rnaiii of the staphylococcus aureus agr system … of the staphylococcus aureus agr system activates...
TRANSCRIPT

Correction
MICROBIOLOGYCorrection for “RNAIII of the Staphylococcus aureus agr systemactivates global regulator MgrA by stabilizing mRNA,” by RaviKr. Gupta, Thanh T. Luong, and Chia Y. Lee, which appearedin issue 45, November 10, 2015, of Proc Natl Acad Sci USA
(112:14036–14041; first published October 26, 2015; 10.1073/pnas.1509251112).The authors note that Fig. 2 appeared incorrectly. The cor-
rected figure and its legend appear below.
www.pnas.org/cgi/doi/10.1073/pnas.1523895113
B A
0123456
Hal
f-life
(min
)
5-nt
reve
rtant
(A22
)
Newm
an agr
(113
46)
5-nt
mut
ant
(129
16)
P1 U
TR
(184
4)P2
UTR
(1
845)
P2 U
TR agr
(125
02)
** * * * ** P1
UTR
agr
(125
01)
Hal
f-life
(min
)P1
UTR
+p
ML1
00
P2 U
TR
+pM
L100
P2 U
TR
+RNA
III
**
P1 U
TR
+RNA
III
0123456
Fig. 2. Stability of mgrA mRNA. (A) Stability in various chromosomal mutants. (B) RNAIII complementation of deletion mutations in mgrA UTR. mRNAstability expressed as half-life in minutes. The numbers in parentheses represent strain number. *P < 0.05, **P < 0.01 (unpaired two-tailed Student t testbetween Newman and each mutant, n = 3).
E7306 | PNAS | December 29, 2015 | vol. 112 | no. 52 www.pnas.org

RNAIII of the Staphylococcus aureus agr systemactivates global regulator MgrA by stabilizing mRNARavi Kr. Gupta, Thanh T. Luong, and Chia Y. Lee1
Department of Microbiology and Immunology, University of Arkansas for Medical Sciences, Little Rock, AR 72205
Edited by Richard P. Novick, New York University School of Medicine, New York, NY, and approved September 9, 2015 (received for review May 12, 2015)
RNAIII, the effector of the agr quorum-sensing system, plays a keyrole in virulence gene regulation in Staphylococcus aureus, but howRNAIII transcriptionally regulates its downstream genes is not com-pletely understood. Here, we show that RNAIII stabilizesmgrAmRNA,thereby increasing the production of MgrA, a global transcriptionalregulator that affects the expression of many genes. The mgrA geneis transcribed from two promoters, P1 and P2, to produce two mRNAtranscripts with long 5′ UTR. Two adjacent regions of themgrAmRNAUTR transcribed from the upstream P2 promoter, but not the P1 pro-moter, form a stable complex with two regions of RNAIII near the5′ and 3′ ends. We further demonstrate that the interaction has sev-eral biological effects. We propose that MgrA can serve as an inter-mediary regulator through which agr exerts its regulatory function.
Staphylococcus aureus | RNAIII | MgrA | virulence regulation
Small regulatory RNAs (sRNAs) play an important role in generegulation in both eukaryotes and prokaryotes. In bacteria,
sRNAs typically act by base pairing with target mRNAs with lim-ited or extended complementarity (antisense mechanism), but theycan also modulate protein activity (1, 2). sRNAs commonly act ontheir target mRNA near the translation initiation site by basepairing to block—and in some cases promote—translation. SomesRNAs cause alterations in the secondary structure of the targetmRNA, thereby enhancing or reducing mRNA degradation (3, 4).Staphylococcus aureus is an important human pathogen that
produces many virulence factors that are regulated by an equallyimpressive number of regulators (5, 6). A key regulatory systeminvolved in virulence regulation in S. aureus is the agr quorum-sensing system. A 514-nt RNA, RNAIII, is the effector throughwhich most genes in the agr regulon are regulated (7, 8). RNAIII isone of the largest known sRNAs that regulates gene expression viaantisense mechanism by base pairing with target mRNAs (5, 9).RNAIII is capable of forming a stable structure that is characterizedby 14 stem-loop motifs and three long-distance interacting helices(9). Several examples of regulation by direct interaction betweenRNAIII and its target mRNAs have been reported (10–14). Acommon denominator in these regulatory interactions is the tar-geting of translation initiation sequences, through base pairing ofthe CU-rich loop domains of RNAIII with regions containing theShine and Dalgarno (SD) sequence or AUG start codon of thetarget mRNA, resulting in negative control of translation (15, 16).A notable exception is the activation of Hla by RNAIII, in whichthe interaction unmasks the SD sequence to facilitate translation ofhla mRNA (10). Although a recent report shows that RNAIIIactivates map expression, the mechanism of activation is un-known (17). More recently, RNAIII has also been shown to neg-atively control sbi translation through direct interaction with thetranslation initiation sequences of the sbi mRNA. However, in thiscase, three distant RNAIII domains that do not include the CU-rich loop domains have been shown to directly interact with threedistinct sites in the sbi mRNA to repress sbi translation (18).Rot is a pleiotropic regulator that affects many toxins and sur-
face proteins (19). The finding that RNAIII represses rot mRNAtranslation has advanced the understanding of agr regulation(12, 13). However, RNAIII and Rot transcriptomes only partiallyoverlap (19, 20). Thus, RNAIII repression of Rot cannot com-pletely account for how RNAIII regulates the majority of itsdownstream genes. This led Geisinger et al. (12) and Boisset et al.
(13) to postulate that RNAIII must interact with one or morepleiotropic regulators. Here, we report that RNAIII can also affectits downstream genes through another important S. aureus globaltranscriptional regulator, MgrA, which has been shown to affectmore than 350 genes involved in virulence, antibiotic resistance,autolysis, and biofilm formation (21–25). Our results suggest thatpart of the MgrA regulon is affected by RNAIII.
ResultsmgrA Gene Is Transcribed by Two Promoters. We have previouslyidentified the mgrA gene (22). To map the mgrA promoter, weperformed primer extension experiments. We found three tran-scriptional start sites: at nucleotides −123, −301, and −303 up-stream from the ATG start codon (Fig. S1), which closely agreewith the nucleotide −124 and −302 sites previously reported (21).Most likely, nucleotides −301 and −303 represent start sites fromthe same promoter with the former being the preferred start site.We then designated the promoters as P1 and P2, as shown in Fig.1. To further define the promoters, we constructed promoter fu-sions to the xylE reporter gene in plasmid pLL38 with successivedeletions of DNA extending from 368 bp upstream of the mgrAATG start codon. Our results showed that deletion of P2 reducedreporter activity by about 33–43%, whereas deletion of both pro-moters resulted in about 92% reduction (Fig. 1), indicating thatboth promoters were functional and that the predicted promotersare most likely correct. It should be noted here that the P2 pro-moter is embedded within the coding region of a divergentlytranscribed upstream gene encoding a putative ABC transporter.
P2 5′ UTR Affects the Stability of mgrA mRNA. The 123-nt and 301/303-nt UTRs transcribed from the P1 and P2 promoters, re-spectively, are unusually long for bacterial mRNAs. For conve-nience, herein we refer to the region between the P2 and P1transcriptional start sites as the P2 UTR and the region between
Significance
In Staphylococcus aureus, the agr quorum-sensing system iscritical for regulation of many virulence genes, primarily throughRNAIII, a small RNA. RNAIII functions by interacting with trans-lational elements of its downstream genes typically by inhibitingtranslation initiation. Only a few direct targets of RNAIII havebeen identified, one of which is global regulator Rot. However,Rot can only partially account for RNAIII regulation. We showhere that RNAIII can affect gene expression through anotherglobal regulator MgrA by enhancing MgrA production via mRNAstabilization, a novel mechanism of RNAIII regulation. Our find-ings provide evidence linking two major master regulators,thereby advancing our understanding of virulence regulation inthis important human and animal pathogen.
Author contributions: R.K.G. and C.Y.L. designed research; R.K.G. and T.T.L. performedresearch; R.K.G., T.T.L., and C.Y.L. analyzed data; and R.K.G. and C.Y.L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1509251112/-/DCSupplemental.
14036–14041 | PNAS | November 10, 2015 | vol. 112 | no. 45 www.pnas.org/cgi/doi/10.1073/pnas.1509251112

the P1 transcriptional start site and the AUG start codon as the P1UTR. To determine whether sequences within the UTR affect theexpression of the mgrA gene, we performed deletion analysis. Asshown in Fig. 1, a deletion of 95 bp within the P1 UTR reduced thereporter activity by about 33%, whereas a deletion of 138 bp withinthe P2 UTR reduced by about 47%, suggesting that the sequenceswithin the UTRs are important for mgrA transcription.One mechanism whereby the 5′ UTR could affect gene tran-
scription is by influencing mRNA stability. To determine whetherthis mechanism is involved, we first constructed a 35-bp deletion inthe P1 UTR (ΔP1 UTR) and a 138-bp deletion in the P2 UTR (ΔP2UTR) in the chromosome. The mutants were tested for mgrAmRNA stability by real-time RT-PCR (qRT-PCR) upon rifampicin-mediated transcription arrest. We found that the ΔP2 UTR muta-tion significantly reduced themgrAmRNA stability, whereas theΔP1UTRmutation had no significant effect on mRNA stability (Fig. 2A).
RNAIII Stabilizes mgrA mRNA by Interacting with mgrA P2 UTR. Theabove results suggest that sequences in the P2 UTR are involvedin stabilization of mgrA mRNA. To investigate the mechanism,we searched for known S. aureus sRNAs using IntaRNA (26) andidentified RNAIII as a potential sRNA that could interact withthe UTR of mgrA mRNA by base paring at two regions (Fig. S2).Both the 33-nt and 14-nt regions in the mgrA transcript are lo-cated within the P2 UTR, whereas the two corresponding com-plementary regions in RNAIII are located at the ends of themolecule (the 5′ end 28 nt and the 3′ end 14 nt). To test whetherRNAIII is involved in stabilizing mgrA mRNA by interactingwith P2 UTR, we first compared the effect of agr deletion onmgrA mRNA stability in the wild-type, ΔP1 UTR, and ΔP2 UTRstrains. The results showed that the deletion of agr (CYL11346)decreased the half-life of mgrA mRNA (Fig. 2A). Furthermore, thedeletion of agr in the ΔP1 UTR strain reduced stability to about thesame level as that of the agr deletion alone (compare 11346 to12501) but had no effect on mRNA stability in the ΔP2 UTR strain(compare 1845 to 12502). These results suggest that agr has an effecton mgrA mRNA stability by targeting the P2 UTR. To furtherconfirm that RNAIII of the agr system is involved in the regulation,we expressed RNAIII under the control of an anhydrotetracycline(aTc)-inducible promoter. As shown in Fig. 2B, the induction ofRNAIII increased (though not statistically significant) the mgrAmRNA stability in the ΔP1 UTR mutant in which the P2 UTR isintact, whereas the stability in the ΔP2 UTR strain remained thesame with or without RNAIII induction. Because the stability testabove using qRT-PCR was unable to distinguish between P1- andP2-derived mgrA mRNA species, we further carried out Northernanalyses and confirmed that agr affected only the P2-derived mRNAspecies (Fig. S3A). In addition, to test the effect in a different geneticbackground, we showed that agr deletion has similar effect in strainUSA300 LAC (JE2), a strain with naturally high RNAIII level (Fig.S3B). These results further suggest that RNAIII is involved in sta-bilizing mgrA mRNA by targeting P2 UTR.As shown in Fig. S1, the mgrA P2 promoter is located within the
divergent NWMN_0656 coding sequence. Overlapping transcriptshave been shown to be degraded by RNase III, thereby affecting
mRNA level of the genes involved (27). Thus, it is possible thatRNAIII could affect NWMN_0656 expression, thereby affectingmgrAmRNA levels. However, we showed that RNAIII had no effecton NWMN_0656 transcription (Fig. S4), indicating that RNAIIIdoes not regulate mgrA mRNA levels through NWMN_0656.
mgrA UTR Interacts with RNAIII at Two Separate Domains. To determinewhether the complementary regions identified above are involved inthe interaction between RNAIII and mgrA P2 UTR, we used RNA-RNA EMSA. RNA corresponding to the mgrA P2 UTR was syn-thesized in vitro and labeled with biotin. Various regions of theRNAIII were also synthesized in vitro. Incubation of the labeledmgrA P2 UTR probe with full-length RNAIII resulted in a shiftedband (Fig. 3A), which could be effectively competed away with 10-to 30-fold excess of cold fragment (lanes 2 and 3 from the right inFig. 3A). An irrelevant RNA fragment derived from the 5′-end ofclpC (negative control) did not result in a band shift. By using partialRNAIII fragments, we showed that deletions of the segmentscontaining either one of the complementary regions in RNAIII stillresulted in a shifted band, but deletions of both regions abolishedthe shift (Fig. 3A and Fig. S5). These results suggest that both the28-nt and 14-nt regions of RNAIII contribute to the interaction ofRNAIII with mgrA P2 UTR and that the interaction is specific.To further confirm the interaction between RNAIII and mgrA
mRNA using EMSA, we labeled RNAIII. As shown in Fig. 3B,fragments containing both the 33-nt and the 14-nt regions ofmgrA UTR resulted in a shifted band. However, deletions of the14-nt region, but not the 33-nt region, abolished the shift. Theseresults did not totally conform to those of Fig. 3A and Fig. S5, inwhich both the 5′ and the 3′ domains of RNAIII contributed tothe binding. However, close examination of the shifted bandsrevealed a much-weaker band intensity when the 14-nt regionwas deleted compared with that of the deletion of the 28-nt re-gion (Fig. 3A and Fig. S5). Thus, based on these results, wespeculate that the perfectly complementary 14-nt domain mayplay a more important role than the 28-nt/33-nt domain in theinteraction between RNAIII and mgrA mRNA.
RNA SHAPE-Directed Chemical Probing of Interaction of mgrA 5′ UTRwith RNAIII.We then used the RNA SHAPE (selective 2′-hydroxylacylation analyzed by primer extension) method (28) to analyzethe mgrA UTR structure, with or without the presence of RNAIII.N-methylisatoic anhydride (NMIA) was used to form a 2′-O-adduct,with flexible nucleotides representing unpaired sequences, whichwere then detected by primer extension. The in vitro synthesized303-nt mgrA UTR with adaptor sequences at both 5′ and 3′ endswas used as the template for the reaction and primer extension. Theraw data were analyzed by QuShape software (28–30). This ex-perimental design resulted in highly reproducible SHAPE reactivitydata of themgrAUTR (Fig. 4A). The reactivity data were then usedto predict the structure of mgrA UTR (Fig. 4B) by using RNAs-tructure and XRNA software programs (31, 32). The proposedstructure indicates some of the nucleotides in the 14-nt (nucleotides
P2 100bp mgrA
P1-35
pTL3221 8.3±2.6 pTL3236 67.7±4.9 pTL3235 65.8±5.0 pTL3219 57.8±2.6 Δ59bp
Δ138bp
pTL3245 66.8±7.8
pTL3271 53.4±10.2
+82 -368
Δ103bp
Δ216bp Δ152bp
Δ95bp
Plasmid Promoter
activity (%)
pTL3289 100±22.3 -492
pTL3234 100±16.3 -10-35 -10
Fig. 1. Analysis of mgrA promoter region. DNA fragments with various de-letions, as indicated, were fused to the xylE reporter in pLL38. All deletionswere derived from pTL3234 with a 450-bp insert containing both promotersand 82 bp of the mgrA coding sequence, except pTL3271, which was derivedfrom pTL3289 with an additional 124 bp of upstream DNA (dashed line; not toscale). XylE activities were assayed in Newman. The promoter activities areexpressed as percentages of the activity of the parent plasmid.
A B
0 1 2 3 4 5 6
Hal
f-life
(min
)
5-nt
reve
rtant
(A22
)
Newm
an
Δagr
(1
1346
)
5-nt
mut
ant
(129
16)
ΔP1
UTR
(184
4)ΔP
2 UT
R (1
845)
ΔP2
UTR Δa
gr
(125
02)
** * * * **
ΔP1
UTR Δa
gr
(125
01)
Hal
f-life
(min
)ΔP
1 UT
R +p
ML1
00
ΔP2
UTR
+pM
L100
ΔP2
UTR
+RNA
III
**
ΔP1
UTR
+RNA
III
0 1 2 3 4 5 6
Fig. 2. Stability ofmgrAmRNA. (A) Stability in various chromosomal mutants.(B) RNAIII complementation of deletion mutations in mgrA UTR. mRNA sta-bility expressed as half-life in minutes. The numbers in parentheses representstrain number. *P < 0.05, **P < 0.01 (unpaired two-tailed Student t test be-tween Newman and each mutant, n = 3).
Gupta et al. PNAS | November 10, 2015 | vol. 112 | no. 45 | 14037
MICRO
BIOLO
GY

96–109) and the 33-nt (nucleotides 57–89) regions are involvedin the formation of secondary structures. However, the majority ofthe nucleotides in these regions are moderately reactive (Fig. 4A),suggesting that the hybrids are not stable. These regions also be-came less accessible to NMIA upon RNAIII addition (Fig. 4A,Lower, and Fig. 4C), suggesting that RNAIII displaces the hybridsto form more stable duplexes at these regions. Besides the changesin reactivity at the RNAIII-interacting regions, we found little re-activity change in the rest of the mgrA UTR molecule except twoshort regions: one near the 5′-end (nucleotides 23–27) and theother near the 3′-end (nucleotides 263–278). The former is near theregions that interact with RNAIII suggesting that the interactionwith RNAIII induces some changes in secondary structure atnearby sequences. The latter includes the region that is predictedto form a long-distance hybrid with the 33-nt domain within themgrA UTR (Fig. 4B) and, therefore, the increase of reactivity at
this region is consistent with the notion that these sequences aredisplaced upon RNAIII interaction.Similarly, we analyzed the RNAIII structure by RNA SHAPE in
the presence or absence of mgrA UTR (Fig. S6A). In the presenceof mgrA UTR, we found the 14-nt (nucleotides 447–460) and the28-nt (nucleotides 8–35) domains of RNAIII, which were predictedto interact with mgrA, became less accessible to NMIA. Few or nochanges were found in the rest of the RNAIII sequences. We alsomodeled the RNAIII structure based on the reactivity data (Fig.S6B), which matches well with the structure of RNAIII predictedusing chemical and enzymatic probing (9). The high similarity be-tween our predicted RNAIII structure and the previously estab-lished structure indirectly validates our RNA SHAPE results. Theseresults from SHAPE analyses are in line with our model that mgrAUTR interacts with RNAIII at the 14-nt and 33-nt/28-nt domains,which further corroborate the EMSA study described above.
RNA Walk Reveals in Vivo Interaction of RNAIII and mgrA UTR. Toinvestigate the interaction of RNAIII and mgrA mRNA in vivo, wemodified the RNA Walk method that had been used to studyRNA–RNA interactions in trypanosomes (33). The method usesAMT (4′-aminomethyl-trioxsalen hydrochloride) to induce cross-linking under UV irradiation, which links two RNA molecules inclose proximity within the cell. The cross-linked molecules werethen mapped by RT-PCR, in which the primers spanning theregion of an interacting domain cannot be copied efficientlybecause of cross-linking. S. aureus Newman, CYL1844 (ΔP1UTR), CYL1845 (ΔP2 UTR), and CYL11346 (Δagrmutant) weretreated with AMT and UV-irradiated. Total RNA from the strains,with or without treatment, were isolated, converted to cDNA, andthen PCR-amplified using primers spanning the P2 UTR con-taining the 14-nt region (the 5′ primer contains a portion of the 14-ntregion sequence for specific amplification of the interacting region)(Table S1). Amplification of the hu gene was included as a control.As shown in Fig. 5A, a 170-bp PCR fragment could be detected instrains with an intact P2 UTR and an intact agr locus when thecultures were not treated. However, no PCR product was detected intreated cultures (Fig. 5A, lanes 1–4). On the other hand, the 170-bpPCR fragment was detected in the agr-deleted strain but not in theΔP2 UTR strain, with or without AMT and UV (Fig. 5A, lanes 5–8).In addition, treatment did not affect amplification of the hu gene inany strain. These results demonstrated that the presence of both agr
B
A
Fig. 3. RNA EMSA. (A) Labeled mgrA P2 UTR reacted with various deletionsof 514-nt RNAIII. clpC RNA was used as a negative control. (B) Labeled RNAIIIreacted with various deletions of mgrA UTR RNA fragments A through F. Redand green boxes represent matched sequences in each RNA species. Trianglesindicate transcriptional start sites.
0 1 2 3
1 11 21 31 41 51 61 71 81 91 101 111 121 131 141 151 161 171 181 191 201 211 221 231 241 251 261 271 281 291 301
33nt 14nt
mgrA UTR
0 1 2 3
1 11 21 31 41 51 61 71 81 91 101 111 121 131 141 151 161 171 181 191 201 211 221 231 241 251 261 271 281 291 301
33nt 14nt
A
mgrA UTR + RNAIII
B
AUAGG
UAUUUAU ACGGAU
A
GUCUG U
G
A U
GA
UUUUCUCUU
AAAAGAGAA
AUAU
UAUA
GAU
CUA
GAUAAC
UUAUUG
CGC
AAUCA UUAGU
CA
GAAAGUUG UUUUCAAC
AA
UU
AA
UUUUGGGCAAUGA
AAAAUCCGUUAUU
A
U
14 nt
33 nt
AUUG
UAAC
10
20
30
40
50
60
70
80
90
100
110
130
140
150
160
170 180
190
210
220 230 24
0
250
260
280
290
300
U
C
Fig. 4. SHAPE analysis of mgrA UTR. (A) Average reactivity of each nucleotide of mgrA UTR in the absence (Upper) or presence (Lower) of RNAIII. Red, highlyreactive (>0.80 unit); orange, moderately reactive (0.40–0.80 unit); black, unreactive (<0.40 unit). n = 2. (B) Predicted structure of mgrA UTR RNA. The sequences ofthe 14-nt domain are in red and the 33-nt domain in green. Solid and open arrowheads indicate increase and decrease, respectively, in nucleotide reactivity in thepresence of RNAIII. (C) Comparison of reactivity of each nucleotide in the RNAIII-binding domains inmgrA P2 UTR. Red trace, with RNAIII; Blue trace, without RNAIII.
14038 | www.pnas.org/cgi/doi/10.1073/pnas.1509251112 Gupta et al.

RNAIII and the P2 UTR was required to form a cross-link adduct,suggesting their specific interaction in vivo.
Mutation in the 14-nt Site Confirms Interaction of RNAIII and mgrAUTR. To further demonstrate that the RNAIII interacts with mgrAmRNA, we constructed a 5-bp substitution mutation within the14-bp domain, in the Newman chromosome. These 5 bp werechosen because of their high GC content and relative central lo-cation in the 14-nt domain (Fig. S2C). The mutation did not affectbacterial growth (Fig. S7). We then assessed whether the mutationaffected mgrA mRNA stability. As shown in Fig. 2, the 5-bp mu-tation (CYL12916) resulted in a significant reduction in mRNAstability, similar to that of the agr deletion. These results suggestthat the 14-nt domain in the P2 UTR is critical for mgrA mRNAstability. Because the 5-bp mutation within the 14-bp domain can-not be complemented with a plasmid, we sought to revert themutation to the wild-type sequence to ensure that the mutantphenotype was not a result of inadvertent secondary mutationsduring the construction of the mutant. The mgrA mRNA stabilityof the resultant revertant (CYLA22) was similar to that of theNewman (Fig. 2A), indicating that the mutant phenotype was aresult of the 5-bp mutation.As the 14-nt domain affects stability of mRNA transcribed from
the P2 promoter, but not from the P1 promoter, we expected thatMgrA production would be partially affected by the mutation inthe 14-nt domain or the agr regulatory system under our ex-perimental conditions. By Western analyses, we found that theamounts of MgrA were similar between the 5-bp mutationmutant and the Δagr mutant, which were both about 60% ofthe wild-type (Fig. 5B), suggesting that agr affects mgrA expressionthrough the 14-nt domain in the P2 UTR. As expected, the revertantproduced a similar amount of MgrA as Newman and the ΔmgrAmutant produced, essentially, an undetectable amount of protein.To directly confirm the interaction, we constructed a 5-nt mu-
tation in RNAIII such that the 14-nt region of the RNAIII iscomplementary to the 5-bp mutation in the 14-nt region of mgrAmRNA in strain CYL12916 described above. The 5-bp mutantRNAIII fragment was cloned in pML100 under an aTc-induciblepromoter (pRGE23). As shown in Fig. 5B, the mutation in RNAIIIrestored the MgrA protein production in CYLA15 (CYL12916Δagr) to a similar level produced in Newman. These results stronglysuggest that RNAIII directly interacts with mgrA mRNA at the14-nt domain to stabilize the mgrA mRNA stability.
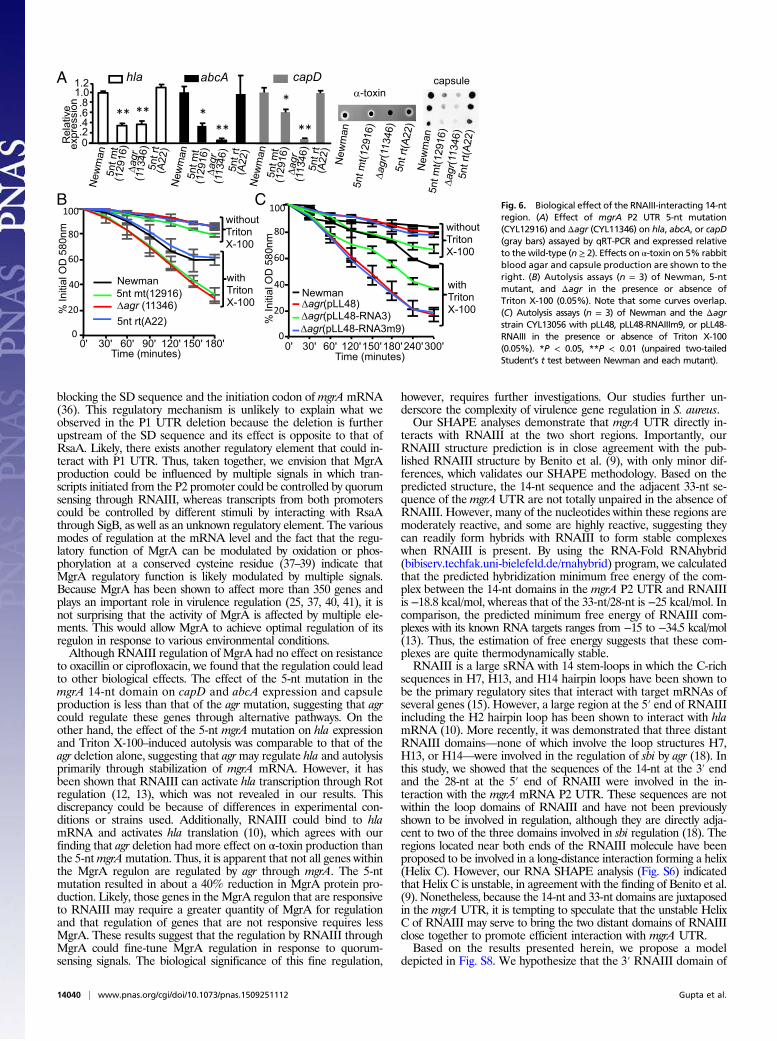
Phenotypic Effects of a 14-nt Domain Mutation in mgrA P2 UTR. MgrAhas been shown to affect capsule, surface proteins, toxins, autolysis,and antibiotic resistance (21–24, 34). To determine whetherRNAIII–mgrA mRNA interaction has an effect on biological
function, we tested a phenotype of CYL12916 containing the 5-ntmutation in the mgrA UTR. We found that hla, which encodesα-toxin, was affected by the 5-nt mutation to the same degree asthe agr deletion (Fig. 6A), suggesting that agr activates hla primarilythrough mgrA in Newman. We also found that abcA, which en-codes an ABC transporter involved in antibiotic resistance (34),and capD, which is one of the capsule genes in the cap operon,were significantly affected by the 5-nt mutation but not as drasti-cally as the agr deletion (Fig. 6A), suggesting that agr affects thesegenes, in part, through interaction with the mgrA mRNA. Ex-pression of all these genes was restored to the wild-type level in the5-nt revertant strain. In addition, we also tested the effect of the5-nt mutation on hemolysin activity and capsule production. Asshown in Fig. 6A, mutation in agr reduced the hemolysin activity toan undetectable level, whereas the 5-nt mutation in the mgrA P2UTR had a moderate reduction, which is consistent with a previousstudy that agr could affect hla at the translational level (10). On theother hand, the effect of the mutation on capsule production wascomparable to the transcriptional effect measured by qRT-PCR.MgrA has been shown to inhibit cell lysis (24). As shown in Fig.
6B, we found increased Triton X-100–induced cell autolysis inCYL12916 compared with Newman. Deletion of agr (CYL11346)increased the autolysis to the same extent as the 5-bp mutation,suggesting that agr could affect autolysis through the 14-nt element.The revertant of the 5-bp mutation had a rate of lysis similar toNewman. To test this further, we complemented an agr deletionmutant (CYL13056) with pLL48 expressing either intact RNAIII(pLL48-RNAIII) or mutated RNAIII (pLL48-RNAIIIm9, whichcontains a substitution of 9-nt in the 14-nt domain in RNAIII). Ourresults (Fig. 6C) showed that pLL48-RNAIIIm9 could not com-plement the agr deletion, whereas pLL48-RNAIII complementedthe deletion although not to the level of Newman. These resultssuggest that RNAIII affects autolysis by stabilizing mgrA mRNAthrough the 14-nt domain.The above results demonstrated that the stabilization of mgrA
mRNA by RNAIII could lead to significant biological effects.However, we found that the minimum inhibitory concentration(MIC) of oxacillin or ciprofloxacin was not affected by the 5-ntmutation, although others have reported thatmgrA mutants havereduced MIC to ciprofloxacin (23) and that overexpression ofmgrA increased the MIC to oxacillin in strain N315 (35). Thus,not all phenotypic changes associated with MgrA regulation areaffected by RNAIII.
DiscussionMicroarray studies showed that Agr and MgrA have similar effectson many virulence genes, suggesting that the two regulators maycoregulate a subset of target genes (25). Here we found that RNAIIIdirectly interacts with the mgrA mRNA. Our results suggest thatRNAIII stabilizes the mgrA mRNA primarily through base pairingbetween the two RNA molecules at two domains. We provide sev-eral lines of evidence supporting this conclusion. RNAIII, as a reg-ulatory effector of the agr system, has so far been shown to affect itstarget genes at the translational level by RNA base pairing that ei-ther masks or unmasks the translation initiation site (10, 11, 13, 14).To our knowledge, the findings presented here are the first to showthat S. aureus RNAIII affects a target gene by mRNA stabilization.The agr system regulates its targets mostly through an RNAIII-
dependent pathway, but direct transcriptional activation of the psmgenes by AgrA, the response regulator of the agr system, has beenreported (8). Among the target genes, rot is the only regulatory genefound to be directly targeted by RNAIII. As such, Rot serves as anintermediary regulator of RNAIII (12, 13). In this study, our findingsstrongly suggest that MgrA is another intermediary regulator ofRNAIII. However, we found that RNAIII only affected the tran-script initiated from the P2 promoter, indicating that it has onlypartial control on mgrA expression. Interestingly, in addition tothe P2 UTR, expression of mgrA is also affected by the sequenceswithin the P1 UTR, suggesting a different regulatory factor (orfactors) is involved (Fig. 1). A recent report has shown that RsaA,a SigB-dependent sRNA, represses translation of MgrA by
A B
Fig. 5. Interaction of RNAIII and mgrA UTR in vivo. (A) RNA Walk. Cross-linkedRNAs were amplified by RT-PCR with primers specific for the 14-nt region inmgrAUTR. Cross-linked adducts prevents amplification. hu primers were used as positivecontrols. (B) MgrAWestern blots were quantified by densitometry, normalized toa Coomassie blue-stained protein band (arrow), and expressed relative to New-man (Nm). n = 3. CYL12916 carries the 5-ntmutation inmgrAUTR and A22 carriesthe corresponding reversion. MgrA complementation of the 5-nt mutation inmgrA UTR (Right) was performed in A15 (CYL12916 Δagr) using pML100 carryingeither the wild-type RNAIII or RNAIII with a 5-nt mutation (RNAIIIm5) that iscomplementary to the five mutated nucleotides in CYL12916. *P < 0.05, **P <0.01 (unpaired two-tailed Student’s t test between Newman and each mutant).
Gupta et al. PNAS | November 10, 2015 | vol. 112 | no. 45 | 14039
MICRO
BIOLO
GY
blocking the SD sequence and the initiation codon ofmgrAmRNA(36). This regulatory mechanism is unlikely to explain what weobserved in the P1 UTR deletion because the deletion is furtherupstream of the SD sequence and its effect is opposite to that ofRsaA. Likely, there exists another regulatory element that could in-teract with P1 UTR. Thus, taken together, we envision that MgrAproduction could be influenced by multiple signals in which tran-scripts initiated from the P2 promoter could be controlled by quorumsensing through RNAIII, whereas transcripts from both promoterscould be controlled by different stimuli by interacting with RsaAthrough SigB, as well as an unknown regulatory element. The variousmodes of regulation at the mRNA level and the fact that the regu-latory function of MgrA can be modulated by oxidation or phos-phorylation at a conserved cysteine residue (37–39) indicate thatMgrA regulatory function is likely modulated by multiple signals.Because MgrA has been shown to affect more than 350 genes andplays an important role in virulence regulation (25, 37, 40, 41), it isnot surprising that the activity of MgrA is affected by multiple ele-ments. This would allow MgrA to achieve optimal regulation of itsregulon in response to various environmental conditions.Although RNAIII regulation of MgrA had no effect on resistance
to oxacillin or ciprofloxacin, we found that the regulation could leadto other biological effects. The effect of the 5-nt mutation in themgrA 14-nt domain on capD and abcA expression and capsuleproduction is less than that of the agr mutation, suggesting that agrcould regulate these genes through alternative pathways. On theother hand, the effect of the 5-nt mgrA mutation on hla expressionand Triton X-100–induced autolysis was comparable to that of theagr deletion alone, suggesting that agrmay regulate hla and autolysisprimarily through stabilization of mgrA mRNA. However, it hasbeen shown that RNAIII can activate hla transcription through Rotregulation (12, 13), which was not revealed in our results. Thisdiscrepancy could be because of differences in experimental con-ditions or strains used. Additionally, RNAIII could bind to hlamRNA and activates hla translation (10), which agrees with ourfinding that agr deletion had more effect on α-toxin production thanthe 5-ntmgrAmutation. Thus, it is apparent that not all genes withinthe MgrA regulon are regulated by agr through mgrA. The 5-ntmutation resulted in about a 40% reduction in MgrA protein pro-duction. Likely, those genes in the MgrA regulon that are responsiveto RNAIII may require a greater quantity of MgrA for regulationand that regulation of genes that are not responsive requires lessMgrA. These results suggest that the regulation by RNAIII throughMgrA could fine-tune MgrA regulation in response to quorum-sensing signals. The biological significance of this fine regulation,
however, requires further investigations. Our studies further un-derscore the complexity of virulence gene regulation in S. aureus.Our SHAPE analyses demonstrate that mgrA UTR directly in-
teracts with RNAIII at the two short regions. Importantly, ourRNAIII structure prediction is in close agreement with the pub-lished RNAIII structure by Benito et al. (9), with only minor dif-ferences, which validates our SHAPE methodology. Based on thepredicted structure, the 14-nt sequence and the adjacent 33-nt se-quence of themgrAUTR are not totally unpaired in the absence ofRNAIII. However, many of the nucleotides within these regions aremoderately reactive, and some are highly reactive, suggesting theycan readily form hybrids with RNAIII to form stable complexeswhen RNAIII is present. By using the RNA-Fold RNAhybrid(bibiserv.techfak.uni-bielefeld.de/rnahybrid) program, we calculatedthat the predicted hybridization minimum free energy of the com-plex between the 14-nt domains in the mgrA P2 UTR and RNAIIIis −18.8 kcal/mol, whereas that of the 33-nt/28-nt is −25 kcal/mol. Incomparison, the predicted minimum free energy of RNAIII com-plexes with its known RNA targets ranges from −15 to −34.5 kcal/mol(13). Thus, the estimation of free energy suggests that these com-plexes are quite thermodynamically stable.RNAIII is a large sRNA with 14 stem-loops in which the C-rich
sequences in H7, H13, and H14 hairpin loops have been shown tobe the primary regulatory sites that interact with target mRNAs ofseveral genes (15). However, a large region at the 5′ end of RNAIIIincluding the H2 hairpin loop has been shown to interact with hlamRNA (10). More recently, it was demonstrated that three distantRNAIII domains—none of which involve the loop structures H7,H13, or H14—were involved in the regulation of sbi by agr (18). Inthis study, we showed that the sequences of the 14-nt at the 3′ endand the 28-nt at the 5′ end of RNAIII were involved in the in-teraction with the mgrA mRNA P2 UTR. These sequences are notwithin the loop domains of RNAIII and have not been previouslyshown to be involved in regulation, although they are directly adja-cent to two of the three domains involved in sbi regulation (18). Theregions located near both ends of the RNAIII molecule have beenproposed to be involved in a long-distance interaction forming a helix(Helix C). However, our RNA SHAPE analysis (Fig. S6) indicatedthat Helix C is unstable, in agreement with the finding of Benito et al.(9). Nonetheless, because the 14-nt and 33-nt domains are juxtaposedin the mgrA UTR, it is tempting to speculate that the unstable HelixC of RNAIII may serve to bring the two distant domains of RNAIIIclose together to promote efficient interaction with mgrA UTR.Based on the results presented herein, we propose a model
depicted in Fig. S8. We hypothesize that the 3′ RNAIII domain of
C B
A
0' 30' 60' 120' 150' 180' 240' 300'
without Triton X-100
with Triton X-100
% In
itial
OD
580
nm
Time (minutes)
Newman Δagr(pLL48)
Δagr(pLL48-RNA3m9)Δagr(pLL48-RNA3)
0
20
40
60
80
100
0' 30' 60' 90' 120' 150' 180'
% In
itial
OD
580
nm
Newman
Δagr (11346)5nt mt(12916)
5nt rt(A22)
without Triton X-100
with Triton X-100
Time (minutes)
New
man
New
man
New
man
5nt m
t(129
16)
5nt m
t(129
16)
Δagr
(113
46)
5nt m
t(1
2916
)Δa
gr(1
1346
)
5nt m
t(1
2916
)Δa
gr(1
1346
)
5nt m
t(1
2916
)
Δagr
(113
46)
5nt r
t(A22
))))
0 .2.4.6.8
1.0
Rel
ativ
e e
xpre
ssio
nhla abcA capD1.2
5nt r
t(A
22)
5nt r
t(A
22)
5nt r
t(A
22)
α-toxin capsule
5nt r
t(A22
)
New
man
New
man
Δagr
(113
46)
Fig. 6. Biological effect of the RNAIII-interacting 14-ntregion. (A) Effect of mgrA P2 UTR 5-nt mutation(CYL12916) and Δagr (CYL11346) on hla, abcA, or capD(gray bars) assayed by qRT-PCR and expressed relativeto the wild-type (n ≥ 2). Effects on α-toxin on 5% rabbitblood agar and capsule production are shown to theright. (B) Autolysis assays (n = 3) of Newman, 5-ntmutant, and Δagr in the presence or absence ofTriton X-100 (0.05%). Note that some curves overlap.(C) Autolysis assays (n = 3) of Newman and the Δagrstrain CYL13056 with pLL48, pLL48-RNAIIIm9, or pLL48-RNAIII in the presence or absence of Triton X-100(0.05%). *P < 0.05, **P < 0.01 (unpaired two-tailedStudent’s t test between Newman and each mutant).
14040 | www.pnas.org/cgi/doi/10.1073/pnas.1509251112 Gupta et al.

14-nt first interacts with the 14-nt sequence of mgrA UTR by basepairing. After the initial interaction, the 28-nt 5′ RNAIII domainthen pairs with the mgrA UTR 33-nt sequence. In Newman, mgrAmRNA is expressed at the highest level in postexponential phase,at which time RNAIII is most abundant (7, 42), allowing forpairing to occur readily. The complex then stabilizes mgrA mRNA,likely by blocking degradation by one or more RNases, leading tomore MgrA protein production, thereby affecting expression of asubset of genes in the MgrA regulon.
Materials and MethodsStrain and Plasmid Construction. Strains and plasmids used in this study arelisted in Table S2. Plasmid and strain construction were carried out usingstandard molecular methods. For details, see SI Materials and Methods.Primers used in this study are listed in Table S1.
mRNA Stability Assay. RNAs from transcription-arrested cultures were isolatedat different time points. mRNA stability was assayed by qRT-PCR or Northernanalysis. Half-lives were calculated by using linear regression analysis. Fordetailed procedures, see SI Materials and Methods.
RNA EMSA. RNA EMSA reactions were performed using RNAs synthesized byin vitro transcription. For details, see SI Materials and Methods.
RNA Walk. RNA Walk was performed essentially as described previously(33). Cross-linking of paired RNAs in vivo was done by adding AMT tocultures and UV exposure. RT-PCR was performed to detect cross-linkedRNA regions. The detailed procedures are provided in the SI Materialsand Methods.
RNA SHAPE Reactions and Analysis. RNA SHAPE experiments were performedaccording to the previously described method using NMIA (Sigma) (43, 44).The detailed procedures are provided in the SI Materials and Methods.
ACKNOWLEDGMENTS. We thank David Cue and Mei Lei for critical reviewand helpful discussions; Mei Lei for construction of pML12570 and tech-nical help; and Wes Sanders and Allain Laederach (University of NorthCarolina) for help with SHAPE analyses. This work was supported byNational Institute of Allergy and Infectious Diseases Grants AI37027 andAI113766. The University of Arkansas for Medical Sciences core facilities aresupported in part by National Institutes of Health Grants P20GM103450 andP30GM103429.
1. Storz G, Vogel J, Wassarman KM (2011) Regulation by small RNAs in bacteria: Expandingfrontiers. Mol Cell 43(6):880–891.
2. Romby P, Vandenesch F, Wagner EG (2006) The role of RNAs in the regulation ofvirulence-gene expression. Curr Opin Microbiol 9(2):229–236.
3. Caron MP, Lafontaine DA, Massé E (2010) Small RNA-mediated regulation at the levelof transcript stability. RNA Biol 7(2):140–144.
4. Podkaminski D, Vogel J (2010) Small RNAs promote mRNA stability to activate thesynthesis of virulence factors. Mol Microbiol 78(6):1327–1331.
5. Novick RP (2003) Autoinduction and signal transduction in the regulation ofstaphylococcal virulence. Mol Microbiol 48(6):1429–1449.
6. Junecko J, et al. (2012) Transcribing virulence in Staphylococcus aureus. World J ClinInfect Dis 2(4):63–76.
7. Novick RP, et al. (1993) Synthesis of staphylococcal virulence factors is controlled by aregulatory RNA molecule. EMBO J 12(10):3967–3975.
8. Queck SY, et al. (2008) RNAIII-independent target gene control by the agr quorum-sensing system: Insight into the evolution of virulence regulation in Staphylococcusaureus. Mol Cell 32(1):150–158.
9. Benito Y, et al. (2000) Probing the structure of RNAIII, the Staphylococcus aureus agrregulatory RNA, and identification of the RNA domain involved in repression of protein Aexpression. RNA 6(5):668–679.
10. Morfeldt E, Taylor D, von Gabain A, Arvidson S (1995) Activation of alpha-toxin trans-lation in Staphylococcus aureus by the trans-encoded antisense RNA, RNAIII. EMBO J14(18):4569–4577.
11. Huntzinger E, et al. (2005) Staphylococcus aureus RNAIII and the endoribonuclease IIIcoordinately regulate spa gene expression. EMBO J 24(4):824–835.
12. Geisinger E, Adhikari RP, Jin R, Ross HF, Novick RP (2006) Inhibition of rot translationby RNAIII, a key feature of agr function. Mol Microbiol 61(4):1038–1048.
13. Boisset S, et al. (2007) Staphylococcus aureus RNAIII coordinately represses the syn-thesis of virulence factors and the transcription regulator Rot by an antisensemechanism. Genes Dev 21(11):1353–1366.
14. Chevalier C, et al. (2010) Staphylococcus aureus RNAIII binds to two distant regions of coamRNA to arrest translation and promote mRNA degradation. PLoS Pathog 6(3):e1000809.
15. Felden B, Vandenesch F, Bouloc P, Romby P (2011) The Staphylococcus aureus RNomeand its commitment to virulence. PLoS Pathog 7(3):e1002006.
16. Romilly C, et al. (2012) Current knowledge on regulatory RNAs and their machineriesin Staphylococcus aureus. RNA Biol 9(4):402–413.
17. Liu Y, et al. (2011) RNAIII activates map expression by forming an RNA-RNA complexin Staphylococcus aureus. FEBS Lett 585(6):899–905.
18. Chabelskaya S, Bordeau V, Felden B (2014) Dual RNA regulatory control of aStaphylococcus aureus virulence factor. Nucleic Acids Res 42(8):4847–4858.
19. Saïd-Salim B, et al. (2003) Global regulation of Staphylococcus aureus genes by Rot.J Bacteriol 185(2):610–619.
20. Dunman PM, et al. (2001) Transcription profiling-based identification of Staphylococcusaureus genes regulated by the agr and/or sarA loci. J Bacteriol 183(24):7341–7353.
21. Ingavale SS, Van Wamel W, Cheung AL (2003) Characterization of RAT, an autolysisregulator in Staphylococcus aureus. Mol Microbiol 48(6):1451–1466.
22. Luong TT, Newell SW, Lee CY (2003) Mgr, a novel global regulator in Staphylococcusaureus. J Bacteriol 185(13):3703–3710.
23. Truong-Bolduc QC, Zhang X, Hooper DC (2003) Characterization of NorR protein, amultifunctional regulator of norA expression in Staphylococcus aureus. J Bacteriol185(10):3127–3138.
24. Trotonda MP, Xiong YQ, Memmi G, Bayer AS, Cheung AL (2009) Role of mgrA andsarA in methicillin-resistant Staphylococcus aureus autolysis and resistance to cellwall-active antibiotics. J Infect Dis 199(2):209–218.
25. Luong TT, Dunman PM, Murphy E, Projan SJ, Lee CY (2006) Transcription Profiling ofthe mgrA Regulon in Staphylococcus aureus. J Bacteriol 188(5):1899–1910.
26. Busch A, Richter AS, Backofen R (2008) IntaRNA: Efficient prediction of bacterial sRNAtargets incorporating target site accessibility and seed regions. Bioinformatics 24(24):2849–2856.
27. Lasa I, et al. (2011) Genome-wide antisense transcription drives mRNA processing inbacteria. Proc Natl Acad Sci USA 108(50):20172–20177.
28. Mortimer SA, Weeks KM (2009) Time-resolved RNA SHAPE chemistry: QuantitativeRNA structure analysis in one-second snapshots and at single-nucleotide resolution.Nat Protoc 4(10):1413–1421.
29. Karabiber F, McGinnis JL, Favorov OV, Weeks KM (2013) QuShape: Rapid, accurate,and best-practices quantification of nucleic acid probing information, resolved bycapillary electrophoresis. RNA 19(1):63–73.
30. Low JT, Weeks KM (2010) SHAPE-directed RNA secondary structure prediction.Methods 52(2):150–158.
31. Reuter JS, Mathews DH (2010) RNAstructure: Software for RNA secondary structureprediction and analysis. BMC Bioinformatics 11:129.
32. Deigan KE, Li TW, Mathews DH, Weeks KM (2009) Accurate SHAPE-directed RNAstructure determination. Proc Natl Acad Sci USA 106(1):97–102.
33. Lustig Y, Wachtel C, Safro M, Liu L, Michaeli S (2010) ‘RNAWalk’ a novel approach to studyRNA-RNA interactions between a small RNA and its target. Nucleic Acids Res 38(1):e5.
34. Villet RA, et al. (2014) Regulation of expression of abcA and its response to envi-ronmental conditions. J Bacteriol 196(8):1532–1539.
35. Cui L, Lian JQ, Neoh HM, Reyes E, Hiramatsu K (2005) DNA microarray-based identi-fication of genes associated with glycopeptide resistance in Staphylococcus aureus.Antimicrob Agents Chemother 49(8):3404–3413.
36. Romilly C, et al. (2014) A non-coding RNA promotes bacterial persistence and decreasesvirulence by regulating a regulator in Staphylococcus aureus. PLoS Pathog 10(3):e1003979.
37. Chen PR, et al. (2006) An oxidation-sensing mechanism is used by the global regulatorMgrA in Staphylococcus aureus. Nat Chem Biol 2(11):591–595.
38. Truong-Bolduc QC, Hooper DC (2010) Phosphorylation of MgrA and its effect onexpression of the NorA and NorB efflux pumps of Staphylococcus aureus. J Bacteriol192(10):2525–2534.
39. Sun F, et al. (2012) Protein cysteine phosphorylation of SarA/MgrA family transcrip-tional regulators mediates bacterial virulence and antibiotic resistance. Proc NatlAcad Sci USA 109(38):15461–15466.
40. Jonsson IM, Lindholm C, Luong TT, Lee CY, Tarkowski A (2008) MgrA regulatesstaphylococcal virulence important for induction and progression of septic arthritisand sepsis. Microbes Infect 10(12-13):1229–1235.
41. Gupta RK, Alba J, Xiong YQ, Bayer AS, Lee CY (2013) MgrA activates expression ofcapsule genes, but not the α-toxin gene in experimental Staphylococcus aureus en-docarditis. J Infect Dis 208(11):1841–1848.
42. Ballal A, Manna AC (2009) Expression of the sarA family of genes in different strainsof Staphylococcus aureus. Microbiology 155(Pt 7):2342–2352.
43. Wilkinson KA, Merino EJ, Weeks KM (2006) Selective 2′-hydroxyl acylation analyzedby primer extension (SHAPE): Quantitative RNA structure analysis at single nucleotideresolution. Nat Protoc 1(3):1610–1616.
44. Mortimer SA, Doudna JA (2013) Unconventional miR-122 binding stabilizes the HCVgenome by forming a trimolecular RNA structure. Nucleic Acids Res 41(7):4230–4240.
45. Sau S, Sun J, Lee CY (1997) Molecular characterization and transcriptional analysis oftype 8 capsule genes in Staphylococcus aureus. J Bacteriol 179(5):1614–1621.
46. Bose JL, Fey PD, Bayles KW (2013) Genetic tools to enhance the study of gene functionand regulation in Staphylococcus aureus. Appl Environ Microbiol 79(7):2218–2224.
47. Chen Z, Luong TT, Lee CY (2007) The sbcDC locus mediates repression of type 5capsule production as part of the SOS response in Staphylococcus aureus. J Bacteriol189(20):7343–7350.
48. Lei MG, Cue D, Roux CM, Dunman PM, Lee CY (2011) Rsp inhibits attachment andbiofilm formation by repressing fnbA in Staphylococcus aureus MW2. J Bacteriol193(19):5231–5241.
49. Luong TT, et al. (2011) Staphylococcus aureus ClpC divergently regulates capsule viasae and codY in strain newman but activates capsule via codY in strain UAMS-1 and instrain Newman with repaired saeS. J Bacteriol 193(3):686–694.
Gupta et al. PNAS | November 10, 2015 | vol. 112 | no. 45 | 14041
MICRO
BIOLO
GY