revista latinoamericana el ambiente y las ciencias 6(13...
TRANSCRIPT

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
72
1Determinación de la actividad de oxidasas producidas por Oxyporus latemarginatus
crecido en fermentación sumergida en presencia y ausencia del colorante amarillo
azo.
Determination of oxidases activity produced by Oxyporus latemarginatus grown in
submerged fermentation in the presence and the absence of the yellow azo dye.
1Iván González Caloch, 2Soley Berenice Nava Galicia, 3Rubén Díaz Godínez, 2Veronica
Garrido Bazán, 1Saúl Tlecuitl Beristain, 2Martha Dolores Bibbins Martínez.
1Ingeniería en Biotecnología-UPTlax, Av Universidad Politécnica No. 1, C.P. 90180, Tlaxcala,
México, Tel. (246) 465 1300.
2Centro de Investigación en Biotecnología Aplicada (CIBA-IPN), Carretera Estatal Tecuexcomac-
Tepetitla km 1.5, C.P. 90700, Tlaxcala, México, Tel (248) 487 0765.
3Centro de Investigación en Ciencias Biológicas (CICB-UATx), Carretera San Martin Texmelucan
km 10.5, C.P. 90120, Tlaxcala, México, Tel (248)481 5482.
RESUMEN. En esta investigación se realizaron fermentaciones líquidas en medio basal y
en presencia del colorante amarillo azo a 500 ppm con el hongo basidiomiceto Oxyporus
latemarginatus. En relación a la cinética de crecimiento, se observó que hay una
disminución en la producción de biomasa máxima (Xmáx) y un aumento en la velocidad
específica de crecimiento () en la fermentación en presencia del colorante, obteniéndose
valores de Xmáx de 2.71 gL-1 y 3.74 gL-1, así como s de 0.019 h-1 y 0.016 h-1 para la
fermentación con y sin colorante, respectivamente. El análisis que se realizó para la
actividad enzimática indicó que cuatro de las cinco enzimas estudiadas (lacasa (Lac),
lignino peroxidasa (LiP), manganeso peroxidasa (MnP) y versátil peroxidasa (VP))
presentaron actividades máximas mayores en la fermentación en presencia del colorante y
sólo la enzima DyP mostró un valor de actividad máxima mayor en la fermentación basal.
Por lo que la actividad enzimática de las cinco oxidasas demuestra que O. latemarginatus
en presencia del colorante amarillo azo induce la producción de enzimas ligninolíticas.
Recibido: Abril, 2015.
Aprobado: Junio, 2015

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
73
ABSTRACT. In this research were performed submerged fermentations of the
basidiomycete fungus Oxyporus latemarginatus grown in basal medium and in the presence
of 500 ppm of the yellow azo textile dye. The growth kinetic shown that there is a decrease
in the maximum biomass production (Xmax) and an increase in the specific growth rate ()
in the fermentation in the presence of the dye, obtaining values of Xmax of 2.71 gL-1 and
3.74 gL-1, as well as s of 0.019 h-1 and h-1 0.016 for fermentation with and without dye
respectively. Analysis carried out for the enzyme activity indicated that four of the five
studied enzymes (laccase (Lac), lignin peroxidase (LiP), manganese peroxidase (MnP) and
versatile (VP) peroxidase,) showed maximum activities higher in the fermentation in the
presence of the dye and just the DyP enzyme showed a value of maximum activity in the
basal fermentation. These results indicate that the yellow azo-dye acts as inducer of the
enzyme activity of the ligninolytic enzymes produced by O. latemarginatus.
Palabras clave: Biorremediación, oxidasas, Oxyporus latemarginatus
Key words: Bioremediation, oxidases, Oxyporus latemarginatus
INTRODUCCIÓN
Más de diez mil diferentes tipos de pigmentos y colorantes sintéticos son usados en
diferentes industrias como la textil, papelera, cosmética y farmacéutica, entre otras
(Anjaneyulu y col., 2005). Los colorantes que están presentes en las aguas residuales
constituyen un problema ambiental y está bien establecido que la contaminación reduce la
calidad de vida en diversos aspectos, afecta a la salud y la duración de la misma, por lo
tanto, la biorremediación de contaminantes para la reducción de sus efectos tóxicos es de
primordial importancia (Phugare y col., 2011). La ventaja que se tiene al trabajar con este
tipo de tecnologías es que, además de la decoloración que se lleva a cabo, se puede alcanzar
la completa mineralización del colorante, gracias a la existencia de un gran número de
microorganismos que tienen la capacidad de suprimir el color que está presente en las aguas
residuales y todo esto se puede llevar a cabo mediante mecanismos de biodegradación
aeróbica o anaeróbica, biosorción y producción de enzimas capaces de catalizar la
decoloración, oxidación y la mineralización de los colorantes presentes en el agua del

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
74
efluente. Por lo que se dice que la degradación de los efluentes textiles utilizando
microorganismos ha sido ampliamente estudiado (Tamboli y col., 2010).
Así mismo, investigaciones de degradación de estos contaminantes utilizando las enzimas
oxidasas del sistema ligninolítico de los hongos de pudrición blanca se han llevado a cabo y
hoy en día se emplean para el desarrollo de técnicas de biorremediación de compuestos
xenobióticos y recalcitrantes debido a que este sistema enzimático es de carácter no
especifico, es decir, tiene la capacidad de catalizar la oxidación o hidrolizar a éstos y otros
tipos de compuestos orgánicos. Una ventaja del enfoque enzimático es que estas moléculas
pueden reaccionar con una amplia gama de compuestos aromáticos en condiciones diluidas.
A veces, la velocidad de decoloración y eliminación es bastante rápido y son menos
sensibles a las perturbaciones operacionales en comparación con la flora microbiana (Khan
y col., 2007).
Oxyporus latemarginatus es un hongo perteneciente a los basidiomicetos que ha sido poco
estudiado en su producción de enzimas ligninolíticas las cuales pueden ser aplicadas para el
desarrollo de nuevos métodos de biorremediación. Por lo que en el presente trabajo se
estudió la producción de cinco oxidasas de O. latemarginatus crecido en fermentación
líquida en presencia y ausencia del colorante amarillo azo.
METODOLOGÍA
Condiciones de cultivo de Oxyporus latemarginatus
La composición del medio de la fermentación sumergida con y sin colorante se preparó tal
y como se muestra a continuación (Tabla 1), ajustando el pH inicial a 6.5 con NaOH.
Tabla 1. Composición del medio de cultivo (Téllez-Téllez y col., 2008).
Componente Concentración en gL-1
Extracto de levadura 5

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
75
Glucosa 10
K2HPO4 0.4
ZnSO4 7H2O 0.001
KH2PO4 0.6
FeSO4 7H2O 0.05
MnSO4 H2O 0.05
MgSO4 7H2O 0.5
CuSO4 5H2O* 0.25
Las fermentaciones se llevaron a cabo en matraces Erlenmeyer de 125 mL, a cada matraz se
le agregó 50 mL de medio y fueron incubados en agitación orbital a 120 rpm a temperatura
de 30 °C.
La diferencia para ambas fermentaciones en cuanto a composición fue que a los matraces
Erlenmeyer de la fermentación en presencia del colorante amarillo azo se les adicionó una
concentración de 500 ppm (p/v) en un volumen final de 51 mL.
Extracto enzimático
Se obtuvo el extracto enzimático a partir de las 48 h y posteriormente cada 24 h hasta
terminar a las 504 h. El medio se filtró en papel Whatman No.1 con un equipo de filtración
al vacío. Las muestras obtenidas fueron conservadas en tubos eppendorf en congelación a -
20 °C.
Biomasa
La biomasa se separó del extracto enzimático por filtración al vacío, posteriormente se secó
en horno a 60°C por 24 h, por lo tanto, el peso seco de la biomasa se reportó como gramos
de biomasa seca (X) por litro de medio (g/L) (Díaz-Godínez y col., 2001).
Parámetros cinéticos de crecimiento del hongo

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
76
Se consideró que la evolución de la biomasa X=X(t) por la ecuación logística (Ec. 1) por la
minimización del error cuadrático y esto con ayuda de la herramienta Solver de la hoja
electrónica de Excel (Microsoft) (Díaz-Godínez y col.,2001; Viniegra-González y col.,
2003)
(Ec. 1) 𝑑𝑋
𝑑𝑡= 𝜇 (1 −
𝑋
𝑋𝑚𝑎𝑥) 𝑋
La solución de la ecuación logística es:
(Ec. 2) 𝑋 =𝑋𝑚𝑎𝑥
(1+𝐶 𝑒𝑥𝑝−𝜇𝑡)
Donde, μ = velocidad específica de crecimiento, Xmax = valor de biomasa máxima o de
equilibrio, C = (Xmax-X0/X0), cuando X=X0 es el valor inicial de la biomasa.
Determinación de la concentración de proteína soluble extracelular
Se utilizó albúmina sérica bovina como proteína estándar. La concentración de proteína
soluble se determinó en el extracto enzimático libre de células por el método de Bradford
(Bradford, 1976). A 100 µL de extracto enzimático se le adicionaron 200 µL del reactivo
de Bradford Biorad, el volumen final de la reacción fue de 300 µL. La absorbancia se leyó
a 595 nm en un espectrofotómetro GENEYSIS UV-Vis.
Determinación del contenido de azúcares
Se empleó como referencia la técnica de DNS (Miller , 1959). A 25 µL de extracto
enzimático se le adicionó 975 µL de agua desionizada y 2 mL de DNS. De esta manera, el
volumen final de la reacción fue de 3 mL y posteriormente las muestras se pusieron a baño
maría durante 5 minutos y la reacción se detuvo sumergiendo los tubos en agua fría. Se
leyó la absorbancia a 575 nm.
Determinación del pH
El pH se determinó por potenciometría en cada uno de los extractos crudos enzimáticos de
ambas fermentaciones iniciando el monitoreo en el tiempo 0 horas y posteriormente a las
hora 48 h, continuando cada 24 h hasta llegar a las 504 h.

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
77
Actividad enzimática extracelular de lacasa
Se determinó utilizando el extracto enzimático empleando 2,6-dimetoxifenol (DMP) como
sustrato. Se modificó la técnica de Díaz-Godínez (2009) para llevar a cabo la mezcla de
reacción, la cual se preparó con 950 µl de DMP 2mM y 50 μL de extracto enzimático en
buffer de fosfatos 0.1 M a pH 6.5. Después de 1 min de incubación a 40 ºC, se midió la
absorbancia a una longitud de onda de 568 nm, (ε568 10000 mol-1 cm-1). Se considera que el
grupo fenol es oxidado por la lacasa y de este modo se forma una semiquinona que provoca
la coloración, también los dos grupos metoxi pueden ser oxidados, lo cual permite que se
lleve a cabo la polimerización. Una unidad de actividad de lacasa (U), es la cantidad de
enzima que provoca un incremento de una unidad de absorbancia por minuto (Téllez–
Téllez y col., 2008).
Actividad enzimática extracelular de lignino peroxidasa
Se evaluó modificando la técnica de Ten Have y col., (1998). La reacción se llevó mediante
la oxidación de alcohol veratrílico a veratrildehído. Se empleó como sustrato alcohol
veratrílico al 2 mM en buffer de tartrato de sodio 0.1 M a pH 3. Se colocaron 950 µL de
sustrato y 20 µL de extracto enzimático, esta mezcla se utilizó como blanco. Después, se
agregaron 30 µL de H2O2 0.5 mM y toda la mezcla de reacción se incubó por 1 minuto a 35
°C. Se leyó a una longitud de onda de 310 nm (ε568 9300 mol-1 cm-1), se considera que una
unidad de actividad de versátil peroxidasa es la cantidad de enzima que oxida 1µmol de
alcohol veratrílico a veratrildehído por minuto bajo las condiciones mencionadas de
reacción.
Actividad enzimática extracelular de manganeso peroxidasa
Se empleó sulfato manganoso como sustrato a 0.5 mM en buffer de malonato de sodio 50
mM a pH 4.5. Se utilizó 950 µL de sustrato y se añadieron 20 µL de extracto enzimático,
de esta manera se midió el blanco a una longitud de onda de 270 nm (ε568 11590 mol-1
cm-1). Posteriormente se adicionaron 30 µL de H2O2 0.05 mM y se incubó a 30 °C por un
minuto (Giardina y col., 2000). Se leyó la muestra de reacción a la misma longitud de onda
que la muestra blanco.
Actividad enzimática extracelular de versátil peroxidasa
Se midió mediante la oxidación directa de Mn+2 a través de la formación del complejo
tartrato (Pérez-Boada y col., 2005). Se utilizó sulfato de manganeso 0.1 mM como sustrato

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
78
en buffer de tartrato de sodio 0.1 M pH 5 (950 µL) en presencia de H2O2 a 0.1 mM (30 µL)
y extracto enzimático (20 µL). Se utilizó como blanco la mezcla del sustrato y extracto
enzimático, posteriormente se adicionó el H2O2 y se incubo toda la mezcla por 1 min a 24
°C. Se leyó a una longitud de onda de 238 nm (ε238 6500 mol-1 cm-1).
Actividad enzimática extracelular de Dye peroxidasa
Para llevar a cabo la actividad enzimática se utilizó y modificó la técnica de Salvachúa y
col., (2013). Se empleó como sustrato oxidativo ABTS al 2.5 mM y H2O2 como activador
de la reacción al 0.1 mM en buffer de tartrato de sodio 0.1 M a pH 3. Para llevar a cabo la
actividad se colocó 980 µL de sustrato y 20 µL extracto enzimático. Se utilizó como blanco
toda la mezcla de reacción misma que posteriormente fue incubada por 1 min a 45 °C, la
longitud de onda se midió a 436 nm (ε436 29300 mol-1 cm-1).
RESULTADOS Y DISCUSIÓN
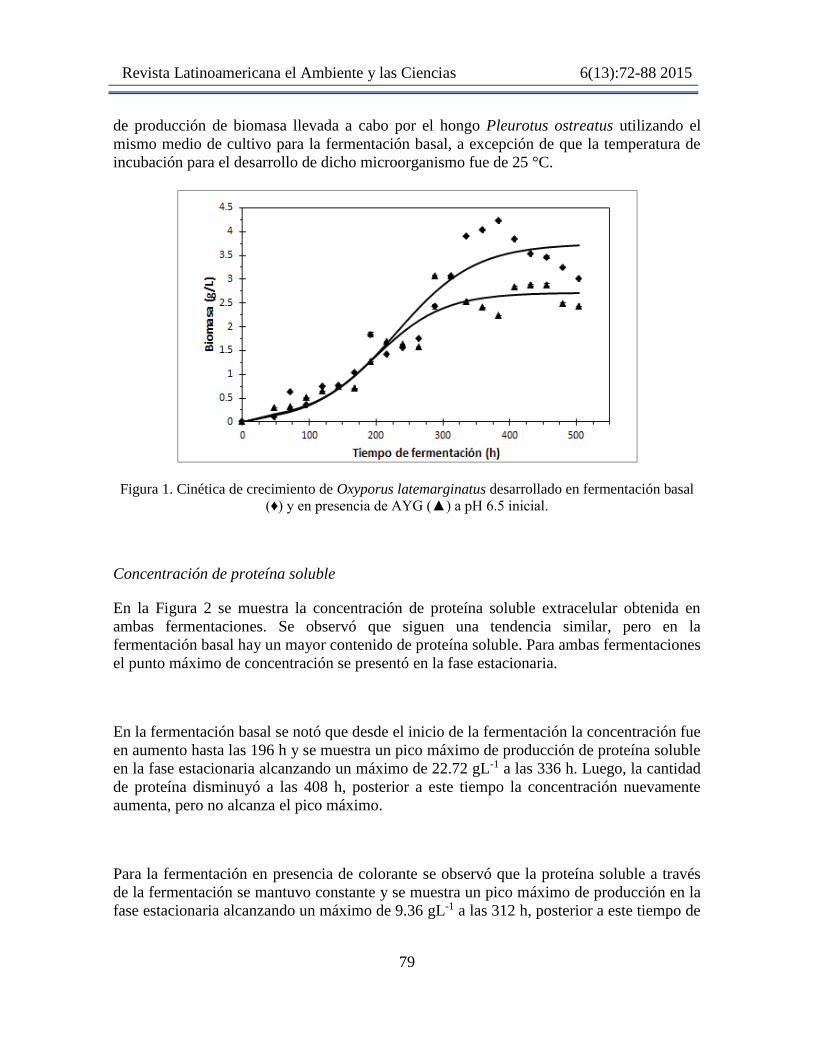
Cinéticas de crecimiento de Oxyporus latemarginatus
En la Figura 1 se muestra la cinética de crecimiento de O. latemarginatus en fermentación
basal y en presencia del colorante amarillo azo. Se observó que en la fermentación basal la
fase de adaptación duró aproximadamente 144 h, posteriormente a este tiempo inició la fase
exponencial de crecimiento que terminó aproximadamente a las 336 h, comenzando la fase
de estacionaria. La Xmáx obtenida fue de 4.2 g/L a las 384 h y la μ fue de 0.016 h-1.
En la fermentación con colorante de observó que el hongo tardó aproximadamente 72 h en
la fase de adaptación y luego comenzó la fase exponencial, misma que terminó
aproximadamente a las 312 h y posteriormente se observó la fase estacionaria. Se puede
apreciar que en el tiempo 264 h hubo una disminución en el crecimiento, este fenómeno
pudiese explicarse como el resultado de la actividad proteolítica, sin embargo, esta
hipótesis tendría que ser comprobada. En esta fermentación la Xmáx obtenida fue de 3.07
g/L a las 312 h y la μ fue de 0.019 h-1.
Los datos obtenidos en esta investigación se puede comparar con los resultados
conseguidos por Téllez-Téllez (2011) quién observó el mismo comportamiento cinético y
Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
79
de producción de biomasa llevada a cabo por el hongo Pleurotus ostreatus utilizando el
mismo medio de cultivo para la fermentación basal, a excepción de que la temperatura de
incubación para el desarrollo de dicho microorganismo fue de 25 °C.
Figura 1. Cinética de crecimiento de Oxyporus latemarginatus desarrollado en fermentación basal
(♦) y en presencia de AYG (▲) a pH 6.5 inicial.
Concentración de proteína soluble
En la Figura 2 se muestra la concentración de proteína soluble extracelular obtenida en
ambas fermentaciones. Se observó que siguen una tendencia similar, pero en la
fermentación basal hay un mayor contenido de proteína soluble. Para ambas fermentaciones
el punto máximo de concentración se presentó en la fase estacionaria.
En la fermentación basal se notó que desde el inicio de la fermentación la concentración fue
en aumento hasta las 196 h y se muestra un pico máximo de producción de proteína soluble
en la fase estacionaria alcanzando un máximo de 22.72 gL-1 a las 336 h. Luego, la cantidad
de proteína disminuyó a las 408 h, posterior a este tiempo la concentración nuevamente
aumenta, pero no alcanza el pico máximo.
Para la fermentación en presencia de colorante se observó que la proteína soluble a través
de la fermentación se mantuvo constante y se muestra un pico máximo de producción en la
fase estacionaria alcanzando un máximo de 9.36 gL-1 a las 312 h, posterior a este tiempo de

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
80
fermentación, la cantidad de proteína disminuyó hasta las 360 h, pero siguió en aumento
después de este tiempo.
Díaz-Godínez (2011) reportó un comportamiento de grafico similar a este, a diferencia de
que él trabajó con Pleurotus ostreatus, el cual igual ya se sabe que es un hongo de
pudrición blanca tal y como lo es O. latemarginatus, pudiéndose deber a que el patrón del
gráfico pertenece a algunos hongos productores de enzimas ligninolíticas
Figura 2. Proteína soluble extracelular de Oxyporus latemarginatus desarrollado en fermentación
basal (♦) y en presencia de AYG (▲) a pH 6.5 inicial.
Consumo de glucosa durante la fermentación
El consumo de glucosa en ambas fermentaciones se presenta en la Figura 3, se puede
observar que para la fermentación basal la concentración de glucosa disminuyó hasta
aproximadamente a 2 gL-1 a las 408 h, y posteriormente se mantuvo constante. Este tiempo
corresponde con la transición de la fase exponencial a fase estacionaria del crecimiento del
hongo.
En la fermentación con colorante la concentración de glucosa disminuyó a un valor mínimo
de 2 gL-1 aproximadamente a las 384 h. El comportamiento se correlaciona con el
crecimiento del hongo, mostrando retardos en el consumo de azúcares en ciertos puntos y
en el desarrollo del hongo. Después de las 384 h la concentración se mantiene más o menos
constante.

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
81
Figura 3. Consumo de glucosa de Oxyporus latemarginatus desarrollado en fermentación basal (♦)
y en presencia de AYG (▲) a pH de 6.5 inicial con extracto de levadura.
Perfil de pH
En la Figura 4 se muestra el comparativo de ambas fermentaciones respecto al pH. Se
observó que ambas fermentaciones siguen la misma tendencia manteniendo valores de pH
en un rango de 5 a 6.5, lo cual sugiere que el medio de cultivo fue incapaz de mantener
constante el pH a lo largo de la fermentación.
Figura 4. Comparación del monitoreo de pH de Oxyporus latemarginatus desarrollado en
fermentación basal (♦) y en presencia de AYG (▲) a pH 6.5 inicial.
0
2
4
6
8
10
12
14
16
18
0 100 200 300 400 500
Co
nce
ntr
ació
n d
e G
luco
sa (
g/L)
Tiempo de fermentación (h)

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
82
Actividad enzimática de lacasa
La actividad de Lac en ambas fermentaciones evaluadas se muestra en la Figura 5. De
acuerdo a los resultados obtenidos se muestra que en la fermentación en presencia de
amarillo azo a las 48 h, presenta un valor de actividad de 59.33 UI/L, es decir, casi tres
veces la actividad enzimática máxima que se presenta en la fermentación basal (20 UI/L).
Inclusive, la actividad mínima que se presenta en la fermentación con colorante (52 UI/L)
es aún más alta en comparación a la actividad determinada en la fermentación basal.
Para el caso de la fermentación basal, se observó que se presentaron incrementos y
descensos de actividad a lo largo de toda la cinética de crecimiento, reportando actividades
bajas. Se observaron dos picos de actividad máxima, uno casi al final de la fermentación
(480 h) con un valor de 18 UI/L, mientas que la mayor actividad se presentó en la fase
estacionaria a las 384 h con 20 UI/L.
En contraste para la fermentación en medio basal, en la del colorante se observa que a las
primeras horas de la fermentación la actividad es alta, pero es en la fase estacionaria donde
se presenta la máxima actividad con un valor de 163.33 UI/L correspondiente a las 336 h.
Figura 5. Actividad de Lac de Oxyporus latemarginatus desarrollado en fermentación basal (♦) y en
presencia de AYG (▲) a pH de 6.5 inicial.
Actividad enzimática de lignino peroxidasa
0
50
100
150
200
0 100 200 300 400 500
Act
ivid
ad e
nzi
mát
ica
(UI/
L)
Tiempo de fermentación (h)

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
83
En la Figura 6 se muestra la actividad de LiP, se aprecia claramente que en la fermentación
con colorante la actividad es mayor, mostrando una actividad máxima de 329.74 UI/L en la
fase estacionaria a diferencia de la basal que mostró un valor máximo de 87.81UI/L en la
misma fase. Del mismo modo, la actividad mínima que se presenta en la fermentación con
colorante es igual mayor que en la basal, mostrando un valor de 50.17 UI/L a diferencia de
14.33 UI/L.
Para la fermentación basal se observó que a las 48 h la actividad empieza de forma
ascendente con 46.59 UI/L, pero después de este tiempo la actividad disminuye y es hasta
la fase estacionaria y al final de la fermentación que se presentan los picos más altos,
siendo a las 504 h donde se lleva a cabo la actividad enzimática más alta. En la otra
fermentación se observó una actividad constante en los primeros tiempos de fermentación,
teniendo un pico máximo de actividad a las 336 h en la fase estacionaria alcanzando un
valor de 329.74 UI/L. Después de este tiempo la actividad de LiP disminuye y al igual que
al inicio, se mantiene constante hasta el término de la fermentación, presentando valores
que van de 114.69 UI/L a 176.70 UI/L.
Figura 6. Actividad de LiP de Oxyporus latemarginatus desarrollado en fermentación basal (♦) y en
presencia de AYG (●) a pH de 6.5 inicial.
Actividad enzimática de manganeso peroxidasa
En la Figura 7 se muestra la actividad de MnP. La menor actividad de MnP registrada para
ambas fermentaciones se presentó a las 48 h en la fase de adaptación con 28.76 UI/L en
basal y 159.62 UI/L en presencia del colorante. De acuerdo al resultado anterior, se puede
observar que la actividad mínima en presencia del colorante es casi cercana a la actividad

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
84
máxima que presenta la fermentación basal con 191.25 UI/L a las 120 h. Se observó que el
punto máximo de actividad enzimática se llevó a cabo en la fase de adaptación, presentando
un valor de 191.25 UI/L (120 h).
En la fermentación con colorante, se observó que la actividad mínima fue de 159.62 UI/L a
las 48 h y la máxima actividad se determinó casi al final de la fermentación, a las 480 h con
533.50 UI/L.
Figura 7. Actividad de MnP de Oxyporus latemarginatus desarrollado en fermentación basal (♦) y
en presencia de AYG (■) a pH de 6.5 inicial.
Actividad enzimática de versátil peroxidasa
La actividad de VP obtenida en ambas fermentaciones, se muestra en la Figura 8. La
actividad no siguió una tendencia similar, excepto al inicio de la fase de adaptación,
coincidiendo con el mismo valor de actividad mínima, 74.35 UI/L a las 48h en basal y en
presencia de colorante a las 72. La actividad máxima se presentó en la fase estacionaria
para basal y en fase exponencial en la fermentación con colorante.
En la fermentación en ausencia del colorante desde el inicio la actividad va en aumento y se
observó que dicho aumento termino en la fase de adaptación a las 144 h, posteriormente la
actividad disminuyó, sin embargo vuelve a aumentar a las 216 h y es hasta la fase
estacionaria donde se presenta el valor máximo de actividad con 966.66 UI/L a las 408 h.

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
85
En la fermentación con colorante la actividad es baja al inicio de la fermentación, siendo la
actividad mínima a las 48 h con un valor de 74.35 UI/L, a partir del tiempo 72 h, la
actividad presentó un patrón no constante de aumento y disminución, sin embargo al final
de la fase exponencial se alcanzó la máxima actividad con un valor de 1443.58 UI/L a las
288 h.
Figura 8. Actividad de VP de Oxyporus latemarginatus desarrollado en fermentación basal (♦) y en
presencia de AYG (▬) a pH de 6.5 inicial.
Actividad enzimática de Dye peroxidasa
En la Figura 9 se muestra la actividad de DyP obtenida en ambas fermentaciones. Se puede
observar que a diferencia de la fermentación basal, la fermentación con el colorante
presentó actividad enzimática durante toda la fermentación. Por otra parte, en la
fermentación basal sólo se detectó actividad en 7 puntos de la fermentación, sin embargo
los valores obtenidos fueron mayores a los determinados en la fermentación con colorante.
Obteniendo un valor máximo de 1312.85 UI/L, en comparación con el valor de 1275.88
UI/L de la fermentación con colorante.
En la fermentación basal se observó que la actividad se presentó al tiempo 288 h y
posteriormente disminuyó en la fase estacionaria a las 432 h. De acuerdo a la actividad
reportada, se observó que la máxima es de 1312.85 UI/L y la mínima presenta un valor de
166.66 UI/L.

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
86
Para la fermentación con colorante se observó que al inicio de la fase de adaptación y
seguida de la fase exponencial, se reportaron valores bajos de actividad a diferencia de los
resultados que se llevaron a cabo en la fase estacionaria, y es en esta fase donde se reportó
el valor máximo con un resultado de 1275.88 UI/L a las 432 h, seguido de este tiempo y
hasta el final de la fermentación la actividad disminuye, pero no alcanza el valor mínimo
porque a las 48 h el resultado fue de 33.56 UI/L correspondiente a la actividad enzimática
mínima.
Figura 9. Actividad de DyP de Oxyporus latemarginatus desarrollado en fermentación basal (♦) y
en presencia de AYG (*) a pH de 6.5 inicial.
CONCLUSIONES
La presencia del colorante amarillo azo no afectó los parámetros cinéticos de crecimiento
de O. latemarginatus (Xmax y ) por lo que no se observó diferencia significativa con los
valores obtenidos en la fermentación basal sin colorante. El colorante amarillo azo induce a
la actividad de las cinco oxidasas evaluadas, Lac, LiP, MnP, DyP y VP y la actividad
máxima alcanzada de 4 de las 5 enzimas evaluadas se presentó en la fermentación con
colorante. Presentando una diferencia de 143.33 UI/L para Lac, 241.93 UI/L para LiP,
342.24 UI/L para MnP y en LiP 476.92 UI/L mayor en comparación con la fermentación
basal. La máxima actividad de las enzimas evaluadas se presenta en la fase estacionaria del
crecimiento de O. latemarginatus. Los análisis de la actividad enzimática realizados
sugieren la participación conjunta y coordinada de las enzimas estudiadas en la oxidación y
probable mineralización del colorante evaluado.

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
87
BIBLIOGRAFÍA
Anjaneyulu, Y.; Sreedhara, C.; Suman, S. (2005); Decolourization of Industrial Effluents –
Available Methods and Emerging Technologies – A Review. Reviews in Environmental
Science and Bio/Technology. 4(4) 245-273.
Bradford, M. (1976); A rapid and sensitive method for the quantitation of microgram of
protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72 248-254.
Diaz, G.; Soriano, J.; Augur, C.; Viniegra, G. (2001); Exopectinases produced by
Aspergillus niger in solid state and submerged fermentation: a comparative study. Journal
Industrial Microbiology Biotechnology. 26(5) 271-275.
Díaz-Godínez R. 2009. Efecto del pH inicial de desarrollo de Pleurotus ostreatus en
fermentación sumergida sobre su actividad de lacasas. Tesis profesional. IPN, Centro de
Investigación en Biotecnología Aplicada. Tlaxcala. México.
Giardina, P.; Palmieri, G.; Fontanella, B.; Rivieccio, V.; Sannia, G. (2000); Manganese
Peroxidase Isoenzymes Produced by Pleurotus ostreatus Grown on Wood Sawdust.
Archives of Biochemistry and Biophysics. 376(1) 171-179.
Khan, A.A.; Husain, Q. (2007); Decolorization and removal of textile and non-textile dyes
from polluted wastewater and dyeing effluent by using potato (Solanum tuberosum) soluble
and immobilized polyphenol oxidase. Bioresource Technology 98(5) 1012-1019.
Miller, G. (1959); Use of Dinitrosalicilic acid for determination of reducing sugar.
Analytical Chemistry. 31(3) 426-428.
Pérez, M.; Ruiz, F.J.; Pogni, R.; Basosi, R.; Choinowski, T.; Martínez, M.J.; Piontek, K.;
Martínez, A.T. (2005); Versatile peroxidase oxidation of high redox potential aromatic
compounds: site-directed mutagenesis, spectroscopic and crystallographic investigation of
three long-range electron transfer pathways. Journal of Molecular Biology. 354(2) 385-
402.
Phugare, S.; Kalyani, D.; Patil, A.; Jadhay, J. (2011); Textile dye degradation by bacterial
consortium and subsequent toxicological analysis of dye and dye metabolites using
cytotoxicity, genotoxicity and oxidative stress studies. Journal of Hazardous Materials.
186(1) 713-723.
Salvachúa, D.; Prieto, A.; Martínez, Á.T.; Martínez, M.J. (2013); Characterization of a
novel dye-decolorizing peroxidase (DyP)-type enzyme from Irpex lacteus and its

Revista Latinoamericana el Ambiente y las Ciencias 6(13):72-88 2015
88
application in enzymatic hydrolysis of wheat straw. Applied and Environmental
Microbiology, 79(14) 4316-4324.
Tamboli, D.P.; Kurade, M.B.; Waghmode, T.R.; Joshi, S.M.; Govindwar, S.P. (2010);
Exploring the ability of Sphingobacterium sp. ATM to degrade textile dye Direct Blue GLL,
mixture of dyes and textile effluent and production of polyhydroxyhexadecanoic acid using
waste biomass generated after dye degradation. Journal of Hazardous Materials. 182(1-3)
169–176.
Téllez, M.; Fernández, F.J.; Montiel, A.M.; Sánchez, C.; Díaz, G. (2008); Growth and
laccase production by Pleurotus ostreatus in submerged and solid-state fermentation.
Applied Microbiology and Biotechnology. 81(4) 675-679.
Téllez-Téllez, M. 2011. Identificación de una lacasa de Pleurotus ostreatus predominante
en condiciones de fermentación líquida y caracterización molecular del gen que la codifica.
Tesis profesional. UAM, División de Ciencias Biológicas y de la Salud. Departamento de
Biotecnología. México D.F.
Ten, R.; Hartmans, S.; Teunissen, P.J.; Field, J.A. (1998); Purication and characterization
of two lignin peroxidase isozymes produced by Bjerkandera sp. strain BOS55. FEBS
Letters. 422(3) 391-394.
Viniegra, G.; Favel, E.; Aguilar, C.N.; Rómero, S.J.; Díaz, G.; Augur, C. (2003);
Advantages of fungal enzyme production in solid state over liquid fermentation systems.
Biochemical Engineering Journal. 13(2-3) 157–167.