research article changes in otx2 and parvalbumin...
TRANSCRIPT

Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 848265 17 pageshttpdxdoiorg1011552013848265
Research ArticleChanges in Otx2 and Parvalbumin Immunoreactivity inthe Superior Colliculus in the Platelet-Derived Growth FactorReceptor-120573 Knockout Mice
Juanjuan Zhao12 Susumu Urakawa3 Jumpei Matsumoto1 Ruixi Li12 Yoko Ishii4
Masakiyo Sasahara4 Yuwen Peng2 Taketoshi Ono3 and Hisao Nishijo1
1 SystemEmotional Science Graduate School ofMedicine and Pharmaceutical Sciences University of Toyama Toyama 930-0194 Japan2Department of Anatomy Histology and Embryology Shanghai Medical School Fudan University Shanghai 200032 China3Department of Judo Physiotherapy Graduate School of Medicine and Pharmaceutical Sciences University of ToyamaToyama 930-0194 Japan
4Department of Pathology Graduate School of Medicine and Pharmaceutical Sciences University of Toyama Toyama 930-0194 Japan
Correspondence should be addressed to Hisao Nishijo nishijomedu-toyamaacjp
Received 17 July 2013 Accepted 30 September 2013
Academic Editor Carlos Tomaz
Copyright copy 2013 Juanjuan Zhao et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The superior colliculus (SC) a relay nucleus in the subcortical visual pathways is implicated in socioemotional behaviorsHomeoprotein Otx2 and 120573 subunit of receptors of platelet-derived growth factor (PDGFR-120573) have been suggested to play animportant role in development of the visual system and development andmaturation of GABAergic neurons Although PDGFR-120573-knockout (KO) mice displayed socio-emotional deficits associated with parvalbumin (PV-)immunoreactive (IR) neurons theiranatomical bases in the SC were unknown In the present study Otx2 and PV-immunolabeling in the adult mouse SC wereinvestigated in the PDGFR-120573KOmice Although there were no differences in distribution patterns of Otx2 and PV-IR cells betweenthe wild type and PDGFR-120573 KO mice the mean numbers of both of the Otx2- and PV-IR cells were significantly reduced in thePDGFR-120573KOmice Furthermore average diameters of Otx2- and PV-IR cells were significantly reduced in the PDGFR-120573KOmiceThese findings suggest that PDGFR-120573 plays a critical role in the functional development of the SC through its effects on Otx2- andPV-IR cells provided specific roles of Otx2 protein and PV-IR cells in the development of SC neurons and visual informationprocessing respectively
1 Introduction
The mammalian superior colliculus (SC) is a seven-layeredstructure on the roof of the midbrain [1] and can be dividedinto two functionally distinct units a superficial subdivisionand a deep subdivision [2 3] The superficial layers zonale(ZO) superficial gray (SGR) and optic (OP) layers receivetheir major inputs from the retina and the visual cortex andare involved exclusively in visual information processingIn contrast the four deep layers intermediate gray (IGR)intermediate white (IW) deep gray (DGR) and deep white(DW) layers which receive auditory somatic and visualinputs from numerous cortical and subcortical areas areinvolved in the control of orientating behaviors of head eye
and ear [1 4ndash6] Furthermore recent studies suggest thatthe subcortical visual pathway including the retina SC andpulvinar is implicated in socioemotional behaviors [7 8]
Several studies have used parvalbumin (PV) asmarkers toreveal details of the parallel subcortical pathways involved invisual processing [9] In the primate lateral geniculate nucleus(LGN) PV is found in the relay cells in the P andM pathways[9ndash11] In the cat SC Mize et al [12] found a band of PV-immunoreactive (IR) cells (PVcells) in the deep SGR andupper OP a region of the cat SC that receives inputs from Yretinal ganglion cells [13] In addition PV is also one of thebiochemical markers of the matrix surrounding some of theIGR compartments in the SC [14] Other studies have asso-ciated PV with specific firing patterns of neurons which are
2 BioMed Research International
often found in ldquofast spikingrdquo neurons with nonadapting trainsof action potentials It has been suggested that PVmight regu-late a Ca2+-activated K+ channel involved in spike adaptation[15] and a particular type of K+ channel was developmentallyregulated with PV and was expressed in about 80 of the PV-containing GABAergic interneurons in the rat hippocampus[16] Furthermore deficits in these PV-containing GABAer-gic interneurons were associated with schizophrenia andautism with social deficits [17 18] and the number of PV-positive neurons was reduced in the SC of an animal modelof autism with prenatal valproic acid exposure [19]
The Otx2 homeobox gene is a vertebrate orthologue ofthe Drosophila orthodenticle gene [20ndash22] members of thisorthology group play a fundamental role in developmentof photoreceptors Otx2 and Crx (cone-rod homeobox) androstral brain regions (Otx1 and Otx2) [23 24] Otx2 alsoappears to play a role in development and function of theretina in which the gene is expressed at both prenatal andpostnatal stages [25ndash29] In addition Otx2 coordinates post-natal PVcell maturation and activates visual cortical plasticity[30] Otx2 is strikingly restricted to relay centers in theprimary visual pathway at birth before PV-cells are evident[31] including the retina LGN and visual cortex (V1)Furthermore exogenous Otx2 infusion accelerated PV-celldevelopment in the V1 area [30] These findings indicate thatOtx2 is related to development of the visual system Howeverto our knowledge no information is available concerningOtx2 expression in the retinocollicular (retinotectal) visualpathway in adult mice It is possible that Otx2 may controlfunctional features of SC neurons in adult mice
Platelet-derived growth factor (PDGF) was originallyreported as a substance in platelets that promote growthof tissue culture cells [32] However the factor is recentlyreported to act as a neuroprotective factor in the centralnervous system (CNS) [33ndash36] PDGF is also involved inthe regulation of cell growth and differentiation duringembryonal development [37ndash39] The family of platelet-derived growth factors (PDGFs) comprises 4 membersmdashPDGF-A -B -C and -Dmdashthat are assembled from disulfide-linked homo- or heterodimers of 2 distinct but related chains(PDGF-AA -AB -BB -CC and -DD) Two receptor subtypesof PDGF (PDGFR-120572 and -120573) can form mature dimericreceptor complexes that can bind to ligands with differ-ent affinities [40] PDGFR-120572 is largely expressed in oligo-dendroglial progenitors while PDGFR-120573 is predominantlyexpressed in neurons [41] and upregulated in the neonatalrat brain [42] PDGF-BB that specifically binds to PDGFR-120573120573 is abundantly expressed in neurons and is upregulatedin neonatal brains [33 43 44] PDGF-B exerts neurotrophiceffects on 120574-aminobutyric acid GABA ergic neurons [42 45]Altering PDGFR-120573 may result in abnormalities during thedevelopment of the CNS [46 47] Consistently conditionalknockout (KO) mice with suppressed expression of neuronalPDGFR-120573 in the CreloxP system (PDGFR-120573 KO mice)displayed autisticschizophrenic traits especially deficits insocioemotional behaviors [48]
Taken together there is a possible link among PV Otx2and PDGFR-120573 and behavioral deficits in the PDGFR-120573
KO mice might be ascribed to changes in PV and Otx2in the SC In the present study to investigate the role ofPDGFR-120573 in Otx2 and PV expression in the SC we analyzedimmunohistochemical alterations in Otx2 and PV expressionin the SC of PDGFR-120573 KOmice
2 Materials and Methods
21 Generation of the Conditional PDGFR-120573 Knockout MiceThe CreloxP system was used to develop conditionalPDGFR-120573 KO mutants A previously established mutantmouse line was used in which exons 4ndash7 of PDGFR-120573 whichencode the extracellular domain of the PDGFR-120573 proteinwere flanked by 2 loxP sequences (floxed) positioned inintrons 3 and 7 [49] After Cre-mediated recombinationdeletion of the loxP-flanking region and resulting frame shiftmutation in the adjoining 31015840 region occurred in PDGFR-120573 To obtain conditional PDGFR-120573 KO we then crossedmutant mice harboring the PDGFR-120573 floxed allele and thoseexpressing Cre recombinase under the control of the nestinpromoter and enhancer (nestin-Cre+ mouse The JacksonLaboratory Bar Harbor ME USA) as previously described[50 51] Before this cross both mutant mice harboringfloxed PDGFR-120573 and nestin-Cre+ were outbred to the miceof C57BL6J (B6J) strain for 14 generations to replace thegenetic background of our mutant mice with that of the B6Jstrain In the present study the following 2 types of 22 to 29weeks old male mice were used mice with the Cre transgeneand floxed PDGFR-120573 (PDGFR-120573KOmice) andmice withoutthe Cre transgene but with floxed PDGFR-120573 (control mice)
Genotypes were confirmed by PCR of tail DNA usingoligonucleotide primers pairs for floxedPDGFR-120573 and for theCre transgene as described previously [50] The genotypingwas confirmed by Western blot of the total lysates of theadult mouse brains to show that the PDGFR-120573 expressiondecreased to undetectable levels in the PDGFR-120573 KO micecompared with that in the control mice [50]
In the present study 6 control mice (24ndash28 weeks old 22ndash29 g body weight) and 4 PDGFR-120573 KO mice (24ndash28 weeksold 22ndash29 g body weight) were used All mice were housedin individual cages in a temperature-controlled environmentwith a 1212 h lightdark cycle (lights were turned on and offat 0800 and 2000 resp) Food and water were supplied adlibitumMice (10 to 16weeks old) were handled for 3 consecu-tive days before the start of the experiments All experimentalprotocols were performed in accordance with the guidelinesfor Care and Use of Laboratory Animals approved by theUniversity of Toyama and the National Institutes of HealthrsquosGuide for the Care and Use of Laboratory Animals andapproved by the Committee for Animal Experiments at theUniversity of Toyama
22 Perfusion and Tissue Processing Under deep anesthesiawith sodium pentobarbital (50mgkg body weight ip) themice were transcardially perfused with heparinized saline(09 wv NaCl) followed by 4 paraformaldehyde dis-solved in 01Mphosphate buffer (PB) pH74After perfusionthe brains were removed from the skull cut coronally into
BioMed Research International 3
small blocks and postfixed in 4 paraformaldehyde forovernight The fixed brain blocks were immersed into 30sucrose in 01M PB until they sank down on the bottomThebrain blocks were freezed in dry ice and coronally cut intosections at the thickness of 40120583mThe sectionswere collectedin 001MPBS and then transferred into a cryoprotectant solu-tion (25 ethylene glycol 25 glycerin and 50 01M PBS)and stored at minus20∘C until immunohistochemical staining
23 Immunohistochemistry of the Sections Three serial sec-tions were collected every 120 120583m two for PV and Otx2-immunocytochemistry and one for Nissl staining with cresylviolet The sections were washed thrice in 001M phosphatebuffer saline (PBS) for 15min blocked with 3 normalhorse serum 025 Triton X-100 in PBS for 30min at roomtemperature and incubated overnight at 4∘C with mousemonoclonal anti-PV (1 10000 dilution Sigma St LouisMO USA) or goat polyclonal anti-Otx2 (1 200 dilutionRampD) antibodies in 1 blocking solution 025 Triton X-100 in PBS These sections were washed thrice with 001MPBS for 5min each time incubated with biotinylated horseanti-mouse IgG or anti-goat IgG (1 250 dilution VectorBurlingame USA) for 1 hr at room temperature and thenafter washing incubated in ABC reagent (Vector) for 50minFinally the PV- and Otx2-immunoreactive elements werevisualized by reacting with 20mg 331015840-diaminobenzidine and20120583L 30 H
2O2in 100mL 005M PBS (pH 76) for 5ndash
8min The sections were then rinsed several times in PBSdehydrated in graded concentrations of ethanol cleared inxylene and cover-slipped with Entellan (Merck DarmstadtGermany) To minimize variability in staining the sectionsfrom the all treatment groups were run in the same immuno-cytochemistry session Negative control sections were treatedidentically except for omission of the primary antibody Noreaction product was observed in any of the control sections
The corresponding adjacent sections were stained withcresyl violet to identify colliculus layers The colliculus layerswere identified by staining and cellular densities and mor-phologies of SC neurons [1] We also identified collicular lay-ers using parvalbumin-labeled sections where the collicularlayers of these proteins have been well described previously[12 52 53]
24 Identification of the SC and Image Capture In eachsection the SC was identified and subdivided into collicularlayers The nomenclature of the mouse atlas of Patrick andWarren (2000) was adopted Labeling was examined usingstandard bright-field microscope (BX 61 Olympus TokyoJapan) with different magnifications (4sim100x) A series ofimages of the SC were captured by a digital camera (DP 70Olympus Tokyo)
25 Data Analysis In the present study 6 coronal sections(minus330 minus362 minus394 minus426 minus458 and minus490mm from thebregma) in each mouse were analyzed for quantification ofOtx2- and PV-IR cells These digital images were analyzedusing the ImageJ software (NIH ImageJ httprsbwebnihgovij) The mean number of cells per section in each layer
was estimated based upon two approache the measurementof pixel intensity and counting of immunopositive cellsFor the pixel intensity of immunopositive cells all imagesused for cell quantification were saved as uncompressed 8-bit grey-scale tiff files and then were thresholded using theautothreshold tool in ImageJ with visual comparison beingmade to the original grey-scale images to ensure that the tooleffectively resolved all labeled cell somas or nucleiThereforeparticle analysis was carried out using the analyze particletool The mask tool was used to confirm that all particles(labeled cell somas or nuclei) were detected and measured
Estimates of cell number in two-dimensional sections aresubject to error and bias which can be partially overcomeby use of the Abercrombie correction [54 55] To enablesuch a correction in the current study sections were cut atthickness of 40120583m and nuclear diameter of Otx2-IR cellsand soma diameters of PV-IR cells (the average of long andshort axes) were computed by measuring the long and shortaxes of cells counted The correction factor was then appliedto all cell measurements The assessments were carried outby a single investigator (JJ Z) blind to group status In orderto furthermore diminish variability rating of images wasperformed on 10 different occasions by the same blindedobserver Great care was taken to match sections throughthe same region of the brain and at the same level using theanatomical landmarks Layers of the SC were identified usingthe counterstained sections of both the control and KO miceand then outlined using commercial software (Corel Draw12) The collicular layers traced as above were analyzed onthe corresponding collicular areas of immunocytologicallyprocessed control and KO mice
We analyzed the mean number of cells in each layer (6sections from each of the 6 control mice and 4KO mice)the diameter of each cell counted (6 sections from each ofthe 4 control mice and 4KO mice) and categorized cells asvertical and horizontal fusiform pyriformmarginal (round)and multipolar cells according to the previous studies [5657] All statistical comparisons (119905-test and repeated measureANOVA) were performed using SPSS 170 software A valueof 119875 lt 005 was considered to be significant
3 Results
31 Distribution of Otx2- and Parvalbumin (PV-) Immunore-active (IR) Cells in the SC Figure 1 shows the location ofthe SC in the coronal (a) sections of the mouse brain atlasand a section stained by cresyl violet (b) indicating differentsubdivisions of the SC Neuroanatomically the SC has twodivisions a superficial subdivision comprising the zonal(ZO) superficial gray (SGR) and optic (OP) layers and a deepsubdivision comprising the intermediate gray intermediatewhite deep gray and deep white layers The two layers ofwhite matter in the deep subdivision are often rather indis-tinct and do not appear to demarcate cellular distributionsTherefore we simply divided the deep subdivision into tworegions the intermediate gray (IGR) layer and the deep layers(DLs) including the deep white deep gray and intermediatewhite layers
4 BioMed Research International
minus41mm
(a)
ZO
SGR
OPIGR
DL
PAG
minus394mm
(b)
Figure 1 Coronal section in the brain atlas (AP minus41mm from the bregma) (a) and representative section stained with cresyl violet (APminus394mm from the bregma) (b) showing the location and laminar pattern of the mouse SC The square in A indicates the SC region ZOzonale SGR superficial gray layer OP optic IGR intermediate gray layer DL deep layer and PAG periaqueductal gray matter Scale bar =200 120583m
Figure 2 illustrates low power photomicrographs of thecontrol mouse SC sections stained with Otx2 antibody (A)and PV antibody (B) indicating the laminar distributionsof Otx2 and PV-IR cells Figure 3(a) shows black and whitephotomicrographs of the rostral SC sections stained withOtx2 (a) and PV (b) antibodies in the control mice as afunction of the threshold intensity The image data werethresholded to highlight the labeled cell somas or nucleiFigure 4(a) indicates high-magnification photomicrographsof the Otx2-labeled sections in the superficial (A) and deepmain (B) layers of the SC in the control mice The patterns ofOtx2 and PV labeling in the present study were different fromeach other Otx2-IR cells were found in all layers of the SCbut more densely in the superficial layers PV-IR cells werealso observed in all layers of the SC but different from that ofOtx2-IR cells (see below in detail)
The dense distribution of Otx2-IR cells was found in thesuperficial layers (ZO SGR and OP) (Figures 2(a) 3(a)(A)and 4(a)(A)) Otx2-IR cells were also observed in the deeplayers (IGR and DL) (Figures 2(a) 3(a)(A) and 4(a)(B))which mainly included lightly labeled small to medium-sized cells (Figure 4(a)(B)) In contrast very few labeled cellswere observed in the areas surrounding the SC The similardistribution patterns of Otx2-IR cells were observed throughthe rostral-caudal extent of the SC (Figure 2(a))
Figure 5(a) indicates high-magnification photomicro-graphs of the PV-labeled sections in the superficial (A)and deep main layers (B C) of the control mouse SCPV-IR cells were concentrated within the superficial layersof the SC with a heavier frequency within SGR than inthe OP (Figure 5(a)(A) Also see Figures 2(b) and 3(a)(B))Labeled IR elements in the dense tier included not only
lightly stained small-to-medium sized somas but also proxi-mal dendrites and small immunoreactive puncta that couldbe axons axon terminals or small dendrites (open tri-angles) (Figure 5(a)(A)) PV-IR cells were also scatteredthroughout the deep layers below the dense tier In thelateral part of the IGR clusters of PV-IR cells (arrows) wereobserved in the immunoreactive fiber patches (open trian-gles) (Figure 5(a)(B)) Scatteredmedium-to-large PV-IR cells(arrows) were found in the DL (Figure 5(a)(C)) including afew that are immediately adjacent to the periaqueductal grayand occasionally in the ZOThe neuropil labeling was patchyin the IGR and moderate in the DL (Figure 2(b))
32 Morphology of the Otx2- and PV-IR Cells in the ControlMice Figure 4(a)(C) represents various types of Otx2-IRcells in the control mouse SC The principal cell types inthe mouse SC labeled with Otx2 antibody were dark or lightround to oval cells These labeled IR elements were located inthe nucleus of the cell body consistent with a previous study[30 58] However ratios of the strongly and weakly stainedcells varied between the superficial and deep layers of the SCIn the superficial layers of the SC a large majority of Otx2-IR cells were composed of the strongly stained cells In thedeep layers of the SC most of the labeled cells were weakerand smaller compared with those observed in the superficiallayers of the SC Furthermore there were some differencesin diameter (the average of long and short axes) betweenthe two divisions The average diameter of Otx2-IR cells inthe superficial layers of the SC ranged from 326 to 900 120583mwith a mean of 601 120583m The average diameter of Otx2-IRcells in the deep layers of the SC ranged from 317 to 850120583mwith a mean of 575120583m Statistical comparison indicated that
BioMed Research International 5
(B)
SL
DL
PAG
(C)
SL
DL
PAG
IGR
IGR
DL
PAG
(A)
SL
IGR
minus394mm
minus458mm
minus33mm
(a) Otx2
SL
IGR
DL
PAG
SL
IGR
DL
PAG
SL
IGR
DL
PAG
(B)
(C)
(A)
minus394mm
minus458mm
minus33mm
(b) PV
Figure 2 Low power photomicrographs of the laminar distribution of Otx2-IR (a) and parvalbumin (PV-) IR (b) elements in the controlmouse SC Two matching series of left coronal sections in 3AP levels (AndashC) are presented Note prominent Otx2-IR cells in the superficiallayers (SLs) and scattered Otx2-IR cells in the intermediate gray layer (IGR) and deep layers (DLs) Also note strong PV-labeling in the SLpatchy PVlabeling in the IGR and scattered andor clustered PV-IR cells in the IGR and DL Number below each section indicate distancefrom the bregma PAG periaqueductal gray matter Scale bar = 200 120583m
6 BioMed Research International
(A) Otx2
IGR
DL
PAG
SGR
SGR
IGR
DL
PAG
(B) PV
(a) Control
(A) Otx2
SGR
IGR
PAG
DL
IGR
DL
PAG
SGR
(B) PV
(b) KO
Figure 3 Distribution of Otx2-IR (A) and parvalbumin (PV-) IR (B) cells in the control (a) and KO (b) mice Image data were masked bythreshold intensity using ImageJ SGR superficial gray IGR intermediate gray layer DL deep layer PAG periaqueductal gray matter Scalebar = 200 120583m
themean sizes of Otx2-IR cells were significantly larger in thesuperficial layers than those in the deep layers (unpaired 119905-test 119875 lt 0001)
Figure 6 represents various types of PV-IR cells in thecontrol mouse SC We categorized the PV-IR cells as verticaland horizontal fusiform pyriform marginal (round) andmultipolar cells according to the previous studies [56 57]Thetypes of PV-IR cells identified in the mouse SC were morevariable than the classes of Otx2-IR cells These labeled cellsconsisted of at least four distinctmorphologiesMarginal cellshad small roundish or stellate soma with few or no labeleddendrites (a) These cells were the most prominent type inthe superficial layers of the mouse SC Pyriform (pyramidal)cells had pear-shaped soma with one tufted dendritic treeextending above the cell body (b) This cell type was locatedmostly in the ventral SGR IGR and DL
Horizontal fusiform cells had horizontally elongated tri-angular or fusiform cell with stout dendrites coursing in thesame plane as the cell body (c)These cells were foundmostlyin the dorsal OP and in the lateral region of the DL Verticalfusiform cells had vertical fusiform soma usually with onedendrite emanating vertically from each pole (d) These cellswere located mostly in the ventral SGR
Fourth type of the PV-IR cells was multipolar cells withlarge lightly stained multipolar soma and several dendritesthat radiated from the soma (e) These cells were observed inthe IGR and DL
Furthermore the three layers had their correspondingdiameters The average diameter of PV-IR cells in the super-ficial layers of the SC ranged from 608 to 2162120583m with amean of 913 120583m In the IGR the diameter of labeled cellsranged from 608 to 2027120583m with a mean of 1076120583m Inthe DL of the SC the diameter of PV-IR cells ranged from703 to 2162 120583m with a mean of 1291120583m Statistical analysisindicated that there was a significant difference in cell sizeamong the 3 laminae (119875 lt 0001 Bonferroni tests afterrepeated measures one-way ANOVA)
33 Otx2 and PV Expression in the PDGFR-120573 KO MiceComparison of the control and KO mice revealed simi-lar distribution patterns of Otx2- and PV-IR cells in theSC (Figure 3) in which the image data were thresholdedto highlight the labeled cell somas or nuclei Thus twodistinctive tiers of Otx2-IR labeling in the KO mouse SC(Figure 4(b)) were virtually identical to those in the control
BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
often found in ldquofast spikingrdquo neurons with nonadapting trainsof action potentials It has been suggested that PVmight regu-late a Ca2+-activated K+ channel involved in spike adaptation[15] and a particular type of K+ channel was developmentallyregulated with PV and was expressed in about 80 of the PV-containing GABAergic interneurons in the rat hippocampus[16] Furthermore deficits in these PV-containing GABAer-gic interneurons were associated with schizophrenia andautism with social deficits [17 18] and the number of PV-positive neurons was reduced in the SC of an animal modelof autism with prenatal valproic acid exposure [19]
The Otx2 homeobox gene is a vertebrate orthologue ofthe Drosophila orthodenticle gene [20ndash22] members of thisorthology group play a fundamental role in developmentof photoreceptors Otx2 and Crx (cone-rod homeobox) androstral brain regions (Otx1 and Otx2) [23 24] Otx2 alsoappears to play a role in development and function of theretina in which the gene is expressed at both prenatal andpostnatal stages [25ndash29] In addition Otx2 coordinates post-natal PVcell maturation and activates visual cortical plasticity[30] Otx2 is strikingly restricted to relay centers in theprimary visual pathway at birth before PV-cells are evident[31] including the retina LGN and visual cortex (V1)Furthermore exogenous Otx2 infusion accelerated PV-celldevelopment in the V1 area [30] These findings indicate thatOtx2 is related to development of the visual system Howeverto our knowledge no information is available concerningOtx2 expression in the retinocollicular (retinotectal) visualpathway in adult mice It is possible that Otx2 may controlfunctional features of SC neurons in adult mice
Platelet-derived growth factor (PDGF) was originallyreported as a substance in platelets that promote growthof tissue culture cells [32] However the factor is recentlyreported to act as a neuroprotective factor in the centralnervous system (CNS) [33ndash36] PDGF is also involved inthe regulation of cell growth and differentiation duringembryonal development [37ndash39] The family of platelet-derived growth factors (PDGFs) comprises 4 membersmdashPDGF-A -B -C and -Dmdashthat are assembled from disulfide-linked homo- or heterodimers of 2 distinct but related chains(PDGF-AA -AB -BB -CC and -DD) Two receptor subtypesof PDGF (PDGFR-120572 and -120573) can form mature dimericreceptor complexes that can bind to ligands with differ-ent affinities [40] PDGFR-120572 is largely expressed in oligo-dendroglial progenitors while PDGFR-120573 is predominantlyexpressed in neurons [41] and upregulated in the neonatalrat brain [42] PDGF-BB that specifically binds to PDGFR-120573120573 is abundantly expressed in neurons and is upregulatedin neonatal brains [33 43 44] PDGF-B exerts neurotrophiceffects on 120574-aminobutyric acid GABA ergic neurons [42 45]Altering PDGFR-120573 may result in abnormalities during thedevelopment of the CNS [46 47] Consistently conditionalknockout (KO) mice with suppressed expression of neuronalPDGFR-120573 in the CreloxP system (PDGFR-120573 KO mice)displayed autisticschizophrenic traits especially deficits insocioemotional behaviors [48]
Taken together there is a possible link among PV Otx2and PDGFR-120573 and behavioral deficits in the PDGFR-120573
KO mice might be ascribed to changes in PV and Otx2in the SC In the present study to investigate the role ofPDGFR-120573 in Otx2 and PV expression in the SC we analyzedimmunohistochemical alterations in Otx2 and PV expressionin the SC of PDGFR-120573 KOmice
2 Materials and Methods
21 Generation of the Conditional PDGFR-120573 Knockout MiceThe CreloxP system was used to develop conditionalPDGFR-120573 KO mutants A previously established mutantmouse line was used in which exons 4ndash7 of PDGFR-120573 whichencode the extracellular domain of the PDGFR-120573 proteinwere flanked by 2 loxP sequences (floxed) positioned inintrons 3 and 7 [49] After Cre-mediated recombinationdeletion of the loxP-flanking region and resulting frame shiftmutation in the adjoining 31015840 region occurred in PDGFR-120573 To obtain conditional PDGFR-120573 KO we then crossedmutant mice harboring the PDGFR-120573 floxed allele and thoseexpressing Cre recombinase under the control of the nestinpromoter and enhancer (nestin-Cre+ mouse The JacksonLaboratory Bar Harbor ME USA) as previously described[50 51] Before this cross both mutant mice harboringfloxed PDGFR-120573 and nestin-Cre+ were outbred to the miceof C57BL6J (B6J) strain for 14 generations to replace thegenetic background of our mutant mice with that of the B6Jstrain In the present study the following 2 types of 22 to 29weeks old male mice were used mice with the Cre transgeneand floxed PDGFR-120573 (PDGFR-120573KOmice) andmice withoutthe Cre transgene but with floxed PDGFR-120573 (control mice)
Genotypes were confirmed by PCR of tail DNA usingoligonucleotide primers pairs for floxedPDGFR-120573 and for theCre transgene as described previously [50] The genotypingwas confirmed by Western blot of the total lysates of theadult mouse brains to show that the PDGFR-120573 expressiondecreased to undetectable levels in the PDGFR-120573 KO micecompared with that in the control mice [50]
In the present study 6 control mice (24ndash28 weeks old 22ndash29 g body weight) and 4 PDGFR-120573 KO mice (24ndash28 weeksold 22ndash29 g body weight) were used All mice were housedin individual cages in a temperature-controlled environmentwith a 1212 h lightdark cycle (lights were turned on and offat 0800 and 2000 resp) Food and water were supplied adlibitumMice (10 to 16weeks old) were handled for 3 consecu-tive days before the start of the experiments All experimentalprotocols were performed in accordance with the guidelinesfor Care and Use of Laboratory Animals approved by theUniversity of Toyama and the National Institutes of HealthrsquosGuide for the Care and Use of Laboratory Animals andapproved by the Committee for Animal Experiments at theUniversity of Toyama
22 Perfusion and Tissue Processing Under deep anesthesiawith sodium pentobarbital (50mgkg body weight ip) themice were transcardially perfused with heparinized saline(09 wv NaCl) followed by 4 paraformaldehyde dis-solved in 01Mphosphate buffer (PB) pH74After perfusionthe brains were removed from the skull cut coronally into
BioMed Research International 3
small blocks and postfixed in 4 paraformaldehyde forovernight The fixed brain blocks were immersed into 30sucrose in 01M PB until they sank down on the bottomThebrain blocks were freezed in dry ice and coronally cut intosections at the thickness of 40120583mThe sectionswere collectedin 001MPBS and then transferred into a cryoprotectant solu-tion (25 ethylene glycol 25 glycerin and 50 01M PBS)and stored at minus20∘C until immunohistochemical staining
23 Immunohistochemistry of the Sections Three serial sec-tions were collected every 120 120583m two for PV and Otx2-immunocytochemistry and one for Nissl staining with cresylviolet The sections were washed thrice in 001M phosphatebuffer saline (PBS) for 15min blocked with 3 normalhorse serum 025 Triton X-100 in PBS for 30min at roomtemperature and incubated overnight at 4∘C with mousemonoclonal anti-PV (1 10000 dilution Sigma St LouisMO USA) or goat polyclonal anti-Otx2 (1 200 dilutionRampD) antibodies in 1 blocking solution 025 Triton X-100 in PBS These sections were washed thrice with 001MPBS for 5min each time incubated with biotinylated horseanti-mouse IgG or anti-goat IgG (1 250 dilution VectorBurlingame USA) for 1 hr at room temperature and thenafter washing incubated in ABC reagent (Vector) for 50minFinally the PV- and Otx2-immunoreactive elements werevisualized by reacting with 20mg 331015840-diaminobenzidine and20120583L 30 H
2O2in 100mL 005M PBS (pH 76) for 5ndash
8min The sections were then rinsed several times in PBSdehydrated in graded concentrations of ethanol cleared inxylene and cover-slipped with Entellan (Merck DarmstadtGermany) To minimize variability in staining the sectionsfrom the all treatment groups were run in the same immuno-cytochemistry session Negative control sections were treatedidentically except for omission of the primary antibody Noreaction product was observed in any of the control sections
The corresponding adjacent sections were stained withcresyl violet to identify colliculus layers The colliculus layerswere identified by staining and cellular densities and mor-phologies of SC neurons [1] We also identified collicular lay-ers using parvalbumin-labeled sections where the collicularlayers of these proteins have been well described previously[12 52 53]
24 Identification of the SC and Image Capture In eachsection the SC was identified and subdivided into collicularlayers The nomenclature of the mouse atlas of Patrick andWarren (2000) was adopted Labeling was examined usingstandard bright-field microscope (BX 61 Olympus TokyoJapan) with different magnifications (4sim100x) A series ofimages of the SC were captured by a digital camera (DP 70Olympus Tokyo)
25 Data Analysis In the present study 6 coronal sections(minus330 minus362 minus394 minus426 minus458 and minus490mm from thebregma) in each mouse were analyzed for quantification ofOtx2- and PV-IR cells These digital images were analyzedusing the ImageJ software (NIH ImageJ httprsbwebnihgovij) The mean number of cells per section in each layer
was estimated based upon two approache the measurementof pixel intensity and counting of immunopositive cellsFor the pixel intensity of immunopositive cells all imagesused for cell quantification were saved as uncompressed 8-bit grey-scale tiff files and then were thresholded using theautothreshold tool in ImageJ with visual comparison beingmade to the original grey-scale images to ensure that the tooleffectively resolved all labeled cell somas or nucleiThereforeparticle analysis was carried out using the analyze particletool The mask tool was used to confirm that all particles(labeled cell somas or nuclei) were detected and measured
Estimates of cell number in two-dimensional sections aresubject to error and bias which can be partially overcomeby use of the Abercrombie correction [54 55] To enablesuch a correction in the current study sections were cut atthickness of 40120583m and nuclear diameter of Otx2-IR cellsand soma diameters of PV-IR cells (the average of long andshort axes) were computed by measuring the long and shortaxes of cells counted The correction factor was then appliedto all cell measurements The assessments were carried outby a single investigator (JJ Z) blind to group status In orderto furthermore diminish variability rating of images wasperformed on 10 different occasions by the same blindedobserver Great care was taken to match sections throughthe same region of the brain and at the same level using theanatomical landmarks Layers of the SC were identified usingthe counterstained sections of both the control and KO miceand then outlined using commercial software (Corel Draw12) The collicular layers traced as above were analyzed onthe corresponding collicular areas of immunocytologicallyprocessed control and KO mice
We analyzed the mean number of cells in each layer (6sections from each of the 6 control mice and 4KO mice)the diameter of each cell counted (6 sections from each ofthe 4 control mice and 4KO mice) and categorized cells asvertical and horizontal fusiform pyriformmarginal (round)and multipolar cells according to the previous studies [5657] All statistical comparisons (119905-test and repeated measureANOVA) were performed using SPSS 170 software A valueof 119875 lt 005 was considered to be significant
3 Results
31 Distribution of Otx2- and Parvalbumin (PV-) Immunore-active (IR) Cells in the SC Figure 1 shows the location ofthe SC in the coronal (a) sections of the mouse brain atlasand a section stained by cresyl violet (b) indicating differentsubdivisions of the SC Neuroanatomically the SC has twodivisions a superficial subdivision comprising the zonal(ZO) superficial gray (SGR) and optic (OP) layers and a deepsubdivision comprising the intermediate gray intermediatewhite deep gray and deep white layers The two layers ofwhite matter in the deep subdivision are often rather indis-tinct and do not appear to demarcate cellular distributionsTherefore we simply divided the deep subdivision into tworegions the intermediate gray (IGR) layer and the deep layers(DLs) including the deep white deep gray and intermediatewhite layers
4 BioMed Research International
minus41mm
(a)
ZO
SGR
OPIGR
DL
PAG
minus394mm
(b)
Figure 1 Coronal section in the brain atlas (AP minus41mm from the bregma) (a) and representative section stained with cresyl violet (APminus394mm from the bregma) (b) showing the location and laminar pattern of the mouse SC The square in A indicates the SC region ZOzonale SGR superficial gray layer OP optic IGR intermediate gray layer DL deep layer and PAG periaqueductal gray matter Scale bar =200 120583m
Figure 2 illustrates low power photomicrographs of thecontrol mouse SC sections stained with Otx2 antibody (A)and PV antibody (B) indicating the laminar distributionsof Otx2 and PV-IR cells Figure 3(a) shows black and whitephotomicrographs of the rostral SC sections stained withOtx2 (a) and PV (b) antibodies in the control mice as afunction of the threshold intensity The image data werethresholded to highlight the labeled cell somas or nucleiFigure 4(a) indicates high-magnification photomicrographsof the Otx2-labeled sections in the superficial (A) and deepmain (B) layers of the SC in the control mice The patterns ofOtx2 and PV labeling in the present study were different fromeach other Otx2-IR cells were found in all layers of the SCbut more densely in the superficial layers PV-IR cells werealso observed in all layers of the SC but different from that ofOtx2-IR cells (see below in detail)
The dense distribution of Otx2-IR cells was found in thesuperficial layers (ZO SGR and OP) (Figures 2(a) 3(a)(A)and 4(a)(A)) Otx2-IR cells were also observed in the deeplayers (IGR and DL) (Figures 2(a) 3(a)(A) and 4(a)(B))which mainly included lightly labeled small to medium-sized cells (Figure 4(a)(B)) In contrast very few labeled cellswere observed in the areas surrounding the SC The similardistribution patterns of Otx2-IR cells were observed throughthe rostral-caudal extent of the SC (Figure 2(a))
Figure 5(a) indicates high-magnification photomicro-graphs of the PV-labeled sections in the superficial (A)and deep main layers (B C) of the control mouse SCPV-IR cells were concentrated within the superficial layersof the SC with a heavier frequency within SGR than inthe OP (Figure 5(a)(A) Also see Figures 2(b) and 3(a)(B))Labeled IR elements in the dense tier included not only
lightly stained small-to-medium sized somas but also proxi-mal dendrites and small immunoreactive puncta that couldbe axons axon terminals or small dendrites (open tri-angles) (Figure 5(a)(A)) PV-IR cells were also scatteredthroughout the deep layers below the dense tier In thelateral part of the IGR clusters of PV-IR cells (arrows) wereobserved in the immunoreactive fiber patches (open trian-gles) (Figure 5(a)(B)) Scatteredmedium-to-large PV-IR cells(arrows) were found in the DL (Figure 5(a)(C)) including afew that are immediately adjacent to the periaqueductal grayand occasionally in the ZOThe neuropil labeling was patchyin the IGR and moderate in the DL (Figure 2(b))
32 Morphology of the Otx2- and PV-IR Cells in the ControlMice Figure 4(a)(C) represents various types of Otx2-IRcells in the control mouse SC The principal cell types inthe mouse SC labeled with Otx2 antibody were dark or lightround to oval cells These labeled IR elements were located inthe nucleus of the cell body consistent with a previous study[30 58] However ratios of the strongly and weakly stainedcells varied between the superficial and deep layers of the SCIn the superficial layers of the SC a large majority of Otx2-IR cells were composed of the strongly stained cells In thedeep layers of the SC most of the labeled cells were weakerand smaller compared with those observed in the superficiallayers of the SC Furthermore there were some differencesin diameter (the average of long and short axes) betweenthe two divisions The average diameter of Otx2-IR cells inthe superficial layers of the SC ranged from 326 to 900 120583mwith a mean of 601 120583m The average diameter of Otx2-IRcells in the deep layers of the SC ranged from 317 to 850120583mwith a mean of 575120583m Statistical comparison indicated that
BioMed Research International 5
(B)
SL
DL
PAG
(C)
SL
DL
PAG
IGR
IGR
DL
PAG
(A)
SL
IGR
minus394mm
minus458mm
minus33mm
(a) Otx2
SL
IGR
DL
PAG
SL
IGR
DL
PAG
SL
IGR
DL
PAG
(B)
(C)
(A)
minus394mm
minus458mm
minus33mm
(b) PV
Figure 2 Low power photomicrographs of the laminar distribution of Otx2-IR (a) and parvalbumin (PV-) IR (b) elements in the controlmouse SC Two matching series of left coronal sections in 3AP levels (AndashC) are presented Note prominent Otx2-IR cells in the superficiallayers (SLs) and scattered Otx2-IR cells in the intermediate gray layer (IGR) and deep layers (DLs) Also note strong PV-labeling in the SLpatchy PVlabeling in the IGR and scattered andor clustered PV-IR cells in the IGR and DL Number below each section indicate distancefrom the bregma PAG periaqueductal gray matter Scale bar = 200 120583m
6 BioMed Research International
(A) Otx2
IGR
DL
PAG
SGR
SGR
IGR
DL
PAG
(B) PV
(a) Control
(A) Otx2
SGR
IGR
PAG
DL
IGR
DL
PAG
SGR
(B) PV
(b) KO
Figure 3 Distribution of Otx2-IR (A) and parvalbumin (PV-) IR (B) cells in the control (a) and KO (b) mice Image data were masked bythreshold intensity using ImageJ SGR superficial gray IGR intermediate gray layer DL deep layer PAG periaqueductal gray matter Scalebar = 200 120583m
themean sizes of Otx2-IR cells were significantly larger in thesuperficial layers than those in the deep layers (unpaired 119905-test 119875 lt 0001)
Figure 6 represents various types of PV-IR cells in thecontrol mouse SC We categorized the PV-IR cells as verticaland horizontal fusiform pyriform marginal (round) andmultipolar cells according to the previous studies [56 57]Thetypes of PV-IR cells identified in the mouse SC were morevariable than the classes of Otx2-IR cells These labeled cellsconsisted of at least four distinctmorphologiesMarginal cellshad small roundish or stellate soma with few or no labeleddendrites (a) These cells were the most prominent type inthe superficial layers of the mouse SC Pyriform (pyramidal)cells had pear-shaped soma with one tufted dendritic treeextending above the cell body (b) This cell type was locatedmostly in the ventral SGR IGR and DL
Horizontal fusiform cells had horizontally elongated tri-angular or fusiform cell with stout dendrites coursing in thesame plane as the cell body (c)These cells were foundmostlyin the dorsal OP and in the lateral region of the DL Verticalfusiform cells had vertical fusiform soma usually with onedendrite emanating vertically from each pole (d) These cellswere located mostly in the ventral SGR
Fourth type of the PV-IR cells was multipolar cells withlarge lightly stained multipolar soma and several dendritesthat radiated from the soma (e) These cells were observed inthe IGR and DL
Furthermore the three layers had their correspondingdiameters The average diameter of PV-IR cells in the super-ficial layers of the SC ranged from 608 to 2162120583m with amean of 913 120583m In the IGR the diameter of labeled cellsranged from 608 to 2027120583m with a mean of 1076120583m Inthe DL of the SC the diameter of PV-IR cells ranged from703 to 2162 120583m with a mean of 1291120583m Statistical analysisindicated that there was a significant difference in cell sizeamong the 3 laminae (119875 lt 0001 Bonferroni tests afterrepeated measures one-way ANOVA)
33 Otx2 and PV Expression in the PDGFR-120573 KO MiceComparison of the control and KO mice revealed simi-lar distribution patterns of Otx2- and PV-IR cells in theSC (Figure 3) in which the image data were thresholdedto highlight the labeled cell somas or nuclei Thus twodistinctive tiers of Otx2-IR labeling in the KO mouse SC(Figure 4(b)) were virtually identical to those in the control
BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
small blocks and postfixed in 4 paraformaldehyde forovernight The fixed brain blocks were immersed into 30sucrose in 01M PB until they sank down on the bottomThebrain blocks were freezed in dry ice and coronally cut intosections at the thickness of 40120583mThe sectionswere collectedin 001MPBS and then transferred into a cryoprotectant solu-tion (25 ethylene glycol 25 glycerin and 50 01M PBS)and stored at minus20∘C until immunohistochemical staining
23 Immunohistochemistry of the Sections Three serial sec-tions were collected every 120 120583m two for PV and Otx2-immunocytochemistry and one for Nissl staining with cresylviolet The sections were washed thrice in 001M phosphatebuffer saline (PBS) for 15min blocked with 3 normalhorse serum 025 Triton X-100 in PBS for 30min at roomtemperature and incubated overnight at 4∘C with mousemonoclonal anti-PV (1 10000 dilution Sigma St LouisMO USA) or goat polyclonal anti-Otx2 (1 200 dilutionRampD) antibodies in 1 blocking solution 025 Triton X-100 in PBS These sections were washed thrice with 001MPBS for 5min each time incubated with biotinylated horseanti-mouse IgG or anti-goat IgG (1 250 dilution VectorBurlingame USA) for 1 hr at room temperature and thenafter washing incubated in ABC reagent (Vector) for 50minFinally the PV- and Otx2-immunoreactive elements werevisualized by reacting with 20mg 331015840-diaminobenzidine and20120583L 30 H
2O2in 100mL 005M PBS (pH 76) for 5ndash
8min The sections were then rinsed several times in PBSdehydrated in graded concentrations of ethanol cleared inxylene and cover-slipped with Entellan (Merck DarmstadtGermany) To minimize variability in staining the sectionsfrom the all treatment groups were run in the same immuno-cytochemistry session Negative control sections were treatedidentically except for omission of the primary antibody Noreaction product was observed in any of the control sections
The corresponding adjacent sections were stained withcresyl violet to identify colliculus layers The colliculus layerswere identified by staining and cellular densities and mor-phologies of SC neurons [1] We also identified collicular lay-ers using parvalbumin-labeled sections where the collicularlayers of these proteins have been well described previously[12 52 53]
24 Identification of the SC and Image Capture In eachsection the SC was identified and subdivided into collicularlayers The nomenclature of the mouse atlas of Patrick andWarren (2000) was adopted Labeling was examined usingstandard bright-field microscope (BX 61 Olympus TokyoJapan) with different magnifications (4sim100x) A series ofimages of the SC were captured by a digital camera (DP 70Olympus Tokyo)
25 Data Analysis In the present study 6 coronal sections(minus330 minus362 minus394 minus426 minus458 and minus490mm from thebregma) in each mouse were analyzed for quantification ofOtx2- and PV-IR cells These digital images were analyzedusing the ImageJ software (NIH ImageJ httprsbwebnihgovij) The mean number of cells per section in each layer
was estimated based upon two approache the measurementof pixel intensity and counting of immunopositive cellsFor the pixel intensity of immunopositive cells all imagesused for cell quantification were saved as uncompressed 8-bit grey-scale tiff files and then were thresholded using theautothreshold tool in ImageJ with visual comparison beingmade to the original grey-scale images to ensure that the tooleffectively resolved all labeled cell somas or nucleiThereforeparticle analysis was carried out using the analyze particletool The mask tool was used to confirm that all particles(labeled cell somas or nuclei) were detected and measured
Estimates of cell number in two-dimensional sections aresubject to error and bias which can be partially overcomeby use of the Abercrombie correction [54 55] To enablesuch a correction in the current study sections were cut atthickness of 40120583m and nuclear diameter of Otx2-IR cellsand soma diameters of PV-IR cells (the average of long andshort axes) were computed by measuring the long and shortaxes of cells counted The correction factor was then appliedto all cell measurements The assessments were carried outby a single investigator (JJ Z) blind to group status In orderto furthermore diminish variability rating of images wasperformed on 10 different occasions by the same blindedobserver Great care was taken to match sections throughthe same region of the brain and at the same level using theanatomical landmarks Layers of the SC were identified usingthe counterstained sections of both the control and KO miceand then outlined using commercial software (Corel Draw12) The collicular layers traced as above were analyzed onthe corresponding collicular areas of immunocytologicallyprocessed control and KO mice
We analyzed the mean number of cells in each layer (6sections from each of the 6 control mice and 4KO mice)the diameter of each cell counted (6 sections from each ofthe 4 control mice and 4KO mice) and categorized cells asvertical and horizontal fusiform pyriformmarginal (round)and multipolar cells according to the previous studies [5657] All statistical comparisons (119905-test and repeated measureANOVA) were performed using SPSS 170 software A valueof 119875 lt 005 was considered to be significant
3 Results
31 Distribution of Otx2- and Parvalbumin (PV-) Immunore-active (IR) Cells in the SC Figure 1 shows the location ofthe SC in the coronal (a) sections of the mouse brain atlasand a section stained by cresyl violet (b) indicating differentsubdivisions of the SC Neuroanatomically the SC has twodivisions a superficial subdivision comprising the zonal(ZO) superficial gray (SGR) and optic (OP) layers and a deepsubdivision comprising the intermediate gray intermediatewhite deep gray and deep white layers The two layers ofwhite matter in the deep subdivision are often rather indis-tinct and do not appear to demarcate cellular distributionsTherefore we simply divided the deep subdivision into tworegions the intermediate gray (IGR) layer and the deep layers(DLs) including the deep white deep gray and intermediatewhite layers
4 BioMed Research International
minus41mm
(a)
ZO
SGR
OPIGR
DL
PAG
minus394mm
(b)
Figure 1 Coronal section in the brain atlas (AP minus41mm from the bregma) (a) and representative section stained with cresyl violet (APminus394mm from the bregma) (b) showing the location and laminar pattern of the mouse SC The square in A indicates the SC region ZOzonale SGR superficial gray layer OP optic IGR intermediate gray layer DL deep layer and PAG periaqueductal gray matter Scale bar =200 120583m
Figure 2 illustrates low power photomicrographs of thecontrol mouse SC sections stained with Otx2 antibody (A)and PV antibody (B) indicating the laminar distributionsof Otx2 and PV-IR cells Figure 3(a) shows black and whitephotomicrographs of the rostral SC sections stained withOtx2 (a) and PV (b) antibodies in the control mice as afunction of the threshold intensity The image data werethresholded to highlight the labeled cell somas or nucleiFigure 4(a) indicates high-magnification photomicrographsof the Otx2-labeled sections in the superficial (A) and deepmain (B) layers of the SC in the control mice The patterns ofOtx2 and PV labeling in the present study were different fromeach other Otx2-IR cells were found in all layers of the SCbut more densely in the superficial layers PV-IR cells werealso observed in all layers of the SC but different from that ofOtx2-IR cells (see below in detail)
The dense distribution of Otx2-IR cells was found in thesuperficial layers (ZO SGR and OP) (Figures 2(a) 3(a)(A)and 4(a)(A)) Otx2-IR cells were also observed in the deeplayers (IGR and DL) (Figures 2(a) 3(a)(A) and 4(a)(B))which mainly included lightly labeled small to medium-sized cells (Figure 4(a)(B)) In contrast very few labeled cellswere observed in the areas surrounding the SC The similardistribution patterns of Otx2-IR cells were observed throughthe rostral-caudal extent of the SC (Figure 2(a))
Figure 5(a) indicates high-magnification photomicro-graphs of the PV-labeled sections in the superficial (A)and deep main layers (B C) of the control mouse SCPV-IR cells were concentrated within the superficial layersof the SC with a heavier frequency within SGR than inthe OP (Figure 5(a)(A) Also see Figures 2(b) and 3(a)(B))Labeled IR elements in the dense tier included not only
lightly stained small-to-medium sized somas but also proxi-mal dendrites and small immunoreactive puncta that couldbe axons axon terminals or small dendrites (open tri-angles) (Figure 5(a)(A)) PV-IR cells were also scatteredthroughout the deep layers below the dense tier In thelateral part of the IGR clusters of PV-IR cells (arrows) wereobserved in the immunoreactive fiber patches (open trian-gles) (Figure 5(a)(B)) Scatteredmedium-to-large PV-IR cells(arrows) were found in the DL (Figure 5(a)(C)) including afew that are immediately adjacent to the periaqueductal grayand occasionally in the ZOThe neuropil labeling was patchyin the IGR and moderate in the DL (Figure 2(b))
32 Morphology of the Otx2- and PV-IR Cells in the ControlMice Figure 4(a)(C) represents various types of Otx2-IRcells in the control mouse SC The principal cell types inthe mouse SC labeled with Otx2 antibody were dark or lightround to oval cells These labeled IR elements were located inthe nucleus of the cell body consistent with a previous study[30 58] However ratios of the strongly and weakly stainedcells varied between the superficial and deep layers of the SCIn the superficial layers of the SC a large majority of Otx2-IR cells were composed of the strongly stained cells In thedeep layers of the SC most of the labeled cells were weakerand smaller compared with those observed in the superficiallayers of the SC Furthermore there were some differencesin diameter (the average of long and short axes) betweenthe two divisions The average diameter of Otx2-IR cells inthe superficial layers of the SC ranged from 326 to 900 120583mwith a mean of 601 120583m The average diameter of Otx2-IRcells in the deep layers of the SC ranged from 317 to 850120583mwith a mean of 575120583m Statistical comparison indicated that
BioMed Research International 5
(B)
SL
DL
PAG
(C)
SL
DL
PAG
IGR
IGR
DL
PAG
(A)
SL
IGR
minus394mm
minus458mm
minus33mm
(a) Otx2
SL
IGR
DL
PAG
SL
IGR
DL
PAG
SL
IGR
DL
PAG
(B)
(C)
(A)
minus394mm
minus458mm
minus33mm
(b) PV
Figure 2 Low power photomicrographs of the laminar distribution of Otx2-IR (a) and parvalbumin (PV-) IR (b) elements in the controlmouse SC Two matching series of left coronal sections in 3AP levels (AndashC) are presented Note prominent Otx2-IR cells in the superficiallayers (SLs) and scattered Otx2-IR cells in the intermediate gray layer (IGR) and deep layers (DLs) Also note strong PV-labeling in the SLpatchy PVlabeling in the IGR and scattered andor clustered PV-IR cells in the IGR and DL Number below each section indicate distancefrom the bregma PAG periaqueductal gray matter Scale bar = 200 120583m
6 BioMed Research International
(A) Otx2
IGR
DL
PAG
SGR
SGR
IGR
DL
PAG
(B) PV
(a) Control
(A) Otx2
SGR
IGR
PAG
DL
IGR
DL
PAG
SGR
(B) PV
(b) KO
Figure 3 Distribution of Otx2-IR (A) and parvalbumin (PV-) IR (B) cells in the control (a) and KO (b) mice Image data were masked bythreshold intensity using ImageJ SGR superficial gray IGR intermediate gray layer DL deep layer PAG periaqueductal gray matter Scalebar = 200 120583m
themean sizes of Otx2-IR cells were significantly larger in thesuperficial layers than those in the deep layers (unpaired 119905-test 119875 lt 0001)
Figure 6 represents various types of PV-IR cells in thecontrol mouse SC We categorized the PV-IR cells as verticaland horizontal fusiform pyriform marginal (round) andmultipolar cells according to the previous studies [56 57]Thetypes of PV-IR cells identified in the mouse SC were morevariable than the classes of Otx2-IR cells These labeled cellsconsisted of at least four distinctmorphologiesMarginal cellshad small roundish or stellate soma with few or no labeleddendrites (a) These cells were the most prominent type inthe superficial layers of the mouse SC Pyriform (pyramidal)cells had pear-shaped soma with one tufted dendritic treeextending above the cell body (b) This cell type was locatedmostly in the ventral SGR IGR and DL
Horizontal fusiform cells had horizontally elongated tri-angular or fusiform cell with stout dendrites coursing in thesame plane as the cell body (c)These cells were foundmostlyin the dorsal OP and in the lateral region of the DL Verticalfusiform cells had vertical fusiform soma usually with onedendrite emanating vertically from each pole (d) These cellswere located mostly in the ventral SGR
Fourth type of the PV-IR cells was multipolar cells withlarge lightly stained multipolar soma and several dendritesthat radiated from the soma (e) These cells were observed inthe IGR and DL
Furthermore the three layers had their correspondingdiameters The average diameter of PV-IR cells in the super-ficial layers of the SC ranged from 608 to 2162120583m with amean of 913 120583m In the IGR the diameter of labeled cellsranged from 608 to 2027120583m with a mean of 1076120583m Inthe DL of the SC the diameter of PV-IR cells ranged from703 to 2162 120583m with a mean of 1291120583m Statistical analysisindicated that there was a significant difference in cell sizeamong the 3 laminae (119875 lt 0001 Bonferroni tests afterrepeated measures one-way ANOVA)
33 Otx2 and PV Expression in the PDGFR-120573 KO MiceComparison of the control and KO mice revealed simi-lar distribution patterns of Otx2- and PV-IR cells in theSC (Figure 3) in which the image data were thresholdedto highlight the labeled cell somas or nuclei Thus twodistinctive tiers of Otx2-IR labeling in the KO mouse SC(Figure 4(b)) were virtually identical to those in the control
BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
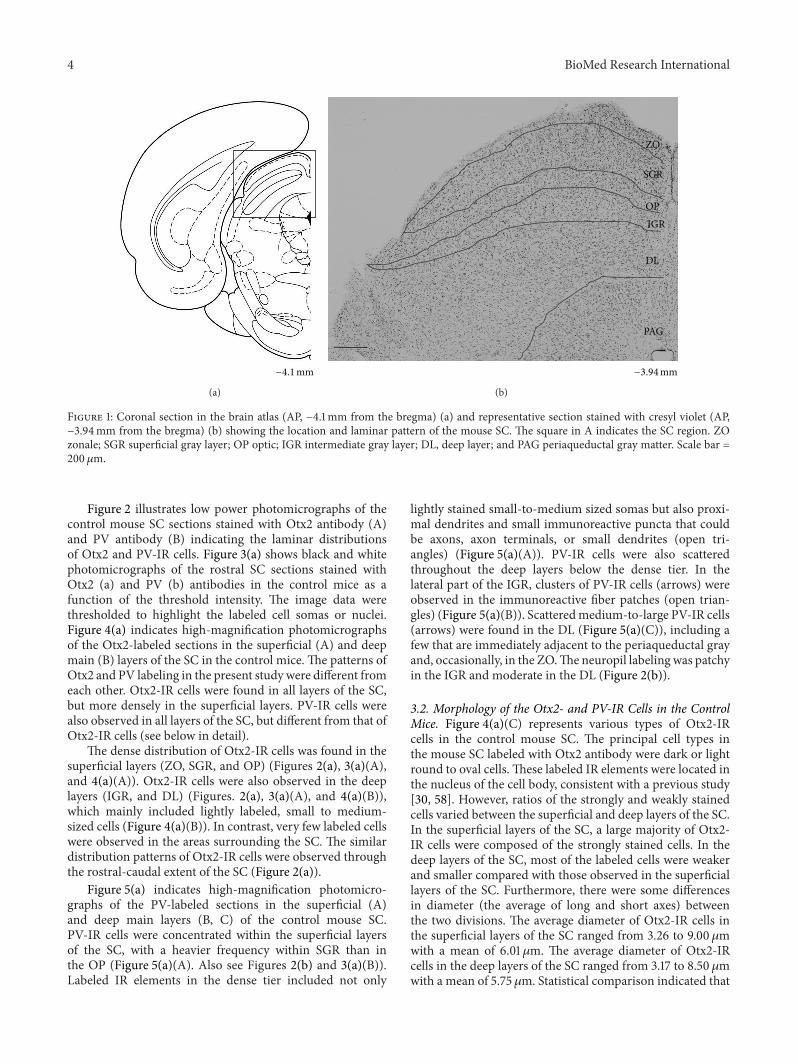
4 BioMed Research International
minus41mm
(a)
ZO
SGR
OPIGR
DL
PAG
minus394mm
(b)
Figure 1 Coronal section in the brain atlas (AP minus41mm from the bregma) (a) and representative section stained with cresyl violet (APminus394mm from the bregma) (b) showing the location and laminar pattern of the mouse SC The square in A indicates the SC region ZOzonale SGR superficial gray layer OP optic IGR intermediate gray layer DL deep layer and PAG periaqueductal gray matter Scale bar =200 120583m
Figure 2 illustrates low power photomicrographs of thecontrol mouse SC sections stained with Otx2 antibody (A)and PV antibody (B) indicating the laminar distributionsof Otx2 and PV-IR cells Figure 3(a) shows black and whitephotomicrographs of the rostral SC sections stained withOtx2 (a) and PV (b) antibodies in the control mice as afunction of the threshold intensity The image data werethresholded to highlight the labeled cell somas or nucleiFigure 4(a) indicates high-magnification photomicrographsof the Otx2-labeled sections in the superficial (A) and deepmain (B) layers of the SC in the control mice The patterns ofOtx2 and PV labeling in the present study were different fromeach other Otx2-IR cells were found in all layers of the SCbut more densely in the superficial layers PV-IR cells werealso observed in all layers of the SC but different from that ofOtx2-IR cells (see below in detail)
The dense distribution of Otx2-IR cells was found in thesuperficial layers (ZO SGR and OP) (Figures 2(a) 3(a)(A)and 4(a)(A)) Otx2-IR cells were also observed in the deeplayers (IGR and DL) (Figures 2(a) 3(a)(A) and 4(a)(B))which mainly included lightly labeled small to medium-sized cells (Figure 4(a)(B)) In contrast very few labeled cellswere observed in the areas surrounding the SC The similardistribution patterns of Otx2-IR cells were observed throughthe rostral-caudal extent of the SC (Figure 2(a))
Figure 5(a) indicates high-magnification photomicro-graphs of the PV-labeled sections in the superficial (A)and deep main layers (B C) of the control mouse SCPV-IR cells were concentrated within the superficial layersof the SC with a heavier frequency within SGR than inthe OP (Figure 5(a)(A) Also see Figures 2(b) and 3(a)(B))Labeled IR elements in the dense tier included not only
lightly stained small-to-medium sized somas but also proxi-mal dendrites and small immunoreactive puncta that couldbe axons axon terminals or small dendrites (open tri-angles) (Figure 5(a)(A)) PV-IR cells were also scatteredthroughout the deep layers below the dense tier In thelateral part of the IGR clusters of PV-IR cells (arrows) wereobserved in the immunoreactive fiber patches (open trian-gles) (Figure 5(a)(B)) Scatteredmedium-to-large PV-IR cells(arrows) were found in the DL (Figure 5(a)(C)) including afew that are immediately adjacent to the periaqueductal grayand occasionally in the ZOThe neuropil labeling was patchyin the IGR and moderate in the DL (Figure 2(b))
32 Morphology of the Otx2- and PV-IR Cells in the ControlMice Figure 4(a)(C) represents various types of Otx2-IRcells in the control mouse SC The principal cell types inthe mouse SC labeled with Otx2 antibody were dark or lightround to oval cells These labeled IR elements were located inthe nucleus of the cell body consistent with a previous study[30 58] However ratios of the strongly and weakly stainedcells varied between the superficial and deep layers of the SCIn the superficial layers of the SC a large majority of Otx2-IR cells were composed of the strongly stained cells In thedeep layers of the SC most of the labeled cells were weakerand smaller compared with those observed in the superficiallayers of the SC Furthermore there were some differencesin diameter (the average of long and short axes) betweenthe two divisions The average diameter of Otx2-IR cells inthe superficial layers of the SC ranged from 326 to 900 120583mwith a mean of 601 120583m The average diameter of Otx2-IRcells in the deep layers of the SC ranged from 317 to 850120583mwith a mean of 575120583m Statistical comparison indicated that
BioMed Research International 5
(B)
SL
DL
PAG
(C)
SL
DL
PAG
IGR
IGR
DL
PAG
(A)
SL
IGR
minus394mm
minus458mm
minus33mm
(a) Otx2
SL
IGR
DL
PAG
SL
IGR
DL
PAG
SL
IGR
DL
PAG
(B)
(C)
(A)
minus394mm
minus458mm
minus33mm
(b) PV
Figure 2 Low power photomicrographs of the laminar distribution of Otx2-IR (a) and parvalbumin (PV-) IR (b) elements in the controlmouse SC Two matching series of left coronal sections in 3AP levels (AndashC) are presented Note prominent Otx2-IR cells in the superficiallayers (SLs) and scattered Otx2-IR cells in the intermediate gray layer (IGR) and deep layers (DLs) Also note strong PV-labeling in the SLpatchy PVlabeling in the IGR and scattered andor clustered PV-IR cells in the IGR and DL Number below each section indicate distancefrom the bregma PAG periaqueductal gray matter Scale bar = 200 120583m
6 BioMed Research International
(A) Otx2
IGR
DL
PAG
SGR
SGR
IGR
DL
PAG
(B) PV
(a) Control
(A) Otx2
SGR
IGR
PAG
DL
IGR
DL
PAG
SGR
(B) PV
(b) KO
Figure 3 Distribution of Otx2-IR (A) and parvalbumin (PV-) IR (B) cells in the control (a) and KO (b) mice Image data were masked bythreshold intensity using ImageJ SGR superficial gray IGR intermediate gray layer DL deep layer PAG periaqueductal gray matter Scalebar = 200 120583m
themean sizes of Otx2-IR cells were significantly larger in thesuperficial layers than those in the deep layers (unpaired 119905-test 119875 lt 0001)
Figure 6 represents various types of PV-IR cells in thecontrol mouse SC We categorized the PV-IR cells as verticaland horizontal fusiform pyriform marginal (round) andmultipolar cells according to the previous studies [56 57]Thetypes of PV-IR cells identified in the mouse SC were morevariable than the classes of Otx2-IR cells These labeled cellsconsisted of at least four distinctmorphologiesMarginal cellshad small roundish or stellate soma with few or no labeleddendrites (a) These cells were the most prominent type inthe superficial layers of the mouse SC Pyriform (pyramidal)cells had pear-shaped soma with one tufted dendritic treeextending above the cell body (b) This cell type was locatedmostly in the ventral SGR IGR and DL
Horizontal fusiform cells had horizontally elongated tri-angular or fusiform cell with stout dendrites coursing in thesame plane as the cell body (c)These cells were foundmostlyin the dorsal OP and in the lateral region of the DL Verticalfusiform cells had vertical fusiform soma usually with onedendrite emanating vertically from each pole (d) These cellswere located mostly in the ventral SGR
Fourth type of the PV-IR cells was multipolar cells withlarge lightly stained multipolar soma and several dendritesthat radiated from the soma (e) These cells were observed inthe IGR and DL
Furthermore the three layers had their correspondingdiameters The average diameter of PV-IR cells in the super-ficial layers of the SC ranged from 608 to 2162120583m with amean of 913 120583m In the IGR the diameter of labeled cellsranged from 608 to 2027120583m with a mean of 1076120583m Inthe DL of the SC the diameter of PV-IR cells ranged from703 to 2162 120583m with a mean of 1291120583m Statistical analysisindicated that there was a significant difference in cell sizeamong the 3 laminae (119875 lt 0001 Bonferroni tests afterrepeated measures one-way ANOVA)
33 Otx2 and PV Expression in the PDGFR-120573 KO MiceComparison of the control and KO mice revealed simi-lar distribution patterns of Otx2- and PV-IR cells in theSC (Figure 3) in which the image data were thresholdedto highlight the labeled cell somas or nuclei Thus twodistinctive tiers of Otx2-IR labeling in the KO mouse SC(Figure 4(b)) were virtually identical to those in the control
BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
(B)
SL
DL
PAG
(C)
SL
DL
PAG
IGR
IGR
DL
PAG
(A)
SL
IGR
minus394mm
minus458mm
minus33mm
(a) Otx2
SL
IGR
DL
PAG
SL
IGR
DL
PAG
SL
IGR
DL
PAG
(B)
(C)
(A)
minus394mm
minus458mm
minus33mm
(b) PV
Figure 2 Low power photomicrographs of the laminar distribution of Otx2-IR (a) and parvalbumin (PV-) IR (b) elements in the controlmouse SC Two matching series of left coronal sections in 3AP levels (AndashC) are presented Note prominent Otx2-IR cells in the superficiallayers (SLs) and scattered Otx2-IR cells in the intermediate gray layer (IGR) and deep layers (DLs) Also note strong PV-labeling in the SLpatchy PVlabeling in the IGR and scattered andor clustered PV-IR cells in the IGR and DL Number below each section indicate distancefrom the bregma PAG periaqueductal gray matter Scale bar = 200 120583m
6 BioMed Research International
(A) Otx2
IGR
DL
PAG
SGR
SGR
IGR
DL
PAG
(B) PV
(a) Control
(A) Otx2
SGR
IGR
PAG
DL
IGR
DL
PAG
SGR
(B) PV
(b) KO
Figure 3 Distribution of Otx2-IR (A) and parvalbumin (PV-) IR (B) cells in the control (a) and KO (b) mice Image data were masked bythreshold intensity using ImageJ SGR superficial gray IGR intermediate gray layer DL deep layer PAG periaqueductal gray matter Scalebar = 200 120583m
themean sizes of Otx2-IR cells were significantly larger in thesuperficial layers than those in the deep layers (unpaired 119905-test 119875 lt 0001)
Figure 6 represents various types of PV-IR cells in thecontrol mouse SC We categorized the PV-IR cells as verticaland horizontal fusiform pyriform marginal (round) andmultipolar cells according to the previous studies [56 57]Thetypes of PV-IR cells identified in the mouse SC were morevariable than the classes of Otx2-IR cells These labeled cellsconsisted of at least four distinctmorphologiesMarginal cellshad small roundish or stellate soma with few or no labeleddendrites (a) These cells were the most prominent type inthe superficial layers of the mouse SC Pyriform (pyramidal)cells had pear-shaped soma with one tufted dendritic treeextending above the cell body (b) This cell type was locatedmostly in the ventral SGR IGR and DL
Horizontal fusiform cells had horizontally elongated tri-angular or fusiform cell with stout dendrites coursing in thesame plane as the cell body (c)These cells were foundmostlyin the dorsal OP and in the lateral region of the DL Verticalfusiform cells had vertical fusiform soma usually with onedendrite emanating vertically from each pole (d) These cellswere located mostly in the ventral SGR
Fourth type of the PV-IR cells was multipolar cells withlarge lightly stained multipolar soma and several dendritesthat radiated from the soma (e) These cells were observed inthe IGR and DL
Furthermore the three layers had their correspondingdiameters The average diameter of PV-IR cells in the super-ficial layers of the SC ranged from 608 to 2162120583m with amean of 913 120583m In the IGR the diameter of labeled cellsranged from 608 to 2027120583m with a mean of 1076120583m Inthe DL of the SC the diameter of PV-IR cells ranged from703 to 2162 120583m with a mean of 1291120583m Statistical analysisindicated that there was a significant difference in cell sizeamong the 3 laminae (119875 lt 0001 Bonferroni tests afterrepeated measures one-way ANOVA)
33 Otx2 and PV Expression in the PDGFR-120573 KO MiceComparison of the control and KO mice revealed simi-lar distribution patterns of Otx2- and PV-IR cells in theSC (Figure 3) in which the image data were thresholdedto highlight the labeled cell somas or nuclei Thus twodistinctive tiers of Otx2-IR labeling in the KO mouse SC(Figure 4(b)) were virtually identical to those in the control
BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
(A) Otx2
IGR
DL
PAG
SGR
SGR
IGR
DL
PAG
(B) PV
(a) Control
(A) Otx2
SGR
IGR
PAG
DL
IGR
DL
PAG
SGR
(B) PV
(b) KO
Figure 3 Distribution of Otx2-IR (A) and parvalbumin (PV-) IR (B) cells in the control (a) and KO (b) mice Image data were masked bythreshold intensity using ImageJ SGR superficial gray IGR intermediate gray layer DL deep layer PAG periaqueductal gray matter Scalebar = 200 120583m
themean sizes of Otx2-IR cells were significantly larger in thesuperficial layers than those in the deep layers (unpaired 119905-test 119875 lt 0001)
Figure 6 represents various types of PV-IR cells in thecontrol mouse SC We categorized the PV-IR cells as verticaland horizontal fusiform pyriform marginal (round) andmultipolar cells according to the previous studies [56 57]Thetypes of PV-IR cells identified in the mouse SC were morevariable than the classes of Otx2-IR cells These labeled cellsconsisted of at least four distinctmorphologiesMarginal cellshad small roundish or stellate soma with few or no labeleddendrites (a) These cells were the most prominent type inthe superficial layers of the mouse SC Pyriform (pyramidal)cells had pear-shaped soma with one tufted dendritic treeextending above the cell body (b) This cell type was locatedmostly in the ventral SGR IGR and DL
Horizontal fusiform cells had horizontally elongated tri-angular or fusiform cell with stout dendrites coursing in thesame plane as the cell body (c)These cells were foundmostlyin the dorsal OP and in the lateral region of the DL Verticalfusiform cells had vertical fusiform soma usually with onedendrite emanating vertically from each pole (d) These cellswere located mostly in the ventral SGR
Fourth type of the PV-IR cells was multipolar cells withlarge lightly stained multipolar soma and several dendritesthat radiated from the soma (e) These cells were observed inthe IGR and DL
Furthermore the three layers had their correspondingdiameters The average diameter of PV-IR cells in the super-ficial layers of the SC ranged from 608 to 2162120583m with amean of 913 120583m In the IGR the diameter of labeled cellsranged from 608 to 2027120583m with a mean of 1076120583m Inthe DL of the SC the diameter of PV-IR cells ranged from703 to 2162 120583m with a mean of 1291120583m Statistical analysisindicated that there was a significant difference in cell sizeamong the 3 laminae (119875 lt 0001 Bonferroni tests afterrepeated measures one-way ANOVA)
33 Otx2 and PV Expression in the PDGFR-120573 KO MiceComparison of the control and KO mice revealed simi-lar distribution patterns of Otx2- and PV-IR cells in theSC (Figure 3) in which the image data were thresholdedto highlight the labeled cell somas or nuclei Thus twodistinctive tiers of Otx2-IR labeling in the KO mouse SC(Figure 4(b)) were virtually identical to those in the control
BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7
(A) Superficial layers
(C)
(B) Deep layers
(a) Control (Otx2)
(A) Superficial layers
(C)
(B) Deep layers
(b) KO (Otx2)
Figure 4 High-magnification photomicrographs of the Otx2-labeled sections in the control (a) and KO (b) mice (a b) Photomicrographsin the superficial (A) and deep main (B) layers of the SC Note that the darklylabeled small-to-medium cells are densely distributed in thesuperficial layers in the SC of the control mouse Similar patterns of distribution of Otx2-IR cells are also observed in the KOmice but fewerOtx2-IR cells are observed in the KO mice Scale bar = 60120583m (C) Higher-magnification photomicrographs of the Otx2-IR cells in the optic(OP) layer Note oval- or round-shaped Otx2-IR cells IRelements are localized in the nucleus of the cell body Orientation of the images isdorsal to the top and lateral to the left Scale bar = 10120583m
mice (Figure 4(a)) The band of densely labeled small-to-medium-sized PV-IR cells in the superficial layers (SGRand OP) was also present in the KO mice (Figure 5(b)(A))Clusters of PV-IR cells surrounded by the patchy fiber in the
IGR were also visible in the KO mice (Figure 5(b)(B)) Wealso observed scattered medium-to-large PV-IR cells in theDLof theKOmouse SC (Figure 5(b)(C)) Tables 1 and 2 revealmean number of Otx2- and PV-IR cells in each layer of the
8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(a) Control (PV)
(A) Superficial layers
(C) Deep layers (DL)
(B) Deep layers (IGR)
(b) KO (PV)
Figure 5 High-magnification photomicrographs of the PV-labeled sections in the superficial layers (A) IGR (B) and DL (C) of the control(A) and KO (B) mouse SC Note a dense band in the deep SGR and the upper OP (open triangles) (A) clusters of PV-IR cells (arrows)and the intense PV-IR neuropils (open triangles) in the IGR (B) and scattered PV-IR cells (arrows) in the DL (C) in the control mouse (a)Similar patterns of distribution of PV-IR cells are also observed in the KO mice (b) but fewer PV-IR cells and less intense PV-IR neuropilsare observed Orientation of the images dorsal to the top and lateral to the left SGR superficial gray OP optic IGR intermediate gray layeris DL and the deep layers Scale bar = 60120583m
mouse SC Cell counts in each layer indicated that frequencydistributions of the labeled cells were similar in the controland KO mice (Tables 1 and 2)
Furthermore Figures 4(b)(C) and 7 show high-magnifi-cation photomicrographs of Otx2- and PV-IR cells in the KOmice respectively All the various morphological Otx2- andPV-IR cell types in the controlmice were also identified in theKOmouse SCThus Otx2 antibody labeling in the KOmouseSC revealed dark or light round to oval cells (Figure 4(b)(C))All the PV-IR cell types found in the SGR and OP in the
control mice were also present in the KO mice (Figure 7)Marginal cells with star-shaped cell bodies and few or nolabeled dendrites were commonly observed (a) and severalpyriform cells were also found in the KOmouse SGR and OP(b c) Vertical fusiform cells were also frequently seen in theSGR regions of the KO mice (d) Horizontal fusiform cellswere observed in the KO mouse OP (f) each having a hori-zontally elongated cell body and two stout dendrites coursingfrom the cell bodyThePV-IR cells in theKOmouse IGRwerelarge and clustered (Figure 5(b)(B)) just as in the control
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
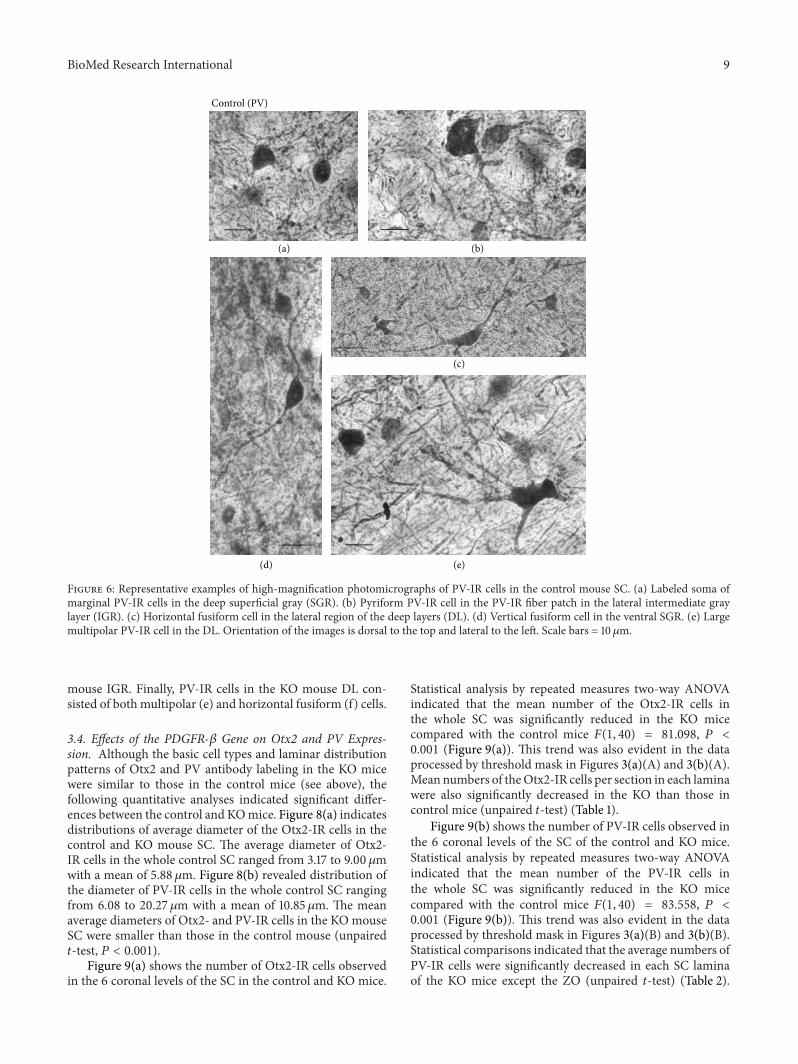
BioMed Research International 9
Control (PV)
(a) (b)
(c)
(d) (e)
Figure 6 Representative examples of high-magnification photomicrographs of PV-IR cells in the control mouse SC (a) Labeled soma ofmarginal PV-IR cells in the deep superficial gray (SGR) (b) Pyriform PV-IR cell in the PV-IR fiber patch in the lateral intermediate graylayer (IGR) (c) Horizontal fusiform cell in the lateral region of the deep layers (DL) (d) Vertical fusiform cell in the ventral SGR (e) Largemultipolar PV-IR cell in the DL Orientation of the images is dorsal to the top and lateral to the left Scale bars = 10120583m
mouse IGR Finally PV-IR cells in the KO mouse DL con-sisted of both multipolar (e) and horizontal fusiform (f) cells
34 Effects of the PDGFR-120573 Gene on Otx2 and PV Expres-sion Although the basic cell types and laminar distributionpatterns of Otx2 and PV antibody labeling in the KO micewere similar to those in the control mice (see above) thefollowing quantitative analyses indicated significant differ-ences between the control and KOmice Figure 8(a) indicatesdistributions of average diameter of the Otx2-IR cells in thecontrol and KO mouse SC The average diameter of Otx2-IR cells in the whole control SC ranged from 317 to 900120583mwith a mean of 588120583m Figure 8(b) revealed distribution ofthe diameter of PV-IR cells in the whole control SC rangingfrom 608 to 2027120583m with a mean of 1085 120583m The meanaverage diameters of Otx2- and PV-IR cells in the KOmouseSC were smaller than those in the control mouse (unpaired119905-test 119875 lt 0001)
Figure 9(a) shows the number of Otx2-IR cells observedin the 6 coronal levels of the SC in the control and KO mice
Statistical analysis by repeated measures two-way ANOVAindicated that the mean number of the Otx2-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 81098 119875 lt0001 (Figure 9(a)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(A) and 3(b)(A)Mean numbers of theOtx2-IR cells per section in each laminawere also significantly decreased in the KO than those incontrol mice (unpaired 119905-test) (Table 1)
Figure 9(b) shows the number of PV-IR cells observed inthe 6 coronal levels of the SC of the control and KO miceStatistical analysis by repeated measures two-way ANOVAindicated that the mean number of the PV-IR cells inthe whole SC was significantly reduced in the KO micecompared with the control mice 119865(1 40) = 83558 119875 lt0001 (Figure 9(b)) This trend was also evident in the dataprocessed by threshold mask in Figures 3(a)(B) and 3(b)(B)Statistical comparisons indicated that the average numbers ofPV-IR cells were significantly decreased in each SC laminaof the KO mice except the ZO (unpaired 119905-test) (Table 2)
10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
Table1Com
paris
onof
numbero
fOtx2-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO2248
2140
2462
2038
2233
23482245plusmn150
1538
1575
1608
15681572plusmn29
minus300
0001
SGR
3788
3587
3760
4010
3918
45423934plusmn331
2405
2712
2310
2393
2455plusmn176
minus376
0001
OP
3255
3857
3990
3303
3500
35473575plusmn295
2693
2800
2475
2610
2645plusmn137
minus252
0001
IGR
3162
3162
3020
2792
3285
2998
3070plusmn172
2147
2290
2207
21672203plusmn63
minus282
0001
DL
1898
1688
2582
1592
2687
27232195plusmn525
1763
1383
1470
1592
1552plusmn165
minus293
004
8Fo
reachlayero
fthe
SCthe
meannu
mbersof
cells
persectio
nin
each
anim
alareind
icatedIneach
anim
al6
sections
weree
xamined
Change
and119875valueind
icate
change
from
thec
ontro
lmicea
ndsta
tistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerZO
zon
allayerSG
Rsuperficialgray
layerOP
optic
layerIG
Rinterm
ediategray
layerandDLdeep
layer
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
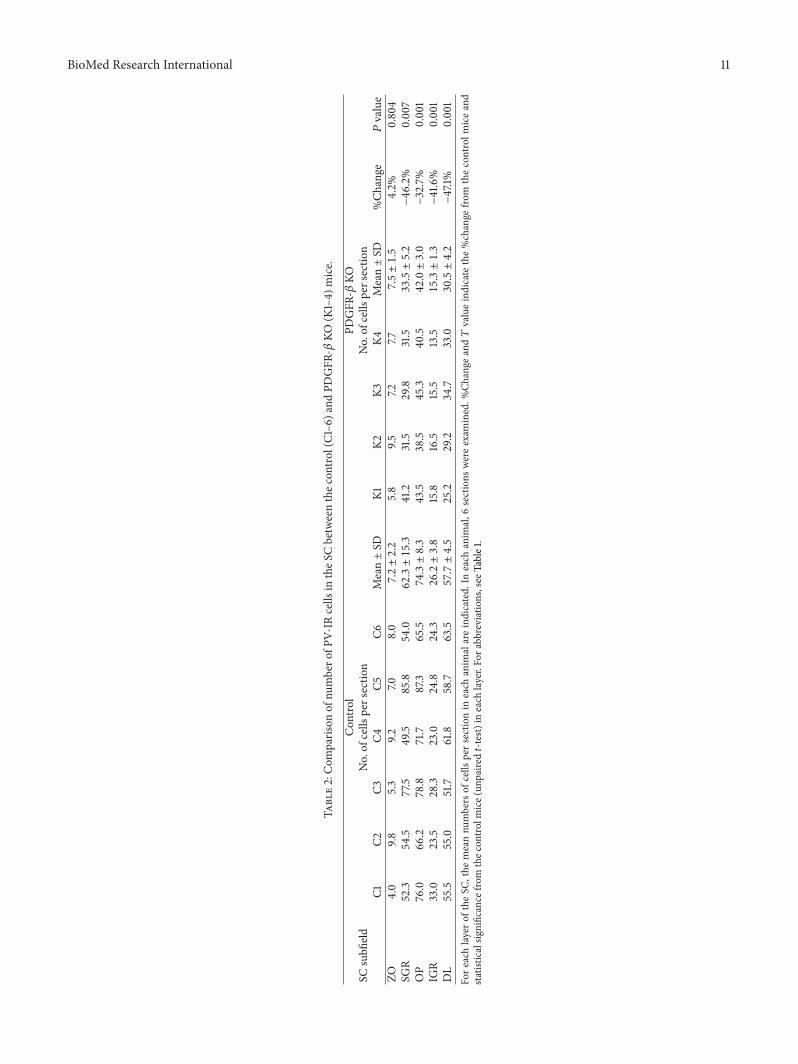
BioMed Research International 11
Table2Com
paris
onof
numbero
fPV-IR
cells
intheS
Cbetweenthec
ontro
l(C1ndash6
)and
PDGFR
-120573KO
(K1ndash4)
mice
SCsubfi
eldCon
trol
PDGFR
-120573KO
Noof
cells
persectio
nNoof
cells
persectio
nC1
C2C3
C4C5
C6MeanplusmnSD
K1K2
K3K4
MeanplusmnSD
Ch
ange
119875value
ZO40
9853
9270
80
72plusmn22
58
9572
7775plusmn15
42
0804
SGR
523
545
775
495
858
540
623plusmn153
412
315
298
315
335plusmn52
minus46
2
0007
OP
760
662
788
717
873
655
743plusmn83
435
385
453
405
420plusmn30
minus327
0001
IGR
330
235
283
230
248
243
262plusmn38
158
165
155
135
153plusmn13
minus416
0001
DL
555
550
517
618
587
635
577plusmn45
252
292
347
330
305plusmn42
minus471
0001
Fore
achlayero
fthe
SCthe
meannu
mbers
ofcells
pers
ectio
nin
each
anim
alareindicated
Ineach
anim
al6
sections
wereexam
ined
Change
and119879valueindicate
thechange
from
thecontrolm
iceand
statistic
alsig
nificance
from
thec
ontro
lmice(un
paire
d119905-te
st)in
each
layerFor
abbreviatio
nssee
Table1
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
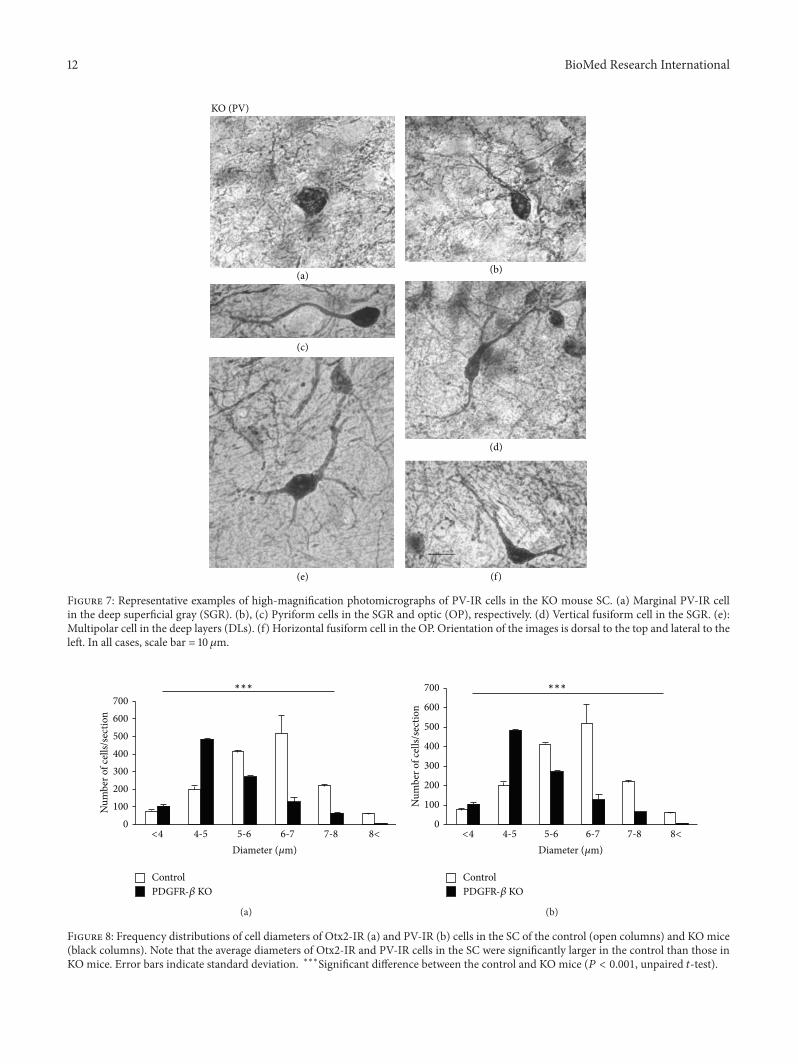
12 BioMed Research International
KO (PV)
(a) (b)
(c)
(d)
(e) (f)
Figure 7 Representative examples of high-magnification photomicrographs of PV-IR cells in the KO mouse SC (a) Marginal PV-IR cellin the deep superficial gray (SGR) (b) (c) Pyriform cells in the SGR and optic (OP) respectively (d) Vertical fusiform cell in the SGR (e)Multipolar cell in the deep layers (DLs) (f) Horizontal fusiform cell in the OP Orientation of the images is dorsal to the top and lateral to theleft In all cases scale bar = 10 120583m
Control
700600500400300200100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
PDGFR-120573 KO
(a)
700
600
500
400
300
200
100
0lt4 4-5 5-6 6-7 7-8 8lt
Num
ber o
f cel
lsse
ctio
n
Diameter (120583m)
lowastlowastlowast
ControlPDGFR-120573 KO
(b)
Figure 8 Frequency distributions of cell diameters of Otx2-IR (a) and PV-IR (b) cells in the SC of the control (open columns) and KOmice(black columns) Note that the average diameters of Otx2-IR and PV-IR cells in the SC were significantly larger in the control than those inKO mice Error bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 unpaired 119905-test)
BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 13
lowastlowastlowast
2500
2000
1500
1000
500
0
Num
ber o
f cel
lsse
ctio
n
minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
ControlPDGFR-120573 KO
(a)
lowastlowastlowast350
300
250
200
150
100
50
0minus33 minus362 minus394 minus426 minus458 minus49
Distance from the bregma (mm)
Num
ber o
f cel
lsse
ctio
n
ControlPDGFR-120573 KO
(b)
Figure 9 Comparison of mean number of Otx2-IR (a) and PV-IR (b) cells in the SC between the control (open columns) and KO mice(black columns) Each value represents the mean plusmn SEM Numbers below the abscissa indicate the rostrocaudal distance from the bregmaError bars indicate standard deviation lowastlowastlowastSignificant difference between the control and KO mice (119875 lt 0001 repeated measures two-wayANOVA)
Furthermore analyses by pixel intensity also revealed similarchanges (data not shown)
4 Discussion
The present study provides the first morphological andquantitative descriptions of Otx2-IR cells in the adult mouseSC Immunohistochemical observations of the mouse SCin the present study demonstrated that Otx2-IR cells formtwo main tiers (superficial and deep layers with dense andlight immunoreactivity resp) whereas PV-IR elements areconcentrated in a dense tier in the superficial layer whichrepresent a pattern of sublimation in the mouse SC Inaddition this study provides the first evidence for the role ofPDGFR-120573 in Otx2 and PV expression in the SC
41 Otx2 Expression in the SC The dense distribution ofOtx2-IR cells is observed in the superficial layers of themouse SC and less frequent distribution was also found inthe deep layers Presence of Otx2 expression in the postnatalSC is consistent with previous studies in rats [30 59] Inthe present experiment intensity of Otx2-IR labeling wasdifferent among the Otx2-IR cells Transcription factorsincluding Otx2 propagate along the visual pathway by trans-synaptic (cell-to-cell) transfer [30 60] and the SC receivesdirect afferents from the retina where the Otx2 protein isstrongly expressed [26 30 61] These findings suggest thatOtx2-immunoactivity in the Otx2-IR cells in the SC mightbe derived from the retinal ganglion cellsTherefore strengthof Otx2-immunoreactivity in individual SC neurons mightbe dependent on synaptic strength between the given SCneurons and retinal ganglion cells Furthermore a diversityof Otx2 proteins and thiermRNA isoforms has been reportedin the mouse brain [62]This finding also supports a diversity
of Otx2-immunoreactivity in the SC Further studies arerequired to clarify these phenomena
The Otx2 expression in the brain is involved in visualinformation processing at both prenatal and postnatal stages[25ndash29 63] and its role in differentiation of the retina [64 65]has been well established However the exact function ofOtx2 in the SC remains unclear The superficial layers ofthe SC receive visual information from the retina and visualcortex and are involved in visual information processing[1 4ndash6] while the deep SC layers play a role in integratingsensory information into motor signals that help to orientthe head and body toward various stimuli and in saccadiceye movements [1 4ndash6] On the other hand Otx2 is involvedin the positioning of the isthmic organizer and control ofneuronal subtype differentiation in the mid-brain duringembryogenesis [60 66 67] Furthermore Otx2 is implicatedin neural plasticity of PV-IR neurons in the postnatal visualcortex [30] and in axonal navigation and shaping neuronalarbors [60 68] These results suggest that Otx2 in theSC might be involved in formation andor maintenance ofprecise topographical neuronal circuits such as retinotopicorganization in the superficial layers [69 70] and in formationandor maintenance of neural circuits for spatially selectiveorientation of the body in the deep layers [71 72]
42 PV Expression in the SC PV-IR elements were rich inall collicular layers and superficial gray (SGR) was homo-geneously stained The staining is attributed to rich varicoseneuropils as well as a population of vertically oriented cellswith a similar shape which were more numerous in thedorsal optic (OP) than in the ventral SGR In contrast tothe staining pattern in the superficial layers the pattern ofimmunoreactivity in the IGR was clustered This patternappears as a row of patches with various holes of different
14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 BioMed Research International
sizes in which numerous marginal and multipolar cellsare mostly found The similar PV-immunolabeling patternsdescribed in the present study are also reported in therat SC [14] In the superficial layers of the SC a diffusesuperposition by both PV- and Otx2-IR substances wasobserved suggesting that some Otx2-IR cells coexpress PVprotein Further studies using double-labeled staining arerequired to confirm colocalization of Otx2- and PV-IR cellsin the adult mouse SC
In the SC about 50 of PV-IR cells were GABAergicand GABAergic neurons in the SC are small to medium insize (less than 15 120583m mean diameter) [73] These findingsare consistent with present studies in which PV-IR cellswith the same size were observed These GABAergic areinvolved in both intralaminar and interlaminar inhibition[74] Consistently electrical stimulation within the superficiallayers in the SC evoked inhibitory postsynaptic current [75]and projection from the superficial layers to deep layers inthe SC is under tonic inhibition by GABAergic activity [74]These GABAergic circuits are suggested to play a role inshaping receptive field inhibitory surrounds as well as visualresponse habituation [76] and sensorimotor processing fororientation [74] Furthermore PV-IR interneurons with fastspiking are involved in gamma oscillation [77 78] whichis induced in the SC by visual stimulation [79 80] Thesefindings suggest that PV-IR cells are involved in gammaoscillation in the mouse SC
On the other hand the remaining half of the PV-IRcells in the SC might be excitatory and colocalize withglutamate [73 81] These PV-IR excitatory neurons couldbe excitatory projection neurons with large cell size [81] orlocal interneurons in the excitatory networks to generatehigh-frequency burst activity in premotor neurons in theintermediate layers [73 82] since these premotor neuronsin the intermediate layers have no intrinsic mechanisms tosupport high-frequency burstic activity [6]
43 Role of PDGFR-120573 in PV and Otx2 Expression the SCPDGF-B chain together with expression of PDGFR-120573 canwidely exert a neurotrophic or regulatory activity on neuronsin an autocrine manner or by neuron to neuron interactionsin the brain [83] In the present study we observed significantreductions in Otx2- and PV-IR cells in the SC of the KOmice Furthermore cell sizes of Otx2- and PV-IR cells weresignificantly reduced in the SC Since Otx2-immunoreactiveelements are usually localized in the nucleus of the cellbody decrease in Otx2-IR cell size might reflect decreasein expression of Otx2 protein These findings suggest thatPDGFR-120573 exerts substantial effects on expression ofOtx2 andPV in the SC
Since PDGF-BPDGFR-120573 signal axis exerts neurotrophiceffects on GABAergic neurons [42 45] a decrease in thenumber of PV-IR cells might be attributed to the loss ofneurotrophic effects of PDGFR-120573 in the PDGFR-120573 KOmiceFurthermore Otx2 and PDGF-BPDGFR-120573 exert similareffects on neurons both Otx2 and PDGF-B exert neu-rotrophic effects on dopaminergic neurons [67 68 84] areinvolved in maturation of GABAergic neurons [30] or exert
neurotrophic effects on GABAergic neurons [42 45] and arealso involved in neurogenesis andmigration of some neurons[51 67 85] These results suggest that effects of PDGFR-120573 might be partially mediated through Otx2 Consistentwith this idea Otx2 expression is controlled by neurotrophicfactors such as basic fibroblast growth factor (bFGF) [86]and PDGFR-120573 signals are indispensable for the bFGF signals[51] The present results along with these previous findingssuggest that PDGF-B might directly affect development ofPV-IR cells andor indirectly affect development of PV-IRcells through its effects on Otx2 Further studies are requiredto elucidate signal transduction pathways among PDGFR-120573Otx2 and PV
5 Conclusions
The present study demonstrates the distribution patterns ofOtx2- and PV-IR cells in the mouse SC Both the Otx2-IRcells and PV-IR elements revealed characteristic patternsThelabeled cells were heterogeneous in diameter Although thesecharacteristic patterns did not differ between the control andPDGFR-120573 KO mice mean numbers of the Otx2- and PV-IRcells were significantly decreased in the PDGFR-120573 KO miceFurthermore Otx2- and PV-IR cell sizes were significantlysmaller in the PDGFR-120573KOmiceThese findings suggest thatPDGFR-120573 KO induces development and maturation deficitsin Otx2- and PV-IR cells in the SC These results furthersuggest that behavioral traits (socioemotional deficits) ofPDGFR-120573 KO mice [48] might be ascribed partly to deficitsin PV-positive GABAergic neurons in the SC of PDGFR-120573KOmiceThis idea is supported by previous studies bilateralSC lesions reduced social behaviors [7 8] and reduction ofnumber of PV-containing GABAergic neuronsin the SC isassociated with autism and schizophrenia with social deficits[17ndash19] Activity in the subcortical visual pathway includingthe SC was altered in autistic patients [87] Further studiesare required to elucidate molecular mechanisms of the role ofPDGFR-120573 in survival and induction of Otx2 and PV proteins
Acknowledgments
This work was supported partly by JSPS Asian Core Programand the Ministry of Education Science Sports and CultureGrant-in-Aid for Scientific Research (B) (25290005)
References
[1] M F Huerta A Frankfurter and J K Harting ldquoStudiesof the principal sensory and spinal trigeminal nuclei of therat projections to the superior colliculus inferior olive andcerebellumrdquoThe Journal of Comparative Neurology vol 220 no2 pp 147ndash167 1983
[2] J K Harting W C Hall I T Diamond and G F MartinldquoAnterograde degeneration study of the superior colliculus inTupaia glis evidence for a subdivision between superficial anddeep layersrdquoThe Journal of Comparative Neurology vol 148 no3 pp 361ndash386 1973
[3] S B Edwards C L Ginsburgh C K Henkel and B E SteinldquoSources of subcortical projections to the superior colliculus in
BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 15
the catrdquo The Journal of Comparative Neurology vol 184 no 2pp 309ndash330 1979
[4] RGrantyn ldquoGaze control through superior colliculus structureand functionrdquo Reviews of Oculomotor Research vol 2 pp 273ndash333 1988
[5] J K Harting M F Huerta T Hashikawa and D P vanLieshout ldquoProjection of the mammalian superior colliculusupon the dorsal lateral geniculate nucleus organization oftectogeniculate pathways in nineteen speciesrdquo The Journal ofComparative Neurology vol 304 no 2 pp 275ndash306 1991
[6] T Isa ldquoIntrinsic processing in the mammalian superior collicu-lusrdquoCurrentOpinion inNeurobiology vol 12 no 6 pp 668ndash6772002
[7] R S Maior E Hori C E Uribe et al ldquoA role for the superiorcolliculus in the modulation of threat responsiveness in pri-mates towards the ontogenesis of the social brainrdquo Reviews inthe Neurosciences vol 22 pp 697ndash706 2012
[8] MNNguyen EHori JMatsumoto AH Tran TOno andHNishijo ldquoNeuronal responses to face-like stimuli in the monkeypulvinarrdquoThe European Journal of Neuroscience vol 37 pp 35ndash51 2013
[9] V A Casagrande ldquoA third parallel visual pathway to primateareaV1rdquoTrends inNeurosciences vol 17 no 7 pp 305ndash310 1994
[10] I T Diamond D Fitzpatrick and D Schmechel ldquoCalciumbinding proteins distinguish large and small cells of the ventralposterior and lateral geniculate nuclei of the prosimian galagoand the tree shrew (Tupaia belangeri)rdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 4 pp 1425ndash1429 1993
[11] J K Johnson and V A Casagrande ldquoDistribution of calcium-binding proteinswithin the parallel visual pathways of a primate(Galago crassicaudatus)rdquoThe Journal of ComparativeNeurologyvol 356 no 2 pp 238ndash260 1995
[12] R R Mize C-J Jeon G D Butler Q Luo and P C EmsonldquoThe calcium binding protein calbindin-D 28K reveals sub-populations of projection and interneurons in the cat superiorcolliculusrdquo The Journal of Comparative Neurology vol 307 no3 pp 417ndash436 1991
[13] D M Berson and J T McIlwain ldquoRetinal Y-cell activation ofdeep-layer cells in superior colliculus of the catrdquo Journal ofNeurophysiology vol 47 no 4 pp 700ndash714 1982
[14] R-B Illing ldquoAssociation of efferent neurons to the compart-mental architecture of the superior colliculusrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol89 no 22 pp 10900ndash10904 1992
[15] P S Chard D Bleakman S Christakos C S Fullmer and RJ Miller ldquoCalcium buffering properties of calbindin D28k andparvalbumin in rat sensory neuronesrdquoThe Journal of Physiologyvol 472 pp 341ndash357 1993
[16] J Du L Zhang M Weiser B Rudy and C J McBainldquoDevelopmental expression and functional characterizationof the potassium-channel subunit Kv31b in parvalbumin-containing interneurons of the rat hippocampusrdquo The Journalof Neuroscience vol 16 no 2 pp 506ndash518 1996
[17] D A Lewis T Hashimoto and DW Volk ldquoCortical inhibitoryneurons and schizophreniardquo Nature Reviews Neuroscience vol6 no 4 pp 312ndash324 2005
[18] D C Rojas K Maharajh P Teale and S J Rogers ldquoReducedneural synchronization of gamma-band MEG oscillations infirst-degree relatives of children with autismrdquo BMC Psychiatryvol 8 article 66 2008
[19] G Dendrinos M Hemelt and A Keller ldquoPrenatal VPA expo-sure and changes in sensory processing by the superior collicu-lusrdquo Frontiers in Integrative Neuroscience vol 5 article 68 2011
[20] R Finkelstein D Smouse T M Capaci A C Spradlingand N Perrimon ldquoThe orthodenticle gene encodes a novelhomeo domain protein involved in the development of theDrosophila nervous system and ocellar visual structuresrdquoGenesamp Development vol 4 no 9 pp 1516ndash1527 1990
[21] A Simeone D Acampora M Gulisano A Stornaiuolo andE Boncinelli ldquoNested expression domains of four homeoboxgenes in developing rostral brainrdquo Nature vol 358 no 6388pp 687ndash690 1992
[22] A Simeone D Acampora A Mallamaci et al ldquoA vertebrategene related to orthodenticle contains a homeodomain of thebicoid class and demarcates anterior neuroectoderm in thegastrulating mouse embryordquo The EMBO Journal vol 12 no 7pp 2735ndash2747 1993
[23] A Simeone E Puelles and D Acampora ldquoThe Otx familyrdquoCurrent Opinion in Genetics and Development vol 12 no 4 pp409ndash415 2002
[24] D Arendt ldquoEvolution of eyes and photoreceptor cell typesrdquoInternational Journal of Developmental Biology vol 47 no 7-8pp 563ndash571 2003
[25] P Bovolenta A Mallamaci P Briata G Corte and EBoncinelli ldquoImplication of OTX2 in pigment epithelium deter-mination and neural retina differentiationrdquo The Journal ofNeuroscience vol 17 no 11 pp 4243ndash4252 1997
[26] D Baas K M Bumsted J A Martinez F M Vaccarino KC Wikler and C J Barnstable ldquoThe subcellular localization ofOTX2 is cell-type specific and developmentally regulated in themouse retinardquoMolecular Brain Research vol 78 no 1-2 pp 26ndash37 2000
[27] J R Martinez-Morales M Signore D Acampora A Simeoneand P Bovolenta ldquoOtx genes are required for tissue specificationin the developing eyerdquo Development vol 128 no 11 pp 2019ndash2030 2001
[28] A S Viczian R Vignali M E Zuber G Barsacchi and W AHarris ldquoXOtx5b and XOtx2 regulate photoreceptor and bipolarfates in the Xenopus retinardquo Development vol 130 no 7 pp1281ndash1294 2003
[29] S Sakami O Hisatomi S Sakakibara J Liu T A Reh andF Tokunaga ldquoDownregulation of Otx2 in the dedifferentiatedRPE cells of regenerating newt retinardquo Developmental BrainResearch vol 155 no 1 pp 49ndash59 2005
[30] S Sugiyama A A di Nardo S Aizawa et al ldquoExperience-dependent transfer of Otx2 homeoprotein into the visual cortexactivates postnatal plasticityrdquo Cell vol 134 no 3 pp 508ndash5202008
[31] F Nothias G Fishell and A Ruiz i Altaba ldquoCooperation ofintrinsic and extrinsic signals in the elaboration of regionalidentity in the posterior cerebral cortexrdquo Current Biology vol8 no 8 pp 459ndash462 1998
[32] R Ross J Glomset B Kariya and L Harker ldquoA plateletdependent serum factor that stimulates the proliferation ofarterial smoothmuscle cells in vitrordquoProceedings of theNationalAcademy of Sciences of the United States of America vol 71 no4 pp 1207ndash1210 1974
[33] A Sasahara J N Kott M Sasahara E W Raines R Rossand L E Westrum ldquoPlatelet-derived growth factor B-chain-like immunoreactivity in the developing and adult rat brainrdquoDevelopmental Brain Research vol 68 no 1 pp 41ndash53 1992
16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

16 BioMed Research International
[34] J Krupinski R Issa T Bujny et al ldquoA putative role for platelet-derived growth factor in anglogenesis and neuroprotection afterischemic stroke in humansrdquo Stroke vol 28 no 3 pp 564ndash5731997
[35] K Iihara N Hashimoto T TsukaharaM Sakata H Yanamotoand T Taniguchi ldquoPlatelet-derived growth factor-BB but not -AA prevents delayed neuronal death after forebrain ischemia inratsrdquo Journal of Cerebral Blood Flow andMetabolism vol 17 no10 pp 1097ndash1106 1997
[36] M Ohno M Sasahara S Narumiya et al ldquoExpressionof platelet-derived growth factor B-chain and 120573-receptor inhypoxicischemic encephalopathy of neonatal ratsrdquo Neuro-science vol 90 no 2 pp 643ndash651 1999
[37] DA Rappolee C A Brenner R Schultz DMark andZWerbldquoDevelopmental expression of PDGF TGF-120572 and TGF-120573 genesin preimplantation mouse embryosrdquo Science vol 241 no 4874pp 1823ndash1825 1988
[38] M Mercola D A Melton and C D Stiles ldquoPlatelet-derivedgrowth factor A chain is maternally encoded in Xenopusembryosrdquo Science vol 241 no 4870 pp 1223ndash1225 1988
[39] A S Goustin C Betsholtz S Pfeifer-Ohlsson et al ldquoCoexpres-sion of the sis and myc proto-oncogenes in developing humanplacenta suggests autocrine control of trophoblast growthrdquoCellvol 41 no 1 pp 301ndash312 1985
[40] L Fredriksson H Li and U Eriksson ldquoThe PDGF family fourgene products formfive dimeric isoformsrdquoCytokine andGrowthFactor Reviews vol 15 no 4 pp 197ndash204 2004
[41] C-H Heldin A Ostman and L Ronnstrand ldquoSignal transduc-tion via platelet-derived growth factor receptorsrdquo Biochimica etBiophysica Acta vol 1378 no 1 pp F79ndashF113 1998
[42] A Smits M Kato B Westermark M Nister C-H Heldinand K Funa ldquoNeurotrophic activity of platelet-derived growthfactor (PDGF) rat neuronal cells possess functional PDGF120573-type receptors and respond to PDGFrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 88 no 18 pp 8159ndash8163 1991
[43] M Sasahara JWU Fries EW Raines et al ldquoPDGFB-chain inneurons of the central nervous system posterior pituitary andin a transgenic modelrdquo Cell vol 64 no 1 pp 217ndash227 1991
[44] M Sasahara S Amano H Sato et al ldquoNormal developing ratbrain expresses a platelet-derived growth factor B chain (c-sis)mRNA truncated at the 51015840 endrdquo Oncogene vol 16 no 12 pp1571ndash1578 1998
[45] A Smits A E Ballagi and K Funa ldquoPDGF-BB exerts trophicactivity on cultured GABA interneurons from the newborn ratcerebellumrdquo European Journal of Neuroscience vol 5 no 8 pp986ndash994 1993
[46] C-H Heldin and B Westermark ldquoMechanism of action andin vivo role of platelet-derived growth factorrdquo PhysiologicalReviews vol 79 no 4 pp 1283ndash1316 1999
[47] J Andrae R Gallini and C Betsholtz ldquoRole of platelet-derived growth factors in physiology and medicinerdquo Genes ampDevelopment vol 22 no 10 pp 1276ndash1312 2008
[48] P T H Nguyen T Nakamura E Hori et al ldquoCognitiveand socio-emotional deficits in platelet-derived growth factorreceptor-120573 gene knockoutmicerdquo PLoSONE vol 6 no 3 ArticleID e18004 2011
[49] Z Gao T Sasaoka T Fujimori et al ldquoDeletion of the PDGFR-120573 gene affects key fibroblast functions important for woundhealingrdquo Journal of Biological Chemistry vol 280 no 10 pp9375ndash9389 2005
[50] Y Ishii T Oya L Zheng et al ldquoMouse brains deficient inneuronal PDGF receptor-120573 develop normally but are vulnerableto injuryrdquo Journal of Neurochemistry vol 98 no 2 pp 588ndash6002006
[51] Y Ishii Y Matsumoto R Watanabe et al ldquoCharacterization ofneuroprogenitor cells expressing the PDGF 120573-receptor withinthe subventricular zone of postnatal micerdquo Molecular andCellular Neuroscience vol 37 no 3 pp 507ndash518 2008
[52] R R Mize and Q Luo ldquoVisual deprivation fails to reducecalbindin 28kD or GABA immunoreactivity in the rhesusmonkey superior colliculusrdquo Visual Neuroscience vol 9 no 2pp 157ndash168 1992
[53] M Behan A Jourdain and G M Bray ldquoCalcium bindingprotein (calbindin D28k) immunoreactivity in the hamstersuperior colliculus ultrastructure and lack of co-localizationwithGABArdquoExperimental Brain Research vol 89 no 1 pp 115ndash124 1992
[54] M Abercrombie ldquoEstimation of nuclear population frommicrotome sectionsrdquo The Anatomical Record vol 94 pp 239ndash247 1946
[55] J C Hedreen ldquoWhat was wrong with the Abercrombie andempirical cell counting methods A reviewrdquo The AnatomicalRecord vol 250 no 3 pp 373ndash380 1998
[56] T P Langer and R D Lund ldquoThe upper layers of the superiorcolliculus of the rat a Golgi studyrdquoThe Journal of ComparativeNeurology vol 158 no 4 pp 418ndash435 1974
[57] A R Labriola and L K Laemle ldquoCellular morphology inthe visual layer of the developing rat superior colliculusrdquoExperimental Neurology vol 55 no 1 pp 247ndash268 1977
[58] K B Larsen M C Lutterodt K Moslashllgard and M MoslashllerldquoExpression of the homeobox genes OTX2 and OTX1 in theearly developing human brainrdquo Journal of Histochemistry andCytochemistry vol 58 no 7 pp 669ndash678 2010
[59] M F Rath E Munoz S Ganguly et al ldquoExpression of theOtx2 homeobox gene in the developing mammalian brainembryonic and adult expression in the pineal glandrdquo Journal ofNeurochemistry vol 97 no 2 pp 556ndash566 2006
[60] A Prochiantz and A Joliot ldquoCan transcription factors functionas cell-cell signallingmoleculesrdquoNature ReviewsMolecular CellBiology vol 4 no 10 pp 814ndash819 2003
[61] F Beby M Housset N Fossat et al ldquoOtx2 gene deletion inadult mouse retina induces rapid RPE dystrophy and slowphotoreceptor degenerationrdquo PLoS ONE vol 5 no 7 Article IDe11673 2010
[62] V Courtois G Chatelain Z-Y Han N le Novere G Brunand T Lamonerie ldquoNewOtx2mRNA isoforms expressed in themouse brainrdquo Journal of Neurochemistry vol 84 no 4 pp 840ndash853 2003
[63] A Nishida A Furukawa C Koike et al ldquoOtx2 homeoboxgene controls retinal photoreceptor cell fate and pineal glanddevelopmentrdquoNature Neuroscience vol 6 no 12 pp 1255ndash12632003
[64] M E Zuber G Gestri A S Viczian G Barsacchi and W AHarris ldquoSpecification of the vertebrate eye by a network of eyefield transcription factorsrdquo Development vol 130 no 21 pp5155ndash5167 2003
[65] T Akagi M Mandai S Ooto et al ldquoOtx2 homeobox geneinduces photoreceptor-specific phenotypes in cells derivedfrom adult iris and ciliary tissuerdquo Investigative Ophthalmologyand Visual Science vol 45 no 12 pp 4570ndash4575 2004
BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 17
[66] E Puelles A Annino F Tuorto et al ldquoOtx2 regulates the extentidentity and fate of neuronal progenitor domains in the ventralmidbrainrdquo Development vol 131 no 9 pp 2037ndash2048 2004
[67] B Vernay M Koch F Vaccarino et al ldquoOtx2 regulates subtypespecification and neurogenesis in the midbrainrdquoThe Journal ofNeuroscience vol 25 no 19 pp 4856ndash4867 2005
[68] C Y Chung P Licznerski K N Alavian et al ldquoThe transcrip-tion factor orthodenticle homeobox 2 influences axonal pro-jections and vulnerability of midbrain dopaminergic neuronsrdquoBrain vol 133 part 7 pp 2022ndash2031 2010
[69] M Cynader and N Berman ldquoReceptive-field organization ofmonkey superior colliculusrdquo Journal of Neurophysiology vol 35no 2 pp 187ndash201 1972
[70] U C Drager and D H Hubel ldquoResponses to visual stimulationand relationship between visual auditory and somatosensoryinputs inmouse superior colliculusrdquo Journal ofNeurophysiologyvol 38 no 3 pp 690ndash713 1975
[71] L R Burnett B E Stein D Chaponis and M T WallaceldquoSuperior colliculus lesions preferentially disrupt multisensoryorientationrdquo Neuroscience vol 124 no 3 pp 535ndash547 2004
[72] S D Crish CMDengler-Crish andCM Comer ldquoPopulationcoding strategies and involvement of the superior colliculus inthe tactile orienting behavior of nakedmole-ratsrdquoNeurosciencevol 139 no 4 pp 1461ndash1466 2006
[73] M BehanK Steinhacker S Jeffrey-Borger andMAMeredithldquoChemoarchitecture of GABAergic neurons in the ferret supe-rior colliculusrdquoThe Journal of Comparative Neurology vol 452no 4 pp 334ndash359 2002
[74] T Isa andWCHall ldquoExploring the superior colliculus in vitrordquoJournal of Neurophysiology vol 102 no 5 pp 2581ndash2593 2009
[75] S A Neale and T E Salt ldquoModulation of GABAergic inhi-bition in the rat superior colliculus by a presynaptic group IImetabotropic glutamate receptorrdquoThe Journal of Physiology vol577 no 2 pp 659ndash669 2006
[76] K E Binns and T E Salt ldquoDifferent roles for GABA(A) andGABA(B) receptors in visual processing in the rat superiorcolliculusrdquo The Journal of Physiology vol 504 part 3 pp 629ndash639 1997
[77] E C Fuchs A R Zivkovic M O Cunningham et al ldquoRecruit-ment of parvalbumin-positive interneurons determines hip-pocampal function and associated behaviorrdquo Neuron vol 53no 4 pp 591ndash604 2007
[78] V S Sohal F Zhang O Yizhar and K Deisseroth ldquoParval-bumin neurons and gamma rhythms enhance cortical circuitperformancerdquo Nature vol 459 no 7247 pp 698ndash702 2009
[79] M Breche R GoebelW Singer andA K Engel ldquoSynchroniza-tion of visual responses in the superior colliculus of awake catsrdquoNeuroReport vol 12 no 1 pp 43ndash47 2001
[80] Q Pauluis S N Baker and E Olivier ldquoPrecise burst synchronyin the superior colliculus of the awake cat during movingstimulus presentationrdquo The Journal of Neuroscience vol 21 no2 pp 615ndash627 2001
[81] R R Mize ldquoNeurochemical microcircuitry underlying visualand oculomotor function in the cat superior colliculusrdquoProgressin Brain Research vol 112 pp 35ndash55 1996
[82] T P Trappenberg M C Dorris D P Munoz and R M KleinldquoA model of saccade initiation based on the competitive inte-gration of exogenous and endogenous signals in the superiorcolliculusrdquo Journal of Cognitive Neuroscience vol 13 no 2 pp256ndash271 2001
[83] M Sasahara H Sato K Iihara et al ldquoExpression of platelet-derived growth factor B-chain in the mature rat brain andpituitary glandrdquoMolecular Brain Research vol 32 no 1 pp 63ndash74 1995
[84] A Othberg P Odin A Ballagi A Ahgren K Funa and OLindvall ldquoSpecific effects of platelet derived growth factor(PDGF) on fetal rat and human dopaminergic neurons in vitrordquoExperimental Brain Research vol 105 no 1 pp 111ndash122 1995
[85] F Inverardi M S Beolchi B Ortino et al ldquoGABA immunore-activity in the developing rat thalamus and Otx2 homeoproteinexpression in migrating neuronsrdquo Brain Research Bulletin vol73 no 1ndash3 pp 64ndash74 2007
[86] L Robel M Ding A J James et al ldquoFibroblast growth factor 2increases Otx2 expression in precursor cells from mammaliantelencephalonrdquo The Journal of Neuroscience vol 15 no 12 pp7879ndash7891 1995
[87] N M Kleinhans T Richards L C Johnson et al ldquoFMRI evi-dence of neural abnormalities in the subcortical face processingsystem in ASDrdquo NeuroImage vol 54 no 1 pp 697ndash704 2011
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology