regulatory role of nkt cells on effects of 1,25(oh)
TRANSCRIPT

The Pennsylvania State University
The Graduate School
The Huck Institutes of Life Sciences
REGULATORY ROLE OF NKT CELLS ON EFFECTS OF
1,25(OH)2D3 IN MICE WITH EAE
A Thesis in
Cell and Developmental Biology
by
Jun Zhao
2012 Jun Zhao
Submitted in Partial Fulfillment of the Requirements
for the Degree of
Master of Science
August 2012

ii
The thesis of Jun Zhao was reviewed and approved* by the following:
Margherita T. Cantorna Professor of Molecular Immunology Thesis Advisor Chair of Intercollege Graduate Degree Program in Immunology and
Infectious Diseases
Na Xiong Associate Professor of Veterinary and Biomedical Sciences
Pamela Hankey Associate Professor of Immunology
Zhi-chun Lai Professor of Biology, Biochemistry and Molecular Biology Chair of Intercollege Graduate Degree Program in Cell and
Developmental Biology
*Signatures are on file in the Graduate School

iii
ABSTRACT
Vitamin D is an immune regulator and has been shown to be involved in the
development and treatment of autoimmune diseases such as multiple sclerosis (MS). The
active form of vitamin D, 1,25(OH)2D3, has been shown to suppress experimental
autoimmune encephalomyelitis (EAE). Invariant Natural Kill T (iNKT) cells have been
proven to be an important suppressor of EAE. The development of iNKT cells was
impaired due to vitamin D and or vitamin D receptor (VDR) deficiency. 1,25(OH)2D3
had less effects in protecting CD1d knockout (KO) mice from EAE than in WT. CD1d
KO mice with 1,25(OH)2D3 supplementation had significantly higher incidence of EAE
and developed significantly higher EAE scores than WT mice with 1,25(OH)2D3
treatment. 1,25(OH)2D3 was also less effective in preventing Jα18 KO mice against EAE
than in WT. Jα18 KO mice on 1,25(OH)2D3 treatment had significantly higher incidence
of EAE and significantly higher cumulative EAE scores than WT mice with 1,25(OH)2D3
treatment. Either 1,25(OH)2D3 treatment or -galactosylceramide (-GalCer)
administration failed to prevent EAE development in Interleukin-4 (IL-4) KO mice.
These data suggests that the protective effects of 1,25(OH)2D3 against EAE might be
regulated by iNKT cells and IL-4 might be a positive regulator in the beneficial effects of
either 1,25(OH)2D3 or -GalCer on EAE inhibition.

iv
TABLE OF CONTENT
Chapter 1 Introduction ........................................................................................................ 1
1.1 Vitamin D............................................................................................................ 1
1.1.1 Metabolism and classical functions of Vitamin D .................................... 1
1.1.2 Vitamin D in immune regulation ............................................................... 1
1.1.3 Vitamin D and NKT cells .......................................................................... 2
1.2 Multiple Sclerosis .................................................................................................. 3
1.2.1 Background and epidemiology .................................................................. 3
1.2.2 Vitamin D and multiple sclerosis .............................................................. 4
1.2.3 EAE and vitamin D ................................................................................... 4
1.2.4 EAE and NKT cells ................................................................................... 6
1.3 Conclusion ............................................................................................................. 7
Chapter 2 Materials and Methods ....................................................................................... 8
2.1 Mice and diets ........................................................................................................ 8
2.2 αGalCer stimulation ............................................................................................... 8
2.3 EAE induction........................................................................................................ 8
2.4 EAE symptoms ...................................................................................................... 9
2.5 Statistical analysis .................................................................................................. 9
Chapter 3 Results .............................................................................................................. 11

v
3.1 1,25(OH)2D3 is ineffective at suppressing EAE in CD1d KO mice .................... 11
3.2 1,25(OH)2D3 is ineffective at suppressing EAE in Jα18 KO mice ...................... 12
3.3 Either 1,25(OH)2D3 or α-GalCer fails to prevent EAE in IL-4KO mice ............. 13
Chapter 4 Discussion ........................................................................................................ 15
References ......................................................................................................................... 25

1
Chapter 1 Introduction
1.1 Vitamin D
1.1.1 Metabolism and classical functions of Vitamin D
Vitamin D is a fat soluble vitamin which can be produced in the body or absorbed
through diets or supplements. The precursor of vitamin D, seven-dehydrocholesterol, is
converted into cholecalciferol (vitamin D3) in the skin by UVB light from sunlight
exposure. Vitamin D3 is then converted into 25-hydroxy vitamin D3 [25(OH)D3] in the
liver, which is the major circulating form of vitamin D in human body. 25-hydroxy
vitamin D3 is then hydroxylated by the Cyp27B1 encoded enzyme 1-hydroxylase into
the active form of vitamin D, 1,25-dihydroxyvitamin D3 [1,25(OH)2D3] in the kidney
(DeLuca 2004).
Vitamin D is known for its classical functions of maintaining calcium and phosphate
homeostasis (DeLuca 2004). Vitamin D could regulate absorption and reabsorption of
calcium and phosphate through tissues such as intestines, bones, kidneys, and the
parathyroid gland. Vitamin D could also directly regulate parathyroid hormone
metabolism and itself (Feldman 1999).
1.1.2 Vitamin D in immune regulation
The vitamin D receptor (VDR) is a member of the class II steroid hormone
superfamily of nuclear receptors (Masuyama 1997). 1,25(OH)2D3 binds to the VDR and

2
the formed complex directly binds to DNA to function as a transcriptional regulator in
target cells (Jones 1998). VDR is expressed in almost all tissues and many types of cells
including CD4+ and CD8+ T cells, B cells (Veldman 2000), macrophages (Helming
2005) and monocytes (Provvedini 1983). This suggests that vitamin D can directly
regulate different aspects of the immune system.
1,25(OH)2D3 could directly affect T helper1/T helper2 (Th1/Th2) cell differentiation
(Boonstra 2001). 1,25(OH)2D3 positively regulated anti-encephalitogenic cytokines and
inhibited autoimmune related cytokines in T cells (Cantorna 1998; Mahon, Wittke et al.
2003). In vitro studies have found that through VDR signaling, 1,25 (OH)2D3 repressed
Th17 and regulatory T cell (Treg) differentiation (Chang, Cha et al. 2010). 1,25(OH)2D3
could also regulate dendritic cell maturation (Piemonti 2000). 1,25(OH)2D3 has also been
shown to positively regulate several murine models of autoimmune diseases such as
multiple sclerosis (MS), inflammatory bowel diseases (IBD) (Cantorna 2006),
inflammatory arthritis (Cantorna 1998) and type I diabetes (Zella 2003).
1.1.3 Vitamin D and NKT cells
Natural Killer T (NKT) cells were originally defined as a subset of T cells that
express NK lineage receptors like NK1.1 (Bendelac 1997). NKT cells are divided into
two different subset, including type I and type II NKT cells (Godfrey, MacDonald et al.
2004). Type I NKT cells are also called invariant NKT cells (iNKT cells). The iNKT
cells have an invariant Vα14-Jα18 T cell receptor (TCR) chain (in mice) and can be
specifically activated by a marine sponge derived-glycolipid -galactosylceramide (-
GalCer) presented by CD1d-expressing antigen-presenting cells (Godfrey, MacDonald et

3
al. 2004; Reilly, Wands et al. 2010). iNKT cells possess effector and immunoregulatory
functions. They have cytotoxic effects and secrete large amounts of cytokines very
rapidly, helping effector cells and also regulating Th1/Th2 cell differentiation (Wilson
and Delovitch 2003). Type II NKT cells (non-classical NKT cells) are also CD1d
dependent, but they express diverse TCR chains and are not reactive to -GalCer
(Godfrey, MacDonald et al. 2004). CD1d KO mice are NKT cell deficient and Jα18 KO
mice are iNKT cell deficient, which were used widely to investigate the functions of
NKT cells. The NKT cells serve as important regulators of the immune responses in
different aspects.
The normal development and functions of mouse iNKT cells require vitamin D
receptor (VDR) (Yu and Cantorna 2008). The number of iNKT cells was significantly
reduced in vitamin D deficient mice compared to vitamin D sufficient mice, and the
reduced cell numbers could not be recovered following 1,25(OH)2D3 supplementation in
either neonatal or adult mice (Yu and Cantorna 2011). Vitamin D and the VDR both
regulated the expansion and proliferation of early iNKT cell precursors (Cantorna, Zhao
et al. 2012).
1.2 Multiple Sclerosis
1.2.1 Background and epidemiology
Multiple sclerosis is a chronic autoimmune demyelinating disease affecting the
central nervous system (CNS). The myelin and axons of the brain and spinal cord are
damaged, which leads to demyelination and scarring (Scheinberg 1987; Compston 2008).

4
MS afflicts about 350,000 people in the United States and about 1.1 million people all the
world (Gross and Jager 2011). The etiology of MS still remains unknown, but evidence
shows that environmental and genetic factors involve roles in MS incidence.
Geographical distribution of MS around the world showed that disease prevalence and
severity was dramatically decreased along with increasing sunlight exposure, which was
explained by the increased vitamin D production through the UV sunlight (Gross and
Jager 2011). Vitamin D is a potential environmental factor affecting MS development.
1.2.2 Vitamin D and multiple sclerosis
Vitamin D has been shown to be involved in the development of autoimmune
diseases. We have already proposed that vitamin D status plays an important role at the
environmental level in regulating the development of autoimmunity (Cantorna 2011).
Previous study demonstrated that deficiency of 25(OH)D3 was found in MS patients
compared to healthy population (Sioka 2009). Low levels of circulating vitamin D were
linked to severe symptoms (increased disability) in MS patients in Tasmania, Australia
(van der Mei 2007). Increase in both sun exposure and vitamin D supplements during
childhood and adolescence were proven to be related with decreased MS incidence in
north of the Arctic Circle, and these factors were also linked to MS onset time (Kampman
2007). Thus, vitamin D is an important factor in regulating MS incidence and severity.
1.2.3 EAE and vitamin D
Experimental autoimmune encephalomyelitis (EAE) is a commonly used murine
model for MS. EAE is a Th1 and Th17 CD4+ T cell mediated autoimmune demyelinating

5
disease, targeting the CNS and inducing inflammation and paralysis to the mice.
Preceding evidence has shown that the deficiency of vitamin D caused earlier onset of
EAE symptoms in mice (Cantorna 1996). Recent studies have found that vitamin D
deficiency reduced EAE severity and delayed EAE onset (Fernandes de Abreu, Ibrahim
et al. 2010; DeLuca and Plum 2011; Fernandes de Abreu, Landel et al. 2011). The
vitamin D hormone 1,25(OH)2D3 could prevent EAE and reversibly block the progression
of the disease (Lemire 1991; Cantorna 1996). 1,25(OH)2D3 increased anti-
encephalitogenic cytokine TGF-β1 and IL-4 with EAE, which correlated with the
inhibition of MS like symptoms in mice (Cantorna 1998; Mahon, Wittke et al. 2003).
Serum TGF-β1 levels were increased in MS patients given a vitamin D supplements
(Mahon 2003). In vivo experiments have shown that 1,25(OH)2D3 failed to protect IL-4
KO mice from developing EAE (Cantorna, Humpal-Winter et al. 2000), suggesting that
in vivo up-regulation of IL-4 is an important factor mediating the 1,25(OH)2D3 effects in
EAE inhibition.
1,25(OH)2D3 treatment decreased production of Th1-associated cytokines IFN-γ, IL-
2 and TNF-α in T cells (Rigby 1987; Mahon, Wittke et al. 2003), suggesting a role for
1,25(OH)2D3 in inhibiting EAE related cytokines. Th17 related-cytokines were also
inhibited by 1,25(OH)2D3 in vivo and in vitro in experimental autoimmune uveitis
experiments (Tang, Zhou et al. 2009). 1,25(OH)2D3 suppressed IL-17A expression in
EAE mouse models through mechanism of transcriptional suppression mediated by VDR
(Joshi, Pantalena et al. 2011).

6
Recent studies using bone marrow chimeric mice with a disrupted VDR and
conditional targeting experiments have shown that VDR was necessary for 1,25(OH)2D3
inhibition of EAE and 1,25(OH)2D3 acted directly through VDR on CD4+ T cells to
inhibit EAE (Mayne, Spanier et al. 2011). Other researchers have demonstrated that the
development of EAE required mediation of vitamin D and VDR, and 1,25(OH)2D3 may
not play a role in the autoimmune response to initiate EAE (Wang, Marling et al. 2012).
Overall, the data showed that 1,25(OH)2D3 prevents EAE onset and reverses the
progression of symptoms partly through inhibiting Th1 and Th17 cells and inducing Th2
cytokine production.
1.2.4 EAE and NKT cells
NKT cells were also shown to play a regulatory role in EAE. SJL mice which are
highly susceptible to EAE have Vα14 NKT cell dysfunction (Taniguchi, Harada et al.
2003). Transgenic mice that have enriched CD1d restricted NKT cells failed to develop
EAE and the encephalitogenic autoantigen-specific IFN- production was inhibited in the
spleen. This suggested that the protective role of CD1d restricted NKT cells against EAE
may work through inhibition of autoreactive Th1 cell cytokine responses (Mars 2002).
Activation of NKT cells with -GalCer at the time of EAE induction in C57BL/6 mice
prevented EAE (Jahng 2001). Prior-activation of NKT cells provided protection to
B10.PL mice against EAE development dependent on IL-4 secretion (Jahng 2001). IL-4
KO C57BL/6 mice exhibited increased susceptibility to EAE, and α-GalCer
administration failed to protect IL-4 KO mice against EAE (Singh 2001; Furlan 2003).
NKT cell activation protected mice from EAE in a CD1d-, IL-4- and IL-10-dependent

7
manner and the protection was associated with suppression of Th1 cytokine IFN-
production and increase of Th2 cytokines IL-4 and IL-10 production (Singh 2001; Furlan
2003). The protection from EAE in NKT-enriched mice was associated with NKT cell
infiltrating the CNS and suppression of encephalitogenic Th1 and Th17 cytokine
production responses in the spleen (Mars 2008). WT iNKT cells inhibit both Th1 and
Th17 responses in suppressing EAE while iNKT cells producing either IL-4 or IL-10 but
not IFN- could only inhibit Th1 responses (Oh and Chung 2011). All these data indicate
that NKT cells play a regulatory role in EAE prevention.
1.3 Conclusion
Vitamin D is an immune regulator and has been shown to be involved in the
development and treatment of autoimmune diseases such as MS. The active form of
vitamin D, 1,25(OH)2D3, has been shown to suppress EAE. The iNKT cells have also
been proven to be an important suppressor of EAE. The development of iNKT cells was
impaired due to vitamin D and or vitamin D receptor deficiency. -GalCer administration
prevents EAE development but not in IL-4 KO mice. 1,25(OH)2D3 is less effective in
preventing EAE symptoms in IL-4 KO mice. The goal of this research was to determine
whether 1,25(OH)2D3 functions to suppress EAE with the production of IL-4 in iNKT
cells.

8
Chapter 2 Materials and Methods
2.1 Mice and diets
C57BL/6 mice with the following genotypes: wild type (WT), CD1d KO (which
lack endogenous NKT cells), Jα18 KO (which lack endogenous iNKT cells) and IL-4 KO
were produced in our colony at Pennsylvania State University (University Park, PA).
Breeding pairs of WT and IL-4 KO mice were purchased from Jackson Laboratories (Bar
Harbor, ME) (Cantorna, Humpal-Winter et al. 2000). CD1d KO and Jα18 KO mice were
kind gifts from Vanderbilt University (Nashville, TN). Mice were fed on purified control
gel diets generated in the laboratory of Dr. Cantorna. 1,25(OH)2D3 supplemented diets
were synthetic gel diets that contained 1,25(OH)2D3 (50ng/day/mouse). Mice with
αGalCer treatment were fed on purified control gel diets. For all EAE experiments, the
diets were fed beginning 1 week prior to the EAE induction and continued throughout the
whole experiment (21 days). Mice were fed with freshly made diets which were replaced
freshly 3 times per week. All described experimental procedures were viewed and
approved by the Office of Research Protections, Institutional Animal Care and Use
Committee at the Pennsylvania State University.
2.2 αGalCer stimulation
αGalCer (Axxora, San Diego, CA) was dissolved in PBS containing 0.5% Tween 20.
(Yu and Cantorna 2008). Mice were given 4.4 μg of αGalCer dissolved in PBS or vehicle
through intraperitoneal (i.p.) injection at the time of EAE induction (day 0) (Jahng 2001).
2.3 EAE induction

9
Six to eight week old sex-matched C57BL/6 mice were immunized subcutaneously
with 100 µl of 200 µg MOG (myelin oligodendrocyte glycoprotein) peptide 35-55
(AnaSpec, Fremont, CA) emulsified in Freund’s adjuvant (IFA; Difco, Detroit, MI)
supplemented with attenuated Mycobacterium tuberculosis H37RA (Difco, Detroit, MI)
to 4mg/ml. Additionally, on the day of immunization and 2 days after (day 0 and 2),
100µl of 200 ng pertussis toxin (List Biological Laboratories, Campbell, CA) dissolved
in PBS was injected i.p. to the mice (Miller and Karpus 2007).
2.4 EAE symptoms
Mice were monitored for signs of EAE until 21 days after immunization. Clinical
EAE scores were recorded daily according to the severity of EAE symptoms using the
following scoring system: 1, limp tail; 2, partial hind limb paralysis; 3, complete hind
limb paralysis; 4, complete hind and some fore limb paralysis; 5, moribund or dead
(Cantorna 1996). Mice with EAE scores of 2 or more were considered EAE positive.
Mice with EAE score of 5 were sacrificed due to humane reasons. Day of onset was
calculated by averaging the day of EAE onset for each mouse that got scored equal or
larger than 2. Peak severity scores were the average of the maximum score for each
mouse. Incidence showed the proportion of number of affected mice out of number of all
mice tested in the experiment. Mortality displayed the proportion of mice that died or
were sacrificed in the experiment. Cumulative disease score (CDI) was calculated by
summing all of the EAE scores for the 21 days of the experiment and dividing by the
number of mice in each group (Cantorna 1996; Becklund, Severson et al. 2010).
2.5 Statistical analysis

10
Experimental data were expressed as mean ± SEM. Statistical analyses were
performed using GraphPad Prism software (GraphPad, La Jolla, CA). Clinical scores and
other measurements were compared by the unpaired Student t test and analysis of
variance (ANOVA), incidence ratio was compared by two sample test for binomial
proportions (Wang, Marling et al. 2012). P-value < 0.05 was considered statistically
significant.

11
Chapter 3 Results
3.1 1,25(OH)2D3 is ineffective at suppressing EAE in CD1d KO mice
WT mice on control diet showed EAE symptoms appeared between 12-13 days after
MOG immunization in, with peak EAE scores from around day 15 till day 21(Fig. 3-1A).
In clear contrast, WT mice on 1,25(OH)2D3 diet were protected from developing EAE,
showing significant lower mean EAE scores than mice on control diet throughout the
course of the experiment (Fig. 3-1A). The cumulative EAE scores for WT mice on
1,25(OH)2D3 diet were significantly lower than that of WT mice on control diet (Fig. 3-
1B). Only one WT mouse on 1,25(OH)2D3 diet developed EAE. The incidence of the
disease was significantly lower (8%) than that of WT mice on control diet (91%). The
mean day of EAE onset was also delayed (day 17) compared to WT controls (Table 3-1).
CD1d KO mice on control diet displayed similar course of EAE development as WT
mice on control diet (Fig. 3-1A). For both WT and CD1d KO mice on control diet, the
measures of EAE severity were similar: the mean day of onset was around day 12-13; the
peak severity EAE score was around 3.2-3.3; the incidence of the disease was above 91%
and the CDI was around 27 (Table 3-1). CD1d KO mice on 1,25(OH)2D3 diet had similar
EAE incidence (81%) to that of CD1d KO mice on control diets (95%), but showed
significantly decreased mean EAE scores from day 11 till day 18 compared to CD1d KO
mice on control diet (Fig. 3-1A). The cumulative EAE scores of CD1d KO mice on
1,25(OH)2D3 diet were also significantly lower compared to CD1d KO mice on control
diet (Fig. 3-1B). CD1d KO mice on 1,25(OH)2D3 diet had delayed mean day of EAE

12
onset and significantly lower peak severity scores compared to control treated CD1d KO
mice (Table 3-1).
The mean EAE scores of CD1d KO mice on 1,25(OH)2D3 diet were significantly
higher than that of WT mice on 1,25(OH)2D3 diet from day 16, day 18 to day 21 (Fig. 3-
1A). Additionally, the cumulative EAE score of CD1d KO mice on 1,25(OH)2D3 diet was
also significantly higher than that of WT mice on 1,25(OH)2D3 diet (Fig. 3-1B). The EAE
incidence of CD1d KO mice on 1,25(OH)2D3 diet (81%) was also statistically higher than
that of WT mice on same diet (8%, Table 3-1).
3.2 1,25(OH)2D3 is ineffective at suppressing EAE in Jα18 KO mice
Jα18 KO mice on control diet displayed similar course of EAE development as WT
mice on control diet. EAE symptoms appeared around 13 days after MOG immunization,
with high EAE scores from around day 15-20 (Fig. 3-2A). For both WT and Jα18 KO
mice on control diet, the measures of EAE severity were similar (Table 3-2).
Jα18 KO mice on 1,25(OH)2D3 diet had similar EAE incidence (81%) to that of Jα18
KO mice on control diets (78%), but showed significantly lower mean EAE scores from
day 14 till day 18 compared to Jα18 KO mice on control diet (Fig. 3-2A). The cumulative
EAE scores of Jα18 KO mice on 1,25(OH)2D3 diet were also significantly lower
compared to Jα18 KO mice on control diet (Fig. 3-2B). But, the mean day of EAE onset
and the peak severity score of Jα18 KO mice on 1,25(OH)2D3 diet was not significantly
different than that of Jα18 KO mice on control diet (Table 3-2). However, when
comparing the EAE symptoms between Jα18 KO and WT mice on 1,25(OH)2D3 diet, the
cumulative EAE score of Jα18 KO mice on 1,25(OH)2D3 diet was significantly higher

13
than that of WT mice on 1,25(OH)2D3 diet (Fig. 3-2B). Additionally, the EAE incidence
of Jα18 KO mice on 1,25(OH)2D3 diet (81%) was also significantly higher than that of
WT mice on same diet (8%, Table 3-2).
CD1d KO mice and Jα18 KO mice on control or 1,25(OH)2D3 diet had similar
course of EAE development (Fig. 3-3A). The cumulative EAE scores of CD1d KO mice
and Jα18 KO mice on control diet did not differ from each other. Also, the cumulative
EAE scores of CD1d KO mice and Jα18 KO mice on 1,25(OH)2D3 diet were similar to
each other (Fig. 3-3B).
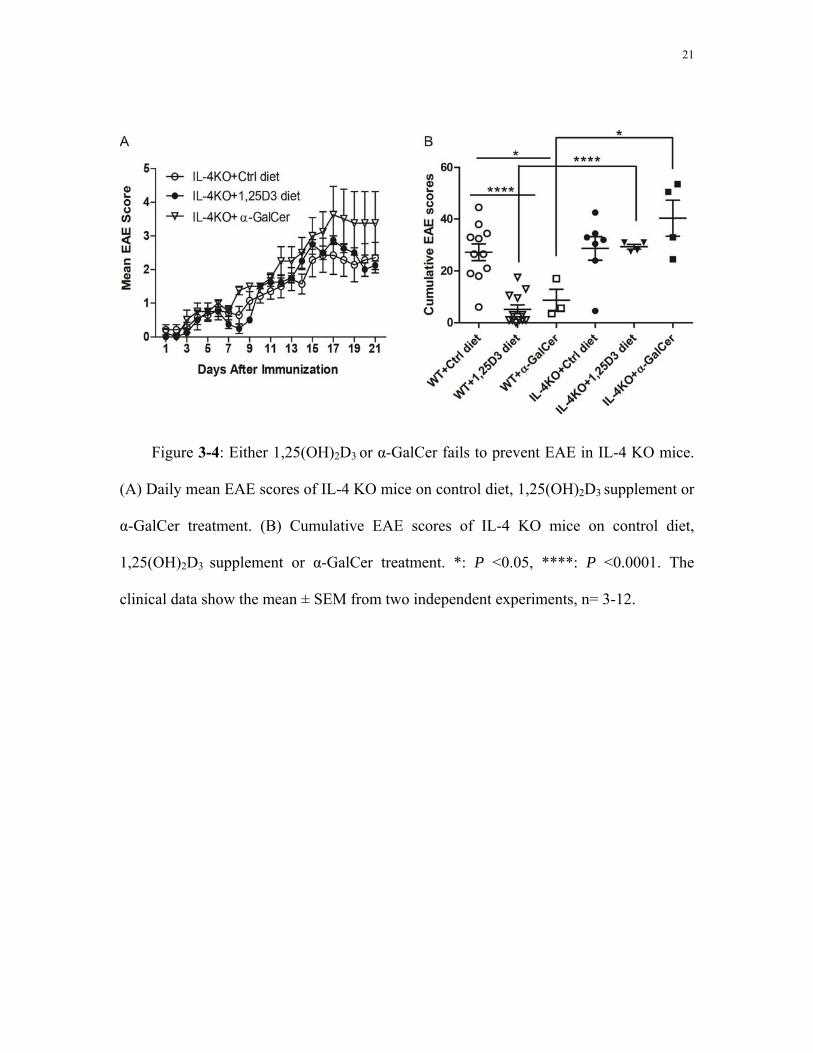
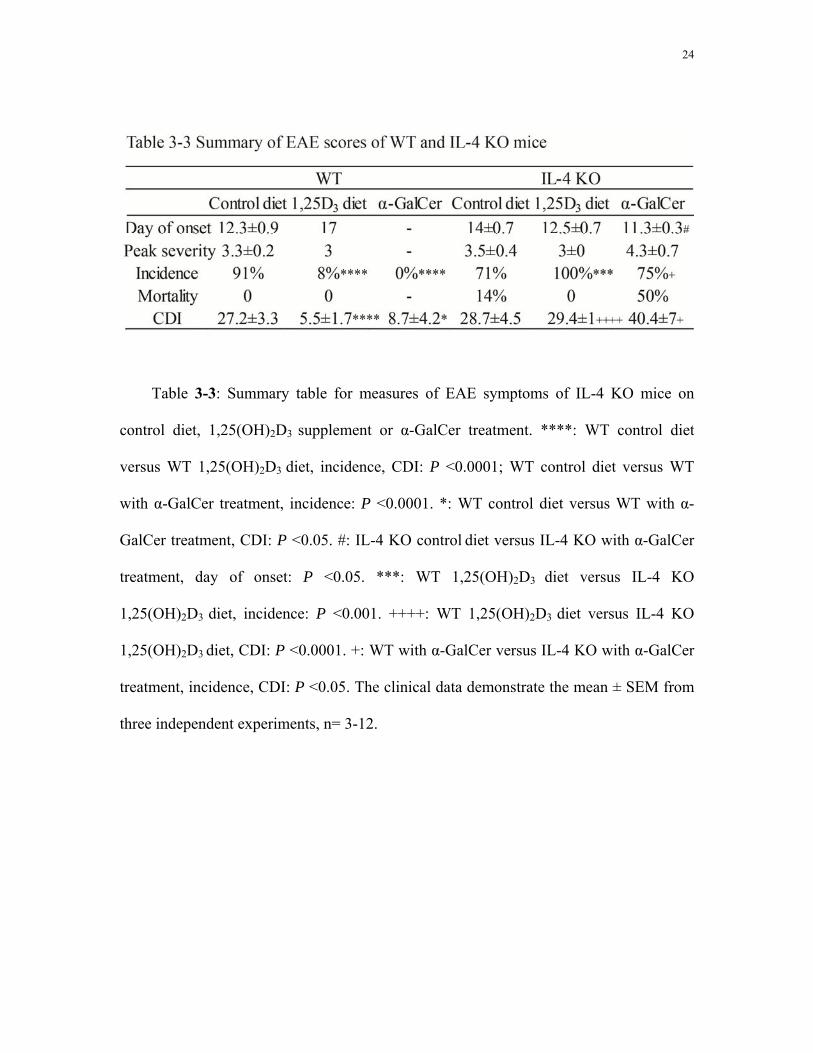
3.3 Either 1,25(OH)2D3 or α-GalCer fails to prevent EAE in IL-4KO mice
IL-4 KO mice on control diet displayed a similar course of EAE development as WT
mice on control diet. EAE symptoms appeared around 14 days after MOG immunization,
with high EAE scores from around day 15-21 (Fig. 3-4A). For both WT and IL-4 KO
mice on control diet, the measures of EAE severity were similar: the mean day of EAE
onset was around day 12-14; the peak severity EAE score was around 3.3-3.5; and the
CDI was around 27-28 (Table 3-3). The mortality rate of IL-4 KO mice on control diet
(14%) was higher than that of WT mice on control diet (0%).
IL-4 KO mice on 1,25(OH)2D3 diet showed similar EAE symptoms as IL-4 KO mice
on control diet. EAE symptoms appeared ~12-14 days after MOG immunization, with
high EAE scores from around day 15 to day 21 (Fig. 3-4A). The cumulative EAE scores
of IL-4 KO mice on 1,25(OH)2D3 diet were not significantly different from that of IL-4
KO mice on control diet (Fig. 3-4B). Peak severity score and incidence were also not
significantly different for IL-4 KO mice on control diet and 1,25(OH)2D3 diet (Table 3-3).

14
Comparing the EAE symptoms of IL-4 KO mice on 1,25(OH)2D3 diet with WT mice on
1,25(OH)2D3 diet, the EAE incidence of IL-4 KO mice on 1,25(OH)2D3 diet (100%) was
significantly higher than that of WT mice on 1,25(OH)2D3 diet (8%, Table 3-3).
IL-4 KO mice on control diet with α-GalCer administration also showed similar
EAE symptoms as IL-4 KO mice on control diet (Fig. 3-4A). The mean day of EAE onset
of IL-4 KO mice on control diet with α-GalCer (day 11) was significantly earlier than
that of IL-4 KO mice on control diet without α-GalCer (day 14, Table 3-3). There were
no significant differences in other measures of EAE symptoms including peak severity,
incidence, mortality and CDI between these two groups (Table 3-3, Fig. 3-4B). On the
contrary, WT mice on control diet with α-GalCer administration were protected from
EAE. The EAE incidence of IL-4 KO mice on control diet with α-GalCer administration
(75%) was significantly higher than that of WT mice on the same treatment (0%, Table 3-
3).

15
Chapter 4 Discussion
Vitamin D status plays an important role in regulating the development of MS. It is
also required for the normal development and function of iNKT cells (Yu and Cantorna
2008). The active form of vitamin D, 1,25(OH)2D3 or iNKT cells were both shown to be
positive regulators of EAE (Cantorna 1996; Jahng 2001; Mars 2002). My data shows that
iNKT cells are required for the full beneficial effects of 1,25(OH)2D3 in EAE, suggesting
a role for iNKT cells in mediating beneficial effects of 1,25(OH)2D3 in mice with EAE.
My results show that the supplementation of 1,25(OH)2D3 in diet reduces the
severity of EAE symptoms but does not decrease the incidence of EAE in CD1d KO
mice. The EAE scores were significantly lower in CD1d KO mice with 1,25(OH)2D3
treatment than in CD1d KO controls. The difference in the EAE clinical measures
between these two treatments is because that the affected mice with 1,25(OH)2D3
treatment didn’t develop as high EAE scores as CD1d KO controls did. However, the
beneficial effects of 1,25(OH)2D3 supplementation on EAE inhibition in CD1d KO mice
were far less than that in WT mice, which suggested that the beneficial effects of
1,25(OH)2D3 in EAE prevention might be mediated by NKT cell regulation. From these
results alone, we are not sure yet whether type I or type II NKT cells or both play the role
to mediate the beneficial effects 1,25(OH)2D3 in EAE prevention.
Further, my data demonstrate that 1,25(OH)2D3 supplementation in diet reduces the
severity of EAE symptoms but does not decrease the incidence of EAE in Jα18 KO mice.
The beneficial effects of 1,25(OH)2D3 in Jα18 KO mice were less than that in WT mice.

16
Besides, 1,25(OH)2D3 has similar effects on the course of EAE development of CD1d KO
mice and Jα18 KO mice and the cumulative EAE scores did not differ from each other.
Since CD1d KO lack both iNKT and type II NKT cells while Jα18 KO mice only lack
iNKT cells, these data suggest that the beneficial effects of 1,25(OH)2D3 in EAE
prevention might be mediated by regulation of iNKT cells but not type II NKT cells.
Previous studies have found that 1,25(OH)2D3 was a positive regulator for the anti-
encephalitogenic Th2 cytokine IL-4 (Cantorna 1998; Mahon, Wittke et al. 2003). It has
been shown that 1,25(OH)2D3 failed to prevent IL-4 KO mice from developing EAE
(Cantorna, Humpal-Winter et al. 2000). α-GalCer administration also failed to protect IL-
4 KO mice on the C57BL/6 background (Falcone 1998; Singh 2001; Furlan 2003). My
results confirm that neither 1,25(OH)2D3 supplementation nor α-GalCer administration
could protect IL-4 KO mice from EAE development. Neither of these treatments had
protective effects against EAE in IL-4 KO mice compared to WT, indicating that in vivo
up-regulation of IL-4 is an important factor in effects of 1,25(OH)2D3 or α-GalCer on
EAE inhibition.
Vitamin D status affects the development of autoimmune diseases like MS. My data
suggest that the preventive effects of 1,25(OH)2D3 in EAE mice might be regulated by
iNKT cells and IL-4 might play an important role in mediating the beneficial effects of
either 1,25(OH)2D3 or α-GalCer on EAE inhibition. The protective effects of
1,25(OH)2D3 against EAE might be regulated by NKT cell derived IL-4. Understanding
the role of NKT cells on the beneficial effects of active vitamin D against EAE might
help us understand more about the protective and pathologic mechanisms of vitamin D

17
involved in human MS development and find new therapeutic targets to serve as
treatment of the disease.

18
Figure 3-1: 1,25(OH)2D3 treatment is ineffective to suppress EAE in CD1d KO mice.
(A) Daily mean EAE scores of WT and CD1d KO mice on control or 1,25(OH)2D3
supplemented diet. *: WT control diet versus WT 1,25(OH)2D3 diet, day 10: P <0.01, day
11,12,14,16-21: P <0.0001, day 13: P <0.05, day 15: P<0.001; #: CD1d KO control diet
versus CD1d KO 1,25(OH)2D3 diet, day 11: P <0.05, day 12,18: P <0.01, day 13:
P<0.001; day14-17: P <0.0001; +: WT 1,25(OH)2D3 diet versus CD1d KO 1,25(OH)2D3
diet, day 16: P <0.05, day 18,19,21: P <0.01, day 20: P<0.001. (B) The cumulative EAE
scores of WT and CD1d KO mice on control or 1,25(OH)2D3 supplemented diet. **: P
<0.01, ****: P <0.0001. The clinical data show the mean ± SEM from three independent
experiments, n= 11-21.

19
Figure 3-2: 1,25(OH)2D3 treatment is ineffective to suppress EAE in Jα18 KO mice.
(A) Daily mean EAE scores of WT and Jα18 KO mice on control or 1,25(OH)2D3
supplemented diet. #: WT control diet versus WT 1,25(OH)2D3 diet, day 10: P <0.01, day
11,12,14,16-21: P <0.0001, day 13: P <0.05, day 15: P<0.001; *: Jα18 KO control diet
versus Jα18 KO 1,25(OH)2D3 diet, day 14: P <0.001, day 15,18: P <0.05, day 16-17:
P<0.01. (B) The cumulative EAE scores of WT and Jα18 KO mice on control or
1,25(OH)2D3 supplemented diet. *: P <0.05, ****: P <0.0001. The clinical data show the
mean ± SEM from three independent experiments, n= 11-22.

20
Figure 3-3: 1,25(OH)2D3 treatment is ineffective to suppress EAE in either CD1d
KO or Jα18 KO mice. (A) Daily mean EAE scores of CD1d KO and Jα18 KO mice on
control or 1,25(OH)2D3 supplemented diet. (B) The cumulative EAE scores of CD1d KO
and Jα18 KO mice on control or 1,25(OH)2D3 supplemented diet. The clinical data show
the mean ± SEM from three independent experiments, n= 18-22.
21
Figure 3-4: Either 1,25(OH)2D3 or α-GalCer fails to prevent EAE in IL-4 KO mice.
(A) Daily mean EAE scores of IL-4 KO mice on control diet, 1,25(OH)2D3 supplement or
α-GalCer treatment. (B) Cumulative EAE scores of IL-4 KO mice on control diet,
1,25(OH)2D3 supplement or α-GalCer treatment. *: P <0.05, ****: P <0.0001. The
clinical data show the mean ± SEM from two independent experiments, n= 3-12.

22
Table 3-1: Summary table for measures of EAE symptoms of WT and CD1d KO
mice on control or 1,25(OH)2D3 diet. ****: WT control diet versus WT 1,25(OH)2D3 diet,
incidence, CDI: P <0.0001; CD1d KO control diet versus CD1d KO 1,25(OH)2D3 diet,
CDI: P <0.0001. **: CD1d KO control diet versus CD1d KO 1,25(OH)2D3 diet, day of
onset: P<0.01.*: CD1d KO control diet versus CD1d KO 1,25(OH)2D3 diet, peak severity:
P <0.05. ++++: WT 1,25(OH)2D3 diet versus CD1d KO 1,25(OH)2D3 diet, incidence: P
<0.0001. ++: WT 1,25(OH)2D3 diet versus CD1d KO 1,25(OH)2D3 diet, CDI: P <0.01.
The clinical data demonstrate the mean ± SEM from three independent experiments, n=
11-21.

23
Table 3-2: Summary table for measures of EAE symptoms of WT and Jα18 KO
mice on control or 1,25(OH)2D3 diet. ****: WT control diet versus WT 1,25(OH)2D3 diet,
incidence, CDI: P <0.0001; **: WT 1,25(OH)2D3 diet versus Jα18 KO 1,25(OH)2D3 diet,
incidence: P <0.01; *: Jα18 KO control diet versus Jα18 KO 1,25(OH)2D3 diet, CDI: P
<0.05; +: WT 1,25(OH)2D3 diet versus Jα18 KO 1,25(OH)2D3 diet, CDI: P <0.05. The
clinical data demonstrate the mean ± SEM from three independent experiments, n= 11-22.
24
Table 3-3: Summary table for measures of EAE symptoms of IL-4 KO mice on
control diet, 1,25(OH)2D3 supplement or α-GalCer treatment. ****: WT control diet
versus WT 1,25(OH)2D3 diet, incidence, CDI: P <0.0001; WT control diet versus WT
with α-GalCer treatment, incidence: P <0.0001. *: WT control diet versus WT with α-
GalCer treatment, CDI: P <0.05. #: IL-4 KO control diet versus IL-4 KO with α-GalCer
treatment, day of onset: P <0.05. ***: WT 1,25(OH)2D3 diet versus IL-4 KO
1,25(OH)2D3 diet, incidence: P <0.001. ++++: WT 1,25(OH)2D3 diet versus IL-4 KO
1,25(OH)2D3 diet, CDI: P <0.0001. +: WT with α-GalCer versus IL-4 KO with α-GalCer
treatment, incidence, CDI: P <0.05. The clinical data demonstrate the mean ± SEM from
three independent experiments, n= 3-12.

25
References
Becklund, B. R., K. S. Severson, et al. (2010). "UV radiation suppresses experimental autoimmune encephalomyelitis independent of vitamin D production." Proc Natl Acad Sci U S A 107(14): 6418-6423.
Bendelac, A. (1997). "Mouse CD1-specific NK1 T cells: development, specificity, and function." Annual review of immunology 15(1): 535-562.
Boonstra, A. (2001). "1alpha,25-Dihydroxyvitamin d3 has a direct effect on naive CD4( ) T cells to enhance the development of Th2 cells." The Journal of immunology (1950) 167(9): 4974-4980.
Cantorna, M. T. (1996). "1,25-Dihydroxyvitamin D3 reversibly blocks the progression of relapsing encephalomyelitis, a model of multiple sclerosis." PNAS : Proceedings of the National Academy of Sciences 93(15): 7861-7864.
Cantorna, M. T. (1998). "1,25-Dihydroxycholecalciferol inhibits the progression of arthritis in murine models of human arthritis." The Journal of nutrition 128(1): 68-72.
Cantorna, M. T. (1998). "1,25-dihydroxyvitamin D3 is a positive regulator for the two anti-encephalitogenic cytokines TGF-beta 1 and IL-4." The Journal of immunology (1950) 160(11): 5314-5319.
Cantorna, M. T. (2006). "Vitamin D and its role in immunology: multiple sclerosis, and inflammatory bowel disease." Progress in biophysics and molecular biology 92(1): 60-64.
Cantorna, M. T. (2011). "Why do T cells express the vitamin D receptor?" Ann N Y Acad Sci 1217: 77-82.
Cantorna, M. T., J. Humpal-Winter, et al. (2000). "In vivo upregulation of interleukin-4 is one mechanism underlying the immunoregulatory effects of 1,25-dihydroxyvitamin D(3)." Arch Biochem Biophys 377(1): 135-138.
Cantorna, M. T., J. Zhao, et al. (2012). "Vitamin D, invariant natural killer T-cells and experimental autoimmune disease." Proc Nutr Soc 71(1): 62-66.
Chang, J. H., H. R. Cha, et al. (2010). "1,25-Dihydroxyvitamin D3 inhibits the differentiation and migration of T(H)17 cells to protect against experimental autoimmune encephalomyelitis." PLoS One 5(9): e12925.
Compston, A. (2008). "Multiple sclerosis." The Lancet (British edition) 372(9648): 1502-1517.
DeLuca, H. F. (2004). "Overview of general physiologic features and functions of vitamin D." Am J Clin Nutr 80(6 Suppl): 1689S-1696S.
DeLuca, H. F. and L. A. Plum (2011). "Vitamin D deficiency diminishes the severity and delays onset of experimental autoimmune encephalomyelitis." Arch Biochem Biophys 513(2): 140-143.
Falcone, M. (1998). "A critical role for IL-4 in regulating disease severity in experimental allergic encephalomyelitis as demonstrated in IL-4-deficient C57BL/6 mice and BALB/c mice." The Journal of immunology (1950) 160(10): 4822-4830.

26
Feldman, D. (1999). "Vitamin D, parathyroid hormone, and calcium: a complex regulatory network." The American journal of medicine 107(6): 637-639.
Fernandes de Abreu, D. A., E. C. Ibrahim, et al. (2010). "Severity of experimental autoimmune encephalomyelitis is unexpectedly reduced in mice born to vitamin D-deficient mothers." J Steroid Biochem Mol Biol 121(1-2): 250-253.
Fernandes de Abreu, D. A., V. Landel, et al. (2011). "Seasonal, gestational and postnatal influences on multiple sclerosis: the beneficial role of a vitamin D supplementation during early life." J Neurol Sci 311(1-2): 64-68.
Furlan, R. (2003). "Activation of invariant NKT cells by alphaGalCer administration protects mice from MOG35-55-induced EAE: critical roles for administration route and IFN-gamma." Eur J Immunol 33(7): 1830-1838.
Godfrey, D. I., H. R. MacDonald, et al. (2004). "NKT cells: what's in a name?" Nat Rev Immunol 4(3): 231-237.
Gross, R. H. and P. L. Jager (2011). "MS: Epidemiology and Genetics." 71-87.
Helming, L. (2005). "1alpha,25-Dihydroxyvitamin D3 is a potent suppressor of interferon gamma-mediated macrophage activation." Blood 106(13): 4351-4358.
Jahng, A. W. (2001). "Activation of natural killer T cells potentiates or prevents experimental autoimmune encephalomyelitis." The Journal of experimental medicine 194(12): 1789-1799.
Jones, G. (1998). "Current understanding of the molecular actions of vitamin D." Physiological reviews 78(4): 1193-1231.
Kampman, M. T. (2007). "Outdoor activities and diet in childhood and adolescence relate to MS risk above the Arctic Circle." Journal of neurology 254(4): 471-477.
Lemire, J. M. (1991). "1,25-dihydroxyvitamin D3 prevents the in vivo induction of murine experimental autoimmune encephalomyelitis." The Journal of clinical investigation 87(3): 1103-1107.
Mahon, B. D. (2003). "Cytokine profile in patients with multiple sclerosis following vitamin D supplementation." Journal of neuroimmunology 134(1-2): 128-132.
Mahon, B. D., A. Wittke, et al. (2003). "The targets of vitamin D depend on the differentiation and activation status of CD4 positive T cells." J Cell Biochem 89(5): 922-932.
Mars, L. T. (2002). "Cutting edge: V alpha 14-J alpha 281 NKT cells naturally regulate experimental autoimmune encephalomyelitis in nonobese diabetic mice." The Journal of immunology (1950) 168(12): 6007-6011.
Masuyama, H. (1997). "Evidence for ligand-dependent intramolecular folding of the AF-2 domain in vitamin D receptor-activated transcription and coactivator interaction." Molecular endocrinology (Baltimore, Md.) 11(10): 1507-1517.

27
Mayne, C. G., J. A. Spanier, et al. (2011). "1,25-Dihydroxyvitamin D3 acts directly on the T lymphocyte vitamin D receptor to inhibit experimental autoimmune encephalomyelitis." Eur J Immunol 41(3): 822-832.
Miller, S. D. and W. J. Karpus (2007). "Experimental autoimmune encephalomyelitis in the mouse." Curr Protoc Immunol Chapter 15: Unit 15 11.
Piemonti, L. (2000). "Vitamin D3 affects differentiation, maturation, and function of human monocyte-derived dendritic cells." The Journal of immunology (1950) 164(9): 4443-4451.
Provvedini, D. M. (1983). "1,25-dihydroxyvitamin D3 receptors in human leukocytes." Science (New York, N.Y.) 221(4616): 1181-1183.
Reilly, E. C., J. R. Wands, et al. (2010). "Cytokine dependent and independent iNKT cell activation." Cytokine 51(3): 227-231.
Rigby, W. F. (1987). "Regulation of lymphokine production and human T lymphocyte activation by 1,25-dihydroxyvitamin D3. Specific inhibition at the level of messenger RNA." The Journal of clinical investigation 79(6): 1659-1664.
Scheinberg, L. (1987). "Rehabilitation of patients with multiple sclerosis." Neurologic clinics 5(4): 585-600.
Singh, A. K. (2001). "Natural killer T cell activation protects mice against experimental autoimmune encephalomyelitis." The Journal of experimental medicine 194(12): 1801-1811.
Sioka, C. (2009). "Multiple sclerosis, osteoporosis, and vitamin D." Journal of the neurological sciences 287(1-2): 1-6.
Tang, J., R. Zhou, et al. (2009). "Calcitriol suppresses antiretinal autoimmunity through inhibitory effects on the Th17 effector response." J Immunol 182(8): 4624-4632.
Taniguchi, M., M. Harada, et al. (2003). "The regulatory role of Valpha14 NKT cells in innate and acquired immune response." Annu Rev Immunol 21: 483-513.
van der Mei, I. A. F. (2007). "Vitamin D levels in people with multiple sclerosis and community controls in Tasmania, Australia." Journal of neurology 254(5): 581-590.
Veldman, C. M. (2000). "Expression of 1,25-dihydroxyvitamin D(3) receptor in the immune system." Archives of biochemistry and biophysics 374(2): 334-338.
Wang, Y., S. J. Marling, et al. (2012). "Development of experimental autoimmune encephalomyelitis (EAE) in mice requires vitamin D and the vitamin D receptor." Proc Natl Acad Sci U S A 109(22): 8501-8504.
Wang, Y., S. J. Marling, et al. (2012). "Development of experimental autoimmune encephalomyelitis (EAE) in mice requires vitamin D and the vitamin D receptor." Proc Natl Acad Sci U S A.
Wilson, S. B. and T. L. Delovitch (2003). "Janus-like role of regulatory iNKT cells in autoimmune disease and tumour immunity." Nat Rev Immunol 3(3): 211-222.
Yu, S. and M. T. Cantorna (2008). "The vitamin D receptor is required for iNKT cell development." Proc Natl Acad Sci U S A 105(13): 5207-5212.

28
Yu, S. and M. T. Cantorna (2011). "Epigenetic reduction in invariant NKT cells following in utero vitamin D deficiency in mice." J Immunol 186(3): 1384-1390.
Zella, J. B. (2003). "Vitamin D and autoimmune diabetes." J Cell Biochem 88(2): 216-222.