research and development - defra, uk - science...
TRANSCRIPT

DEPARTMENT for ENVIRONMENT, FOOD and RURAL AFFAIRS CSG 15Research and Development
Final Project Report(Not to be used for LINK projects)
Two hard copies of this form should be returned to:Research Policy and International Division, Final Reports UnitDEFRA, Area 301Cromwell House, Dean Stanley Street, London, SW1P 3JH.
An electronic version should be e-mailed to [email protected]
Project title Conserved protein/energy-rich forages in sustainable livestock production: rumen function and animal response
DEFRA project code LS0302
Contractor organisation and location
Institute of Grassland and Environmental Research, Plas Gogerddan, Aberystwyth, Ceredigion, SY23 3EB
Total DEFRA project costs £ 751,044
Project start date 01/07/97 Project end date 30/06/02
Executive summary (maximum 2 sides A4)
This project addresses DEFRA policy objectives related to exploitation of natural, home-grown (alternative) forages to replace concentrate supplements, in the context of creating sustainable agricultural systems for the ruminant sector. The final report is based on a 5-year programme and consists of data from ruminant production and digestion studies and laboratory-based experiments to examine the efficiency of utilisation of nutrients from high protein and/or energy, alternative forage crops. The Milk Development Council were joint funders for the first 3 years of the programme and a full report was submitted to them in August 1997. The main objectives of the first 3 years of LS0302 were: (1) to determine milk yield and milk quality responses associated with the feeding of ensiled alternative forages as complete diets for dairy cows; and (2) evaluate the effect of ensiled alternative forages on rumen function and protein utilisation by dairy and beef cattle. The programme was extended (2000-2002) to look at strategies for exploiting ensiled high sugar grasses to manipulate energy supply and improve N use in cattle fed diets based mainly on legume silages. New approaches were also developed during the latter part of the programme to explore plant- and microbially-mediated mechanisms of nutrient supply and their influence on microbial colonisation of feed boli in grazing cattle. Project LS0302 is one of three projects on Alternative Forage Crops which which also includes LS03 01and 03.
1. The nutritive quality and efficiency of protein utilisation of ensiled, protein-rich forages for dairy cattleDairy cow experiments were conducted to examine the nutritive value of legume silages in comparison with grass silage or legume/grass mixtures; the effect of level of concentrate feeding was also examined. Silages were prepared from pure stands of ryegrass, lucerne, white and red clover as bales using a silage inoculant. All silages were fed alone and grass was also fed as a mixture with red and white clover silages (50:50; DM basis). A standard concentrate was included at a flat rate of (8 kg/day in Experiment 1 and 4, or 8 kg/day in Experiment 2). As the level of inclusion of legume silage in the diet increased, silage dry matter intake and milk yield (range 24.9 kg/day for grass silage vs from 28.1and 31.5 for red and white clover, respectively) and milk solids increased, whilst efficiency of conversion of feed-N into milk-N declined (18-20% for pure legumes vs 25.4% for grass). Milk production was high and increased, despite the lower digestibility of red clover and lucerne silages in Experiment 1. Milk output per unit of silage intake was reduced by one third when concentrate levels were reduced from 8 to 4 kg/head/day although, when expressed per unit of total ration, CSG 15 (Rev. 6/02) 1

Projecttitle
Conserved protein/energy-rich forages in sustainable livestock production: rumen function and animal response
DEFRAproject code
LS0302
values were similar, irrespective of concentrate level. Clover silages led to increased levels of PUFA in milk, but the increased α-linolenic acid content of milk from cows fed red clover silage was greater than would be expected from intake effects, suggesting that some other mechanism was operating, probably by reducing biohydrogenation. The use of silage inoculant and baling technology allows the reliable production of well fermented legume silages that support high levels of production in dairy cattle.
2. In vitro and in vivo rumen function experiments involving beef and dairy cattle to examine the efficiency of protein and energy utilisation
A. In vitroA rumen simulation technique (Rusitec) was used to compare digestion and microbial N-flow in response to a range of alternative forage silages (red clover prepared with and without a silage inoculant, and untreated white clover, lucerne, lotus, wholecrop barley/pea, kale) in comparison with grass silage. Microbial protein production and non ammonia-N flows from Rusitec cultures were highest with red clover and kale; grass and barley/pea silages giving the poorest values, with intermediate values for lucerne and lotus. Microbial growth efficiency was highest with red clover silage, particularly after inoculation. Differences in fermentation patterns observed between the silages suggest that differences in energy availability may provide a partial explanation for the range of values obtained. The relative effects of the rate of release of the energy and N fractions of lucerne and red clover silages on microbial growth and efficiency were further explored. The rate of DM loss was similar in both silages, but ammonia-N release was faster with lucerne than red clover, suggesting that more rapid and extensive protein degradation occurred. This was coupled with a higher rate of VFA production and thus energy supply with red clover silage. These factors combined to give a 15% higher value for efficiency of microbial protein synthesis in the early stages of fermentation with red clover and suggest that, compared to lucerne, red clover provided a more balanced supply of nitrogen and energy and a more efficient use of N by the rumen microbial population.
B. In vivo The mechanisms driving the observed differences in feed intake, N utilisation and milk yield and composition in dairy cows fed legume silages, were examined. Silages were prepared from pure stands of ryegrass, lucerne, white and red clover as bales using a silage inoculant. All silages were fed alone and grass silage was also fed as a mixture with red and white clover silages (50 :50; DM basis). A standard concentrate was included at a flat rate of 8 kg/day. High voluntary intakes of lucerne and white clover silage were obtained despite low rumen volumes, and were accompanied by higher passage rates from the rumen. Grass silage had the lowest intakes and highest rumen fills. The high intake characteristics of different legume silages appear to result from different mechanisms; high rates of particle breakdown and passage with lucerne, and high rates of fermentation and passage with white clover, with red clover behaving similarly to grass. There was some evidence of reduced N degradation with red clover, possibly due to the action of plant polyphenol oxidase activity and resultant binding of polyphenols to protein during ensilage; with white clover, reduced N degradation was probably as a consequence of high rates of passage from the rumen. High passage rates would also increase rumen microbial growth efficiency, although there were no significant differences between silages in this experiment. In an experiment with beef steers, diets based on clover silages again led to significantly higher DM intakes than for grass silage. Microbial nitrogen flow at the duodenum was significantly higher when feeding clover silage, particularly that made from white clover. A large part of this effect was due to the DM intake effect as with dairy cows, and again the efficiency of rumen microbial protein synthesis was not different between grass and red or white clover, or the mixtures.
3. In vitro rumen function studies with red clover silage, with and without high sugar or control grass silages The effect of additive treatment on ensilage of high sugar or a control grass was examined in a laboratory experiment. Each grass was ensiled in 1 kg glass jars, alone or with red clover, with or without acid or inoculant treatment. The residual water soluble carbohydrate content (WSC) of high sugar grass silage and mixtures of high sugar grass and red clover silages were significantly higher than for the control grass or red clover silages. Silage quality was also significantly improved by additive treatment, irrespective of the grass used, with formic acid treatment preserving more sugar and inoculant treatment preserving more protein. These results demonstrate the benefits of using additives on silage quality even under good ensiling conditions. They also highlight the potential of high sugar grass for improving the quality of red clover silage. The in vitro rumen simulation techniques (Rusitec) was used to examine how rumen function is influenced by feeding silages prepared from high sugar or control grass varieties, with or without red clover. Microbial N flows were significantly greater for high sugar grass silage (86 g WSC/kg DM) than for control grass silage (30 g WSC/kg DM), and the efficiency of conversion of feed N into microbial N was considerably higher for high sugar grass silage. A graded response was found for both microbial N flows and efficiency values where red clover was included in the diet, with the highest values being seen when the ratio of red clover : high sugar grass silage was in the ratio 70 : 30 on a DM basis. This approach provides a forage-based strategy for balancing the N and energy supply for rumen microbial growth in ruminants fed both grass and legume silages, and offers an option for reducing N pollution to the environment.
4. In vivo rumen function and digestion studies with cattle fed red clover silage, with or without high sugar or control grass silagesThe benefit of conserving a high level of residual sugar in silages prepared from high sugar grasses on efficiency of N and energy utilisation in beef cattle fed grass silages or mixtures of grass and red clover silage was examined. Silages were prepared as bales from pure stands of high sugar and control ryegrass varieties and red clover, using a silage inoculant. All silages were well preserved and there was a 3.5 percentage unit difference in WSC content between the high sugar grass and control grass silages (90.5 v. 55.3 g
CSG 15 (Rev. 6/02) 2

Projecttitle
Conserved protein/energy-rich forages in sustainable livestock production: rumen function and animal response
DEFRAproject code
LS0302
WSC/kg DM respectively). All silages (grasses and red clover) were fed alone and the two grass silages were also fed as mixtures with red clover silage (50:50; DM basis). Silage intake was highest with pure red clover alone or when it was mixed with grass silage, but intakes were similar with these diets to that for high sugar grass silage alone. Feeding high sugar grass silage led to greater flows of microbial nitrogen to the duodenum and values for the efficiency of rumen microbial protein synthesis (in terms of organic matter digested) than for the control grass silage and was also higher for mixtures of control grass and high sugar grass silage than for the red clover silage alone. The efficiency of N use for microbial growth in the rumen increased from 48% with the control silage to 65% with the high sugar grass silage, and small increases were also observed with the grass/clover mixtures in comparison with red clover. These results suggest that if the difference in level of residual sugar in grass silage is above a threshold value of approximately 4 percentage units a substantial reduction in N losses from animals fed diets based on grass and/or red clover can be achieved. Both observations have implications for reducing environmental pollution and the sustainability of livestock agriculture.
5. Bolus biology and microbial ecology studiesA technique was developed to obtain freshly ingested herbage boli from the rumens of dairy cows prior to contact with rumen digesta. Down boli contained large (> 107 cfu/g FM) populations of rumen bacteria, presumably via oesophageal inoculation, but viable fungal populations were not detected using conventional culture techniques. Odd chain fatty acid profiling and molecular techniques were developed and used to study population diversity in the bolus material before and after incubation in the rumen for up to 72 h. Two odd-chain fatty acids (iso C15:0 and iso C17:0) were not detected in grass or white clover from pasture, but were found in washed bacterial preparations obtained from the rumen, indicating that they could be used as microbial markers. The concentration of the odd-chain fatty acid C15:0 accumulated linearly in relation to DM disappearance with both grass and white clover, which suggests rapid and continued microbial colonisation had occurred. Initial results also gave an indication that colonisation occurred sequentially, with different genera appearing at different time points, and there appeared to be interactions with individual animal and plant types. [NB: Subsequent work leads us to question some of the data relating to bacterial, but not fungal, colonisation as the bacterial primers also showed a positive alignment to chloroplastic DNA.] Molecular analysis of anaerobic gut fungal populations was also made on bolus material recovered after incubation in the rumens of animals fed on the same herbages. The four major anaerobic gut fungal genera (Anaeromyces, Neocallimastix, Piromyces and Orpinomyces) were found associated with both grass and white clover boli. Variations in predominant isolates appeared to be host-specific rather than diet-specific. Analysis of bolus samples revealed very early colonisation by anaerobic fungi, with detection at low levels on plant material that had been chewed, swallowed and recovered before contact with rumen digesta. For grass boli, the subsequent growth of anaerobic fungi was immediate and colonised isolates were well established after 1 h of incubation and increased up to 24 h. For white clover boli, colonisation was as rapid as for grass but subsequent growth was slower, displaying a lag (~8 h), before more extensive growth proceeded. Anaerobic gut fungal populations were maximal between 24 and 72 h on white clover boli.
SDS-PAGE techniques were used to fractionate grass and white clover polypeptides and study the rate of ruminal breakdown of proteins in grass and white clover boli. Differences were found between both the rates of disappearance of different proteins and also between plant types. For example, Rubisco disappeared within 2-4 h of incubation, but light harvesting protein was relatively stable for in excess of 24 h, and whilst the breakdown products of grass proteins accumulated after 4 h, those from white clover did not. These results highlight important differences in the digestion of proteins with the same physiological function but from different herbage species. A simple exclusion system was devised to demonstrate if proteases of plant origin were active in the degradation of plant proteins in the rumen environment under in vivo conditions. The three treatments described allowed complete access of micro-organisms to plant material (grass), exclusion of rumen micro-organisms or allowed interaction of fermentation products with plant material while preventing ingress of rumen fluid proteases during in sacco incubation of grass in the rumen. Significant dry matter loss and substantial changes in polypeptide profile of the grass residues was observed during incubation and indicated significant protein breakdown in all treatments. The similar profile in the Dacron bag treatment compared with the dialysis tubing treatment suggests that plant proteases were active in the early stages of digestion and provides in vivo data in support of our previous findings made under in vitro conditions.
An in vitro examination of how 'processing' of herbage during bolus formation influences nutrient release and rate of digestion of fresh herbage demonstrated how rapidly substrate (water soluble carbohydrate) becomes available from the bolus compared to chopped grass and freeze-dried, ground grass; the latter is conventionally used in studies of forage digestion. These findings supported the concept of using the feed bolus to study microbial colonisation, as tissue damage that occurs during ingestion clearly influenced nutrient availability in the early stages of digestion.
In conclusion, the approaches described above, will form the basis of a 'toolkit' for studies of the early stages of digestion of fresh forages, leading to a more precise understanding of the initial events during colonisation and degradation of fresh forage, in particular feed boli. Further development is required but the techniques show considerable promise for examining the response of the rumen microbial population to changes in herbage type, particularly species bred for characteristics that may enhance nutrient use efficiency in the rumen or to improve food quality and/or safety.
Technology TransferThe research findings of this project have been fully communicated to the DEFRA Livestock Knowledge Transfer and MDC Technology Transfer programmes and details of these and publications associated with the work are in the Appendix.
CSG 15 (Rev. 6/02) 3

Projecttitle
Conserved protein/energy-rich forages in sustainable livestock production: rumen function and animal response
DEFRAproject code
LS0302
CSG 15 (Rev. 6/02) 4

Projecttitle
Conserved protein/energy-rich forages in sustainable livestock production: rumen function and animal response
DEFRAproject code
LS0302
Scientific report (maximum 20 sides A4)
CONSERVED PROTEIN/ENERGY-RICH FORAGES IN SUSTAINABLE LIVESTOCK PRODUCTION: RUMEN FUNCTION AND ANIMAL RESPONSE
There is limited information available on the effects on production response, rumen function and N utilisation when ensiled alternative, protein/energy-rich forages are offered as complete diets to ruminants, either alone or in combination with grass silage. The research reported here was carried out to advance scientific knowledge and technical knowhow on the effects of feeding ensiled alternative, protein/energy-rich forages to cattle, with the aim of demonstrating their potential contribution to sustainable livestock systems. The following experiments were carried out to complete the objectives of years 1-3 of the programme: (1) determine milk yield and milk quality responses associated with the use of ensiled, protein-rich alternative forages fed alone or with ensiled grass as complete diets for dairy cows, and (2) evaluate the effect of ensiled alternative forages on rumen function and protein utilisation by dairy and beef cattle using both in vitro and in vivo approaches. The programme in years 4-5 (2001-2002) introduced new concepts for increasing the efficiency of N use in the rumen in animals fed alternative forage silages and, in addition, examined the effect of forage type on nutrient supply and microbial colonisation of ingested feed in relation to rumen function in cattle grazing fresh forage.
1. THE NUTRITIVE QUALITY AND EFFICIENCY OF PROTEIN UTILISATION OF ENSILED, PROTEIN-RICH FORAGES IN DAIRY CATTLE
1.1 Intake and milk production responses to legume silages offered to Holstein-Friesian cows - Experiment 11.1.1 Introduction Work by Thomas et al. (1985) showed that silage made from red clover had considerable potential for milk production, although low digestibility and ensiling difficulties were problems that needed to be addressed. Advances in legume breeding and conservation technology, as well as a renewed emphasis on extensive production systems, meant that it was timely to re-evaluate the potential of legume silages for milk production.
1.1.2 Materials and methods Pure stands of red clover (cv. Milvus), white clover (cv. Aran) and lucerne (cv.Vertus), as well as an area of ryegrass (mixture of cvs AberElan, AberComo and Augusta), were established in late summer 1997. Each crop was harvested over 3 cuts during 1998. Crops were mowed using a disc mower fitted with rubber rollers, left in the swath until shortly before baling and wilted aiming for a dry matter content of 30-35% (maximum 48 hours). Crops were baled using a round baler with a biological additive (Ecosyl; Ecosyl Products Ltd., Billingham, UK) applied according to the manufacturer's recommendations at 1.5 litres per tonne of crop. Mixtures of the 3 cuts, in proportion to yields, were used in feeding experiments in order to make the results more representative of the season’s production. Eighteen Holstein-Friesian dairy cows in early- to mid-lactation were used in a 3-period incomplete changeover design experiment involving 6 treatments: grass silage (G), red clover silage (RC), white clover silage (WC), lucerne silage (L), and 50/50 (DM basis) mixture of G and RC (GRC) and G and WC (GWC). Six of the cows had rumen and duodenal cannulae and were used for the 4-period experiment described below (see Section 2.4). Cows were given a flat-rate of 8 kg/day of a standard concentrate (starch: 229 g/kg DM; neutral detergent fibre (NDF): 247 g/kg DM; crude protein (CP): 220 g/kg DM) and had ad libitum access to the forages either through roughage intake control feeders (Insentec B.V., The Netherlands) or in individual stalls. Feed intake, milk yield and milk composition were recorded throughout and values from the final week of each 4-week period were used in the statistical analysis. DM digestibilities were measured using 6-day total collections of faeces from the fistulated cows in the third week of each period. Results were analysed using REML (Genstat 5; Lawes Agricultural Trust, 1987) with a fixed model of ‘diet’ and a random model of ‘period + cow’.
1.1.3 Results and discussion All silages were well preserved, aerobically stable and acceptable to the animals, despite the wide range of DM contents achieved at baling. Chemical analysis of G, RC, WC and L silages gave the following values: for freeze-DM: 345, 376, 258 and 359 g/kg; for CP: 146, 205, 278 and 244 g/kg DM; for NDF: 548, 439, 287 and 458 g/kg DM; for lactic acid: 66, 74, 99 and 60 g/kg DM; and for pH: 4.45, 4.24, 3.85 and 4.68 respectively. The effects of treatments on intake and production are shown in Table 1.1.1.
CSG 15 (Rev. 6/02) 5

Table 1.1.1 Effects of legume silages on feed intake, diet digestibility and milk production G GRC RC GWC WC L s.e.d. Sig.
DM intake (kg/day) 18.4 19.3 20.4 19.7 19.8 20.6 0.66 **Diet digestibility (g/g) 0.720 0.687 0.649 0.719 0.687 0.640 0.0094 ***Silage DM intake (kg/day) 11.5 12.4 13.5 12.8 12.9 13.7 0.65 **Diet digestibility (%) 72.0 68.7 64.9 71.9 68.7 64.0 0.94 ***Milk yield (kg/day) 24.9 28.6 28.1 27.9 31.5 27.7 1.81 *Milk fat (g/kg) 44.5 46.0 45.2 46.6 43.9 44.2 2.49 NSMilk protein (g/kg) 32.6 32.1 31.4 32.2 32.0 32.6 0.54 NSMilk lactose (g/kg) 47.1 47.2 46.8 47.4 47.1 46.6 0.41 NSN efficiency (milk N/Feed N) 0.254 0.249 0.209 0.225 0.204 0.181 0.0146 ***NS Not significant ( P > 0.1) *, P< 0.05; ** P < 0.01; *** P < 0.001
The results support earlier work, with higher intakes and higher milk production for the legumes, particularly white clover, and legume:grass silage mixtures, despite their lower digestibility values (Thomas et al., 1985). Milk composition was not significantly different across all diets. However, in this experiment where concentrates were fed at a flat rate across treatments, the efficiency of N incorporation into milk with the legume silages was generally low (associated with higher total intake of N) in comparison with grass silage alone. This may in part be related to a shortfall in readily available energy in the diet to support rumen microbial growth. The use of biological inoculants and preparation of legume silages in big bales with minimal handling and chopping allowed the reliable production of well-fermented and stable legume silages and confirmed their ability to support high levels of milk production in dairy cattle.
1.2 Evaluation of legume silages offered to Holstein-Friesian cows with small amounts of concentrates - Experiment 2 1.2.1 Introduction The high intake characteristics of red clover silage have been recognised for many years (e.g. Thomas et al., 1985). The previous study (see 1.1) demonstrated the considerable intake and milk production potential of clover silages prepared using a new approach based on preparation of big-bales from wilted material with the use of a silage inoculant. The objective of this study was to verify the positive results obtained with red and white clover silages using herbage taken from a further series of cuts made in a subsequent year and to evaluate the silages with two levels (8 and 4 kg/day) of concentrate feeding.
1.2.2 Materials and methods Stands of pure red clover (cv. Milvus), white clover (cv. Aran) and ryegrass (mixture of cvs AberElan, AberComo and Augusta) were harvested in 1999. Crops were mowed using a disc mower fitted with rubber rollers, left in the swath until shortly before baling and wilted aiming for a DM content of 30% (maximum 48 hours). Crops were baled using a round baler with a biological additive (Ecosyl; Ecosyl Products, Billingham) applied according to the manufacturers recommendations at 1.5 litres per tonne of crop. Mixtures of the three/four cuts, in proportion to production, were used in feeding experiments in order to make the results representative of the season’s production. Twenty-one Holstein-Friesian dairy cows in early- to mid-lactation were used in a 3-period incomplete changeover design experiment with 7 treatments, based on 4 forages (grass silage (G), red clover silage (RC), white clover silage (WC) and a 50/50 mixture (DM basis) of grass silage and red clover silage (GRC)), with 2 levels of concentrates (4 or 8 kg/day). The standard concentrate had the following analysis: starch: 231 g/kg DM; neutral detergent fibre (NDF): 246 g/kg DM; crude protein (CP): 250 g/kg DM). The cows had ad libitum access to the forages through roughage intake control feeders. Feed intake, milk yield and milk composition were recorded continuously and values from the final week of each 4-week period were used for the statistical analysis. Results were analysed using REML (Genstat 5; Lawes Agricultural Trust, 1987) with a fixed model of ‘diet’ and a random model of ‘period + cow’.
1.2.3 Results and discussion Chemical analysis of G, RC and WC silages gave the following values: for oven-DM: 284, 221 and 284 g/kg; for CP: 151, 190 and 230 g/kg DM; for NDF: 571, 428 and 323 g/kg DM; and for pH: 4.22, 4.38 and 3.98 respectively. The effects of treatments on intake and production are in Table 1.2.1.

Table 1.2.1 Effects of legume silages on feed intake and milk production Treatment: s.e.d. Sig
G4 G8 GRC4 GRC8 RC4 RC8 WC8Silage DM intake (kg/d) 14.0 12.6 14.5 13.5 16.6 15.2 15.9 0.61 ***Total DM intake (kg/d) 17.4 19.3 17.9 20.3 20.0 22.0 22.6 0.62 NSDiet digestibility (%) 65.5 68.2 69.0 69.0 66.5 66.9 67.6 67.8 *Milk (kg/day) 23.5 27.5 23.7 28.6 25.6 30.2 33.2 0.83 ***Milk fat (g/kg) 37.3 41.0 36.7 37.9 39.1 37.4 35.2 1.86 NSMilk protein (g/kg) 29.8 30.4 29.8 31.1 29.4 29.7 31.7 0.49 ***Milk lactose (g/kg) 46.2 45.9 45.6 46.0 46.2 46.1 46.0 0.49 ***N efficiency + 24.3 23.6 24.0 23.4 18.8 19.7 20.4 0.80 ***+ Milk N/feed N; NS Not significant ( P > 0.1) *, P< 0.05; *** P < 0.001
Considering the treatments based on grass and red clover silages, there were no significant interactions between forage and concentrate level in their effects on both forage intake and milk yield. Feed intake and milk production responses were very similar to those observed in the previous year (See Section 1), despite an increased cutting frequency for grass and red clover. Silage intake increased at the lower level of concentrate supplementation and was associated with a fall in milk output across all silage types. Milk output (g/g silage intake) for the pure grass and red clover and corresponding mixtures ranged from 1.6-1.8 at 4 kg concentrates/day to 2.1-3.2 at 8 kg/day, but when expressed in terms of total feed intake (silage plus concentrate), values were very similar. The efficiency of N incorporation into milk was again poor and lower with legumes, but when changing from grass silage to a mixture of grass silage and red clover silage, little change in N efficiency occurred, suggesting a synergistic effect of the forage mixture on rumen efficiency. Further work is needed to design forage mixtures, and protein and energy supplements to optimise N efficiency with diets based on high levels of legume silages. Clover silages have high intake characteristics, which are partially offset by lower digestibility in the case of red clover. Whilst yields of white clover grown as a pure stand were disappointingly low (less than half of the yields from grass), these results show that encouraging the presence of white clover in swards will add intake and milk production potential to grass silages.
1.3 The effect of legumes silages on milk fatty acid composition
1.3.1 Introduction Ruminant products have been criticised for their high content of saturated fatty acids and this has contributed to declining consumption. However, milk and beef are also important dietary sources of polyunsaturated fatty acids (PUFA), such as α-linolenic acid and conjugated linoleic acids, which are regarded as beneficial in the human diet because of their effects on cardio-vascular function and possible anti-cancer effects. Rumen bacteria have an extremely high level of biohydrogenation activity against PUFA so that recovery from diet into products is very low. Recent studies have shown increased delivery of PUFA into products through chemical treatment of oils or oilseeds (e.g. Goodridge et al., 2001), though these products are expensive and treatments are not always successful. Although forages contain relatively low levels of fatty acids, they are highly unsaturated and offer a possible low-cost approach to manipulating milk fatty acids. The aim of this work was to examine the effects of legume silages on milk fatty acids from the two dairy feeding experiments reported above in Sections 1.2 and 1.3.
1.3.2 Materials and methods The silages fed and experimental details are as described for Experiments 1 and 2 reported above in Sections 1.1.2 and 1.2.2, respectively. Samples of milk were taken from one block of cows in Experiment 1 and all cows in Experiment 2 and analysed for fatty acid content using the procedure of Sukhija and Palmquist (1988).
1.3.3 Results and discussion All silages were well preserved, aerobically stable and well accepted by the cows. The concentration of fatty acids in the different silages is given in Table 1.3.1. The effects of dietary treatments on silage DM intake, milk yield and the proportions of PUFA are given in Tables 1.3.2 and 1.3.3 for Experiments 1 and 2, respectively. There were no significant effects on milk fat content (mean 45.1 and 37.8 g/kg for Experiments 1 and 2, respectively).
CSG 15 (1/00) 7

Table 1.3.1 Fatty acid concentrations in the silages (g/kg DM)Experiment 1 Experiment 2
G GRC RC GWC WC L G GRC RC WCTotal fatty acids 14.6 14.1 14.3 18.1 21.8 13.9 13.2 15.1 20.0 23.3C16:0 2.9 3.0 3.2 3.5 4.1 3.5 2.9 3.1 3.8 4.2C18:2 2.4 2.8 3.3 2.9 3.7 3.0 2.2 2.7 4.0 4.3C18:3 8.0 6.7 6.0 9.9 12.0 5.7 7.1 7.9 10.4 12.8
Table 1.3.2 Effects on intake, milk production and milk fatty acids (% of total fatty acids; Expt. 1).G GRC RC GWC WC L s.e.d. Sig.
Silage DMI (kg/day) 11.1 11.0 13.5 11.9 12.1 13.6 0.80 P<0.001Milk yield (kg/day) 24.9 28.6 28.1 27.9 31.5 27.7 1.81 P<0.05linoleic acid; C18:2 1.44 1.69 1.82 1.44 1.74 1.51 0.15 P<0.05-linolenic acid; C18:3 0.43 0.53 0.84 0.52 1.04 0.57 0.12 P<0.001
Table 1.3.3 Effects on intake, milk production and milk fatty acids (% of total fatty acids; Expt. 2) G4 G8 GRC4 GRC8 RC4 RC8 WC8 s.e.d. Sig.
Silage DMI (kg/d) 12.8 11.5 13.4 12.5 15.8 14.5 14.6 0.57 P<0.001Milk yield (kg/day) 23.5 27.5 23.7 28.6 25.6 30.2 33.2 0.83 P<0.001linoleic acid; C18:2 0.90 1.05 1.08 1.18 1.47 1.58 1.54 0.047 P<0.001CLA (c-9, t-11) 0.37 0.36 0.45 0.39 0.42 0.41 0.34 0.032 P<0.05α-linolenic acid; C18:3 0.48 0.40 0.77 0.64 1.51 1.28 0.96 0.044 P<0.001
In both experiments clover silages led to increased levels of PUFA in milk compared to grass silage (Tables 1.3.2 and 1.3.3). In the case of white clover silage, the increase was partly explained by the increased silage DM intake and the higher content of PUFA in the silage (Table 1.3.1). Increased rumen passage rates with white clover silage (see Section 2.5) may have reduced rumen biohydrogenation to some extent. The increased α-linolenic acid content in milk from cows consuming red clover silage was greater than would be expected from intake effects, suggesting that some other mechanism was operating, probably by reducing biohydrogenation.
2. THE EFFICIENCY OF PROTEIN AND ENERGY UTILISATION IN IN VITRO AND IN VIVO BEEF AND DAIRY COW RUMEN FUNCTION EXPERIMENTS
A. In vitro studies Microbial protein synthesis in the rumen of the dairy cow supplies at least 50% and often up to 80% of the amino acids absorbed in the duodenum. The efficiency of microbial protein synthesis is therefore a key parameter to measure when attempting to address problems of protein supply by feeding home-grown forages. A number of factors affect the efficiency of microbial protein synthesis, including energy and protein supply to the rumen and the balance in the supply of these two fractions to the rumen microbial population (Dewhurst et al., 2000). The rumen simulation technique (Rusitec) developed by Czerkawski and Breckenridge (1977), with some adaptation, offered an in vitro technique for evaluating factors influencing microbial protein synthesis, in particular assessing the nutritive value to rumen micro-organisms of fresh silages made from alternative forages, unlike other systems where dried and ground feeds are generally used. Experiments were carried out to examine (i) the Rusitec system as a model for investigating rumen function with fresh silages, and (ii) the effect of alternative forage silages (untreated or inoculated) on rumen function.
2.1 Establishment of the procedures to adapt the Rumen Simulation Technique for use with fresh forages
2.1.1 Introduction Previous work (Davies et al., 1999; Lowman et al., 2002) has indicated that the drying and grinding of forages can markedly influence their fermentation characteristics and consequently rumen microbial growth. The aim of this experiment was to adapt Rusitec for use with fresh chopped material instead of the more commonly used freeze-dried and ground feeds, and assess the reproducibility of results on a daily basis, which could become a problem due to the heterogeneous nature of fresh intact and unprocessed silage. Thus an experiment was conducted to evaluate the changes required to ensure the system was suitable for such studies.
CSG 15 (1/00) 8

2.1.2 Methodology for the Rumen SimulationTechnique Feed was fermented in Rusitec under the conditions described below. Each silage ( 12 g DM contained within a Dacron bag) was placed into a perforated container in each of eight incubation vessels at time zero. At the same time an equivalent amount of the pre-digested fibrous fraction of rumen contents was placed into a second bag. The fermentation vessels were charged with strained rumen fluid (from a silage-fed cow) and artificial saliva in the proportion 2.5:1 v/v was continuously infused (700 ml per day representing a dilution rate of 3%/h) over a period of 10 days. From day 3 15N-labelled ammonium sulphate was infused continuously in the artificial saliva. The vessel containing the feed bags was mechanically driven to give a reciprocating action that simulated movement and mixing in the rumen. On each subsequent day a new bag of feed replaced those which had spent 48 h in the vessel. Effluent flowing from the vessel was collected daily in ice-cooled vessels, the volume measured and samples taken and prepared for analysis of VFA, ammonia-N, Total N, dry matter and microbial N. On the last 2 days of each experiment a portion of the residue remaining in the bag after 48 h of digestion was retained for 15N analysis and the remainder washed and used for DM and OM determination. On these days samples of effluent were also taken and mixed rumen bacteria harvested from the remainder by differential centrifugation. 15N and total N analysis was carried out on both effluent and bacteria to allow estimates of microbial protein flow to be made. On the last day of the experiment aliquots (5 ml) of Rusitec fluid were removed from each vessel at intervals up to 24 h from the time that a new bag of feed was placed in the vessel to follow the diurnal pattern of VFA and ammonia concentrations. Experiments were replicated three times using different rumen contents on each occasion to inoculate the vessels and with random distribution of silages between vessels. This general method was adopted in all experiments. In this experiment a farm clamp perennial ryegrass silage (from the IGER farm at Trawsgoed) was used as the feed for the Rusitec. Effluent volume and VFA concentrations in the effluent were analysed daily and the composition and the extent of silage dry matter digestion. 2.1.3 Results and Discussion Results for total VFA, pH and % DM loss are shown in Table 2.1.1. As anticipated there was an initial drop in the VFA concentration over the first 2–3 days after charging the system with rumen digesta, but thereafter until the end of the experiment (after 8 days) fairly constant total VFA concentrations ( Table 2.1.1) and relative molar concentrations of individual VFAs (Table 2.1.2) (within the physiological range of those expected in the rumen) were maintained (Tables 2.1.1 and 2.1.2). At between 86 to 89%, dry matter digestion was also constant over the duration of the experiment, once stable conditions had been attained (i.e., after day 2). At ca 7.0, the effluent pH remained stable throughout the experimental period. The relative concentrations of the different VFAs are shown in Table 2.1.2. Again, after the initial stabilisation period, they remained relatively constant at 46, 19 and 12 mM for acetate, propionate and n-butyrate, respectively.
Table 2.1.1 Daily pH values, total VFA concentrations and dry matter loss where grass silage was fed as the sole feed to the RusitecDay Total VFA mM pH % DM loss
1 105.07 (6.95) 6.54 (0.12) NA2 100.82 (6.44) 6.90 (0.07) 87.46 (1.82)3 92.40 (8.68) 6.93 (0.04) 87.34 (2.03)4 91.29 (12.26) 6.93 (0.09) 89.06 (0.96)5 81.33 (3.86) 6.99 (0.06) 89.59 (0.75)6 85.22 (4.08) 6.95 (0.06) 88.20 (2.54)7 88.02 (5.96) 6.89 (0.06) 88.55 (1.17)
8 85.35 (3.81) 7.02 (0.05) 86.21 (2.02)NA= Not applicable. Each value represents the mean (s.e.) of 8 vessels. The same grass silage was fed in each vessel.
Table 2.1.2 Daily volatile fatty acid profile where grass silage was fed as the sole feed to the RusitecDay acetate mmol/l propionate mmol/l n-butyrate
mmol/li-butyrate mmol/l
n-valeratemmol/l
i-valerate mmol/l
1 59.1 (4.28) 19.1 (1.36) 21.2 (1.05) 1.2 (0.15) 2.0 (0.12) 2.5 (0.25)2 55.8 (3.45) 19.9 (1.18) 18.1 (1.63) 1.5 (0.18) 2.4 (0.17) 4.0 (0.54)3 51.7 (5.46) 18.9 (1.72) 15.4 (2.37) 1.3 (0.11) 2.3 (0.29) 4.3 (0.60)4 49.0 (6.17) 19.5 (2.98) 13.9 (2.37) 1.3 (0.22) 2.4 (0.39) 5.3 (0.61)5 41.75 (2.20) 18.6 (1.21) 12.6 (0.97) 1.0 (0.10) 2.5 (0.24) 4.9 (0.32)6 43.5 (2.08) 19.7 (1.69) 12.5 (0.71) 1.2 (0.08) 2.9 (0.19) 5.6 (0.45)7 46.0 (2.94) 20.9 (1.58) 11.3 (1.23) 1.3 (0.12) 2.9 (0.16) 5.7 (0.57)8 45.9 (2.40) 18.9 (1.22) 9.9 (0.60) 1.3 (0.11) 2.8 (0.17) 6.5 (0.56)
Each value represents the mean (s.e.) of 8 vessels with the same grass silage feed in each vessel.
In conclusion, the results from this experiment indicated that despite the heterogeneous nature of fresh silage the reproducibility of fermentation characteristics on a daily basis was acceptable and that Rusitec is a sutable system for evaluating rumen function parameters in response to feeding fresh silage.
CSG 15 (1/00) 9

2.2 The influence of alternative forage silages on efficiency of rumen function under in vitro conditions
2.2.1 Introduction Much information is available on the efficiency of utilisation of grass silages in the rumen, but there is little published on alternative high protein/energy forages, particularly under conditions where no concentrate supplementation was offered. A number of alternative forage silages were examined to establish the efficiency of rumen function and in particular microbial protein synthesis when offered to the Rusitec as the sole feed. This experiment was used to identify the forages with the greatest potential for improving nutrient utilisation by the ruminant.
2.2.2 Methods One grass silage and 5 alternative forage silages were taken as cores from round bales 90 days after ensiling. The silages were part of both the detailed chemical studies and sheep digestibility trials reported in LS0301. All silages were prepared as bales in 1997 and included second cut untreated grass silage, second maturity untreated Barley/Pea (60 : 40) bi-crop, second cut untreated Lotus, second cut untreated Lucerne, first cut untreated Red Clover and first maturity untreated Kale. Red clover silage was also prepared according to the manufaturer's instructions using an inoculant (Powerstart, Genus plc). These silages were fed unsupplemented to Rusitec using the conditions described in Section 2.1.
2.2.3 Results and Discussion
2.2.3.1 Silage CompositionSome values for the chemical composition of the silages used in this experiment are shown in Table 2.2.1.
Table 2.2.1 Chemical composition of a range of alternative forage silages (all values g/kg DM unless stated)Barley/Pea Lotus Lucerne Red Clover Kale
DM (g/kg FM) 273 325 352 279 138pH 4.33 5.14 5.3 5.3 4.05Ammonia N(g/kg TN) 2.02 31.8 50 95 102.4Crude protein 114 230 207 233 118Lactic acid 44.0 9.4 13 38 145Acetic acid 6.14 5.6 8.3 15 29.1
2.2.3.2 Digestion, pH and VFA production Values for the in vitro DM digestibility (DMD) and OM digestibility (OMD) of the silages and the daily effluent volume, pH and VFA concentrations are presented in Tables 2.2.2 and 2.2.3. Effluent outflow volumes were not significantly different across all treatments, thus any differences in rumen function values were due to forage effects and not differences in microbial growth due to dilution rate. Values for in vitro DMD were consistently high for all crops, in fact higher than the respective in vivo figures for whole tract digestibility in sheep (Akyol et al., 1999). This is largely explained by the 48 h digestion in the in vitro system, compared to the animal trials where retention time of feed in the rumen may have been shorter. The procedure used to remove buffer salts from feed residues could also contribute to the difference, due to small losses of particles from the dacron bag during washing. DMD and OMD values for kale were significantly higher than the other forages (Table 2.2.2), closely followed by red clover, which was significantly greater than the rest. The barley/pea bi-crop had significantly lower DMD than for the other forages, followed closely by lucerne. The general trend, however, in terms of relative digestibility (ranking order) was similar to that found in the in vivo studies). Values for OMD (Table 2.2.2) reflected those for DMD as would be expected. In terms of effluent pH, the feeding of legume silages led to significantly higher values than for the other crops, with kale having significantly lower values than the grass and barley/pea silages, which were intermediate. Total concentrations of VFAs in the vessels (Table 2.2.3) were within the physiological range found in vivo ranging from 71 to 91 mmol/l, respectively, for lucerne and kale silages and the relative concentrations of the individual VFA produced were similar to that generally observed under in vivo conditions for high forage diets. Where the barley/pea and kale diets were fed, the value for propionate was highest, probably due to the higher starch content of these silages. The acetate : propionate ratio was significantly higher for the red clover than all other diets. On the other hand, a significantly lower ratio was found with the barley/pea diet. All other silages had intermediate values. The diurnal variation in VFA concentrations in the vessel was measured and the highest concentrations were seen with the kale silage diet (see Appendix - Figure 2.2.1), reflecting the higher digestibility of this silage. When VFA flows were expressed in terms of OM digested, values were similar with few significant differences (Table 2.2.2), but although this suggests that energy availability was similar, the pattern of production was clearly different (Figure 2.2.1) which has implications for the efficiency of microbial growth and protein synthesis.
Table 2.2.2 Effect of different silages on effluent flow, pH, DMD, OMD, and VFA concentrations in RusitecBarley/
PeaGrass Kale Red
CloverLotus Lucerne s.e.m.
CSG 15 (1/00) 10

Effluent (ml/d) 718a 699a 723a 709a 723a 701a 7.2pH 6.69b 6.70b 6.50a 6.84c 6.83c 6.84c 0.013DMD (%) 65.97a 70.09c 84.30f 78.58e 76.11d 67.80b 0.358OMD (%) 65.53a 69.46a 82.88d 78.45c 72.91b 66.67a 0.746VFA (mol/kgOMAD) 7.43ab 6.93a 7.81b 6.86a 7.28ab 7.05ab 0.190Each value represents the mean of 3 replicated experiments, and values within rows with different superscripts differ significantly (P < 0.05).
Table 2.2.3 Effect of different silages on daily production of individual VFAs (mmol/l) in RusitecBarley/
PeaGrass Kale Red
CloverLotus Lucerne s.e.m.
Acetate 40.78a 45.66bc 55.42d 49.21c 45.63bc 43.02ab 1.032Propionate 18.12c 17.32bc 21.38d 16.08b 16.35b 13.24a 0.346n-Butyrate 10.33d 7.00bc 8.31c 6.44ab 7.11bc 5.46a 0.300i-Butyrate 1.03ab 0.81a 0.87a 1.46c 1.34bc 1.23bc 0.074n-Valerate 2.11a 2.00a 2.02a 2.22a 2.90b 2.95b 0.126i-Valerate 2.73a 2.68a 3.05a 4.33b 5.18c 5.00c 0.126Total VFA 75.09ab 75.45ab 91.06c 79.73b 78.50b 70.91a 1.626Acetate/Propionate ratio 2.26a 2.64bc 2.59b 3.06d 2.78c 3.26e 0.036Each value represents the mean of 3 replicated experiments, and values within rows with different superscripts differ significantly (P < 0.05).
2.2.3.3 Changes in nitrogen fractionsFlows of microbial-N, ammonia-N and non ammonia-N and values for the efficiency of microbial protein synthesis are shown in Table 2.2.4. Flows of microbial protein and non ammonia–N (NAN) were highest with red clover and kale and, in general, grass and barley/pea gave the poorest values, with intermediate values for lucerne and lotus. However, in our studies we did not measure the contribution of ruminal undegraded by-pass protein, which could make a significant contribution to the overall nitrogen status in lotus-fed ruminants. NAN flowing from the vessel was highest for red clover silage, closely followed by values for the lotus silage diet. Ammonia-N concentrations in the vessel were also measured to study diurnal variations (values not shown) and again a similar pattern emerged, with the group of legumes giving the highest values and the bi-crop and kale the lowest. This appeared to be directly related to the input of feed N, which was consistently higher with the legumes, but would also be expected to be related to protein degradability (both in the silo and rumen) and the efficiency of incorporation of ammonia into microbial protein. The pattern of ammonia production was similar for all silages, with a peak forming between 4 and 8 h after feed input to the vessel. There was, however, a lag with lucerne silage, although the highest peak concentrations were observed with this crop, reflecting the refractile nature of lucerne protein. The patterns of ammonia concentration for lotus and red clover were more stable throughout the fermentation period, which reflects the reported resistance of protein from these forages to degradation in the rumen. Overall, the efficiency of microbial protein synthesis (microbial-N/ kg OMAD) was significantly greater for the red clover and kale silages than for the other silage diets and grass silage had a significantly lower value than all other diets. Efficiency was also improved by treatment of legumes with a silage inoculant, for example with red clover ( 9.8 g MN/kg OMAD) it was approximately 13% higher compared to untreated silage (8.2 MN/kg OMAD). In conclusion, the results of this in vitro study gave support to the view that red clover silage, particularly that prepared using a silage inoculant, possessed characteristics which in terms of rumen function efficiency would contribute to enhanced animal performance.
CSG 15 (1/00) 11

Table 2.2.4 Effect of different silages flows of nitrogen fractions on efficiency of microbial protein synthesis Barley/Pea Grass Kale Red Clover Lotus Lucerne s.e.m.
Microbial N Flow (mg/d) 35.63a 29.69a 66.32c 67.69c 49.84b 43.80b 1.721Ammonia N Flow (mg/d) 80.0b 83.8b 53.8a 173.9c 183.1c 188.4c 3.73Non ammonia N Flow (mg/d) 34.5a 30.8a 61.0c 85.6d 53.1c 44.4b 1.76Microbial N (g/kg OMAD) 4.92b 3.89a 7.87d 8.19d 6.38c 6.08c 0.225
Each value represents the mean of 3 replicated experiments, and values within rows with different superscripts differ significantly (P < 0.05). OMAD = Organic matter apparently digested.
2.3 Comparative effects of red clover and lucerne silages on rumen function and microbial N flow and efficiency
2.3.1 Introduction Lack of balance between the form and amount of energy and protein is often cited as the reason why certain forages do not realise their full potential as a sole feed and require concentrate supplementation. The aim of the experiment was to investigate the effect of rates of release of the energy and N fractions of lucerne and red clover and their synchrony on rumen microbial growth and efficiency.
2.3.2 Materials and Methods Red clover and lucerne silages were fed (15 g DM/day) to a Rusitec in vitro system for a period of 10 days. Artificial saliva was infused continuously to give a liquid outflow rate of 2.9%/h and from day 3 (N 15-
H4)2SO4 was included in the buffer as a marker of microbial N flow and daily samples of effluent collected in an ice-cooled vessel. On the last 3 days of each experiment, samples were taken for measurements of rumen function parameters and bacteria were harvested by differential centrifugation from the effluent and also washings derived from the residual feed solids. On the final day of the experiment, a time course study involving destructive sampling of the vessel and its contents at 3, 6, 12 and 24 h post feeding was carried out, with sampling of the liquid fraction in the vessel and residual solid fraction. At the end of this period, bacteria were harvested from the washings derived from the residual solids and also a combined sample of the vessel liquid phase and effluent. The experiment was replicated 3 times. Analysis of the liquid fraction included VFAs ammonia-N, TN and microbial protein. Analysis of the solid fraction included TN, DM and OM loss and microbial protein. Changes in protein and energy (soluble and cell wall sugar) fractions were also followed.
2.3.3 Results and Discussion Some rumen function parameters are presented in Table 2.3.1. There was no difference between the two silages in terms of the rate of DM loss. The results also indicate that approximately 50% of the DM had been degraded in the first 3 h. The rate of ammonia-N release was higher in the cultures fed lucerne silage than for those fed clover silage (18.4 vs 14.3 m mol/l, respectively), suggesting that more rapid and extensive degradation of protein had occurred. However, as lucerne silage contained more protein and ammonia-N concentrations were net values (the product of release and uptake by rumen micro-organisms) it is possible also that the rate of prioteolysis was the similar for both silages. This was coupled with a higher VFA production rate in the red clover-fed cultures than for those fed lucerne, suggesting an improved supply of energy from the red clover. These two factors combined to improve the efficiency of microbial protein synthesis by approximately 15% in the early stages of fermentation with red clover silage (55.3 vs 48.2 g microbial N/kg DM apparently digested for lucerne at 3 h). The results suggest that compared to lucerne, red clover provided a more balanced supply of nitrogen
Table 2.3.1 Diurnal variation in some rumen function parameters for red clover (RC) and lucerne (Luc) silagesTime (h) DM loss (g) Ammonia-N (mM) VFA (mM) Microbial N (g/kg DMAD)
RC Luc RC Luc RC Luc RC Luc0 0.00 0.00 11.43 13.89 60.10 53.83 0.00 0.003 4.56 4.54 14.27 18.44 68.37 59.23 55.30 48.206 5.18 5.14 15.80 21.75 68.0 64.27 55.40 53.27
12 6.93 6.63 13.30 20.54 77.63 72.50 50.00 45.3024 8.62 8.32 12.13 17.82 75.20 72.20 37.37 37.57
DMAD = Dry matter apparent ly digested
and energy, resulting in more efficient use of N by the rumen microbial population. Thus, patterns of release of dietary nitrogen and energy from red clover silage appears to provide a more balanced nutrient supply than for lucerne silage, but further studies are needed to examine energy /nitrogen interactions in more detail.
B. In vivo studies In vitro studies permitted the comparison of large number of treatments and more detailed examination of certain aspects of rumen function in the absence of the effects of absorption and passage, although ultimately in vivo experimentation is a
CSG 15 (1/00) 12

necessary adjunct. A series of in vivo experiments with dairy cows and beef cattle examined the effect of legume silages on rumen function and digestion, with particular emphasis on red clover silage.
2.5 Effects of legume silages on rumen function in Holstein-Friesian cows
2.5.1 Introduction Legume silages have a lower average digestibility than grass silages, and yet often have higher intake characteristics. Thus, poor animal performance associated with feeding silages of low digestibility results partly from the reduced nutrient yield per unit intake, but also from the associated lower intakes, which are presumed to be a consequence of rumen fill effects and patterns of particle size reduction. These factors may also influence the efficiency of microbial protein synthesis in the rumen and microbial N flow to the duodenum. The objective of this work was to understand the rumen mechanisms contributing to the differences observed between animals fed grass and/or legume silage-based diets and the effect of these diets on microbial protein supply to the small intestine.
2.5.2 Materials and methods Six Holstein-Friesian dairy cows (mean initial live-weight 577 kg) with simple rumen and duodenal cannulae were used in a 4-period incomplete changeover design experiment with 28-day periods. Cows were held in individual stalls, offered a flat-rate of 8 kg/day of a standard concentrate and had ad libitum access to one of 6 silages: grass silage (G), red clover silage (RC), white clover silage (WC), lucerne silage (L), and 50/50 (DM basis) mixtures of G and RC (GRC) and G and WC (GWC). The agronomy, ensiling procedures and concentrates were as described in Section 1.1. Fresh forage was offered at 09:00 h each day and concentrates were given in equal portions at milking times (twice daily). A series of measurements of rumen function parameters was made in order to investigate the mechanisms of dietary effects on intake and N-use efficiency as those in Experiment 1, (section 1.1). Measurements included estimates of apparent rumen digestion and microbial protein synthesis in a duodenal flow experiment. Each experimental period lasted for 28 days and measurements were made in the final week of each period. Sample of rumen fluid were taken on day 1 for measurement of rumen parameters and over the following 2 days duodenal digesta was collected. Ytterbium acetate was infused into the rumen (mean 650 mg Yb/day) as a marker to allow estimation of digesta flows at the duodenum. Liquid-phase rumen bacteria were separated from rumen contents by differential centrifugation (1600 × g for 15 min and 30,000 × g for 25 min). A further sample of rumen contents was washed in saline to remove loosely-adherent bacteria, pummeled in a stomacher to detach solid-associated bacteria then isolated by the differential centrifugation as described above. On the penultimate day of each period, rumen contents were emptied by baling, sub-sampled (every twentieth lot) and weighed before being returned to the animal just prior to the morning feed (09:00 h). Rumen contents were again weighed and sampled at 13:00 h on the final day of each period (4 h after feeding). Particle size distribution of rumen digesta was determined by wet sieving (Jones & Mosely, 1977). Results were analysed by analysis of variance using the REML (residual maximum likelihood) directive of Genstat for Windows (Genstat, 2000). The analysis generally used mean values for each cow and period and adopted a fixed model of ‘Diet’ and a random model of ‘Period’ + ‘Cow’. A fixed model of ‘Diet’ × ‘Time’ was used for the analysis of the rumen emptying study (rumen fill and particle size distribution).
2.5.3 Results The effects of forage treatments on feed intake, milk production and milk composition were very similar to those reported in Experiment 1 (Section 1.1) viz. legume silages generally led to increased intakes and milk production, with the highest yields obtained with white clover silage. There were only small effects on rumen VFA concentrations and VFA proportions and the higher N content of the diets based on legume silage resulted in significantly higher rumen ammonia concentrations (data not shown). The results of the rumen emptying study are shown in Table 2.4.1. There were no significant time × time interaction effects on rumen fill. There was a significantly lower quantity of rumen contents and DM when cows were offered alfalfa and, particularly, white clover silages. These two forages were also associated with significant changes in the distribution of particle sizes within rumen contents. White clover silage led to a high proportion of large particles (> 2 mm) and a low proportion of intermediate particles (0.1-2 mm). By contrast, alfalfa silage led to a low proportion of large particles and a high proportion of intermediate particles.
CSG 15 (1/00) 13

Table 2.4.1 Effects of legume silages of the amount, dry matter and particle size distributions of rumen contentsG GRC RC GWC WC L s.e.d. Sig.
Rumen contents (kg) 91.1 87.5 86.1 87.9 69.4 72.9 5.33 ***DM of rumen contents (g/kg) 119 120 129 123 124 129 3.71 *Rumen contents (kg DM) 11.0 10.6 11.2 10.8 8.5 9.3 0.73 ***Rumen contents (kg / kg DM intake) 0.55 0.58 0.57 0.49 0.45 0.43 0.040 ***Proportion of particles >2mm (g/g DM) 0.430 0.374 0.399 0.415 0.440 0.331 0.0235 ***Proportion of particles 0.1-2mm (g/g) 0.227 0.236 0.238 0.195 0.161 0.307 0.0149 ***Proportion of particles <0.1mm (g/g) 0.338 0.390 0.362 0.396 0.398 0.363 0.0174 ***NS = Not significant; *, P< 0.05; *** P < 0.001
The effects of forage treatments on nutrient flows from the rumen are shown in Table 2.4.2. All the legume silages led to increased flows of non-ammonia nitrogen at the duodenum, particularly for white clover silage. There was an indication, although not significant, that red clover N was less degradable than for other silages. Microbial N flow was significantly higher for white clover than all other silages but, although microbial efficiency was higher for both red and white clover, values were not significantly different from that for grass silage. Outflow rates from the rumen were calculated by expressing duodenal DM flow as a proportion of rumen DM pool, from the rumen emptying studies. Both alfalfa silage and white clover silage led to significantly higher outflow rates from the rumen, whilst grass silage led to lowest rumen outflow rates.
Table 2.4.2 Effects of legume silages on flows of nutrients and microbial protein to the duodenumG GRC RC GWC WC L s.e.d. Sig.
Microbial N (g/d) 190 165 182 181 257 171 38.8 †Microbial N (g/kg ADOMR) 22.8 19.1 23.0 17.3 24.1 18.6 4.99 NSDuodenal non-ammonia N (g d-1) 317 370 434 424 571 389 49.4 ***N degradability (%) 69.6 67.4 65.8 74.1 67.1 72.4 5.41 NSDM outflow rate from the rumen 4.26 4.93 4.78 4.84 6.50 6.14 6.14 ***ADOMR = Organic matter apparently digested in the rumen. G = Grass silage; R = Red clover silage; WC = White clover silage; L = Lucerne silage; GRC = mixture of G and RC (50/50 DM basis); mixture of G and W (50/50 DM basis). NS = Not significant; †, P < 0.1*; P< 0.05; *** P < 0.001. † Estimates assume that endogenous-N was 2.8 g/kg DM intake.
2.5.4 Discussion It is clear that the usual relationships between diet digestibility and intake of grass silages do not apply with legume silages, since the least digestible forage, lucerne, had very high intakes. Interestingly, the rumen contents tended to be lowest for cows offered lucerne and particularly white clover. It seems that the high rate of fermentation of white clover means that intake is not constrained by fill, so that very high intakes can be attained without cows becoming full. Other studies have shown the very high intake characteristics and milk production from white clover, even as the sole feed (Castle et al., 1984; Auldist et al., 1999), which probably reflects its low rumen fill effect. The rumen emptying studies suggest that the beneficial effects of white clover on passage rates may be neutralised by grass silage, and it is possible that particles of other forages become trapped in the recalcitrant mat formed from grass silage. The distribution of particle sizes in the rumens of cows fed ryegrass silage and lucerne silage were similar to those observed by Waghorn et al. (1989) for the fresh forages. It is suggested that high intakes of white clover reflect its high degradability, whilst high intakes of lucerne are made possible by a high rate of particle clearance from the rumen. . There was some evidence of reduced N degradation with red clover, possibly due to the action of plant polyphenol oxidase activity and resultant binding of polyphenols to protein during ensilage; with white clover, reduced N degradation was probably as a consequence of high rates of passage from the rumen. All of the legume silages led to an increase in the flow of non-ammonia-nitrogen at the duodenum. Some of this increase is explained by the tendency for increased flows of microbial nitrogen. There was no significant treatment effects on microbial energetic efficiency (g microbial N/kg apparently digested OM in the rumen), though values were highest for treatments with highest rumen passage rates.This latter effect would be expected according to conventional microbial energetics (Pirt, 1965), if high rumen passage rates of DM are associated with higher microbial growth rates. Increases in Metabolisable Protein (duodenal non-ammonia N) supply result from the increased supply of undegraded N from legume silages, particularly red clover.
2.6 Effects of legume silages on rumen function in Hereford x Friesian steers.
2.6.1 Introduction The high intakes and milk production from legumes silages documented in the dairy cow experiments reported in Section 1.1 were reflected by differences in microbial N flow and but not efficiency, although this may have been masked by the feeding of a flst rate of concentrates. This experiment was carried out to examine the effect of legume silages on rumen function and microbial protein synthesis in steers when silage was offered as the sole feed.
CSG 15 (1/00) 14

2.6.2 Materials and methods Ten Hereford x Friesian steers 362 (se 7.7) kg, prepared with rumen and duodenal cannulae were allocated at random to receive one 5 silage diets ad libitum; grass (G); grass and white clover (60 : 40 DM basis; GW); grass and red clover (60 : 40 DM basis; GR); white clover (W) and red clover (R). The experiment was conducted as a 2 period changeover design within each legume treatment, with grass silage as an experimental control. Animals allocated to red clover or white clover remained on the same legume treatment, but the percentage of legume changed across the periods. Animals allocated to grass silage remained on that feed in both periods. Each period lasted 24 days, with a 14-day adaptation period to the diets, followed by a 10-day measurement period. Digesta flow at the duodenum and ileum were estimated using a dual-phase marker system with ytterbium acetate (YbAc) and chromium ethylene diamine tetra-acetic acid (CrEDTA) as the particulate and liquid markers, respectively. Digesta flows at the duodenum and ileum were estimated using a dual-phase marker system with ytterbium acetate (YbAc) and chromium ethylene diamine tetra-acetic acid (CrEDTA) as the particulate and liquid markers, respectively. On days 21 and 22 duodenal digesta was collected every 4 h over a 24 h period. Samples of rumen fluid were taken every 2 h over a 24 h period (day 23) and on day 24 rumen fluid was sampled, 2 h post feeding, for microbial isolations by differential centrifugation. Statistical analysis was carried out using REML, with silage as the fixed effect and animal + period × period as the random effect (Genstat 5; Lawes Agricultural Trust, 1987).
2.6.3 Results Nutrient intakes and duodenal flows of nutrients are presented in Table 2.5.1.
Table 2.5.1 Nutrient intake and duodenal flow in steers fed the experimental silages: (G) grass; (R) red clover; (GR) red clover and grass; (W) white clover and (GW) white clover and grass.
G R GR W GW S.e.d. P*Intake (kg/d) Dry Matter 4.15a 7.02b 6.36b 8.48c 8.43c 0.641 * Organic Matter 3.75a 6.28b 5.75b 7.73c 7.66c 0.592 * Total Nitrogen 1.07a 2.24cd 1.70bc 2.93e 2.47de 0.233 ***
Duodenal Flow (kg/d) Dry Matter 2.36a 4.13ab 3.54a 4.81b 4.38b 0.899 * Organic Matter 1.68a 3.19b 2.58ab 3.47b 3.15b 0.580 * Total Nitrogen 0.84a 1.78bc 1.34ab 2.25c 1.84bc 0.404 * Microbial Nitrogen 0.581a 0.901ab 0.822ab 1.05b 1.10b 0.190 * EMPS† (gMN‡/kg OMAD§) 28.5 30.5 25.3 24.7 26.5 6.01 NS
Duodenal flow per unit intake (kg/kg DMI║) Dry Matter 0.581 0.589 0.554 0.556 0.517 0.0621 NS Organic Matter 0.415 0.457 0.403 0.402 0.378 0.0451 NS Total Nitrogen 0.208 0.253 0.210 0.259 0.219 0.0303 NS Microbial Nitrogen 0.141 0.132 0.121 0.133 0.142 0.0232 NS† Efficiency of microbial protein synthesis; ‡ Microbial nitrogen; § Organic matter apparently digested;║ Dry matter intake; Values with different superscripts differ significantly.
Intakes of all diets based on clover silage were significantly higher than for the grass silage diet. Total nitrogen flows were significantly higher for the pure legume diets, which is not surprising in view of the higher N content and intakes of these silages. Legume silages, particularly white clover, led to significantly greater flows of microbial nitrogen, though there were no significant differences in the efficiency of microbial protein synthesis (EMPS) on any of the treatments. Duodenal flows of microbial N expressed per unit of intake were also similar and not significantly different. There were no significant effects of diet on rumen characteristics (not shown) except for the ruminal concentration of n-butyrate, which was significantly higher when the white clover silage was fed.
2.6.4 Discussion The diets based on clover silages led to significantly higher DM intakes than the grass silage. This effect was consistent with the results of earlier studies with beef steers (Thomas et al., 1981). The higher DM intakes with the clover silages may relate to their lower fibre content (Thornton & Minson, 1972) and increased rates of particle breakdown (Dewhurst et al., 2001). Grass silage intake is often limited by the long retention time of grass fibre in the rumen. Of all the silages used in this work, grass silage had the worst fermentation profile (highest pH, ammonia-N and n-butyric acid concentrations) and this may have contributed to lower intakes (Gordon et al., 1998). All of the diets were composed entirely of forage and the analysis of rumen fluid showed extremely stable rumen conditions, with high pH and modest VFA concentrations, with little difference between treatment groups. The only significant effect on rumen measurements was an increased level of n-butyric acid with the diets that included white clover silage. Similarly, there were no differences in the diurnal patterns of rumen parameters. These results are in agreement with the findings of Dewhurst et al. (2001) who fed similar forages to dairy cows, although with concentrate supplementation. Microbial
CSG 15 (1/00) 15

nitrogen flow at the duodenum was significantly higher when feeding clover silage, particularly white clover silage. However, a substantial part of this effect related to the DM intake effect noted above and the efficiency of microbial protein synthesis (EMPS) was not significantly different across the four treatments, which supports the observations in dairy cows (see Section 2.5). The mean EMPS value across treatments was 27.1 g MN/kg ADOMR, which is similar to the average value (30) recognised by ARC (1980).
2.7 General conclusions on in vivo rumen function studies in cattleIncreased feed intake appears to be a major driver behind the effects of legume silages on animal production, and effects can largely be explained by the degradation characteristics of the different silages in the rumen. Although there were no differences in microbial efficiency in terms of organic matter digested across all silages examined (including grass silage), the poorer efficiency of N use for production with legume silages could in part be addressed by manipulating the form and amount of readily available energy in the diet. Further studies are needed to balance the levels and types of complementary forages or supplements for legume silages. Forage breeders also need to consider ways in which digestibility is achieved (i.e. low vs high rates of fermentation and passage) in order to ensure high digestibility, high intakes and consequent maximum production from forage.
CONTINUATION PROGRAMME FOR YEARS 4-5 (2000-2002)The programme for years 4 and 5 was divided into two discrete parts (A and B) with the following objectives:
A To identify the best ensiling practice (relative contribution of legume: high sugar grass) to maximise red clover silage quality, in particular the residual protein and WSC content of silages, and the test combination of red clover silage to mix with high sugar ryegrass silage in order to maximise rumen microbial protein synthesis and efficiency of N use.
B To investigate plant and microbial activity in grazing ruminants (bolus microbiology and biochemistry; in association with LS0303); develop novel procedures to examine the biochemistry and microbiology of ingested forage boli and examine microbial succession on ingested boli in vivo.
3. ENSILING AND RUMEN FUNCTION STUDIES WITH GRASSES AND LEGUMES
3.1 Ensiling of red clover with and without high sugar grasses
3.1.1 Introduction Research on the silage fermentation has generally been targeted at maximising the efficiency of use of water-soluble carbohydrate (WSC) for lactic acid production, and inoculants containing lactic acid bacteria (LAB) are often used. Novel perennial ryegrass lines with exceptionally high WSC content provide opportunities to retain both high quality protein and high levels of WSC in the silage. The aim of this experiment was to examine the effect of different additives on the quality of silages prepared from ryegrasses containing either high or standard levels of WSC, ensiled either alone or in combination with red clover.
3.1.2 Materials and Methods The herbages used were a first cut of either perennial ryegrass with a high WSC content (experimental variety Ba11353) or a standard WSC content (cv. Aberelan) or red clover (cv. Melle). All herbages were wilted overnight and chopped with a precision chop forage harvester, prior to treatment and packing. Herbages were ensiled either pure or as mixtures with red clover and the respective grass in a ratio of 70 : 30 (clover:grass on a fresh matter basis). Each treatment was ensiled in triplicate, in 1 kg glass laboratory silos. Ensilage methods were as detailed by Cussen et al. (1995). All herbages were ensiled either untreated, inoculated (Powerstart applied according to the manufacturer's recommendations; Genus Plc, Crewe, Cheshire, UK) or acidified (Add Safe at 6 l tonne -1; BP Nutrition, Northwich, UK). Silos were opened after 90 days and analysis of WSC, lactate, ammonia-N, total N and pH was carried out on sub-samples of the silage, as detailed in Cussen et al. (1995). Differences in silages as a result of treatment and herbage type were determined using two-way analysis of variance (Genstat 5; Lawes Agricultural Trust, 1987).
3.1.3 Results and Discussion The results presented in Table 3.1.1 show that for all chemical analyses there were highly significant treatment, herbage and interaction effects (P<0.001). Ensiling ryegrass with a high WSC content resulted in silages with significantly higher levels of WSC than that produced from the control grass. Inclusion of red clover with the high WSC ryegrass (for acidified and inoculated silages) also significantly improved residual WSC concentrations, in comparison with silage produced from the WSC grass mixed with red clover. Two other ratios of grass to red clover (50:50 and 70:30 were examined but similar trends were observed and results are not included here. With respect to the 3 treatments used, acidified silage has a higher WSC concentration than inoculated silage, which in turn was higher than for untreated silage. Ammonia N concentrations for untreated silage were higher than for acidified silages, which in turn were higher than for inoculated silages. Thus, in terms of silage quality, direct acidification with formic acid preserved more WSC, whereas inoculation preserved more protein.
Table 3.1.1 Chemical composition of silages at 90 days after ensiling.
CSG 15 (1/00) 16

Herbage Type Treatment pH WSC g/kg DM Lactate g/kg DM NH3-N g/kg TN
CSG 15 (1/00) 17

Red clover Untreated 5.57d 11.48a 10.96b 179.3f
Inoculated 3.93a 19.81a 74.32f 17.2a
Acidified 4.56c 89.17ef 24.32c 37.6bcd
High WSC grass Untreated 4.31c 49.34b 44.60e 65.9e
Inoculated 3.74a 121.52g 91.54h 25.4abc
Acidified 4.61c 266.61h 15.77b 50.4de
Control grass Untreated 5.59d 6.55a 1.66a 178.7f
Inoculated 3.98ab 15.04a 83.28g 41.8cd
Acidified 4.22bc 70.48cd 47.91e 51.0de
Red clover:High WSC grass (70:30)
Untreated 5.69de 8.93a 2.47a 205.0g
Inoculated 3.73a 58.66bc 95.07I 22.4ab
Acidified 4.32c 97.13f 35.48d 48.4de
Red clover:Control grass (70:30)
Untreated 5.94e 8.24a 2.53a 222.2g
Inoculated 3.86a 17.80a 86.39gh 26.9abc
Acidified 4.23bc 77.18de 40.42de 39.7bcd
s.e.d. 0.134 6.980 3.680 9.07Treatment means within columns with different superscripts differ (P<0.05).
The results presented here indicate that with judicious selection of additive treatment, well fermented silages with low ammonia-N contents (reduced protein degradation) can be prepared from high WSC grasses, but with the added benefit of exceptionally high residual WSC content. This latter benefit also extends to the ensiling of red clover, where mixing of this herbage with high sugar grass prior to ensiling confers similar benefits on silage quality. The highest WSC values were observed in silages treated with high levels of formic acid. The relative benefits of this extra WSC has to be weighed against the more environmentally-conscious biological approach using inoculants, where WSC contents were still exceptionally high.
3.2 In vitro studies of rumen function in continuous cultures fed red clover silage, high sugar grass silage or corresponding mixtures
3.2.1 Introduction An insufficient supply of readily available sugar relative to a surplus of soluble protein in grass and legume silages can often lead to inefficient use of silage-N for rumen microbial growth and protein synthesis. Novel perennial ryegrass cultivars (Humphreys, 1989) bred for elevated levels of water soluble carbohydrate (WSC), present an opportunity to increase the available energy supply to the rumen and improve microbial growth efficiency. As observed in Section 3.1 a high proportion of this extra sugar can be retained in silage prepared from grass with high WSC content (Davies et al., 2002). The objective of this experiment was to determine how grass silages with high levels of residual WSC influence rumen microbial growth and the efficiency of use of grass and legume silage N under in vitro conditions.
3.2.2 Materials and Methods Primary growths of 2 perennial ryegrass varieties, (i) an experimental cultivar (Ba11353) bred for high sugar content, or (ii) AberElan, a control grass and also a red clover (cv. Milvus) were cut, wilted and precision chopped. They were all treated with an inoculant (Powerstart; Genus plc) containing Lactobacillus plantarum and Lactococcus lactis (106 colony forming units/g FM) and ensiled in glass jars (1 kg) for 90 days. An 8-vessel in vitro simulated rumen culture system (Rusitec; Czerkawski & Breckenridge, 1977) was fed (11.1 g DM daily) with the above silages or mixtures; high sugar grass (HSG), control grass (CG), red clover (RC), HSG:RC at ratios of 70:30, 50:50 or 30:70 (dry matter basis), or CG:RC at ratios of 70:30 and 30:70. Artificial saliva was infused continuously into the culture vessels at a rate of 0.7 volumes/day and feed was introduced every 24 h. ( 15NH4)2 SO4 was added to the artificial saliva and infused as a microbial marker and samples of washed bacteria, vessel contents and effluent were taken 7 days after addition of (NH4)2S04, when steady state had been reached. Flows of organic matter and ammonia-N and VFA concentration in the effluents were estimated and daily microbial N flows and efficiency were calculated from values for 15N enrichment of the effluent and harvested microbial fractions. Differences between silages were determined using two-way analysis of variance (Genstat 5; Lawes Agricultural Trust, 1987).
3.2.3 Results and discussion Before ensiling, the high sugar grass (HSG), control grass (CG) and the red clover (RC) had WSC contents of 244, 126 and 118 and total N contents of 19.0, 22.1 and 30.3 g/kg DM, respectively. Some chemical characteristics of the silages produced from these forages as fed are shown in Table 3.2.1. The silages all had pH values of < 4.0 and high lactic acid contents with ammonia-N concentration being lowest in the high sugar grass silage.
Table 3.2.1 Chemical composition of grass and red clover silages [Mean of 3 replicates (SE)].DM
(g/kg FM)pH Lactic acid TN WSC Ammonia-N
(g/kg TN)(g /kg DM)
CSG 15 (1/00) 18

HSG 24.3 (0.04) 3.58 (0.06) 161.3 (1.95) 22.1 (0.30) 85.7(4.07) 79.0 (1.35)CG 23.6 (0.04) 3.72 (0.12) 150.9 (0.55) 24.6 (0.16) 30.0(2.55) 116.7 (1.80)RC 23.3 (0.28) 3.77 (0.06) 139.5 (0.55) 32.9 (0.06) 30.1 (0.25) 94.6 (1.75)
The WSC content of the HSG silage was approximately 3 times higher than for the CG silage, which had a similar content to the RC silage. Thus, a range in the ratio of WSC:TN was achieved across the treatments examined. These silages were fed to in vitro rumen cultures and values obtained for ammonia-N and volatile fatty acid (VFA) concentrations in the effluents, organic matter apparently digested (OMAD) and microbial N flow and efficiency (Table 3.2.2).
Table 3.2.2 Microbial N production and fermentation parameters in rumen cultures fed high sugar grass, control grass and red clover silages or corresponding mixtures.
Ammonia-N(mmol/l)
VFA (mmol/l)
OMAD(g/d)
MN(g/d)
EMPS(g MN/kg OMAD)
High Sugar Grass (HSG) 7.1 88.3 9.2 0.217 23.6Red Clover (RC) 12.5 77.8 8.5 0.197 23.170% HSG:30% RC 9.1 89.9 9.1 0.235 25.850% HSG:50% RC 10.8 80.9 8.9 0.243 27.330% HSG:70% RC 11.1 81.1 8.8 0.252 28.7Control Grass (CG) 9.1 87.2 8.3 0.179 21.470% CG:30% RC 10.5 81.1 8.4 0.198 23.630% HSG:70% RC 12.5 86.7 8.5 0.213 25.3s.e.m. 0.67 3.23 0.10 0.0093 0.95l.s.d 2.040 9.80 0.319 0.0283 2.909OMAD = Organic Matter Apparently Digested; MN = Microbial N; EMPS = Efficiency of Microbial Protein Synthesis
Only small differences in ammonia-N concentration were observed between the different silages, with a trend for significantly lower values with the HSG and HSG/RC mixed silages than for RC, CG and the corresponding CG/RC mixtures. Values for organic matter apparently digested (OMAD) were significantly higher with HSG compared to the CG and RC silages, but were highest where > 50% of the mixed silage was HSG. Microbial N (MN) flows were significantly higher for HSG than CG silage but the efficiency of microbial protein synthesis (EMPS; g MN/kg OMAD) was higher, but not significantly different. However, the efficiency of conversion of feed N into microbial N (0.85g MN/g N input for HSG vs O.65 for CG) was significantly (p<0.001) higher for HSG silage. Mixing of RC silage with HSG silage significantly increased MN flows and EMPS values, with a graded response, the highest values being seen where HSG silage was included at the 30% level. This approach provides a forage-based strategy for balancing the N and energy supply for rumen microbial growth and increasing N use efficiency when feeding legume silage to ruminants.
3.3 Effects of red clover silage, high sugar grass silage or corresponding mixtures on rumen function and digestion in Hereford x Friesian steers.
3.3.1 Introduction The in vitro findings reported in Section 3.2 suggested that microbial N flow and efficiency of microbial synthesis in the rumen, both in terms of energy and in particular N supply can be improved using high sugar grasses to prepare silages with high residual WSC content. There was also a suggestion that the mixing of high sugar grass silages with red clover could improve microbial growth efficiency and N use with diets based largely on red clover. This experiment was carried out to confirm the in vitro findings and demonstrate the advantages of grass silages containing high levels of residual WSC in increasing N use efficiency in the rumen.
3.3.2 Materials and methodsA first regrowth of two perennial ryegrass swards; Ba11353, an experimental variety bred to express high levels of WSC (HSG), and Fenemma, the control (CG), were cut in June 2001, wilted for 24 h and harvested with a precision-chop forage harvester and ensiled in big bales. A second re-growth sward of a monoculture of red clover (Trifolium pratense) was cut in August 2001 and also wilted for 24 h and ensiled in big bales. A silage inoculant (Powerstart; Genus plc, Crewe) was applied to all crops at a rate of 2 l/tonne FM. Six Hereford × Friesian steers, initial liveweight 163 (s.e. 5.9) kg, prepared with a rumen cannulae and simple ‘T’-piece cannulae in the proximal duodenum were used. The experiment was conducted as a 5 x 5 incomplete Latin square with an additional randomly repeated sequence. There were four experimental periods with six animals being allocated to one of the five diets: high WSC grass silage (HSG); control grass silage (CG); high WSC grass and red clover silage (50:50 DM basis; HGR); control grass and red clover silage (50:50 DM basis; CGR); and red clover (R). Each period lasted 24 days, with a 14 day adaptation period to the diets,
CSG 15 (1/00) 19

followed by a 10 day measurement period for collection of rumen and duodenal fluid. Silages were offered ad libitum in two meals at 09:00 h and 16:00 h.
3.3.3 Results The WSC contents of the high sugar and control grasses after a 24 h wilt were 210.4 and 163.7 g/kg DM, respectively. The chemical composition of all silages and silage mixtures are shown in Table 3.3.1. All silages were well preserved with pH values ranging from 4.04 - 4.12. Both grass silages were treated with a silage inoculant and the rapid fermentation led to high residual WSC contents, but it was significantly higher in HSG silage (by approximately 3.5 percentage units) than for CG silage. The mixtures of HSG and CG silages with red clover silage had correspondingly lower WSC values due to the dilution effect. The ADF and NDF contents of HSG and CG silages were not significantly different and ammonia N values were also very similar.
Table 3.3.1. Chemical composition of the experimental silages: (HSG) high WSC grass; (CG) control grass; (HGR) high WSC and red clover; (CGR) control grass and red clover; (R) red clover.
HSG CG HGR CGR R S.e.d. PDry Matter (g/kg) 280.8ab 306.6b 277.8a 258.8a 284.4ab 1.32 0.046Organic Matter (g/kg DM†) 918.7cd 924.0d 908.0b 913.7bc 894.7a 8.02 0.001pH 4.08ab 4.12b 4.11b 4.04a 4.12b 0.023 0.020ADF‡ (g/kg DM) 327.8b 333.3b 308.0a 320.3ab 317.2a 6.14 0.013NDF§ (g/kg DM) 551.0bc 587.0c 459.0ab 481.9ab 397.0a 47.8 0.001Ether Extract (g/kg DM) 30.0 31.6 33.9 39.2 32.4 3.18 NSWSC║ (g/kg DM) 90.5d 55.3bc 66.5c 43.7ab 29.3a 8.23 0.001Gross Energy (MJ/kg DM)Ammonia-N (g/kg total-N) 110.8 110.5 116.0 108.7 113.0 6.15 NSTotal Nitrogen (g/kg DM) 24.5a 25.4a 29.4b 29.2b 32.2c 1.13 0.001Lactic acid (g/kg DM) 81.5 79.1 76.6 79.1 77.8 4.65 NS† Dry matter; ‡ Acid detergent fibre; § Neutral detergent fibre;║Water soluble carbohydrate; Values with different superscripts differ significantly at the P values shown.
The nutrient intakes and duodenal flows of nutrients are presented in Table 3.3.2. Results with P values of < 0.10 were considered to be significantly different
CSG 15 (1/00) 20

Table 3.3.2 Nutrient intake and duodenal flow in steers fed the experimental silages: (HSG) high WSC grass; (CG) control grass; (HGR) high WSC and red clover; (CGR) control grass and red clover; (R) red clover.
HSG CG HGR LGR R S.e.d. PIntake Dry Matter (kg/d) 4.44b 3.55a 4.70b 4.57b 4.51b 0.439 0.07 Organic Matter (kg/d) 4.08ab 3.29a 4.27b 4.17ab 4.04ab 0.400 0.102 Total Nitrogen (g/d) 96.8a 88.6a 137.2b 133.3b 145.5b 12.20 0.001
Duodenal Flow Dry Matter (kg/d) 3.15b 2.47a 3.38b 3.30b 3.11b 0.299 0.044 Organic Matter (kg/d) 2.48b 1.84a 2.58b 2.55b 2.43b 0.283 0.073 Total Nitrogen (g/d) 107.0b 77.8a 116.0b 110.0b 112.7b 10.84 0.015 Microbial Nitrogen (g/d) EMPS (g MN/kg OMAD)
66.0c
40.6b40.2a
29.9a 58.5bc
35.5ab 59.7bc
37.3ab 42.8ab
27.6a 8.08 4.75
0.0180.077
† EMPS = Efficiency of microbial protein synthesis; MN = Microbial nitrogen; OMAD = Organic matter apparently digested in the rumen;║ Dry matter intake; Values with different superscripts differ significantly at P values shown.
Intakes of diets based on RC silage and also the pure HSG silage were significantly higher (p<0.070) than for the control grass silage diet. Total nitrogen flows were highest for the pure legume diets and the mixtures, which is not surprising in view of the higher N content and intakes of these silages. HSG silage led to significantly greater flows of microbial nitrogen than for CG silage (p<0.015) and also increases in the efficiency of microbial protein synthesis (p<0.077) in terms of OM digested in the rumen. The efficiency of microbial protein synthesis was also higher, although not significantly different for the mixtures of CG and HSG with RC silage, in comparison with red clover silage alone. There was a significant effect of diet on rumen ammonia-N concentration (not shown), with HSG having a lower value than CG and the mixtures (HGR and LGR) both being higher than for pure red clover silage. When the microbial efficiency of N use in the rumen was calculated in terms of microbial N flow per unit of N intake, nearly 70% of the high sugar grass silage-N was incorporated into microbial N, compared to 45% with the control grass silage and the mixtures and <30% for red clover alone.
3.3.4 General discussion and conclusions for in vitro and in vivo studiesThe results of these in vitro and in vivo experiments support the view that N use efficiency in the rumen in animals fed grass silages can be markedly enhanced if the high levels of WSC in IGER grass varieties can be retained in the silo. There is also an indication that if the difference in level of sugar is above a threshold value of 4 percentage units the mixing of grass silage with red clover silage could reduce the N losses seen in other work with legume-based diets (Dewhurst et al., 2003). Both observations have implications for reducing environmental pollution and the sustainability of livestock agriculture.
4. BOLUS BIOLOGY AND MICROBIAL ECOLOGY STUDIES
Plant enzymes have been shown to play a role in protein degradation in the early stages of digestion of fresh forage in the rumen (Kingston-Smith & Theodorou, 2002), a process previously assumed to be mediated by gut micro-organisms. Thus, some re-evaluation of current feeding systems and particularly protein rationing schemes may be needed to take into account plant enzyme mediated processes and the role that they play in modifying nutrient supply and utilisation in the rumen.
Much of the digestive activity in the rumen (both microbially and plant driven; see Report for LS0303, Section 1.7) occurs within the first 3 to 6 h after ingestion of plant biomass, which has focused our attention on the implications that this has for nutrient supply and thus the successional microbiology associated with the colonisation of fresh forages. An understanding of how different forage types perturb the rumen ecosystem and influence rumen microbial ecology and thus rumen function will provide opportunities to use plant-based strategies for manipulating rumen function and N use efficiency, whilst reducing the level of environmental pollution. The aim of this work was to investigate and develop a series of ‘tools’ to study the early stages of digestion of fresh forages, in order to obtain a more precise understanding of the initial events during colonisation and degradation of fresh forage, in particular feed boli. These included (a) a procedure to enable capture of freshly ingested forage boli, in vitro and in vivo (in sacco), (b) procedures for studying rates of digestion of bolus material, (c) microbial marker and molecular techniques for following the temporal colonisation and succession of microbial populations under in vivo conditions and (d) a novel approach for examining plant mediated proteolysis in the rumen under in vivo conditions.
4.1 Development of a bolus capture technique and its use for studying microbial colonisation of boli from freshly ingested herbages
CSG 15 (1/00) 21

4.1.1 Introduction Plant proteins are degraded extremely rapidly in the rumen by both plant enzyme and microbial activity and the accompanying microbial colonisation process which presumably takes place in response to release of nutrients is probably equally fast. Little work has been done to study these early stages of digestion, particularly with ruminants grazing fresh forage. The concept of 'bolus biology', or using the feed bolus that is formed during ingestion of fresh herbage represents a novel approach to studying early events in the digestion and microbial colonisation process. We propose that incubation of boli in the rumen in sacco could provide insights into microbial colonisation and succession of different types of herbage. The aim of this work was i) to develop a procedure for capturing feed boli and ii) to examine a combination of fatty acid profiling (of rumen bacteria) and denaturing gradient gel electrophoresis (DGGE) for studying microbial colonisation of boli from two contrasting fresh forages. This study is complementary to that concerned with plant- and microbially-mediated protein degradation described in Section 4.2. The fatty acid marker technique permits broad characterisation of gross shifts in bacterial populations, based on appearance of particular 'marker' fatty acids that are specific to different bacterial species. The DGGE approach is a molecular technique in which total genomic DNA (plant and microbial) is isolated from forage bolus samples and subjected to PCR using anaerobic fungal- or bacterial-specific primers, creating a population of rDNA fragments representative of the anaerobic gut fungal and bacterial populations present in these samples. This allows the temporal sequence of microbial colonisation of fresh herbage to be observed during incubation in the rumen.
4.1.2 Materials and Methods Friesian/Holstein dairy cows with rumen fistulas were used as the donor animals to develop a method for collecting the down boli formed immediately after ingestion of fresh grass . The cows were denied access to feed for 30 min, and the rumen emptied manually. Once the rumen had been emptied the cows were allowed access to zero-grazed grass. Upon chewing and bolus formation in the mouth, the grass bolus was swallowed and captured (by hand) at the oesophageal/rumen aperture, immediately prior to making contact with the rumen wall or any residual rumen digesta. Approximately 3 kg of rumen boli were collected from each animal. Bacterial and fungal numbers were enumerated on the bolus material using an anaerobic most probable numbers procedure (Theodorou et al., 1990) and 150 g portions were weighed immediately into pre-weighed Dacron bags and incubated in the rumen for 0, 1, 2, 4, 8, 24 and 72 h. After removal from the rumen, bags were gently rinsed in cold water, freeze-dried and weighed for estimation of DM disappearance. Fatty acid methyl esters were prepared (methanolic HCl, 5%), extracted and determined by gas chromatography using tricosanoic acid (C23:0) as the internal standard. Genomic DNA from dacron bag residues was obtained using Fast DNA® Kit (BIO 101 Protocol, Catalog# 6540-400, CA92008, USA) in combination with the FastPrep® instrument. Anaerobic fungal-specific PCR primers were designed and used to amplify the variable internal transcribed spacer 1 (ITS1) region of the anaerobic gut fungal ribosomal DNA (rDNA) repeat. This region of the rDNA repeat displays size and sequence heterogeneity between anaerobic gut fungal isolates. Size variability of the PCR amplified rDNA fragments were then used to electrophoretically separate the population using high-resolution Spreadex gels. Subsequent retrieval of sequence information enabled positive identification of anerobic fungal community members. Initial development and optimisation of the precedure used axenic cultures and anaerobic fungi found in ruminant faeces. For bacteria the V3 variable region of 16S rDNA was amplified by PCR using rumen bacterial primers specific for conserved regions of the 16S rRNA genes as described by Muyzer et al. (1993). The reaction mixture contained approximately 125 ng of genomic DNA, 25 pmol of each primer, 4 l of dNTP mixture, 5 l of 10 x ExTaq buffer and 0.5 l of TaKaRa Ex Taq polymerase (TaKaRa Shuzo, Otsu, Japan). The DGGE was performed using a Bio-Rad D-Code System (Hercules, CA). And amplicons were separated using 35-60% linear gradient of denaturant.
4.1.3 Results and Discussion Anaerobic bacteria (1x107 CFU g-1 FM) were detected on the bolus, which is ca 1000-fold fewer than for the average rumen microbial population. Anaerobic fungi were not detected on the bolus using the MPN technique, although molecular approaches (see below) were successful in the isolating fungal DNA from boli. These results suggest that herbage is ‘inoculated’ (but not colonised) by coming into contact with micro-organisms in the oesophagus prior to contact with rumen fluid. These micro-organims are on the oesophagus wall, presumably because of prior ‘ruminations’. It is not surprising that population sizes of anaerobic fungi were lower in the oesophagus as in this experiment they were probably outside of the detectable range of the MPN technique. Earlier studies (Lee et al., 1999) identified distinctive odd-chain fatty acids that could be used as microbial markers. Two of these fatty acids ( iso C15:0 and iso C17:0) were not detected in grass or white clover from pasture (Table 4.2.1), but were found in washed bacterial preparations obtained from the rumen, indicating that they could be used as microbial markers.
Table 4.2.1 Fatty acid content of fresh herbages Fatty acid (mg/kg DM) Grass White cloverC15:0 21 55iso C15:0 trace 0anteiso C15:0 926 297
CSG 15 (1/00) 22

C17:0 28 62iso C17:0 0 0anteiso C17:0 34 36
The concentration of both these fatty acids in harvested boli of both forages increased with incubation time. The relationship between the concentration of iso C15:0 (mg/kg DM) and DM disappearance (g/g DM) was described by linear regression: for the grass: Y = 317.3X – 104.5 (r2=86.9, s.e. 19.8, P<0.01); and for white clover: Y = 96.1X – 13.2 (r2=66.4, s.e. 18.0, P<0.01), respectively (see Appendix - Figure 4.2.2). The results suggest rapid and continued microbial colonisation of both grass and white clover, concomitant with loss of digestible DM. The bacterial PCR-DGGE analysis demonstrated rapid (within 2 h) and continued colonisation of both grass and white clover (see Appendix - Figure 4.2.3). Bacteria appeared to colonise plant substrates in succession and diversity differed according to both plant type and individual animal. However, further analysis of sequences retrieved from the gels revealed that many bands excised from the gel were of plant chloroplastic origin (or originated from plant chloroplasts), especially at the early sample collection times (0-4 h). Subsequently, several standard primer sets for the study of bacterial diversity based on the 16S rDNA molecule were compared with the sequences of the maize chloroplast using BLASTn and most were found to be very similar. The primer set targeting the V3 region of 16S rDNA is therefore unsuitable for the study of plant-microbe interactions in the rumen. Further analysis with a range of different primer sets is in progress to resolve this problem.
Anaerobic gut fungal populations were also analysed in bolus material recovered after incubation in the rumens of animals fed on grass or white clover diets. The four major anaerobic gut fungal genera (Anaeromyces, Neocallimastix, Piromyces and Orpinomyces) were all found associated with both grass and white clover boli (see Appendix - Figure 4.1.3). Variations in predominant isolates appeared to be host-specific rather than diet-specific, athough there is not yet enough data to be absolutely certain. Analysis of bolus samples revealed very early colonisation by anaerobic fungi. Analysis of bolus samples revealed very early colonisation by anaerobic fungi, with detection at low levels on plant material that had been chewed, swallowed and recovered before contact with rumen digesta (Appendix - Figure 4.1.3; t0
bolus samples). For grass boli, the subsequent growth of anaerobic fungi was immediate and colonised isolates were well established after 1 h of incubation and increased up to 24 h. For white clover boli, colonisation was as rapid as for grass but subsequent growth was slower, displaying a lag (~8 h), before more extensive growth proceeded. Anaerobic gut fungal populations were maximal between 24 and 72 h on white clover boli.
In conclusion, these studies indicate that a combination of microbial marker and molecular techniques provide useful tools for gaining a more precise understanding of the initial events occurring during colonisation and degradation of fresh forage, in particular feed boli. Further development is required, particularly in relation to the choice of suitable bacterial primers but the techniques show considerable promise for examining the response of the rumen microbial population to changes in herbage type, particularly species bred for characteristics that may enhance nutrient use efficiency in the rumen or to improve food quality and/or safety.
4.2 Degradation of protein from freshly ingested herbage (boli) in the rumen of dairy cows
4.2.1 Introduction Protein breakdown in the rumen is generally considered to be a microbially mediated process in which microbial proteases cleave plant protein into the peptides and amino acids required for microbial growth and protein synthesis. However, previous research at IGER, Aberystwyth (Zhu et al., 1999) suggested that intrinsic plant proteases contribute to the initial stages of proteolysis of freshly ingested herbages. We have conducted numerous studies in vitro that support our plant protease hypothesis (see LS0303). In this work we used the method developed for retrieving feed boli (partially masticated herbage) from the rumen of cows grazing fresh herbages (Sections 4.1 and 4.2) to examine the pattern of protein breakdown from freshly-ingested herbages in the rumen.
4.2.2 Materials and methods Holstein-Friesian cows (2 per treatment) fitted with rumen cannulae were fed either a basal diet of perennial ryegrass or white clover ad libitum. Fresh feed boli were harvested as described in Section 4.1.2, immediately after feeding of a handful of either fresh grass or white clover cut on the morning of the experiment. Approximately 3 kg of rumen boli were collected from each animal and sub-samples (150 g) immediately weighed into dacron bags and incubated in duplicate in the rumen for 0, 1, 2, 4, 8 and 24 h. After removal from the rumen, the bags containing the residues were rinsed in cold water, freeze-dried and weighed to estimate dry matter disappearance. Proteins in the dacron bag residues were extracted using the method described by Mae et al. (1993). The polypeptides in the extracts were separated by SDS-PAGE (Laemmli, 1970). Gels were loaded on an equal protein basis (10 µg per track) after determination of the protein contents of the extracts by the method of Bradford (1976). For Western Blotting analysis, the proteins were separated by electrophoresis and then electrotransferred to nitrocellulose membranes and cross-reacted with heterologous antisera prepared against the large and small subunits of ribulose 1,5 bisphosphatase carboxylase/oxygenase (Rubisco; Beha et al., 2002).
CSG 15 (1/00) 23

4.2.3 Results and discussion The disappearances of plant dry matter (a) and protein (b) during incubation in dacron bags were compared using the retrieved boli from the two contrasting herbages at each incubation time (see Appendix - Figure 4.4.1). Protein values were corrected for dry matter disappearance during incubation and presented in terms of the initial dry matter placed in the bag. [N.B. the hypothesis was that if there was no proteolysis, protein content per g DM placed into bags would remain constant over the incubation time.] Dry matter loss was in general higher from the white clover boli than for grass (see Appendix - Figure 4.4.1a) and this could be in part associated with protein degradation (see Appendix - Figure 4.4.1b) in the herbage in the bag.
Analysis of polypeptides extracted from the residues and separated by SDS-PAGE showed that protein degradation occurred in grass and white clover boli. The most abundant proteins at zero time were the large subunit (LSU) of Rubisco (Appendix - Figure 4.4.2, arrow A) and the light harvesting protein (LHP, Appendix - Figure 4.4.2, arrow C). A proportion of the Rubisco LSU remained until 2 h of incubation, while the polypeptide corresponding to LHP appeared to be relatively stable for up to 24 h of incubation in the rumen. This supports the view that Rubisco is susceptible to plant protease attack /microbial degradation in the rumen, whereas LHP is relatively undegradable.
Further analysis of Rubisco by Western blotting showed a decline in the amounts of Rubisco with time (Appendix - Figure 4.4.3), the majority disappearing within 24 h of incubation. The small subunit (SSU) of Rubisco appeared to be more slowly degraded and was still present after 8 h. Breakdown products of Rubisco accumulated after 4 h of incubation of grass (Appendix - Figure 4.4.3a), but not white clover (Appendix - Figure 4.4.3b), suggesting that some differences in the mechanism could exist between plant species. These differences could alter patterns of succession by the rumen microbial population (see Section 4.1) but more molecular microbial data is needed for any conclusions to drawn. The LHP polypeptide was much more stable than the LSU of Rubisco. This is possibly because of protection afforded by the intra-membrane localisation of LHP. The reasons behind stability of SSU are unclear but could be related to this polypeptide being nuclear encoded, whereas LSU is chloroplast encoded. This could provide a breeding trait for future exploitation and needs further investigation.
4.3 In vivo evidence of plant mediated proteolysis in the rumen of dairy cows
4.3.1 Introduction Extensive proteolysis has previously been observed as a result of in vitro incubations of grass or legumes in the presence or absence of rumen micro-organisms (Zhu et al., 1999; Kingston-Smith & Theodorou, 2000), indicating the potential activity of plant proteases under rumen-like conditions. The aim of the present study was to provide supportive in vivo evidence of plant mediated activity in experiments where ryegrass was incubated in the rumens of dairy cows and to assess the degree to which ruminal proteolysis was under the influence of plant and/or microbial proteases.
4.3.2 Materials and methods Freshly harvested grass was chopped into 1-2 cm lengths. Aliquots of 3 g fresh weight (FW) were placed directly in Dacron bags (10M pore size; Sericol, Industrial Fabrics Division, Pycons Road, Broadstairs, Kent) or suspended in 20 ml artificial saliva (McDougal, 1948) and placed in either polythene and heat sealed or dialysis tubing (10,000 Da cut-off; Spectrum, USA) which were then placed in Dacron bags for support during incubation in the rumen. As a negative control, dialysis tubing (also held in a Dacron bag) was filled with 20 ml artificial saliva but no grass. These approaches are subsequently referred to on a collective basis as in sacco. Two Dacron bags for each treatment (Db, dacron bag; Pb, polythene bag; Dt, dialysis tubing) incubated in the rumen of each of 4 dry fistulated Holstein cows grazing a grass sward. After 16 h incubation the bags were removed and the herbage residues recovered by straining through 3 layers of muslin. Herbage was placed in sintered funnels and rinsed with 100 ml distilled water. Aliquots of herbage residues were used to determine dry matter disappearance, or stored at -80 C until analysis of protein content. Volatile fatty acids (VFA), were performed on rumen fluid samples taken before and after incubation of herbage, and on liquid recovered from polythene and dialysis bags. These liquid fractions were also used to quantify protease activity using a gel diffusion assay. Zones of clearing were calibrated by comparison with reference discs of standard area. Protein was extracted from herbage residues and the protein content of extracts was determined by a modified Lowry reaction and the polypeptides contained in aliquots separated by denaturing gel electrophoresis. Gels were stained with Coomassie Brilliant Blue, photographed and the bands quantified by densitometry. As appropriate ANOVA (Excel or Genstat) or a two tailed Student’s t-test (Bailey 1976) was used to evaluate the significance of differences between treatment results.
4.3.3 Results Incubation of grass in Dacron bags simulated normal rumen conditions, with the rumen micro-organisms having unrestricted access to the herbage via the 10 M pores. Incubation of grass in a polythene bag in artificial saliva provided a normal rumen temperature, but excluded rumen micro-organisms, microbial extra-cellular proteases and all other components of the rumen fluid. Grass enclosed in bag made of dialysis tubing allowed entry of small molecules in rumen fluid (ammonia, salts, short chain fatty acids, etc.) but rumen micro-organisms and all but the very small
CSG 15 (1/00) 24

extracellular enzymes (<10kDa) were excluded due to the molecular weight cut-off of the tubing (10,000 Da). Measurements were made of some indicators of microbial and plant activity in rumen fluid and compared with corresponding values in fluid obtained from the polythene bag and dialysis tubing treatments. Some VFAs accumulated in the polythene bag fluid (total VFA concentration of 32 mM), but this was considerably lower than in rumen fluid collected before incubation started and 85% was acetate, so was probably derived from plant rather than microbial activity. TVFA concentrations in the dialysis tubing sacs containing either grass plus buffer (111.0 mM), or buffer alone (negative control; 115.0 mM) were similar to that in rumen fluid collected before incubation (114 mM) This finding and similar corresponding values for ammonia gave confidence that dialysis tubing allowed a 'normal' rumen environment in the absence of rumen micro-organisms. Measurements of protease clearing activity showed that both rumen fluid and polythene bag fluid contained significant activities (60 and 29 mm2 clearing zones, respectively), while a lesser but measurable amount was present in the fluid from treatment bags made of dialysis tubing (8 mm2). Protease activity was not recovered from dialysis tubing sacs that did not contain plant biomass, demonstrating that the tubing was effective in excluding rumen fluid proteases. Dry matter (DM) disappearance from herbage was observed in all treatments, but this was significantly (P < 0.05) greater in herbage retained by Dacron bags (46.3%) compared to polythene bags (36.2%) or bags made of dialysis tubing (38.1%). As microbial enzymes could not penetrate dialysis tubing or polythene, all DM loss related to solubilisation of plant constituents, i.e. about 35% of DM loss, did not involve micro-organisms and was due to entirely physical or plant-based enzymatic activities. Gel electrophoresis revealed compositional changes in the herbage protein pool as a result of incubation in the rumen (Figure 4.3.1). There were marked changes in the polypeptide profile in all herbages after 16 h incubation in the rumen as compared with the polypeptides extracted from freshly cut grass (Figure 1). In contrast, there was little difference in the polypeptide profile of residual protein replicates recovered from bags after 16h, irrespective of treatment. Quantification by densitometry showed that the large subunit of the most abundant soluble protein, Rubisco (ribulose 1,5 bisphosphate carboxylase/oxygenase) was degraded to the same extent regardless of treatment (Figure 2). In contrast, the Rubisco small subunit and the membrane-associated protein LHCPII (light-harvesting complex proteins of photosystem 2) were relatively undegraded in all treatments (Figure 2).
4.3.4 Discussion We have described a simple exclusion system which was devised in order to demonstrate if proteases of plant origin were active in the degradation of plant proteins in the rumen environment under in vivo conditions. The 3 treatments described allowed complete access of micro-organisms to plant material (Dacron), exclusion of rumen micro-organisms (polythene) or allowed interaction of fermentation products with plant material while preventing ingress of rumen fluid proteases (dialysis tubing). Significant dry matter loss was observed and substantial changes in polypeptide profile of residues compared with that from untreated grass indicated significant protein breakdown in all treatments. The similar profile in the Dacron bag treatment compared with the dialysis tubing treatment suggests that plant proteases were active in the early stages of digestion and provides in vivo data in support of our findings under in vitro conditions.
4.4 Bolus biology and digestion under in vitro conditions
4.4.1 Introduction Boli formed during ingestion of herbage are truly representative of material that enters the rumen, unlike either freshly harvested and unprocessed grass, or ground and dried grass, the substrate that is generally used in both in vitro and in sacco Dacron bag digestibility studies. Patterns of nutrient supply with these substrates is likely to be markedly different, as suggested by the results of experiments where the fermentation rates of freeze-dried and ground silages were compared (Davies et al., 1999; Lowman et al., 2002). The objective of this experiment was to examine how the effects of herbage 'processing' during bolus formation influences the release of nutrients and rate of digestion of fresh herbage and compare this with corresponding fresh or ground and dried material as a first step towards developing an in vitro model for studying the affect of tissue damage on nutrient supply to the microbial population.
4.4.2 Materials and methods Perennial ryegrass was harvested at 8.30 am, 2 days prior to carrying out the experiment described below and immediately frozen and freeze-dried and ground to pass through a 2 mm sieve. Two days later grass was harvested from the same plot at the same time and used to feed to a Friesian/Holstein dairy cow equipped with a rumen fistula This cow was used as the donor animal for collection of the initial down boli as described in Section 4.1.2. In addition, a portion of the same grass was cut by hand into 2 cm lengths using scissors. The collected boli were well mixed and used as substrates in an in vitro fermentation and digestibility experiment in comparison with the original fresh, chopped grass and corresponding freeze-dried and ground grass. Triplicate samples (1 g of DM) of each of the grass treatments was mixed with anaerobic buffer, inoculated with rumen contents and incubated at 39 oC for up to 72 h using the approach described for the gas production procedure of Theodorou et al. (1994). Cumulative gas production was monitored throughout the incubation period and three replicate batch cultures were destructively sampled after 0, 2, 4, 6, 8, 12, 20, 28 and 70 h of incubation and the residue prepared by filtration through a sintered crucible. These samples were analysed for dry matter loss and residual water soluble carbohydrate (WSC) concentration. VFA concentrations were measured in the supernatants from the 72 h sampling time.
CSG 15 (1/00) 25

4.4.3 Results and Discussion The amount of DM remaining in the residues of the grass treatments after different times of incubation are shown in Figure 4.3.1. Values at zero time ranged from 56% with freeze-dried grass to 86% with fresh grass with an intermediate value for the bolus. As the amount of grass DM added for all treatments was similar, these differences reflect the relative ease of movement of soluble components of the DM into the liquid phase, which occurred immediately after immersion in the buffer/rumen fluid mixture. After this time freeze dried, ground material was degraded most rapidly, closely followed by the bolus material, with the fresh cut grass having ca 20% more dry matter remaining at most time points. The final DM digestibility values were 80, 83 and 70%, respectively, for bolus, freeze-dried ground and fresh grass. The cumulative gas production profiles (not shown) reflected the pattern of DM disappearance shown in Figure 4.3.1, but the differences between the three treatments were less pronounced. The final VFA concentrations in the supernatants were 135, 146 and 147 mM, respectively, for bolus, freeze-dried ground and fresh grass. No differences were observed in this experiment in terms of VFA molar proportions (viz. 54.6:18.7:16.1:10.6; acetate:propionate:butyrate:valerate). Immediately prior to immersion of the substrates into the anaerobic buffer the mean WSC concentration of the bolus, fresh grass and freeze dried ground grass was 211 3.1 g/kg DM. The rates of WSC disappearance from the grass treated in these 3 different ways are shown in Figure 4.3.2. As with the DM values for the zero timepoint, there were large differences in WSC concentration in the 3 residues at this time. This reflected the loss of WSC immediately after immersion due to relative degrees of damage, with less than 25% remaining for freeze-dried grass but 75% for the bolus and nearly 100% for chopped grass. Thereafter there was a very rapid rate of loss of WSC from both the freeze-dried and bolus material, but considerably slower from fresh grass. By 6 h the freeze-dried and bolus material had similar WSC contents (between 30 and 40 g/kg DM) whereas the fresh grass still had a concentration in excess of 130 g/kg DM. These initial results demonstrate how rapidly substrate becomes available from dried and ground herbage, compared to the bolus and the other extreme, chopped grass, where only cut ends are exposed. These findings support the concept of using the bolus to study microbial colonisation, as tissue damage occurring during ingestion clearly influences nutrient availability in the early stages of digestion. Further research is required to confirm these findings and elucidate whether other mechanical treatments can simulate the precesses involved in bolus formation in order to develop an in vitro model for studying comparative rates of digestion of fresh forages by plant- and microbially-mediated processes.
4.5 General conclusions for bolus biology and microbial ecology studies In conclusion, the approaches described above, will form the basis of a 'toolkit' for studies of the early stages of digestion of fresh forages, leading to a more precise understanding of the initial events during colonisation and degradation of fresh forage and, in particular, feed boli.
5.0 Publications and technology transfer The attached Appendix gives details specific to publications and technology transfer for LS0302. The overall communication of project LS03 (01, 02 and 03) was delivered to the industry by means of 3 main systems. Direct technology transfer by IGER mechanisms, communication by means of the DEFRA Livestock Knowledge Transfer and, thirdly, by preparation of technical factsheets and oral presentations on behalf of the industry Levy Boards. Details of these can be seen in the Report for LS0301, Appendices 1-4.
Future research Future research areas that address DEFRA policy objectives on improving efficiency of N use/unit of animal product
and reducing N losses to the environment in sustainable agricultural systems include: Optimising rumen function and digestion to reduce the problem of low N-use efficiency encountered with legume-based diets. For example the (i) use of low-protein concentrates and/or readily available energy supplies (high sugar grasses, whole crop cereals) and (ii) the kinetics of physical and chemical breakdown of forages in the rumen, both of which have important effects on feed intake and digestion.
Plant traits that present forage breeding targets and plant based strategies for reducing protein degradation in the rumen e.g., polyphenol oxidase and/or plant proteases activities, plant light harvesting protein.
Bolus biology and associated molecular techniques to determine how plant and microbial activities influence microbial colonisation of fresh forages as a means of manipulating the microbial population and efficiency of nutrient utilisation via the plant based strategies.
ReferencesAkyol, I. , Fraser, M. D. , Griffith, G. W. & Davies, D. R. (1999). Comparison of in vivo and in vitro digestibility of silages made from a range of alternative crops Proceedings of the British Society of Animal Science (BSAS) Annual Meeting, Scarborough, UK, 22-24 March 1999 97 BSAS, Penicuik, UK.
Auldist, D.E., Atkinson, K.L., Silvapulle, M.J., Dellow, D.W. & McDowell, G.H. (1999). Utilisation of white clover silage fed alone or with maize silage by lactating dairy cows. Australian Journal of Experimental Agriculture, 39, 237-246.
CSG 15 (1/00) 26
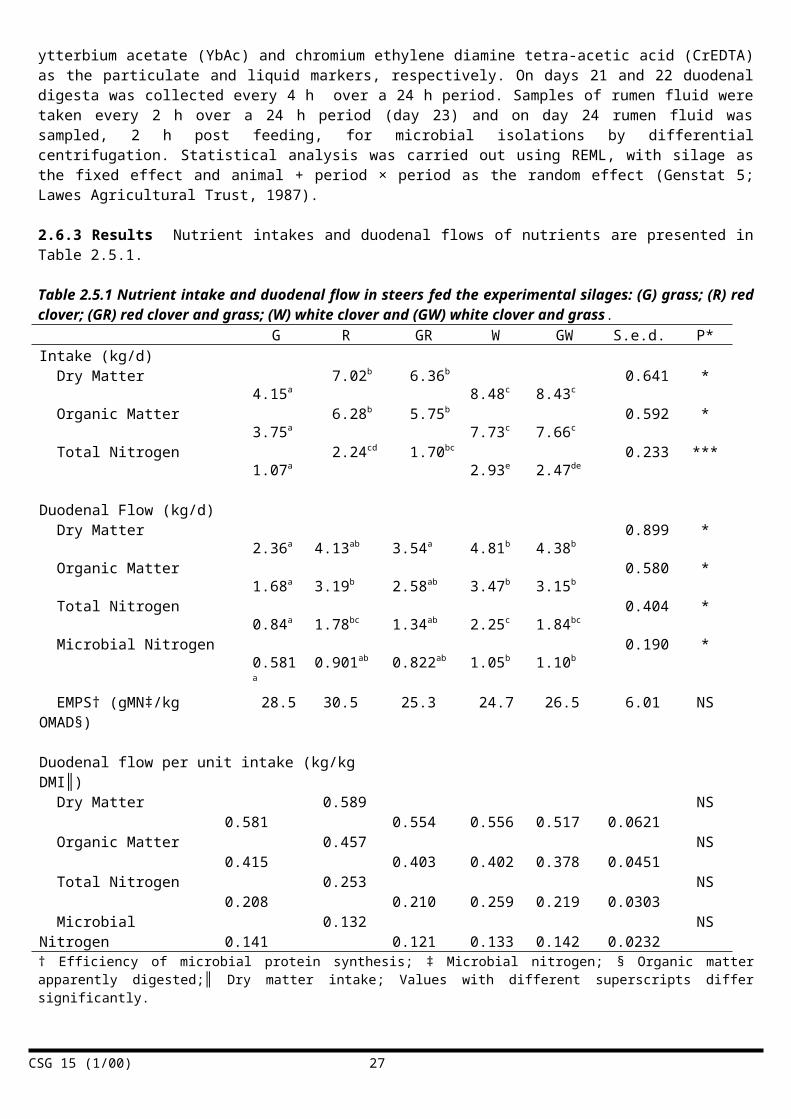
Beha, E. M., Theodorou, M. K., Thomas, B. J. & Kingston-Smith, A. H. (2002) Grass cells ingested by ruminants undergo autolysis which differs from senescence: implications for grass breeding targets and livestock production Plant Cell & Environment 25, 1299-1312
Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Analytical Biochemistry 72, 248-254.
Castle, M.E., Reid, D. & Watson, J.N. 1984. Silage and milk production: a comparison between supplements of barley and soybean meal offered with white clover silage. Grass and Forage Science, 39, 287-289.Cussen R.F., Merry, R.J., Williams, A.P. & Tweed, J.K.S. (1995) The effect of additives on the ensilage of forage of differing perennial ryegrass and white clover content. Grass and Forage Science 50, 249 – 258.Czerkawski, J & Breckenridge, G (1977). Design and development of a long-term rumen simulation technique (Rusitec). British Journal of Nutrition 38, 371 - 385.
Davies, Z.S., Brooks, A.E., Theodorou, M.K., Griffith, G.W. & Merry, R.J. (1999). Effect of inoculant and enzyme additives on fermentation characteristics and gas production of grass silage. Proceedings of the British Society of Animal Science Annual Meeting, Scarborough, March 22-24, 1999, pp. 145. BSAS, Penicuik, Scotland.
Davies D.R.,. Leemans D.K & Merry R.J. (2002) Improving silage quality by ensiling perennial ryegrasses high in water soluble carbohydrate content, either with or without different additives. Proceeding of the XIIIth Silage Conference, Ayr, pp. 386-387. SAC, Ayr.
Dewhurst, R. J., Davies, D. W. R., Fisher, W. J., Bertilsson, J. & Wilkins, R. J. (2000). Intake and milk production responses to legume silages offered to Holstein-Friesian cows. Proceedings of the British Society of Animal Science p.11.
Dewhurst, R. J., Merry, R. J., Bertilsson, J.& Tuori, M. (2001): Effects of clover silages on diet digestibility and rumen function. Landbauforschung Voelkenrode 234, 47-53.
Genstat (1987) Genstat 5. Lawes Agricultural Trust: Clarendon Press.
Genstat Release 6.1 Reference Manual (2002). Procedures. pp. 372-376. Oxford:VSN International. Goodridge, J., Ingalls, J.R. & Crow, G.H. (2001) Transfer of omega-3 linolenic acid and linoleic acid to milk fat from flaxseed or Linola protected with formaldehyde. Canadian Journal of Animal Science, 81, 525-532.
Gordon, F. J., Cooper, K. M., Park, R. S. & Steen, R. W. J. (1998). The prediction of intake potential and organic matter digestibility of grass silages near infrared spectroscopy analysis of undried samples. Animal Feed Science and Technology 70, 339-351.
Humphreys, M.O. (1989). Water soluble carbohydrates in perennial ryegrass breeding. II. Cultivar and hybrid progeny performance in cut plots. Grass and Forage Science, 44, 237-244.
Jones & Mosely,1977. Laboratory Practice, 26, 687-689.
Kingston-Smith, A.H. and Theodorou M.K. (2000). Post-ingestion metabolism of fresh forage. Tansley Review No.19. New Phytologist, 148, 37-55.
Lee, S. S., Chang, M. B., Scollan, N. D., Merry, R. J., Dhanoa, M. S., Hobbs, P. J., Theobald, V. J., Maeng, W. J. and Dewhurst, R. J. (1999). The fatty acid composition of solid- and liquid-associated rumen bacteria isolated from cows. Proceedings of the British Society of Animal Science, p.30.
Lowman, R.S., Theodorou, M.K. and Cuddeford, D (2002) The effect of sample processing on gas production profiles otained using the pressure transducer technique. Animal Feed Science and Technology, 97, 221-237.
Mae, T., Thomas, H., Gay, A.P., Makino, A. and Hidema, J. (1993). Leaf development in Lolium temulentum: photosynthesis and photosynthetic proteins in leaves senescing under different irradiances. Plant Cell Physiology 34, 391-399. Muyzer, G., de Waal, E. & Uitterlinden, A. (1993). Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain-reaction-amplified genes coding for 16S rRNA. Applied and Environmental Microbiology59, 695-700.
Sukhija, P.S. & Palmquist, D.L. (1988) Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. Journal of Agricultural and Food Chemistry, 36, 1202-1206.
Theodorou, M. K. , Gill, M. , King-Spooner, C. & Beever, D. E. (1990). Enumeration of anaerobic Chytridiomycetes as thallus-forming units: novel method for quantification of fibrolytic fungus populations from the digestive tract ecosystem Applied Environmental Microbiology, 56, 1073-1078.
Theodorou M.K. Williams, B.A., Dhanoa, M.S., McAllan, A.B. & France., J. (1994). A simple gas production method using a pressure transducer system to determine the fermentation kinetics of ruminant feeds. Animal Feed Science and Technology, 48, 185-197.
Thomas, C., Aston, K. & Daley, S. R. 1985. Milk production from silage. 3. A comparison of red clover with grass silage. Animal Production 41, 23-31.
CSG 15 (1/00) 27

Thomas. C., Aston, K., Gibbs, B.G. & Tayler (1981). Beef production from silage. 1. The volunrtary intake and liveweight gain of beef cattle given red clover silage. Animal Production, 32, 143-148.Thomas, C. & Thomas, P.C. (1985). Factors affecting the nutritive value of grass silages. In: Recent advances inanimal nutrition. Eds. W. Haresign and D.J.A. Cole, Butterworths. pp. 223-256.Thornton, R. F. & Minson, D. J. (1972). The relationship between voluntary intake and mean apparent retention time in the rumen. Australian Journal of Agricultural Research 23, 871-877.
Waghorn, G. C., Shelton, I. D. & Thomas, V. J. 1989. Particle breakdown and rumen digestion of fresh ryegrass ( Lolium perenne L.) and lucerne (Medicago sativa L.) fed to cows during a restricted feeding period. British Journal of Nutrition 61, 409-423.
Zhu, W.-Y., Kingston-Smith, A.H., Troncoso, D., Merry, R.J., Davies, D.R., Pichard, G., Thomas, H & Theodorou, M.K. (1999). Evidence of a role for plant proteases in the degradation of herbages proteins in the rumen of grazing cattle. Journal of Dairy Science 82, 2651-2658.
CSG 15 (1/00) 28