence indicates that.motor commands alter threshold length,an effect analogous to cranking the rack and pinion of the motoservo analog.
One concludes.that descending motor commands control neither mus-cle forces nor movements but rather~the threshold length of the springthat characterizes the properties of the,motor servo (the threshold of thestretch reflex). A controlled change in threshold length leads to movement
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

SKELETOMOTOR REGULATION OF STIFFNESS 107
if (or when) the mechanical load yields; if the load is isometric, the samemotor command controls the force of contraction. This effect of load on theperipheral outcome of a given central motor command is illustrated inFigure 2 by the three trajectories. The one labeled a corresponds to pullingon an immovable object, b to pulling on a spring, and c to pulling againsta constant force load. The absence of load compensation is illustrated bythe fact that the length change resulting from the same motor commanddiffers so markedly in the three cases.
ABSENCE OF EVIDENCE FOR THE CONTROLOF MOTOSERVO GAIN
Other types of neural control signals sent by central motor processes mightoperate by controlling the gain of the reflex loops, which would alter thestiffness that is then regulated by motoservo action. This mode of controlcould form the efferent mechanism of an adaptive control process in thebrain that adjusts the mechanical impedance of the neuromuscular ap-paratus to match the requirements of the task at hand. A rigid control ofmuscle length might be desirable in many tasks, while a more compliantsystem would improve the suspension of the body during locomotion overrough terrain (el 28, 36). Thus, there is reason enough to expect gaincontrol, and, indeed, numerous authors have suggested that it occurs.
Most of the available studies that are thought to demonstrate gain controlsuffer from technical limitations. Unnatural stimuli and abnormal spinalstates have already been mentioned. A second problem is that nonlinearitiesin the properties of motoservo components can be confused with changesin gain thought to be controlled by a central adaptive mechanism. Althoughgain variations having this origin may subserve some useful function [e.g.compensation for fatigue, as suggested in (37)], they are of no direct concernto ccntral motor processes, since they cannot bc controlled in any indepen-dent fashion but occur in an obligatory manner along with motor com-mands that control movements. Thc possibility that observed changes inresponsiveness result from motoservo nonlinearities is minimized if initiallength, force, and motoneuronal output arc held constant while testing fora gain change of central origin.
A. third problem (a special case of the second) arises if the relevantmotoncuronal pools are biased below the threshold for motor output, whichhappcns whenevcr a limb is initially unloaded. The experimenter then losescontrol over the initial excitability of the motoncurons. Subthresholdchanges in excitability produce apparent changes in reflex responsiveness,but these changes can bc duc either to motor commands that do not reachthreshold or to control signals that actually change gain. While changes in
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

108 HOUK
responsiveness under passive conditions (e.g. 12, 14) cannot be taken evidence for gain control, they may contribute to the preparedness of asubject for a transition from an inactive to an active state.
A fourth and major problem concerns the admixture of reaction-timemovements with motoservo responses. The former are not servoregulatoryactions, yet their presence in one response and absence in another might beinterpreted (falsely) as the result of a change in motoservo gain. Since themovements are delayed by a reaction time, many authors have simplyrestricted their analysis to events occurring at times prior to the supposedonset of a reaction. The two disadvantages of this approach are that itneglects later phases of reflex response and it relies heavily on a validmeasure of the minimal reaction time. This measure is dit~cult to obtainsince reaction times depend upon several parameters, such as the intensityand morality of the stimulus, the number of choices in the task, and thedegree of attention and practice (59). The usual procedure in stretch-reflexstudies has been to measure a reaction time to stimuli other than musclestretch, or to small tendon taps that provoke no appreciable reflex, and toassume that the latency of a reaction in response to a large stretch wouldbe no shorter (e.g. 23, 37, 43). Unfortunately this is not true.
An alternative procedure has been proposed by Evarts & Granit (12),who suggested that the time at which EMG activity recommences afterinitially being silenced by an unloading reflex is indicative of the onset ofa reaction-time movement ("intended movement" in their terminology).They reported a latency as brief as 70 reset. A potential problem with thismethod is that an enhancement of EMG activity during unloading mightalso be explained on the basis of a segmental reflex action (3, 8). Further-more, the method does not provide a means for studying later (beyond 70msee) phases of reflex response.
In addition to a delayed onset, reaction-time movements have severalspecial properties useful in distinguishing them from motoservo actions (8).For example, the latency of a reaction becomes longer and more variablewhen the subject is required to choose between alternative responses, ascontrasted with performance in a simple reaction-time task. Also, subjectsin a choice-reaction task will occasionally make errors; if the task is appro-priately designed, the errors will appear as responses of opposite direction,which are easily detected in movement traces.
Very few studies of adaptive responsiveness meet all of the objectionsraised in this section; if only those that do are considered (5, 8), one findsno evidence at present that demonstrates an ability of the central nervoussystem to control the gain of the motor servo in an adaptive manner. Thisresult is surprising since neural mechanisms for gain change are available,as evidenced by authentic alterations that have been observed in reducedanimal preparations (46, 52) and that may occur also in human pathology.
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

SKELETOMOTOR REGULATION OF STIFFNESS 109
While it is premature to conclude that there are no situations in whichnormal humans adaptively alter the gain of their motoservo loops, theavailable evidence does suggest that gain change is not a usual mode ofcontrol. Ordinarily the motor servo operates in a rather stereotypic mannerwhen it is brought to action by a central motor command.
EVIDENCE CONCERNING SUPRASPINALSERVOREGULATORY LOOPS
The recent hypothesis that a supraspinal servo loop is largely responsiblefor the stretch reflex can be traced to Hammond’s (23) observations that thefirst major temporal component of EMG response to an abrupt stretch hasa latency of 50-60 msec and that its amplitude can be influenced by priorinstructions. Hammond suggested that this latency is too long to representsegmental reflex action and too short to represent a reaction-time move-ment. He explained both the latency and the modifiability by proposing along servo loop that ascends to the brain where a modification of the gainof reflex transmission was supposed to occur. This interpretation of theso-called "long-loop reflex" has been accepted by several subsequent investi-gators. For example, the same arguments led Phillips (48) to propose thata pathway from primary spindle receptors to cortical area 3a constitutes theafferent limb of a transcortical servo loop that was supposed to providenegative feedback compensation for changes in load. Evarts (11) supportedPhil!ips’ proposal in reporting that single pyramidal tract neurons in behav-ing primates responded at short latency to load perturbations, and, later,that the responses of these neurons are subject to the influence of ’set’ (13).
We now know that reaction times can be as short as 50-60 msec in man(of 29) and even shorter in trained eats (19). The modifiable aspect of stretchresponse seems to be accounted for by the versatility of the reaction-timemovements superimposed upon motoservo actions (8, 29). It is also wellknown that the segmental reflex pathways that participate in the operationsof the motor servo are both monosynaptic and polysynaptic, and thattransmission through these pathways, particularly when natural stimuli areemployed, will not necessarily be as rapid as conduction through themonosynaptie pathway (42).
A second influential observation by Hammond (23) indicated that themonosynaptic component of EMG response is not followed by a large forceresponse. Because of this, and because it was assumed that the later compo-nent at 50 msec was unrelated to spinal mechanisms, Hammond suggestedthat the segmental stretch reflex might be unimportant in man, perhapsbeing replaced by the supraspinal servo loop with adjustable gain mentionedearlier. This proposal, reinforced by Phillips (48) and by Melvill, Jones Watt (43), has been dit~cult to reconcile with the large body of literature
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

110 HOUK
illustrating the importance of the segmental mechanisms (42). One shouldconsider first that the size of the stretch reflex response is not an adequatemeasure of motoservo action, since the response to a load change reflectsinhibitory as well as excitatory components. Also, the segmental reflexresponse in man undoubtedly continues beyond the time of the monosynap-tic peak, which only signals its onset.
Some arguments in support of a supraspinal servo loop are independentof the previous criticisms. Evarts (11) pointed out that the initial perturba-tion response of some pyramidal tract neurons occurs sufficiently early topostulate that it is conducted to spinal motoneurons in advance of thesignals that initiate reaction-time movements. This short-duration phase ofdischarge is sensitive to the direction of the perturbation, whereas a later,longer-lasting phase is sensitive to the direction of the intended movement(14). There is also evidence in support of the assumption that the perturba-tion-sensitive neurons project to the relevant spinal motoneurons (45).While this and other evidence is consistent with the hypothesis that atranscortical servo loop contributes to motoservo action, the efficacy of theproposed loop has not been demonstrated, nor has it been compared withthe efficacy of segmental pathways.
Marsden, Merton & Morton (38) reported that an early peak in the EMGresponse to stretch has longer latencies in muscles of the lower limb thanin those of the upper limb. They pointed out that this observation is consis-tent with a supraspinal pathway; however, there are numerous problemsassociated with the interpretation of EMG peaks. Ghez & Shinoda (18)showed recently that the peaks of longer latency formerly attributed tosupraspinal pathways are, in fact, present also in spinalized cats.
Another argument is based on the effects of lesions and cooling of supras-pinal structures on stretch reflex function (e.g. I, 56, 57). The difficulty that changes in motoservo actions can be interpreted in two different ways.They may indicate interference with supraspinal loops or they may reflecta depression or enhancement of purely segmental mechanisms caused by aremoval of the normal tonic activity in descending pathways. The latterinterpretation is reinforced by the observations that normal stretch reflexes,which disappear after cord transection, are observed in chronic spinalanimals and in acute preparations after the administration of 5-HTP orL-DOPA (2, 20). These drugs are known to reverse the generalized depres-sion of spinal reflex transmission caused by interruption of descendingpathways.
In an earlier section I compared motoservo actions in decerebrate ani-mals (which appear not to differ from those in spinal animals as mentionedabove) with those in normal subjects, and concluded that there is a greatdeal of similarity between the two. The only major difference is that human
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

SKELETOMOTOR REGULATION OF STIFFNESS 111
motoservo action sometimes assists length change, whereas this has notbeen observed in the deeerebrate cat preparation. A likely explanation ofthis difference is that inhibitory feedback from Golgi tendon organs has ahigher gain in intact man than in the decerebrate cat (8). While the highgain~ might be attributed to an inhibitory supraspinal loop, a more par-simonious explanation is that the Ib segmental pathway is relatively de-
.pressed in the decerebrate, as contrasted with intact animals; moreover,there is independent evidence favoring this interpretation (9, 27, 31).
In summary, it appears likely that the motor servo is in large part asegmental reflex mechanism. If this reflex system does employ supraspinalloops, it is not clear what they add to the servoregulatory actions knownto be mediated by segmental pathways. Formerly it was believed that im-proved load compensation and adaptive responsiveness might be added, butit now seems clear that these features result from reaction-time responsesrather than from the servoregulatory actions of the stretch reflex.
CONCLUSIONS
Stretch and unloading reflexes are controlled by the combined actions ofseveral autogenetic reflex pathways, which are part of a negative feedbacksystem referred to as the motor servo. This fundamental skeletomotor reflexsystem is organized segmentally within the spinal cord; recent claims thatthe segmental system is dominated by supraspinal servo loops are found tobe weak when evaluated critically.
The older view that this system functions to regulate muscle length andto provide load compensation is not supported by recent experimentalstudies, nor does it have a sound theoretical basis. Length feedback frommuscle spindles when combined with force feedback from tendon organsshould give rise to stiffness regulation, and experimental studies have dem-onstrated that motoservo actions are, in fact, effective in compensating forknown variations in the inherent mechanical stiffness of skeletal muscle.
Descending motor commands could act to shift the threshold length ofthe motor servo (i.e. the threshold of the stretch reflex), and they could actalso to modify the stiffness regulated by length and force feedback (anadaptive type of control that requires a change in gain of the feedbackpathways). Current experimental evidence clearly supports the former ac-tion, while there is no good evidence that the latter mode of control isactually used by normal intact subjects. However, gain changes probablydo occur in pathology.
These results provide an answer to the often-posed question concerningwhich mechanical variable, muscle length or force, or some derivative ofthese variables, is actually controlled by descending motor commands. The
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

112 HOUK
answer appears to be none of these individually, since a controlled change
in threshold length acts to shift the entire relationship between length and
force that is then regulated locally by length and force feedback. The actual
values of length and force resulting from a central motor command depend
on the mechanical load to which the muscle is attached.One advantage of stiffness regulation is that it provides a spring-like
interface between the body and its mechanical environment. The compli-
ance of this interface probably serves to absorb the impacts of abrupt
changes in load, thus attenuating their transmission to the body and head.
For example, in the standing posture with bent knees, stiffness regulationprobably insures a good suspension system for the body.
The apparent absence of an adaptive control of stiffness might be viewed
as a disadvantage, since flexibility is clearly sacrificed. On the other hand,
the relative constancy of motoservo properties should simplify the task of
higher motor control processes that must generate appropriate motor com-
mands. Such commands must be adjusted to account for the properties of
the system they control, and, if these properties are constant, the adjust-
ments can be made routine.
Literature Cited
1. Adam, J., Marsden, C. D., Merton, P.A., Morton, H. B. 1975. The effect oflesions in the internal capsule and thesensodmotor cortex on servo action inthe human thumb. J. Physiol. London254:27P
2. Ahlman, H., Gdllner, S., Udo, M. 1971.The effect of 5-HTP on the staticfusimotor activity and the tonic stretchreflex of an extensor muscle. Brain Re~27:393-96
3. Alston, W., Angel, R. W., Fink, F. S.,Hoffman, W. W. 1967. Motor activityfollowing the silent period in humanmuscle. J. Physiol. London 190:189-202
4. Appelberg, B., Johansson, H., Kalis-tratov, G. 1977. The inituencc of GroupII muscle afferents and low thresholdskin aiferents on dynamic fusimotorneurons to the triceps surae of the cat.Brain Res. 132:153-58
5. Asatryan, D. G., Fel’dman, A. G. 1965.Functional tuning of the nervous sys-tem with control of movement or main-tenance of posture-I. Mechanographieanalysis of the work of the joint on exe-cution of a postural task. Biophysics10:925-35
6. Bizzi, E., Polit, A., Morasso, P. 1976.Mechanisms underlying achievement offinal head position. J. Neurophysiol.39:435-44
7. Bizzi, E., Dev, P., Morasso, P., Polit, A.1978. Effect of load disturbances duringcentrally initiated movements. ZNeurophysiol. 41:542-57
8. Crago, P. E., Houk, J. C., Hasan, Z.1976. Regulatory actions of the humanstretch reflex. J. Neurophysiol. 39:925-35
9. Eccles, R. M., Lundberg, A. 1959. Su-praspinal control of interneurons me-diating spinal reflexes. J. Physiol. Lon-don 147:565-84
10. Ellaway, P. H., Trott, J. R. 1976. Reflexconnections from muscle stretch recep-tors to their own fusimotor neurons.Progr. Brain Res. 44:113-21
11. Evarts, E. V. 1973. Motor cortexreflexes associated with learned move-ments. Science 179:501-3
12. Evarts, E. V., Granit, R. 1976. Rela-tions of reflexes and intended move-ments. Progr. Brain Res. 44:1-14
13. Evarts, E. V., Tanji, J. 1974. Gating ofmotor cortex reflexes by prior instruc-tion. Brain Rez 71:479-94
14. Evarts, E. V., Tanji, J. 1976. Reflex andintended responses in motor pyramidaltract neurons of monkey. J. Neuro-physiol. 39:1069-80
15. Fel’dman, A. G. 1966. Functional tun-ing of the nervous system with controlof movement or maintenance of a
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

SKELETOMOTOR REGULATION OF STIFFNESS 113
steady posture. II. Controllable param-eters of the muscle. Biophysics11:565-78
16. Fel’dman, A. G., Orlovsky, G. N. 1972.The influence of different descendingsystems on the tonic stretch reflex in thecat. Exp. Neurol. 37:481-94
17. Fromm, C., Haase, J., Noth, J. 1974.Length dependent autogenetie inhibi-tion of extensor gamma motoneurons inthe decerebrate cat. Pfluegers Arch.346:251-62
18. Ghez, C., Shinoda, Y. 1978. Spinalmechanisms of the functional stretchreflex. Brain Res. 3~2:55-6g
19. Ghez, C., Vicario, D. 1978. The controlof rapid limb movement in the cat. I.Response latency. Brain Rez 33:173-89
20. Goodwin, G. M., MeGrath, G. J., Mat-thews, P. B. C. 1973. The tonic vibra-tion reflex seen in the acute spinal eatafter treatment with DOPA. Brain Res.49:463-66
21. C~ranit, R. 1975. The functional role ofthe muscle spindles--facts and hypoth-eses. Brain 98:531-56
22. Gdllner, S. 1972. A role for muscularstiffness in meeting the changing pos-tural and locomotor requirements forforce development by the ankle exten-sors. Acta Physio£ Scand. 86:92-108
23. Hammond, P.H. 1960. An experimen-tal study of servo action in human mus-cular control. In Proc III Int. Congr.Med. Electron., pp. 190-99. London:Inst. Electr. Eng.
24. Harker, D. W., Jami, L, Laporte, Y.,Petit, J. 1977. Fast-conduction skeleto-fusimotor axons supplying intrafusalchain fibers in the cat peroneus tertiusmuscle. J. Neurophysiol. 40:791-99
25. Hasan, Z., Houk, J. C. 1975. The transi-tion in the sensitivity of spindle recep-tors that occurs when the muscle isstretched more than a fraction of a mil-limeter. J.. NeurophysioL 38:673-89
26. Henneman, E., Somjen, G., Carpenter,D. O. 1965. Functional signifieanee ofcell size in spinal motoneurons. J.Neurophysiol. 28:599-620
27. Hongo, T., Jankowska, E., Lundberg,A. 1969. The rubrospinal tract. II. Fa-cilitation of interneuronal-transmissionin reflex paths to motoneurons. Exp.Brain Re~ 7:365-91
28. Houk, J. C. 1972. The phylogeny ofmuscular control configurations. InBiocybernetics, ed. H. Drischel, P. Dett-mar, 4:125-44. Jena: Fisher
29. Houk, J. C. 1978. Participation of reflexmechanisms and reaction-time pro-
cesses in the compensatory adjustmentsto mechanical disturbances. Progr. Clir~Neurophysiol. 4:193-215
30. Houk, J. C., Rymer, W. Z., Crago, P. E.1977. Complex velocity dependence ofthe eleetromyographie component ofthe stretch reflex. Proc. XXVl11 lnt.Congr. Physiol. Sci., Paris. (Abstr.:~:981)
31. Houk, J. C., Singer, J. J., Goldman, M.R. 1970. An evaluation of length andforce feedback to soleus muscles ofdeeerebrate eats. J. Neurophysiol. 33:784-811
32. Hunt, C. C. 1951. The reflex activity ofmammalian small-nerve fibres. J.Physiol. London 115:456-69
33. Joyce, 12. (L, Rack, P. M. H., Westbury,D. F. 1969. The mechanical propertiesof cat soleus muscle during controlledlengthening and shortening movements.Z PhysioL London 204:461-74
34. Kanda, K., Rymer, W. Z. 1977. An esti-mate of the secondary spindle receptorafferent contribution to the stretchreflex in extensor muscle of the deeere-brate eat. J. Physiol. London 264:63-87
35. Lundberg, A. 1967. The supraspinalcontrol of transmission in spinal reflexpathways. Electroencephalogr. Clin.Neurophysiol. Suppl. 25:35-46
36. Lundberg, A. 1969. Reflex control ofstepping. No.sen Mere. Lect., I~., pp. 1-42. Oslo: Uaiversitetsfodaget
37. Marsden, C. D., Merton, P. A., Mor-ton, H. B. 1976. Servo action in the hu-man thumb. £ Physiol. London257:1-44
38. Marsden, (2. D., Merton, P. A., Mor-ton, H. B. 1976. Stretch reflexes andservo actions in a variety of humanmuscles. J. Physiol. London 259:531--60
39. Matthews, P. B. C. 1959. The depen-dence of tension upon extension in thestretch reflex of the soleus muscle of thedeeerebrate cat. J. Physiol. London147:521-46
40. Matthews, P. B. C. 1959. A study ofcertain factors influencing the stretchreflex of the decerebrate eat. Z Physiol.London 147:547-64
41. Matthews, P. B. C. 1969. Evidence thatthe secondary as well as the primaryendings of the muscle spindles may beresponsible for the tonic stretch reflex ofthe decerebrate eat. Z Physiol. London204:365-93
42. Matthews, P. B. C. 1972. MammalianMuscle Receptors and Their Central Ac-tion~ Baltimore: Williams and Wilkins.
43. Melvill Jones, G., Watt, D. G. D. 197i.
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

114 HOUK
Observations on the control of steppingand hopping movements in man. J.Physiol. London 291:709-27
44. Merton, P. A. 1953. Speculations on theservo control of movement. The SpinalCord, pp. 183-98. Boston: Little,Brown
45. Murphy, J. T., Wong, Y. C., Kwan, H.C. 1975. Afferent-efferent linkages inmotor cortex for single forelimb mus-cles. J. Neurophysiol. 38:990-1014
46. Nichols, T. R. 1974. Soleus muscle stiff-ness and its reflex control. Ph.D. thesis,Harvard University, Cambridge, Mass.
47. Nichols, T. R., Houk, J. C. 1976. Theimprovement in linearity and the regu-lation of stiffness that results from theactions of the stretch reflex. J. Neuro-physiol. 39:119--42
48. Phillips, C. G. 1969. Motor apparatusof the baboon’s hand. Proc. R. Soc. Lon-don Ser. B 173:141-74
49. Pompeiano, O. 1960. Alpha types of"re-lease" studied in tension-extension dia-grams from cat’s forelimb triceps mus-cle. ,~rch. Ital. Biol. 98:92-117
50. Rack, P. M. H. 1970. The significanceof mechanical properties of muscle inthe reflex control of posture. In Excita-tory Synaptic Mechanisms, ed. P. An-dersen, J. K. S. Jansen, pp. 317-21.Oslo: Universitetsforlaget
51. Rymer, W. Z., Houk, L C., Crago, P. E.1976. Vibratory occlusion of compensa-tion for muscle yielding. Neuroscl~lbstr. #758
52. Rymer, W. Z., Houk, J. C., Crago, P. E.
1977. Mechanisms of the clasp-knifereflex. Neuroscl. ~4bstr. #887
53. Rymer, W. Z., Houk, J. C., Crago, P. E.1977. The relation between dynamic re-sponse and velocity sensitivity for mus-cle spindle receptors. Proc. XXVIIIIn~Congr. Physiol. ScL Paris (Abstr.# 1922)
54. Stauffer, E. K., Watt, D. G. D., Taylor,A., Reinking, R. M., Stuart, D. B. 1976.Analysis of muscle receptor connec-tions by spike-triggered averaging. 2.Spindle group II afferents. J. Neuro-physiol. 39:1393-1402
55. Stein, R. B. 1974. Peripheral control ofmovement. Physiol. Rev. 54:215-43
56. Tatton, W. G., Forner, S. D., Gerstein,G. L., Chambers, W. W., Liu, C. N.1975. The effect of postcentral corticallesions on motor respoases to suddenupper limb displacement in monkeys.Brain Rex 96:108--13
57. Texzuolo, C. A., Viviani, P. 1974.Movements’ parameters and EMG ac-tivities during some simple motor tasksin normal subjects and cerebellar pa-tients. In Cerebellum, Epilepsy and Be-havior, ed. I. Cooper, M. Riklan, R. S.Snider, pp. 173-215. London: Perga-mon
58. Vallbo, A. B. 1974. Human musclespindle discharge during isometric vol-untary contractions. Amplitude rela-tions between spindle frequency andtorque. ,4cta Physiol. Scand. 90:319-36
59. Welford, A. T. 1968. Fundamentals ofSkill. London: Methuen. pp. 27-104
www.annualreviews.org/aronlineAnnual Reviews
Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.

Ann
u. R
ev. P
hysi
ol. 1
979.
41:9
9-11
4. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by N
OR
TH
WE
STE
RN
UN
IVE
RSI
TY
- L
aw L
ibra
ry (
Chi
cago
Cam
pus)
on
12/3
1/08
. For
per
sona
l use
onl
y.