review article evaluating the importance of the carotid...
TRANSCRIPT

Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 893506 18 pageshttpdxdoiorg1011552013893506
Review ArticleEvaluating the Importance of the Carotid Chemoreceptors inControlling Breathing during Exercise in Man
M J Parkes
School of Sport Exercise amp Rehabilitation Sciences University of Birmingham Edgbaston Birmingham B15 2TT UK
Correspondence should be addressed to M J Parkes mjparkesbhamacuk
Received 25 April 2013 Accepted 8 July 2013
Academic Editor Maxime Cannesson
Copyright copy 2013 M J Parkes This is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Only the carotid chemoreceptors stimulate breathing during hypoxia in Man They are also ideally located to warn if the brainrsquosoxygen supply falls or if hypercapnia occurs Since their discovery sim80 years ago stimulation ablation and recording experimentsstill leave 3 substantial difficulties in establishing how important the carotid chemoreceptors are in controlling breathing duringexercise inMan (i) they are in the wrong location to measure metabolic rate (but are ideally located to measure any mismatch) (ii)they receive no known signal during exercise linking them with metabolic rate and no overt mismatch signals occur and (iii) theirdenervation inMan fails to prevent breathingmatchingmetabolic rate in exercise New research is needed to enable recording fromcarotid chemoreceptors in Man to establish whether there is any factor that rises with metabolic rate and greatly increases carotidchemoreceptor activity during exercise Available evidence so far in Man indicates that carotid chemoreceptors are either one oftwo mechanisms that explain breathing matching metabolic rate or have no importance We still lack key experimental evidenceto distinguish between these two possibilities
1 Introduction
Following earlier studies of the anatomy sensory innervationand cardiovascular functions of the carotid bifurcation regionby De Castro [1] the respiratory functions of the carotidchemoreceptors were discovered in 1927 by accident [2] andwon Corneille Heymans the 1938 Nobel Prize in medicineThe carotid chemoreceptors remain the only chemoreceptorsknown in Man to be stimulated by hypoxia and that in turnstimulate breathing It has always been believed thereforethat they must be key to explaining how breathing matchesmetabolic rate so well Yet since 1927 the experimental evi-dence has failed to reveal whether they have any importancein Man
Between rest and maximum exercise in Man metabolicrate (the rate of oxygen consumption VO
2) increases by up to
sim23-fold and breathing increases by up to sim35-fold to match(a hyperpnea) or exceed (a hyperventilation) metabolic rateIf breathing matches metabolic rate the systemic arterialblood gases (the partial pressures of oxygen [PaO
2] and
of carbon dioxide [PaCO2]) should remain constant and
the stability of PaCO2is the simplest indication of how
well matching occurs Since they do match during exercise(Figures 1 and 2) there is no arterial blood gas signal to tellthe carotid chemoreceptors what metabolic rate is This hasalways been a fundamental problem in exercise physiology
Julius Comroe wrote in 1944
the history of respiratory physiology for the last60 years has been a succession of attempts toexplain all deviations from normal respirationby one theory but no one theory yet offeredcan account for all or even a large part of thehyperpnea of muscular exercise and each of thesetheories has failed to gain general acceptancebecause of its inability to explain this the mostcommon and most powerful of all respiratoryadjustments [3]
Despite many attempts to address this problem and toevaluate the numerous other possible mechanisms involved(eg [4ndash10]) and despite the outstanding work on Man ofWasserman Whipp et al [11ndash14] the remarkable situationexists in 2012 where two reviews in the same journal still drawvery different conclusions Thus
2 BioMed Research International
the plasticity of the sensory response of the carotidbody are critical for ventilatory adaptations as well as during exercise [15]
but
stimuli postulated to act at carotid chemore-ceptors are not primary mediators of the hyperp-nea [16]
This review identifies the key experimental evidenceavailable in Man and what is still required to establish howimportant the carotid chemoreceptors are in controllingbreathing in exercise in Man the species in which wehave most interest It deliberately follows Bronowskirsquos [17]dictum
students are not here to worship what is knownbut to question it
2 Terminology
Here the terms ldquobreathingrdquo ldquoventilationrdquo and ldquominute ven-tilation ( Ve)rdquo will be used interchangeably and measuredas minute ventilation ( Ve in litres air BTPSsdotminminus1) Mea-surements of metabolic rate ( VO
2) are made at the mouth
by indirect calorimetry in kilowatts (where kW = LO2
STPDsdotminminus1 x an assumed fuel conversion factor of 03366)and rate of CO
2production ( VCO
2) ismeasured at themouth
as LCO2STPDsdotminminus1 External work rate is indicated as
Watts (Wew) Measurements are cited as means plusmn SEM andthe number of subjects is indicated by 119899 =
3 Carotid Chemoreceptors asMetabolic Rate or Mismatch Sensors andTheir Ideal Location
The simplest system to explain how breathing matchesmetabolic rate so well requires something acting as ametabolic rate sensor This sensor does not have to bea chemoreceptor If it is ideally it would somehow mea-sure how much O
2has been extracted from arterial blood
(the systemic arteriovenous difference in O2content) and
metabolic rate could be derived by combining this withknowledge of cardiac output (via the Fick principle) Thefundamental difficulty however is that the known peripheralchemoreceptors are located only on the systemic arterialside They are therefore in the wrong location to measuremetabolic rate because O
2has not yet been extracted and
because CO2has already been removed Figures 1 and 3 show
that it is the systemic venous not arterial blood gases thatchange proportionately with metabolic rate Yet no cardiacpulmonary nor mixed venous chemoreceptors have yet beenfound in Man [11 18]
Although in the wrong place to measure metabolic rateitself carotid chemoreceptors are ideally located to indicate ifany breath fails to match metabolic rate and hence to providethe necessary signal to fine-tune each breath with metabolicrate A single mismatching breath would cause arterial blood
80
70
60
50
40
00 05 10 15 20
PCO2
(torr
)
LAT
Mixed venous
Arterial
VO2 VO2LAT
Figure 1 PaCO2fails to rise above the normal range during exercise
(unlike PvCO2) Mean plusmn se PaCO
2(brachial artery) and mixed
venous PvCO2(pulmonary artery) during incremental exercise to
maximum normalized to theirmean lactic acidosis threshold (LAT)of 20 plusmn 00 LO
2minminus1 07 kW in 5 healthy subjects Reproduced
with permission from Sun et al [75]
gases to deviate from normal and provide a blood gas errorsignal to modify the next breath The bigger the mismatchthe bigger the error signal Although this error signal isinherently ambiguous (it cannot distinguish a mismatchcaused by an inappropriate breath from a change inmetabolicrate or from disproportionate changes in both) the systemappears to be wired on the assumption that any mismatch iscaused by hypoventilation rather than raised metabolic rateThis is because arterial hypoxia or hypercapnia stimulatesbreathing rather than reduce metabolic rate The carotidchemoreceptors undoubtedly have the capability to signal anymismatch and have a response time fast enough to fine-tuneeach breath All this is of clinical relevance [19] But as shownbelow there is no evidence yet that they do perform this rolenor that this role makes any important contribution to thecontrol of breathing in exercise
4 What Is Necessary to Establish IfCarotid Chemoreceptors Are the MetabolicRate Sensor
Classical scientific method in neurophysiology (originatingwith Bernard [20] and Mill [21] and see also [22ndash26])proposes that 3 conditions must be satisfied to establishwhether any one sensor alone is the keymetabolic rate sensor
(1) stimulation at rest of this sensor should mimic themaximum increase in breathing (100ndash150 L airsdotminminus1)at maximummetabolic rate (sim5 LO
2sdotminminus1)
(2) ablation of this sensor should prevent breathingmatching metabolic rate
(3) recorded activity of this sensor must increase anddecrease proportionately with metabolic rate
BioMed Research International 3
06 kW
120
80
40
0minus1 0 2 4 6 minus1 0 2 4 6 minus1 0 2 4 6
09 kW 11 kW
Time (min) Time (min) Time (min)
(L ai
r BTP
Smiddotminminus1)
Ve
(a)
40
minus4
minus8
ΔpH
a
000
008
minus016
minus0240 1 2 3 4 5 6 0 1 2 3 4 5 6 0 1 2 3 4 5 6
ΔPa
CO2
(mm
Hg)
lowast lowast
lowastlowast lowast
lowast lowast
lowast
(b)
ΔPa
CO2
(mm
Hg)
6
4
2
0
minus20 20 40 60 0 20 40 60 0 20 40 60
06 kW 09 kW 11 kW
Time (s) Time (s) Time (s)
lowast lowast
lowast lowast lowast
lowast lowast lowast
(c)
Figure 2 No detectable PaCO2rise above the normal range during exercise even when frequently sampled (a) Mean breathing (b) Mean plusmn
se change in brachial PaCO2from resting values (ΔPaCO
2) andmean change in pHa (ΔpHa) during rest followed by the 6min exercise period
(average values for the 8 subjects displayed in 10 s time bins) with cycling at sim06 kW (80 of their lactic acidosis threshold) sim09 kW andsim11 kW (mean VO
2max was 36 L O
2minminus1 = 12 kW) in 8 subjects (c) Mean plusmn se ΔPaCO
2during the first minute only lowast119875 lt 005 versus
rest (repeated measures ANOVA) Reproduced with permission from Stringer et al [68]
4 BioMed Research International
(a) Rest
Cardiac output = 5 L blood per min
PaCO2 = 40mmHg(052L CO2 per L arterial blood)
Right heartLeft heart
Systemic venous chemoreceptors
Mixed Pv CO2 = 40 + 6mmHg(052 + 025L CO2 per L venous blood)
Systemic veins
CO2
CO2
Lungs
muscleSkeletal
Systemic arteries
Cardiac output = 25 L blood per min
PaCO2 = 35mmHg(049 L CO2 per L arterial blood)
Carotidchemoreceptors
Centralchemoreceptors
Mixed Pv CO2 = 35 + 45mmHg(049 + 4525L CO2 per L venous blood)
Each L venous blood gains 004 L CO2
times 5 Lmin cardiac output = 02L CO2 carriedEach L venous blood gains 018 L CO2
times 25 Lmin cardiac output = 45L CO2 carried
(b) Exercise
= 02 L CO2min = 45 L CO2min
= 45 L CO2min= 02 L CO2min
VCO2 VCO2
VCO2 VCO2
Figure 3 Systemic arterial chemoreceptors are ideally located anatomically to measure any mismatch but are in the wrong place to measuremetabolic rate or metabolically produced CO
2 Diagram of typical values of cardiac output systemic arterial PaCO
2and mixed PvCO
2
and of blood CO2content at rest (a) and maximum exercise (b) with data from multiple sources For clarity the corresponding numbers
for metabolic rates are omitted Comparison of (a) and (b) shows that systemic arterial chemoreceptors are in the wrong place to measuremetabolically produced CO
2 because metabolically produced CO
2is carried via the systemic veins frommuscle to air without PaCO
2rising
To establish the carotid chemoreceptors as the metabolicrate sensor all three conditions must be satisfied Eitheralone is inadequate for example (1) alone might also beachieved by an artificial stimulus for example a drug (2)alone might merely represent some traumatic or accidentalside effect and (3) alone establishes only correlation butnot causation This review will consider each condition forcarotid chemoreceptors but before doing so some furtherpoints must first be considered
5 The Redundancy Summation andMultiplication Beliefs
All ablation experiments in exercise so far fail to abolishbreathing matching metabolic rate Rather than concludingthat the ablated factors are irrelevant it is usually concludedinstead that at least two sensor mechanisms must exist eachof which could mediate matching on its own but at anymoment one or more must contribute (ie both duplicate orare redundant) This is one of two common beliefs
(i) the redundancy (or duplication) belief
nature would not have designed such an impor-tant control system to rely on only one mechanism so at least two factors must control
(ii) the multiplication belief
hyperpnea of exercise due to multiplicationof a great number of discrete impulses whichare individually small but collectively large
These beliefs are automatically applied when interpretingcarotid chemoreceptor experiments and are never chal-lenged Both are scientifically testable in Man but there is nosubstantial experimental evidence yet for either
The redundancy belief means that wemust already acceptthat at least twomechanisms sense metabolic rate (ie one inaddition to carotid chemoreceptors) This in turn comes upagainst Occamrsquos razor
(essentia non sunt multiplicanda praeter necessi-tatem)
it is naıve to propose multiple mechanisms before firstestablishing one Yet so far we cannot even establish one Nostimulation of a singlemechanismhas yet increased breathingby more than sim15 Lsdotmin-1 [27 28] that is to nowherenear that of maximum exercise The principal interest in theredundancy belief however is the difficulty that it creates ininterpreting the negative effect of an ablation experiment inMan as further explained in Section 6
The multiplication belief predicts that the more multipli-cation the interaction requires themore vulnerable breathing
BioMed Research International 5
must be to ablation of any singlemechanismNeither ablationof carotid chemoreceptors nor of any other mechanism hasyet detected any such vulnerability of breathing in Man seefor example [29 30]
6 Can Stimulation Ablation and RecordingEstablish That Carotid Chemoreceptors AreNot the Key Sensor in Man
Falsifying any one of these three conditions could establishthat carotid chemoreceptors are not the key sensor Falsifi-cation however is difficult for two reasons First it is moredifficult to deal with negatives (no change in response) Forexample it is not yet possible to record neural activity directlyfrom the carotid chemoreceptors in Man and hence KarlSaganrsquos
ldquoabsence of evidence is not evidence of absencerdquoUltimately negatives are validated only by discovering whatmechanism does satisfy all three conditions Secondly thesafer conclusion from a negative result is initially ldquoincon-clusive without further experimentsrdquo because either someexperimental error occurred (eg the ablation was techni-cally unsuccessful or some measurement error occurred)or because the ablated mechanism is either irrelevant orredundant
Resolving ldquoinconclusiverdquo requires independent verifica-tion that ablation was successfully achieved that no experi-mental errors occurred and then identification and ablationof the second (or more) mechanism(s) There are equal dif-ficulties in identifying what might be the second duplicatingmechanism (and the quest to identify the second mechanismis beyond the scope of this review)Only if ablating the secondmechanism alone prevents matching could this finally estab-lish the irrelevance of the firstmechanism and the importanceof the second Whereas if matching was prevented onlyby ablating both both are important and duplicate Thisreview will also demonstrate that we are equally unable toestablish definitively that carotid chemoreceptors are not thekey metabolic rate sensor in Man
7 The Ideal Experimental Design with CarotidChemoreceptor Experiments
It is assumed that the same control system explains matchingfrom rest to maximum exercise rather than exercise beingsome special case This is important for two reasons Firstwhatever explains matching should be greater hence easierto detect as metabolic rate increases Secondly whateverexplains matching must explain it over the large increments(sim35- sim23- and sim21-fold) in the key variables ( Ve VO
2 and
VCO2) up to maximum exercise (ge19 kW)
There is also the scientific tenet that all experimentalresults should be confirmed by independent laboratories(every ldquofactrdquo should be referenced at least twice) Many of thecrucial carotid chemoreceptor experiments cited here haveonly ever been attempted once Furthermore there is noagreement on the ideal number of subjects per study Few usemore than 10 (often for good practical reasons) and many do
not even use 5 Statistical analysis can help (but not all applyit) although statistical power calculations are limited becausethey are for design rather than analysis and because they toodepend on the number of subjects
Positive results that are statistically significant (becausethe observed effect size is so large and its variance is so small)evenwith only two subjects are important and have sufficientpower But even these have two problems First there cannotbe complete confidence in variance being estimated fromonly 2 subjects Secondly there can be difficulty in convincingthe scientific community that these 2 subjects are sufficientlyrepresentative of the population
The particular problem with many of the key carotidchemoreceptor experiments onMan reviewed here howeveris that negative results occurred few subjects were usedthe observed effect size is small and the variance is largeFor example with only 2 subjects the observed effect sizeis often not statistically significant The difficulty then is indistinguishing a lack of power (false negative) from a truenegative Strictly statistical power analysis indicates only thatthe design requires more subjects which only confirms ouroriginal concern about low subject numbers
8 Recording Experiments fromHuman Arterial Blood
The principal physiological stimulants of the carotid bodiesare hypoxia hypercapnia and acidity Without the ability torecord from carotid chemoreceptor activity in Man mea-suring PaCO
2 PaO
2 and pHa levels is the only means of
investigating the recording condition for carotid chemore-ceptors that is are carotid chemoreceptors stimulated byraising metabolic rate
A fundamental requirement of a classical control systemusing feedback from a metabolic rate sensor to drive breath-ing is that arterial blood gases must change appropriatelyand proportionately with metabolic rate To produce theldquoerrorrdquo signal that drives breathing they must change in theappropriate directionmdashPaO
2falling and PaCO
2rising they
should change in proportion to exercise intensity and theymust remain changed as long as metabolic rate is increased inorder to sustain breathing at this new metabolic rate
Figures 4(a) and 4(b) show how large the changes inarterial blood gasesmust be for arterial blood gases to providethe key metabolic rate signal driving breathing at maximumexercise (breathing gt 100 Lsdotminminus1) But at maximum exercise(even in endurance trained athletes [31]) PaO
2decreases
neither transiently nor consistently enough to reach even theFiO2ldquothresholdrdquo in Figure 4(a) that stimulates breathing
Similarly PaCO2fails to rise enough Figures 1 and 2
show that there is barely any reproducible or sustained risein PaCO
2during moderate exercise and PaCO
2actually
falls during severe exercise So there is no PaCO2rise
during exercise sufficient to stimulate breathing even to the20 Lsdotminminus1 of Figure 4(b) So far the only observed bloodgas changes suitable to provide a metabolic rate signal are insystemic venous blood (Figure 1) Thus at present there is noobvious arterial factor that rises with metabolic rate and isknown to stimulate carotid chemoreceptors in Man
6 BioMed Research International
Equivalent altitude (km) calculated by Dripps0 5 10 15 20 25
40
30
20
10
100 209 18 16 14 12 10 8 6 4
500
400
300
200
100
Percent oxygen in inspired gas
102 80 66 51 40
Resp
irato
ry m
inut
evo
lum
e(L
min
)
Estimated equivalent PaO2 (mmHg)
Perc
enta
ge o
f nor
mal
resp
irato
ry m
inut
e vol
ume
(a)
20
18
16
12
8
4
32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62
PaCO2 (mmHg)
55 CO2
43 CO2
22 CO2
Air
plusmn1 se from mean respiratory minute volume
Ventilation (LBTPSmiddotminminus1)
(b)
Figure 4 (a) Relative insensitivity of breathing at rest to artificially lowering PO2in Man Minute ventilation (plusmn se solid bars plusmn sd open
bars) in normal subjects [87] as inspired oxygen is artificially lowered (strictly hypocapnic hypoxia exists once hyperventilation occurs)Equivalent PaO
2points are aligned on the FiO
2scale with PaO
2estimated before hyperventilation occurs using the alveolar gas equation
(assuming 760mmHg barometric pressure RQ = 08 PaO2= PAO
2 and PaCO
2= PACO
2) and the point afterwards is estimated based on
dynamic forcing experiments in isocapnia (courtesy of Dr G A Balanos) Reproducedwith permission fromDripps et al [87] (b) Sensitivityof breathing at rest to artificially raising PaCO
2in Man Minute ventilation and PaCO
2(femoral) in 8 healthy men [88] while inhaling 0ndash6
CO2in air at atmospheric pressure (mean slope is 25 Lsdotminminus1mmHgminus1 artificial PaCO
2rise) Reproduced with permission from Lambertsen
et al [88]
9 Absence of Suitable PaCO2Oscillations in Man
Another possibility is that to detect and signal metabolic ratecarotid chemoreceptors might detect some other propertyof O2or CO
2 such as the variability in PaO
2or PaCO
2
rather than their absolute levels Alveolar PCO2in Man does
oscillate systematically within the respiratory cycle at rest andis mirrored as oscillations in arterial blood pH and PaCO
2
[32ndash34] It was therefore thought that carotid chemoreceptorsmight estimate metabolic rate from the rate of rise or someother mathematical function of the PaCO
2oscillation But
there are several difficulties [18] with the CO2oscillation
hypothesis Thus
(i) It has not been established whether such oscillationscause the breathing pattern or whether the breathingpattern causes such oscillations
(ii) Such oscillations apparently disappear when breath-ing frequency rises during exercise Band et al [32]did find larger pH oscillations at exercise than at restbut only in one subject and only in the first minute ofexercise as tidal volume (but not breathing frequency)increased Whereas Murphy et al [33] found thatsuch oscillations disappeared in 4 subjects during thefirst minute of exercise (006 kW) when breathingat 20 breaths per minute (suggesting that it is slowbreathing that causes these oscillations and not viceversa)
(iii) The oscillation hypothesis is not supported by thedemonstration [35] that brief hypoxic pulses (N
2in
25ndash33 of the tidal volume) to the inspirate in 4subjects that cause detectable arterial desaturationat the carotid region (earlobe) failed to alter thefirst relevant respiratory cycle either at rest or inexercise (ge125Wew) and even if hypoxia (PAO
2
of 60mmHg) was used to increase chemoreceptoractivity
(iv) Denervation of carotid chemoreceptors (see [2930]mdash Section 13) apparently fails to prevent breathingmatchingmetabolic rate inMan yet these are the onlyknown chemoreceptors fast enough to detect suchoscillations in Man
There is no good reason to doubt any of these data Afurther difficulty is that it is not yet possible to test whetherblood gas changes precede a change in each breath becausesufficiently fast responding arterial electrodes are not gener-ally available [33] Using breath-by-breath measurement ofend tidal PetCO
2or PetO
2to estimate their corresponding
arterial levels preceding each breath is problematic becauseend tidal levels are not always reliable estimates of arteriallevels at rest and become progressively less so as systemicPvCO
2increases with metabolic rate (see Figure 1)
BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
the plasticity of the sensory response of the carotidbody are critical for ventilatory adaptations as well as during exercise [15]
but
stimuli postulated to act at carotid chemore-ceptors are not primary mediators of the hyperp-nea [16]
This review identifies the key experimental evidenceavailable in Man and what is still required to establish howimportant the carotid chemoreceptors are in controllingbreathing in exercise in Man the species in which wehave most interest It deliberately follows Bronowskirsquos [17]dictum
students are not here to worship what is knownbut to question it
2 Terminology
Here the terms ldquobreathingrdquo ldquoventilationrdquo and ldquominute ven-tilation ( Ve)rdquo will be used interchangeably and measuredas minute ventilation ( Ve in litres air BTPSsdotminminus1) Mea-surements of metabolic rate ( VO
2) are made at the mouth
by indirect calorimetry in kilowatts (where kW = LO2
STPDsdotminminus1 x an assumed fuel conversion factor of 03366)and rate of CO
2production ( VCO
2) ismeasured at themouth
as LCO2STPDsdotminminus1 External work rate is indicated as
Watts (Wew) Measurements are cited as means plusmn SEM andthe number of subjects is indicated by 119899 =
3 Carotid Chemoreceptors asMetabolic Rate or Mismatch Sensors andTheir Ideal Location
The simplest system to explain how breathing matchesmetabolic rate so well requires something acting as ametabolic rate sensor This sensor does not have to bea chemoreceptor If it is ideally it would somehow mea-sure how much O
2has been extracted from arterial blood
(the systemic arteriovenous difference in O2content) and
metabolic rate could be derived by combining this withknowledge of cardiac output (via the Fick principle) Thefundamental difficulty however is that the known peripheralchemoreceptors are located only on the systemic arterialside They are therefore in the wrong location to measuremetabolic rate because O
2has not yet been extracted and
because CO2has already been removed Figures 1 and 3 show
that it is the systemic venous not arterial blood gases thatchange proportionately with metabolic rate Yet no cardiacpulmonary nor mixed venous chemoreceptors have yet beenfound in Man [11 18]
Although in the wrong place to measure metabolic rateitself carotid chemoreceptors are ideally located to indicate ifany breath fails to match metabolic rate and hence to providethe necessary signal to fine-tune each breath with metabolicrate A single mismatching breath would cause arterial blood
80
70
60
50
40
00 05 10 15 20
PCO2
(torr
)
LAT
Mixed venous
Arterial
VO2 VO2LAT
Figure 1 PaCO2fails to rise above the normal range during exercise
(unlike PvCO2) Mean plusmn se PaCO
2(brachial artery) and mixed
venous PvCO2(pulmonary artery) during incremental exercise to
maximum normalized to theirmean lactic acidosis threshold (LAT)of 20 plusmn 00 LO
2minminus1 07 kW in 5 healthy subjects Reproduced
with permission from Sun et al [75]
gases to deviate from normal and provide a blood gas errorsignal to modify the next breath The bigger the mismatchthe bigger the error signal Although this error signal isinherently ambiguous (it cannot distinguish a mismatchcaused by an inappropriate breath from a change inmetabolicrate or from disproportionate changes in both) the systemappears to be wired on the assumption that any mismatch iscaused by hypoventilation rather than raised metabolic rateThis is because arterial hypoxia or hypercapnia stimulatesbreathing rather than reduce metabolic rate The carotidchemoreceptors undoubtedly have the capability to signal anymismatch and have a response time fast enough to fine-tuneeach breath All this is of clinical relevance [19] But as shownbelow there is no evidence yet that they do perform this rolenor that this role makes any important contribution to thecontrol of breathing in exercise
4 What Is Necessary to Establish IfCarotid Chemoreceptors Are the MetabolicRate Sensor
Classical scientific method in neurophysiology (originatingwith Bernard [20] and Mill [21] and see also [22ndash26])proposes that 3 conditions must be satisfied to establishwhether any one sensor alone is the keymetabolic rate sensor
(1) stimulation at rest of this sensor should mimic themaximum increase in breathing (100ndash150 L airsdotminminus1)at maximummetabolic rate (sim5 LO
2sdotminminus1)
(2) ablation of this sensor should prevent breathingmatching metabolic rate
(3) recorded activity of this sensor must increase anddecrease proportionately with metabolic rate
BioMed Research International 3
06 kW
120
80
40
0minus1 0 2 4 6 minus1 0 2 4 6 minus1 0 2 4 6
09 kW 11 kW
Time (min) Time (min) Time (min)
(L ai
r BTP
Smiddotminminus1)
Ve
(a)
40
minus4
minus8
ΔpH
a
000
008
minus016
minus0240 1 2 3 4 5 6 0 1 2 3 4 5 6 0 1 2 3 4 5 6
ΔPa
CO2
(mm
Hg)
lowast lowast
lowastlowast lowast
lowast lowast
lowast
(b)
ΔPa
CO2
(mm
Hg)
6
4
2
0
minus20 20 40 60 0 20 40 60 0 20 40 60
06 kW 09 kW 11 kW
Time (s) Time (s) Time (s)
lowast lowast
lowast lowast lowast
lowast lowast lowast
(c)
Figure 2 No detectable PaCO2rise above the normal range during exercise even when frequently sampled (a) Mean breathing (b) Mean plusmn
se change in brachial PaCO2from resting values (ΔPaCO
2) andmean change in pHa (ΔpHa) during rest followed by the 6min exercise period
(average values for the 8 subjects displayed in 10 s time bins) with cycling at sim06 kW (80 of their lactic acidosis threshold) sim09 kW andsim11 kW (mean VO
2max was 36 L O
2minminus1 = 12 kW) in 8 subjects (c) Mean plusmn se ΔPaCO
2during the first minute only lowast119875 lt 005 versus
rest (repeated measures ANOVA) Reproduced with permission from Stringer et al [68]
4 BioMed Research International
(a) Rest
Cardiac output = 5 L blood per min
PaCO2 = 40mmHg(052L CO2 per L arterial blood)
Right heartLeft heart
Systemic venous chemoreceptors
Mixed Pv CO2 = 40 + 6mmHg(052 + 025L CO2 per L venous blood)
Systemic veins
CO2
CO2
Lungs
muscleSkeletal
Systemic arteries
Cardiac output = 25 L blood per min
PaCO2 = 35mmHg(049 L CO2 per L arterial blood)
Carotidchemoreceptors
Centralchemoreceptors
Mixed Pv CO2 = 35 + 45mmHg(049 + 4525L CO2 per L venous blood)
Each L venous blood gains 004 L CO2
times 5 Lmin cardiac output = 02L CO2 carriedEach L venous blood gains 018 L CO2
times 25 Lmin cardiac output = 45L CO2 carried
(b) Exercise
= 02 L CO2min = 45 L CO2min
= 45 L CO2min= 02 L CO2min
VCO2 VCO2
VCO2 VCO2
Figure 3 Systemic arterial chemoreceptors are ideally located anatomically to measure any mismatch but are in the wrong place to measuremetabolic rate or metabolically produced CO
2 Diagram of typical values of cardiac output systemic arterial PaCO
2and mixed PvCO
2
and of blood CO2content at rest (a) and maximum exercise (b) with data from multiple sources For clarity the corresponding numbers
for metabolic rates are omitted Comparison of (a) and (b) shows that systemic arterial chemoreceptors are in the wrong place to measuremetabolically produced CO
2 because metabolically produced CO
2is carried via the systemic veins frommuscle to air without PaCO
2rising
To establish the carotid chemoreceptors as the metabolicrate sensor all three conditions must be satisfied Eitheralone is inadequate for example (1) alone might also beachieved by an artificial stimulus for example a drug (2)alone might merely represent some traumatic or accidentalside effect and (3) alone establishes only correlation butnot causation This review will consider each condition forcarotid chemoreceptors but before doing so some furtherpoints must first be considered
5 The Redundancy Summation andMultiplication Beliefs
All ablation experiments in exercise so far fail to abolishbreathing matching metabolic rate Rather than concludingthat the ablated factors are irrelevant it is usually concludedinstead that at least two sensor mechanisms must exist eachof which could mediate matching on its own but at anymoment one or more must contribute (ie both duplicate orare redundant) This is one of two common beliefs
(i) the redundancy (or duplication) belief
nature would not have designed such an impor-tant control system to rely on only one mechanism so at least two factors must control
(ii) the multiplication belief
hyperpnea of exercise due to multiplicationof a great number of discrete impulses whichare individually small but collectively large
These beliefs are automatically applied when interpretingcarotid chemoreceptor experiments and are never chal-lenged Both are scientifically testable in Man but there is nosubstantial experimental evidence yet for either
The redundancy belief means that wemust already acceptthat at least twomechanisms sense metabolic rate (ie one inaddition to carotid chemoreceptors) This in turn comes upagainst Occamrsquos razor
(essentia non sunt multiplicanda praeter necessi-tatem)
it is naıve to propose multiple mechanisms before firstestablishing one Yet so far we cannot even establish one Nostimulation of a singlemechanismhas yet increased breathingby more than sim15 Lsdotmin-1 [27 28] that is to nowherenear that of maximum exercise The principal interest in theredundancy belief however is the difficulty that it creates ininterpreting the negative effect of an ablation experiment inMan as further explained in Section 6
The multiplication belief predicts that the more multipli-cation the interaction requires themore vulnerable breathing
BioMed Research International 5
must be to ablation of any singlemechanismNeither ablationof carotid chemoreceptors nor of any other mechanism hasyet detected any such vulnerability of breathing in Man seefor example [29 30]
6 Can Stimulation Ablation and RecordingEstablish That Carotid Chemoreceptors AreNot the Key Sensor in Man
Falsifying any one of these three conditions could establishthat carotid chemoreceptors are not the key sensor Falsifi-cation however is difficult for two reasons First it is moredifficult to deal with negatives (no change in response) Forexample it is not yet possible to record neural activity directlyfrom the carotid chemoreceptors in Man and hence KarlSaganrsquos
ldquoabsence of evidence is not evidence of absencerdquoUltimately negatives are validated only by discovering whatmechanism does satisfy all three conditions Secondly thesafer conclusion from a negative result is initially ldquoincon-clusive without further experimentsrdquo because either someexperimental error occurred (eg the ablation was techni-cally unsuccessful or some measurement error occurred)or because the ablated mechanism is either irrelevant orredundant
Resolving ldquoinconclusiverdquo requires independent verifica-tion that ablation was successfully achieved that no experi-mental errors occurred and then identification and ablationof the second (or more) mechanism(s) There are equal dif-ficulties in identifying what might be the second duplicatingmechanism (and the quest to identify the second mechanismis beyond the scope of this review)Only if ablating the secondmechanism alone prevents matching could this finally estab-lish the irrelevance of the firstmechanism and the importanceof the second Whereas if matching was prevented onlyby ablating both both are important and duplicate Thisreview will also demonstrate that we are equally unable toestablish definitively that carotid chemoreceptors are not thekey metabolic rate sensor in Man
7 The Ideal Experimental Design with CarotidChemoreceptor Experiments
It is assumed that the same control system explains matchingfrom rest to maximum exercise rather than exercise beingsome special case This is important for two reasons Firstwhatever explains matching should be greater hence easierto detect as metabolic rate increases Secondly whateverexplains matching must explain it over the large increments(sim35- sim23- and sim21-fold) in the key variables ( Ve VO
2 and
VCO2) up to maximum exercise (ge19 kW)
There is also the scientific tenet that all experimentalresults should be confirmed by independent laboratories(every ldquofactrdquo should be referenced at least twice) Many of thecrucial carotid chemoreceptor experiments cited here haveonly ever been attempted once Furthermore there is noagreement on the ideal number of subjects per study Few usemore than 10 (often for good practical reasons) and many do
not even use 5 Statistical analysis can help (but not all applyit) although statistical power calculations are limited becausethey are for design rather than analysis and because they toodepend on the number of subjects
Positive results that are statistically significant (becausethe observed effect size is so large and its variance is so small)evenwith only two subjects are important and have sufficientpower But even these have two problems First there cannotbe complete confidence in variance being estimated fromonly 2 subjects Secondly there can be difficulty in convincingthe scientific community that these 2 subjects are sufficientlyrepresentative of the population
The particular problem with many of the key carotidchemoreceptor experiments onMan reviewed here howeveris that negative results occurred few subjects were usedthe observed effect size is small and the variance is largeFor example with only 2 subjects the observed effect sizeis often not statistically significant The difficulty then is indistinguishing a lack of power (false negative) from a truenegative Strictly statistical power analysis indicates only thatthe design requires more subjects which only confirms ouroriginal concern about low subject numbers
8 Recording Experiments fromHuman Arterial Blood
The principal physiological stimulants of the carotid bodiesare hypoxia hypercapnia and acidity Without the ability torecord from carotid chemoreceptor activity in Man mea-suring PaCO
2 PaO
2 and pHa levels is the only means of
investigating the recording condition for carotid chemore-ceptors that is are carotid chemoreceptors stimulated byraising metabolic rate
A fundamental requirement of a classical control systemusing feedback from a metabolic rate sensor to drive breath-ing is that arterial blood gases must change appropriatelyand proportionately with metabolic rate To produce theldquoerrorrdquo signal that drives breathing they must change in theappropriate directionmdashPaO
2falling and PaCO
2rising they
should change in proportion to exercise intensity and theymust remain changed as long as metabolic rate is increased inorder to sustain breathing at this new metabolic rate
Figures 4(a) and 4(b) show how large the changes inarterial blood gasesmust be for arterial blood gases to providethe key metabolic rate signal driving breathing at maximumexercise (breathing gt 100 Lsdotminminus1) But at maximum exercise(even in endurance trained athletes [31]) PaO
2decreases
neither transiently nor consistently enough to reach even theFiO2ldquothresholdrdquo in Figure 4(a) that stimulates breathing
Similarly PaCO2fails to rise enough Figures 1 and 2
show that there is barely any reproducible or sustained risein PaCO
2during moderate exercise and PaCO
2actually
falls during severe exercise So there is no PaCO2rise
during exercise sufficient to stimulate breathing even to the20 Lsdotminminus1 of Figure 4(b) So far the only observed bloodgas changes suitable to provide a metabolic rate signal are insystemic venous blood (Figure 1) Thus at present there is noobvious arterial factor that rises with metabolic rate and isknown to stimulate carotid chemoreceptors in Man
6 BioMed Research International
Equivalent altitude (km) calculated by Dripps0 5 10 15 20 25
40
30
20
10
100 209 18 16 14 12 10 8 6 4
500
400
300
200
100
Percent oxygen in inspired gas
102 80 66 51 40
Resp
irato
ry m
inut
evo
lum
e(L
min
)
Estimated equivalent PaO2 (mmHg)
Perc
enta
ge o
f nor
mal
resp
irato
ry m
inut
e vol
ume
(a)
20
18
16
12
8
4
32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62
PaCO2 (mmHg)
55 CO2
43 CO2
22 CO2
Air
plusmn1 se from mean respiratory minute volume
Ventilation (LBTPSmiddotminminus1)
(b)
Figure 4 (a) Relative insensitivity of breathing at rest to artificially lowering PO2in Man Minute ventilation (plusmn se solid bars plusmn sd open
bars) in normal subjects [87] as inspired oxygen is artificially lowered (strictly hypocapnic hypoxia exists once hyperventilation occurs)Equivalent PaO
2points are aligned on the FiO
2scale with PaO
2estimated before hyperventilation occurs using the alveolar gas equation
(assuming 760mmHg barometric pressure RQ = 08 PaO2= PAO
2 and PaCO
2= PACO
2) and the point afterwards is estimated based on
dynamic forcing experiments in isocapnia (courtesy of Dr G A Balanos) Reproducedwith permission fromDripps et al [87] (b) Sensitivityof breathing at rest to artificially raising PaCO
2in Man Minute ventilation and PaCO
2(femoral) in 8 healthy men [88] while inhaling 0ndash6
CO2in air at atmospheric pressure (mean slope is 25 Lsdotminminus1mmHgminus1 artificial PaCO
2rise) Reproduced with permission from Lambertsen
et al [88]
9 Absence of Suitable PaCO2Oscillations in Man
Another possibility is that to detect and signal metabolic ratecarotid chemoreceptors might detect some other propertyof O2or CO
2 such as the variability in PaO
2or PaCO
2
rather than their absolute levels Alveolar PCO2in Man does
oscillate systematically within the respiratory cycle at rest andis mirrored as oscillations in arterial blood pH and PaCO
2
[32ndash34] It was therefore thought that carotid chemoreceptorsmight estimate metabolic rate from the rate of rise or someother mathematical function of the PaCO
2oscillation But
there are several difficulties [18] with the CO2oscillation
hypothesis Thus
(i) It has not been established whether such oscillationscause the breathing pattern or whether the breathingpattern causes such oscillations
(ii) Such oscillations apparently disappear when breath-ing frequency rises during exercise Band et al [32]did find larger pH oscillations at exercise than at restbut only in one subject and only in the first minute ofexercise as tidal volume (but not breathing frequency)increased Whereas Murphy et al [33] found thatsuch oscillations disappeared in 4 subjects during thefirst minute of exercise (006 kW) when breathingat 20 breaths per minute (suggesting that it is slowbreathing that causes these oscillations and not viceversa)
(iii) The oscillation hypothesis is not supported by thedemonstration [35] that brief hypoxic pulses (N
2in
25ndash33 of the tidal volume) to the inspirate in 4subjects that cause detectable arterial desaturationat the carotid region (earlobe) failed to alter thefirst relevant respiratory cycle either at rest or inexercise (ge125Wew) and even if hypoxia (PAO
2
of 60mmHg) was used to increase chemoreceptoractivity
(iv) Denervation of carotid chemoreceptors (see [2930]mdash Section 13) apparently fails to prevent breathingmatchingmetabolic rate inMan yet these are the onlyknown chemoreceptors fast enough to detect suchoscillations in Man
There is no good reason to doubt any of these data Afurther difficulty is that it is not yet possible to test whetherblood gas changes precede a change in each breath becausesufficiently fast responding arterial electrodes are not gener-ally available [33] Using breath-by-breath measurement ofend tidal PetCO
2or PetO
2to estimate their corresponding
arterial levels preceding each breath is problematic becauseend tidal levels are not always reliable estimates of arteriallevels at rest and become progressively less so as systemicPvCO
2increases with metabolic rate (see Figure 1)
BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
06 kW
120
80
40
0minus1 0 2 4 6 minus1 0 2 4 6 minus1 0 2 4 6
09 kW 11 kW
Time (min) Time (min) Time (min)
(L ai
r BTP
Smiddotminminus1)
Ve
(a)
40
minus4
minus8
ΔpH
a
000
008
minus016
minus0240 1 2 3 4 5 6 0 1 2 3 4 5 6 0 1 2 3 4 5 6
ΔPa
CO2
(mm
Hg)
lowast lowast
lowastlowast lowast
lowast lowast
lowast
(b)
ΔPa
CO2
(mm
Hg)
6
4
2
0
minus20 20 40 60 0 20 40 60 0 20 40 60
06 kW 09 kW 11 kW
Time (s) Time (s) Time (s)
lowast lowast
lowast lowast lowast
lowast lowast lowast
(c)
Figure 2 No detectable PaCO2rise above the normal range during exercise even when frequently sampled (a) Mean breathing (b) Mean plusmn
se change in brachial PaCO2from resting values (ΔPaCO
2) andmean change in pHa (ΔpHa) during rest followed by the 6min exercise period
(average values for the 8 subjects displayed in 10 s time bins) with cycling at sim06 kW (80 of their lactic acidosis threshold) sim09 kW andsim11 kW (mean VO
2max was 36 L O
2minminus1 = 12 kW) in 8 subjects (c) Mean plusmn se ΔPaCO
2during the first minute only lowast119875 lt 005 versus
rest (repeated measures ANOVA) Reproduced with permission from Stringer et al [68]
4 BioMed Research International
(a) Rest
Cardiac output = 5 L blood per min
PaCO2 = 40mmHg(052L CO2 per L arterial blood)
Right heartLeft heart
Systemic venous chemoreceptors
Mixed Pv CO2 = 40 + 6mmHg(052 + 025L CO2 per L venous blood)
Systemic veins
CO2
CO2
Lungs
muscleSkeletal
Systemic arteries
Cardiac output = 25 L blood per min
PaCO2 = 35mmHg(049 L CO2 per L arterial blood)
Carotidchemoreceptors
Centralchemoreceptors
Mixed Pv CO2 = 35 + 45mmHg(049 + 4525L CO2 per L venous blood)
Each L venous blood gains 004 L CO2
times 5 Lmin cardiac output = 02L CO2 carriedEach L venous blood gains 018 L CO2
times 25 Lmin cardiac output = 45L CO2 carried
(b) Exercise
= 02 L CO2min = 45 L CO2min
= 45 L CO2min= 02 L CO2min
VCO2 VCO2
VCO2 VCO2
Figure 3 Systemic arterial chemoreceptors are ideally located anatomically to measure any mismatch but are in the wrong place to measuremetabolic rate or metabolically produced CO
2 Diagram of typical values of cardiac output systemic arterial PaCO
2and mixed PvCO
2
and of blood CO2content at rest (a) and maximum exercise (b) with data from multiple sources For clarity the corresponding numbers
for metabolic rates are omitted Comparison of (a) and (b) shows that systemic arterial chemoreceptors are in the wrong place to measuremetabolically produced CO
2 because metabolically produced CO
2is carried via the systemic veins frommuscle to air without PaCO
2rising
To establish the carotid chemoreceptors as the metabolicrate sensor all three conditions must be satisfied Eitheralone is inadequate for example (1) alone might also beachieved by an artificial stimulus for example a drug (2)alone might merely represent some traumatic or accidentalside effect and (3) alone establishes only correlation butnot causation This review will consider each condition forcarotid chemoreceptors but before doing so some furtherpoints must first be considered
5 The Redundancy Summation andMultiplication Beliefs
All ablation experiments in exercise so far fail to abolishbreathing matching metabolic rate Rather than concludingthat the ablated factors are irrelevant it is usually concludedinstead that at least two sensor mechanisms must exist eachof which could mediate matching on its own but at anymoment one or more must contribute (ie both duplicate orare redundant) This is one of two common beliefs
(i) the redundancy (or duplication) belief
nature would not have designed such an impor-tant control system to rely on only one mechanism so at least two factors must control
(ii) the multiplication belief
hyperpnea of exercise due to multiplicationof a great number of discrete impulses whichare individually small but collectively large
These beliefs are automatically applied when interpretingcarotid chemoreceptor experiments and are never chal-lenged Both are scientifically testable in Man but there is nosubstantial experimental evidence yet for either
The redundancy belief means that wemust already acceptthat at least twomechanisms sense metabolic rate (ie one inaddition to carotid chemoreceptors) This in turn comes upagainst Occamrsquos razor
(essentia non sunt multiplicanda praeter necessi-tatem)
it is naıve to propose multiple mechanisms before firstestablishing one Yet so far we cannot even establish one Nostimulation of a singlemechanismhas yet increased breathingby more than sim15 Lsdotmin-1 [27 28] that is to nowherenear that of maximum exercise The principal interest in theredundancy belief however is the difficulty that it creates ininterpreting the negative effect of an ablation experiment inMan as further explained in Section 6
The multiplication belief predicts that the more multipli-cation the interaction requires themore vulnerable breathing
BioMed Research International 5
must be to ablation of any singlemechanismNeither ablationof carotid chemoreceptors nor of any other mechanism hasyet detected any such vulnerability of breathing in Man seefor example [29 30]
6 Can Stimulation Ablation and RecordingEstablish That Carotid Chemoreceptors AreNot the Key Sensor in Man
Falsifying any one of these three conditions could establishthat carotid chemoreceptors are not the key sensor Falsifi-cation however is difficult for two reasons First it is moredifficult to deal with negatives (no change in response) Forexample it is not yet possible to record neural activity directlyfrom the carotid chemoreceptors in Man and hence KarlSaganrsquos
ldquoabsence of evidence is not evidence of absencerdquoUltimately negatives are validated only by discovering whatmechanism does satisfy all three conditions Secondly thesafer conclusion from a negative result is initially ldquoincon-clusive without further experimentsrdquo because either someexperimental error occurred (eg the ablation was techni-cally unsuccessful or some measurement error occurred)or because the ablated mechanism is either irrelevant orredundant
Resolving ldquoinconclusiverdquo requires independent verifica-tion that ablation was successfully achieved that no experi-mental errors occurred and then identification and ablationof the second (or more) mechanism(s) There are equal dif-ficulties in identifying what might be the second duplicatingmechanism (and the quest to identify the second mechanismis beyond the scope of this review)Only if ablating the secondmechanism alone prevents matching could this finally estab-lish the irrelevance of the firstmechanism and the importanceof the second Whereas if matching was prevented onlyby ablating both both are important and duplicate Thisreview will also demonstrate that we are equally unable toestablish definitively that carotid chemoreceptors are not thekey metabolic rate sensor in Man
7 The Ideal Experimental Design with CarotidChemoreceptor Experiments
It is assumed that the same control system explains matchingfrom rest to maximum exercise rather than exercise beingsome special case This is important for two reasons Firstwhatever explains matching should be greater hence easierto detect as metabolic rate increases Secondly whateverexplains matching must explain it over the large increments(sim35- sim23- and sim21-fold) in the key variables ( Ve VO
2 and
VCO2) up to maximum exercise (ge19 kW)
There is also the scientific tenet that all experimentalresults should be confirmed by independent laboratories(every ldquofactrdquo should be referenced at least twice) Many of thecrucial carotid chemoreceptor experiments cited here haveonly ever been attempted once Furthermore there is noagreement on the ideal number of subjects per study Few usemore than 10 (often for good practical reasons) and many do
not even use 5 Statistical analysis can help (but not all applyit) although statistical power calculations are limited becausethey are for design rather than analysis and because they toodepend on the number of subjects
Positive results that are statistically significant (becausethe observed effect size is so large and its variance is so small)evenwith only two subjects are important and have sufficientpower But even these have two problems First there cannotbe complete confidence in variance being estimated fromonly 2 subjects Secondly there can be difficulty in convincingthe scientific community that these 2 subjects are sufficientlyrepresentative of the population
The particular problem with many of the key carotidchemoreceptor experiments onMan reviewed here howeveris that negative results occurred few subjects were usedthe observed effect size is small and the variance is largeFor example with only 2 subjects the observed effect sizeis often not statistically significant The difficulty then is indistinguishing a lack of power (false negative) from a truenegative Strictly statistical power analysis indicates only thatthe design requires more subjects which only confirms ouroriginal concern about low subject numbers
8 Recording Experiments fromHuman Arterial Blood
The principal physiological stimulants of the carotid bodiesare hypoxia hypercapnia and acidity Without the ability torecord from carotid chemoreceptor activity in Man mea-suring PaCO
2 PaO
2 and pHa levels is the only means of
investigating the recording condition for carotid chemore-ceptors that is are carotid chemoreceptors stimulated byraising metabolic rate
A fundamental requirement of a classical control systemusing feedback from a metabolic rate sensor to drive breath-ing is that arterial blood gases must change appropriatelyand proportionately with metabolic rate To produce theldquoerrorrdquo signal that drives breathing they must change in theappropriate directionmdashPaO
2falling and PaCO
2rising they
should change in proportion to exercise intensity and theymust remain changed as long as metabolic rate is increased inorder to sustain breathing at this new metabolic rate
Figures 4(a) and 4(b) show how large the changes inarterial blood gasesmust be for arterial blood gases to providethe key metabolic rate signal driving breathing at maximumexercise (breathing gt 100 Lsdotminminus1) But at maximum exercise(even in endurance trained athletes [31]) PaO
2decreases
neither transiently nor consistently enough to reach even theFiO2ldquothresholdrdquo in Figure 4(a) that stimulates breathing
Similarly PaCO2fails to rise enough Figures 1 and 2
show that there is barely any reproducible or sustained risein PaCO
2during moderate exercise and PaCO
2actually
falls during severe exercise So there is no PaCO2rise
during exercise sufficient to stimulate breathing even to the20 Lsdotminminus1 of Figure 4(b) So far the only observed bloodgas changes suitable to provide a metabolic rate signal are insystemic venous blood (Figure 1) Thus at present there is noobvious arterial factor that rises with metabolic rate and isknown to stimulate carotid chemoreceptors in Man
6 BioMed Research International
Equivalent altitude (km) calculated by Dripps0 5 10 15 20 25
40
30
20
10
100 209 18 16 14 12 10 8 6 4
500
400
300
200
100
Percent oxygen in inspired gas
102 80 66 51 40
Resp
irato
ry m
inut
evo
lum
e(L
min
)
Estimated equivalent PaO2 (mmHg)
Perc
enta
ge o
f nor
mal
resp
irato
ry m
inut
e vol
ume
(a)
20
18
16
12
8
4
32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62
PaCO2 (mmHg)
55 CO2
43 CO2
22 CO2
Air
plusmn1 se from mean respiratory minute volume
Ventilation (LBTPSmiddotminminus1)
(b)
Figure 4 (a) Relative insensitivity of breathing at rest to artificially lowering PO2in Man Minute ventilation (plusmn se solid bars plusmn sd open
bars) in normal subjects [87] as inspired oxygen is artificially lowered (strictly hypocapnic hypoxia exists once hyperventilation occurs)Equivalent PaO
2points are aligned on the FiO
2scale with PaO
2estimated before hyperventilation occurs using the alveolar gas equation
(assuming 760mmHg barometric pressure RQ = 08 PaO2= PAO
2 and PaCO
2= PACO
2) and the point afterwards is estimated based on
dynamic forcing experiments in isocapnia (courtesy of Dr G A Balanos) Reproducedwith permission fromDripps et al [87] (b) Sensitivityof breathing at rest to artificially raising PaCO
2in Man Minute ventilation and PaCO
2(femoral) in 8 healthy men [88] while inhaling 0ndash6
CO2in air at atmospheric pressure (mean slope is 25 Lsdotminminus1mmHgminus1 artificial PaCO
2rise) Reproduced with permission from Lambertsen
et al [88]
9 Absence of Suitable PaCO2Oscillations in Man
Another possibility is that to detect and signal metabolic ratecarotid chemoreceptors might detect some other propertyof O2or CO
2 such as the variability in PaO
2or PaCO
2
rather than their absolute levels Alveolar PCO2in Man does
oscillate systematically within the respiratory cycle at rest andis mirrored as oscillations in arterial blood pH and PaCO
2
[32ndash34] It was therefore thought that carotid chemoreceptorsmight estimate metabolic rate from the rate of rise or someother mathematical function of the PaCO
2oscillation But
there are several difficulties [18] with the CO2oscillation
hypothesis Thus
(i) It has not been established whether such oscillationscause the breathing pattern or whether the breathingpattern causes such oscillations
(ii) Such oscillations apparently disappear when breath-ing frequency rises during exercise Band et al [32]did find larger pH oscillations at exercise than at restbut only in one subject and only in the first minute ofexercise as tidal volume (but not breathing frequency)increased Whereas Murphy et al [33] found thatsuch oscillations disappeared in 4 subjects during thefirst minute of exercise (006 kW) when breathingat 20 breaths per minute (suggesting that it is slowbreathing that causes these oscillations and not viceversa)
(iii) The oscillation hypothesis is not supported by thedemonstration [35] that brief hypoxic pulses (N
2in
25ndash33 of the tidal volume) to the inspirate in 4subjects that cause detectable arterial desaturationat the carotid region (earlobe) failed to alter thefirst relevant respiratory cycle either at rest or inexercise (ge125Wew) and even if hypoxia (PAO
2
of 60mmHg) was used to increase chemoreceptoractivity
(iv) Denervation of carotid chemoreceptors (see [2930]mdash Section 13) apparently fails to prevent breathingmatchingmetabolic rate inMan yet these are the onlyknown chemoreceptors fast enough to detect suchoscillations in Man
There is no good reason to doubt any of these data Afurther difficulty is that it is not yet possible to test whetherblood gas changes precede a change in each breath becausesufficiently fast responding arterial electrodes are not gener-ally available [33] Using breath-by-breath measurement ofend tidal PetCO
2or PetO
2to estimate their corresponding
arterial levels preceding each breath is problematic becauseend tidal levels are not always reliable estimates of arteriallevels at rest and become progressively less so as systemicPvCO
2increases with metabolic rate (see Figure 1)
BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
(a) Rest
Cardiac output = 5 L blood per min
PaCO2 = 40mmHg(052L CO2 per L arterial blood)
Right heartLeft heart
Systemic venous chemoreceptors
Mixed Pv CO2 = 40 + 6mmHg(052 + 025L CO2 per L venous blood)
Systemic veins
CO2
CO2
Lungs
muscleSkeletal
Systemic arteries
Cardiac output = 25 L blood per min
PaCO2 = 35mmHg(049 L CO2 per L arterial blood)
Carotidchemoreceptors
Centralchemoreceptors
Mixed Pv CO2 = 35 + 45mmHg(049 + 4525L CO2 per L venous blood)
Each L venous blood gains 004 L CO2
times 5 Lmin cardiac output = 02L CO2 carriedEach L venous blood gains 018 L CO2
times 25 Lmin cardiac output = 45L CO2 carried
(b) Exercise
= 02 L CO2min = 45 L CO2min
= 45 L CO2min= 02 L CO2min
VCO2 VCO2
VCO2 VCO2
Figure 3 Systemic arterial chemoreceptors are ideally located anatomically to measure any mismatch but are in the wrong place to measuremetabolic rate or metabolically produced CO
2 Diagram of typical values of cardiac output systemic arterial PaCO
2and mixed PvCO
2
and of blood CO2content at rest (a) and maximum exercise (b) with data from multiple sources For clarity the corresponding numbers
for metabolic rates are omitted Comparison of (a) and (b) shows that systemic arterial chemoreceptors are in the wrong place to measuremetabolically produced CO
2 because metabolically produced CO
2is carried via the systemic veins frommuscle to air without PaCO
2rising
To establish the carotid chemoreceptors as the metabolicrate sensor all three conditions must be satisfied Eitheralone is inadequate for example (1) alone might also beachieved by an artificial stimulus for example a drug (2)alone might merely represent some traumatic or accidentalside effect and (3) alone establishes only correlation butnot causation This review will consider each condition forcarotid chemoreceptors but before doing so some furtherpoints must first be considered
5 The Redundancy Summation andMultiplication Beliefs
All ablation experiments in exercise so far fail to abolishbreathing matching metabolic rate Rather than concludingthat the ablated factors are irrelevant it is usually concludedinstead that at least two sensor mechanisms must exist eachof which could mediate matching on its own but at anymoment one or more must contribute (ie both duplicate orare redundant) This is one of two common beliefs
(i) the redundancy (or duplication) belief
nature would not have designed such an impor-tant control system to rely on only one mechanism so at least two factors must control
(ii) the multiplication belief
hyperpnea of exercise due to multiplicationof a great number of discrete impulses whichare individually small but collectively large
These beliefs are automatically applied when interpretingcarotid chemoreceptor experiments and are never chal-lenged Both are scientifically testable in Man but there is nosubstantial experimental evidence yet for either
The redundancy belief means that wemust already acceptthat at least twomechanisms sense metabolic rate (ie one inaddition to carotid chemoreceptors) This in turn comes upagainst Occamrsquos razor
(essentia non sunt multiplicanda praeter necessi-tatem)
it is naıve to propose multiple mechanisms before firstestablishing one Yet so far we cannot even establish one Nostimulation of a singlemechanismhas yet increased breathingby more than sim15 Lsdotmin-1 [27 28] that is to nowherenear that of maximum exercise The principal interest in theredundancy belief however is the difficulty that it creates ininterpreting the negative effect of an ablation experiment inMan as further explained in Section 6
The multiplication belief predicts that the more multipli-cation the interaction requires themore vulnerable breathing
BioMed Research International 5
must be to ablation of any singlemechanismNeither ablationof carotid chemoreceptors nor of any other mechanism hasyet detected any such vulnerability of breathing in Man seefor example [29 30]
6 Can Stimulation Ablation and RecordingEstablish That Carotid Chemoreceptors AreNot the Key Sensor in Man
Falsifying any one of these three conditions could establishthat carotid chemoreceptors are not the key sensor Falsifi-cation however is difficult for two reasons First it is moredifficult to deal with negatives (no change in response) Forexample it is not yet possible to record neural activity directlyfrom the carotid chemoreceptors in Man and hence KarlSaganrsquos
ldquoabsence of evidence is not evidence of absencerdquoUltimately negatives are validated only by discovering whatmechanism does satisfy all three conditions Secondly thesafer conclusion from a negative result is initially ldquoincon-clusive without further experimentsrdquo because either someexperimental error occurred (eg the ablation was techni-cally unsuccessful or some measurement error occurred)or because the ablated mechanism is either irrelevant orredundant
Resolving ldquoinconclusiverdquo requires independent verifica-tion that ablation was successfully achieved that no experi-mental errors occurred and then identification and ablationof the second (or more) mechanism(s) There are equal dif-ficulties in identifying what might be the second duplicatingmechanism (and the quest to identify the second mechanismis beyond the scope of this review)Only if ablating the secondmechanism alone prevents matching could this finally estab-lish the irrelevance of the firstmechanism and the importanceof the second Whereas if matching was prevented onlyby ablating both both are important and duplicate Thisreview will also demonstrate that we are equally unable toestablish definitively that carotid chemoreceptors are not thekey metabolic rate sensor in Man
7 The Ideal Experimental Design with CarotidChemoreceptor Experiments
It is assumed that the same control system explains matchingfrom rest to maximum exercise rather than exercise beingsome special case This is important for two reasons Firstwhatever explains matching should be greater hence easierto detect as metabolic rate increases Secondly whateverexplains matching must explain it over the large increments(sim35- sim23- and sim21-fold) in the key variables ( Ve VO
2 and
VCO2) up to maximum exercise (ge19 kW)
There is also the scientific tenet that all experimentalresults should be confirmed by independent laboratories(every ldquofactrdquo should be referenced at least twice) Many of thecrucial carotid chemoreceptor experiments cited here haveonly ever been attempted once Furthermore there is noagreement on the ideal number of subjects per study Few usemore than 10 (often for good practical reasons) and many do
not even use 5 Statistical analysis can help (but not all applyit) although statistical power calculations are limited becausethey are for design rather than analysis and because they toodepend on the number of subjects
Positive results that are statistically significant (becausethe observed effect size is so large and its variance is so small)evenwith only two subjects are important and have sufficientpower But even these have two problems First there cannotbe complete confidence in variance being estimated fromonly 2 subjects Secondly there can be difficulty in convincingthe scientific community that these 2 subjects are sufficientlyrepresentative of the population
The particular problem with many of the key carotidchemoreceptor experiments onMan reviewed here howeveris that negative results occurred few subjects were usedthe observed effect size is small and the variance is largeFor example with only 2 subjects the observed effect sizeis often not statistically significant The difficulty then is indistinguishing a lack of power (false negative) from a truenegative Strictly statistical power analysis indicates only thatthe design requires more subjects which only confirms ouroriginal concern about low subject numbers
8 Recording Experiments fromHuman Arterial Blood
The principal physiological stimulants of the carotid bodiesare hypoxia hypercapnia and acidity Without the ability torecord from carotid chemoreceptor activity in Man mea-suring PaCO
2 PaO
2 and pHa levels is the only means of
investigating the recording condition for carotid chemore-ceptors that is are carotid chemoreceptors stimulated byraising metabolic rate
A fundamental requirement of a classical control systemusing feedback from a metabolic rate sensor to drive breath-ing is that arterial blood gases must change appropriatelyand proportionately with metabolic rate To produce theldquoerrorrdquo signal that drives breathing they must change in theappropriate directionmdashPaO
2falling and PaCO
2rising they
should change in proportion to exercise intensity and theymust remain changed as long as metabolic rate is increased inorder to sustain breathing at this new metabolic rate
Figures 4(a) and 4(b) show how large the changes inarterial blood gasesmust be for arterial blood gases to providethe key metabolic rate signal driving breathing at maximumexercise (breathing gt 100 Lsdotminminus1) But at maximum exercise(even in endurance trained athletes [31]) PaO
2decreases
neither transiently nor consistently enough to reach even theFiO2ldquothresholdrdquo in Figure 4(a) that stimulates breathing
Similarly PaCO2fails to rise enough Figures 1 and 2
show that there is barely any reproducible or sustained risein PaCO
2during moderate exercise and PaCO
2actually
falls during severe exercise So there is no PaCO2rise
during exercise sufficient to stimulate breathing even to the20 Lsdotminminus1 of Figure 4(b) So far the only observed bloodgas changes suitable to provide a metabolic rate signal are insystemic venous blood (Figure 1) Thus at present there is noobvious arterial factor that rises with metabolic rate and isknown to stimulate carotid chemoreceptors in Man
6 BioMed Research International
Equivalent altitude (km) calculated by Dripps0 5 10 15 20 25
40
30
20
10
100 209 18 16 14 12 10 8 6 4
500
400
300
200
100
Percent oxygen in inspired gas
102 80 66 51 40
Resp
irato
ry m
inut
evo
lum
e(L
min
)
Estimated equivalent PaO2 (mmHg)
Perc
enta
ge o
f nor
mal
resp
irato
ry m
inut
e vol
ume
(a)
20
18
16
12
8
4
32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62
PaCO2 (mmHg)
55 CO2
43 CO2
22 CO2
Air
plusmn1 se from mean respiratory minute volume
Ventilation (LBTPSmiddotminminus1)
(b)
Figure 4 (a) Relative insensitivity of breathing at rest to artificially lowering PO2in Man Minute ventilation (plusmn se solid bars plusmn sd open
bars) in normal subjects [87] as inspired oxygen is artificially lowered (strictly hypocapnic hypoxia exists once hyperventilation occurs)Equivalent PaO
2points are aligned on the FiO
2scale with PaO
2estimated before hyperventilation occurs using the alveolar gas equation
(assuming 760mmHg barometric pressure RQ = 08 PaO2= PAO
2 and PaCO
2= PACO
2) and the point afterwards is estimated based on
dynamic forcing experiments in isocapnia (courtesy of Dr G A Balanos) Reproducedwith permission fromDripps et al [87] (b) Sensitivityof breathing at rest to artificially raising PaCO
2in Man Minute ventilation and PaCO
2(femoral) in 8 healthy men [88] while inhaling 0ndash6
CO2in air at atmospheric pressure (mean slope is 25 Lsdotminminus1mmHgminus1 artificial PaCO
2rise) Reproduced with permission from Lambertsen
et al [88]
9 Absence of Suitable PaCO2Oscillations in Man
Another possibility is that to detect and signal metabolic ratecarotid chemoreceptors might detect some other propertyof O2or CO
2 such as the variability in PaO
2or PaCO
2
rather than their absolute levels Alveolar PCO2in Man does
oscillate systematically within the respiratory cycle at rest andis mirrored as oscillations in arterial blood pH and PaCO
2
[32ndash34] It was therefore thought that carotid chemoreceptorsmight estimate metabolic rate from the rate of rise or someother mathematical function of the PaCO
2oscillation But
there are several difficulties [18] with the CO2oscillation
hypothesis Thus
(i) It has not been established whether such oscillationscause the breathing pattern or whether the breathingpattern causes such oscillations
(ii) Such oscillations apparently disappear when breath-ing frequency rises during exercise Band et al [32]did find larger pH oscillations at exercise than at restbut only in one subject and only in the first minute ofexercise as tidal volume (but not breathing frequency)increased Whereas Murphy et al [33] found thatsuch oscillations disappeared in 4 subjects during thefirst minute of exercise (006 kW) when breathingat 20 breaths per minute (suggesting that it is slowbreathing that causes these oscillations and not viceversa)
(iii) The oscillation hypothesis is not supported by thedemonstration [35] that brief hypoxic pulses (N
2in
25ndash33 of the tidal volume) to the inspirate in 4subjects that cause detectable arterial desaturationat the carotid region (earlobe) failed to alter thefirst relevant respiratory cycle either at rest or inexercise (ge125Wew) and even if hypoxia (PAO
2
of 60mmHg) was used to increase chemoreceptoractivity
(iv) Denervation of carotid chemoreceptors (see [2930]mdash Section 13) apparently fails to prevent breathingmatchingmetabolic rate inMan yet these are the onlyknown chemoreceptors fast enough to detect suchoscillations in Man
There is no good reason to doubt any of these data Afurther difficulty is that it is not yet possible to test whetherblood gas changes precede a change in each breath becausesufficiently fast responding arterial electrodes are not gener-ally available [33] Using breath-by-breath measurement ofend tidal PetCO
2or PetO
2to estimate their corresponding
arterial levels preceding each breath is problematic becauseend tidal levels are not always reliable estimates of arteriallevels at rest and become progressively less so as systemicPvCO
2increases with metabolic rate (see Figure 1)
BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
must be to ablation of any singlemechanismNeither ablationof carotid chemoreceptors nor of any other mechanism hasyet detected any such vulnerability of breathing in Man seefor example [29 30]
6 Can Stimulation Ablation and RecordingEstablish That Carotid Chemoreceptors AreNot the Key Sensor in Man
Falsifying any one of these three conditions could establishthat carotid chemoreceptors are not the key sensor Falsifi-cation however is difficult for two reasons First it is moredifficult to deal with negatives (no change in response) Forexample it is not yet possible to record neural activity directlyfrom the carotid chemoreceptors in Man and hence KarlSaganrsquos
ldquoabsence of evidence is not evidence of absencerdquoUltimately negatives are validated only by discovering whatmechanism does satisfy all three conditions Secondly thesafer conclusion from a negative result is initially ldquoincon-clusive without further experimentsrdquo because either someexperimental error occurred (eg the ablation was techni-cally unsuccessful or some measurement error occurred)or because the ablated mechanism is either irrelevant orredundant
Resolving ldquoinconclusiverdquo requires independent verifica-tion that ablation was successfully achieved that no experi-mental errors occurred and then identification and ablationof the second (or more) mechanism(s) There are equal dif-ficulties in identifying what might be the second duplicatingmechanism (and the quest to identify the second mechanismis beyond the scope of this review)Only if ablating the secondmechanism alone prevents matching could this finally estab-lish the irrelevance of the firstmechanism and the importanceof the second Whereas if matching was prevented onlyby ablating both both are important and duplicate Thisreview will also demonstrate that we are equally unable toestablish definitively that carotid chemoreceptors are not thekey metabolic rate sensor in Man
7 The Ideal Experimental Design with CarotidChemoreceptor Experiments
It is assumed that the same control system explains matchingfrom rest to maximum exercise rather than exercise beingsome special case This is important for two reasons Firstwhatever explains matching should be greater hence easierto detect as metabolic rate increases Secondly whateverexplains matching must explain it over the large increments(sim35- sim23- and sim21-fold) in the key variables ( Ve VO
2 and
VCO2) up to maximum exercise (ge19 kW)
There is also the scientific tenet that all experimentalresults should be confirmed by independent laboratories(every ldquofactrdquo should be referenced at least twice) Many of thecrucial carotid chemoreceptor experiments cited here haveonly ever been attempted once Furthermore there is noagreement on the ideal number of subjects per study Few usemore than 10 (often for good practical reasons) and many do
not even use 5 Statistical analysis can help (but not all applyit) although statistical power calculations are limited becausethey are for design rather than analysis and because they toodepend on the number of subjects
Positive results that are statistically significant (becausethe observed effect size is so large and its variance is so small)evenwith only two subjects are important and have sufficientpower But even these have two problems First there cannotbe complete confidence in variance being estimated fromonly 2 subjects Secondly there can be difficulty in convincingthe scientific community that these 2 subjects are sufficientlyrepresentative of the population
The particular problem with many of the key carotidchemoreceptor experiments onMan reviewed here howeveris that negative results occurred few subjects were usedthe observed effect size is small and the variance is largeFor example with only 2 subjects the observed effect sizeis often not statistically significant The difficulty then is indistinguishing a lack of power (false negative) from a truenegative Strictly statistical power analysis indicates only thatthe design requires more subjects which only confirms ouroriginal concern about low subject numbers
8 Recording Experiments fromHuman Arterial Blood
The principal physiological stimulants of the carotid bodiesare hypoxia hypercapnia and acidity Without the ability torecord from carotid chemoreceptor activity in Man mea-suring PaCO
2 PaO
2 and pHa levels is the only means of
investigating the recording condition for carotid chemore-ceptors that is are carotid chemoreceptors stimulated byraising metabolic rate
A fundamental requirement of a classical control systemusing feedback from a metabolic rate sensor to drive breath-ing is that arterial blood gases must change appropriatelyand proportionately with metabolic rate To produce theldquoerrorrdquo signal that drives breathing they must change in theappropriate directionmdashPaO
2falling and PaCO
2rising they
should change in proportion to exercise intensity and theymust remain changed as long as metabolic rate is increased inorder to sustain breathing at this new metabolic rate
Figures 4(a) and 4(b) show how large the changes inarterial blood gasesmust be for arterial blood gases to providethe key metabolic rate signal driving breathing at maximumexercise (breathing gt 100 Lsdotminminus1) But at maximum exercise(even in endurance trained athletes [31]) PaO
2decreases
neither transiently nor consistently enough to reach even theFiO2ldquothresholdrdquo in Figure 4(a) that stimulates breathing
Similarly PaCO2fails to rise enough Figures 1 and 2
show that there is barely any reproducible or sustained risein PaCO
2during moderate exercise and PaCO
2actually
falls during severe exercise So there is no PaCO2rise
during exercise sufficient to stimulate breathing even to the20 Lsdotminminus1 of Figure 4(b) So far the only observed bloodgas changes suitable to provide a metabolic rate signal are insystemic venous blood (Figure 1) Thus at present there is noobvious arterial factor that rises with metabolic rate and isknown to stimulate carotid chemoreceptors in Man
6 BioMed Research International
Equivalent altitude (km) calculated by Dripps0 5 10 15 20 25
40
30
20
10
100 209 18 16 14 12 10 8 6 4
500
400
300
200
100
Percent oxygen in inspired gas
102 80 66 51 40
Resp
irato
ry m
inut
evo
lum
e(L
min
)
Estimated equivalent PaO2 (mmHg)
Perc
enta
ge o
f nor
mal
resp
irato
ry m
inut
e vol
ume
(a)
20
18
16
12
8
4
32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62
PaCO2 (mmHg)
55 CO2
43 CO2
22 CO2
Air
plusmn1 se from mean respiratory minute volume
Ventilation (LBTPSmiddotminminus1)
(b)
Figure 4 (a) Relative insensitivity of breathing at rest to artificially lowering PO2in Man Minute ventilation (plusmn se solid bars plusmn sd open
bars) in normal subjects [87] as inspired oxygen is artificially lowered (strictly hypocapnic hypoxia exists once hyperventilation occurs)Equivalent PaO
2points are aligned on the FiO
2scale with PaO
2estimated before hyperventilation occurs using the alveolar gas equation
(assuming 760mmHg barometric pressure RQ = 08 PaO2= PAO
2 and PaCO
2= PACO
2) and the point afterwards is estimated based on
dynamic forcing experiments in isocapnia (courtesy of Dr G A Balanos) Reproducedwith permission fromDripps et al [87] (b) Sensitivityof breathing at rest to artificially raising PaCO
2in Man Minute ventilation and PaCO
2(femoral) in 8 healthy men [88] while inhaling 0ndash6
CO2in air at atmospheric pressure (mean slope is 25 Lsdotminminus1mmHgminus1 artificial PaCO
2rise) Reproduced with permission from Lambertsen
et al [88]
9 Absence of Suitable PaCO2Oscillations in Man
Another possibility is that to detect and signal metabolic ratecarotid chemoreceptors might detect some other propertyof O2or CO
2 such as the variability in PaO
2or PaCO
2
rather than their absolute levels Alveolar PCO2in Man does
oscillate systematically within the respiratory cycle at rest andis mirrored as oscillations in arterial blood pH and PaCO
2
[32ndash34] It was therefore thought that carotid chemoreceptorsmight estimate metabolic rate from the rate of rise or someother mathematical function of the PaCO
2oscillation But
there are several difficulties [18] with the CO2oscillation
hypothesis Thus
(i) It has not been established whether such oscillationscause the breathing pattern or whether the breathingpattern causes such oscillations
(ii) Such oscillations apparently disappear when breath-ing frequency rises during exercise Band et al [32]did find larger pH oscillations at exercise than at restbut only in one subject and only in the first minute ofexercise as tidal volume (but not breathing frequency)increased Whereas Murphy et al [33] found thatsuch oscillations disappeared in 4 subjects during thefirst minute of exercise (006 kW) when breathingat 20 breaths per minute (suggesting that it is slowbreathing that causes these oscillations and not viceversa)
(iii) The oscillation hypothesis is not supported by thedemonstration [35] that brief hypoxic pulses (N
2in
25ndash33 of the tidal volume) to the inspirate in 4subjects that cause detectable arterial desaturationat the carotid region (earlobe) failed to alter thefirst relevant respiratory cycle either at rest or inexercise (ge125Wew) and even if hypoxia (PAO
2
of 60mmHg) was used to increase chemoreceptoractivity
(iv) Denervation of carotid chemoreceptors (see [2930]mdash Section 13) apparently fails to prevent breathingmatchingmetabolic rate inMan yet these are the onlyknown chemoreceptors fast enough to detect suchoscillations in Man
There is no good reason to doubt any of these data Afurther difficulty is that it is not yet possible to test whetherblood gas changes precede a change in each breath becausesufficiently fast responding arterial electrodes are not gener-ally available [33] Using breath-by-breath measurement ofend tidal PetCO
2or PetO
2to estimate their corresponding
arterial levels preceding each breath is problematic becauseend tidal levels are not always reliable estimates of arteriallevels at rest and become progressively less so as systemicPvCO
2increases with metabolic rate (see Figure 1)
BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
Equivalent altitude (km) calculated by Dripps0 5 10 15 20 25
40
30
20
10
100 209 18 16 14 12 10 8 6 4
500
400
300
200
100
Percent oxygen in inspired gas
102 80 66 51 40
Resp
irato
ry m
inut
evo
lum
e(L
min
)
Estimated equivalent PaO2 (mmHg)
Perc
enta
ge o
f nor
mal
resp
irato
ry m
inut
e vol
ume
(a)
20
18
16
12
8
4
32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62
PaCO2 (mmHg)
55 CO2
43 CO2
22 CO2
Air
plusmn1 se from mean respiratory minute volume
Ventilation (LBTPSmiddotminminus1)
(b)
Figure 4 (a) Relative insensitivity of breathing at rest to artificially lowering PO2in Man Minute ventilation (plusmn se solid bars plusmn sd open
bars) in normal subjects [87] as inspired oxygen is artificially lowered (strictly hypocapnic hypoxia exists once hyperventilation occurs)Equivalent PaO
2points are aligned on the FiO
2scale with PaO
2estimated before hyperventilation occurs using the alveolar gas equation
(assuming 760mmHg barometric pressure RQ = 08 PaO2= PAO
2 and PaCO
2= PACO
2) and the point afterwards is estimated based on
dynamic forcing experiments in isocapnia (courtesy of Dr G A Balanos) Reproducedwith permission fromDripps et al [87] (b) Sensitivityof breathing at rest to artificially raising PaCO
2in Man Minute ventilation and PaCO
2(femoral) in 8 healthy men [88] while inhaling 0ndash6
CO2in air at atmospheric pressure (mean slope is 25 Lsdotminminus1mmHgminus1 artificial PaCO
2rise) Reproduced with permission from Lambertsen
et al [88]
9 Absence of Suitable PaCO2Oscillations in Man
Another possibility is that to detect and signal metabolic ratecarotid chemoreceptors might detect some other propertyof O2or CO
2 such as the variability in PaO
2or PaCO
2
rather than their absolute levels Alveolar PCO2in Man does
oscillate systematically within the respiratory cycle at rest andis mirrored as oscillations in arterial blood pH and PaCO
2
[32ndash34] It was therefore thought that carotid chemoreceptorsmight estimate metabolic rate from the rate of rise or someother mathematical function of the PaCO
2oscillation But
there are several difficulties [18] with the CO2oscillation
hypothesis Thus
(i) It has not been established whether such oscillationscause the breathing pattern or whether the breathingpattern causes such oscillations
(ii) Such oscillations apparently disappear when breath-ing frequency rises during exercise Band et al [32]did find larger pH oscillations at exercise than at restbut only in one subject and only in the first minute ofexercise as tidal volume (but not breathing frequency)increased Whereas Murphy et al [33] found thatsuch oscillations disappeared in 4 subjects during thefirst minute of exercise (006 kW) when breathingat 20 breaths per minute (suggesting that it is slowbreathing that causes these oscillations and not viceversa)
(iii) The oscillation hypothesis is not supported by thedemonstration [35] that brief hypoxic pulses (N
2in
25ndash33 of the tidal volume) to the inspirate in 4subjects that cause detectable arterial desaturationat the carotid region (earlobe) failed to alter thefirst relevant respiratory cycle either at rest or inexercise (ge125Wew) and even if hypoxia (PAO
2
of 60mmHg) was used to increase chemoreceptoractivity
(iv) Denervation of carotid chemoreceptors (see [2930]mdash Section 13) apparently fails to prevent breathingmatchingmetabolic rate inMan yet these are the onlyknown chemoreceptors fast enough to detect suchoscillations in Man
There is no good reason to doubt any of these data Afurther difficulty is that it is not yet possible to test whetherblood gas changes precede a change in each breath becausesufficiently fast responding arterial electrodes are not gener-ally available [33] Using breath-by-breath measurement ofend tidal PetCO
2or PetO
2to estimate their corresponding
arterial levels preceding each breath is problematic becauseend tidal levels are not always reliable estimates of arteriallevels at rest and become progressively less so as systemicPvCO
2increases with metabolic rate (see Figure 1)
BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7
10 Absence of Suitable Changes inCO2 Sensitivity in Man
Another possibility is that whatever initiates exercise mightinstantly change the sensitivity of arterial chemoreceptors toblood gases [36] There are two difficulties however with theCO2sensitivity hypothesis
First since PaCO2fails to rise consistently by more
than sim2mmHg in exercise the CO2sensitivity of breathing
would have to reach at least 50 Lsdotminminus1sdotmmHgminus1 PaCO2to
explain breathing reaching 100 Lsdotminminus1 Yet numerous studies(eg [37ndash41]) show that the measured CO
2sensitivity to
artificially induced CO2rises in exercise either falls does
not change or rises to no more than sim7 Lsdotminminus1sdotmmHgminus1PaCO
2 Secondly when faced with the arterial hypocapnia of
severe exercise that apparently lowers PaCO2even to below
the apnea threshold [42] any important mechanism relyingsolely on increased CO
2sensitivity should cause apnea Yet
clearly apnea does not occur in severe exerciseFuture experiments could still find some new function
of blood gas levels or of arterial blood (eg [H+]amdashseeSection 16) that does change appropriately with metabolicrate and hence could still confirm carotid chemoreceptors asthe keymetabolic rate sensor driving breathing But until theydo there remains no variable in arterial blood that rises withmetabolic rate and stimulates carotid chemoreceptors
11 Is There an Unknown Substancein Arterial Blood That Rises withMetabolic Rate and Stimulates CarotidChemoreceptors to Control Breathingin Exercise
Numerous other blood-borne substances have been con-sidered as candidates to link carotid chemoreceptors tometabolic rate and the experiments with potassium (K+)levels provide a good example of the difficulties with all othersuitable candidates meeting the stimulation ablation andrecording criteria Thus while arterial (and venous) plasmaK+ levels in Man do rise from sim4 to 7mM between rest andexercise [43] lowering plasma K+ levels in Man from 7 to5mM fails to alter breathing [44] Infusing K+ intravenouslyis too dangerous in Man but infusion to mimic the riseto 5ndash8mM in Man produces little increase in breathingin monkeys [45] and none in goats [46] Furthermore asdiscussed below the fact that carotid chemoreceptor dener-vation fails to prevent breathing matching metabolic rate inMan is evidence that no arterial blood-borne ldquofactor 119883rdquo thatstimulates carotid chemoreceptors has been missed
12 Carotid ChemoreceptorStimulation Experiments
The failure to detect a consistent rise in PaCO2or fall in PaO
2
during exercise has led some to question the relevance toexercise of any experiments that artificially induce hypoxiaand hypercapnia to stimulate carotid chemoreceptors Nev-ertheless such experiments are extensively pursued These
show that artificially manipulating the inspired CO2or O2
levels to stimulate carotid chemoreceptors fails to causehyperventilation to the Ve levels anywhere near those seenat maximum exercise (100ndash150 Lsdotminminus1)
Figure 4(a) shows that even lowering FiO2levels to
around 4 in resting subjects barely raisedminute ventilationgt30 Lsdotminminus1 Yet this is the severest hypoxia tolerable sincehumans start to pass out at FiO
2levels below 6 PaO
2at
maximum exercise does not consistently fall at all and thelowest levels recorded (75mmHg [31]) do not even reach theldquothresholdrdquo to stimulate breathing in Figure 4(a)
Clearly breathing inMan ismore sensitive to small rises inPaCO
2than to falls in PaO
2 for example artificially lowering
eupneic PaCO2by more than sim5mmHg at rest can cause
apnea [42] when untangled from the additional voluntarydrive to breathe [47] Figure 4(b) shows that artificiallyraising PaCO
2to 50mmHg causes a hyperventilation to only
about 20 Lsdotminminus1 Yet Figures 1 and 2 show thatwithmoderateexercise PaCO
2fails to rise consistently above sim45mmHg
and it actually falls at maximum exerciseStrictly the breathing response to hypoxia (Figure 4(a)) is
reduced by hypocapnia [48] and accentuated by hypercapniathat to hypercapnia (Figure 4(b)) is accentuated by hypoxia[49] and the combination of artificially induced hypoxiaand hypercapnia (ie asphyxia) is a much more potentstimulant of breathing than either alone Nevertheless thefailure to detect asphyxic blood gas changes during normalexercise questions the value of attempting to find out what isthe maximum minute ventilation achievable with asphyxiaFor the same reason further investigation of Figure 4(a)outside the isocapnic range and of Figure 4(b) outside theiso-oxic range does not appear relevant Thus not only hasno suitable factor that stimulates carotid chemoreceptorsbeen found to rise in arterial blood but also artificiallyinducing large changes in arterial oxygen or carbon dioxidelevels in Man to deliberately provide intense stimulation ofcarotid chemoreceptors fails to stimulate breathing to levelsanywhere near those found at maximum exercise
13 Carotid ChemoreceptorAblation Experiments
Carotid bodies exist in Man [50] There is not a doubtthat hypoxia and hypercapnia stimulate them and rapidlystimulate breathing (Figures 4(a) and 4(b)) In Man aftersurgical denervation of carotid chemoreceptors bilaterally[51ndash55] both the rapid component of the hyperventilationto raised PaCO
2and the hyperventilation following hypoxia
appear absent (although how such hypoxia relates to theFiO2levels in Figure 4(a) is not always clear) Moreover the
hyperventilation to an FiO2of 8 is temporarily abolished
by injection of local anaesthetic to block their afferent nerves[56] Unlike in other species this abolition of the ventilatoryresponse is permanent inMan [53 54] although intriguinglysuch patients retain however a small and residual responseto the Dejours test [53 57] which therefore ldquois not whollyattributable to suppression of carotid bodiesrdquo [57] The initialrapid time course of the increased breathing to raised PaCO
2
8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
fits a single exponential [51 52] so is assumed to involve onlyone sensor mechanism the carotid chemoreceptors
Available evidence indicates that carotid chemorecep-tor ablation in Man does not prevent breathing matchingmetabolic rate either at rest or during moderate exerciseIf carotid chemoreceptors did cause matching the simplestconsequence of their ablation should be marked instabilityof both blood gases and breathing on a breath-by-breathbasis both at rest and during exercise Blood gas levels shouldchange so much that other responses should be triggered forexample unconsciousness at PaO
2below sim25mmHg andor
PaCO2above sim90mmHg and hypocapnic tetany at PaCO
2
levels below 20mmHg (acute hyperoxia being innocuous)Breathing both at rest and during exercise should also behighly unstable and oscillate between periods of apnea andsevere hyperventilation Yet after such ablation in Man noconsistent change has been observed in either arterial bloodgases or breathing neither at rest nor during exercise
131 Effects on Breathing of Carotid Chemoreceptor Ablation atRest Of the most recent studies Fatemian et al [52] foundthat resting PetCO
2levels (45 plusmn 1mmHg) in patients with
bilateral carotid body removal for glomus tumours (chemod-ectoma) were 5 plusmn 1mmHg higher than in healthy volunteerswhereas other studies with resting patients after bilateralcarotid body removal (to treat asthma) found no changein PaCO
2or PetCO
2versus healthy controls [30 53 54]
despite asthma itself if severe enough undoubtedly causingCO2retention that is PaCO
2levels at up to 6mmHg higher
than in controls [58 59] Dahan et al [51] found that restingPetCO
2levels in 3 chemodectoma patients were 7mmHg
higher (at 41 plusmn 1mmHg) after than before denervation Evenif Dahanrsquos study is accepted as the best evidence that PaCO
2
does rise slightly their PetCO2of 41mmHg is still within the
normal range in healthy subjects [60]It is not therefore obvious that any substantial mismatch
between breathing and metabolic rate exists in such patientsat rest nor is their breathing pattern at rest obviouslydifferent although new and more detailed studies of breath-by-breath variability at rest might still reveal subtle deficits
132 Effects on Breathing of Carotid Chemoreceptor Ablationduring Exercise No study has established that breath-by-breath control of PaCO
2nor the hyperpnea of exercise
is obviously abolished after carotid chemodenervation Sofar only two groups have tested such patients in exerciseHonda et al [29] found that subjects could still exerciseto 50Wew but PCO2 rose by 3mmHg (to 46 plusmn 2mmHg)whereas Wasserman et al [30] found (Figure 5) that bilateraldenervation during more intense exercise did not preventbreathing from matching metabolic rate that is
ldquothe carbon dioxide blood gas tensions remained unchanged in exerciserdquo at50119882119890119908(119899 = 7)
and
ldquoa marked transient overshoot in the PaCO2
prior to attaining a phase III response in which
PaCO2was indistinguishable from resting levelsrdquo
[11] at 98W119890119908
exercise (119899 = 6) [30]
Note precisely what the PaCO2levels are in Figure 5(a)
and compare them with Figures 1 2(b) and 2(c)This negative result appears validated by their positive
result [54] that the stimulation of breathing with an FiO2
of 12 was absent in their carotid body denervated patientsIn fact Figure 4(a) indicates that using an FiO
2of sim8 [56]
would be definitive but the precise FiO2ldquothresholdrdquo at which
the stimulation of breathing first occurs appears never tohave been deliberately measured with modern measurementand statistical analysis techniques Thus ablation appearstechnically successful and no obvious measurement errorsoccurred
It is not reasonable to propose that such ablation makesthe control system too abnormal to be relevant because it isnot particularly abnormal Moreover although such ablationapparently abolishes the stimulation of breathing by hypoxiaits ablation is no hindrance since such hypoxia does notoccur during exercise
Correct interpretation of these experiments is crucialTheusual interpretation is that ldquocarotid chemoreceptors must stillbe important and redundancy must existrdquo But a more realisticinterpretation is ldquoinconclusive without further experimentsrdquobecause these results demonstrate that either the chemore-ceptors must be one of at least two important mechanismsthat sense metabolic rate (ie are duplicating) or have noimportance
In either case this shows that at least one mechanismother than carotid chemoreceptors must also account forbreathing matching metabolic rate This other mechanismremains unidentified and until it is we cannot pursueexperiments to distinguish its role from that of carotidchemoreceptors and hence to distinguish between carotidchemoreceptors being redundant or irrelevant
14 Aortic and Central Chemoreceptors TooFail to Sustain Breathing after Ablation ofCarotid Chemoreceptors in Man
How do carotid chemodenervated patients maintain rela-tively normal breathing at rest during sleep wakefulness andexercise This question is still unanswered but it appearsunlikely that the second mechanism involves either aorticor central chemoreceptors acting as the key metabolic ratesensor because they too have no known means of measuringmetabolic rate
Aortic bodies do exist in humans [50] theymay have car-diovascular effects [61] but have no known effects on breath-ing in Man Therefore there is no evidence that they canact as the metabolic sensor Thus carotid chemodenervation(while the aortic chemoreceptors are still intact) apparentlyabolishes the breathing response to hypoxia permanently[51 52 54 55] and that additional bilateral anaesthesia ofthe vagus nerve (in which aortic afferents travel) produces nofurther deficits [56] So not only do aortic chemoreceptorsnot affect breathing in Man but also they too have no known
BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
70 anaerobic threshold 150 anaerobic threshold
740
735
730
740
735
730
pH
45
40
35
45
40
35
PaCO
2(m
mH
g)
10 20 10 2030Minutes of exercise Minutes of exercise
ControlCBR
ControlCBR
Rest uarrRest uarr
(a)
70 anaerobic threshold 150 anaerobic threshold
40
30
10
0
40
30
20
10
0Control Denervated Control Denervated
lowast
ns
VeV
CO2
(b)
70 anaerobic threshold 150 anaerobic threshold
100
50
0
100
50
00 2 4 60 2 4 6 8
Minutes of exercise Minutes of exercise
Normal
CBRNormal
CBR
(per
cent
age o
f to
tal c
hang
e)Ve
(c)
Figure 5 Continued
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
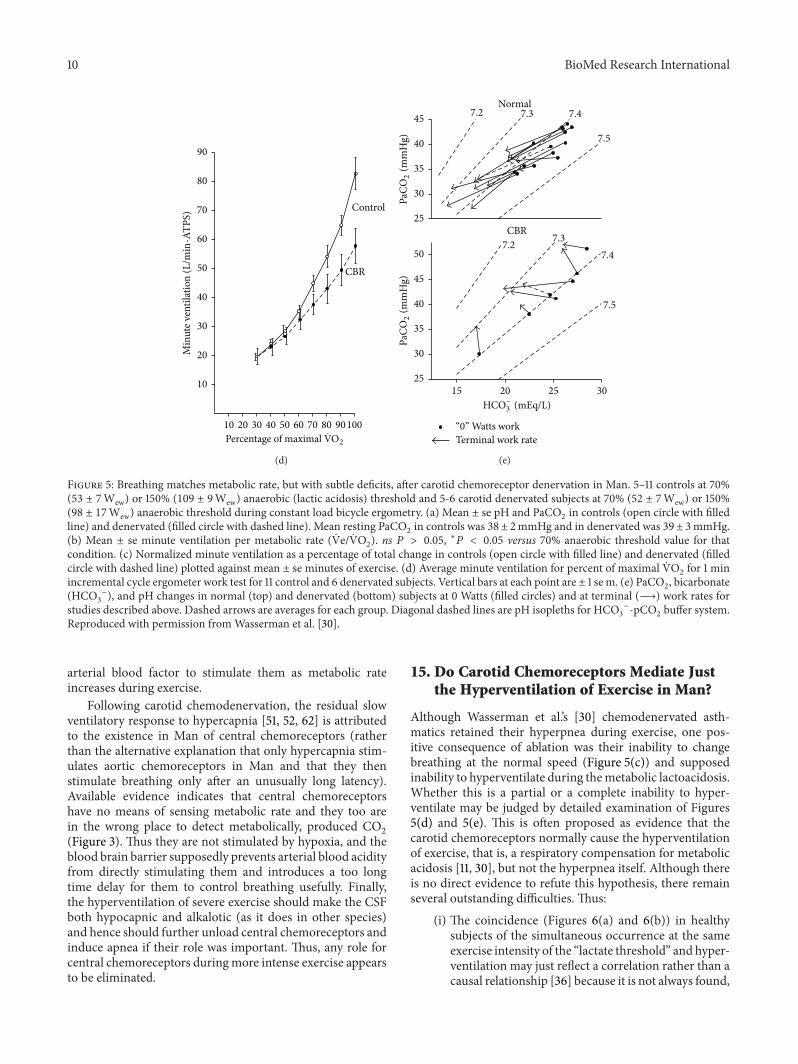
10 BioMed Research International
90
80
70
60
50
40
30
20
10
10 20 30 40 50 60 70 80 90100
Min
ute v
entil
atio
n (L
min
-ATP
S)
Control
CBR
Percentage of maximal VO2
(d)
45
40
35
30
25
50
45
40
35
30
25
72 73 74
75
72 7374
75
Normal
CBR
HCOminus3 (mEqL)
Terminal work rate
15 20 25 30Pa
CO2
(mm
Hg)
PaCO
2(m
mH
g)
ldquo0rdquo Watts work
(e)
Figure 5 Breathing matches metabolic rate but with subtle deficits after carotid chemoreceptor denervation in Man 5ndash11 controls at 70(53 plusmn 7Wew) or 150 (109 plusmn 9Wew) anaerobic (lactic acidosis) threshold and 5-6 carotid denervated subjects at 70 (52 plusmn 7Wew) or 150(98 plusmn 17Wew) anaerobic threshold during constant load bicycle ergometry (a) Mean plusmn se pH and PaCO
2in controls (open circle with filled
line) and denervated (filled circle with dashed line) Mean resting PaCO2in controls was 38 plusmn 2mmHg and in denervated was 39 plusmn 3mmHg
(b) Mean plusmn se minute ventilation per metabolic rate ( Ve VO2) ns 119875 gt 005 lowast119875 lt 005 versus 70 anaerobic threshold value for that
condition (c) Normalized minute ventilation as a percentage of total change in controls (open circle with filled line) and denervated (filledcircle with dashed line) plotted against mean plusmn se minutes of exercise (d) Average minute ventilation for percent of maximal VO
2for 1 min
incremental cycle ergometer work test for 11 control and 6 denervated subjects Vertical bars at each point are plusmn 1 se m (e) PaCO2 bicarbonate
(HCO3
minus) and pH changes in normal (top) and denervated (bottom) subjects at 0 Watts (filled circles) and at terminal (997888rarr) work rates for
studies described above Dashed arrows are averages for each group Diagonal dashed lines are pH isopleths for HCO3
minus-pCO2buffer system
Reproduced with permission fromWasserman et al [30]
arterial blood factor to stimulate them as metabolic rateincreases during exercise
Following carotid chemodenervation the residual slowventilatory response to hypercapnia [51 52 62] is attributedto the existence in Man of central chemoreceptors (ratherthan the alternative explanation that only hypercapnia stim-ulates aortic chemoreceptors in Man and that they thenstimulate breathing only after an unusually long latency)Available evidence indicates that central chemoreceptorshave no means of sensing metabolic rate and they too arein the wrong place to detect metabolically produced CO
2
(Figure 3) Thus they are not stimulated by hypoxia and theblood brain barrier supposedly prevents arterial blood acidityfrom directly stimulating them and introduces a too longtime delay for them to control breathing usefully Finallythe hyperventilation of severe exercise should make the CSFboth hypocapnic and alkalotic (as it does in other species)and hence should further unload central chemoreceptors andinduce apnea if their role was important Thus any role forcentral chemoreceptors duringmore intense exercise appearsto be eliminated
15 Do Carotid Chemoreceptors Mediate Justthe Hyperventilation of Exercise in Man
Although Wasserman et alrsquos [30] chemodenervated asth-matics retained their hyperpnea during exercise one pos-itive consequence of ablation was their inability to changebreathing at the normal speed (Figure 5(c)) and supposedinability to hyperventilate during themetabolic lactoacidosisWhether this is a partial or a complete inability to hyper-ventilate may be judged by detailed examination of Figures5(d) and 5(e) This is often proposed as evidence that thecarotid chemoreceptors normally cause the hyperventilationof exercise that is a respiratory compensation for metabolicacidosis [11 30] but not the hyperpnea itself Although thereis no direct evidence to refute this hypothesis there remainseveral outstanding difficulties Thus
(i) The coincidence (Figures 6(a) and 6(b)) in healthysubjects of the simultaneous occurrence at the sameexercise intensity of the ldquolactate thresholdrdquo and hyper-ventilation may just reflect a correlation rather than acausal relationship [36] because it is not always found
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
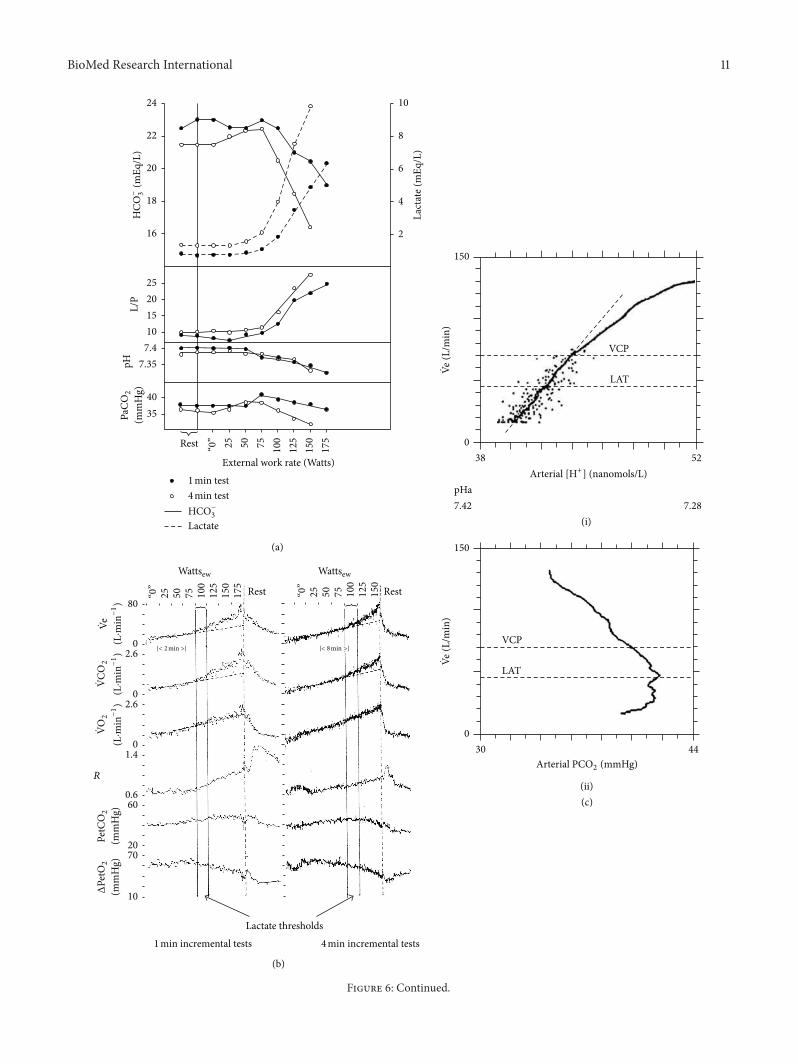
BioMed Research International 11
24
22
20
18
16
25201510
10
8
6
4
2
74735
4035
HCO
minus 3(m
EqL
)L
PpH
PaCO
2
(mm
Hg)
ldquo0rdquo 25 50 75 100
125
150
175Rest
External work rate (Watts)1 min test4 min testHCOminus
3
Lactate
Lact
ate (
mEq
L)
(a)
(c)
150
038 52
(Lm
in)
VCP
LAT
(i)
150
0
(Lm
in)
VCP
LAT
(ii)
30 44Arterial PCO2 (mmHg)
Wattsew Wattsew
25 50 75 100
125
150
175 Rest Restldquo0rdquo
25 50 75 100
125
150
ldquo0rdquo
|lt 2 min gt| |lt 8 min gt|
80
026
0
0
26
14
0660
2070
10
Lactate thresholds
(b)
(Lmiddotm
inminus1)
(Lmiddotm
inminus1)
PetC
O2
(mm
Hg)
ΔPe
tO2
(mm
Hg)
R
pHa742 728
(Lmiddotm
inminus1)
Arterial [H+] (nanomolsL)
1 min incremental tests 4 min incremental tests
VeVe
VeVC
O2
VO2
Figure 6 Continued
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
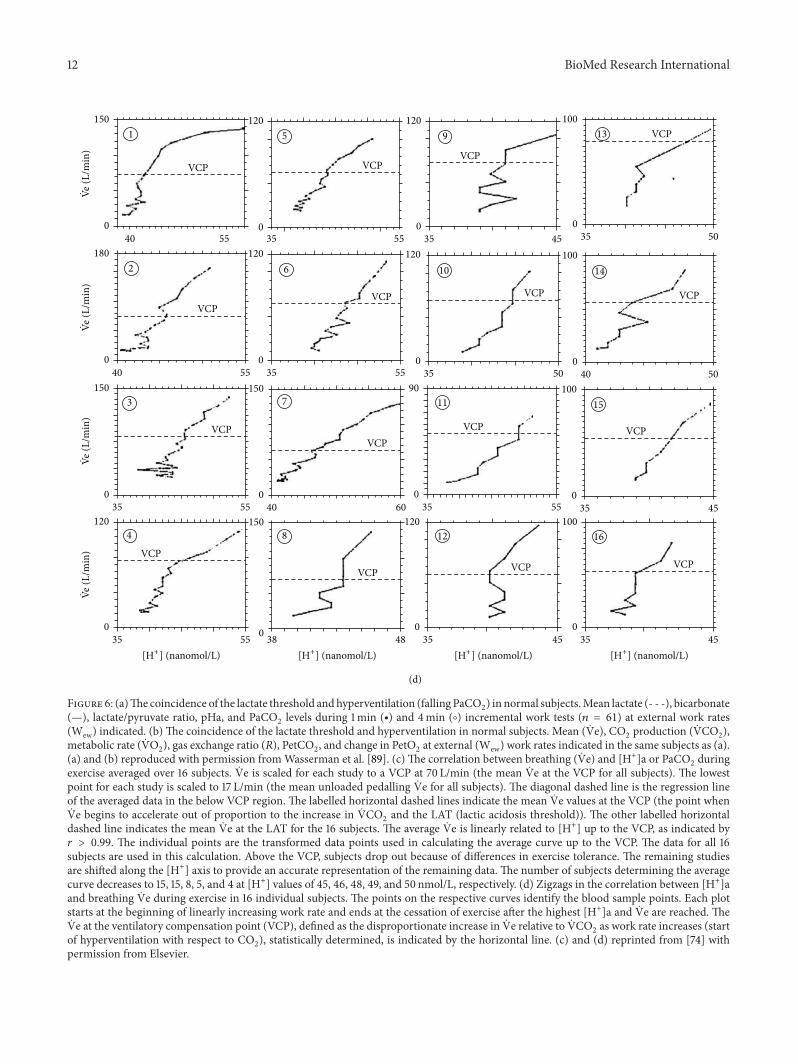
12 BioMed Research International(L
min
)(L
min
)(L
min
)(L
min
)
VCP VCPVCP
VCP
VCP
VCP
VCPVCP
VCP
VCPVCP
VCP
VCP
VCP
VCP
VCP
150
0 0
0 0 0
0
0
0 0
0
0
0
0
0 0
0
180
120
120
120
150 150
150
120
120
90
120
100
100
100
100
40
50
50
50
55
40 55 55
55
45
35
35
5535
5535
35
4535 4535
4535
35
35
5535
40
40
60
38 48
(d)
[H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL) [H+] (nanomolL)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
VeVe
VeVe
Figure 6 (a)The coincidence of the lactate threshold andhyperventilation (falling PaCO2) in normal subjectsMean lactate (- - -) bicarbonate
(mdash) lactatepyruvate ratio pHa and PaCO2levels during 1min (∙) and 4min (∘) incremental work tests (119899 = 61) at external work rates
(Wew) indicated (b) The coincidence of the lactate threshold and hyperventilation in normal subjects Mean ( Ve) CO2production ( VCO
2)
metabolic rate ( VO2) gas exchange ratio (119877) PetCO
2 and change in PetO
2at external (Wew)work rates indicated in the same subjects as (a)
(a) and (b) reproduced with permission fromWasserman et al [89] (c) The correlation between breathing ( Ve) and [H+]a or PaCO2during
exercise averaged over 16 subjects Ve is scaled for each study to a VCP at 70 Lmin (the mean Ve at the VCP for all subjects) The lowestpoint for each study is scaled to 17 Lmin (the mean unloaded pedalling Ve for all subjects) The diagonal dashed line is the regression lineof the averaged data in the below VCP region The labelled horizontal dashed lines indicate the mean Ve values at the VCP (the point whenVe begins to accelerate out of proportion to the increase in VCO
2and the LAT (lactic acidosis threshold)) The other labelled horizontal
dashed line indicates the mean Ve at the LAT for the 16 subjects The average Ve is linearly related to [H+] up to the VCP as indicated by119903 gt 099 The individual points are the transformed data points used in calculating the average curve up to the VCP The data for all 16subjects are used in this calculation Above the VCP subjects drop out because of differences in exercise tolerance The remaining studiesare shifted along the [H+] axis to provide an accurate representation of the remaining data The number of subjects determining the averagecurve decreases to 15 15 8 5 and 4 at [H+] values of 45 46 48 49 and 50 nmolL respectively (d) Zigzags in the correlation between [H+]aand breathing Ve during exercise in 16 individual subjects The points on the respective curves identify the blood sample points Each plotstarts at the beginning of linearly increasing work rate and ends at the cessation of exercise after the highest [H+]a and Ve are reached TheVe at the ventilatory compensation point (VCP) defined as the disproportionate increase in Ve relative to VCO
2as work rate increases (start
of hyperventilation with respect to CO2) statistically determined is indicated by the horizontal line (c) and (d) reprinted from [74] with
permission from Elsevier
BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 13
nor does it always persist after training [63] and thereis no correlation between the severity of the acidosisand the size of the hyperventilation [64]
(ii) The zigzag in the relationship between [H+]aand Ve in individuals occurring in 1316 subjects(Figure 6(d)) is inconvenient
(iii) Patients who cannot produce lactic acid (McArdlersquossyndrome) appear to have a similar hyperventilation(ie have a nonlinear increase in breathing) at aboutthe same relative exercise intensity [65] Neverthelesssince VO
2max in such patients is however so much
lower (25ndash46 of those in healthy controls) thevalidity of this comparison may be questioned
(iv) Experimentally induced changes in acid productiondo not always produce appropriate changes in thehyperventilation [18 66]
(v) This proposal predicts that the hyperventilationshould be abolished during exercise by unloading thecarotid chemoreceptors with sufficient hyperoxia butthis has not yet been demonstrated [67]
(vi) The metabolic acidosis of exercise is never fullycompensated by the hyperventilation [67 68] It is notobvious why carotid chemoreceptors should fail toachieve complete respiratory compensation whereasthis failure is irrelevant if the primary purpose of thehyperventilation has nothing to do with respiratorycompensation
Furthermore there remain several alternative explanationsfor the results in Figure 5 Thus
(i) This positive effect of ablationmdashthe failure to hyper-ventilatemdashmight have been some property of theseparticular asthmatics It was never established by howmuch these particular asthmatics could hyperventi-late before their chemodenervation (and there appearto be no definitive studies showing that asthma alonecannot also cause this) although such experimentsare still possible as this operation is still performed[51]
(ii) The failure to hyperventilate might be because braintemperature failed to rise as much in the chemoden-ervated asthmatics To allow for their lower ldquolactatethresholdrdquo they were exercised at a 10 lower inten-sity (98 plusmn 17Wew) than the intact controls (109 plusmn9Wew) [30] It is possible that it is hyperthermiainstead that causes the hyperventilation to increaseevaporative heat loss [69] Any additional ventilatoryloss of metabolic acid may be merely fortuitous(and this would explain why the hyperventilationnever fully compensates for the metabolic acidosis)Exercise (90Wew or more) can raise CNS (tym-panic membrane) temperature by sim1∘C [70 71] Suchhyperthermia induced at rest can itself cause suffi-cient hyperventilation in Man to lower PaCO
2below
25mmHg [71 72] Moreover preventing temperaturerising during exercise at 1 kW prevents PaCO
2falling
from 40 to 30mmHg [73]
All these difficulties could easily be dismissed by a newstudy demonstrating that such patients do hyperventilatebefore denervation (but not afterwards) and that they havethe same CNS temperature rises during exercise as thosein intact subjects But without such a study it is surpris-ingly difficult to provide definitive evidence that the carotidchemoreceptors mediate the hyperventilation of exercise
16 Could New Evidence for Acidemia belowthe Lactate Threshold Provide the Stimulusto Carotid Chemoreceptors That DrivesBreathing in Exercise
It has always been believed that metabolic acidosis itselfcauses the hyperventilation even if not the hyperpnea ofexerciseThus Figure 6(a) shows that acidemia did not occurin low intensity exercise that is below the lactate thresholdwhen only hyperpnea occurred Here the rate of arterialblood gas sampling was however unspecified Moreover afterexercise the decline in breathing did not apparently correlatewith the changes in arterial blood acidity (eg [68]) But thelatest work of Wasserman et al [74] involving rapid arterialsampling during exercise (6 per minute) and improved bloodgas measurement technology and expression of acidity in[H+]a units rather than the traditional but less sensitive pHnotation now challenges this belief It shows (see Figure 6(c))that theremay after all be a rise in [H+]a in exercise below thelactate threshold This may also be indicated in their earlierdata (see Figure 2(b) and [75]) This relationship however isstill only an intriguing correlation and does not establish anycausal relationship between [H+]a carotid chemoreceptorsand breathing during rest or exercise Could [H+]a be themissing link betweenmetabolic rate and carotid chemorecep-tors To establish a causal relationship new stimulation andablation experiments are still required to test whether
(i) infusing H+ stimulates breathing at rest (analogous tothe ldquoinfusionsrdquo of O
2ampCO
2of Figures 4(a) and 4(b))
(ii) infusion at rest to achieve the [H+]a levels inFigure 6(c) will mimic the breathing levels inFigure 6(c)
(iii) preventing the [H+]a rise during exercise prevents theincrease in breathing
The main problem for this hypothesis too however is stillthe fact that since carotid body denervation fails to preventbreathing matching metabolic rate during exercise how can[H+]a acting on carotid chemoreceptors alone provide thelink between metabolic rate and breathing during exercise
17 Possible Outcomes with Future RecordingExperiments in Man
All the above difficulties could be resolved if a new techniqueis found to record neural activity directly from peripheralarterial chemoreceptors in Man Intriguingly all the aboveevidence could fit either of the two possible outcomes fromsuch a recording experiment
14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 BioMed Research International
(A) Recording might indicate that their neural activityduring maximum exercise is barely different from that atrest In this case combining these results with the availableevidence from the other 2 key conditions (the absence of anyarterial stimulus to carotid chemoreceptors and the inabilityof carotid chemoreceptor ablation to prevent matching)would indicate that carotid chemoreceptors are not the keymetabolic rate sensor But even these negatives would befinally validated only when the mechanism that does act likea metabolic rate sensor is discovered
(B) Recording might indicate instead that their neu-ral activity during exercise does increase in proportionto metabolic rate Combining the results of the availableevidence from the 3 key experimental conditions describedabove would confirm the opposite conclusion that carotidchemoreceptors are the keymetabolic rate sensor In this casethe absence of a carotid arterial stimuluswould be an exampleof ldquoabsence of evidence is not evidence of absencerdquo It wouldsuggest that a carotid arterial stimulus must exist and will befound when new research adopts different approaches
The failure of carotid chemoreceptor denervation toprevent matching now favours duplication that is they andat least one other mechanism both act as key metabolic ratesensors It would then be necessary to test whether matchingis prevented by ablation only of the second mechanism(carotid chemoreceptors therefore being irrelevant) or onlyby ablation of both (carotid chemoreceptors therefore beingimportant but duplicating)
The correlation between their neural activity and meta-bolic rate implies causation which could then be establishedfrom the appropriate outcome of the other 2 conditions
18 Do Carotid Chemoreceptors ActOnly to Fine-Tune Breathing duringExercise in Man
Although at least one factor other than carotid chemorecep-tors must act equally well as carotid chemoreceptors as ametabolic rate sensor carotid chemoreceptors still could beand are often proposed [76] to operate as mismatch sensorsto fine-tune every breath on a breath-by-breath basis Whilethey have the potential to do so there is no evidence yet thatthey do act in this way
Carotid chemoreceptors potentially have this abilitybecause as already discussed artificially inducing hypoxia orhypercapnia always stimulates breathing Moreover carotidchemoreceptor discharge in anaesthetized or unanaes-thetised animals changes fast enough to detect changes inPaCO
2between breaths [77] and artificially applying large
changes in PaO2does affect breathing during exercise Thus
(i) Hypoxia (FiO2sim12) accelerates the speed of the
breathing response [78]
(ii) Hyperoxia ([79] the ldquoDejours testrdquo) causes a smallreduction and slowing of the kinetics of breathing[78 80] (although it has never been established howequivalent is the Dejours test to a proper carotidchemoreceptor denervation)
(iii) Carotid denervated patients may have a slowerbreathing response to exercise (see Figure 5(c)) havereduced sensitivity to artificially raising CO
2 and fail
to respond to the Dejours test [14 81]
Such evidence has always sustained the belief that some-how carotid chemoreceptors ldquomustrdquo be important in exercise[76] And the mismatch hypothesis is particularly attractivesince the required blood gas changes during exercise needonly involve PaCO
2and could only be of a few mmHg
Furthermore mismatch need not occur consistently becausewhatever sensed metabolic rate might be so successfulthat mismatch rarely occurs during exercise and thereforecarotid chemoreceptors normally have little to do
The key question however is whether they do act asmismatch sensors that make any important contributionto breath-by-breath control of breathing during exerciseEstablishing this requires new experiments measuring theincidence of breathsmismatchingmetabolic rate during exer-cise demonstrating that each mismatched breath is followedby a PaCO
2change and demonstrating that this PaCO
2
change causes a subsequent augmented breath (or two) torestore PaCO
2 At present the available evidence does not
support the hypothesis that they do act to fine-tune everybreath during exercise Thus
(i) Carotid denervation experiments failed to reveal anyobvious changes in breathing pattern or blood gasesduring exercise [29 30] that is removal of thereceptors necessary to mediate fine-tuning appearsnot to disturb greatly the control of breathing duringexercise
(ii) Strange-Petersen et alrsquos [35] stimulation experimentsin 4 subjects using brief pulses of hypoxia failed toalter the next breath (see Section 9)
(iii) The best recording experiment available so far totest this hypothesis by making frequent and sub-tle blood gas measurements (at least breath-by-breath) using intra-arterial pH electrodes [33]mdashseeSection 9mdashreveals no evidence of a signal whichcarotid chemoreceptors could use for fine-tuning
19 Are Carotid ChemoreceptorsImportant in Learning PostnatallyHow to Control Breathing
There is one last and ingenious hypothesis [82 83] Sincebreathing does match metabolic rate so well in adults ismatching present at birth or only learnt postnatally Iflearnt there has to be an early postnatal period for learningwhen breathing fails to match metabolic rate well Episodesof pronounced mismatch should therefore be more readilydetectable in neonates Such mismatch would result in pro-portionate blood gas error signals [82 83] and the carotidchemoreceptors would be the essential mismatch sensorsto detect them and to stimulate breathing accordingly Thebrain could then learn to become a metabolic rate sensor byanticipating the breathing required for every possible physical
BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 15
exertion in order to minimize these mismatch error signalsSuch learning may become so successful that by adulthoodmismatch almost never occurs Hence the carotid chemore-ceptors would not normally be stimulated in exercise whilealways being available to signal if mismatch ever occurredObviously no experiments have yet been devised to testthis hypothesis in Man and interpretation of such tests inneonates of other species is complicated by the fact that inother species carotid chemoreceptors may not be the onlyperipheral chemoreceptors that stimulate breathing
20 Difficulties in InterpretingRecent Animal Experiments StudyingCarotid Chemoreceptors
Ultimately it is not acceptable to combine key argumentsbased on experiments across species for example a stimu-lation experiment from a dog a recording experiment froma goat and an ablation experiment from a cat Unfortunatelythis often has to be done [15 16] because only informationacross species is available Ultimately the case for whateveracts as the key metabolic rate sensor must be establishedby stimulation ablation and recording in each species sep-arately as this review considers forManThere are difficultiesfor all other mammalian species (for reviews see Section 1)in establishing definitively if carotid chemoreceptors are thekey metabolic rate sensors that mediate matching Recentand ingenious experiments on dogs show some of the equaldifficulties in establishing a role for carotid chemoreceptorsin other species
Blain et al [84] performed a series of experiments on4 unanaesthetised and resting dogs with carotid bodies onone side denervated and the other side being experimen-tally unloaded by extra corporeal perfusion with hyperoxic(PaO2gt 500mmHg) and hypocapnic (PaCO
2sim20mmHg)
blood Unloading transiently decreased ventilation but whileunloading persisted ventilation partially recovered (after50ndash70 s) Furthermore in 8 resting dogs such unloadingdecreased the slope (by 19) of the breathing response toartificially lowering PetCO
2(by 10ndash12mmHg below its eup-
neic level [85]) whereas stimulation at rest with artificiallyhypoxic arterial blood (PaO
2of 40mmHg) increased the
slope by 223Detecting any effects on breathing with such manipu-
lations is all the more remarkable because aortic chemore-ceptors also stimulate breathing in dogs and enhance thebreathing response to carotid body stimulation [86] Thusinformation from intact aortic chemoreceptors will contra-dict that from such manipulated carotid chemoreceptors
It is still unclear precisely what these ingenious exper-iments reveal about the role of carotid chemoreceptors asmetabolic rate or mismatch sensors in resting dogs If it werealready established definitively that carotid chemoreceptorsare the key metabolic rate sensors in dogs these experimentsprovide elegant confirmation But this is still unclear in dogsWithout an arterial blood gas (or other) change to link carotidchemoreceptors with metabolic rate in dogs these results areequally consistent with carotid chemoreceptors not being the
key metabolic rate sensors Instead they also confirm merelythat breathing in dogs is alterable by withdrawing or addinganother stimulus and provide no evidence about their role inbreathing matching metabolic rate during exercise
21 Conclusions
The only compelling evidence that carotid chemoreceptorsare important in controlling breathing during exercise inManis the fact that they are the only peripheral chemoreceptorsever found in Man that are stimulated by hypoxia whichin turn stimulates breathing Since neither arterial hypoxianor hypercapnia occurs during exercise to stimulate carotidchemoreceptors in Man and bilateral carotid chemoreceptordenervation in Man fails to prevent breathing matchingmetabolic rate we still can neither confirm nor refute thebelief that carotid chemoreceptors are important in control-ling breathing during exercise in Man
There is still huge controversy over whether carotidchemoreceptors have any importance in controlling breath-ing during exercise despite sim80 years of research Remark-ably two reviews in the same journal in 2012 [15 16] stilldraw very different conclusions about their role This reviewidentifies what evidence is necessary to establish how impor-tant are carotid chemoreceptors in controlling breathing inexercise in Man the species in which we have most interestWithout being able to record from carotid chemoreceptorsduring exercise the best conclusion so far is that we stillcannot distinguish between carotid chemoreceptors beingone of two important but duplicated mechanisms or havingno importance in controlling breathing during exercise inMan
Conflict of Interests
The author has no conflict of interests with companies ofmanufacturers who might benefit from the results of thepresent study
References
[1] F De Castro ldquoTowards the sensory nature of the carotid bodyHering De Castro and Heymansrdquo Frontiers in Neuroanatomyvol 3 pp 1ndash11 2009
[2] C Heymans ldquoPharmacology in old and modern medicinerdquoAnnual Review of Pharmacology vol 7 pp 1ndash15 1967
[3] J H Comroe ldquoThe hypernea of muscular exerciserdquo Physiologi-cal Reviews vol 24 pp 319ndash339 1944
[4] T G Waldrop F L Eldridge G A Iwamoto and J HMitchell ldquoCentral neural control of respiration and circulationduring exerciserdquo inHandbook of Physiology Section 12 ExerciseRegulation and Integration of Multiple Systems L B Rowell andJ T Shepherd Eds pp 353ndash380 Oxford University Press NewYork NY USA 1996
[5] S A Ward ldquoAssessment of peripheral chemoreflex contribu-tions to exercise hyperpnea in humansrdquo Medicine and Sciencein Sports and Exercise vol 26 no 3 pp 303ndash310 1994
16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

16 BioMed Research International
[6] N R Prabhakar and Y-J Peng ldquoPeripheral chemoreceptors inhealth and diseaserdquo Journal of Applied Physiology vol 96 no 1pp 359ndash366 2004
[7] P Dejours ldquoControl of respiration in muscular exerciserdquo inHandbook of Physiology Respiration Section 1 W O Fenn andH Rahn Eds pp 631ndash648 American Physiological SocietyWashington DC USA 1964
[8] J H Comroe Physiology of Respiration An Introductory TextYear Book Chicago Ill USA 2nd edition 1974
[9] J A Dempsey and B J Whipp ldquoThe respiratory systemrdquo inExercise Physiology People and Ideas CM Tipton Ed pp 138ndash187 American Physiological Society Bethesda Md USA 2003
[10] G SDawes and JHComroe Jr ldquoChemoreflexes from the heartand lungsrdquoPhysiological reviews vol 34 no 2 pp 167ndash201 1954
[11] K Wasserman B J Whipp and R Casaburi ldquoRespiratorycontrol during exerciserdquo in Handbook of Physiology Section 3the Respiratory System II the Control of Breathing pp 595ndash619American Physiological Society Bethesda Md USA 1986
[12] B J Whipp ldquoThe control of exercise hyperpneardquo in Regulationof Breathing Part II T F Hornbein Ed pp 1069ndash1139 MarcelDekker New York NY USA 1981
[13] B J Whipp ldquoVentilatory control during exercise in humansrdquoAnnual Review of Physiology vol 45 pp 393ndash413 1983
[14] B J Whipp and J A Davis ldquoPeripheral chemoreceptorsand exercise hyperpneardquo Medicine and Science in Sports andExercise vol 11 no 2 pp 204ndash212 1979
[15] P Kumar and N R Prabhakar ldquoPeripheral chemoreceptorsfunction and plasticity of the carotid bodyrdquo ComprehensivePhysiology vol 2 pp 141ndash219 2012
[16] H V Forster P Haouzi and J A Dempsey ldquoControl ofbreathing during exerciserdquoComprehensive Physiology vol 2 pp743ndash777 2012
[17] J BronowskiThe Ascent of Man BBC London UK 1973[18] H V Forster and L G Pan ldquoControl of breathing during exer-
ciserdquo inThe Lung R G Crystal J B West E R Weibel and PJ Barnes Eds pp 2001ndash2010 Lippincott-Raven PhiladelphiaPa USA 2nd edition 1997
[19] E Nattie ldquoWhy do we have both peripheral and centralchemoreceptorsrdquo Journal of Applied Physiology vol 100 no 1pp 9ndash10 2006
[20] C Bernard An Introduction to the Study of ExperimentalMedicine Dover Publications New York NY USA 1865
[21] J S Mill A System of Logic Ratiocinative and Inductive Beinga Connected View of the Principles of Evidence and the Methodsof Scientific Investigation Longmans London UK 9th edition1875
[22] M I Cohen and S C Wang ldquoRespiratory neuronal activity inpons of catrdquo Journal of Neurophysiology vol 22 no 1 pp 33ndash501959
[23] M J Parkes ldquoEndocrine factors in fetal growthrdquo in Fetal andNeonatal Growth F Cockburn Ed pp 33ndash48 John Wiley ampSons Chichester UK 1988
[24] J F Stein An Introduction to Neurophysiology BlackwellsOxford UK 1982
[25] J F Stein and C J StoodleyNeuroscience an Introduction JohnWiley amp Sons Chichester UK 2006
[26] F M R Walshe ldquoOn the interpretation of experimental studiesof cortical motor function with special reference to the ldquooper-ational viewrdquo of experimental proceduresrdquo Brain vol 74 no 3pp 249ndash266 1951
[27] H Galbo M Kjaer and N H Secher ldquoCardiovascular ventila-tory and catecholamine responses tomaximal dynamic exercisein partially curarized manrdquo Journal of Physiology vol 389 pp557ndash568 1987
[28] B K Hutt S M Horvath and G B Spurr ldquoInfluence of varyingdegrees of passive limb movements on respirationrdquo Journal ofApplied Physiology vol 12 no 2 pp 297ndash300 1958
[29] Y Honda S Myojo S Hasegawa T Hasegawa and J WSeveringhaus ldquoDecreased exercise hyperpnea in patients withbilateral carotid chemoreceptor resectionrdquo Journal of AppliedPhysiology Respiratory Environmental and Exercise Physiologyvol 46 no 5 pp 908ndash912 1979
[30] K Wasserman B J Whipp S N Koyal and M G ClearyldquoEffect of carotid body resection on ventilatory and acid basecontrol during exerciserdquo Journal of Applied Physiology vol 39no 3 pp 354ndash358 1975
[31] J A Dempsey and P D Wagner ldquoExercise-induced arterialhypoxemiardquo Journal of Applied Physiology vol 87 no 6 pp1997ndash2006 1999
[32] D M Band C B Wolff J Ward G M Cochrane and J PriorldquoRespiratory oscillations in arterial carbon dioxide tension as acontrol signal in exerciserdquo Nature vol 283 no 5742 pp 84ndash851980
[33] K Murphy R P Stidwill B A Cross et al ldquoIs hypercapnianecessary for the ventilatory response to exercise in manrdquoClinical Science vol 73 no 6 pp 617ndash625 1987
[34] C S Poon and C Tin ldquoMechanism of augmented exercisehyperpnea in chronic heart failure and dead space loadingrdquoRespiratory Physiology amp Neurobiology vol 186 pp 114ndash1302013
[35] E Strange-Petersen B J Whipp D B Drysdale and D JCunningham ldquoCarotid arterial blood gas oscillations and thephase of the respiratory cycle during exercise in man testing amodelrdquoAdvances in ExperimentalMedicine and Biology vol 99pp 335ndash342 1978
[36] J HMateika and J Duffin ldquoA review of the control of breathingduring exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 71 no 1 pp 1ndash27 1995
[37] T J Clark and S Godfrey ldquoThe effect of CO2on ventilation and
breath-holding during exercise and while breathing through anadded resistancerdquo Journal of Physiology vol 201 no 3 pp 551ndash566 1969
[38] A R C Cummin J Alison M S Jacobi V I Iyawe and KB Saunders ldquoVentilatory sensitivity to inhaled carbon dioxidearound the control point during exerciserdquo Clinical Science vol71 no 1 pp 17ndash22 1986
[39] J Duffin and G V McAvoy ldquoThe peripheral-chemoreceptorthreshold to carbon dioxide in manrdquo Journal of Physiology vol406 pp 15ndash26 1988
[40] M Miyamura H Folgering R A Binkhorst F D J Smoldersand F Kreuzer ldquoVentilatory response to CO
2at rest and during
positive andnegativework in normoxia and hyperoxiardquoPflugersArchiv European Journal of Physiology vol 364 no 1 pp 7ndash151976
[41] C S Poon and J G Greene ldquoControl of exercise hyperpneaduring hypercapnia in humansrdquo Journal of Applied Physiologyvol 59 no 3 pp 792ndash797 1985
[42] A K Datta S A Shea R L Horner and A Guz ldquoThe influenceof induced hypocapnia and sleep on the endogenous respiratoryrhythm in humansrdquo Journal of Physiology vol 440 pp 17ndash331991
BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 17
[43] D J Paterson P A Robbins and J Conway ldquoChanges in arterialplasma potassium and ventilation during exercise in manrdquoRespiration Physiology vol 78 no 3 pp 323ndash330 1989
[44] D J Paterson J S Friedland D O Oliver and P A RobbinsldquoThe ventilatory response to lowering potassium with dextroseand insulin in subjects with hyperkalaemiardquo Respiration Physi-ology vol 76 no 3 pp 393ndash398 1989
[45] D J Paterson K L Dorrington D H Bergel et al ldquoEffect ofpotassium on ventilation in the rhesus monkeyrdquo ExperimentalPhysiology vol 77 no 1 pp 217ndash220 1992
[46] M M Warner and G S Mitchell ldquoVentilatory responses tohyperkalemia and exercise in normoxic and hypoxic goatsrdquoRespiration Physiology vol 82 no 2 pp 239ndash250 1990
[47] H E Cooper T H Clutton-Brock and M J Parkes ldquoContri-bution of the respiratory rhythm to sinus arrhythmia in normalunanesthetized subjects during positive-pressure mechanicalhyperventilationrdquo American Journal of PhysiologymdashHeart andCirculatory Physiology vol 286 no 1 pp H402ndashH411 2004
[48] S Corne K Webster and M Younes ldquoHypoxic respiratoryresponse during acute stable hypocapniardquo American Journal ofRespiratory and Critical Care Medicine vol 167 no 9 pp 1193ndash1199 2003
[49] RMohan and JDuffin ldquoThe effect of hypoxia on the ventilatoryresponse to carbon dioxide in manrdquo Respiration Physiology vol108 no 2 pp 101ndash115 1997
[50] P BockThe Paraganglia Springer Heidelberg Germany 1982[51] A Dahan D Nieuwenhuijs and L Teppema ldquoPlasticity of cen-
tral chemoreceptors effect of bilateral carotid body resection oncentral CO
2sensitivityrdquo PLoSMedicine vol 4 no 7 article e239
2007[52] M Fatemian D J F Nieuwenhuijs L J Teppema et al
ldquoThe respiratory response to carbon dioxide in humans withunilateral and bilateral resections of the carotid bodiesrdquo Journalof Physiology vol 549 no 3 pp 965ndash973 2003
[53] Y Honda S Watanabe I Hashizume et al ldquoHypoxicchemosensitivity in asthmatic patients two decades after carotidbody resectionrdquo Journal of Applied Physiology Respiratory Envi-ronmental and Exercise Physiology vol 46 no 4 pp 632ndash6381979
[54] R Lugliani B J Whipp C Seard and K Wasserman ldquoEffectof bilateral carotid-body resection on ventilatory control atrest and during exercise in manrdquo The New England Journal ofMedicine vol 285 no 20 pp 1105ndash1111 1971
[55] J G Wade C P Larson Jr R F Hickey W K Ehrenfeld and JW Severinghaus ldquoEffect of carotid endarterectomy on carotidchemoreceptor and baroreceptor function in manrdquo The NewEngland Journal of Medicine vol 282 no 15 pp 823ndash829 1970
[56] A Guz M I M Noble J G Widdicombe D Trenchardand W W Mushin ldquoPeripheral chemoreceptor block in manrdquoRespiration Physiology vol 1 no 1 pp 38ndash40 1966
[57] B J Whipp S A Ward V C Baum and B Winter ldquoTheinfluence of hyperoxia on ventilatory kinetics during moderateexercise in carotid-body-resected humansrdquo Journal of Physiol-ogy vol 475 article 20 1994
[58] J W Severinghaus ldquoCarotid body resection for COPDrdquo Chestvol 95 no 5 pp 1128ndash1129 1989
[59] M S Stulbarg andW RWinn ldquoBilateral carotid body resectionfor the relief of dyspnea in severe chronic obstructive pul-monary disease physiologic and clinical observations in threepatientsrdquo Chest vol 95 no 5 pp 1123ndash1128 1989
[60] S A Shea andA Guz ldquoPersonnalite ventilatoiremdashan overviewrdquoRespiration Physiology vol 87 no 3 pp 275ndash291 1992
[61] J J Rutherford T H Clutton-Brock and M J ParkesldquoHypocapnia reduces the T wave of the electrocardiogramin normal human subjectsrdquo American Journal of PhysiologymdashRegulatory Integrative and Comparative Physiology vol 289 no1 pp R148ndashR155 2005
[62] J W Bellville B J Whipp R D Kaufman G D Swanson KA Aqleh and D MWiberg ldquoCentral and peripheral chemore-flex loop gain in normal and carotid body-resected subjectsrdquoJournal of Applied Physiology Respiratory Environmental andExercise Physiology vol 46 no 4 pp 843ndash853 1979
[63] J A Dempsey H V Forster and D M Ainsworth ldquoRegulationof hyperpnea hyperventilation and respiratory muscle recruit-ment during exerciserdquo inRegulation of Breathing J A Dempseyand A I Pack Eds pp 1065ndash1132 Marcel Dekker New YorkNY USA 1995
[64] J A Dempsey E H Vidruk and G S Mitchell ldquoPulmonarycontrol systems in exercise updaterdquo Federation Proceedings vol44 no 7 pp 2260ndash2270 1985
[65] J M Hagberg E F Coyle J E Carroll J M Miller WH Martin and M H Brooke ldquoExercise hyperventilation inpatients with McArdlersquos diseaserdquo Journal of Applied PhysiologyRespiratory Environmental and Exercise Physiology vol 52 no4 pp 991ndash994 1982
[66] R Jeyaranjan R Goode and J Duffin ldquoThe effect of metabolicacid-base changes on the ventilatory changes at the end ofheavy exerciserdquo European Journal of Applied Physiology andOccupational Physiology vol 58 no 4 pp 405ndash410 1989
[67] S M Rausch B J Whipp K Wasserman and A HuszczukldquoRole of the carotid bodies in the respiratory compensationfor the metabolic acidosis of exercise in humansrdquo Journal ofPhysiology vol 444 pp 567ndash578 1991
[68] W Stringer R Casaburi and KWasserman ldquoAcid-base regula-tion during exercise and recovery in humansrdquo Journal of AppliedPhysiology vol 72 no 3 pp 954ndash961 1992
[69] R G G Hanson ldquoRespiratory heat loss at increased coretemperaturerdquo Journal of Applied Physiology vol 37 no 1 pp103ndash107 1974
[70] P Chappuis P Pittet and E Jequier ldquoHeat storage regulation inexercise during thermal transientsrdquo Journal of Applied Physiol-ogy vol 40 no 3 pp 384ndash392 1976
[71] J D MacDougall W G Reddan C R Layton and J ADempsey ldquoEffects of metabolic hyperthermia on performanceduring heavy prolonged exerciserdquo Journal of Applied Physiologyvol 36 no 5 pp 538ndash544 1974
[72] P F Iampietro M Mager and E B Green ldquoSome physiologicalchanges accompanying tetany induced by exposure to hot wetconditionsrdquo Journal of Applied Physiology vol 16 pp 409ndash4121961
[73] J A Dempsey J M Thomson S C Alexander H V Forsterand L W Chosy ldquoRespiratory influences on acid-base statusand their effects on O
2transport during prolonged muscular
workrdquo in Metabolic Adaptation to Prolonged Physical ExerciseH Howard and J R Poortmans Eds pp 56ndash64 BirkhauserBasel Switzerland 1975
[74] K Wasserman W L Beaver X-G Sun and W W StringerldquoArterial H+ regulation during exercise in humansrdquo RespiratoryPhysiology amp Neurobiology vol 178 no 2 pp 191ndash195 2011
[75] X-G Sun J E HansenWW Stringer H Ting and KWasser-man ldquoCarbon dioxide pressure-concentration relationship in
18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

18 BioMed Research International
arterial and mixed venous blood during exerciserdquo Journal ofApplied Physiology vol 90 no 5 pp 1798ndash1810 2001
[76] D J C Cunningham ldquoStudies on arterial chemoreceptors inmanrdquo Journal of Physiology vol 384 pp 1ndash26 1987
[77] S Lahiri ldquoPhysiologic responses peripheral chemoreflexesrdquo inThe Lung Scientific Foundations R G Crystal and J B WestEds pp 1333ndash1340 Raven Press New York NY USA 1991
[78] T L Griffiths L C Henson and B J Whipp ldquoInfluence ofinspired oxygen concentration on the dynamics of the exercisehypernoea in manrdquo Journal of Physiology vol 380 pp 387ndash4031986
[79] J V Weil and G D Swanson ldquoPeripheral chemoreceptors andthe control of breathingrdquo in Exercise Pulmonary Physiology andPathophysiology B J Whipp and K Wasserman Eds pp 371ndash403 Marcel Dekker New York NY USA 1991
[80] R Jeyaranjan R Goode S Beamish and J Duffin ldquoThe con-tribution of peripheral chemoreceptors to ventilation duringheavy exerciserdquo Respiration Physiology vol 68 no 2 pp 203ndash213 1987
[81] KWasserman ldquoTesting regulation of ventilation with exerciserdquoChest vol 70 no 1 pp 173ndash178 1976
[82] G G Somjen ldquoThe missing error signalmdashregulation beyondnegative feedbackrdquo News in Physiological Sciences vol 7 pp184ndash185 1992
[83] H E Wood M Fatemian and P A Robbins ldquoA learnedcomponent of the ventilatory response to exercise in manrdquoJournal of Physiology vol 553 no 3 pp 967ndash974 2003
[84] G M Blain C A Smith K S Henderson and J A DempseyldquoContribution of the carotid body chemoreceptors to eupneicventilation in the intact unanesthetized dogrdquo Journal of AppliedPhysiology vol 106 no 5 pp 1564ndash1573 2009
[85] G M Blain C A Smith K S Henderson and J A DempseyldquoPeripheral chemoreceptors determine the respiratory sensitiv-ity of central chemoreceptors to CO
2rdquo Journal of Physiology vol
588 no 13 pp 2455ndash2471 2010[86] M Daly and A Ungar ldquoComparison of the reflex responses
elicited by stimulation of the separately perfused carotid andaortic body chemoreceptors in the dogrdquo Journal of Physiologyvol 182 no 2 pp 379ndash403 1966
[87] R D Dripps and J H Comroe ldquoThe effect of the inhalation ofhigh and low oxygen concentrations on respiration pulse rateballistocardiogram and arterial oxygen saturation (oximeter) ofnormal individualsrdquo American Journal of Physiology vol 149pp 277ndash291 1947
[88] C J Lambertsen R H Kough D Y Cooper G L Emmel HH Loeschcke and C F Schmidt ldquoComparison of relationshipof respiratory minute volume to pCO
2and pH or arterial blood
and internal jugular blood in normal Man during hyperventi-lation produced by low concentrations of CO
2at 1 atmosphere
and byO2at 30 atmospheresrdquo Journal of Applied Physiology vol
5 pp 255ndash263 1953[89] K Wasserman B J Whipp S N Koyal and W L Beaver
ldquoAnaerobic threshold and respiratory gas exchange duringexerciserdquo Journal of Applied Physiology vol 35 no 2 pp 236ndash243 1973
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology