role of acid ph and deficient efflux of pyrazinoic acid in ... · pzase into pyrazinoic acid...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY,0021-9193/99/$04.0010
Apr. 1999, p. 2044–2049 Vol. 181, No. 7
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Role of Acid pH and Deficient Efflux of Pyrazinoic Acid in UniqueSusceptibility of Mycobacterium tuberculosis to Pyrazinamide
YING ZHANG,1* ANGELO SCORPIO,1† HIROSHI NIKAIDO,2 AND ZHONGHE SUN1
Department of Molecular Microbiology and Immunology, School of Hygiene and Public Health,Johns Hopkins University, Baltimore, Maryland 21205,1 and Department of Molecular
and Cell Biology, University of California, Berkeley, California 947202
Received 25 November 1998/Accepted 29 January 1999
Pyrazinamide (PZA) is an important antituberculosis drug. Unlike most antibacterial agents, PZA, despiteits remarkable in vivo activity, has no activity against Mycobacterium tuberculosis in vitro except at an acidic pH.M. tuberculosis is uniquely susceptible to PZA, but other mycobacteria as well as nonmycobacteria are intrinsic-ally resistant. The role of acidic pH in PZA action and the basis for the unique PZA susceptibility of M. tuber-culosis are unknown. We found that in M. tuberculosis, acidic pH enhanced the intracellular accumulation ofpyrazinoic acid (POA), the active derivative of PZA, after conversion of PZA by pyrazinamidase. In contrast,at neutral or alkaline pH, POA was mainly found outside M. tuberculosis cells. PZA-resistant M. tuberculosiscomplex organisms did not convert PZA into POA. Unlike M. tuberculosis, intrinsically PZA-resistant M. smeg-matis converted PZA into POA, but it did not accumulate POA even at an acidic pH, due to a very active POAefflux mechanism. We propose that a deficient POA efflux mechanism underlies the unique susceptibility ofM. tuberculosis to PZA and that the natural PZA resistance of M. smegmatis is due to a highly active efflux pump.These findings may have implications with regard to the design of new antimycobacterial drugs.
The antituberculosis drug pyrazinamide (PZA) is an ana-logue of nicotinamide, which is a vitamin B3 (nicotinic acid;also called niacin) precursor. The discovery of PZA in the late1940s as a powerful drug against tuberculosis (TB) was basedon the serendipitous observation that nicotinamide, curiously,had activity against tubercle bacilli in animal models (19). Sub-sequent synthesis of analogues of nicotinamide led to the iden-tification of PZA as the most active derivative against Myco-bacterium tuberculosis (12). PZA is an important component ofthe current 6-month short-course TB chemotherapy. This ther-apy, which consists of isoniazid, rifampin, PZA, and ethambu-tol and is also called DOTS (for directly observed treatment,short-course), is recommended by the World Health Organi-zation for treatment of every TB patient (30). PZA plays aunique role in shortening the therapy from a period of 9 to 12months down to 6 months, because PZA kills a population ofsemidormant tubercle bacilli, residing in an acidic environment(occurring during active inflammation), which are not killed byother TB drugs (6, 14). Unlike other TB drugs, PZA, despiteits remarkable activity in vivo (10), has no activity againsttubercle bacilli in vitro in normal culture medium (26) exceptunder acidic-pH conditions (e.g., pH 5.5) (11).
M. tuberculosis is uniquely susceptible to PZA, whose MICfor this bacterium is about 16 to 50 mg/ml (11). In contrast, oth-er mycobacteria, and all nonmycobacteria, are completely in-sensitive to PZA (5). In M. tuberculosis, the susceptibility toPZA correlates with the presence of a single enzyme with nico-tinamidase and pyrazinamidase (PZase) activities (7). Strainsof M. tuberculosis that are resistant to PZA are often defectivein PZase activity (7, 9, 28), and we have recently cloned theM. tuberculosis PZase gene (pncA) (21) and shown that muta-
tion of pncA is a major mechanism of PZA resistance in M. tu-berculosis (22). However, the correlation between PZase activ-ity and PZA susceptibility does not exist for nontuberculousmycobacteria, since they have ample PZase activity but arenevertheless intrinsically resistant to PZA (5, 25). Despite theperformance of many studies, the mode of action of PZA inM. tuberculosis is unknown. Just as isoniazid requires activationby the M. tuberculosis catalase-peroxidase (32), PZA, as a pro-drug, needs to be activated by the bacterial nicotinamidase-PZase into pyrazinoic acid (POA) (7, 21), the active form ofthe drug, in bacterial cells. Yet, the active derivative POA isnot directly used to treat TB patients, because the bactericidalactivity of POA, when given orally to mice infected with M. tu-berculosis, was found to be not as significant as that of theprodrug PZA, presumably due to poor absorption through thegastrointestinal tract and to significant serum binding (7).However, the reasons why PZA requires an acidic environmentto show activity and why M. tuberculosis is uniquely susceptibleto PZA were unknown. In this study, we show that the roleof acidic pH is to enhance the accumulation of POA and thatM. tuberculosis has a defective efflux mechanism for POAwhereas the naturally PZA-resistant bacterium M. smegmatishas a much more active POA efflux mechanism.
MATERIALS AND METHODS
Chemicals and radiochemicals. Carbonyl cyanide m-chlorophenylhydrazone(CCCP), nigericin, valinomycin, reserpine, N,N9-dicyclohexyl carbodiimide(DCCD), sodium salicylate, [14C]benzoic acid, and [14C]salicylic acid were ob-tained from Sigma Chemical Co. [carbonyl-14C]PZA (specific activity, 52 mCi/mmol) was kindly supplied by the National Institutes of Health AIDS ReagentsProgram, Rockville, Md. The amount of [14C]PZA used in the various experi-ments was between 1 and 2 mCi/ml, which is equivalent to 2.4 to 4.8 mg of PZA/ml; these PZA concentrations are much lower than its MIC for M. tuberculosisand are thus unlikely to cause cidal effects. [14C]POA was generated by incubat-ing [14C]PZA with purified M. tuberculosis PZase, overexpressed in Escherichiacoli, for about 60 min at 37°C, conditions under which [14C]PZA was convertedto [14C]POA completely as judged by thin-layer chromatography (TLC) analysis(see below).
Effect of pH on [14C]PZA conversion and [14C]POA accumulation in M. tuber-culosis. Late-log-phase M. tuberculosis H37Ra cultures (2 to 3 week old), grownin 7H9 liquid medium supplemented with albumin-dextrose-catalase, were cen-
* Corresponding author. Mailing address: Department of MolecularMicrobiology and Immunology, School of Hygiene and Public Health,Johns Hopkins University, 615 N. Wolfe St., Baltimore, MD 21205.Phone: (410) 614-2975. Fax: (410) 955-0105. E-mail: [email protected].
† Present address: Virus Research Institute, Cambridge, MA 02138.
2044
on August 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

trifuged, and the cells were resuspended to a density of about 5 3 109/ml in 7H9medium adjusted to various pH values. [14C]PZA was added to a concentrationof 2.5 mCi/ml. Following incubation at 37°C for about 16 h, both the supernatantfluids and bacterial lysates, prepared by sonication of concentrated bacterial cellswashed with phosphate-buffered saline, were analyzed by TLC followed by au-toradiography (see below). To test the effect of pH on POA accumulation in theM. tuberculosis cells, [14C]POA was added to the bacterial suspensions, at variouspH values, at a concentration of 1 mCi/ml. At various times, 50-ml portions wereremoved, filtered through 0.45-mm-pore-size nitrocellulose membranes, andwashed with 0.1 M potassium phosphate buffer (pH 7.0) containing 0.1 M LiCl.In this as well as other experiments, the radioactivity associated with cells wasmeasured by scintillation counting. The intracellular concentration of POA wascalculated by assuming that 1 mg of dry cells is equivalent to 3 ml of internalwater (31).
TLC. For TLC, 2-ml portions of radioactive supernatants or lysates werespotted onto a 0.25-mm-thick silica G gel 60 plate with an aluminum backing(Whatman). The TLC plate was developed in 1-butanol–10% ammonia (5:1).The plate was then air dried and exposed to X-ray film for autoradiography.
Determination of intracellular pH. M. tuberculosis H37Ra cells were grown in7H9 liquid medium (pH 6.6) to late log phase. The culture was centrifuged, andthe cells were resuspended in Sauton’s medium (pH 5.0) at a density of 5 3109/ml. POA was added to 0.4 or 4 mM (about 500 mg/ml). Salicylate, used as apositive control, was also added to 4 mM. Each sample was tested in triplicate.[14C]benzoic acid or [14C]salicylic acid (Sigma Chemical Co.), as a pH probe, wasadded to a final concentration of 1 mCi/ml at time zero, 50-ml portions wereremoved at various times and filtered through 0.45-mm-pore-size nitrocellulosemembranes, and the membranes were washed twice with 2 ml of 7H9 medium.Scintillation cocktail (3 ml) was then added to each filter in a counting vial, andthe radioactivity was measured with a scintillation counter. The internal cellvolume was measured by using 3H2O and [14C]taurine, and the internal pH wascalculated according to the method of Rottenberg (18).
Isolation of membrane and cytoplasmic fractions from M. tuberculosis. Alate-log-phase culture of M. tuberculosis H37Ra (50 ml) was harvested, and thecells were washed twice with 40 mM potassium phosphate buffer (pH 6.5)containing 1 mM EDTA and then resuspended in 5 ml of 40 mM potassiumphosphate buffer (pH 6.5) containing 0.3 mM phenylmethylsulfonyl fluoride,1,000 U of DNase I, 0.5 mg of RNase A, and 2 mM magnesium chloride. The cellsuspension (5 ml) was sonicated for 10 to 15 min on ice and then centrifuged at13,000 rpm for 15 min to remove large cellular debris and unbroken cells. Thesupernatant was then spun at 32,000 rpm for 1 h to separate the membrane andcytosolic fractions. The supernatant fraction (cytosolic fraction) was saved. Thepellet (membrane fraction) was washed with 40 mM potassium phosphate buffer(pH 6.5) containing 0.3 mM phenylmethylsulfonyl fluoride and 1 mM EDTA andthen dissolved in 40 mM potassium phosphate buffer (pH 6.5) containing 1%Triton X-100. Both the supernatant and pellet fractions were tested for PZaseactivity by incubating them with [14C]PZA (1 to 2 mCi) in a volume of 30 ml for7 h, and the degree of conversion of [14C]PZA to [14C]POA was monitored byTLC as described above.
PZA accumulation and conversion in PZA-susceptible and -resistant M. tu-berculosis complex organisms. Two- to 3-week-old M. tuberculosis H37Ra andM. bovis BCG cultures, grown in Sauton’s medium, were harvested and thenwashed with Sauton’s medium, and the cell pellets were resuspended in Sauton’smedium (pH 6.6) at 5 3 109 cells/ml. [14C]PZA was added to these cell suspen-sions to a concentration of 1 mCi/ml, and the cell mixtures were incubated at37°C. At different time points, 50-ml portions were removed and washed withSauton’s medium by filtration on 0.45-mm-pore-size nitrocellulose filters by theuse of a vacuum pump. The amount of radioactivity associated with the bacterialcells was determined by scintillation counting.
Effect of reserpine and valinomycin on accumulation of POA in M. smegmatisand M. tuberculosis. [14C]PZA was added to a concentrated bacterial suspension(5 3 109 cells/ml) in 7H9 liquid medium at pH 6.6 to a final concentration of 2mCi/ml. A sublethal concentration of reserpine (20 mM) was added to theM. smegmatis cells after they had been incubated with [14C]PZA for 1 min,allowing a substantial amount of PZA to be taken up by the cells and convertedto POA. At various times after the addition of reserpine, 50-ml portions wereremoved and spotted onto 0.45-mm-pore-size nitrocellulose membranes under avacuum. Because washing the M. smegmatis cells with buffer tends to remove[14C]POA associated with the cells, the supernatant was removed by vacuumfiltration without washing. The membrane area where the cell suspension wasspotted was cut out, and the radioactivity was determined. The effect of valino-mycin (1 mM; sublethal concentration) was determined in the same manner,using 10 mM potassium. The effect of reserpine and valinomycin on [14C]POAaccumulation in M. tuberculosis was examined in a similar manner with thefollowing modifications. Reserpine (50 mM) and valinomycin (1 mM) were added2 h after addition of [14C]PZA to allow sufficient conversion of PZA to POA.Portions (50 ml) of suspension were filtered, and the radioactive cells on themembrane were washed twice with 2 ml of 0.1 M potassium phosphate buffer(pH 7.0) containing 0.1 M LiCl.
RESULTS
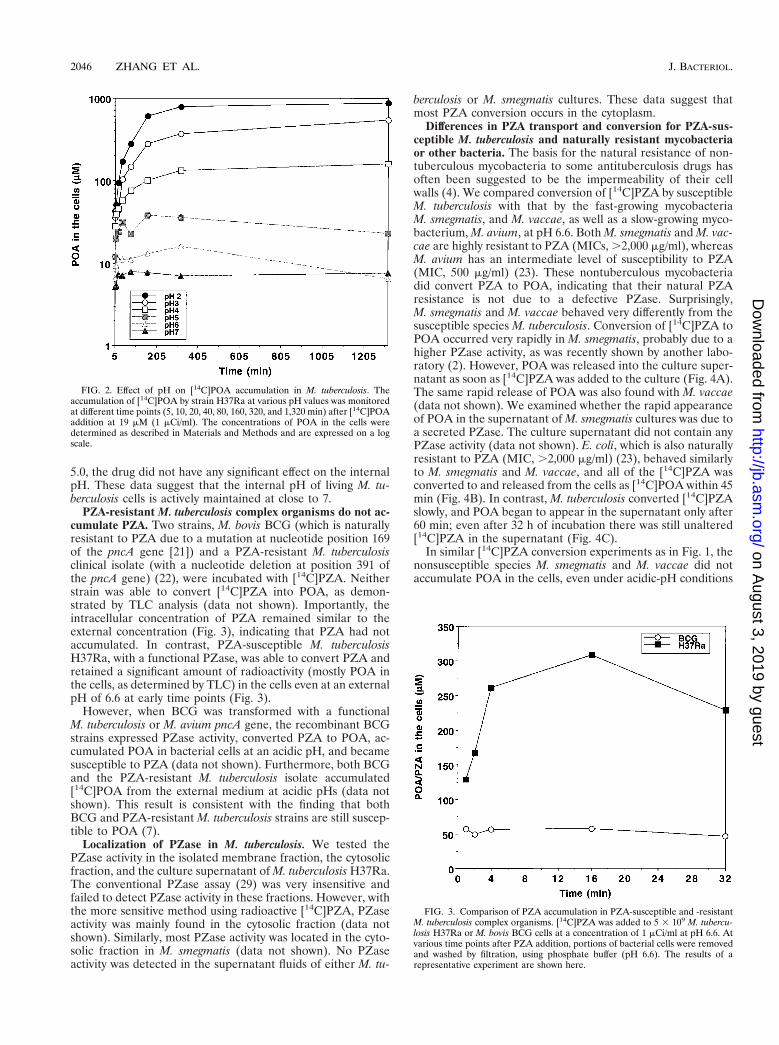
Acidic pH enhances accumulation of POA in M. tuberculosis.We examined the conversion of PZA to POA and the accu-mulation of POA in M. tuberculosis H37Ra by incubating bac-terial cells with [14C]PZA for 16 h under various pH condi-tions. Culture supernatants and bacterial lysates were analyzedby TLC followed by autoradiography (Fig. 1). [14C]PZA wasconverted to POA, which was not further converted into othercomponents (Fig. 1). At neutral or alkaline pHs, there waslittle POA associated with the bacterial cells and POA wasfound mainly in the supernatant. In contrast, at acidic pHs,there was much more [14C]POA associated with the bacterialcells (Fig. 1), although the conversion of [14C]PZA was some-what reduced, judging from the increased amount of unaltered[14C]PZA in the supernatant. This was apparently due to acidinhibition of the PZase (data not shown).
We then tested whether the increased accumulation of POAin the M. tuberculosis cells under acidic-pH conditions was dueto passive distribution of POA between the bacterial cellsand the medium. Using [14C]POA, we found that the effect ofacidic pH was indeed to enhance the accumulation of POA (aweak acid with a pKa of 2.9 [3]) in M. tuberculosis cells and thatthe POA accumulation was more pronounced at lower pHvalues (Fig. 2). This observation is consistent with the expec-tation that weak acids are accumulated intracellularly underacidic-pH conditions because an equilibrium is reached whenthe concentration of the uncharged form or conjugate acid,which permeates through the membrane by passive diffusion,becomes equal on both sides of the membrane (16, 18). It isnoteworthy that there was a major difference in between POAaccumulation at pH 6.6 and that at pH 2 to 5 (Fig. 1 and 2). AtpH 6.6, conditions under which PZA has no antituberculosisactivity (26), the POA concentration in the M. tuberculosis cellswas similar to the external concentration; by contrast, the POAconcentrations in the cells at acidic pH values, conditions un-der which PZA shows activity, were several-hundred-fold high-er than the external concentration, as expected from the pas-sive distribution theory (Fig. 1 and 2).
The internal pH. We measured the internal pH of M. tuber-culosis H37Ra cells at an external pH of 5.0, using [14C]benzoicacid or [14C]salicylic acid as a probe. The internal pH of H37Racells was close to 7 and did not decrease upon exposure to anacidic pH of 5.0 for 5 to 6 h. Addition of salicylate (4 mM)lowered the internal pH quickly, as expected. POA at 4 mM(500 mg/ml), however, did not significantly lower the internalpH of the M. tuberculosis cells, probably because at pH 5.0,according to the Henderson-Hasselbach equation, only 0.8%of the POA is uncharged and therefore POA crosses the mem-brane too slowly. When M. tuberculosis H37Ra cells were in-cubated with PZA at a concentration of 50 mg/ml for 2 h at pH
FIG. 1. Effect of pH on [14C]PZA conversion in M. tuberculosis. [14C]PZAwas incubated with M. tuberculosis H37Ra cells for 16 h under various pHconditions. [14C]PZA conversion and accumulation of [14C]POA by H37Ra inboth supernatant and cell lysate were analyzed by TLC.
VOL. 181, 1999 MODE OF ACTION OF PYRAZINAMIDE IN M. TUBERCULOSIS 2045
on August 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
5.0, the drug did not have any significant effect on the internalpH. These data suggest that the internal pH of living M. tu-berculosis cells is actively maintained at close to 7.
PZA-resistant M. tuberculosis complex organisms do not ac-cumulate PZA. Two strains, M. bovis BCG (which is naturallyresistant to PZA due to a mutation at nucleotide position 169of the pncA gene [21]) and a PZA-resistant M. tuberculosisclinical isolate (with a nucleotide deletion at position 391 ofthe pncA gene) (22), were incubated with [14C]PZA. Neitherstrain was able to convert [14C]PZA into POA, as demon-strated by TLC analysis (data not shown). Importantly, theintracellular concentration of PZA remained similar to theexternal concentration (Fig. 3), indicating that PZA had notaccumulated. In contrast, PZA-susceptible M. tuberculosisH37Ra, with a functional PZase, was able to convert PZA andretained a significant amount of radioactivity (mostly POA inthe cells, as determined by TLC) in the cells even at an externalpH of 6.6 at early time points (Fig. 3).
However, when BCG was transformed with a functionalM. tuberculosis or M. avium pncA gene, the recombinant BCGstrains expressed PZase activity, converted PZA to POA, ac-cumulated POA in bacterial cells at an acidic pH, and becamesusceptible to PZA (data not shown). Furthermore, both BCGand the PZA-resistant M. tuberculosis isolate accumulated[14C]POA from the external medium at acidic pHs (data notshown). This result is consistent with the finding that bothBCG and PZA-resistant M. tuberculosis strains are still suscep-tible to POA (7).
Localization of PZase in M. tuberculosis. We tested thePZase activity in the isolated membrane fraction, the cytosolicfraction, and the culture supernatant of M. tuberculosis H37Ra.The conventional PZase assay (29) was very insensitive andfailed to detect PZase activity in these fractions. However, withthe more sensitive method using radioactive [14C]PZA, PZaseactivity was mainly found in the cytosolic fraction (data notshown). Similarly, most PZase activity was located in the cyto-solic fraction in M. smegmatis (data not shown). No PZaseactivity was detected in the supernatant fluids of either M. tu-
berculosis or M. smegmatis cultures. These data suggest thatmost PZA conversion occurs in the cytoplasm.
Differences in PZA transport and conversion for PZA-sus-ceptible M. tuberculosis and naturally resistant mycobacteriaor other bacteria. The basis for the natural resistance of non-tuberculous mycobacteria to some antituberculosis drugs hasoften been suggested to be the impermeability of their cellwalls (4). We compared conversion of [14C]PZA by susceptibleM. tuberculosis with that by the fast-growing mycobacteriaM. smegmatis, and M. vaccae, as well as a slow-growing myco-bacterium, M. avium, at pH 6.6. Both M. smegmatis and M. vac-cae are highly resistant to PZA (MICs, .2,000 mg/ml), whereasM. avium has an intermediate level of susceptibility to PZA(MIC, 500 mg/ml) (23). These nontuberculous mycobacteriadid convert PZA to POA, indicating that their natural PZAresistance is not due to a defective PZase. Surprisingly,M. smegmatis and M. vaccae behaved very differently from thesusceptible species M. tuberculosis. Conversion of [14C]PZA toPOA occurred very rapidly in M. smegmatis, probably due to ahigher PZase activity, as was recently shown by another labo-ratory (2). However, POA was released into the culture super-natant as soon as [14C]PZA was added to the culture (Fig. 4A).The same rapid release of POA was also found with M. vaccae(data not shown). We examined whether the rapid appearanceof POA in the supernatant of M. smegmatis cultures was due toa secreted PZase. The culture supernatant did not contain anyPZase activity (data not shown). E. coli, which is also naturallyresistant to PZA (MIC, .2,000 mg/ml) (23), behaved similarlyto M. smegmatis and M. vaccae, and all of the [14C]PZA wasconverted to and released from the cells as [14C]POA within 45min (Fig. 4B). In contrast, M. tuberculosis converted [14C]PZAslowly, and POA began to appear in the supernatant only after60 min; even after 32 h of incubation there was still unaltered[14C]PZA in the supernatant (Fig. 4C).
In similar [14C]PZA conversion experiments as in Fig. 1, thenonsusceptible species M. smegmatis and M. vaccae did notaccumulate POA in the cells, even under acidic-pH conditions
FIG. 2. Effect of pH on [14C]POA accumulation in M. tuberculosis. Theaccumulation of [14C]POA by strain H37Ra at various pH values was monitoredat different time points (5, 10, 20, 40, 80, 160, 320, and 1,320 min) after [14C]POAaddition at 19 mM (1 mCi/ml). The concentrations of POA in the cells weredetermined as described in Materials and Methods and are expressed on a logscale.
FIG. 3. Comparison of PZA accumulation in PZA-susceptible and -resistantM. tuberculosis complex organisms. [14C]PZA was added to 5 3 109 M. tubercu-losis H37Ra or M. bovis BCG cells at a concentration of 1 mCi/ml at pH 6.6. Atvarious time points after PZA addition, portions of bacterial cells were removedand washed by filtration, using phosphate buffer (pH 6.6). The results of arepresentative experiment are shown here.
2046 ZHANG ET AL. J. BACTERIOL.
on August 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

(data not shown). Consistent with this observation, when [14C]POA was added to cells at pH 5.5, little POA was found to beassociated with the M. smegmatis cells, whereas increasingamounts of externally added [14C]POA entered into M. tuber-culosis over time (Fig. 5). In the case of M. avium, both the rateof [14C]PZA conversion and the amount of [14C]POA associ-ated with the M. avium cells under acidic pH conditions wereintermediate between the values for M. tuberculosis andM. smegmatis (data not shown). This finding is consistent withthe intermediate level of susceptibility of M. avium to PZA(23).
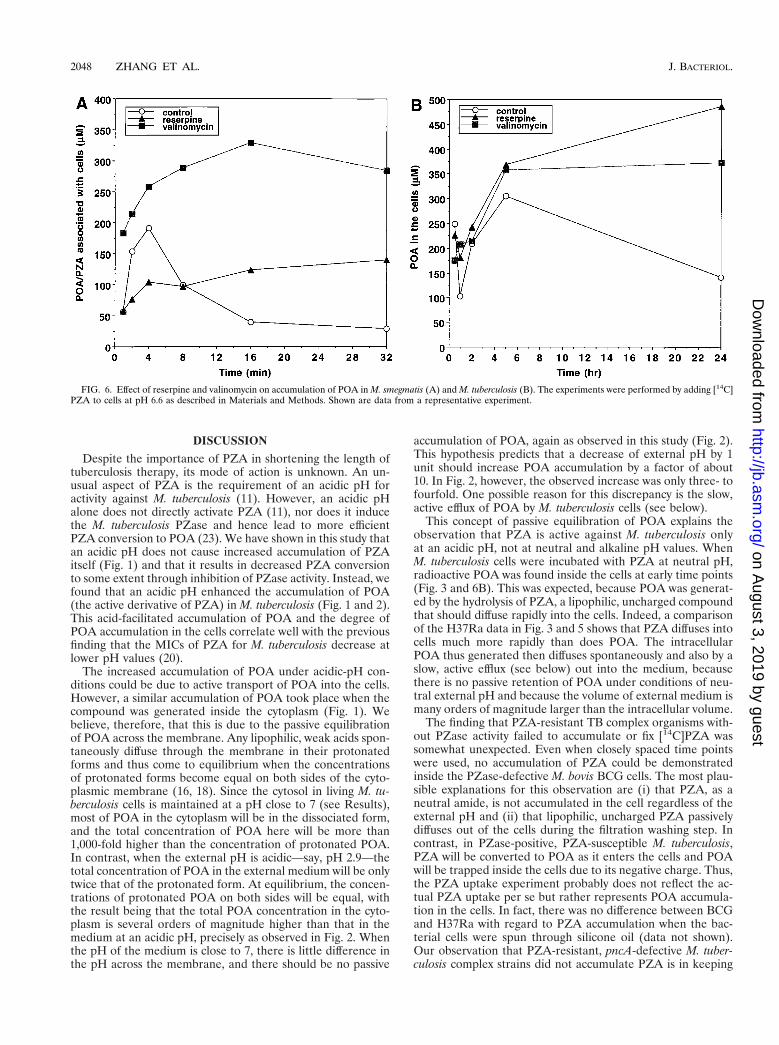
POA efflux in M. smegmatis and M. tuberculosis. POA is ex-pected to be accumulated passively under acidic-pH condi-tions, but it did not accumulate in M. smegmatis (Fig. 5); wesuspected that this was due to an active efflux mechanismpumping POA out of the cells. To prove this, we tested variousinhibitors of energy metabolism, including CCCP (8, 27), ni-gericin (17), valinomycin (17), and DCCD (1), as well as re-serpine, a plant alkaloid that inhibits the Bacillus subtilis mul-tidrug efflux pump (15), by incubating M. smegmatis cells with[14C]PZA at pH 6.6. Interestingly, reserpine at a sublethalconcentration (20 mM) allowed significant accumulation ofPOA in M. smegmatis cells (Fig. 6A), although it inhibited PZAconversion to some degree because of its inhibition of PZaseactivity (data not shown). M. smegmatis cells extruded POAvery rapidly (Fig. 6A). Since at the internal pH of 7 only 0.01%of the POA is expected to be in the membrane-permeable un-charged form, this rapid efflux strongly suggests that M. smeg-matis has an active efflux mechanism that pumps out POA,a conclusion also supported by the reserpine inhibition data.Among the energy inhibitors, valinomycin at a sublethal con-centration (0.85 to 1 mM) caused significant retention of [14C]POA in M. smegmatis cells (Fig. 6A).
To determine whether there is an efflux mechanism for POAin M. tuberculosis, we tested the effects of reserpine and vali-nomycin on the accumulation of [14C]POA in M. tuberculosisH37Rv by adding [14C]PZA at pH 6.6. Reserpine caused POAaccumulation in H37Rv cells (Fig. 6B), suggesting that M. tu-berculosis also has an active POA efflux mechanism. Valino-mycin also caused significant retention of POA in M. tubercu-losis cells (Fig. 6B). However, the rate of POA extrusion (about0.3 pmol/mg/min [Fig. 6B]) was more than 2 orders of magni-tude lower than that found in M. smegmatis (about 70 pmol/mg/min [Fig. 6A]).
FIG. 4. Comparison of [14C]PZA conversion and [14C]POA release amongM. smegmatis, E. coli, and M. tuberculosis. Bacterial cell suspensions (about 5 3109 cells/ml) were prepared from early-stationary-phase cultures and resus-pended in an appropriate medium at pH 6.6. [14C]PZA (1 mCi/ml) was added tothe bacterial suspensions, and the radioactive cell mixtures were incubated at37°C for various periods of time up to 1 h for M. smegmatis (A) and E. coli (B)and up to 32 h for M. tuberculosis H37Ra (C). The extent of [14C]PZA conversionto [14C]POA in the supernatant fluids was monitored by TLC.
FIG. 5. Differences in POA accumulation between M. tuberculosis (M. tb)and M. smegmatis (M. smeg). [14C]POA (2.5 mCi/ml) was added to bacterialsuspensions with a density of 5 3 109 cells/ml at an acidic pH of 5.5, and the cellswere incubated at 37°C for various lengths of time up to 7 h. Portions of thebacterial cell suspensions (50 ml) were removed and washed with Sauton’s me-dium by filtration under a vacuum. The internal POA concentrations in thebacterial cells were determined as described in Materials and Methods.
VOL. 181, 1999 MODE OF ACTION OF PYRAZINAMIDE IN M. TUBERCULOSIS 2047
on August 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
DISCUSSION
Despite the importance of PZA in shortening the length oftuberculosis therapy, its mode of action is unknown. An un-usual aspect of PZA is the requirement of an acidic pH foractivity against M. tuberculosis (11). However, an acidic pHalone does not directly activate PZA (11), nor does it inducethe M. tuberculosis PZase and hence lead to more efficientPZA conversion to POA (23). We have shown in this study thatan acidic pH does not cause increased accumulation of PZAitself (Fig. 1) and that it results in decreased PZA conversionto some extent through inhibition of PZase activity. Instead, wefound that an acidic pH enhanced the accumulation of POA(the active derivative of PZA) in M. tuberculosis (Fig. 1 and 2).This acid-facilitated accumulation of POA and the degree ofPOA accumulation in the cells correlate well with the previousfinding that the MICs of PZA for M. tuberculosis decrease atlower pH values (20).
The increased accumulation of POA under acidic-pH con-ditions could be due to active transport of POA into the cells.However, a similar accumulation of POA took place when thecompound was generated inside the cytoplasm (Fig. 1). Webelieve, therefore, that this is due to the passive equilibrationof POA across the membrane. Any lipophilic, weak acids spon-taneously diffuse through the membrane in their protonatedforms and thus come to equilibrium when the concentrationsof protonated forms become equal on both sides of the cyto-plasmic membrane (16, 18). Since the cytosol in living M. tu-berculosis cells is maintained at a pH close to 7 (see Results),most of POA in the cytoplasm will be in the dissociated form,and the total concentration of POA here will be more than1,000-fold higher than the concentration of protonated POA.In contrast, when the external pH is acidic—say, pH 2.9—thetotal concentration of POA in the external medium will be onlytwice that of the protonated form. At equilibrium, the concen-trations of protonated POA on both sides will be equal, withthe result being that the total POA concentration in the cyto-plasm is several orders of magnitude higher than that in themedium at an acidic pH, precisely as observed in Fig. 2. Whenthe pH of the medium is close to 7, there is little difference inthe pH across the membrane, and there should be no passive
accumulation of POA, again as observed in this study (Fig. 2).This hypothesis predicts that a decrease of external pH by 1unit should increase POA accumulation by a factor of about10. In Fig. 2, however, the observed increase was only three- tofourfold. One possible reason for this discrepancy is the slow,active efflux of POA by M. tuberculosis cells (see below).
This concept of passive equilibration of POA explains theobservation that PZA is active against M. tuberculosis onlyat an acidic pH, not at neutral and alkaline pH values. WhenM. tuberculosis cells were incubated with PZA at neutral pH,radioactive POA was found inside the cells at early time points(Fig. 3 and 6B). This was expected, because POA was generat-ed by the hydrolysis of PZA, a lipophilic, uncharged compoundthat should diffuse rapidly into the cells. Indeed, a comparisonof the H37Ra data in Fig. 3 and 5 shows that PZA diffuses intocells much more rapidly than does POA. The intracellularPOA thus generated then diffuses spontaneously and also by aslow, active efflux (see below) out into the medium, becausethere is no passive retention of POA under conditions of neu-tral external pH and because the volume of external medium ismany orders of magnitude larger than the intracellular volume.
The finding that PZA-resistant TB complex organisms with-out PZase activity failed to accumulate or fix [14C]PZA wassomewhat unexpected. Even when closely spaced time pointswere used, no accumulation of PZA could be demonstratedinside the PZase-defective M. bovis BCG cells. The most plau-sible explanations for this observation are (i) that PZA, as aneutral amide, is not accumulated in the cell regardless of theexternal pH and (ii) that lipophilic, uncharged PZA passivelydiffuses out of the cells during the filtration washing step. Incontrast, in PZase-positive, PZA-susceptible M. tuberculosis,PZA will be converted to POA as it enters the cells and POAwill be trapped inside the cells due to its negative charge. Thus,the PZA uptake experiment probably does not reflect the ac-tual PZA uptake per se but rather represents POA accumula-tion in the cells. In fact, there was no difference between BCGand H37Ra with regard to PZA accumulation when the bac-terial cells were spun through silicone oil (data not shown).Our observation that PZA-resistant, pncA-defective M. tuber-culosis complex strains did not accumulate PZA is in keeping
FIG. 6. Effect of reserpine and valinomycin on accumulation of POA in M. smegmatis (A) and M. tuberculosis (B). The experiments were performed by adding [14C]PZA to cells at pH 6.6 as described in Materials and Methods. Shown are data from a representative experiment.
2048 ZHANG ET AL. J. BACTERIOL.
on August 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

with the previous finding that E. coli mutants defective in pncA(encoding nicotinamidase) also failed to accumulate nicotin-amide (13).
The data presented in Fig. 4 show the kinetic differencesin PZA uptake, conversion to POA, and potential POA effluxamong various bacteria. We believe that the differences reflectdifferences in PZase enzyme activity, POA efflux, and, to amuch lesser extent, cell wall permeability, because PZA is aneutral amide and should passively diffuse into different cellsrelatively easily.
In this study, we found that the naturally PZA-resistantM. smegmatis has a highly active efflux mechanism for POAwhich can be inhibited by reserpine and valinomycin (Fig. 6A).Although nigericin and CCCP did not show inhibition, thiscould be due to the inadequate entry of these agents throughthe rather impermeable cell wall or to their active efflux. Thevalinomycin effect thus suggests that the pump is energized bythe proton motive force or one of its components. M. tubercu-losis also has a POA efflux mechanism, as demonstrated byincreased accumulation of POA at neutral pH in the presenceof reserpine and valinomycin (Fig. 6B). The M. tuberculosisefflux mechanism is much weaker than that of M. smegmatis, asevidenced by the orders-of-magnitude-slower kinetics of POAextrusion (compare Fig. 6B and A). Thus, in M. tuberculosis, atan acidic external pH, the rate of passive transmembrane equi-librium of POA apparently overwhelms that of active efflux,resulting in a huge accumulation of POA in the cells. The POAefflux mechanism in M. smegmatis appears to be different fromthe recently identified M. smegmatis MDR pump LfrA (24),since insertion of the lfrA gene into M. tuberculosis H37Ra didnot cause enhanced efflux of POA or increased resistance toPZA or POA (unpublished data). Studies designed to identifythe POA efflux mechanisms in M. smegmatis and M. tubercu-losis are under way.
While both susceptible M. tuberculosis and other nonsuscep-tible mycobacteria have PZases to convert PZA to POA, thespecificity of PZA for M. tuberculosis appears to be conferredat the stage of POA efflux, which is much weaker in M. tuber-culosis than in the nonsusceptible M. smegmatis. It is notewor-thy that the two types of PZA resistance, the acquired PZAresistance found in susceptible M. tuberculosis and the intrinsicPZA resistance found in nontuberculous mycobacteria, arecaused by very different mechanisms. Acquired PZA resistancein susceptible M. tuberculosis is caused by mutations in thepncA gene which render the organisms unable to convert theprodrug PZA to bactericidal POA. In contrast, the intrinsicPZA resistance in M. smegmatis, and probably in many othernontuberculous mycobacteria, is due to a much more activePOA efflux mechanism which does not allow accumulation ofPOA in the cells.
ACKNOWLEDGMENTSWe thank Peter Maloney for helpful discussions; Diane Griffin,
Barbara Laughon, and Denis Mitchison for encouragement; and theNational Institutes of Health (NIH) AIDS Reagents Program for[14C]pyrazinamide.
This work was supported by research grants from the American LungAssociation, the Potts Memorial Foundation, and NIH (RO1AI40584)to Y.Z.
REFERENCES1. Azzi, A., R. P. Casey, and M. J. Nalecz. 1984. The effect of N,N9-dicyclo-
hexylcarbodiimide on enzymes of bioenergetic relevance. Biochim. Biophys.Acta 768:209–226.
2. Boshoff, H. I. M., and V. Mizrahi. 1998. Purification, gene cloning, targetedknockout, overexpression, and biochemical characterization of the majorpyrazinamidase from Mycobacterium smegmatis. J. Bacteriol. 180:5809–5814.
3. Budavari, S. 1989. The Merck index, 11th ed. Merck & Co., Inc., Rahway,N.J.
4. David, H. 1981. Basis for lack of drug susceptibility of atypical mycobacteria.Rev. Infect. Dis. 3:878–884.
5. Good, R. C., V. A. Silcox, J. O. Kilburn, and B. D. Plikaytis. 1985. Identifi-cation and drug susceptibility test results for Mycobacterium spp. Clin. Mi-crobiol. Newslett. 7:133–136.
6. Heifets, L., and P. Lindholm-Levy. 1992. Pyrazinamide sterilizing activity invitro against semi-dormant Mycobacterium tuberculosis bacterial populations.Am. Rev. Respir. Dis. 145:1223–1225.
7. Konno, K., F. M. Feldman, and W. McDermott. 1967. Pyrazinamide suscep-tibility and amidase activity of tubercle bacilli. Am. Rev. Respir. Dis. 95:461–469.
8. Li, X.-Z., D. M. Livermore, and H. Nikaido. 1994. Role of efflux pump(s) inintrinsic resistance of Pseudomonas aeruginosa: resistance to tetracycline, chlor-amphenicol, and norfloxacin. Antimicrob. Agents Chemother. 38:1732–1741.
9. McClatchy, J. K., A. Y. Tsang, and M. S. Cernich. 1981. Use of pyrazinami-dase activity in Mycobacterium tuberculosis as a rapid method for determi-nation of pyrazinamide susceptibility. Antimicrob. Agents Chemother. 20:556–557.
10. McCune, R. M., R. Tompsett, and W. McDermott. 1956. The fate of Myco-bacterium tuberculosis in mouse tissues as determined by the microbial enu-meration technique. J. Exp. Med. 104:763–802.
11. McDermott, W., and R. Tompsett. 1954. Activation of pyrazinamide andnicotinamide in acidic environment in vitro. Am. Rev. Tuberc. 70:748–754.
12. McKenzie, D., L. Malone, S. Kushner, J. J. Oleson, and Y. Subbarow. 1948.The effect of nicotinic acid amide on experimental tuberculosis of whitemice. J. Lab. Clin. Med. 33:1249–1253.
13. McLaren, J., D. T. C. Ngo, and M. Olivera. 1973. Pyridine nucleotide me-tabolism in Escherichia coli. J. Biol. Chem. 248:5144–5159.
14. Mitchison, D. A. 1985. The action of antituberculosis drugs in short coursechemotherapy. Tubercle 66:219–225.
15. Neyfakh, A. A., V. E. Bidnenko, and L. B. Chen. 1991. Efflux-mediatedmultidrug resistance in Bacillus subtilis: similarities and dissimilarities withthe mammalian system. Proc. Natl. Acad. Sci. USA 88:4781–4785.
16. Nikaido, H., and D. G. Thanassi. 1993. Penetration of lipophilic agents withmultiple protonation sites into bacterial cells: tetracyclines and fluoroquino-lones as examples. Antimicrob. Agents Chemother. 37:1393–1399.
17. Ramos, S., S. Schuldiner, and H. R. Kaback. 1976. The electrochemicalgradient of protons and its relationship to active transport in Escherichia colimembrane vesicles. Proc. Natl. Acad. Sci. USA 73:1892–1896.
18. Rottenberg, H. 1979. The measurement of membrane potential and DpH incells, organelles, and vesicles. Methods Enzymol. 55:547–569.
19. Ryan, F. 1993. The forgotten plague, p. 345–347. Little, Brown and Com-pany, Boston, Mass.
20. Salfinger, M., and L. B. Heifets. 1988. Determination of pyrazinamide MICsfor Mycobacterium tuberculosis at different pHs by the radiometric method.Antimicrob. Agents Chemother. 32:1002–1004.
21. Scorpio, A., and Y. Zhang. 1996. Mutations in pncA, a gene encoding pyrazi-namidase/nicotinamidase, cause resistance to the antituberculous drug py-razinamide in tubercle bacillus. Nat. Med. 2:662–667.
22. Scorpio, A., P. Lindholm-Levy, L. Heifets, R. Gilman, S. Siddiqi, M. Cyna-mon, and Y. Zhang. 1997. Characterization of pncA mutations in pyrazin-amide-resistant Mycobacterium tuberculosis. Antimicrob. Agents Chemother.41:540–543.
23. Sun, A., A. Scorpio, and Y. Zhang. 1997. The pncA gene from naturallypyrazinamide-resistant Mycobacterium avium encodes pyrazinamidase andconfers pyrazinamide susceptibility to resistant M. tuberculosis complex or-ganisms. Microbiology 143:3367–3373.
24. Takiff, H. E., M. Cimino, M. C. Musso, T. Weisbrod, R. Martinez, M. B.Delgado, L. Salazar, B. R. Bloom, and W. R. Jacobs. 1996. Efflux pump of theproton antiporter family confers low-level fluoroquinolone resistance in My-cobacterium smegmatis. Proc. Natl. Acad. Sci. USA 93:362–366.
25. Tarnok, I., and E. Rohrscheidt. 1976. Biochemical background of someenzymatic tests used for the differentiation of mycobacteria. Tubercle 57:145–150.
26. Tarshis, M. S., and W. A. Weed. 1953. Lack of significant in vitro sensitivityof Mycobacterium tuberculosis to pyrazinamide on three different solid me-dia. Am. Rev. Tuberc. 67:391–395.
27. Thanassi, D. G., L. W. Cheng, and H. Nikaido. 1997. Active efflux of bile saltsby Escherichia coli. J. Bacteriol. 179:2512–2518.
28. Trivedi, S. S., and S. G. Desai. 1987. Pyrazinamidase activity of Mycobacte-rium tuberculosis—a test of sensitivity to pyrazinamide. Tubercle 68:221–224.
29. Wayne, L. G. 1974. Simple pyrazinamidase and urease tests for routineidentification of mycobacteria. Am. Rev. Respir. Dis. 109:147–151.
30. World Health Organization. 1995. WHO report on the tuberculosis epi-demic: stop TB at the source. Tuberculosis Programme, World Health Or-ganization, Geneva, Switzerland.
31. Youatt, J., and S. H. Tham. 1969. Radioactive content of Mycobacteriumtuberculosis after exposure to 14C-isoniazid. Am. Rev. Respir. Dis. 100:77–78.
32. Zhang, Y., B. Heym, B. Allen, D. Young, and S. Cole. 1992. The catalase-peroxidase and isoniazid resistance in Mycobacterium tuberculosis. Nature358:591–593.
VOL. 181, 1999 MODE OF ACTION OF PYRAZINAMIDE IN M. TUBERCULOSIS 2049
on August 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from