restoration of the subepidermal tissues and scale regeneration after wounding a cichlid...
TRANSCRIPT

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 281:305–327 (1998)
© 1998 WILEY-LISS, INC.
Restoration of the Subepidermal Tissues and ScaleRegeneration After Wounding a Cichlid Fish,Hemichromis bimaculatus
ALEXANDRA QUILHAC AND JEAN-YVES SIRE*Université Paris 7 - Denis Diderot, CNRS URA 1137, France
ABSTRACT Experimental scale regeneration in fish is used as a means to study the epidermal-dermal interactions controlling organogenesis. Scales and epidermis were removed from a largesurface area (1 cm2) of the flank in the cichlid Hemichromis bimaculatus, and scale regeneration andrestoration of the subepidermal tissues were studied using light and electron microscopy. In addi-tion, cell proliferation in these regenerating dermal tissues was studied using tritiated thymidineincorporation. The original squamation pattern was entirely restored in the wound region, but adelay was observed in the central region compared to the peripheral regions in which normal regen-eration had occurred. This delay was the consequence of the osmotic shock in wound regions thatwere exposed to the external environment for a long time because of the late covering of the woundby the healing epidermis. However, the osmotic shock was not as severe as expected because twomeans of protection of the skin are involved: the scale-pocket lining (SPL) cells function as a barrierresisting the osmotic shock, and an exudate produced by the wounded tissues inhibits water pen-etration. Scale regeneration was initiated in each scale pocket where the healing epidermis enteredin contact with the SPL cells, and it proceeded centripetally from the margins to the center of thewound, as did the healing of the epidermis. The shape and size of the regenerated scales reflectedthis centripetal re-epithelialization, suggesting that the first signal for scale regeneration came fromthe epidermis. In the peripheral regions, which were rapidly protected from osmotic shock, the scalesregenerated in a few days, whereas in the central regions, the dermis had to be restored beforescales were able to regenerate, approximately 14 days after surgery. Healing of the dermis involvedclassical processes including migration, and proliferation of fibroblast cells, followed by extracellularmatrix deposition (mainly collagen). The regenerated scales formed in the central region of the wounddiffered from normal in that new scale-forming cells had to be recruited from the unwounded sur-rounding regions of the dermis, because of destruction of the SPL cell populations. J. Exp. Zool.281:305–327, 1998. © 1998 Wiley-Liss, Inc.
*Correspondence to: Dr. Jean-Yves Sire, Université Paris 7,Laboratoire d’Anatomie Comparée, Case 7077, 2, place Jussieu, 75251Paris cedex 05, France. E-mail: [email protected]
Received 8 October 1997; Accepted 19 February 1998
Organogenesis is largely regulated by inductivetissue interactions that take place mostly betweenepithelial and mesenchymal tissues (see reviewin Thesleff et al., ’95). In fish, such tissue inter-actions are involved during development of thedermal skeleton (Smith and Hall, ’90, ’93; Aki-menko et al., ’95). The numerous data availableon the structure, development, and regenerationof the scales in teleost fish strongly suggest thatepidermal-dermal interactions are involved at thetime of scale morphogenesis, differentiation, andgrowth (e.g., Sire and Géraudie, ’83, ’84; Sire, ’88,’89; Sire et al., ’90). We have chosen fish scales asa model to study the tissue interactions control-ling scale development and regeneration. Fishscales have numerous advantages mainly becauseof the facilities they present for in vivo experi-mentations (Sire, ’87). In normal conditions (scaleontogeny and regeneration), the developmental
events take place quickly in such a way that it isdifficult to understand the roles played by the epi-thelial and the mesenchymal cells. By removingscales and epidermis of a large area (1 cm2) of theskin in the cichlid Hemichromis bimaculatus, thedevelopmental processes leading to scale regen-eration can be slowed, permitting (1) descriptionof the dynamics of re-epithelialization with par-ticular focus on the differentiation of the epider-mal basal layer cells, and (2) the acquisition ofknowledge of the manner in which the subepider-mal tissues are restored and scale regenerationis allowed.
The dynamics of the re-epithelialization of the

306 A. QUILHAC AND J.-Y. SIRE
wound surface of the cichlid skin—from the timeof wounding until complete restoration of the epi-dermis—has recently been described (Quilhac andSire, submitted) and may be summarized as fol-lows: Re-epithelialization of the large wound oc-curred quickly. The 1 cm2 surface was completelycovered by the healing epidermis within a periodof 9 h, and the epidermis was completely restoredapproximately 4 days after surgery. Re-epithelial-ization involves a well-defined, repetitive sequenceof events including proliferation, migration, spread-ing, attachment, and differentiation. During theprocess of re-epithelialization, relationships are pro-gressively established between the differentiatingepidermal basal cells and the dermis below,strongly suggesting that interactive processes aretaking place.
Our goals were (1) to find indications of inter-actions between the differentiated basal epider-mal cells and the cells regenerating the scales orrestoring the dermis, and (2) to estimate the ef-fects of such a wound on scale regeneration in timeand space. In addition, this experiment could gen-erate data about how fish skin is able to resistthe adverse effects of the external milieu whensuch a large wound occurs. In the present paper,we describe the events (e.g., differentiation andproliferation) that occur in the subepidermal tis-sues from the time of wounding to the time of scaleregeneration, using light and transmission elec-tron microscopy and autoradiography after incor-poration of tritiated thymidine.
MATERIALS AND METHODS
AnimalsOne year-old adult specimens (46 to 70 mm
standard length, SL) of the African cichlid Hemi-chromis bimaculatus were used. Fish were bredat a constant temperature of 25 ± 1°C with anartificial 12-h light period and fed daily withChironomus larvae and Tetramin powder. Fifty-six fish were used for the experiment (two speci-mens for each stage studied): 10 specimens forthe reconstitution of the scale pattern, 26 speci-mens for histological and ultrastructural studies.and 20 specimens for autoradiography.
MethodsFish were anaesthetized by immersion in a so-
lution of 0.002% MS 222 (tricaine methane sul-phonate, Sandoz, Basel, Switzerland). Both inexperimental and control animals, adjacent scaleswere removed from their pockets on a 1 cm2 area
of the pectoral region (Figs. 1, 2). In the experi-mental fish, the remaining dermal strips support-ing the epidermal remnants were cut through toremove all the epidermal cells from the woundsurface, and this was consistently done in thesame way for each pocket to avoid variation inthe regeneration process (Fig. 2c). In a 70-mm-SL specimen, such a wound involved 35 scales,representing five rows (I to V) and seven columns(1 to 7) (Fig. 3). After the operation, each fish wasisolated in a 10-liter tank, without addition of an-tibiotics, and allowed to regenerate for the appro-priate time.
Reconstitution of the squamation patternsControl fish with scale removal only. The scales
regenerate rapidly from a thin layer of cells calledthe scale-pocket lining (SPL), which remains atthe deep surface of the scale pockets after scaleremoval (Fig. 2b). The scale matrix starts to bedeposited after 2 days; the regenerating scales canbe removed from their pocket after 5 days; andthey reach the same surface as a normal scaleafter 14 days (see also Sire, ’89). The control fishwere again anaesthetized, and the regeneratedsquamation patterns were reconstructed after 7days (one specimen) and 14 days (two specimens):the regenerated scales were carefully removedfrom their pockets using fine forceps and wereplaced on a glass slide with respect to their origi-nal position in the skin.
Experimental fish. The experimental fish wereanaesthetized at the appropriate time of regen-eration (7 days, three specimens; 14 days, threespecimens; and 30 days, one specimen). All theregenerated scales were carefully removed fromthe area of the operation, and the squamation pat-terns were reconstructed as described above.
For both control and experimental animals, eachsquamation pattern is composed of five rows andseven columns delimited on each side by a row ofnormal scales (Fig. 3). The observations and pho-tographs of the squamation patterns were facili-
Fig. 1. Left flank of the cichlid fish Hemichromis bimacu-latus showing the experimental zone. The double arrow indi-cates the orientation of the sections. Scale bar = 1 cm.

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 307
tated by staining the scales for 2 h in a solutionof 0.1% alizarin red S in 1% KOH.
Histology and TEMThe entire operated regions, plus a row of nor-
mal scales at the margins (i.e., a piece of skinapprox. 1 cm2), were carefully dissected at the fol-lowing intervals: 0, 3, 6, 9, 12 h and 1, 2, 3, 4, 5,7, 10, and 14 days after the operation. Sampleswere fixed for 2 h at room temperature in a solu-tion of 1.5% glutaraldehyde and 1.5% paraform-aldehyde in 0.1 M cacodylate buffer (pH 7.4) anddecalcified for 7 days at 4°C in the fixative solu-tion to which 0.1 M EDTA was added. The sampleswere washed with the same buffer containing 10%sucrose, postfixed for 2 h at room temperature in
1% osmium tetroxide, then dehydrated in a gradedseries of ethanol, embedded in epon, and sec-tioned. One µm-thick longitudinal sections werestained with toluidine blue. Thin sections (ca. 80nm) were contrasted with uranyl acetate and leadcitrate and examined under a Philips 201 EM op-erating at 80 kV.
AutoradiographyFor each chosen time of regeneration (i.e., 3, 6,
9, 12 h and 1, 2, 3, 4, 7 days), 20 µCi tritiatedthymidine (i.e., 5 µCi/g wet body weight, specificactivity: 4 Ci/mM, Amersham, Les Ulis, France)were injected into the skin immediately below theoperated zone, three hours before sacrifice.
In order to search for proliferation sites in theunwounded epidermis, two unwounded specimenswere injected in the same region following thesame procedure and used as controls.
The operated and the control zones were dis-sected, and the skin samples were fixed, dehy-drated, and embedded in epon as described above.Semi-thin sections (2 µm-thick) were mounted onslides, dried, then covered with Ilford K5 nuclearemulsion, exposed for 5 weeks at 4°C, developedin Kodak D19, dried overnight, and stained withtoluidine blue.
RESULTSNormal regeneration, i.e., after scale removal
only, has been documented previously in Hemi-chromis bimaculatus (Sire and Géraudie, ’84; Sireet al., ’90). In brief, when a scale is removed fromits pocket, a large part of the covering epidermisand the scale-forming cells are also removed. Nev-ertheless, a small piece of epidermis remains at-tached to the posterior region of the loose dermalstrips surrounding the scale-pocket (Fig. 2b).Epidermal cells belonging to these epidermal rem-nants spread and migrate rapidly over the scale-pocket, so that the wound surface is coveredwithin approximately 2 h in H. bimaculatus (Sireet al., ’90). The epithelial cells spread on a par-ticular bilayered mesenchymal layer called thescale-pocket lining (SPL) by Whitear et al. (’80).In H. bimaculatus, the SPL is composed of twolayers (called here outer and inner SPL layers) offlat, elongated fibrocyte-like cells attached to oneanother by numerous desmosomes. Before scaleremoval, the SPL cells are in a resting phase. Theyare characterized by scarcity in organelles andabundant microfilaments, which explains the elec-tron density of their cytoplasm at the EM level.Numerous caveolae intracellulares, with an elec-
Fig. 2. Schematical drawings from 1 µm-thick longitudi-nal sections of the skin through the experimental zone. (a)Before scale removal; (b) after scale removal; (c) after cut-ting the dermal strips to remove the overlaying epidermis(double arrows).
AbbreviationsDD dense dermisE epidermisER epidermal remnantsLD loose dermisS scaleSP scale-pocketSPL scale-pocket lining

308 A. QUILHAC AND J.-Y. SIRE
Figures 3 and 4.

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 309
tron-lucent content, are disposed along the cellmembrane. The SPL is linked to the surface ofthe subjacent dense dermis (stratum compactum)by numerous hemidesmosomes and is separatedfrom the scale-forming cells above by a basementmembrane (Whitear et al., ’80; Sire, ’89). Facingthe differentiated basal layer epidermal cells, theSPL cells proliferate, differentiate, and form adermal papilla in which a new scale is formed(Sire and Géraudie, ’84; Sire et al., ’90). The em-bryonic tissue origin of the SPL cells and that ofthe scale-forming cells is still unknown, but bothare derived from the same cell population duringontogeny (Sire, ’89).
Reconstruction of the squamation patternsControl fish: regeneration after removal ofscales only
Seven days after the operation, all the scaleshave regenerated (Fig. 3a) but have not yet at-tained the size and the thickness of normal scales(compare Fig. 4a and b). Except for their irregu-lar outline, all the 7-day-regenerated scales havethe same size and shape, and the shape is com-parable to that of normal scales surrounding thewound.
Fourteen days after scale removal, all the re-generated scales in control fish have the same sizeand shape (Fig. 3b), which are similar to those ofnormal scales (compare Fig. 4a and c). These 14-day-regenerated scales differ from normal scalesonly by their large, central region, compared tothe small focus of a normal scale. This regener-
ated “focus” is easily recognizable by its irregularornamentation (Fig. 4c).
Experimental fish: regeneration afterremoval of scales and epidermis
Seven days after the operation, the only regen-erated scales that are sufficiently developed tobe removed with forceps are situated in the pe-ripheral regions of the wound. These scales be-long to rows I and V, to column 1 and some tocolumn 7 (Fig. 3c). They are smaller than the con-trol 7-day-regenerated scales (compare Fig. 3c anda) and show differences in size and shape depend-ing on their location along the wound margins.
Fourteen days after wounding, all the scaleshave regenerated in the experimental fish, evenin the central region of the wound (Fig. 3d). Thelargest regenerated scales are situated along thewound margins but they are still not the samesize as the controls. The size of the scales de-creases from the peripheral regions towards thecentral region of the wound (compare Fig. 3b andd). Moreover, the shape of the regenerated scalesvaries, depending on their location in the oper-ated zone. The scales of rows I and V are dors-oventrally asymmetrical, and the scales of columns1 and 7 are anteroposteriorly asymmetrical, i.e.,the region of the scale situated closer to thewound margin is more developed than the oppo-site region (Figs. 3d, 4d–g). Towards the centralregion of the wound, the shape of the scales be-comes more and more irregular, and two smallscales are often found in a single pocket (Fig. 3dand detail in Fig. 4h). In one specimen, scaleswere still lacking at the center of the wound (ornot sufficiently regenerated to be removed).
One month after the operation, the squama-tion pattern on the wound (Fig. 3e) looks nor-mal. The regenerated scales are roughly thesame size and have a normal thickness, but theiroutlines are still somewhat irregular. Moreover,in the central region, some 30-day-regeneratedscales possess two foci (Fig. 4i). This suggeststhat the regeneration of several scales had beeninitiated in the same pocket at the onset of theregeneration process, and that these anlagensubsequently fused.
Restoration of the subepidermal tissues
The reconstructions of the squamation patternsreported above clearly show that in the experi-mental fish the regeneration process is progres-sively delayed from the peripheral to the central
Fig. 3. Reconstructions of the squamation patterns. Allthe scales situated in the experimental zone and the normalscales located in the rows and columns surrounding the woundwere removed 7 days (a) and 14 days (b) after scale removalonly (= controls), and 7 days (c), 14 days (d), and 30 days (e)after scale and epidermis removal. I to V and 1 to 7 refer torow and column numbers, respectively. Arrows point to re-generated scales detailed in Figure 4 (d–h). The scales thatare indicated by an arrow in (d) and (e) are detailed in Fig-ure 4. Scale bars = 1 cm.
Fig. 4. Fourteen-day (d–h = arrowed scales in Figure 3d)and 30-day (i) regenerated scales after scales and epidermiswere removed compared to (a) a normal scale, (b) a control7-day regenerated scale and (c) a control 14-day regeneratedscale. (d) Scale from row I (arrow in Fig. 3d); (e) scale fromrow V (arrow in Fig. 3d); (f) scale from column 7 (arrow inFig. 3d); (g) scale from column 1 (arrow in Fig. 3d); (h) one ofthe small scales from the center of the operated zone (arrowin Fig. 3d); (i) scale with two foci from the central region ofthe wound (arrow in Fig. 3e). For rows and column numbers,see Figure 3a. Scale bar = 1 mm.

310 A. QUILHAC AND J.-Y. SIRE
region, compared to normal regeneration. To un-derstand the reason for this delay, and to com-pare the events occurring in the subepidermaltissues until scale regeneration is complete, we havechosen to divide the wound surface into three re-gions to facilitate descriptions and comparisons: (1)a peripheral region, in which the scales seem toregenerate normally; (2) an intermediate region, inwhich scale regeneration is more or less delayed;and (3) a central region in which some scales aresmall, or lacking, even 14 days after surgery.
In the experimental fish, 1 cm2 surface of themesenchyme is exposed to the external environ-ment immediately after surgery. This wound sur-face is composed of two types of substrate onwhich the healing epidermis will spread: (1) large,homogeneous surfaces representing the scale-pocket lining (SPL) which still covers the bottomof the scale-pockets, and (2) the heterogeneous ex-tremities of the cut dermal strips that separatedthe scale-pockets from one another. The dermalstrips are formed of components of the loose der-mis (stratum laxum), i.e., bundles of collagenfibrils among which fibroblasts, pigment cells, andcapillary blood vessels are located. Given theirstructure and composition, these two substrateshave a different resistance to exposure to the ex-ternal environment. On one hand, a large part ofthe SPL keeps its integrity, whereas, on the otherhand, sectioning of the dermal strips creates gapsallowing the subjacent dermal tissues to be im-mediately accessible to the external environment.Below, we will describe the events occurring inthe regions covered by the SPL and those occur-ring at the level of the cut dermal strips, in theperipheral, intermediate, and central regions men-tioned above, from the time of wounding until theinitiation of scale regeneration in each region.
Scale-pocket lining (SPL)Immediately after surgery. Observation of sec-
tions confirms that a large part of the SPL is leftintact at the wound surface and that the organi-zation of the scale pockets was not disturbed bysuch an experimental wound. Nevertheless, theouter SPL layer cells look more electron-lucentthan before the operation, and than the inner SPLlayer cells bordering the dense dermis, and someintercellular spaces have been created (Fig. 5). TheSPL surface is covered by a thin, 250 nm thick,layer of finely fibrous extracellular matrix whichcontains material of the basement membrane thatwas located between the SPL and the scale-form-ing cells, which were removed along with the scale.
Three hours. In the peripheral region of thewound, the SPL is already covered by two layersof elongated epidermal cells that have spread fromthe unwounded epidermis bordering the operatedzone (Fig. 6). In both SPL layers, the cells areelectron-dense because of the large number of mi-crofilaments that fill their cytoplasm. Caveolae aredisposed along the cell membranes facing the der-mis and the epidermis. A space, varying from 100to 250 nm in width, separates the SPL surfacefrom the healing epithelium. This space containsa material similar to that which covered the SPLsurface after the operation (i.e., the basementmembrane components). The cells of the outer SPLlayer have cytoplasmic extensions facing the epi-thelial cells, which also extend short cytoplasmicprolongations towards the SPL cells. In the inter-mediate and central regions of the wound, the SPLis not covered by the healing epithelium, but theextracellular material is still present on its sur-face (Fig. 7). The cells of the upper SPL layer havea necrotic aspect as shown by their electron-lu-cent cytoplasm containing rounded organelles, butare still linked to one another and to the innerlayer cells by desmosomes. The cells of the innerSPL layer have conserved their initial electron-dense aspect and their cytoplasm contains numer-ous caveolae lining the cell membrane (Fig. 7).
Six hours. In the peripheral region, the SPLcells show the same features as described in theprevious stage, but the cells of both layer are lesselectron-dense (Fig. 8). The SPL is now coveredby three to four epithelial cell layers. The inter-mediate region of the wound is covered by thehealing epithelium, but the SPL cells look dam-aged. The outer layer cells show a more or lessadvanced stage of necrosis, with an electron-lu-cent cytoplasm and broken mitochondria. Some
Fig. 5. Immediately after surgery. TEM micrograph of a lon-gitudinal section of the skin surface. The bilayered scale-pocketlining located at the surface of the dense dermis is covered bya fine granular matrix (open arrow). The arrowheads indicatedesmosomes. The arrows point to caveolae located along thecell membrane facing the dense dermis. Scale bar = 500 nm.
Fig. 6. Three hours. In the peripheral region of the op-erated zone, the SPL is covered by the healing epidermis.Cytoplasmic extensions from the SPL (white arrows) andfrom the epidermal cells penetrate the epidermal-dermalspace (star), which is filled with a fine, granular extracellu-lar matrix. Numerous caveolae (black arrows) are presentalong the plasmalemma of the SPL cells. Arrowheads pointto desmosomes. Scale bar = 500 nm.
Fig. 7. Three hours. In the central region of the wound,
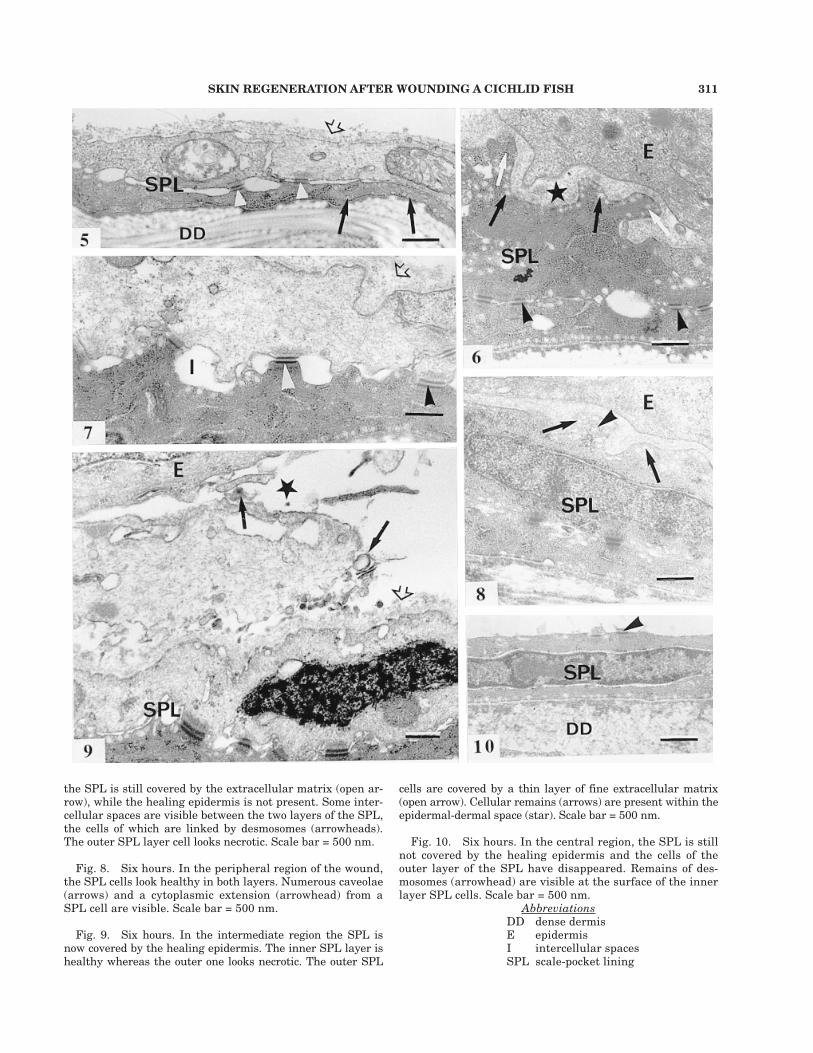
SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 311
cells are covered by a thin layer of fine extracellular matrix(open arrow). Cellular remains (arrows) are present within theepidermal-dermal space (star). Scale bar = 500 nm.
Fig. 10. Six hours. In the central region, the SPL is stillnot covered by the healing epidermis and the cells of theouter layer of the SPL have disappeared. Remains of des-mosomes (arrowhead) are visible at the surface of the innerlayer SPL cells. Scale bar = 500 nm.
AbbreviationsDD dense dermisE epidermisI intercellular spacesSPL scale-pocket lining
the SPL is still covered by the extracellular matrix (open ar-row), while the healing epidermis is not present. Some inter-cellular spaces are visible between the two layers of the SPL,the cells of which are linked by desmosomes (arrowheads).The outer SPL layer cell looks necrotic. Scale bar = 500 nm.
Fig. 8. Six hours. In the peripheral region of the wound,the SPL cells look healthy in both layers. Numerous caveolae(arrows) and a cytoplasmic extension (arrowhead) from aSPL cell are visible. Scale bar = 500 nm.
Fig. 9. Six hours. In the intermediate region the SPL isnow covered by the healing epidermis. The inner SPL layer ishealthy whereas the outer one looks necrotic. The outer SPL

312 A. QUILHAC AND J.-Y. SIRE
cells of the inner layer are also becoming necrotic,but most of them have an electron-dense cyto-plasm and look healthy (Fig. 9). The layer of ex-tracellular material is still present on the SPLsurface, even though the SPL cells are necrotic.The epidermal-dermal space is irregular and con-tains cellular debris, some of which results fromthe necrosis of epidermal cells (Fig. 9). In the cen-tral region, the healing epithelium is still not cov-ering the SPL. Here, all the cells of the outer SPLlayer and the layer of extracellular matrix havedisappeared from the wound surface. Some of theinner-layer SPL cells lining the surface of the der-mis can be necrotic but generally have an elec-tron-dense cytoplasm filled with a large numberof microfilaments (Fig. 10).
Nine hours. The healing epithelium has nowcovered the entire surface of the wound. In theperipheral region, the epidermis is thick, multi-layered, with well-differentiated basal layer cells.The cells of both SPL layers have conserved thesame aspect as in the previous stages. In the in-termediate region, either the cells of the outerlayer only or those of both SPL layers are necrotic(Fig. 11). In some areas, the outer SPL cells havedisappeared but the SPL inner layer cells are gen-erally electron-dense and look healthy. The epi-dermal-dermal space is generally narrow (ca. 200nm) and filled with extracellular matrix. Never-theless, it is enlarged in some areas because ofthe presence of macrophages (Fig. 12). The epi-dermis is now four cell layers thick, and the basallayer cells are differentiated. In the central re-gion, most of the SPL cells are necrotic or evenmissing, and the epidermal-dermal space containscellular debris of various sorts and an extracellu-lar matrix on which the epidermal basal layer cellshave spread.
One day. In the peripheral region, the outerSPL cells, which are facing the differentiated epi-dermal basal layer cells, show a large nucleuswith patches of condensed chromatin (Fig. 13).Their cytoplasm contains numerous free ribo-somes, although it is still rich in microfilamentsand caveolae. The epidermal-dermal space is ca.200 nm wide and contains a homogeneous finematrix. In the intermediate region, the healingepidermis is composed of several layers of elon-gated cells. Below, the outer SPL layer cells areeither necrotic or they are no longer present,whereas some inner layer cells look healthy. Inthe central region, the healing epithelium is threeto four cell layers thick. The basal layer epider-mal cells have irregular contours, and they are
often close to the surface of the stratum com-pactum of the dermis. The SPL cells have eitherdisappeared or are present only as cell debris.Numerous macrophages have invaded the epider-mal-dermal space in which they are seen engulf-ing cell debris (Fig. 14).
Two days. In the peripheral region, the bi-layered organization of the SPL cells facing thedifferentiated epidermal basal layer cells has dis-appeared (Fig. 15). There are now several layersof elongated, flat cells distributed within an en-larged epidermal-dermal space and separated bylarge intercellular spaces containing dispersedpatches of extracellular matrix. The cytoplasm ofthese cells contains mainly free ribosomes, somemitochondria, and elongated RER cisternae. Thecells are linked by short, inconspicuous desmo-somes. In the intermediate region, the SPL hasnot changed compared to the previous stage. Inthe central region, from which the SPL cells havebeen lost, the healing epithelium is thick and itsbasal layer cells are well differentiated. They areseparated from the collagen matrix of the dermisby a 250–300-nm wide space entirely filled witha fine granular material (Fig. 16).
Three days. In the peripheral region, regener-ating scale papillae have formed (Fig. 17). Theyare made of two layers of polarized scleroblasts,constituting an elongated bag in which the firstcomponents of the scale matrix are visible. In theintermediate region, the basal layer epidermalcells are differentiated and cuboidal. They consti-tute an uninterrupted, one-cell-layer-thick epithe-lium facing the dense dermis, on the surface ofwhich some rare SPL cells have differentiated(Fig. 18). In the central region, some elongated,mesenchymal cells are present at the surface ofthe dermis, close to the basal layer epidermal cells.These dermal cells possess a large nucleus whichoccupies most of the cytoplasm, in which howeversome mitochondria and RER cisternae are visible(Fig. 19). Some of these cells are close to cyto-
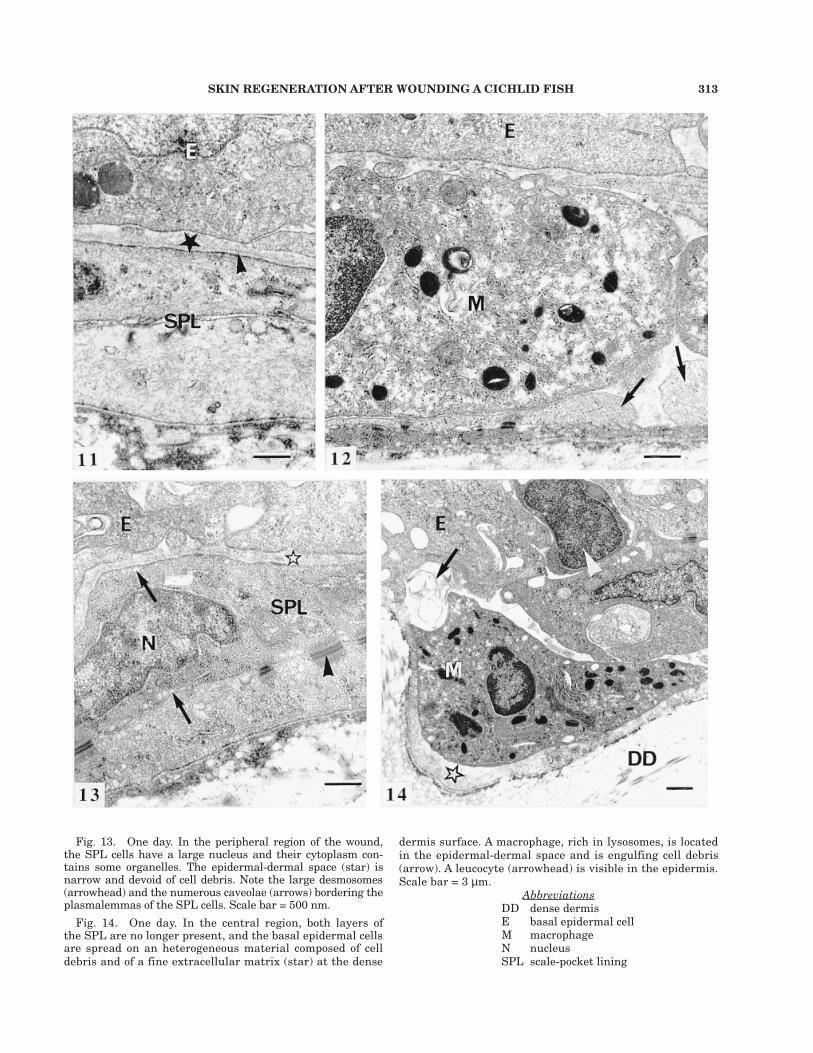
Fig. 11. Nine hours. In the intermediate region, more to-wards the wound center, cells from both SPL layers show anecrotic aspect. Note the narrow epidermal-dermal space (star)and the basement membrane-like aspect of the granular ma-trix at the SPL surface (arrowhead). Scale bar = 500 nm.
Fig. 12. Nine hours. Same region as in Figure 11. A mac-rophage and cellular debris are present in the epidermal-der-mal space. The macrophage lies at the surface of the upperSPL cells which show an electron-lucent cytoplasm (arrows).Scale bar = 500 nm.
SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 313
dermis surface. A macrophage, rich in lysosomes, is locatedin the epidermal-dermal space and is engulfing cell debris(arrow). A leucocyte (arrowhead) is visible in the epidermis.Scale bar = 3 µm.
AbbreviationsDD dense dermisE basal epidermal cellM macrophageN nucleusSPL scale-pocket lining
Fig. 13. One day. In the peripheral region of the wound,the SPL cells have a large nucleus and their cytoplasm con-tains some organelles. The epidermal-dermal space (star) isnarrow and devoid of cell debris. Note the large desmosomes(arrowhead) and the numerous caveolae (arrows) bordering theplasmalemmas of the SPL cells. Scale bar = 500 nm.
Fig. 14. One day. In the central region, both layers ofthe SPL are no longer present, and the basal epidermal cellsare spread on an heterogeneous material composed of celldebris and of a fine extracellular matrix (star) at the dense
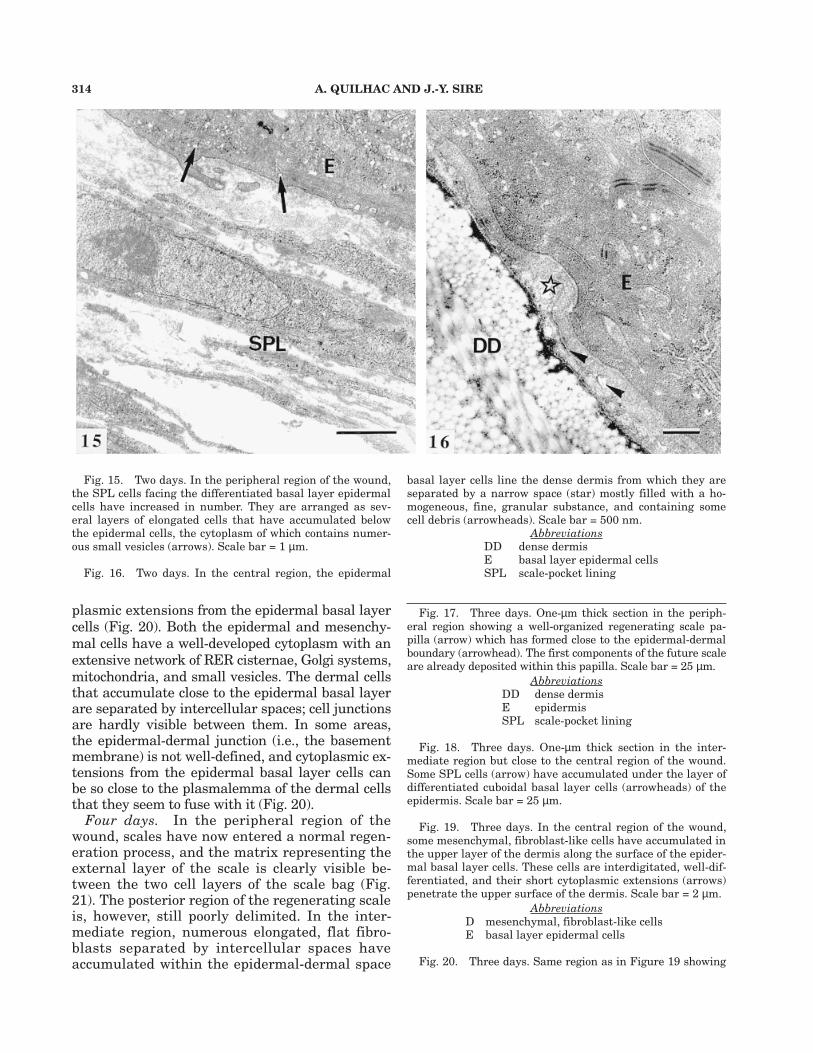
314 A. QUILHAC AND J.-Y. SIRE
plasmic extensions from the epidermal basal layercells (Fig. 20). Both the epidermal and mesenchy-mal cells have a well-developed cytoplasm with anextensive network of RER cisternae, Golgi systems,mitochondria, and small vesicles. The dermal cellsthat accumulate close to the epidermal basal layerare separated by intercellular spaces; cell junctionsare hardly visible between them. In some areas,the epidermal-dermal junction (i.e., the basementmembrane) is not well-defined, and cytoplasmic ex-tensions from the epidermal basal layer cells canbe so close to the plasmalemma of the dermal cellsthat they seem to fuse with it (Fig. 20).
Four days. In the peripheral region of thewound, scales have now entered a normal regen-eration process, and the matrix representing theexternal layer of the scale is clearly visible be-tween the two cell layers of the scale bag (Fig.21). The posterior region of the regenerating scaleis, however, still poorly delimited. In the inter-mediate region, numerous elongated, flat fibro-blasts separated by intercellular spaces haveaccumulated within the epidermal-dermal space
Fig. 15. Two days. In the peripheral region of the wound,the SPL cells facing the differentiated basal layer epidermalcells have increased in number. They are arranged as sev-eral layers of elongated cells that have accumulated belowthe epidermal cells, the cytoplasm of which contains numer-ous small vesicles (arrows). Scale bar = 1 µm.
Fig. 16. Two days. In the central region, the epidermal
basal layer cells line the dense dermis from which they areseparated by a narrow space (star) mostly filled with a ho-mogeneous, fine, granular substance, and containing somecell debris (arrowheads). Scale bar = 500 nm.
AbbreviationsDD dense dermisE basal layer epidermal cellsSPL scale-pocket lining
Fig. 17. Three days. One-µm thick section in the periph-eral region showing a well-organized regenerating scale pa-pilla (arrow) which has formed close to the epidermal-dermalboundary (arrowhead). The first components of the future scaleare already deposited within this papilla. Scale bar = 25 µm.
AbbreviationsDD dense dermisE epidermisSPL scale-pocket lining
Fig. 18. Three days. One-µm thick section in the inter-mediate region but close to the central region of the wound.Some SPL cells (arrow) have accumulated under the layer ofdifferentiated cuboidal basal layer cells (arrowheads) of theepidermis. Scale bar = 25 µm.
Fig. 19. Three days. In the central region of the wound,some mesenchymal, fibroblast-like cells have accumulated inthe upper layer of the dermis along the surface of the epider-mal basal layer cells. These cells are interdigitated, well-dif-ferentiated, and their short cytoplasmic extensions (arrows)penetrate the upper surface of the dermis. Scale bar = 2 µm.
AbbreviationsD mesenchymal, fibroblast-like cellsE basal layer epidermal cells
Fig. 20. Three days. Same region as in Figure 19 showing

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 315
several cytoplasmic extensions (arrows) from a basal layer epi-dermal cell directed towards a well-differentiated dermal cell.Note the absence of a basement membrane on the cytoplasmicextensions. Scale bar = 1 µm. Inset: Detail of a direct contact(arrow) between a cytoplasmic extension of an epidermal basallayer cell and the plasmalemma of a dermal cell. The arrow-head points to a secretory vesicle. Scale bar = 100 nm.
AbbreviationsD a well-differentiated dermal cellE basal layer epidermal cells
Fig. 21. Four days. One-µm-thick section in the periph-
eral region of the wound showing a regenerating scale (ar-row) surrounded by scale-forming cells. The surrounding der-mis has a loose texture but contains numerous fibroblasts.Scale bar = 25 µm.
AbbreviationE epidermis
Fig. 22. Four days. One-µm-thick section in the interme-diate region showing the accumulation of numerous fibroblasts(star) in an enlarged space between the epidermis and thedense dermis. Note the well-differentiated, cuboidal, basallayer cells of the epidermis. Scale bar = 25 µm.

316 A. QUILHAC AND J.-Y. SIRE
in some areas (Fig. 22). These cell populations,which are considered progenitors of the scale pa-pilla cells, are facing the layer of cuboidal, well-differentiated basal epidermal cells. A detailedexamination of consecutive serial sections fromone intermediate region has revealed that (1) thescale papillae do not form at exactly the sameplace when one compares different but adjacentscale-pockets, and (2) sometimes two scale papil-lae can differentiate within a single scale-pocket.In the central region, the epidermal-dermal spacedoes not show such cell accumulations, but themesenchymal cells of the upper layer of the der-mis are clearly differentiating.
Seven days. The regenerating scales situated inthe peripheral region of the wound have enlargedand thickened. Their associated cells are activemainly at their marginal and deep surfaces (Fig.23). In the intermediate region, scale papillae haveformed, and the first scale components are depos-ited (Figs. 24, 25). Here, the scale papillae have apoorly defined outline. They are differentiating atthe surface of the dense dermis, at a distance fromthe epidermal-dermal boundary (Fig. 24). Ultra-structural observations, however, indicate thatthese scales develop like normal ones, i.e., withina scale-bag with differentiated, highly active cells,called elasmoblasts, located along their deep sur-face (Fig. 25). The reorganization of the dermis be-low (see later section) does not appear to have aninfluence on scale regeneration. In the central re-gion, accumulations of dermal cells in the vicinityof the epidermis are clearly recognizable as pro-genitors of scale papilla cells.
Fourteen days (not illustrated). All the scalesin the operated region have regenerated. In theperipheral region, they have roughly the struc-ture of normal 14-day regenerated scales, and anepidermal fold has formed around their posteriormargin (as illustrated in Figure 2a). In the inter-mediate region, the scales are smaller than nor-mal 14-day regenerated ones, but their featuresreflect a normal regeneration process. In the cen-tral region, the irregularly shaped regeneratingscales are smaller than those in the intermediateregion. Nevertheless, they have already been dif-ferentiating for a few days, as revealed by theirthickness. The surrounding tissues look similarto those found in normal skin except for the re-generating loose dermis (but see below).
Events occurring in the dermisBoth the loose and the dense dermis are af-
fected in the experimental fish, and both are sub-
jected to osmotic shock, mostly in the central re-gion because of the long exposure to the surround-ing water.
In H. bimaculatus, normal scales are obliquelyinserted into the dermis. Their deep anterior re-gion meets the dense dermis, and it is covered bythe loose dermis (= dermal strip), which is com-posed of a network of collagen fibrils holdingmainly fibroblasts, pigment cells, nerve bundles,and capillary blood vessels. This region of der-mal strips is not damaged in the experimentalfish (Fig. 2b). The dense dermis, which is locatedbelow the scale-pocket lining (SPL), is composedmainly of several alternating thin layers of col-lagen fibrils arranged compactly in a plywood-likemanner. It also contains fibroblasts and elasticfibres and is penetrated by some nerves, bloodcapillaries, and pigment cells. When cuttingthrough a large part of the dermal strips to re-move the epidermal cells entirely, the anteriorpart of the dermal strips remains (Fig. 2c) and isimmediately exposed to the external environmentand to osmotic shock by water penetration intothe tissue (Figs. 26, 27). However, this part ofthe loose dermis will regenerate to reconstitutethe dermal strips that will eventually cover thefully regenerated scales. The dense dermis is pro-tected by the SPL cells, which, in turn, are rap-idly covered by the epidermis in the peripheralregions. However, in the central region of thewound, the dense dermis is exposed to strong os-motic shock because of the progressive destruc-tion of the SPL cells before they are covered bythe epidermis.
Three to six hours after surgery. Three hoursafter wounding, the cut dermal strip regions showincreased interfibrillar spaces (Fig. 28) comparedto the initial stage (Fig. 26). In the peripheralregions, the healing epidermis rapidly protects thesurface of the cut dermal strips against a pro-longed osmotic shock. In contrast, in the centralregion of the wound, these dermal strips are ex-posed to a longer osmotic shock, because the cov-ering by the healing epidermis is delayed forseveral hours (Fig. 28). In these regions, an exu-date, appearing as a finely granular, electron-dense substance, is visible at the surface of thestrips (Fig. 29) as well as within the interfibril-lar spaces. This exudate was not present on thesurface of the SPL described above. The upperregion of the dense dermis located close to thecut dermal strips also shows enlarged interfibril-lar spaces. Moreover, its upper collagen layers areembedded within an electron-dense substance

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 317
that is not seen in healthy regions. The deep re-gion of the dermis looks normal.
From nine hours to two days. When the epi-dermis has completely covered the wound (sinceapproximately the ninth hour) and is well recon-stituted, the collagenous dermal strips are stilldisorganized, especially near the cut surface. In-terfibrillar spaces persist and the exudate is stillpresent between the epidermis and the cut der-mal strips. Cellular debris and numerous mac-rophages are present within the disorganizedcollagen fibrils (Fig. 30).
Four days. The first indication that regenera-tion of the loose dermis has started is an accu-mulation of numerous fibroblasts within theremaining dermal strips (Fig. 31) and in the up-per part of the dense dermis. Facing these re-gions the epidermal basal layer cells are well
Fig. 23. Seven days. One-µm-thick section in the periph-eral region of the wound showing the anterior edge of a re-generated scale and the accumulation of the fibroblasts inthe loose dermis. Scale bar = 25 µm.
AbbreviationsDD dense dermisE epidermisEl elasmoblastsLD loose dermisS regenerated scaleSPL scale-pocket lining
Fig. 24. Seven days. One-µm-thick section in the inter-mediate region showing a regenerating scale (arrow), whichforms in the deep part of the loose dermis, close to the densedermis. Scale bar = 25 µm.
Fig. 25. Seven days. Detail of the regenerated scale shownin Figure 24. The scale is already composed of two layers,and its deep surface is lined by well-differentiated elasmo-blasts. The SPL below is partially reconstructed and delimitsthe scale-pocket from the upper layers of the dense dermiswhich are subjected to remodeling. Scale bar = 2 µm.
differentiated. The fibroblasts of the healing der-mis lie roughly parallel to the skin surface withthe long axis parallel to the anterior-posterioraxis of the fish. Their cytoplasm contains nu-merous organelles, among which RER cisternaeare prominent, and the oval nucleus occupies alarge part of the cell volume (Fig. 32). Some ofthese fibroblasts are close to newly synthesizedbundles of collagen fibrils parallel to each otherand to the skin surface (Fig. 33). Some enlargedspaces within the cut dermal strips are occupiedby blood capillaries, a situation which is foundmore and more common towards the central re-gion of the wound.
Five days. In the healing dermal strips, nu-merous elongated fibroblasts are regularly ar-ranged parallel to the skin surface, and to theanterior-posterior axis of the fish. In the central

318 A. QUILHAC AND J.-Y. SIRE
Figures 26–33.

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 319
trix (Fig. 35), and they show the same organelles(mainly RER) as in the previous stage.
From 5 to 10 days. The loose dermis contin-ues to regenerate. The dermal strips are progres-sively reconstructed, extending over the entiresurface of the regenerated scales. These regen-erating dermal strips are easily recognizable bythe large number of cells (fibroblasts, macroph-ages, blood cells) they contain, compared to theadjacent unwounded dense dermis (Fig. 36). Theinterfibrillar spaces are progressively reducedfrom the first to the more recently regeneratedzone. This increase in fibroblast density permitsthe place where the dermal strips have been cutto be identified easily. The region where thedense dermis has been damaged also appearsmore contrasted than the undamaged adjacenttissue. At the end of our study, i.e., 14 days af-ter surgery, the number of fibroblasts had onlyslightly decreased in these regions (Fig. 37).
Dermal cell proliferationCell proliferation was monitored during the last
3 h for all stages of wound healing in control andexperimental fish. In control fish, autoradiographsof serially sectioned skin samples reveal that (1)tritiated thymidine is rarely incorporated in thedense dermis and in the loose dermal strips, and(2) no signal was observed in the scale-pocket lin-ing cells (SPL).
Proliferation in the SPL and inscale-associated cells
Until 48 h after surgery, the only cells thatare labelled are some SPL cells of the periph-eral regions of the wound where scale regenera-tion has started. From 48 h to 4 days, thenumber of labelled dermal cells increases mainlyin these peripheral regions. Four days after sur-gery, most of the labelled nuclei are located atthe level of the scale papillae and in the regionimmediately above. The posterior region isheavily labelled compared to the anterior one(Fig. 38). In the intermediate region of thewound, some elongated SPL cells that have ac-cumulated along the epidermal surface are la-belled. In the central region of the wound, fromwhich the SPL cells have been lost, numerousfibroblasts located in the upper region of the der-mis have also proliferated (Fig. 39).
From 4 days to 7 days, the number of labelledscale-associated cells decreases, and proliferationis confined to the scale margins, mainly in theirposterior region. No labelling was found in the
Fig. 26. One-µm-thick longitudinal section of the skinin the operated zone showing a cut dermal strip immedi-ately after surgery. Arrows indicate blood cells. Scale bar= 50 µm.
AbbreviationsDD dense dermisLD cut dermal stripMu muscle cellSPL scale-pocket lining
Fig. 27. Detail of a cut dermal strip as in Figure 26. TheSPL is interrupted (arrowhead) and the collagen fibrils (ar-rows) are directly exposed to the water. Scale bar = 3 µm.
Fig. 28. Three hours after surgery. One-µm-thick longitu-dinal section of the skin in the central region of the wound.Numerous interfibrillar spaces are present within thewounded dermal strips (arrows). Scale bar = 50 µm.
Fig. 29. Two days. Detail of the surface of a cut dermalstrip in the central region of the wound showing the fine fibril-lar exudate, some material of which is dispersed within thedisorganised collagen network at the dermis surface. Scalebar = 500 nm.
AbbreviationsE basal epidermal cellEx exudateF fibroblastLD loose dermisM macrophageS scales
Fig. 30. Two days. A macrophage is located within theupper collagen layers (arrow) of a cut dermal strip coveredby the healing epidermis. Scale bar = 3 µm.
Fig. 31. Four days. One µm-thick longitudinal section inthe intermediate region of the wound. A dermal strip is start-ing to regenerate between two scales. Numerous fibroblasts(arrows) have accumulated. Note the folding of the epider-mis around the posterior edge of the regenerating scale (ar-rowhead). Scale bar = 25 µm.
Fig. 32. Four days. Details of a regenerating dermal stripin the intermediate region. The fibroblasts are surroundedby loosely-arranged collagen fibrils. Scale bar = 2 µm.
Fig. 33. Four days. Fibroblasts surrounded by parallel col-lagen fibrils (arrow) in a regenerating collagen bundle of theloose dermal strip. The cytoplasm of these fibroblasts con-tains numerous organelles. Scale bar = 1 µm.
region of the wound, blood capillaries are alsopresent (Fig. 34). This regenerating tissue fur-ther on called “granulation tissue” was describedin dermal wound healing in other vertebrates.Similarly disposed fibroblasts are seen at the sur-face of the upper region of the wounded densedermis. In both cases, the fibroblasts are sur-rounded by a loosely arranged extracellular ma-

320 A. QUILHAC AND J.-Y. SIRE
Fig. 34. Five days. One-µm-thick longitudinal section inthe central region of the wound showing the granulation tis-sue formed of numerous layers of accumulated fibroblasts andcapillary blood vessels (arrows) in a regenerating dermal strip.Scale bar = 25 µm.
Fig. 35. Five days. Detail of fibroblasts in a region asshown in Figure 34. Their cytoplasm is rich in RER and mi-tochondria and they are regularly arranged parallel to theskin surface. The large intercellular spaces are filled with afibrillar and granular material. Scale bar = 2 µm.
AbbreviationsF fibroblastsRER rough endoplasmic reticulumS scale
Fig. 36. Seven days. One µm-thick longitudinal sectionin the central region of the wound close to the anterior re-gion of a regenerating scale. In the superficial part of theregenerating dermis, the fibroblasts and capillary blood ves-sels are numerous, and the intercellular spaces are reduced.Scale bar = 25 µm.
Fig. 37. Ten days. Final step of the regeneration of a der-mal strip in the intermediate region of the wound. A scale isalready well developed. In the regenerating dermal strip thenumber of fibroblasts has slightly decreased and new collagenbundles are formed. This zone is however distinguishable fromthe adjacent, undamaged dermis by the presence of these nu-merous fibroblasts. Scale bar = 50 µm.
SPL. After 7 days of regeneration, cells surround-ing the scales are rarely labelled.
Proliferation in the dermal strips and in thedense dermis
Until 72 h after the operation, labelled fibro-blast nuclei are rarely observed throughout thecut dermal strips on the entire surface of thewound (Fig. 40). From 72 h onwards, the numberof labelled fibroblasts in these regions increases.Four days after surgery, the labelled cells are ran-domly dispersed within the upper region of thehealing dermal strips (Fig. 41) and throughout
the entire thickness of the damaged dense der-mis located below (Figs 38, 39).
From 4 days to 7 days, some labelled fibroblastsare still visible in the healing dermal strips andin the dense dermis (Fig. 42).
DISCUSSIONThis new experiment of scale regeneration on
a large surface of the skin in a teleost fish hasshown that: (1) there is a clear relation betweenthe shape and size of the regenerating scales andthe epidermal migration front on the wound sur-face; (2) two scales can be initiated in a single

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 321
scale-pocket, indicating that the healing epider-mis has probably made contact with the mesen-chyme (SPL) surface in two different loci of thesame scale-pocket; (3) the delay in scale regen-eration observed in the central region of thewound is due to the entire destruction of the SPLcells, and thus to the need to recruit mesenchy-mal cells to regenerate the scale (in this, scaleregeneration resembles scale ontogeny); (4) thereis morphological evidence for epidermal-dermalinteractions during the initiation of the regen-eration process; and (5) the skin benefits fromtwo efficient protections against osmotic shock:the SPL cells that are able to resist the expo-
sure to the water for a long time and the exu-date produced at the level of the cut dermalstrips to prevent water from entering the skin.All these results support this experimentalmodel as a good tool for further studies dealingwith the genetic control of epidermal-dermal in-teractions leading to scale morphogenesis anddifferentiation.
Reconstruction of the squamation patternand scale regeneration
The squamation patterns obtained after scaleremoval alone (i.e., without removing epidermalremnants), even on such a large surface, are in
Figs. 38–42. Autoradiographs from different regions of thewounded skin during healing, 3 h after incorporation of triti-ated thymidine. Two µm-thick longitudinal sections. No pig-ment cells are visible on these sections. All the black dots aresilver grains. All scale bars = 25 µm.
AbbreviationsDD dense dermisLD loose dermisS scale
Fig. 38. Four days. In the dermis of the intermediateregion of the wound, the labelled cells are situated aroundthe posterior region (arrows) of the regenerated scale, inthe surrounding loose and dense dermis, and in various re-gions of the epidermis.
Fig. 39. Four days. Formation of a scale papilla in thecentral region of the wound. The labelled cells are regularlydistributed among the scale papilla cells (arrows), and epi-dermal cells are still proliferating.
Fig. 40. Two days. In the damaged loose dermis, the la-belled cells (arrow) are scarce.
Fig. 41. Four days. Labelled fibroblasts (arrow) are mainlyvisible in the superficial part of the regenerating dermal stripsin the intermediate region of the wound.
Fig. 42. Seven days. Labelled fibroblasts (arrow) are stillvisible in a regenerated dermal strip above a well-formed scalefrom the intermediate region of the wound.

322 A. QUILHAC AND J.-Y. SIRE
accordance with the previous descriptions of scaleregeneration (Sire, ’82; Sire and Géraudie, ’84).Once a scale is removed from its pocket, the epi-thelial cells issuing from the epidermal remnantsspread onto the mesenchymal surface, i.e., theSPL cells. The latter differentiate and rapidly re-generate a new scale (Sire et al., ’90). Under nor-mal conditions of regeneration, this process occurssimultaneously in each scale-pocket, whatever thenumber of adjacent scales removed (Sire, ’82). InH. bimaculatus, within a short delay of 14 days,all the regenerated scales have the same shapeand size throughout the wound surface, as in thecontrols. Rapid repair after injury has also beendocumented for fin-ray regeneration in other te-leosts (e.g., Géraudie and Singer, ’92) and for limbregeneration in urodele amphibians (e.g., Goss,’69) and mouse (McCluskey and Martin, ’95).However, the rate of scale regeneration is so highthat the entire organ is reconstructed within aperiod of 7 to 14 days, which is not the case forthe fins and limbs, which require more than 1month to be totally replaced. The rapidity of scaleregeneration is probably due to the presence ofthe SPL cells, a cell population that has noequivalent in fin-rays and tetrapod limbs. TheSPL is an endothelial layer (sensu Whitear et al.,’80) which is composed of quiescent cells derivedfrom the same cell population as the scale-form-ing cells (Sire, ’89). When these cells are de-stroyed, mesenchymal cells must be recruitedfrom adjacent, unwounded regions, a process simi-lar to that occurring during fin-ray and limb re-generation (Géraudie and Singer, ’92; McCluskeyand Martin, ’95).
In contrast to the results in controls, the squa-mation patterns obtained after scale and epider-mis removal have revealed (1) that the shape ofthe scales that regenerate in the peripheral re-gions is first asymmetrical, and (2) that scale re-generation is progressively delayed from theperiphery towards the center of the wound.However, 30 days after surgery, the squamationpattern is roughly similar to the normal one,i.e., each scale-pocket contains only one scale,and all regenerated scales have approximatelythe same shape.
What occurred during the first days of regen-eration, in both the peripheral and central re-gions of the wound, is worthy of further interest.In a previous paper, we observed that the heal-ing epidermis migrates centripetally over thewound surface, i.e., from the margins toward thecenter of the wound, and that the complete re-
epithelialization requires approximately 9 h(Quilhac and Sire, submitted). We also showedthat this delay provoked some damage in theskin as a result of osmotic shock. Another studyhas been suggested that the differentiation ofthe mesenchymal cells (SPL), which will regen-erate the scale in each pocket, is probably acti-vated at the place where the epithelial cells haveestablished a contact with the mesenchymal cells(Sire et al., ’90). In the present work, the asym-metrically regenerating scales strongly supportthis hypothesis; indeed, a clear relationship ex-ists between the centripetal re-epithelializationof the wound and the shape of the regeneratingscales. Epithelial-mesenchymal interactions havebeen also reported for fin-ray regeneration in te-leosts and for limb regeneration in urodele am-phibians (Salpeter and Singer, ’60; Géraudie andSinger, ’92). Indeed, here also, the basal layercells of the healing epidermis have been reportedto be in direct contact with the mesenchymalblastema, and this suggests exchanges betweenthe two tissues. The epithelial influence prob-ably changes the course of the proliferative ac-tivity in the wounded tissues (fin-rays or limbs)from a process of repair to one of epimorphic re-generation (Mescher, ’96). A similar epithelial in-fluence is probably involved in the onset of scaleregeneration, as illustrated in the central regionof the wound by the occasional presence of twofoci of regeneration in a single scale-pocket. Inthis region, the healing epidermis, which is mi-grating from several fronts, probably contactsseveral places in a single scale-pocket wheremost of the SPL cells have been lost after theosmotic shock. This situation never occurs in theperipheral regions in which the SPL has main-tained its integrity. Here only one scale regen-erated in each scale-pocket although severalparts of the same pocket can be in contact withthe migrating epithelium. This leads us to thinkthat, where the epidermis has first contacted themesenchymal SPL cells, a signal has probablybeen transmitted to each neighbouring cell toprevent another scale from developing in thesame scale-pocket, if the healing epidermis con-tacts another region of this pocket. Conversely,in the central region of the wound, the initia-tion of two scales in a single pocket can be ex-plained either as the result of two simultaneousactivations of the newly differentiated SPL cellsby the healing epidermis arriving at the sametime in separate regions or, more probably, bythe discontinuity of scale-forming cell popula-

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 323
tions where the SPL cells have been destroyedafter the osmotic shock. Interestingly, even if twoscales are initially formed in the same scalepocket, the final result is a single regeneratedscale which obviously comes from the fusion ofthe two initial scale foci. This observation alsoimplies (1) the existence of cell-cell communica-tions from one scale-forming cell population tothe other, and (2) the final shape of the scale isprobably related to the available, well-delimited,surface of the scale pocket.
Restoration of the subepidermal tissuesOnce the re-epithelialization of the wound has
ensured efficient protection against further os-motic shock and pathogen attack, the subepider-mal tissues are subjected to several influencesleading to scale regeneration and reconstructionof the surrounding dermal connective tissue. Thepresent study clearly shows that these recoveryprocesses can vary, depending on the region ofthe wound surface. They are rapid in the periph-eral regions and are delayed in the central re-gion because of the severe osmotic shock enduredby the dermal cells. It appears that in the pe-ripheral regions where the SPL cells are undam-aged or in regions where they are able to recoverfrom a short exposure to the water, scale regen-eration occurs as described in normal regenera-tion (Sire and Géraudie, ’84; Sire et al., ’90). Onthe contrary, where the SPL cells have beendestroyed because of long exposure to the exter-nal environment, new mesenchymal cells haveto be recruited to re-initiate scale development.In a second phase, fibroblasts proliferate and dif-ferentiate to reconstruct the dermal strips, theregeneration of which is thus substantiallydelayed. Under both conditions, the picturesstrongly suggest a control through epidermal-dermal interactions.
SPL and scale regenerationIn the peripheral region of the wound, the SPL
cells are not damaged because they benefit fromrapid epidermal protection. They differentiate intoscale-forming cells that organize into a dermalpapilla in which a new scale is formed. This pro-cess is similar, both in time and space, to a nor-mal process of regeneration (Sire and Géraudie,’84; Sire et al., ’90, ’97). In the experimental ani-mals, the absence of the posterior region of thedermal strips, and as a consequence of its SPLcovering, appears to have no influence on scaleregeneration. This suggests that initiation of re-
generation requires only interactions between thehealing epidermis and the competent mesenchy-mal (SPL) cells and that patterning of the skinhas been maintained even after destruction of theSPL cells. The area occupied by the SPL cell popu-lation within a scale-pocket is the only factor thatprimarily governs the extent of the regeneratingscale papilla (Sire, ’82). In the intermediate andcentral regions of the wound, the delay observedin scale regeneration is related to the more orless severe destruction of the SPL cells becauseof the long exposure to the external environment.Indeed, in the experimental fish, re-epithelializa-tion occurs rapidly, and the epidermal basal layercells also differentiate quickly (Quilhac and Sire,submitted). The SPL cells can resist osmotic shockfor several hours, as observed in the intermedi-ate regions, where the regenerated scales looknormal. Although the SPL has clearly suffered,as illustrated by the EM micrographs, it is ableto recover its original capacity to regenerate ascale. This also can occur when only the outerSPL layer is destroyed. The inner layer SPL cellshave the same origin as the outer ones (Sire, ’89)and can also interact with the epidermal cells.Some teleost species are known to possess an SPLonly one cell layer thick (Whitear, ’86; Sire et al.,’97), and their scales regenerate normally.
In the central region of the wound, the longdelay observed in scale regeneration (at least 10days) is the consequence of the SPL destruction.A previous study has shown that the re-epithe-lialization of this region is rapidly followed bythe differentiation of the basal layer epidermalcells, i.e., a layer of juxtaposed, cuboidal cells.These cells show an obvious secretory activity in-terpreted as a possible secretion of signaling mol-ecules towards the dermis below (Quilhac andSire, submitted). The mere presence of well-dif-ferentiated epidermal cells facing the dermis isnot sufficient to activate the scale-regenerationprocess in the absence of competent mesenchy-mal cells. The epidermal-dermal interactions arecertainly delayed during the first days followingthe wound. Light and TEM observations haveclearly shown that the only cells in the mesen-chyme able to regenerate scales rapidly are SPLcells. The absence of these cells has also resultedin damage within the subepidermal tissues, in-creasing the delay in scale regeneration evenmore. However, we have shown that even in thesehighly damaged regions, the skin is able to re-generate scales after approximately 10 days ofdelay. Mesenchymal cells, progenitors of the

324 A. QUILHAC AND J.-Y. SIRE
scale-forming cells, probably recruited from thevicinity of the damaged zone, migrate towardsthe wounded region and accumulate beneath thebasal lamina of the epidermal-dermal boundary.This recruitment phase may be similar to whatoccurs during fin-ray regeneration in teleost, dur-ing limb regeneration in amphibians (Géraudie andSinger, ’92; review in Stocum, ’95), and in embry-onic wound healing in mammals (McCluskey andMartin, ’95). Indications of epidermal-dermal in-teractions are morphologically detected at this levelthrough the accumulation of numerous smallvesicles in the cytoplasm of the epidermal cells andthrough the numerous cell to cell contacts be-tween the two tissues (Quilhac and Sire, sub-mitted). The mesenchymal cells that have beenrecruited differentiate into scale-forming cells,which constitute a scale bag in which the firstscale elements are produced. In this recruitment,the process resembles scale development (Sireand Géraudie, ’83; Sire et al., ’97) but not scaleregeneration. This leads us to the conclusionthat, through this technique of experimentalscale regeneration, we are able to provoke there-initiation of the processes of scale develop-ment in adult specimens. Nevertheless the ques-tions of the origin of the mesenchymal cells andof the nature of the signals involved in their re-cruitment remain unanswered.
We have shown previously that the reconstruc-tion of the healing epidermis in H. bimaculatusdoes not depend on the type of substrate it cov-ers (Quilhac and Sire, submitted). Moreover, thepresence of a competent mesenchymal cell popu-lation is not a prerequisite for the differentia-tion of the basal layer epidermal cells, either inthe margins or in the center of the wound. Itappears that the differentiation of the epider-mal basal layer cells depends on the state ofdifferentiation of the epidermis only. On the con-trary, the presence of a well-differentiated epi-dermal basal cell layer seems to be a prerequisitefor the differentiation of the mesenchymal SPLcells: they never differentiate unless the epider-mal basal layer cells are well-differentiated.Thus, the epidermal basal layer cells would playthe first role in the initiation of the scale regen-eration as suggested already by Sire et al. (’90).Such a role for the epithelial cells has alreadybeen demonstrated for tooth initiation in mam-mals, but there the signal seems to be limitedto the so-called dental lamina, a thickening ofthe basal region of the epidermis (Kollar andBaird, ’69; Thesleff et al., ’95). Interestingly, our
study reveals that, once differentiated, the epi-dermal basal layer cells in the central region ofthe wound are able to maintain their special-ized role (i.e., for production of signal molecules)during a long period until the recruitment of ap-propriate mesenchymal cells does occur.
Even though the upper SPL layer cells are dam-aged by osmotic shock, this particular layer is anefficient barrier against the injuries resultingfrom such a large wound. Indeed, the experimen-tal fishes have never shown infections or sufferedfrom weakness after surgery (even though therewere no antibiotics in the tank water). The pro-tection afforded by the fish skin benefits from arapid re-epithelialization of the wound and fromthe production of an exudate in the regions ex-posed to water and potential infection.
DermisDermal wound healing involves considerable
reorganization, notably with regard to the der-mal strips. These have not only been severelydamaged by cutting but have been subsequentlysubmitted to osmotic shock as well. The processof destruction continues for a few hours follow-ing the wound, but it is rapidly stopped by thepresence of the healing epidermis in the periph-eral regions and by the production of an exudatein the intermediate and central regions of thewound. The loose organization of the collagenfibrils and the presence of numerous interfibril-lar spaces and of numerous cell debris reflect theseverity of the injury. A few hours after the op-eration, fibroblast-like cells have colonized thewound and are surrounded by a thin fibrillar sub-stance, suggesting that they have deposited somenew collagen fibrils. In mammals, the soft tis-sues at the wounding site undergo histolysis andretract. In addition, during the initial step ofwound healing, the basement membrane wouldplay a stabilizing role as demonstrated duringnewt limb regeneration (Neufeld et al., ’96). InH. bimaculatus, the tissues at the wound mar-gin did not retract, but histolysis occurred firstas a consequence of osmotic shock. The basementmembrane components, which are well preservedon the SPL surface for several hours, favour epi-thelial cell migration (Quilhac and Sire, submit-ted). In mammals, collagenase activity is highin the wound dermis (Agren, ’94), and in amphib-ians a gelatinase/collagenase has been identifiedas playing a role in the initiation and rapidgrowth of wound healing (Yang and Bryant, ’94).Metalloproteinases could be involved during the

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 325
cleaning phase following the wound closure andwould favour cell migration during the first stepsof dermal strip regeneration in H. bimaculatus,but this needs further investigations. In H.bimaculatus, the reparative phase requires mi-gration and proliferation of fibroblasts followedby synthesis and deposition of collagen matrix,as seen in other teleost species (Mittal and DattaMunshi, ’74) and in mammals (e.g., Bereiter-Hahn, ’86). The collagen matrix deposition in H.bimaculatus is particularly important from thefourth day after surgery onwards, i.e., when thefibroblast proliferation slows down. At this stage,the fibroblasts show clear indications of activeprotein synthesis (numerous Golgi systems, mi-tochondria, and dilated RER cisternae).
The source of the fibroblast cells colonizing thecentral region of the wound are apparently un-differentiated, quiescent mesenchymal cells. Theyare first activated, and then attracted by an un-known mechanism to invade the wound, wherethey differentiate. This process has been de-scribed in other fish (Mittal et al., ’78) and hasbeen observed in amphibians (Weiss, ’61) and inhuman wounds (Aho et al., ’83; Eckerley andDudley, ’88). The origin of the fibroblasts hasbeen debated (Peacock, ’84), but they probablyoriginate from the connective tissues surround-ing the wound (Ross and Odland, ’68; Mittal andDatta Munshi, ’74).
A rich network of blood vessels is visible in thehighly damaged central region, 5 days aftersurgery. They permit the supply of oxygen andnutrients in the healing zone, in view of the in-creased needs by the actively proliferating cellsin the wound region. Formation of this granula-tion tissue seems to be a general phenomenon inwound repair in terrestrial vertebrates (Lacourand Ortonne, ’83; Vanable, ’89), and it has beenobserved in teleosts as well (Mittal and Munshi,’74; Anderson and Roberts, ’75; Mittal et al., ’78).During wound healing in the rabbit ear cham-ber, fibroblasts play a role in angiogenesis as pro-genitors of endothelial cells in newly formedblood vessels (Kon and Fujiwara, ’94), but nu-merous other players are involved, includingcapillary tips, macrophage-derived chemical at-tractants, and components of the extracellularmatrix (Pettet et al., ’96). Fibroblast-dependentangiogenesis is quite favourable for wound heal-ing because the proliferation rate of endothelialcells has been reported to be too slow to accountfor the rapid growth of the vascular network(Fujimoto et al., ’87).
Macrophages are seen invading the damagedtissues of the skin, and are especially numerousin the loose dermal strips during wound repair.During fin-ray regeneration, cleaning of the de-bris is minimal due to the small size of the bro-ken bone (Géraudie and Singer, ’92), whereas inurodele limb regeneration, there are numerousmacrophages because of the large wound (Mes-cher, ’96). In mammals, macrophages invadewounds within about 6 h (review in Riches, ’88),and they are attracted by the substances pro-duced by the degradation of the collagen (Lacourand Ortonne, ’83) and by programmed cell death(Hopkinson-Woolley et al., ’94). These cells havea role of phagocytosis of tissue debris and for-eign particles (Winter, ’73), but macrophages areknown to promote both fibroblast migration to-wards the wounded regions and enhancement ofproliferation by producing growth factors andcytokines (Turck et al., ’87; review in Martin etal., ’92). Cell proliferation in the dermis is highin the regenerating dermal strips but not as im-portant as in the epidermis. It occurs around thethird day after the operation, as the number offibroblasts increases. This shows that most of thefibroblasts are derived by proliferation from fi-broblasts which have previously migrated to-wards the damaged zone.
Wound contraction does not occur in Hemi-chromis after scale and epidermis removal, inspite of serious damage at the center of thewound. The wound surface is still visible aftercomplete healing and has the same size as justafter wounding. In mammals, as soon as thegranulation tissue is visible, wound contractionbegins and leads to wound closure with littlescarring (Grinnel, ’94). This process is probablybrought about by the activity of specialised fi-broblasts, which have contractile properties(Gabbiani et al., ’72).
In conclusion, this experimental wound in acichlid fish appears to be a good model for fur-ther studies dealing with (1) the genetic controlof epidermal-dermal interactions leading to scaleregeneration, including initiation, morphogen-esis, and differentiation, and (2) the processes ofskin healing, especially learning which moleculesare involved in preventing water from enteringthe wounded skin.
ACKNOWLEDGMENTSThe authors are grateful to Mary Whitear
(Tavistok, UK), Prof. Ann Huysseune (Universityof Ghent, Belgium), and Prof. Armand de Ricqlès

326 A. QUILHAC AND J.-Y. SIRE
(Collège de France, Paris) for helpful suggestionsand critical reading of the manuscript. We thankMiss F. Allizard for her excellent technical assis-tance. TEM observations and photographic workhave been done in the Centre Interuniversitairede Microscopie Electronique, CIME Jussieu,Universités Paris 6-Paris 7.
LITERATURE CITEDAgren, M.S. (1994) Gelatinase activity during wound heal-
ing. Br. J. Dermatol., 131:634–640.Aho, H.J., J. Viljanto, J. Raekallio, and L.J. Pelliniemi (1983)
Ultrastructural characteristics of cells in human woundcollected by cellstic device. J. Surg. Res., 35:498–506.
Akimenko, M.A., S.L. Johnson, M. Westerfield, and M. Ekker(1995) Differential induction of four msx homeobox genesduring fin development and regeneration in zebrafish. De-velopment, 121:347–357.
Anderson, C.D., and R.J. Roberts (1975) A comparison ofthe effects of temperature on wound healing in a tropicaland temperate teleost. J. Fish Biol., 7:173–182.
Bereiter-Hahn, J. (1986) Epidermal cell migration and woundrepair. In: Biology of the Integument. 2. Vertebrates. J.Bereiter-Hahn, A.G. Matoltsy, and K.S. Richards, eds.Springer-Verlag, Berlin, pp. 443–471.
Eckerley, J.R.T., and H.A.F. Dudley (1988) Wounds andwound healing. Br. Med. Bull., 44:423–436.
Fujimoto, S., K. Yamamoto, H. Kagawa, M. Yoshizuka, T.Nomiyama, and T. Maruyama (1987) Neovascularizationin the pre- and post-natal rabbit corpora cavernosa penis:Light and electron microscopy and autoradiography. Anat.Rec., 218:30–39.
Gabbiani, G., B.J. Hirschel, G.B. Ryan, P.R. Statkov, and G.Majno (1972) Granulation tissue as a contractile organ: astudy of structure and function. J. Exp. Med., 135:719–734.
Géraudie, J., and M. Singer (1992) The fish fin regenerate.In: Keys for Regeneration. C.H. Taban and B. Boilly, eds.Monogr. Dev. Biol., Karger, Basel. pp. 62–72.
Goss, R.J. (1969) Principles of Regeneration. Academic Press,New York.
Grinnel, F. (1994) Mini-review on the cellular mechanismsof disease. Fibroblasts, myofibroblasts and wound contrac-tion. J. Cell Biol., 124:401–404.
Hopkinson-Woolley, J., D. Hughes, S. Gordon, and P. Mar-tin (1994) Macrophage recruitment during limb develop-ment and wound healing in the embryonic and foetalmouse. J. Cell Sci., 107:1159–1167.
Kollar, E.J., and G.R. Baird (1969) The influence of the den-tal papilla on the development of tooth shape in embryonicmouse tooth germs. J. Embryol. Exp. Morphol., 21:131–148.
Kon, K., and T. Fujiwara (1994) Transformation of fibroblastsinto endothelial cells during angiogenesis. Cell Tissue Res.,278:625–628.
Lacour, J.P., and J.P. Ortonne (1983) La cicatrisation. Ann.Dermatol. Venereol., 110:159–173.
Martin, P., J. Hopkinson-Woolley, and J. McCluskey (1992)Growth factors and cutaneous wound repair. Prog. GrowthFactor Res., 4:25–44.
McCluskey, J., and P. Martin (1995) Analysis of tissue move-ments of embryonic wound healing- DiI studies in the limbbud stage mouse embryo. Dev. Biol., 170:102–114.
Mescher, A.L. (1996) The cellular basis of limb regenerationin urodeles. Int. J. Dev. Biol., 40:785–795.
Mittal, A.K., and J.S. Datta Munshi (1974) On the regen-eration and repair of superficial wounds in the skin of Ritarita (Ham) (Bagridae, Pisces). Acta.Anat., 88:424–442.
Mittal, A.K., A.K. Rai, and T.K. Banerjee (1978) Studies onthe pattern of healing of wounds in the skin of a cat-fishHeteropneustes fossilis (Bloch) (Heteropneustidae, Pisces).Z. Mikrosk-Anat. Forsch (Leipz.) 91:270–286.
Neufeld, D.A., A.D. Frances, and H.E. Settles (1996) Stabi-lizing role of the basement membrane and dermal fibersduring newt limb regeneration. Anat. Rec., 245:122–127.
Peacock, E.E. (1984) Wound Repair, Ed 3. W.B. Saunders,Philadelphia.
Pettet, G., M.A.J. Chaplain, D.L.S. McElwain, and H.M.Byrne (1996) On the role of angiogenesis in wound heal-ing. Proc. R. Soc. Lond. B, 263:1487–1493.
Riches, D.W.H. (1988) The multiple roles of macrophages inwound healing. In: The Molecular and Cellular Biology ofWound Repair.R.A.F. Clark and P.M. Henson, eds. PlenumPress, New York, pp. 213–239.
Ross, R., and G. Odland (1968) Human wound repair. II.Inflammatory cells, epithelial-mesenchymal interrelations,and fibrogenesis. J. Cell Biol., 39:152–168.
Salpeter, M.M., and M Singer (1960) Differentiation of thesubmicroscopic adepidermal membrane during limb regen-eration in adult Triturus, including a note on the termbasement membrane. Anat. Rec., 136:27–34.
Sire, J.-Y. (1982) Régénération des écailles d’un Cichlidé,Hemichromis bimaculatus (Gill) (Téléostéen Perciforme).I. Morphogenèse, structure et minéralisation. Ann. Sci.Nat. Zool., Paris, 4:153–169.
Sire, J.-Y. (1987) Structure, formation et régénération desécailles d’un poisson téléostéen, Hemichromis bimaculatus(Perciforme, Cichlidé). Thèse de Doctorat d’Etat, UniversitéParis 7, Arch. Doc. Inst. Ethnol., Micro-edition, Mus. Hist.Nat., SN 87600449.
Sire, J.-Y. (1988) Evidence that mineralized spherules areinvolved in the formation of the superficial layer of theelasmoid scale in cichlids Cichlasoma octofasciatum andHemichromis bimaculatus (Pisces, Teleostei): an epidermalactive participation. Cell Tissue Res., 253:165–172.
Sire, J.-Y. (1989) The same lineage is involved in scaleformation and regeneration in the teleost fish Hemi-chromis bimaculatus (Perciformes, Cichlidae). TissueCell, 21:447–462.
Sire, J.-Y., and J. Géraudie (1983) Fine structure of the de-veloping scale in the cichlid Hemichromis bimaculatus (Pi-sces, Teleostei, Perciformes). Acta Zool., 64:1–8.
Sire, J.-Y., and J. Géraudie (1984) Fine structure of theregenerating scales and their associated cells in thecichlid Hemichromis bimaculatus (Gill). Cell Tissue Res.,237:537–547.
Sire, J.-Y., H. Boulekbache, and C. Joly (1990) Epidermal-dermal and fibronectin cell-interactions during fish scaleregeneration: immunofluorescence and TEM studies. Biol.Cell, 68:147–158.
Sire, J.-Y., F. Allizard, O. Babiar, J. Bourguignon, and A.Quilhac (1997) Scale development in zebrafish (Daniorerio). J Anat., 190:545–561.
Smith, M.M., and B.K. Hall (1990) Development and evolu-tionary origins of vertebrate skeletogenic and odontogenictissues. Biol. Rev., 65:277–374.
Smith, M.M., and B.K. Hall (1993) A developmental modelfor evolution of the vertebrate exoskeleton and teeth: therole of the cranial and trunk neural crest. Evol. Biol.,27:387–448.

SKIN REGENERATION AFTER WOUNDING A CICHLID FISH 327
Stocum, D.L. (1995) Wound Repair, Regeneration, and Arti-ficial Tissues. R.G. Landes, Austin, Tex.
Thesleff, I., A. Vaahtokari, P. Kettunen, and T. Aberg (1995)Epithelial-mesenchymal signaling during tooth develop-ment. Connect. Tissue Res., 32:9–15.
Turck, C.V., J.G. Dohlman, and E.J. Goetz (1987) Immuno-logical mediators of wound healing and fibrosis. J. CellPhysiol., Suppl. 5:89–93.
Vanable, J.W. (1989) Integumentary potentials and woundhealing. In: Electric Fields in Vertebrates Repair, 5:171–224.
Weiss, P. (1961) The biological foundations of wound repair.Harvey Lect. Ser., 55:13–42.
Whitear, M. (1986) The skin of fishes including cyclostomes:Dermis. In: Biology of the integument. (2) Vertebrates. J.Bereiter-Hahn, A.G. Matoltsy, and S. Richards, eds.Springer, Heidelberg, pp. 8–64.
Whitear, M., A.K. Mittal, and E.B. Lane (1980) Endotheliallayers in fish skin. J. Fish Biol., 17:43–65.
Winter, D.G. (1973) Epidermal regeneration studied in thedomestic pig. In: Epidermal Wound Healing. H.I. Maibachand D.T. Rovee, eds. Year Book Medical Publ., Chicago,pp. 71–112.
Yang, E.V., and S.V. Bryant (1994) Developmental regula-tion of a matrixmetalloproteinase during regeneration ofaxolotl appendages. Dev. Biol., 166:696–703.