role of alpha- and beta-adrenoreceptors in rat monocyte/macrophage function at rest and acute...
TRANSCRIPT

ORIGINAL PAPER
Role of alpha- and beta-adrenoreceptors in ratmonocyte/macrophage function at rest and acute exercise
Juliane da Silva Rossato & Mauricio Krause & Augustus Joli Martins Fernandes &
João Roberto Fernandes & Isis Lenhard Seibt & Anderson Rech &
Paulo Ivo Homem de Bittencourt Jr.
Received: 27 August 2013 /Accepted: 22 December 2013# University of Navarra 2014
Abstract Previous studies fromour laboratory have dem-onstrated that a single bout of moderate exercise stimulatesmacrophage function, increasing phagocytic capacity, andproduction of hydrogen peroxide and nitric oxide (NO˙)through nuclear factor kappa B activation. In this work, weinvestigated the role of α- and β-adrenoreceptors on thefunction of monocyte/macrophages during rest and exer-cise. Adult male Wistar rats were i.p. administered(100μL/100 g) with specific adrenergic antagonists beforean acute moderate exercise bout: prazosin (α1-specificantagonist 2 mg/kg), propranolol (unspecific β1/β2 antag-onist 10 mg/kg), double blockade (α1 and β1/β2), orphosphate-buffered saline (control). Acute exerciseconsisted in a single swimming session of moderate inten-sity (5 % body weight overload on the chest) for 60 min.Control groups (rest) received the same antagonists and
were killed 60 min after drug administration. Exerciseincreased phagocytic capacity (1.7-fold, p<0.05), NO˙production (5.24 fold, p<0.001), and inducible nitric oxidesynthase (NOS2) expression (by 58.1 %), thus suggestingmacrophage activation. The β-adrenoreceptor blockadedid not change this behavior. In resting animals, α1 antag-onist, as well as the double (α1/β) blockade, however,further increased phagocytic capacity (by up to 261 %,p<0.001), NO˙ production (by up to 328 %, p<0.001),and the expressions of NOS2 (by 182 %, p<0.001) andHSP70 (by 42.5 %, p<0.01) suggesting a tonic inhibitoryeffect of α1 stimulation on macrophage activation. Inexercised animals,α1-blockade showed similar enhancingeffect on phagocytic indices and expressions of NOS andHSP70, particularly in double-blocked groups, althoughNO˙ production was found to be reduced in exercisedanimals submitted to both α- and β-blockade. Redox(glutathione) status and lipoperoxidation were evaluatedin all test groups and approximately paralleled macro-phage NO˙ production. We suggest the prevalence of aperipheral α1-adrenoreceptor inhibitory tonus that limitsmacrophage responsiveness but operates differently afterphysical exercise.
Keywords Adrenoreceptors . Monocyte/macrophagefunction . Exercise . Nitric oxide and phagocytosis
Introduction
Monocytes are blood cells with a wide range of func-tions such as phagocytosis, antigen presentation, and
J Physiol BiochemDOI 10.1007/s13105-013-0310-3
Juliane da Silva Rossato and Mauricio Krause equally contributedto this work.
J. da Silva Rossato :M. Krause (*) :A. J. M. Fernandes :J. R. Fernandes : I. L. Seibt :A. Rech :P. I. Homem de Bittencourt Jr. (*)Laboratory of Cellular Physiology, Department of Physiology,Institute of Basic Health Sciences, Federal University of RioGrande do Sul, Rua Sarmento Leite 500 (ICBS),90050-170 Porto Alegre, Rio Grande do Sul, Brazile-mail: [email protected]: [email protected]
J. da Silva Rossato :M. Krause :A. J. M. Fernandes :J. R. Fernandes : I. L. Seibt :A. Rech :P. I. Homem de Bittencourt Jr.National Institute of Hormones and Women’s Health,Porto Alegre, RS, Brazil

cytokine secretion, playing a central role for innate andacquired immunological responses [20, 21]. Physicalexercise is known to modulate monocyte responses byinducing nuclear factor kappa B (NFκB) activation,leading to pro-inflammatory cytokine production(TNF-α and IL-1) and inducible nitric oxide synthase(NOS2) expression. In fact, moderate physical exerciseenhances macrophage phagocytosis and hydrogen per-oxide (H2O2) and nitric oxide (NO˙) production [45].Nevertheless, the underlying mechanisms for this im-munological activation are still under debate.
Both sympathetic nervous system and hypothala-mus–pituitary–adrenal axis are required for the generalstress responses, by modulating the immunological ac-tivation through the release of peripheral signaling mol-ecules, such as adrenaline and noradrenaline, that canact directly through specific adrenoreceptors within theimmune cells [45]. During exercise, for example, incre-ments in monocytes chemotaxis and phagocytosis fromhumans and animals have been demonstrated to bedependent on α- and β-adrenoreceptor activation [15,39–41]. To date, there are only a few published studieslooking at the role of pharmacological adrenoreceptormanipulation in vivo during the exercise.
In this work, we aimed to investigate the effects of apharmacological adrenergic blockade (α and β antago-nists) during rest and after single bout of moderateexercise (swimming, 5 % body weight overload) onthe macrophage responses such as phagocytic capacity,glutathione metabolism, NOS2 and the 70-kDa familyof heat shock protein (HSP70) expression, nitric oxideproduction, and lipoperoxidation.
Materials and methods
Animals Adult male Wistar rats (3–4 months of age, n=40) obtained from The Institute of Basic HealthSciences Animal Care Facility (CREAL-ICBS) werehoused in pairs with food and water ad libitum. Theanimals were maintained under a 12-h light period(lights on at 7:00 am), at 22±2 °C, and relative humidityof 60%. Care and use of the animals were in accordancewith protocols approved by the Federal University ofRio Grande do Sul Animal Utilization EthicsCommission (protocol #23078.037562/08–41) whichadheres to guidelines of The Brazilian NationalCouncil for the Control of Animal Experimentation.
Experimental groups and adrenergic antagonistdrugs Animals were allocated in four groups (n=10each) based on the drug received: no drug group (salinephosphate-buffered saline (PBS)), prazosin group (α1
antagonist), propranolol group (β1, β2 antagonist), anddouble blockage group (α and β blockage). Five ani-mals in each group were submitted to an acute moderateexercise bout (named Exercise), and five did not per-form any type of exercise (named Control). Specificsubclasses of adrenergic receptors antagonists were ad-ministered (to the respective group) intraperitoneally(100 μL/100 g) 30 min before exercise protocol: saline(PBS), prazosin (α1 antagonist 2 mg/kg), propranolol(β1,β2 antagonist 10 mg/kg), and double blockade (αandβ) [1, 30]. In addition to these groups, we also testeda separate group using phentolamine (α1, α2 antagonist,10 mg/kg); however, since this drug induced a massivehypoglycemia (data not shown), we excluded this groupfrom the analysis. Indeed, phentolamine, a non-specificα antagonist, has an inhibitory action in α2 receptors,induced insulin increase, and, consequently, reducedblood glucose concentration during the exercise [47].
Acute exercise protocol Previously to the acute exercisesession, animals were adapted to the swimming tanks inthree separated sessions (in different days) of 10, 15, and20 min of swimming without load attached, whichcharacterizes a low intensity exercise and did not induceany training adaptation [26, 33, 45]. Drugs were admin-istered (to the respective group) intraperitoneally 30 minbefore the exercise. Rats were individually submitted toan exercise protocol (exercise groups only) consisting ofan acute swimming session (60 min), in warmed water(29–30 °C) [24, 38]. Flotation was prevented by fixingan overweight (5 % body weight) to the animal chest[19], characterizing a moderate intensity exercise. Thecontrol groups (no exercise) were kept in a 5-cm layer ofwarmed water for the same period. After 60 min, exer-cise and control animals were killed by decapitation andthe blood prepared for the analysis.
Measurement of blood lactate and glucose concentrationsBlood samples for lactate and glucose determina-tions were collected before and just after the exer-cise. The lactate and glucose concentrations were deter-mined using an automated system Accutrend Plus®(Roche) [30].
J. Silva Rossato et al.

Lipoperoxide measurement (by xylenol orange assay)After killing, blood samples were immediatelycollected into heparinized tubes and placed on ice.Plasma was obtained by centrifugation at 2,000×g for10 min at 4 °C. Butylated hydroxytoluene (BHT) inmethanol was added to a final concentration of20 μmol/L to avoid further oxidative reactions thatmay cause falsely elevated results. The plasma sampleswere stored at −86 °Cwithin 1 h after sample collection.Final ferrous oxidation in xylenol orange (FOX) reagentcomprised the reagent 2 mM of xylenol orange and40 mM of BHT in methanol 90 % (vol/vol).Ammonium ferrous sulfate (10 mM) was dissolved in1 mL of 25 mM sulfuric acid and was added to themethanol solution. Aliquots (90 μL) of plasma weretransferred into microcentrifuge vials (1.5 mL), and10 μL of 10 mM triphenylphosphine in methanol wasadded to half of the vials to reduce ROOHs, therebygenerating blanks. Methanol (10 μL) was added to theremaining other half of vials to produce test samples. Allvials were then mixed and incubated at room tempera-ture for 30 min. After mixing, the samples were incu-bated at room temperature for another 30 min with FOXreagent. Absorbance of the supernatant was measured at570 nm (Benchmark model, Bio-Rad). Plasma proteincontent was spectrophotometrically assessed byBradford method [7] in microplates [4, 29, 46].
GSH and GSSG in peripheral blood erythrocytesErythrocytes were prepared as described in Silveiraet al. [45]. Briefly, peripheral blood was collectedjust after killing into heparinized tubes and centrifugedat 2,000×g for 4 min at 4 °C. The erythrocytespellet (1 mL) was disrupted in 8 mL of 5 % (w/v)metaphosphoric acid (MPA) at 4 °C. The erythrocytessamples were centrifuged (15,000×g for 5 min at 4 °C),and the supernatant fractions were then assessed forglutathione (GSH) and glutathione disulfide (GSSG)content using an adaptation of the GSSG reductaserecycling method, with N-ethylmaleimide (Fluka) con-jugating technique for GSSG sample preparation [2, 3,31]. The samples, for GSH and GSSG, were assayed at415 nm in a 105-μL final volume in 96-well polystyreneplates (Corning) at 37 °C in the presence of 10 mM 5,5′-dithio-bis 2-nitrobenzoic acid, 0.017 mM β-NADPH(dissolved in 0.5 % (w/v) NaHCO3), and 0.5 U/mL GSSG reductase (EC 1.6.4.2, Sigma). Resultswere expressed in nanomoles per milliliter of redblood cell (RBC).
Monocyte/macrophage preparations Soon after killing,the blood for phagocytic assay was collected in hepa-rinized tubes, and mononuclear cells were separated bylayering blood onto a Histopaque 1077 (Sigma) discon-tinuous gradient, following the manufacturer’s instruc-tions. After centrifugation (400×g for 30 min at roomtemperature), the turbid interface containing mononu-clear leukocytes was then washed and resuspended inPBS as previously described [45]. Afterward, mono-cytes were seeded in RPMI1640 culture media supple-mented with 10% (v/v) fetal calf serum (FCS) onto glasscover slips in six-well sterile culture plates (Nunc,Denmark) to produce a final concentration of 5×105
cells in each well. Cells were adhered at 37 °C for60 min in a 5 % CO2 atmosphere in filtered air (Harrismicro-processed CO2 humidified water-jacked incuba-tor). Non-adherent cells were removed by washing threetimes with PBS. This monocyte preparation was imme-diately used in the experiments measuring phagocytosisand NO˙. For the studies assessing the expression ofNOS2 and HSP70, monocyte/macrophage preparationswere left in RPMI1640 culture supplemented with anti-biotics and 10 % (v/v) FCS in six-well dishes for 6 hbefore protein extraction and electrophoresis in order toallow for protein expression peaks to be reached.
Phagocytic activity assay Monocyte/macrophage phago-cytic capacity was estimated by the phagocytosis ofzymosan particles (Saccharomyces cerevisae) as previ-ously described [45]. Fifty milligrams of zymosan(S. cerevisae ghosts, Sigma) was diluted in 100 mL ofPBS, boiled for 30 min, washed three times (by centri-fugation at 15,000×g for 10 s at room temperature), andresuspended in 2.5 mL of PBS which were mixed with2.5 mL of autologous rat serum. The latter mixture wasthen incubated for 30 min at 37 °C under agitation inorder to promote opsonization of zymosan particles bythe serum complement. Blood monocytes in RPMI me-dium were replaced by 1 mL of pre-warmed Hanks’balanced salt solution (HBSS; Sigma) containing20mg/mL defatted BSA, and phagocytosis was initiatedby the addition of 200 mL of opsonized zymosan parti-cles. After 30-min incubation at 37 °C, phagocytosiswas interrupted by placing the culture dishes on ice for10 min. Phagocytic activity was assessed by countingthe total amount of engulfed zymosan particles in 100randomly assigned macrophages in different fields in aNeubauer’s chamber under light microscopy (×1,000magnification, with an immersion objective in a phase
Adrenoreceptors and monocyte/macrophage function in exercise

contrast light microscope, Olympus ix81). This gavethe number of total phagocytosed zymosan parti-cles (ZT) and the percentage of phagocytosing cell(PT) in relation to the total number of cells scored(NT). Phagocytic index (phI), as previouslyassigned [25], was calculated as follows: numberof phagocytosed particles (ZT) divided by the num-ber of cells scored (NT) multiplied by the percentphagocytosing cells (PT), so that phI=(ZT/NT)×100 %×(PT/NT)=[(ZT×PT)/(NT)
2]×100 %.
Determination of nitrite production The NO˙ producedby actived monocytes reacts rapidly with oxygen toproduce nitrite (NO2
−) ions. Therefore, the effect ofexercise on NO˙ production by unstimulated macro-phages obtained from circulating blood monocyteswas assessed by measuring the production of NO2
− bymonocytes/macrophages incubated for 60 min in HBSS(see above) at 37 °C. Before this incubation, the cellswere plaqued in 96-well plates and left to attach for30 min in RPMI1640 medium in the presence of 10 %(v/v) FCS. After incubations in HBSS, NO2
− producedby the cells (2–5×105 per well) were directly assayed byusing the Griess reaction essentially as described bySilveira et al. [45]. The reactions were performed intriplicate by addition of 50 μL of fresh supernatant to50 μL of Griess reagent [0.1 % naphthalenediaminedihydrochloride/1 % sulfanilamide in 5 % (w/v)H3PO4] (Sigma). After incubation at room temperaturefor 10 min, the absorbance was read at 540 nm [34, 35].The NO2
− concentration was determined by comparisonto a sodium nitrite standard curve.
SDS-PAGE and western blot analysis of NOS2 andHSP70 protein expression Protein separation and im-munoblotting analyses were performed as adapted fromelsewhere [32]. Monocytes/macrophages were preparedand 6-h incubated in RPMI1640 culture medium, asdescribed above after that cells were washed three timeswith PBS and collected in cold PBS by using polyeth-ylene cell scraper (Sigma). The samples were centri-fuged (15,000×g for 10 min at room temperature), andthe pelleted cells were lysed in 0.1 % (w/v) sodiumdodecyl sulfate solution containing 2 μg/mL leupeptinand 100 mM PMSF and centrifuged at 15,000×g for2 min at 4 °C. After protein determination [7] (40 μg),the samples were prepared and electroforesed in a 10 %polyacrylamide minigel (Bio-Rad). The proteins weretransferred onto nitrocellulose membranes (GE Health
Care-Amersham) according to the electrotransfer (Bio-Rad) manufacturer’s instructions, prepared for immuno-blotting and incubated for 2 h with anti-HSP70 mono-clonal antibody (Sigma H5147, ascites), which recog-nizes both the 73-kDa constitutive/cognate HSC70(encoded by the hspa8 gene) and the 72-kDa inducibleHSP72 (encoded by the hspa1a gene) forms ofHSP70 at 1:1,000 dilution or with antibodies againstthe 130-kDa inducible iNOS (NOS2) rabbit anti-rat/human NOS2 IgG fraction of antiserum (Sigma) usedat 1:500 dilution. Afterward, membranes were probedwith horseradish peroxidase-labeled secondary antibod-ies (Sigma, 1:1,000) for 2 h at room temperature, and theproteins were visualized in the presence enhancedchemiluminescence (ECL) by using ECL Plus (GEHealth Care-Amersham RPN2132). Protein bands werequantified in the VDS GE Health Care-Amersham sys-tem and accompanying Image Master software. β-Actinanalysis was performed to ensure equal sample loading,and after image digitalization and quantification, proteinexpression was normalized in terms of β-actin.
Statistical analysis For not-normally distributed data, alogarithmic transformation was applied (Log10).ANOVA general linear model was performed tocompare the sub-groups. Post hoc Tukey test wasfurther applied when appropriate. The alpha levelwas set atP<0.05. Data were analyzed usingMinitab 16software.
Results
Metabolites With the exception of the post-exercisepropranolol group, no changes in glycemia were foundbetween the groups and treatments (Table 1). Bloodlactate values increased after the exercise confirmingthe effects of a moderate intensity exercise. No changesin lactate were detected in the control (rest) groups uponadrenoreceptor blockade.
Macrophage activity Our results confirmed that exer-cise stimulates phagocytosis and microbicide capacity(Fig. 1). Phagocytosis indices were increased 1.7-fold inthe exercise-PBS as compared to control-PBS group(p<0.05), corroborating previous results from our labo-ratory [45]. Interestingly, however, phagocytic capacitywas even more elevated in control animals treated with
J. Silva Rossato et al.

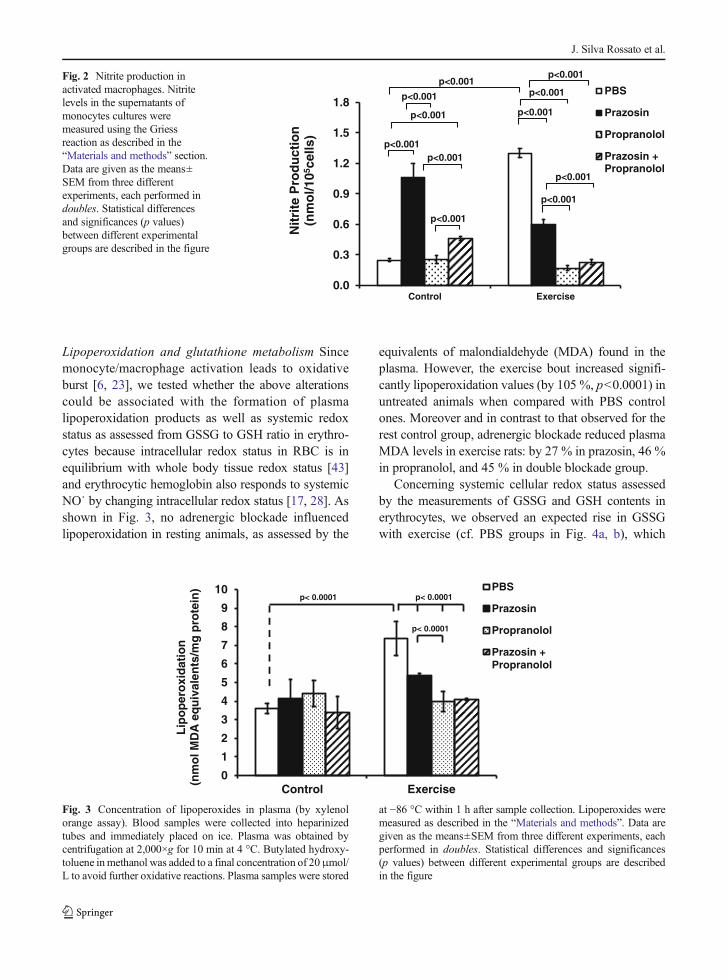
either prazosin (3.61-fold, p<0.001) or a combination ofprazosin and propranolol (2.28-fold, p<0.05) as com-pared to controls treated just with PBS, whereas pro-pranolol alone showed no effect. Nevertheless, inexercised animals, no single pharmacological interven-tion could further change the rise in phagocytic capacityimposed by exercise in comparison with nonexercisedcontrols. However, the association of prazosin and pro-pranolol treatment did increase phagocytic capacity inexercised animals (2.02-fold, as compared to exercise-PBS group, p<0.05) to a level (phI=16.67 %) similar tothat found in resting prazosin-treated rats (phI=17.41 %). Regarding NO˙ production (Fig. 2), wefound, again, a marked effect of exercise, in that nitriteaccumulation in macrophage incubation medium was5.24-fold higher than that observed in resting untreatedPBS group (p<0.001). Also similarly to that observedfor the phagocytic capacity, prazosin increased NO2
−
production 4.28-fold in relation to resting controlanimals, whereas the association of prazosin andpropranolol evoked a 1.84-fold augmentation inNO2
− production as compared to PBS group.Differently to that found in phagocytic assays, adren-ergic blocking treatments resulted in marked decrease(p<0.001) in NO2
− production in comparison withexercised PBS-treated animals (prazosin, by 54.3 %;propranolol, by 86.8 %; double blockade), whichsuggests that in exercised animals both α1- and β-adrenoreceptors participate in NO production bymacrophages. Thus, α1-adrenoreceptor alone has noeffect on exercise-induced phagocytic capacity butstimulates NO˙ production by macrophages, whereasβ-adrenoreceptor reduces the exercise effect on thephagocytosis only if combined with α1-receptorstimulation but stimulates NO˙ production both aloneand in co-operation with α1-adrenoreceptors.
Table 1 Blood glucose and lactate concentrations after experimental sessions and adrenoreceptor blockade
Treatment Glycemia (mg/dL) Lactate (mmol/L)
Post-rest Post-exercise Post-rest Post-exercise
Control (PBS) 78.2±2.6 78.3±0.8 2.9±0.1 4.6±0.1*
Prazosin 86.6±4.7 93±2.4 2.6±0.14 3.6±0.6*
Propranolol 69.3±3.4 67.3±5.1** 2.76±0.22 4±0.2*
Prazosin+propranolol 72±1.4 77±3 2.61±0.14 3.4±0.3*
Rats were adapted to swimming protocol and treated with adrenoreceptor blockers (or PBS controlled) as described in the “Materials andmethods” section. After 60 min, exercised and control (resting) animals were killed by decapitation and the blood prepared for the analysis.Data are expressed as the means±SD (n=10 per group)
*p<0.05 (different from rest values of the same group); **p<0.05 (different from the other drugs post exercise values)
0
5
10
15
20
25
Control Exercise
ph
ago
cytc
ind
ex
PBS
Prazosin
Propranolol
Prazosin+Propranolol
p<0.001
p<0.05
p<0.05
p<0.05
p<0.01p<0.05
p<0.05
p<0.001Fig. 1 Phagocytosis index.Monocyte/macrophagephagocytic capacity estimated bythe phagocytosis of serum-opsonized zymosan particles(S. cerevisae ghosts). Data aregiven as the means±SEM fromthree different experiments, eachperformed in doubles. Statisticaldifferences and significances(p values) between differentexperimental groups aredescribed in the figure
Adrenoreceptors and monocyte/macrophage function in exercise
Lipoperoxidation and glutathione metabolism Sincemonocyte/macrophage activation leads to oxidativeburst [6, 23], we tested whether the above alterationscould be associated with the formation of plasmalipoperoxidation products as well as systemic redoxstatus as assessed from GSSG to GSH ratio in erythro-cytes because intracellular redox status in RBC is inequilibrium with whole body tissue redox status [43]and erythrocytic hemoglobin also responds to systemicNO˙ by changing intracellular redox status [17, 28]. Asshown in Fig. 3, no adrenergic blockade influencedlipoperoxidation in resting animals, as assessed by the
equivalents of malondialdehyde (MDA) found in theplasma. However, the exercise bout increased signifi-cantly lipoperoxidation values (by 105 %, p<0.0001) inuntreated animals when compared with PBS controlones. Moreover and in contrast to that observed for therest control group, adrenergic blockade reduced plasmaMDA levels in exercise rats: by 27 % in prazosin, 46 %in propranolol, and 45 % in double blockade group.
Concerning systemic cellular redox status assessedby the measurements of GSSG and GSH contents inerythrocytes, we observed an expected rise in GSSGwith exercise (cf. PBS groups in Fig. 4a, b), which
0.0
0.3
0.6
0.9
1.2
1.5
1.8
Control ExerciseN
itri
te P
rod
uct
ion
(nm
ol/1
05ce
lls)
PBS
Prazosin
Propranolol
Prazosin +Propranolol
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
p<0.001
Fig. 2 Nitrite production inactivated macrophages. Nitritelevels in the supernatants ofmonocytes cultures weremeasured using the Griessreaction as described in the“Materials and methods” section.Data are given as the means±SEM from three differentexperiments, each performed indoubles. Statistical differencesand significances (p values)between different experimentalgroups are described in the figure
0
1
2
3
4
5
6
7
8
9
10
Control Exercise
Lip
op
ero
xid
atio
n
(nm
ol M
DA
eq
uiv
alen
ts/m
g p
rote
in) PBS
Prazosin
Propranolol
PropranololPrazosin +
p< 0.0001
p< 0.0001
p< 0.0001
Fig. 3 Concentration of lipoperoxides in plasma (by xylenolorange assay). Blood samples were collected into heparinizedtubes and immediately placed on ice. Plasma was obtained bycentrifugation at 2,000×g for 10 min at 4 °C. Butylated hydroxy-toluene inmethanol was added to a final concentration of 20μmol/L to avoid further oxidative reactions. Plasma samples were stored
at −86 °C within 1 h after sample collection. Lipoperoxides weremeasured as described in the “Materials and methods”. Data aregiven as the means±SEM from three different experiments, eachperformed in doubles. Statistical differences and significances(p values) between different experimental groups are describedin the figure
J. Silva Rossato et al.

0.0
4.0
8.0
12.0
16.0
20.0
Control Exercise
GS
H c
on
ten
t (
mo
l/mL
of
RB
C)
PBS
Prazosin
Propranolol
Prazosin +Propranolol
c
0.00
0.50
1.00
1.50
2.00
2.50
Control Exercise
GS
SG
co
nte
nt
(m
ol/m
L o
f R
BC
)
PBS
Prazosin
Propranolol
Prazosin +Propranolol
c
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Control Exercise
RB
C R
edu
xS
tatu
s([
GS
SG
]/[G
SH
] ra
tio
PBS
Prazosin
Propranolol
Prazosin +Propranolol
c
b
ap< 0.0001
p< 0.0001
p< 0.0001
p< 0.0001
p< 0.0001
p< 0.0001
p< 0.001
p< 0.0001
p< 0.0001
p< 0.05
p< 0.0001
p< 0.01 p< 0.001
p< 0.0001
p< 0.0001 p< 0.0001
p<0.0001
p<0.0001
p<0.0001
p<0.05
p<0.0001 p<0.0001
p <0.0001
Fig. 4 Glutathione metabolismin rat erythrocytes. Peripheralblood was collected inheparinized tubes and centrifugedat 2,000×g for 4 min at 4 °C. Theerythrocyte pellet (1 mL) wasdisrupted in 8 mL of 5 % MPA at4 °C. The erythrocyte sampleswere centrifuged (15,000×g for5 min at 4 °C), and thesupernatant fractions were thenassessed for glutathione (GSH; a)and glutathione disulphide(GSSG; b) content as described inthe “Materials and methods”section. Red blood cell (RBC)GSSG to GSH ratio, an index ofintracellular redox status, was alsogiven (c). Data are given as themeans±SEM from three differentexperiments, each performed indoubles. Statistical differencesand significances (p values)between different experimentalgroups are described in the figure
Adrenoreceptors and monocyte/macrophage function in exercise

was counterbalanced by an enhancement in the amountof GSH in RBC, leading to an unaltered systemic redoxstatus, since GSSG to GSH ratio did not change withexercise bouts (Fig. 4c). On the other hand, bothα- and β-blockers changed this scenario.Accordingly, in control resting rats, prazosin en-hanced RBC redox status by 80 %, this meaningthat α1-blockade led to a marked oxidative stress(Fig. 4c), whereas β-blockade caused an impres-sive oxidative challenge as evaluated by the 3.9-fold rise in [GSSG]/[GSH]. Curiously, the admin-istration of prazosin to propranolol-treated rats(double blockade) completely abolished this effectin nonexercised animals. A similar effect was not-ed in exercised animals: only a marginal rise in[GSSG]/[GSH] ratio in prazosin group, in compar-ison with PBS treatment, and a conspicuous 7.4-fold elevation in propranolol group, an effect that wasdismissed by the double blockade, suggesting an antag-onizing effect between these two adrenoreceptors(Fig. 4c).
NOS2 and HSP70 expression in macrophages The ex-pression of NOS2 (Fig. 5a), which is induced in macro-phages in a NFκB activation-dependent manner, wasfound to rise (by 58 %, p<0.05) in exercised-rat macro-phages after 6 h post-bout, as previously results fromthis laboratory have demonstrated [45]. Moreover,prazosin enhanced this effect even more (2.1-fold,p<0.01), and propranolol did not influence the effectof exercise bout on NOS2 expression, neither alone norin combination with prazosin. This was not the case inunexercised animals, in which both prazosin and pro-pranolol conspicuously enhanced NOS2 expression(2.8- and 2.4-fold, respectively). The association of bothadrenoreceptor blockers, however, produced no addi-tional effect (Fig. 5a). On the other hand, the expressionof HSP70, whose 72-kDa form (HSP72) is inducedby a wide variety of stress stimuli, including NFκBactivation, was not affect by the exercise bout(Fig. 5b), confirming our previous report [45].HSP70 have a protective activity as help the correc-tion of misfolded proteins (by oxidative stress, forexample), and for this reason, we used this proteinas a marker for oxidative damage. Nevertheless, it isof note that prazosin was able to increase monocyteHSP70 expression both in unexercised (by 43 %) andexercised (by 77 %) rats (p<0.05), an effect that was
not altered by the association with propranolol (dou-ble blockade) and propranolol alone either (Fig. 5b).
Discussion
Monocytes and macrophages are known to express bothα and β adrenergic receptors, and their activation mod-ulates the immune cell function [22]. The main findingof this study was that, at rest, α1 blockage increasesmacrophage activity, i.e., phagocytic capacity (Fig. 1),alongside NOS2 and HSP70 expression (Fig. 5), thussuggesting the existence of a tonic inhibitory effect ofcatecholamines over monocyte/macrophage activationvia α1-adrenoreceptor. Nevertheless, the production ofNO˙ soon after the exercise bout seemed to be differ-ently affected by α and β adrenergic stimulation(Fig. 2), consistent with the notion that catecholaminesinhibit macrophage nitric oxide production through β1
and β2 adrenoreceptors in LPS-elicited macrophages[44].
Confirming previous results [45], it was found anincrease in phagocytic activity in monocytes/macro-phages after the exercise. Curiously, however, therewas a marked increase in phagocytic capacity after theα1-blockade, independent on the exercise bout.Similarly, double blockage (α1 and β) induced the sameresult. Our data suggest different mechanisms of regu-lation of phagocytosis induced by catecholamines, witha prevalent inhibitory action (with anti-inflammatorycharacteristics), via α1 receptors under resting condi-tions [11, 39, 40]. Indeed, catecholamines acting viaα1 receptors have been shown to impose a suppressiveeffect on the production of H2O2 in peritoneal macro-phages in vitro, this being reversed by α-adrenergicblockade [10].
As previously reported, administration of prazosin, atdifferent doses in vivo, promotes an increase in theproduction of granulocytes and macrophages, suggest-ing a tonic sympathetic inhibition in myelopoiesis viaα1 receptors, which would be present in bone marrowcells, therefore preventing the growth and differentiationof colony-forming units of granulocytes and monocytes.In addition, it has been reported that α-adrenergic re-ceptors can stimulate chemotaxis and activation ofphagocytosis [39, 40]. In vitro studies have demonstrat-ed that administration of physiological doses of nor-adrenaline (10−9 M) results in an increase in the
J. Silva Rossato et al.

phagocytic capacity of monocytes/macrophages, whichis reversed by the use of α and β blockers [22, 39, 40,49].
Herein, in exercised animals, in which norepineph-rine levels may double from ca. 2.5 to 5.5 nM [9], singleα1-blockade was able to increase only NOS2 andHSP70 expression, whereas phagocytic capacity wasnot affected. Considering, in addition, that unspecificβ-blockage alone showed no influence on phagocytosisand expressions of NOS2 and HSP70 either, whiledouble blockade enhanced all these three variables, wespeculate that, firstly, expressions of NOS2 and HSP70and macrophage phagocytic activity are differently
influenced by catecholamines (and their exercise-dependent concentrations) and, secondly, phagocytosisseems to be also modulated by α2 adrenoreceptors dur-ing exercise.
Norepinephrine is a catecholamine which has prefer-ence, at low concentrations, for α receptors, whichpresent high affinity for the neurotransmitter, while athigher concentrations (such as during exercise) the bind-ing to β-adrenoreceptors is much enhanced [10]. Thismay explain the different effects of the adrenoreceptorblockage found in our study, such as the divergences onthe nitrite production. The α1-adrenergic blockage gen-erated an increase in phagocytic ability, production of
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Control ExerciseN
OS
-2/a
ctin
exp
ress
ion
(arb
itra
ry u
nit
s)
PBS
Prazosin
Propranolol
Prazosin +Propranolol
a
b
p<0.001
p<0.01
p<0.05
p<0.01
p<0.01
a
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Control Exercise
HS
P70
/act
in e
xpre
ssio
n(a
rbit
rary
un
its)
PBS
Prazosin
Propranolol
Prazosin +Propranololp<0.01
p<0.001p<0.01
p<0.05p<0.05
130 kDa
72 kDa
p<0.05
b
p<0.05
Fig. 5 Expression of NOS2 andHSP70 protein in macrophages.Cell isolation and treatment forwestern blot is described in the“Materials and methods” section.Protein bands were digitalizedand quantified in VDS GE HealthCare-Amersham system andaccompanying Image Mastersoftware. Sample loading wasmonitored by β-actin analysiswhich was used for proteinexpression normalization. Dataare given as the means±SEMfrom three different experiments,each performed in doubles.Statistical differences andsignificances (p values) betweendifferent experimental groups aredescribed in the figure
Adrenoreceptors and monocyte/macrophage function in exercise

nitrite, and expression of NOS2, clearly indicating theactivation of this cell. Additionally, the β-adrenergicreceptors appear to contribute in regulating the immedi-ate production of nitrite held by monocyte/macrophagedirectly by controlling the expression of NOS2 [10, 12,48, 49].
Physical exercise is a knownmodulator of monocyte/macrophage responses. This activation can be seen evenafter a short duration exercise [40], through generatingthe release of catecholamines which stimulate pro-inflammatory cytokine production [36, 39] by inducingNFκB activation, which, in turn, may trigger sustainedNOS2 and HSP70 expression several hours after exer-cise sessions. An acute session of moderate aerobicexercise can also increase the production of free radi-cals, which may result in an increase in the oxidativestress [13, 27], thus inducing the activation of NFκB-dependent pathways. In our work, under the effect of αand β adrenergic blockers, the animals that performedexercise showed values of plasma lipoperoxidation(Fig. 3) similar to their unexercised controls, as previ-ously reported [45]. Thus, it can be suggested thatcatecholamines exert a stimulatory effect on ROS syn-thesis increasing lipoperoxidation.
As we have previously debated [33], GSSG is aprimary glutathione S-conjugate physiologically pro-duced within the cells, and its accumulation induces asevere state of oxidative stress, inasmuch as the intracel-lular redox potential, i.e., the redox “voltage,” given bythe Nernst equation, is ε=εo–(RT/nF).ln (aGSSG/aGSH),where aGSSG and aGSH are the chemical activities ofGSSG and GSH, respectively. Assuming that intracellu-lar concentrations of GSSG and GSH (respectively[GSSG] and [GSH]) are sufficiently diluted, it is reason-able to write ε=εo− (RT/nF).ln ([GSSG]/[GSH]).Because of this, [GSSG]/[GSH] ratio is considered theindex of cellular redox status [2] and gives a panorama ofhow intracellular redox reactions may be affected if oneor both of the components of this quotient have changedas in the case of such adrenoreceptor blocking interven-tions, despite any acute production of lipoperoxides to-ward the plasma. Accordingly, it was found that [GSSG]/[GSH] ratio (Fig. 4) was markedly increased only in thegroups with β-blockade, both in rest and in exercisetests. This may be explained by the observation thatGSSG was increased while GSH was reduced by52 %, in both control and exercise animals. In addition,β-adrenergic receptors are known modulators of theantioxidant production in vitro [16]; therefore, the use
of β-blockers such as propranolol could generate apro-oxidative condition [37], as observed in ourresults.
Many other factors are suggested to modulate theimmune function in exercise, e.g., the release of HSP70(by hepatocytes and immune cells) and also the activa-tion of sympathetic corticotropin-releasing hormone(CRH)–histamine axis, both dependent on catechol-amine action. The production of extracellular HSP72(eHSP72), for example, is mediated by a noradrenergicstimulation via of α1 receptors in the liver, the mainorgan responsible for its release [8, 14, 30]. TheeHSP72 plays an important role in promoting the phago-cytic activity and production of mediators of the inflam-matory response via TLR2 and TLR4 (Toll-like recep-tors) [5, 18, 50]. Finally, the activation of the sympatheticCRH–histamine axis may occur in situations of acutestress such as exercise when CRH and histamine periph-erally produced in response to the first act as immune-stimulating factor and a pro-inflammatory agent [42]may participate in this process. These possibilities arecurrently under investigation in our laboratory.
Conclusions
We conclude that α1 blockage induces increments inthe phagocytosis, NO˙ production, expression ofNOS2 and HSP70 (essentially HSP72) in rest andafter exercise. This may indicate a tonic inhibitoryeffect produced by the activation of α1-adrenoreceptors, since the blockage of this receptorsleads to an increase in the responsiveness of thiscells. The participation of α2-adreonoreceptors andother factors such as eHSP70 and the CRH-histamine axis needs to be investigated. The reasonwhy NO˙ production can be so rapidly stimulated(within 1 h) after exercise is also a matter of furtherinvestigation. In summary, we suggest the prevalenceof a peripheral α1-adrenoreceptor inhibitory tonus thatlimits macrophage responsiveness but operates differ-ently after physical exercise.
Acknowledgments JSR, MSK, AJMF, JRF, ILS, and AR com-pleted all the experiments described in this manuscript. All authorswere involved in analyzing the results. JSR, MSK, and PIHBJdesigned the study and co-wrote the paper. PIHBJ was responsiblefor grant support with respect to Brazilian National Council forScientific and Technological Development (CNPq, grants 551097/
J. Silva Rossato et al.

2007-8, 402626/2012-5, and 402364/2012-0). All authors hadfinal approval of the submitted and published versions.
Conflict of interest The authors declare no conflict of interest.
References
1. Aarnio P, Lauritsen T, Dela F (2001) Insulin secretion andglucose kinetics during exercise with and without pharmaco-logical alpha(1)- and alpha(2)-receptor blockade. Diabetes 50:1834–1843
2. Akerboom TP, Sies H (1981) Assay of glutathione, glutathi-one disulfide, and glutathione mixed disulfides in biologicalsamples. Methods Enzymol 77:373–382
3. Anderson ME (1985) Determination of glutathione and gluta-thione disulfide in biological samples. Methods Enzymol 113:548–555
4. Arab K, Steghens JP (2004) Plasma lipid hydroperoxidesmeasurement by an automated xylenol orange method. AnalBiochem 325:158–163
5. Asea A, Rehli M, Kabingu E, Boch JA, Bare O, Auron PE,Stevenson MA, Calderwood SK (2002) Novel signal trans-duction pathway utilized by extracellular HSP70: role of toll-like receptor (TLR) 2 and TLR4. J Biol Chem 277:15028–15034
6. Babior BM, Kipnes RS, Curnutte JT (1973) Biological de-fense mechanisms. The production by leukocytes of superox-ide, a potential bactericidal agent. J Clin Invest 52:741–744
7. Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal Biochem 72:248–254
8. Campisi J, Fleshner M (2003) Role of extracellular HSP72 inacute stress-induced potentiation of innate immunity in activerats. J Appl Physiol 94:43–52
9. Couturier K, Servais S, Koubi H, Sempore B, Cottet-EmardJM, Guigas B, Lavoie JM, Favier R (2004) Metabolic andhormonal responses to exercise in the anti-obese Lou/C rats.Int J Obes Relat Metab Disord 28:972–978
10. Dimitrijevic M, Pilipovic I, Stanojevic S, Mitic K, RadojevicK, Pesic V, Leposavic G (2009) Chronic propranolol treatmentaffects expression of adrenoceptors on peritoneal macro-phages and their ability to produce hydrogen peroxide andnitric oxide. J Neuroimmunol 211:56–65
11. Elenkov IJ, Chrousos GP (1999) Stress, cytokine patterns andsusceptibility to disease. Baillieres Best Pract Res ClinEndocrinol Metab 13:583–595
12. Farmer P, Pugin J (2000) beta-Adrenergic agonists exert their“anti-inflammatory” effects in monocytic cells through theIkappaB/NF-kappaB pathway. Am J Physiol Lung Cell MolPhysiol 279:L675–L682
13. Fisher-Wellman K, Bloomer RJ (2009) Acute exercise andoxidative stress: a 30 year history. Dyn Med 8:1
14. Fleshner M, Johnson JD (2005) Endogenous extra-cellularheat shock protein 72: releasing signal(s) and function. Int JHyperthermia 21:457–471
15. Garcia JJ, del Carmen Saez M, De la Fuente M, Ortega E(2003) Regulation of phagocytic process of macrophages by
noradrenaline and its end metabolite 4-hydroxy-3-metoxyphenyl-glycol . Role of a lpha- and beta-adrenoreceptors. Mol Cell Biochem 254:299–304
16. Gillissen A, Wickenburg D, van Zwoll D, Schultze-Werninghaus G (1997) beta-2-Agonists have antioxidantfunction in vitro. 2. The effect of beta-2-agonists on oxidant-mediated cytotoxicity and on superoxide anion generated byhuman polymorphonuclear leukocytes. Respiration 64:23–28
17. Gladwin MT, Lancaster JR Jr, Freeman BA, Schechter AN(2003) Nitric oxide's reactions with hemoglobin: a viewthrough the SNO-storm. Nat Med 9:496–500
18. Gleeson M, McFarlin B, Flynn M (2006) Exercise and Toll-like receptors. Exerc Immunol Rev 12:34–53
19. Gobatto CA, de Mello MA, Sibuya CY, de Azevedo JR, dosSantos LA, Kokubun E (2001) Maximal lactate steady state inrats submitted to swimming exercise. Comp Biochem PhysiolA Mol Integr Physiol 130:21–27
20. Gordon S (2007) Macrophage heterogeneity and tissue lipids.J Clin Invest 117:89–93
21. Gordon S (2007) The macrophage: past, present and future.Eur J Immunol 37(Suppl 1):S9–S17
22. Gosain A, Muthu K, Gamelli RL, DiPietro LA (2007)Norepinephrine suppresses wound macrophage phagocyticefficiency through alpha- and beta-adrenoreceptor dependentpathways. Surgery 142:170–179
23. Gwinn MR, Vallyathan V (2006) Respiratory burst: role insignal transduction in alveolar macrophages. J ToxicolEnviron Health B Crit Rev 9:27–39
24. Harri M, Kuusela P (1986) Is swimming exercise or coldexposure for rats? Acta Physiol Scand 126:189–197
25. Hishikawa T, Cheung JY, Yelamarty RV, Knutson DW (1991)Calcium transients during Fc receptor-mediated and nonspecificphagocytosis by murine peritoneal macrophages. J Cell Biol 115:59–66
26. Hoffman-Goetz L, Spagnuolo PA, Guan J (2008) Repeatedexercise in mice alters expression of IL-10 and TNF-alpha inintestinal lymphocytes. Brain Behav Immun 22:195–199
27. Ji LL (1999) Antioxidants and oxidative stress in exercise.Proc Soc Exp Biol Med 222:283–292
28. Jia L, Bonaventura C, Bonaventura J, Stamler JS (1996) S-Nitrosohaemoglobin: a dynamic activity of blood involved invascular control. Nature 380:221–226
29. Jiang ZY,Woollard AC,Wolff SP (1991) Lipid hydroperoxidemeasurement by oxidation of Fe2+ in the presence of xylenolorange. Comparison with the TBA assay and an iodometricmethod. Lipids 26:853–856
30. Johnson JD,Campisi J, SharkeyCM,KennedySL,NickersonM,Fleshner M (2005) Adrenergic receptors mediate stress-inducedelevations in extracellular Hsp72. J Appl Physiol 99:1789–1795
31. Kolberg A, Rosa TG, Puhl MT, Scola G, da Rocha Janner D,Maslinkiewicz A, Lagranha DJ, Heck TG, Curi R, de BittencourtPI Jr (2006) Low expression of MRP1/GS-X pump ATPase inlymphocytes of Walker 256 tumour-bearing rats is associatedwith cyclopentenone prostaglandin accumulation and cancer im-munodeficiency. Cell Biochem Funct 24:23–39
32. Krause M, Bittencourt A, Homem de Bittencourt PI,McClenaghan NH, Flatt PR, Murphy C, Newsholme P(2012) Physiologic concentrations of IL-6 directly promoteinsulin secretion, signal transduction, nitric oxide release andredox status in a clonal pancreatic beta-cell line and mouseislets. J Endocrinol 214:301–311
Adrenoreceptors and monocyte/macrophage function in exercise

33. Krause MS, Oliveira LP Jr, Silveira EM, Vianna DR, RossatoJS, Almeida BS, RodriguesMF, Fernandes AJ, Costa JA, CuriR et al (2007) MRP1/GS-X pump ATPase expression: is thisthe explanation for the cytoprotection of the heart againstoxidative stress-induced redox imbalance in comparison toskeletal muscle cells? Cell Biochem Funct 25:23–32
34. Kroner EE, Peskar BA, Fischer H, Ferber E (1981) Control ofarachidonic acid accumulation in bone marrow-derived mac-rophages by acyltransferases. J Biol Chem 256:3690–3697
35. Kurose I, Wolf R, GrishamMB, Aw TY, Specian RD, GrangerDN (1995) Microvascular responses to inhibition of nitricoxide production. Role of active oxidants. Circ Res 76:30–39
36. Madden KS, Sanders VM, Felten DL (1995) Catecholamineinfluences and sympathetic neural modulation of immuneresponsiveness. Annu Rev Pharmacol Toxicol 35:417–448
37. Moniri NH, Daaka Y (2007) Agonist-stimulated reactive ox-ygen species formation regulates beta2-adrenergic receptorsignal transduction. Biochem Pharmacol 74:64–73
38. Niess AM, Dickhuth HH, Northoff H, Fehrenbach E (1999)Free radicals and oxidative stress in exercise—immunologicalaspects. Exerc Immunol Rev 5:22–56
39. Ortega E, Giraldo E, Hinchado MD, Martin L, Garcia JJ, De laFuente M (2007) Neuroimmunomodulation during exercise: roleof catecholamines as ‘stress mediator’ and/or ‘danger signal’ forthe innate immune response. Neuroimmunomodulation 14:206–212
40. Ortega E, Giraldo E, Hinchado MD, Martinez M, Ibanez S,Cidoncha A, Collazos ME, Garcia JJ (2006) Role of Hsp72 andnorepinephrine in the moderate exercise-induced stimulation ofneutrophils’ microbicide capacity. Eur J Appl Physiol 98:250–255
41. Ortega E, Marchena JM, Garcia JJ, Barriga C, Rodriguez AB(2005) Norepinephrine as mediator in the stimulation ofphagocytosis induced by moderate exercise. Eur J ApplPhysiol 93:714–718
42. Ralph JA, Zocco D, Bresnihan B, Fitzgerald O, McEvoyAN, Murphy EP (2007) A role for type 1alphacorticotropin-releasing hormone receptors in mediatinglocal changes in chronically inflamed tissue. Am J Pathol170:1121–1133
43. Schafer FQ, Buettner GR (2001) Redox environment of the cellas viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med 30:1191–1212
44. Sigola LB, Zinyama RB (2000) Adrenaline inhibits macro-phage nitric oxide production through beta1 and beta2 adren-ergic receptors. Immunology 100:359–363
45. Silveira EM, Rodrigues MF, Krause MS, Vianna DR,Almeida BS, Rossato JS, Oliveira LP Jr, Curi R, deBittencourt PI Jr (2007) Acute exercise stimulates mac-rophage function: possible role of NF-kappaB pathways.Cell Biochem Funct 25:63–73
46. Sodergren E, Nourooz-Zadeh J, Berglund L, Vessby B (1998)Re-evaluation of the ferrous oxidation in xylenol orange assayfor the measurement of plasma lipid hydroperoxides. JBiochem Biophys Methods 37:137–146
47. Straub RH, Schaller T, Miller LE, von Horsten S, Jessop DS,FalkW, Scholmerich J (2000) Neuropeptide Y cotransmissionwith norepinephrine in the sympathetic nerve-macrophageinterplay. J Neurochem 75:2464–2471
48. Ye RD (2000) beta-Adrenergic agonists regulate NF-kappaBactivation through multiple mechanisms. Am J Physiol LungCell Mol Physiol 279:L615–L617
49. Zhong H, SuYang H, Erdjument-Bromage H, Tempst P,Ghosh S (1997) The transcriptional activity of NF-kappaB isregulated by the IkappaB-associated PKAc subunit through acyclic AMP-independent mechanism. Cell 89:413–424
50. Zhou J, An H, Xu H, Liu S, Cao X (2005) Heat shock up-regulates expression of Toll-like receptor-2 and Toll-likereceptor-4 in human monocytes via p38 kinase signal path-way. Immunology 114:522–530
J. Silva Rossato et al.