rising temperature may negate the stimulatory effect of ... · rising temperature may negate the...
TRANSCRIPT

Rising temperature may negate the stimulatory effect of rising CO2
on growth and physiology of Wollemi pine (Wollemia nobilis)
James D. LewisA,B, Nathan G. PhillipsA,C, Barry A. LoganA,D, Renee A. SmithA,Iker Aranjuelo E, Steve ClarkeF, Catherine A. OffordG, Allison FrithG,H, Margaret BarbourH,Travis HuxmanI and David T. TissueA,J
AUniversity of Western Sydney, Hawkesbury Institute for the Environment, Richmond, NSW 2753, Australia.BFordham University, Louis Calder Center – Biological Field Station, Center for Urban Ecology and Departmentof Biological Sciences, Armonk, NY 10504, USA.
CDepartment of Geography and Environment, Boston University, Boston, MA 02215, USA.DDepartment of Biology, Bowdoin College, Brunswick, ME 04011, USA.EPlant Biology and Ecology Department, Science and Technology Faculty, University of the Basque Country,Barrio Sarriena, 48940 Leioa, Spain.
FUniversity of Western Sydney, Capital Works and Facilities, Richmond, NSW 2753, Australia.GThe Royal Botanic Gardens and Domain Trust, The Australian PlantBank, The Australian Botanic Garden,Mount Annan, NSW 2567, Australia.
HFaculty of Agriculture and Environment, The University of Sydney, NSW 2006, Australia.IEcology and Evolutionary Biology, University of California, Irvine, CA 92697, USA.JCorresponding author. Email: [email protected]
Abstract. Rising atmospheric [CO2] is associated with increased air temperature, and this warming may drive many rareplant species to extinction. However, to date, studies on the interactive effects of rising [CO2] andwarming have focussed onjust a few widely distributed plant species. Wollemi pine (Wollemia nobilisW.G.Jones, K.D.Hill, & J.M.Allen), formerlywidespread in Australia, was reduced to a remnant population of fewer than 100 genetically indistinguishable individuals.Here, we examined the interactive effects of three [CO2] (290, 400 and 650 ppm) and two temperature (ambient,ambient + 4�C) treatments on clonally-propagated Wollemi pine grown for 17 months in glasshouses under well-watered and fertilised conditions. In general, the effects of rising [CO2] and temperature on growth and physiologywere not interactive. Rising [CO2] increased shoot growth, light-saturated net photosynthetic rates (Asat) and net carbon gain.Higher net carbongainwasdue to increasedmaximumapparent quantumyield and reducednon-photorespiratory respirationin the light, which also reduced the light compensation point. In contrast, increasing temperature reduced stem growth andAsat. Compensatory changes inmesophyll conductance and stomatal regulation suggest a narrow functional range of optimalwater and CO2 flux co-regulation. These results suggest Asat and growth of the surviving genotype of Wollemi pinemay continue to increase with rising [CO2], but increasing temperatures may offset these effects, and challenges tophysiological and morphological controls over water and carbon trade-offs may push the remnant wild population ofWollemi pine towards extinction.
Additional keywords: elevated [CO2], growth, photosynthesis, photosynthetic capacity, photosynthetic light response,
pre-industrial [CO2], stomatal conductance.
Received 18 September 2014, accepted 4 May 2015, published online 24 June 2015
Introduction
Rising atmospheric [CO2] and increasing temperatures maysubstantially alter plant physiology and growth, and lead torange shifts or evolution in amenable populations. However,although tree responses to elevated [CO2] have beenextensively studied (Saxe et al. 1998; Ainsworth and Long2005; Norby et al. 2005), much of the work to date hasfocussed on relatively few common species, and the responses
of rare species remain poorly understood, even though rarespecies are more likely to be driven to extinction by climatechange than common species. Further, less is known aboutresponses to past rises in [CO2] (Gerhart and Ward 2010;Lewis et al. 2010; Franks et al. 2013), although the rapidincrease in atmospheric [CO2] over the last 200 years, from along-term range of ~180–280 ppm to 400 ppm at present,suggests that many plant species may be adapted to lower
CSIRO PUBLISHING
Functional Plant Biologyhttp://dx.doi.org/10.1071/FP14256
Journal compilation � CSIRO 2015 www.publish.csiro.au/journals/fpb

[CO2] than they currently experience (Cowling and Sage 1998;Körner 2006; Gerhart et al. 2012; Tissue and Lewis 2012).And, comparatively little is known about the interactive effectsof rising [CO2] and temperature on trees (Ghannoum et al. 2010a;Smith et al. 2012), despite predicted temperature increases of1.7 to 4.4�C by the year 2100 (Solomon et al. 2007). Clearly,predicting the responses of rare tree species to climate changerequires a better understanding of their responses to pre-industrial, modern and future [CO2] and higher air temperature.
Long-lived plant species with low genetic variation andreproductive rates may be particularly susceptible to extinctionas a result of climatic changes because they lack the capacityto sufficiently evolve or migrate in response to rapid increasesin [CO2] or temperature. For example, members of theAraucariaceae, which were once dominant in Australia andinitially evolved under conditions of comparatively high [CO2](Haworth et al. 2011), are thought to have been reduced toremnant stands in part by low [CO2] (Robinson 1994) andprevious changes in climate (Kershaw 1994; Gallagher et al.2003); projected climate change may drive these C3 speciesto extinction (Offord 2011). Only three extant genera ofAraucariaceae remain, and one species, Wollemi pine(Wollemia nobilis W.G.Jones, K.D.Hill, & J.M.Allen), existsas fewer than 100 individuals in the wild. Wollemi pine may beparticularly constrained in its response to climate change, in partdue to its small population size. Although Wollemi pines dateback ~100million years (MacPhail et al. 1995; Zonneveld 2012),all living individuals are genetically indistinguishable (Setoguchiet al. 1998; Peakall et al. 2003), with some of these trees400–450 years old (Banks 2002). Accordingly, Wollemi pinehas limited genetic variation to support evolutionary responses tofuture climate change (Kirschbaum 1994).
The limited genetic variation and pre-industrial origins ofliving Wollemi pine suggests the species has undergone a severegenetic bottleneck where most genotypes failed to survive, butthe surviving genotype had sufficient physiological plasticityto cope with increases in [CO2] and temperature over the last400–450years. Photosynthesis plays a central role in plant carbondynamics, and abundant research has established the key rolesof stomatal conductance (gs), leaf nutrient concentrations, leaftemperature and light availability in regulating photosyntheticresponses to long-term increases in [CO2]. Elevated [CO2]generally is associated with reduced gs (Medlyn et al. 2001;Lewis et al. 2002a) and leaf nitrogen concentrations ([N]) (Lewiset al. 2004); although the relative effectsmay vary among species(Medlyn et al. 2001;Ainsworth andRogers 2007), reduced gs hasbeen observed in Wollemi pine grown under elevated [CO2](Haworth et al.2011).
The effects of rising [CO2] on photosynthetic responsesto temperature and light are more ambiguous, although thetheoretical basis for predicting the interactive effects of rising[CO2] and temperature on photosynthesis is well established.Increasing temperature is predicted to increase the relativeresponse of photosynthesis to elevated [CO2] as a result ofinteractive effects that increase the carboxylation efficiency ofribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco)(Long 1991). However, although some studies report increasesin temperature optima (Lewis et al. 2001;Ghannoum et al. 2010b)and shifts in the light response of photosynthesis (Lewis et al.
1999;Bunce 2014), other studies report no clear effect of elevated[CO2] on photosynthetic responses to temperature (Tjoelker et al.1998; Stinziano and Way 2014) or light (Long and Drake 1991;Grulke et al. 1993). Similarly, empirical studies of the interactiveeffects of rising [CO2] and temperature on these relationshipsin tree seedlings have yielded mixed results (Lewis et al. 1999;Lewis et al. 2004; Ghannoum et al. 2010b), and little is knownabout how past increases in [CO2] have affected theserelationships (Tissue and Lewis 2012).
Our primary objective in this study was to examine the effectsof rising [CO2] and temperature onWollemi pine plants to betterunderstand potential effects of climate change on a species withlimited genetic diversity. More specifically, we examined thegrowth and physiology of Wollemi pine under pre-industrial[CO2], and determined the effects of predicted future increasesin [CO2] and temperature on these traits. We hypothesised that:(i) rising [CO2] and increasing temperature would interactivelyaffect plant growth; (ii) rising [CO2] and increasing temperaturewould interactively affect net photosynthetic rates (A); and(iii) photosynthetic responses to rising [CO2] and increasingtemperature would be associated with changes in therelationships between A and gs, mesophyll conductance (gm),light, temperature and leaf [N]. Climate change effects onseedlings have a much larger impact on population dynamicsand survival compared with effects on larger trees (Lewis et al.2013).Accordingly, our focuson themost susceptible life stage inWollemi pine (Zimmer et al. 2014) is likely to provide crucialinformation for predicting the effects of future climate change onremnant populations of this species.
Materials and methodsPlant material and growth conditions
Wollemi pine (Wollemia nobilis W.G.Jones, K.D.Hill & J.M.Allen) plants were obtained as one-year-old, clonally-propagated, potted plants from the Australian Botanic Garden(Mount Annan, NSW,Australia). Plants were planted one per potin 2.4 L plastic pots filled with a potting mix consisting of1.5 : 0.5 : 1 (v/v/v) coir peat, sand and Perlite, respectively,supplemented with lime, dolomite, iron sulfate, Micromax andNutricote (N:13; Sungro Horticulture, Bellevue,WA, USA). Theplants were randomly assigned to six adjacent, naturally sun-litand temperature-controlled glasshouse compartments. Plantswere grown under 50% shade cloth to provide optimal lightintensities for ex situ Wollemi pine growth (Pohio et al. 2005).Pots were watered twice a week to through-flow with tap water.During the growing season (December–February), pots werewatered ~every 6 weeks with Aquasol fertiliser (N: 23%, P:4%, K:18%, Zn: 0.05%, Cu: 0.06%, Mo: 0.0013%, Fe: 0.06%,Bo: 0.01%; Yates Australia, Padstow, NSW, Australia) at aconcentration of 2 g L–1. Plants were spot sprayed withConfidor pesticide (Bayer CropScience, Melbourne, Vic,Australia), at a concentration of 1 g L–1, when pests (goldenmealy bug (Nipaecoccus aurilanatus) and aphids (Aphis spp.))were detected.
The six glasshouse bays were separated into six factorialcombinations of two temperature and three [CO2] regimes.Temperature and [CO2] were maintained as described in workby Ghannoum et al. (2010a). In brief, three bays tracked local
B Functional Plant Biology J. D. Lewis et al.

(Richmond, NSW, Australia) ambient temperature (i.e. ambienttemperature treatment) and three bays were maintained atambient + 4�C (i.e. elevated temperature treatment). Airtemperature was kept constant during the night and middayperiods; temperature was stepped up and down in two 4�C stepsbefore and after the midday period. Average temperatures for theambient and elevated temperature treatments were 26/18 and30/22�C (day/night) respectively. The temperature set point wasmaintained within +4�C and �2�C for 100% of the time. Withineach temperature treatment, bays were automatically regulatedto maintain pre-industrial (280ppm), ambient (400ppm) andelevated (640ppm) [CO2]. Actual average daytime [CO2]during the study period for the pre-industrial, ambient andelevated treatments was 290, 400 and 650 ppm respectively.The CO2 set point was maintained within� 20 ppm of the target82–86%of the time across bays.Mixing bottledCO2with ambientair (d13C ~ –9.83‰) resulted in a 13CO2 isotopic composition ofd13C of about –5.51 ‰ and –17.02 ‰ under pre-industrial andelevated [CO2] respectively. Air d13C corresponded to averagevaluesdetermined (for thecorresponding treatment) throughout theexperiment, coinciding with gas exchange analyses. Averagevapour pressure deficits in the bays over the growing season inthe ambient temperature treatment were 1.49, 1.69 and 1.62 kPa inthe pre-industrial, ambient and elevated [CO2] treatments,respectively, and in the elevated temperature treatment were1.17, 1.67 and 1.67 kPa in the pre-industrial, ambient andelevated [CO2] treatments respectively. Plants were rotatedwithin glasshouse bays on a fortnightly basis and betweenglasshouse bays on a bi-monthly basis to minimise the impactofglasshousebayenvironmental variationonplant responses to the[CO2] and temperature treatments.
Growth measurementsSeventeen months (April 2010) after the initiation of [CO2] andtemperature treatments, six plants from each of the treatmentcombinations were harvested and separated into leaf and stemtissue. Leaves were further separated into new growth,corresponding to leaves produced since the initiation oftreatments, and initial leaf tissue, corresponding to leavesproduced before the initiation of treatments. Total projectedleaf area of initial leaf tissue and new growth was measuredusing a portable leaf area meter (LI-3100A, Li-Cor BiosciencesInc.,Lincoln,NE,USA).Harvested tissuewasoven-dried at 70�Cuntil completely dry (approximately 1 week) and then massed.For each plant, leaf mass per unit leaf area (LMA) was calculatedas total leaf dry mass/total leaf area. Leaf area per unit shoot drymass (LAR)was calculated as total leaf area/total shoot drymass.
Leaf physiology
Light-saturated net photosynthetic rates (Asat) and stomatalconductance (gs) were measured in November 2008, February,March and June 2009, and January and February 2010 using aportable open gas exchange system (LI-6400,Li-CorBiosciencesInc.) supplying light by a built-in blue and red light-emittingdiode source mounted above the leaf cuvette (LI-6400–02B). Ateach measurement period, measurements were made on matureleaves of apical whorls of 5–10 randomly selected plants per[CO2]� temperature treatment combination. All measurements
(except November 2008) were made at saturating irradiance (Q,700mmolm–2 s–1, 400–700 nm), the target growth [CO2], middaygrowth temperature (26 or 30�C) and a leaf-to-air vapour pressuredeficit (VPD) of 0.9–2.0 kPa, reflecting ambient VPD in theglasshouse compartments. Conditions for measurements inNovember 2008 were the same except that irradiance was1000mmolm–2 s–1. All measurements (except February 2010)were recorded after ~10min, after photosynthesis hadequilibrated. In February 2010, gas-exchange data at the growth[CO2] were determined from measurements of the response ofphotosynthesis (A) to intercellular [CO2] (Ci) and irradiance (Q)(see below).
Photosynthetic CO2 response curvesACi response curves were measured 15 months (February 2010)after treatments were initiated. Curves were measured on 9–10plants in each treatment using protocols previously described (see‘Leaf physiology’ above), except that external [CO2] (Ca) wasapplied after equilibration at the ambient growth [CO2] and thecuvette temperature was maintained at 20�C; Ca was varied 11times between 75 ppm and 2000ppm. Measurements wereautomatically recorded at each Ca set-point when photosynthesishad equilibrated, which was typically 1–2min after a stableCa set-point had been achieved. Gas exchange measurements at thegrowth Ca were used to determine Ls (relative stomatallimitation of photosynthesis). Ls was calculated as Ls = (A0 – A)/A0, where A0 denotes the net rate of CO2 assimilation if stomatalresistance were zero (i.e. Ci =Ca) (Farquhar and Sharkey 1982).ACi curves were also used to estimate Amax (the maximum Aobserved at saturating Q and Ca), Vcmax (apparent, maximalRubisco-limited rate of photosynthesis), Jmax (apparent,maximal electron transport-limited rate of photosynthesis) andTPU (triose phosphate utilisation limitation of photosynthesis).Vcmax, Jmax and TPU were estimated following Sharkey et al.(2007) using the temperature parameterisation described in workby Bernacchi et al. (2003) and without constraining mesophyllconductance.
Photosynthetic light response curvesAQ curves were measured 15 months after initiation of [CO2]and temperature treatments. AQ curves were measured on nine to10 plants in each treatment using protocols described for leafphysiology measurements (see ‘Leaf physiology’ above) at othermeasurement periods except that Q was adjusted in 12 steps,decreasing from 1000 to 0mmolm–2 s–1. Measurements wereautomatically recorded at each Q set-point whenphotosynthesis had equilibrated, which was typically 1–2minafter a stable Q set-point had been achieved.
The AQ curves were individually analysed following Lewiset al. (2000) using the non-rectangular hyperbolic model (Priouland Chartier 1977; Leverenz and Jarvis 1979; Marshall andBiscoe 1980):
QA2�AðF PPFDþ AsatÞ þF PPFD Asat ¼ 0, ð1Þwhere PPFD is the incident photosynthetic photon flux density,A is the photosynthetic rate,F is themaximum apparent quantumyield (the initial slope of the curve),Q is the convexity of the lightresponse curve, andAsat is the light-saturated photosynthetic rate.
Wollemi pine: back to the future Functional Plant Biology C

To obtain a correct intercept on the y-axis (Shapiro et al. 2004),an additional termwas added to the photosynthetic rate. This termwas used as an estimate of the daytime non-photorespiratoryrespiration rate in the light (RL). For some curves, themodelfittedthe curve poorly at low irradiances. For these curves, F and RL
were calculated by linear regression analysis of data below aPPFD of 100mmolm–2 s–1. The light compensation point (G) wascalculated for each curve as the PPFD at which the netphotosynthetic rate equalled zero.
Photosynthetic temperature response curves
The response of A to leaf temperature (TL) was measured at14 months after the initiation of [CO2] and temperaturetreatments. Temperature response curves were measured asdescribed above (see ‘Leaf physiology’) except that five plantswere randomly selected from each treatment and, for each plant,leaf temperaturewas adjusted in seven steps, increasing from~10to 40�C. Temperature was adjusted as described in Ghannoumet al. (2010b). In brief, the A/TL curves were measured by raisingthe block temperature of the leaf cuvette in seven steps (10, 15, 20,25, 30, 35 and 40�C); all plants were measured at the sametemperature before the block was stepped up to the nexttemperature, and plants were allowed to equilibrate for 30minbefore the nextmeasurement period. Concurrent to increasing theblock temperature, the temperature of the glasshouse roomwhereplants were measured was also raised to the same targettemperature, to keep the entire plant at the same temperatureas the measurement leaves.
The rate of O2 evolution under CO2- and light-saturatedconditions is a common and direct measure of the capacity forphotosynthesis (Logan et al. 2010). We measured the responseof photosynthetic O2 evolution at different temperatures onleaves similar in development and exposure to those used forphotosynthetic gas exchange measurements. Detached leaves ofknown area were held in covered Petri dishes with moist filterpaper and transported to the laboratory, where measurementswere initiated 5 to 20min after collection. Measurements wereconducted at 1600mmol photons m–2 s–1 using a gas-phase O2
electrode (Model LD-2 equipped with LS-2 light source,Hansatech, King’s Lynn, UK) according to the protocols inwork by Delieu and Walker (1981). We increased themeasurement temperature in four steps, from 10 to 40�C, bycirculating water, at the target temperature, around the electrodechamber. The air in the chamber was gently replaced with expiredair before measurements to achieve CO2 saturation, as in Li et al.(2000), Wheelwright and Logan (2004) and Logan et al. (2009).
Stomatal behaviour and mesophyll conductance
In order to understand the potential coordination of leaf-levelphotosynthetic biochemistry within the hydraulic structure andcontrol over environmental fluxes in intact leaves, the sensitivityof stomatal conductance to variation in light was evaluated usingGain analysis, as outlined in Huxman and Monson (2003).Briefly, restricted-range light (50–750mmolm–2 s–1) and CO2
(150–800 ppm) response curves were utilised with a focus ondeconstructing the direct and indirect response of stomata to achange in light intensity. Thedirect and indirect responses operateeither through sensing and localised metabolism in guard cells
(Zieger 1983), or the integrated photosynthetic dynamics of themesophyll and information transfer to stomata (Wong et al. 1978)respectively. Over finite intervals of Ci and Q, we calculatedthe overall and component stomatal responses as constrained byinternal CO2 sensitivities and changes in stomatal conductancewhile holding Ci constant (e.g. see Huxman and Monson2003). We then used these functions to evaluate a hypotheticalchange in conductance arising from a 100mmolm–2 s–1 drop inirradiance.
Similarly, we evaluated the impacts of changes in the internalstructures of leaves (mesophyll conductance, gm) from long-termexposure to our environmental treatments for components ofthe CO2 diffusion pathway that are important in controllingphotosynthetic fluxes of CO2. We measured gm through theuse of steady-state isotope fluxes coupled with photosyntheticcalculations from open-flow gas exchange (e.g. see Barbour et al.2007; Jahan et al. 2014). In brief, while measuring CO2 fluxeswith a LI-6400 photosynthetic system, we sampled gases into atunable diode laser (Campbell Scientific, Logan, UT, USA) tomeasure the individual concentrations of varying isotopes of CO2
and compare the measured value of discrimination to theexpectation derived from the model developed by Farquharet al. (1982), and employed in Jahan et al. (2014).
C and N isotope and content analysis
Subsamples of leaf, shoot and root tissue were collected at theend of the experiment (April 2010). Samples were dried at 60�Cfor 48 h before analysis. Samples were analysed to determine theC and N isotope composition using a Flash 1112 ElementalAnalyser (Carbo Erba Instrumentazione, Milan, Italy) coupledto an IRMS Delta C isotope ratio mass spectrometer through aConflo III Interface (Thermo-Finnigan, Dreieich, Germany).Results of carbon isotope ratio analyses are reported in per mil(‰) on the relative d-scale, as d13C, and referenced to theinternational standard V-PDB (Vienna Pee Dee Belemnite)according to the following equation:
d13C ¼�
Rsample
Rstandard
�� 1, ð2Þ
where R is the 13C/12C ratio. The presence of labelled C (CL)in leaves, stems and roots of plants grown at 290 and 650 ppm[CO2] refers to the difference between their TOM d13C andthe value of plants grown under 400 ppm (ambient [CO2];considered as non-labelled plants).
N and C content were determined on leaf, shoot and rootsamples collected from three plants in each treatment. Afterdrying at 60�C for 48 h, samples were ground to powderbefore analysis. N and C content were determined at theServeis Cientifico-Técnics of the University of Barcelona(Barcelona, Spain) using an elemental analyser (EA1108,Series 1; Carbo Erba Instrumentazione).
Air samples from each greenhouse compartment werecollected daily. Air samples were collected using 50mLsyringes (SGE International Pty Ltd, Melbourne, Australia),and stored in 10mL vacutainers (BD Vacutainers, Plymouth,UK), before analysis by gas chromatography-combustion-isotope ratio mass spectrometry (GC-C-IRMS).
D Functional Plant Biology J. D. Lewis et al.
Statistical analysesThis study tested the interactive effects of growth [CO2] andtemperature on growth and photosynthesis in Wollemi pineplants. Statistical tests were conducted using R version 2.10.1(R Core Development Team, Vienna, Austria). Two-wayANOVA was used to test the main and interactive effects ofgrowth [CO2] and temperature on variables measured at only oneperiod, with growth [CO2] treated as a fixed, quantitative factorand growth temperature treated as a fixed, categorical factor. Forphotosynthetic measurements that were conducted at multipletime periods (i.e. Asat, gs,Ci/Ca), three-way ANOVAwas used totest the main and interactive effects of growth [CO2], growthtemperature and measurement period on these variables, withgrowth [CO2] and measurement period treated as fixed,quantitative factors, and growth temperature treated as a fixed,categorical factor. Themeasurement period was treated as a maineffect rather than a repeated-measure because photosyntheticmeasurements were conducted on randomly-selected plants ateach time period. Normal probability plots and plots of residualsversus predicted values were used to assess whether data violatedthe assumptions of normality and homogeneity of variances. Forsignificant two- and three-way interactions, pair-wisecomparisons of means were conducted using Tukey’s HSDtest. In all analyses, test results were considered significantwhen P� 0.05.
Results
In this study, we examined the main and interactive effects ofrising [CO2] and elevated temperature on growth andphotosynthetic properties of Wollemi pine. In general, rising[CO2] increased growth and carbon gain (Table 1). Elevatedtemperature had fewer effects, and these generally were negative(Table 1). Further, there were very few significant interactionsbetween rising [CO2] and temperature (Table 1), indicating theeffects of rising [CO2] generally were consistent acrosstemperature treatments, and the effects of elevated temperaturewere consistent across [CO2] treatments. Responses of specificvariables to rising [CO2] and elevated temperature are outlinedbelow.
Growth
Stem dry mass, new leaf dry mass (the dry mass of leavesproduced after onset of the treatments), total leaf dry mass andtotal shoot dry mass increased with rising [CO2] (Tables 1, 2).New leaf, total leaf and shoot drymasses did not varywith growthtemperature, although stem dry mass was 29% less in plantsgrown in elevated temperature compared with those grown inambient temperature (Tables 1, 2). Initial leaf dry mass (the drymass of leaves produced before the onset of treatments) and totalleaf area did not clearly vary with rising [CO2] or betweentemperature treatments (Tables 1, 2).
The effect of rising [CO2] on leaf mass per unit leaf area(LMA) and leaf area per unit shoot dry mass (LAR) significantlyvaried with growth temperature (Tables 1, 2). LMA increasedwith rising [CO2] in both temperature treatments, but the increasewas larger in the ambient temperature treatment compared withthe elevated temperature treatment. LAR decreased with rising[CO2] in both temperature treatments, and the decrease was
greater in the ambient temperature treatment compared with theelevated temperature treatment. There were no other significantinteractions between [CO2] and temperature treatment on growth(Table 1).
Leaf physiology
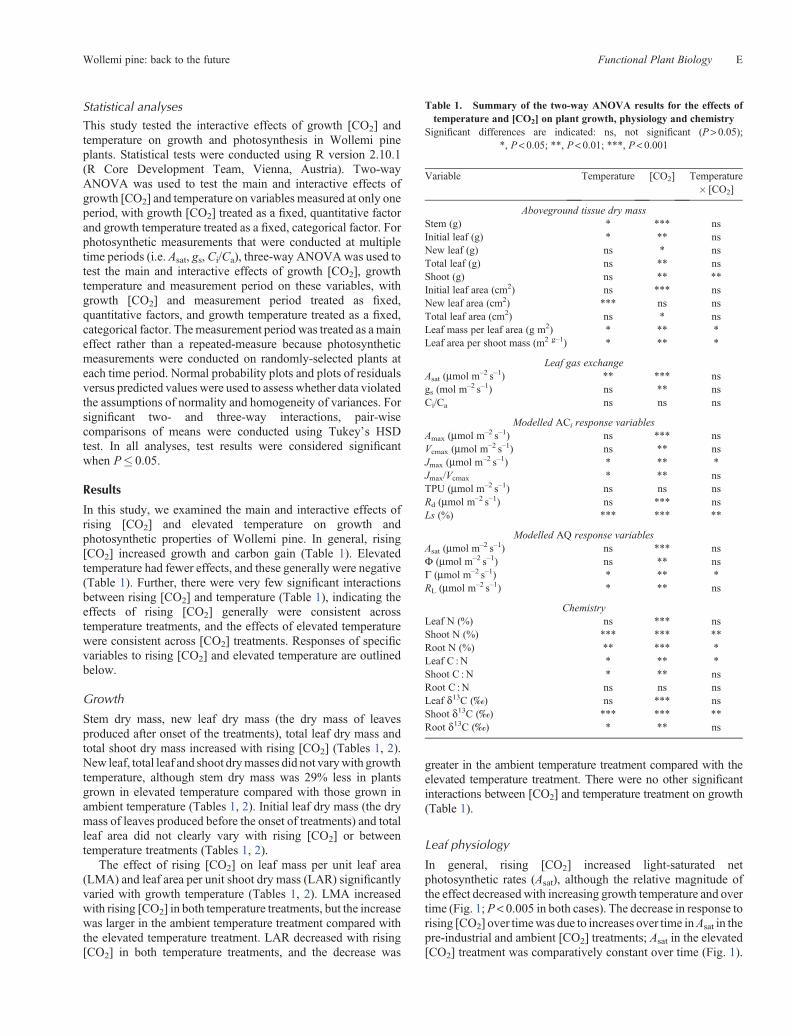
In general, rising [CO2] increased light-saturated netphotosynthetic rates (Asat), although the relative magnitude ofthe effect decreasedwith increasing growth temperature and overtime (Fig. 1; P< 0.005 in both cases). The decrease in response torising [CO2] over timewas due to increases over time inAsat in thepre-industrial and ambient [CO2] treatments; Asat in the elevated[CO2] treatment was comparatively constant over time (Fig. 1).
Table 1. Summary of the two-way ANOVA results for the effects oftemperature and [CO2] on plant growth, physiology and chemistry
Significant differences are indicated: ns, not significant (P> 0.05);*, P< 0.05; **, P< 0.01; ***, P< 0.001
Variable Temperature [CO2] Temperature� [CO2]
Aboveground tissue dry massStem (g) * *** nsInitial leaf (g) * ** nsNew leaf (g) ns * nsTotal leaf (g) ns ** nsShoot (g) ns ** **Initial leaf area (cm2) ns *** nsNew leaf area (cm2) *** ns nsTotal leaf area (cm2) ns * nsLeaf mass per leaf area (g m2) * ** *Leaf area per shoot mass (m2 g–1) * ** *
Leaf gas exchangeAsat (mmol m–2 s–1) ** *** nsgs (mol m–2 s–1) ns ** nsCi/Ca ns ns ns
Modelled ACi response variablesAmax (mmol m–2 s–1) ns *** nsVcmax (mmol m–2 s–1) ns ** nsJmax (mmol m–2 s–1) * ** *Jmax/Vcmax * ** nsTPU (mmol m–2 s–1) ns ns nsRd (mmol m–2 s–1) ns *** nsLs (%) *** *** **
Modelled AQ response variablesAsat (mmol m–2 s–1) ns *** nsF (mmol m–2 s–1) ns ** nsG (mmol m–2 s–1) * ** *RL (mmol m–2 s–1) * ** ns
ChemistryLeaf N (%) ns *** nsShoot N (%) *** *** **Root N (%) ** *** *Leaf C :N * ** *Shoot C :N * ** nsRoot C :N ns ns nsLeaf d13C (‰) ns *** nsShoot d13C (‰) *** *** **Root d13C (‰) * ** ns
Wollemi pine: back to the future Functional Plant Biology E
Although there appeared to be some temporal variation in theinteractive effects of [CO2] and temperature onAsat, this variationwas not significant (P = 0.361).
The effect of [CO2] on stomatal conductance (gs) varied overtime and with temperature treatment (P < 0.015 in both cases;Fig. 1). Data showed that gs increased over time in preindustrial[CO2], butwas relatively constant over time in the elevated [CO2]treatment. As a result, while gs increased with rising [CO2] atthe start of the experiment, gs decreased with rising [CO2] at theend of the experiment; this pattern was consistent between bothtemperature treatments (Table 1). Averaged across measurementperiods, elevated temperature was associated with a largerreduction in gs with rising [CO2] compared with the ambienttemperature treatment.
Rising [CO2] reduced Ci/Ca (Table 1; Fig. 1). In contrast tothe effects of rising [CO2] on Asat and gs, the effect on Ci/Ca wasconsistent across measurement periods (P = 0.801). However,Ci/Ca declined more over time in the ambient temperaturetreatment compared with the elevated temperature treatment(P = 0.020).
Photosynthetic CO2 response curves
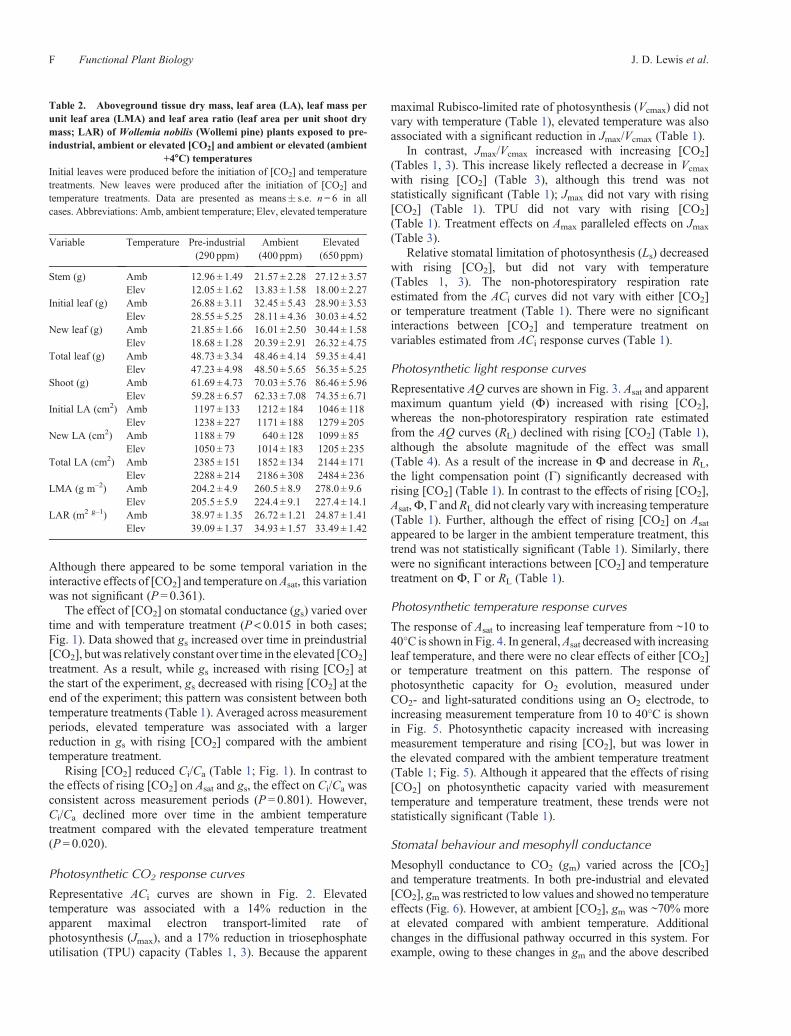
Representative ACi curves are shown in Fig. 2. Elevatedtemperature was associated with a 14% reduction in theapparent maximal electron transport-limited rate ofphotosynthesis (Jmax), and a 17% reduction in triosephosphateutilisation (TPU) capacity (Tables 1, 3). Because the apparent
maximal Rubisco-limited rate of photosynthesis (Vcmax) did notvary with temperature (Table 1), elevated temperature was alsoassociated with a significant reduction in Jmax/Vcmax (Table 1).
In contrast, Jmax/Vcmax increased with increasing [CO2](Tables 1, 3). This increase likely reflected a decrease in Vcmax
with rising [CO2] (Table 3), although this trend was notstatistically significant (Table 1); Jmax did not vary with rising[CO2] (Table 1). TPU did not vary with rising [CO2](Table 1). Treatment effects on Amax paralleled effects on Jmax
(Table 3).Relative stomatal limitation of photosynthesis (Ls) decreased
with rising [CO2], but did not vary with temperature(Tables 1, 3). The non-photorespiratory respiration rateestimated from the ACi curves did not vary with either [CO2]or temperature treatment (Table 1). There were no significantinteractions between [CO2] and temperature treatment onvariables estimated from ACi response curves (Table 1).
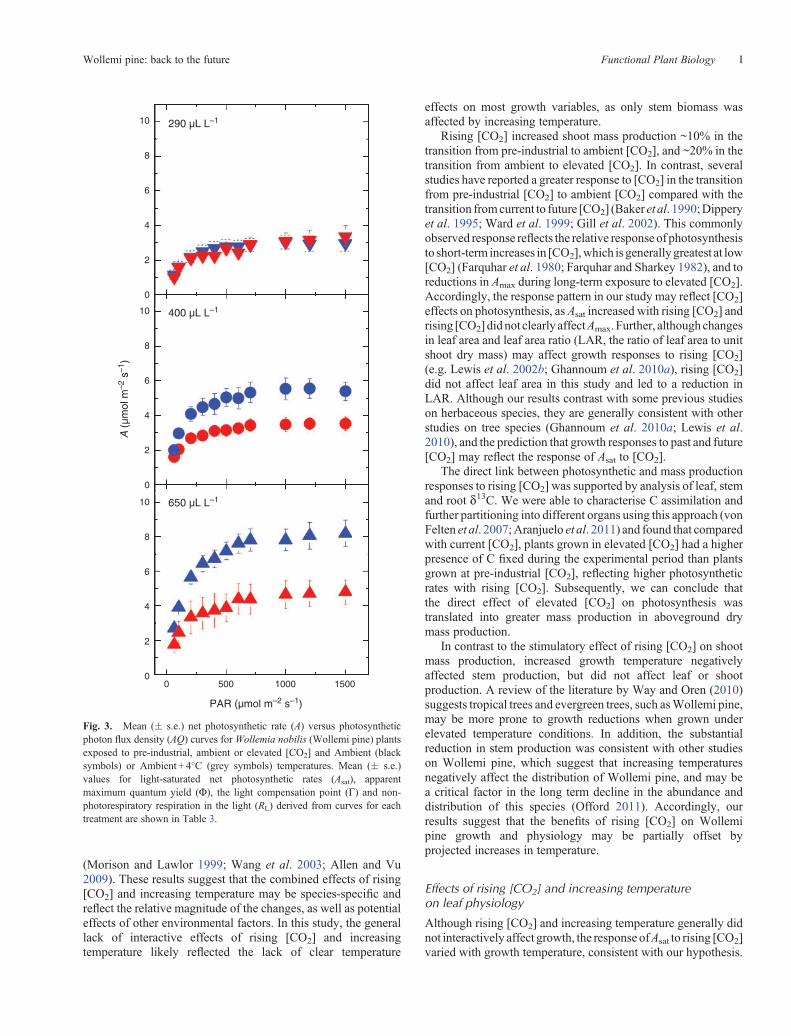
Photosynthetic light response curves
Representative AQ curves are shown in Fig. 3. Asat and apparentmaximum quantum yield (F) increased with rising [CO2],whereas the non-photorespiratory respiration rate estimatedfrom the AQ curves (RL) declined with rising [CO2] (Table 1),although the absolute magnitude of the effect was small(Table 4). As a result of the increase in F and decrease in RL,the light compensation point (G) significantly decreased withrising [CO2] (Table 1). In contrast to the effects of rising [CO2],Asat,F,G andRL did not clearly vary with increasing temperature(Table 1). Further, although the effect of rising [CO2] on Asat
appeared to be larger in the ambient temperature treatment, thistrend was not statistically significant (Table 1). Similarly, therewere no significant interactions between [CO2] and temperaturetreatment on F, G or RL (Table 1).
Photosynthetic temperature response curves
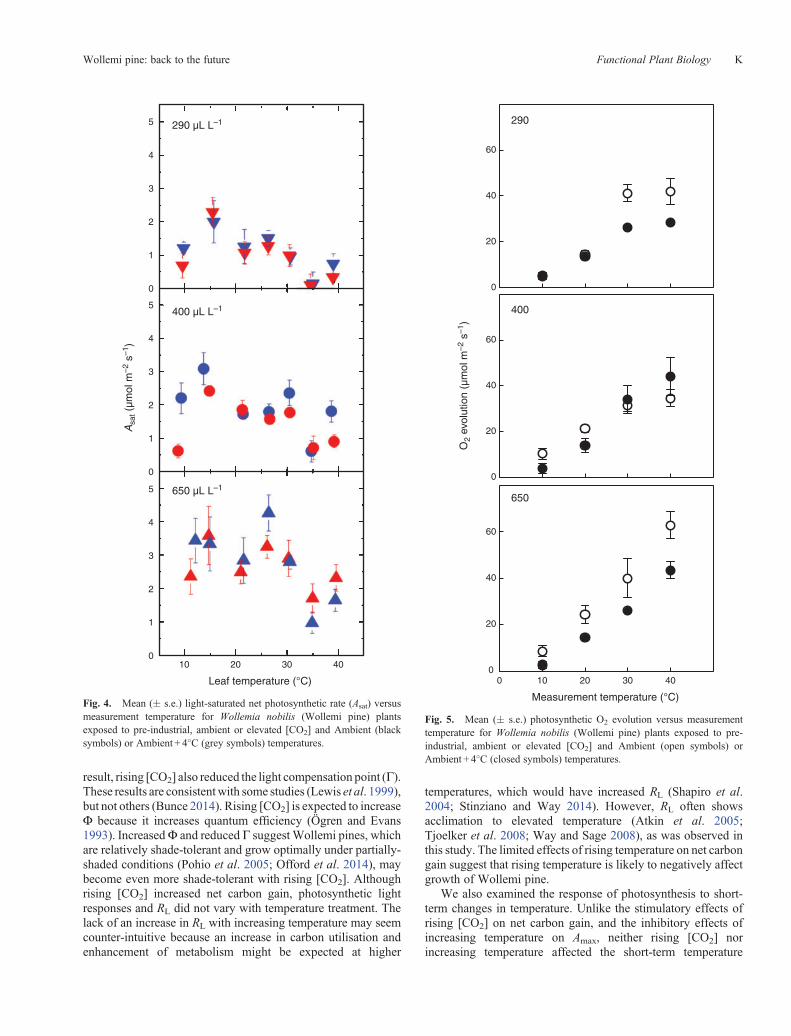
The response of Asat to increasing leaf temperature from ~10 to40�C is shown in Fig. 4. In general,Asat decreasedwith increasingleaf temperature, and there were no clear effects of either [CO2]or temperature treatment on this pattern. The response ofphotosynthetic capacity for O2 evolution, measured underCO2- and light-saturated conditions using an O2 electrode, toincreasing measurement temperature from 10 to 40�C is shownin Fig. 5. Photosynthetic capacity increased with increasingmeasurement temperature and rising [CO2], but was lower inthe elevated compared with the ambient temperature treatment(Table 1; Fig. 5). Although it appeared that the effects of rising[CO2] on photosynthetic capacity varied with measurementtemperature and temperature treatment, these trends were notstatistically significant (Table 1).
Stomatal behaviour and mesophyll conductance
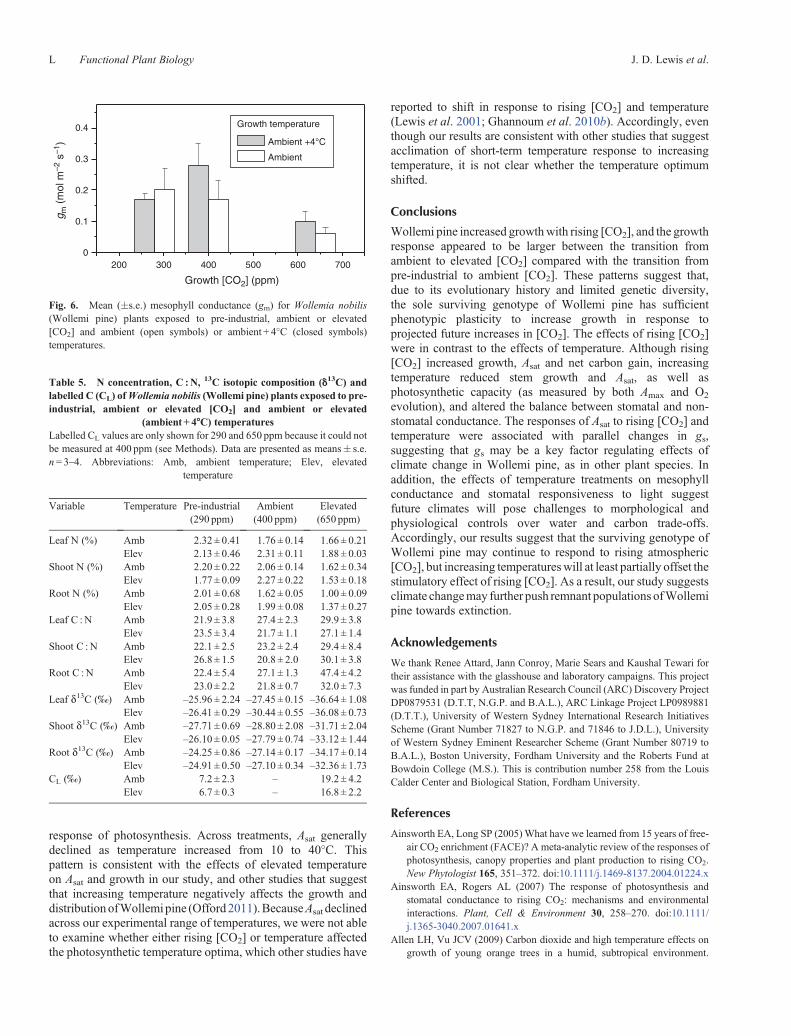
Mesophyll conductance to CO2 (gm) varied across the [CO2]and temperature treatments. In both pre-industrial and elevated[CO2], gmwas restricted to low values and showed no temperatureeffects (Fig. 6). However, at ambient [CO2], gm was ~70% moreat elevated compared with ambient temperature. Additionalchanges in the diffusional pathway occurred in this system. Forexample, owing to these changes in gm and the above described
Table 2. Aboveground tissue dry mass, leaf area (LA), leaf mass perunit leaf area (LMA) and leaf area ratio (leaf area per unit shoot drymass; LAR) of Wollemia nobilis (Wollemi pine) plants exposed to pre-industrial, ambient or elevated [CO2] and ambient or elevated (ambient
+48C) temperaturesInitial leaves were produced before the initiation of [CO2] and temperaturetreatments. New leaves were produced after the initiation of [CO2] andtemperature treatments. Data are presented as means� s.e. n= 6 in allcases. Abbreviations: Amb, ambient temperature; Elev, elevated temperature
Variable Temperature Pre-industrial Ambient Elevated(290 ppm) (400 ppm) (650 ppm)
Stem (g) Amb 12.96 ± 1.49 21.57 ± 2.28 27.12± 3.57Elev 12.05 ± 1.62 13.83 ± 1.58 18.00± 2.27
Initial leaf (g) Amb 26.88 ± 3.11 32.45 ± 5.43 28.90± 3.53Elev 28.55 ± 5.25 28.11 ± 4.36 30.03± 4.52
New leaf (g) Amb 21.85 ± 1.66 16.01 ± 2.50 30.44± 1.58Elev 18.68 ± 1.28 20.39 ± 2.91 26.32± 4.75
Total leaf (g) Amb 48.73 ± 3.34 48.46 ± 4.14 59.35± 4.41Elev 47.23 ± 4.98 48.50 ± 5.65 56.35± 5.25
Shoot (g) Amb 61.69 ± 4.73 70.03 ± 5.76 86.46± 5.96Elev 59.28 ± 6.57 62.33 ± 7.08 74.35± 6.71
Initial LA (cm2) Amb 1197 ± 133 1212 ± 184 1046± 118Elev 1238 ± 227 1171 ± 188 1279± 205
New LA (cm2) Amb 1188 ± 79 640 ± 128 1099± 85Elev 1050 ± 73 1014 ± 183 1205± 235
Total LA (cm2) Amb 2385 ± 151 1852 ± 134 2144± 171Elev 2288 ± 214 2186 ± 308 2484± 236
LMA (g m–2) Amb 204.2 ± 4.9 260.5 ± 8.9 278.0 ± 9.6Elev 205.5 ± 5.9 224.4 ± 9.1 227.4 ± 14.1
LAR (m2 g–1) Amb 38.97 ± 1.35 26.72 ± 1.21 24.87± 1.41Elev 39.09 ± 1.37 34.93 ± 1.57 33.49± 1.42
F Functional Plant Biology J. D. Lewis et al.
photosynthetic dynamics, there was a strong temperature effectonCi at growth irradiancewith rising [CO2].Ci increased from135(� 5; mean� s.e.) to 199 (� 23) to 313 (� 25) ppm in the ambienttemperature treatment, but only increased from 147 (� 4;mean� s.e.) to 153 (� 35) to 178 (� 36) ppm for plants grownat elevated temperature (for pre-industrial, ambient and elevated[CO2], respectively). In concert with this change in diffusionpathway dynamics, gs responses to light differed amongtreatments. In general, gs was quite small and fairly invariant,but the fractional indirect response of [CO2] to light wassignificantly larger for plants grown at ambient [CO2] andambient temperature (41� 8%) when compared with ~12%grand mean across all other treatments. This effect was suchthat a change in Ci with a change in light intensity (associatedwith combined photosynthetic and stomatal responses) was much
lower in plants with greater gm (dCi/dQ=0.3386� gm – 0.1181;R2 = 0.44; P< 0.05), which may highlight a decoupling of thecontrol of stomata in maintaining optimal water and carbon fluxesoutside of the recent environmental conditions (e.g. outside ofambient [CO2] and the two temperature regimes).
N concentration and C isotope composition (d13C)
Across temperature treatments, leaf, shoot and root [N]decreased with rising [CO2] (Tables 1, 5). In contrast, across[CO2] treatments, leaf and root [N] increased with increasedtemperature, although shoot [N] did not clearly vary withtemperature (Tables 1, 5). As a result, rising [CO2] increasedC :N in leaves, shoots and roots, butC : Ndidnot clearlyvarywithtemperature (Tables 1, 5). Although it appeared that rising [CO2]
0
2
4
6
8
(a) (b)
(c) (d)
(e) (d)
Asa
t (µm
ol m
–2 s
–1)
g s (
mol
m–2
s–1
)C
i/Ca
(%)
Asa
t (µm
ol m
–2 s
–1)
g s (
mol
m–2
s–1
)C
i/Ca
(%)
0
0.02
0.04
0.06
0.08
Oct Dec
2008 2009 2010 2008 2009 2010
Feb Apr Jun Aug Oct Dec Feb0
20
40
60
80
0
2
4
6
8
0
0.02
0.04
0.06
0.08
0
20
40
60
80
Oct Dec Feb Apr Jun Aug Oct Dec Feb
(f )
Fig. 1. Effects of growth [CO2] (290, open circles; 400, closed circles; 650 ppm, open squares) on mean (� s.e.)light-saturated net photosynthetic rate (Asat), stomatal conductance (gs) and the ratio of apparent intercellular [CO2]to external [CO2] (Ci/Ca) ofWollemia nobilis (Wollemi pine) plants grown at Ambient (a, c, e) and Ambient + 4�C(b, d, f ) temperature.
Wollemi pine: back to the future Functional Plant Biology G
and increasing temperature interactively affected leaf, shootand root [N], the interactive effects were not significant (Table 1).
Growth [CO2] affected tissue d13C across leaves, shoots androots. The presence of labelled C (CL) was larger in plants inelevated [CO2] compared with pre-industrial [CO2].
Discussion
Rising [CO2] increased shoot mass production, Asat and netcarbon gain of Wollemi pine plants over the 17 monthtreatment period. Increased shoot production reflected
increased stem and leaf mass, while rising [CO2] affected Asat
directly, as Amax did not vary with growth [CO2]. Net carbongain increased with rising [CO2] due to increased F andreduced RL, which, in turn, reduced the light compensationpoint (G). In contrast, increasing temperature reduced stemmass production and Asat, while altering components of thediffusional pathway for photosynthetic gases, reducing theindirect response of stomata to changes in light, and increasingmesophyll conductance. Reductions in Asat with increasingtemperature were associated with reductions in photosyntheticcapacity, as measured both in the presence (Amax) and absence(O2 evolution) of photorespiration, as well as the temperatureresponse of photosynthesis. Moreover, the negative effects ofelevated temperature generally were consistent among [CO2]treatments. These results suggest that while rising [CO2] mayincrease net carbon gain and mass production of Wollemi pineunder current climate conditions, increasing temperatures mayoffset the positive effects of rising atmospheric [CO2] and alterthe coordination of photosynthetic gas exchange regulationin the diffusional pathway between stomatal and non-stomatalcomponents.
Effects of rising [CO2] and increasing temperatureon growth
Rising [CO2] and increasing temperature did not interactivelyaffect growth, in contrast to our hypothesis that the effects wouldbe interactive. Although some studies observe interactive effectsof rising [CO2] and increasing temperature on growth(Ghannoum et al. 2010a), the lack of interactive effects in ourstudy are consistent with studies on a wide range of species
16
12
16
12
8
4
0
16
12
8
4
0
0 100 200 300 400 500
Ci (µL L–1)
600
8
4
0
Asa
t (µm
ol m
–2 s
–1)
290 µL L–1
400 µL L–1
Ls = 0.52
Ls = 0.45
Ls = 0.45
Ls = 0.47
Ls = 0.37
Ls = 0.31
650 µL L–1
Fig. 2. Mean (� s.e.) light-saturated net photosynthetic rates (Asat) versusapparent intercellular [CO2] (ACi) curves forWollemia nobilis (Wollemi pine)plants exposed to pre-industrial, ambient or elevated [CO2] and Ambient(black symbols) orAmbient + 4�C(greysymbols) temperatures.Mean (� s.e.)values for apparent maximal Rubisco-limited rate of photosynthesis (Vcmax)and apparentmaximal electron transport-limited rate of photosynthesis (Jmax),ameasureof thebalancebetweenJmax andVcmax (Jmax/Vcmax), triosephosphateutilisation capacity (TPU), non-photorespiratory respiration (Rd) and relativestomatal limitation (Ls) derived from curves for each treatment are shown inTable 2.
Table 3. Values for maximumCO2 assimilation at saturating light andCO2 (Amax), modelled values for apparent maximal Rubisco-limited rateof photosynthesis (Vcmax) and apparent maximal electron transport-limited rate of photosynthesis (Jmax), a measure of the balancebetween Jmax and Vcmax (Jmax/Vcmax), triosephosphate utilisationcapacity (TPU), non-photorespiratory respiration (Rd) and relativestomatal limitation (Ls) of Wollemia nobilis (Wollemi pine) plantsexposed to pre-industrial, ambient or elevated [CO2] and ambient or
elevated (ambient + 48C) temperaturesData are presented as means� s.e. n= 8–10. Abbreviations: Amb, ambient
temperature; Elev, elevated temperature
Variable Temperature Pre-industrial Ambient Elevated(290 ppm) (400 ppm) (650 ppm)
Amax
(mmol m–2 s–1)Amb 15.8 ± 0.7 16.9 ± 0.8 17.1 ± 0.8Elev 13.5 ± 0.8 14.9 ± 0.6 12.6 ± 0.6
Vcmax
(mmol m–2 s–1)Amb 26.42 ± 1.91 24.97 ± 1.83 24.69 ± 1.24Elev 25.05 ± 1.17 25.11 ± 0.94 21.68 ± 1.35
Jmax
(mmol m–2 s–1)Amb 74.09 ± 4.04 75.11 ± 3.50 79.18 ± 3.75Elev 62.89 ± 3.08 71.56 ± 2.22 59.53 ± 3.27
Jmax/Vcmax Amb 2.84 ± 0.10 3.06 ± 0.12 3.22 ± 0.09Elev 2.54 ± 0.14 2.88 ± 0.12 2.78 ± 0.16
TPU(mmol m–2 s–1)
Amb 5.86 ± 0.31 5.90 ± 0.26 6.28 ± 0.28Elev 4.72 ± 0.27 5.57 ± 0.18 4.63 ± 0.31
Rd (mmol m–2 s–1) Amb 1.83 ± 0.34 1.89 ± 0.36 1.86 ± 0.26Elev 1.68 ± 0.17 2.19 ± 0.26 1.71 ± 0.21
Ls (%) Amb 51.6 ± 2.6 44.7 ± 2.5 36.8 ± 3.4Elev 45.4 ± 2.2 46.5 ± 2.2 30.5 ± 4.6
H Functional Plant Biology J. D. Lewis et al.
(Morison and Lawlor 1999; Wang et al. 2003; Allen and Vu2009). These results suggest that the combined effects of rising[CO2] and increasing temperature may be species-specific andreflect the relative magnitude of the changes, as well as potentialeffects of other environmental factors. In this study, the generallack of interactive effects of rising [CO2] and increasingtemperature likely reflected the lack of clear temperature
effects on most growth variables, as only stem biomass wasaffected by increasing temperature.
Rising [CO2] increased shoot mass production ~10% in thetransition from pre-industrial to ambient [CO2], and ~20% in thetransition from ambient to elevated [CO2]. In contrast, severalstudies have reported a greater response to [CO2] in the transitionfrom pre-industrial [CO2] to ambient [CO2] compared with thetransition fromcurrent to future [CO2] (Baker et al. 1990;Dipperyet al. 1995; Ward et al. 1999; Gill et al. 2002). This commonlyobserved response reflects the relative response of photosynthesisto short-term increases in [CO2],which is generally greatest at low[CO2] (Farquhar et al. 1980; Farquhar and Sharkey 1982), and toreductions in Amax during long-term exposure to elevated [CO2].Accordingly, the response pattern in our study may reflect [CO2]effects on photosynthesis, as Asat increased with rising [CO2] andrising [CO2]didnot clearly affectAmax. Further, althoughchangesin leaf area and leaf area ratio (LAR, the ratio of leaf area to unitshoot dry mass) may affect growth responses to rising [CO2](e.g. Lewis et al. 2002b; Ghannoum et al. 2010a), rising [CO2]did not affect leaf area in this study and led to a reduction inLAR. Although our results contrast with some previous studieson herbaceous species, they are generally consistent with otherstudies on tree species (Ghannoum et al. 2010a; Lewis et al.2010), and the prediction that growth responses to past and future[CO2] may reflect the response of Asat to [CO2].
The direct link between photosynthetic and mass productionresponses to rising [CO2] was supported by analysis of leaf, stemand root d13C. We were able to characterise C assimilation andfurther partitioning into different organs using this approach (vonFelten et al. 2007;Aranjuelo et al. 2011) and found that comparedwith current [CO2], plants grown in elevated [CO2] had a higherpresence of C fixed during the experimental period than plantsgrown at pre-industrial [CO2], reflecting higher photosyntheticrates with rising [CO2]. Subsequently, we can conclude thatthe direct effect of elevated [CO2] on photosynthesis wastranslated into greater mass production in aboveground drymass production.
In contrast to the stimulatory effect of rising [CO2] on shootmass production, increased growth temperature negativelyaffected stem production, but did not affect leaf or shootproduction. A review of the literature by Way and Oren (2010)suggests tropical trees and evergreen trees, such asWollemi pine,may be more prone to growth reductions when grown underelevated temperature conditions. In addition, the substantialreduction in stem production was consistent with other studieson Wollemi pine, which suggest that increasing temperaturesnegatively affect the distribution of Wollemi pine, and may bea critical factor in the long term decline in the abundance anddistribution of this species (Offord 2011). Accordingly, ourresults suggest that the benefits of rising [CO2] on Wollemipine growth and physiology may be partially offset byprojected increases in temperature.
Effects of rising [CO2] and increasing temperatureon leaf physiology
Although rising [CO2] and increasing temperature generally didnot interactively affect growth, the response ofAsat to rising [CO2]varied with growth temperature, consistent with our hypothesis.
290 µL L–1
400 µL L–1
650 µL L–1
10
8
6
4
2
0
10
8
6
4
2
0
10
8
6
4
2
00 500 1000 1500
A (
µmol
m–2
s–1
)
PAR (µmol m–2 s–1)
Fig. 3. Mean (� s.e.) net photosynthetic rate (A) versus photosyntheticphoton flux density (AQ) curves for Wollemia nobilis (Wollemi pine) plantsexposed to pre-industrial, ambient or elevated [CO2] and Ambient (blacksymbols) or Ambient + 4�C (grey symbols) temperatures. Mean (� s.e.)values for light-saturated net photosynthetic rates (Asat), apparentmaximum quantum yield (F), the light compensation point (G) and non-photorespiratory respiration in the light (RL) derived from curves for eachtreatment are shown in Table 3.
Wollemi pine: back to the future Functional Plant Biology I

Other studies have reported amixof additive (Tjoelkeret al. 1998;Lewis et al. 2001; Ghannoum et al. 2010b; Stinziano and Way2014) and interactive (Kellomäki and Wang 1996; Lewis et al.1996) effects of rising [CO2] and temperature on photosynthesis,suggesting that the combined effects of rising [CO2] andincreasing temperature on photosynthesis may reflect bothgenetic and other environmental factors (Stinziano and Way2014). Although most studies have compared temperatureeffects between ambient and elevated [CO2], we comparedacross pre-industrial to elevated [CO2], enabling us to examinewhether increasing temperature has differential effects underrecent and projected increases in [CO2]. Our results suggestthat increasing temperature had similar effects on Wollemipine under past and future [CO2].
Elevated temperature reduced the effect of rising [CO2] onAsat
likely due to parallel effects on gs, as evidenced by the relativelyconstant relative stomatal limitation of photosynthesis (Ls) andCi/Ca across temperature treatments. Plants grownunder elevatedtemperature exhibited lowerLs andCi/Cadata, indicating reducedAsat in those plants reflected reduced gs across the study period.The apparent discrepancy in this pattern in elevated [CO2] plantsmay reflect that at an initial stage, elevated [CO2] increased gs.As an integrator of plant response across the entire treatmentperiod, the fact that (compared with the corresponding ambienttemperature treatment), plants exposed to elevated [CO2] andelevated temperature had lower Asat and labelled C in their totalorganic matter indicates those leaves were composed of a largeramount of pre-label C (with higher d13C values). Taken together,our results suggest that the response of Asat to rising [CO2] andincreased temperature primarily reflected effects of temperatureon gs rather than changes in photosynthetic machinery.
In addition to direct effects on gs, elevated temperature alsoaltered the light sensitivity of gs, and altered the response ofmesophyll conductance (gm) to the [CO2] treatments. Thesechanges altered the balance between stomatal and non-stomatal constraints on [CO2] diffusion, altering both Ci andthe contribution of gs to water-use efficiency in the pre-industrialand elevated [CO2] treatments. The so-called ‘CO2 feedbackloop’, whereby changes in mesophyll metabolism influencestomata, is a potentially important component of the manner inwhich gs contributes to water-use efficiency (e.g. Huxman and
Monson 2003). In the ambient [CO2] treatment, the observedreduction in gswith increasing temperaturewas partially offset bychanges in gm. However, the lack of coordination between gmand gs in the other [CO2] treatments may highlight a decouplingof control of stomata for maintaining optimal water andcarbon fluxes under [CO2] and temperature conditions outsideof recent environmental conditions. These negative effects ofelevated temperature are similar to the negative effects observedon stem growth, and provide additional evidence that the benefitsof rising [CO2] on Wollemi pine physiology may be at leastpartially offset by rising temperatures.
While Amax did not clearly vary with rising [CO2], increasingtemperature was associated with a significant reduction in Amax
across [CO2] treatments, particularly in the apparent maximumelectron transport capacity (Jmax), such that the Jmax/Vcmax ratioalso declinedwith increasing temperature. Reductions inAmax areoften associated with rising [CO2] (Wong et al. 1992; Turnbullet al. 1998; Tissue et al. 1999; Evans et al. 2000; Ellsworth et al.2004), reflecting in part reductions in leaf [N] (Griffin et al.2000; Lewis et al. 2004; Ainsworth and Rogers 2007) or leafC source–sink imbalance (Moore et al. 1999; Lewis et al.2002b; Aranjuelo et al. 2013). The lack of [CO2] effect onphotosynthesis despite a general decline in leaf [N] with rising[CO2] may reflect increased C sink capacity associated withdeclines in LAR. According to the source–sink hypothesis,increases in C sink capacity relative to source activity (e.g.decreases in LAR) may enable plants to maintain Amax as[CO2] rises (Moore et al. 1999; Lewis et al. 2002b; Aranjueloet al. 2013). In our study, despite the decline in leaf [N], leafC : N did not vary with rising [CO2], suggesting that leaves didnot accumulate carbohydrates. Further, rising [CO2] did notreduce either Jmax or TPU, indicating plants in elevated [CO2]maintained higher photosynthetic rates due to higher (comparedwith the corresponding ambient [CO2] treatment) Ci values.Taken together, these results suggest that reductions in LAR,and potentially other shifts in source : sink balance, minimisedcarbohydrate accumulation associated with higher Asat, enablingplants to maintain Amax as [CO2] increased.
In contrastwith the lack of a [CO2] effect onAmax, rising [CO2]increased net carbon gain under low light, due to both increasedapparent maximum quantum yield (F) and reduced RL. As a
Table 4. Modelled values, from AQ curves, for light-saturated net photosynthetic rates (Asat), apparent maximumquantum yield (F), the light compensation point (G) and non-photorespiratory respiration in the light (RL) of Wollemianobilis (Wollemi pine) plants exposed to pre-industrial, ambient or elevated [CO2] andambient or elevated (ambient + 48C)
temperaturesData are presented as means� s.e. n= 7–10. Abbreviations: Amb, ambient temperature; Elev, elevated temperature
Variable Temperature Pre-industrial Ambient Elevated(290 ppm) (400 ppm) (650 ppm)
Asat (mmol m–2 s–1) Amb 2.15 ± 0.19 4.01 ± 0.38 4.86 ± 0.56Elev 3.02 ± 0.39 3.94 ± 0.56 4.79 ± 0.40
F (mmol m–2 s–1) Amb 0.0470 ± 0.0043 0.0941± 0.0108 0.0822 ± 0.0115Elev 0.0543 ± 0.0094 0.0698± 0.0055 0.0963 ± 0.0065
G (mmol m–2 s–1) Amb 7.57 ± 1.56 4.16 ± 0.75 3.23 ± 0.92Elev 7.23 ± 0.96 4.88 ± 0.69 2.16 ± 0.57
RL (mmol m–2 s–1) Amb 0.37 ± 0.08 0.36 ± 0.05 0.22 ± 0.06Elev 0.36 ± 0.05 0.34 ± 0.05 0.20 ± 0.05
J Functional Plant Biology J. D. Lewis et al.
result, rising [CO2] also reduced the light compensation point (G).These results are consistentwith some studies (Lewis et al. 1999),but not others (Bunce 2014). Rising [CO2] is expected to increaseF because it increases quantum efficiency (Ögren and Evans1993). IncreasedF and reduced G suggest Wollemi pines, whichare relatively shade-tolerant and grow optimally under partially-shaded conditions (Pohio et al. 2005; Offord et al. 2014), maybecome even more shade-tolerant with rising [CO2]. Althoughrising [CO2] increased net carbon gain, photosynthetic lightresponses and RL did not vary with temperature treatment. Thelack of an increase in RL with increasing temperature may seemcounter-intuitive because an increase in carbon utilisation andenhancement of metabolism might be expected at higher
temperatures, which would have increased RL (Shapiro et al.2004; Stinziano and Way 2014). However, RL often showsacclimation to elevated temperature (Atkin et al. 2005;Tjoelker et al. 2008; Way and Sage 2008), as was observed inthis study. The limited effects of rising temperature on net carbongain suggest that rising temperature is likely to negatively affectgrowth of Wollemi pine.
We also examined the response of photosynthesis to short-term changes in temperature. Unlike the stimulatory effects ofrising [CO2] on net carbon gain, and the inhibitory effects ofincreasing temperature on Amax, neither rising [CO2] norincreasing temperature affected the short-term temperature
60
290
400
650
40
20
0
60
40
20
0
60
40
20
00 10 20 30
Measurement temperature (°C)
O2
evol
utio
n (µ
mol
m–2
s–1
)
40
Fig. 5. Mean (� s.e.) photosynthetic O2 evolution versus measurementtemperature for Wollemia nobilis (Wollemi pine) plants exposed to pre-industrial, ambient or elevated [CO2] and Ambient (open symbols) orAmbient + 4�C (closed symbols) temperatures.
5
4
3
2
1
0
5
4
3
2
1
0
5
4
3
2
1
010 20 30
Leaf temperature (°C)
40
Asa
t (µm
ol m
–2 s
–1)
290 µL L–1
400 µL L–1
650 µL L–1
Fig. 4. Mean (� s.e.) light-saturated net photosynthetic rate (Asat) versusmeasurement temperature for Wollemia nobilis (Wollemi pine) plantsexposed to pre-industrial, ambient or elevated [CO2] and Ambient (blacksymbols) or Ambient + 4�C (grey symbols) temperatures.
Wollemi pine: back to the future Functional Plant Biology K
response of photosynthesis. Across treatments, Asat generallydeclined as temperature increased from 10 to 40�C. Thispattern is consistent with the effects of elevated temperatureon Asat and growth in our study, and other studies that suggestthat increasing temperature negatively affects the growth anddistributionofWollemipine (Offord2011).BecauseAsat declinedacross our experimental range of temperatures, we were not ableto examine whether either rising [CO2] or temperature affectedthe photosynthetic temperature optima, which other studies have
reported to shift in response to rising [CO2] and temperature(Lewis et al. 2001; Ghannoum et al. 2010b). Accordingly, eventhough our results are consistent with other studies that suggestacclimation of short-term temperature response to increasingtemperature, it is not clear whether the temperature optimumshifted.
Conclusions
Wollemi pine increased growthwith rising [CO2], and the growthresponse appeared to be larger between the transition fromambient to elevated [CO2] compared with the transition frompre-industrial to ambient [CO2]. These patterns suggest that,due to its evolutionary history and limited genetic diversity,the sole surviving genotype of Wollemi pine has sufficientphenotypic plasticity to increase growth in response toprojected future increases in [CO2]. The effects of rising [CO2]were in contrast to the effects of temperature. Although rising[CO2] increased growth, Asat and net carbon gain, increasingtemperature reduced stem growth and Asat, as well asphotosynthetic capacity (as measured by both Amax and O2
evolution), and altered the balance between stomatal and non-stomatal conductance. The responses of Asat to rising [CO2] andtemperature were associated with parallel changes in gs,suggesting that gs may be a key factor regulating effects ofclimate change in Wollemi pine, as in other plant species. Inaddition, the effects of temperature treatments on mesophyllconductance and stomatal responsiveness to light suggestfuture climates will pose challenges to morphological andphysiological controls over water and carbon trade-offs.Accordingly, our results suggest that the surviving genotype ofWollemi pine may continue to respond to rising atmospheric[CO2], but increasing temperatureswill at least partially offset thestimulatory effect of rising [CO2]. As a result, our study suggestsclimate changemay further push remnant populations ofWollemipine towards extinction.
Acknowledgements
We thank Renee Attard, Jann Conroy, Marie Sears and Kaushal Tewari fortheir assistance with the glasshouse and laboratory campaigns. This projectwas funded in part by Australian Research Council (ARC) Discovery ProjectDP0879531 (D.T.T, N.G.P. and B.A.L.), ARC Linkage Project LP0989881(D.T.T.), University of Western Sydney International Research InitiativesScheme (Grant Number 71827 to N.G.P. and 71846 to J.D.L.), Universityof Western Sydney Eminent Researcher Scheme (Grant Number 80719 toB.A.L.), Boston University, Fordham University and the Roberts Fund atBowdoin College (M.S.). This is contribution number 258 from the LouisCalder Center and Biological Station, Fordham University.
References
Ainsworth EA, Long SP (2005)What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses ofphotosynthesis, canopy properties and plant production to rising CO2.New Phytologist 165, 351–372. doi:10.1111/j.1469-8137.2004.01224.x
Ainsworth EA, Rogers AL (2007) The response of photosynthesis andstomatal conductance to rising CO2: mechanisms and environmentalinteractions. Plant, Cell & Environment 30, 258–270. doi:10.1111/j.1365-3040.2007.01641.x
Allen LH, Vu JCV (2009) Carbon dioxide and high temperature effects ongrowth of young orange trees in a humid, subtropical environment.
Table 5. N concentration, C :N, 13C isotopic composition (d13C) andlabelled C (CL) ofWollemia nobilis (Wollemi pine) plants exposed to pre-industrial, ambient or elevated [CO2] and ambient or elevated
(ambient + 48C) temperaturesLabelled CL values are only shown for 290 and 650 ppm because it could notbe measured at 400 ppm (see Methods). Data are presented as means� s.e.n= 3–4. Abbreviations: Amb, ambient temperature; Elev, elevated
temperature
Variable Temperature Pre-industrial Ambient Elevated(290 ppm) (400 ppm) (650 ppm)
Leaf N (%) Amb 2.32 ± 0.41 1.76 ± 0.14 1.66 ± 0.21Elev 2.13 ± 0.46 2.31 ± 0.11 1.88 ± 0.03
Shoot N (%) Amb 2.20 ± 0.22 2.06 ± 0.14 1.62 ± 0.34Elev 1.77 ± 0.09 2.27 ± 0.22 1.53 ± 0.18
Root N (%) Amb 2.01 ± 0.68 1.62 ± 0.05 1.00 ± 0.09Elev 2.05 ± 0.28 1.99 ± 0.08 1.37 ± 0.27
Leaf C : N Amb 21.9 ± 3.8 27.4 ± 2.3 29.9 ± 3.8Elev 23.5 ± 3.4 21.7 ± 1.1 27.1 ± 1.4
Shoot C : N Amb 22.1 ± 2.5 23.2 ± 2.4 29.4 ± 8.4Elev 26.8 ± 1.5 20.8 ± 2.0 30.1 ± 3.8
Root C : N Amb 22.4 ± 5.4 27.1 ± 1.3 47.4 ± 4.2Elev 23.0 ± 2.2 21.8 ± 0.7 32.0 ± 7.3
Leaf d13C (‰) Amb –25.96 ± 2.24 –27.45± 0.15 –36.64± 1.08Elev –26.41 ± 0.29 –30.44± 0.55 –36.08± 0.73
Shoot d13C (‰) Amb –27.71 ± 0.69 –28.80± 2.08 –31.71± 2.04Elev –26.10 ± 0.05 –27.79± 0.74 –33.12± 1.44
Root d13C (‰) Amb –24.25 ± 0.86 –27.14± 0.17 –34.17± 0.14Elev –24.91 ± 0.50 –27.10± 0.34 –32.36± 1.73
CL (‰) Amb 7.2 ± 2.3 – 19.2 ± 4.2Elev 6.7 ± 0.3 – 16.8 ± 2.2
0.1
0200 300 400 500 600 700
0.2
0.3
0.4
g m (
mol
m–2
s–1
)
Growth temperature
Ambient +4°C
Ambient
Growth [CO2] (ppm)
Fig. 6. Mean (�s.e.) mesophyll conductance (gm) for Wollemia nobilis(Wollemi pine) plants exposed to pre-industrial, ambient or elevated[CO2] and ambient (open symbols) or ambient + 4�C (closed symbols)temperatures.
L Functional Plant Biology J. D. Lewis et al.

Agricultural and Forest Meteorology 149, 820–830. doi:10.1016/j.agrformet.2008.11.002
Aranjuelo I, Ebbets AL, Evans RD, Tissue DT, Nogués S, van Gestel N,Payton P, Ebbert V, Adams III WW, Nowak RS, Smith SD (2011)Maintenance of C sinks sustains enhanced C assimilation during long-term exposure to elevated [CO2] in Mojave Desert shrubs. Oecologia167, 339–354. doi:10.1007/s00442-011-1996-y
Aranjuelo I, Sanz-SáezÁ, Jauregui I, Irigoyen JJ, Araus JL, Sánchez-DíazM,Erice G (2013) Harvest index, a parameter conditioning responsivenessof wheat plants to elevated CO2. Journal of Experimental Botany 64,1879–1892. doi:10.1093/jxb/ert081
Atkin OK, Bruhn D, Hurry VM, Tjoelker MG (2005) Evans Review No. 2:The hot and the cold: unravelling the variable response of plant respirationto temperature. Functional Plant Biology 32, 87–105. doi:10.1071/FP03176
Baker JT, Allen LH Jr, Boote KJ (1990) Growth and yield responses of rice tocarbon dioxide concentration. The Journal of Agricultural Science 115,313–320. doi:10.1017/S0021859600075729
Banks JCG (2002)Wollemi pine: treefind of the 20thCentury. In ‘Australia’sever-changing forests. Proceedings of the fifth national conference onAustralian forest history’. (Eds JGDargavel,BLibbis) pp. 85–89. (CRES:Canberra)
BarbourMM, Farquhar GD, HansonDT, Bickford CP, Powers H,McDowellNG (2007) A new measurement technique reveals temporal variation ind18O of leaf-respired CO2. Plant, Cell & Environment 30, 456–468.doi:10.1111/j.1365-3040.2007.01633.x
Bernacchi CJ, Pimentel C, Long SP (2003) In vivo temperature responsefunctions of parameters required to model RuBP-limited photosynthesis.Plant, Cell & Environment 26, 1419–1430. doi:10.1046/j.0016-8025.2003.01050.x
Bunce JA (2014) Limitations to soybean photosynthesis at elevated carbondioxide in free-air enrichment and open top chamber systems. PlantScience 226, 131–135. doi:10.1016/j.plantsci.2014.01.002
Cowling SA, Sage RF (1998) Interactive effects of low atmospheric CO2 andelevated temperature on growth, photosynthesis and respiration inPhaseolus vulgaris. Plant, Cell & Environment 21, 427–435.doi:10.1046/j.1365-3040.1998.00290.x
Delieu T, Walker DA (1981) Polarographic measurement of photosyntheticoxygen evolution by leaf discs. New Phytologist 89, 165–178.doi:10.1111/j.1469-8137.1981.tb07480.x
Dippery JK, Tissue DT, Thomas RB, Strain BR (1995) Effects of low andelevated CO2 on C3 and C4 annuals. I. Growth and biomass allocation.Oecologia 101, 13–20. doi:10.1007/BF00328894
Ellsworth DS, Reich PB, Naumburg ES, Koch GW, Kubiske ME, Smith SD(2004) Photosynthesis, carboxylation and leaf nitrogen responses of 16species to elevated pCO2 across four free-air CO2 enrichment experimentsin forest, grassland and desert. Global Change Biology 10, 2121–2138.doi:10.1111/j.1365-2486.2004.00867.x
Evans JR, Schortemeyer M, McFarlane N, Atkin OK (2000) Photosyntheticcharacteristics of 10 Acacia species grown under ambient and elevatedatmospheric CO2. Australian Journal of Plant Physiology 27, 13–25.
Farquhar GD, Sharkey TD (1982) Stomatal conductance and photosynthesis.Annual Review of Plant Physiology and Plant Molecular Biology 33,317–345. doi:10.1146/annurev.pp.33.060182.001533
Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemical model ofphotosynthetic CO2 assimilation in leaves of C3 species. Planta 149,78–90. doi:10.1007/BF00386231
Farquhar GD, O’Leary MH, Berry JA (1982) On the relationship betweencarbon isotope discrimination and the intercellular carbon dioxideconcentration in leaves. Australian Journal of Plant Physiology 9,121–137. doi:10.1071/PP9820121
Franks PJ, Adams MA, Amthor JS, Barbour MM, Berry JA, Ellsworth DS,FarquharGD,GhannoumO,Lloyd J,McDowellN,NorbyRJ, TissueDT,von Caemmerer S (2013) Sensitivity of plants to changing atmospheric
CO2 concentration: from the geological past to the next century. NewPhytologist 197, 1077–1094. doi:10.1111/nph.12104
Gallagher SJ, Greenwood DR, Taylor D, Smith AJ, Wallace MW, HoldgateGR (2003) The Pliocene climatic and environmental evolution ofsoutheastern Australia: evidence from the marine and terrestrial realm.Palaeogeography, Palaeoclimatology, Palaeoecology 193, 349–382.doi:10.1016/S0031-0182(03)00231-1
Gerhart LM, Ward JK (2010) Plant responses to low [CO2] of the past. NewPhytologist 188, 674–695. doi:10.1111/j.1469-8137.2010.03441.x
Gerhart LM, Harris JM, Nippert JB, Sandquist DR, Ward JK (2012) Glacialtrees from the La Brea tar pits show physiological constraints of lowCO2.New Phytologist 194, 63–69. doi:10.1111/j.1469-8137.2011.04025.x
Ghannoum O, Phillips NG, Conroy JP, Smith RA, Attard RD, Woodfield R,Logan BA, Lewis JD, Tissue DT (2010a) Exposure to preindustrial,current and future atmospheric CO2 and temperature differentially affectsgrowth and photosynthesis in Eucalyptus. Global Change Biology 16,303–319. doi:10.1111/j.1365-2486.2009.02003.x
Ghannoum O, Phillips NG, Sears MA, Logan BA, Lewis JD, Conroy JP,Tissue DT (2010b) Photosynthetic responses of two eucalypts toindustrial-age changes in atmospheric [CO2] and temperature. Plant,Cell & Environment 33, 1671–1681. doi:10.1111/j.1365-3040.2010.02172.x
Gill RA, Polley HW, Johnson HB, Anderson LJ, Maherali H, Jackson RB(2002)Nonlinear grassland responses to past and future atmosphericCO2.Nature 417, 279–282. doi:10.1038/417279a
Griffin KL, Tissue DT, Turnbull MH, Whitehead D (2000) The onset ofphotosynthetic acclimation to elevatedCO2partial pressure infield-grownPinus radiata D.Don. after 4 years. Plant, Cell & Environment 23,1089–1098. doi:10.1046/j.1365-3040.2000.00622.x
Grulke NE, Hom JL, Roberts SW (1993) Physiological adjustment of twofull-sib families of ponderosa pine to elevated CO2. Tree Physiology 12,391–401. doi:10.1093/treephys/12.4.391
HaworthM,Elliott-KingstonC,McElwain JC (2011)The stomatalCO2proxydoes not saturate at high atmospheric CO2 concentrations: evidence fromstomatal index responses of Araucariaceae conifers. Oecologia 167,11–19. doi:10.1007/s00442-011-1969-1
Huxman TE, Monson RK (2003) Stomatal responses of C3, C3-C4 and C4
Flaveria species to light and intercellularCO2 concentration: implicationsfor the evolution of stomatal behaviour. Plant, Cell & Environment 26,313–322. doi:10.1046/j.1365-3040.2003.00964.x
Jahan E, Amthor J, Farquhar G, Trethowan R, BarbourMM (2014) Variationin mesophyll conductance among Australian wheat genotypes.Functional Plant Biology 41, 568–580. doi:10.1071/FP13254
Kellomäki S, Wang K (1996) Photosynthetic response to needle waterpotentials in Scots pine after a four-year exposure to elevated CO2
and temperature. Tree Physiology 16, 765–772. doi:10.1093/treephys/16.9.765
Kershaw AP (1994) Pleistocene vegetation of the humid tropics ofnortheastern Queensland, Australia. Palaeogeography,Palaeoclimatology, Palaeoecology 109, 399–412. doi:10.1016/0031-0182(94)90188-0
Kirschbaum MUF (1994) The sensitivity of C3 photosynthesis to increasingCO2concentration: a theoretical analysis of its dependenceon temperatureand background CO2 concentration. Plant, Cell & Environment 17,747–754. doi:10.1111/j.1365-3040.1994.tb00167.x
KörnerC (2006)PlantCO2 responses: an issue of definition, time and resourcesupply. New Phytologist 172, 393–411. doi:10.1111/j.1469-8137.2006.01886.x
Leverenz JW, Jarvis PG (1979) Photosynthesis in Sitka spruce. VIII. Theeffects of light flux density and direction on the rate of net photosynthesisand the stomatal conductance of needles. Journal of Applied Ecology 16,919–932. doi:10.2307/2402865
Lewis JD, Tissue DT, Strain BR (1996) Seasonal response of photosynthesisto elevatedCO2 in loblollypine (Pinus taedaL)over twogrowingseasons.
Wollemi pine: back to the future Functional Plant Biology M

Global Change Biology 2, 103–114. doi:10.1111/j.1365-2486.1996.tb00055.x
Lewis JD, Olszyk D, Tingey DT (1999) Seasonal patterns of photosyntheticlight response in Douglas-fir seedlings subjected to elevated atmosphericCO2 and temperature. Tree Physiology 19, 243–252. doi:10.1093/treephys/19.4-5.243
Lewis JD, McKane RB, Tingey DT, Beedlow PA (2000) Vertical gradientsin photosynthetic light response within an old-growth Douglas-fir andwestern hemlock canopy. Tree Physiology 20, 447–456. doi:10.1093/treephys/20.7.447
Lewis JD, Lucash M, Olszyk D, Tingey DT (2001) Seasonal patterns ofphotosynthesis in Douglas fir seedlings during the third and fourth year ofexposure to elevated CO2 and temperature. Plant, Cell & Environment24, 539–548. doi:10.1046/j.1365-3040.2001.00700.x
Lewis JD, LucashM, Olszyk DM, Tingey DT (2002a) Stomatal responses ofDouglas-fir seedlings to elevated carbon dioxide and temperature duringthe third and fourth years of exposure. Plant, Cell & Environment 25,1411–1421. doi:10.1046/j.1365-3040.2002.00923.x
Lewis JD, Wang XZ, Griffin KL, Tissue DT (2002b) Effects of age andontogeny on photosynthetic responses of a determinate annual plant toelevated CO2 concentrations. Plant, Cell & Environment 25, 359–368.doi:10.1046/j.0016-8025.2001.00815.x
Lewis JD, Lucash M, Olszyk DM, Tingey DT (2004) Relationships betweenneedle nitrogen concentration and photosynthetic responses of Douglas-fir seedlings to elevated CO2 and temperature. New Phytologist 162,355–364. doi:10.1111/j.1469-8137.2004.01036.x
Lewis JD, Ward JK, Tissue DT (2010) Phosphorus supply drives nonlinearresponses of cottonwood (Populus deltoides) to increases in CO2
concentration from glacial to future concentrations. New Phytologist187, 438–448. doi:10.1111/j.1469-8137.2010.03307.x
Lewis JD, Smith RA, Ghannoum O, Logan BA, Phillips NG, Tissue DT(2013) Industrial-age changes in atmospheric [CO2] and temperaturedifferentially alter responses of faster- and slower-growing Eucalyptusseedlings to short-term drought. Tree Physiology 33, 475–488.doi:10.1093/treephys/tpt032
LiX-P,BjorkmanO, ShihC,GrossmanAR,RosenquistM, JanssonS,NiyogiKK (2000) A pigment-binding protein essential for regulation ofphotosynthetic light harvesting. Nature 403, 391–395. doi:10.1038/35000131
LoganBA,CombsA,MyersK,KentR, StanleyL,TissueDT(2009)Seasonalresponse of photosynthetic electron transport and energy dissipation inthe eighth year of exposure to elevated atmospheric CO2 (FACE) inPinus taeda (loblolly pine). Tree Physiology 29, 789–797. doi:10.1093/treephys/tpp019
Logan BA, Hricko CR, Lewis JD, Ghannoum O, Phillips NG, Smith R,Conroy JP, Tissue DT (2010) Examination of pre-industrial and future[CO2] reveals the temperature-dependent CO2 sensitivity of lightenergy partitioning at PSII in eucalypts. Functional Plant Biology 37,1041–1049. doi:10.1071/FP10113
Long SP (1991) Modification of the response of photosynthetic productivityto rising temperature by atmospheric CO2 concentrations: has itsimportance been underestimated? Plant, Cell & Environment 14,729–739. doi:10.1111/j.1365-3040.1991.tb01439.x
Long SP, Drake BG (1991) Effect of the long-term elevation of CO2
concentration in the field on the quantum yield of photosynthesis ofthe C3 sedge, Scirpus olneyi. Plant Physiology 96, 221–226. doi:10.1104/pp.96.1.221
MacPhail MK, Hill K, Partridge AD, Truswell EM (1995) ‘Wollemi Pine’ -old pollen records for a newly discovered genus of gymnosperms.Geology Today 11, 48–50.
Marshall B, Biscoe PV (1980) A model for C3 leaves describing thedependence of net photosynthesis on irradiance: II. Application to theanalysis of flag leaf photosynthesis. Journal of Experimental Botany 31,41–48. doi:10.1093/jxb/31.1.41
Medlyn BE, Barton CVM, BroadmeadowMSJ, Ceulemans R, De Angelis P,Forstreuter M, Freeman M, Jackson SB, Kellomaki S, Laitat E, Rey A,RoberntzP,SigurdssonBD,Strassemeyer J,WangK,CurtisPS, JarvisPG(2001) Stomatal conductance of forest species after long-term exposure toelevated CO2 concentration: a synthesis. New Phytologist 149, 247–264.doi:10.1046/j.1469-8137.2001.00028.x
Moore BD, Cheng SH, Sims D, Seemann JR (1999) The biochemical andmolecular basis for photosynthetic acclimation to elevated atmosphericCO2. Plant, Cell & Environment 22, 567–582. doi:10.1046/j.1365-3040.1999.00432.x
Morison JIL, Lawlor DW (1999) Interactions between increasing CO2
concentration and temperature on plant growth. Plant, Cell &Environment 22, 659–682. doi:10.1046/j.1365-3040.1999.00443.x
Norby RJ, DeLucia EH, Gielen B, Calfapietra C, Giardina CP, King JS,Ledford J, McCarthy HR,Moore DJP, Ceulemans R, De Angelis P, FinziAC, Karnosky DF, Kubiske ME, Lukac M, Pregitzer KS, Scarascia-Mugnozza GE, Schlesinger WH, Oren R (2005) Forest response toelevated CO2 is conserved across a broad range of productivity.Proceedings of the National Academy of Sciences of the United Statesof America 102, 18052–18056. doi:10.1073/pnas.0509478102
Offord CA (2011) Pushed to the limit: consequences of climate change forthe Araucariaceae: a relictual rain forest family. Annals of Botany 108,347–357. doi:10.1093/aob/mcr135
Offord CA, Meagher PF, Zimmer HC (2014) Growing up or growing out?How soil pH and light affect seedling growth of a relictual rainforest tree.AoB Plants 6, plu011. doi:10.1093/aobpla/plu011
Ögren E, Evans JR (1993) Photosynthetic light response curves. I. Theinfluence of CO2 partial pressure and leaf inversion. Planta 189,182–190. doi:10.1007/BF00195075
Peakall R, Ebert D, Scott LJ, Meagher PF, Offord CA (2003) Comparativegenetic study confirms exceptionally low genetic variation in the ancientand endangered relictual conifer, Wollemia nobilis (Araucariaceae).Molecular Ecology 12, 2331–2343. doi:10.1046/j.1365-294X.2003.01926.x
PohioKE,WallaceHM, Peters RF, Smith TE, Trueman SJ (2005) Cuttings ofWollemi pine tolerate moderate photoinhibition and remain highlycapable of root formation. Trees 19, 587–595. doi:10.1007/s00468-005-0418-3
Prioul JL, Chartier P (1977) Partitioning of transfer and carboxylationcomponents of intracellular resistance to photosynthetic CO2 fixation:a critical analysis of the methods used. Annals of Botany 41, 789–800.
Robinson JM (1994) Speculations on carbon dioxide starvation, latetertiary evolution of stomatal regulation and floristic modernization.Plant, Cell & Environment 17, 345–354. doi:10.1111/j.1365-3040.1994.tb00303.x
Saxe H, Ellsworth DS, Heath J (1998) Tree and forest functioning in anenriched CO2 atmosphere. New Phytologist 139, 395–436. doi:10.1046/j.1469-8137.1998.00221.x
Setoguchi H, Asakawa Osawa T, Pintaud J-C, Jaffré T, Veillon J-M (1998)Phylogenetic relationships within Araucariaceae based on rbcL genesequences. American Journal of Botany 85, 1507–1516. doi:10.2307/2446478
Shapiro JB, Griffin KL, Lewis JD, Tissue DT (2004) Response of Xanthiumstrumarium leaf respiration in the light to elevated CO2 concentration,nitrogen availability and temperature. New Phytologist 162, 377–386.doi:10.1111/j.1469-8137.2004.01046.x
Sharkey TD, Bernacchi CJ, Farquhar GD, Singsaas EL (2007) Fittingphotosynthetic carbon dioxide response curves for C3 leaves. Plant,Cell & Environment 30, 1035–1040. doi:10.1111/j.1365-3040.2007.01710.x
Smith RA, Lewis JD, Ghannoum O, Tissue DT (2012) Leaf structuralresponses to pre-industrial, current and elevated atmospheric [CO2]and temperature affect leaf function in Eucalyptus sideroxylon.Functional Plant Biology 39, 285–296. doi:10.1071/FP11238
N Functional Plant Biology J. D. Lewis et al.

Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M,Miller HL (2007) ‘Climate change 2007. The physical science basis.Contribution of Working Group I to the fourth assessment report of theIntergovernmental Panel on Climate Change’. (Cambridge UniversityPress: Cambridge, UK)
Stinziano JR, Way DA (2014) Combined effects of rising [CO2] andtemperature on boreal forests: growth, physiology and limitations.Botany 92, 425–436. doi:10.1139/cjb-2013-0314
Tissue DT, Lewis JD (2012) Learning from the past: how low [CO2] studiesinform plant and ecosystem response to future climate change. NewPhytologist 194, 4–6. doi:10.1111/j.1469-8137.2012.04081.x
Tissue DT, Griffin KL, Ball JT (1999) Photosynthetic adjustment in field-grown ponderosa pine trees after six years of exposure to elevated CO2.Tree Physiology 19, 221–228. doi:10.1093/treephys/19.4-5.221
Tjoelker MG, Oleksyn J, Reich PB (1998) Seedlings of five boreal treespecies differ in acclimation of net photosynthesis to elevated CO2 andtemperature. Tree Physiology 18, 715–726. doi:10.1093/treephys/18.11.715
Tjoelker MG, Oleksyn J, Reich PB, Zytkowiak R (2008) Coupling ofrespiration, nitrogen, and sugars underlies convergent temperatureacclimation in Pinus banksiana across wide-ranging sites andpopulations. Global Change Biology 14, 782–797. doi:10.1111/j.1365-2486.2008.01548.x
Turnbull MH, Tissue DT, Griffin KL, Rogers GND, Whitehead D (1998)Photosynthetic acclimation to long-term exposure to elevated CO2
concentration in Pinus radiata D.Don. is related to age of needles.Plant, Cell & Environment 21, 1019–1028. doi:10.1046/j.1365-3040.1998.00374.x
von Felten S, Hattenschwiler S, Saurer M, Siegwolf R (2007) Carbonallocation in shoots of alpine treeline conifers in a CO2 enrichedenvironment. Trees – Structure and Function 21, 283–294. doi:10.1007/s00468-006-0118-7
Wang KY, Kellomäki S, Zha T (2003) Modifications in photosyntheticpigments and chlorophyll fluorescence in 20-year-old pine trees after a
four-year exposure to carbon dioxide and temperature elevation.Photosynthetica 41, 167–175. doi:10.1023/B:PHOT.0000011948.00870.db
Ward JK, Tissue DT, Thomas RB, Strain BR (1999) Comparative responsesof model C3 and C4 plants to drought in low and elevated CO2. GlobalChange Biology 5, 857–867. doi:10.1046/j.1365-2486.1999.00270.x
Way DA, Oren R (2010) Differential responses to changes in growthtemperature between trees from different functional groups andbiomes: a review and synthesis of data. Tree Physiology 30, 669–688.doi:10.1093/treephys/tpq015
Way DA, Sage RF (2008) Thermal acclimation of photosynthesis in blackspruce (Picea mariana (Mill.) B.S.P.). Plant, Cell & Environment 31,1250–1262. doi:10.1111/j.1365-3040.2008.01842.x
Wheelwright NT, Logan BA (2004) Previous-year reproduction reducesphotosynthetic capacity and slows lifetime growth in females of aneotropical tree. Proceedings of the National Academy of Sciences ofthe United States of America 101, 8051–8055. doi:10.1073/pnas.0402735101
Wong SC, Cowan IR, Farquhar GD (1978) Leaf conductance in relationto assimilation in Eucalyptus pauciflora Sieb. Ex Spenge. Influenceof irradiance and partial pressure of carbon dioxide. Plant Physiology62, 670–674. doi:10.1104/pp.62.4.670
Wong SC, Kriedemann PE, Farquhar GD (1992) CO2 x nitrogen interactionon seedling growth of four species of Eucalypt. Australian Journal ofBotany 40, 457–472. doi:10.1071/BT9920457
Zieger E (1983) The biology of stomatal guard cells. Annual Review ofPlant Physiology 34, 441–474. doi:10.1146/annurev.pp.34.060183.002301
Zimmer HC, Auld TD, Benson J, Baker PJ (2014) Recruitment bottlenecksin the rare Australian conifer Wollemia nobilis. Biodiversity andConservation 23, 203–215. doi:10.1007/s10531-013-0593-2
Zonneveld BJM (2012) Genome sizes of all 19 Araucaria species arecorrelated with their geographical distribution. Plant Systematics andEvolution 298, 1249–1255. doi:10.1007/s00606-012-0631-7
Wollemi pine: back to the future Functional Plant Biology O
www.publish.csiro.au/journals/fpb