role of the n-terminal region in g protein coupled...
TRANSCRIPT

1521-0111/85/1/127–138$25.00 http://dx.doi.org/10.1124/mol.113.089086MOLECULAR PHARMACOLOGY Mol Pharmacol 85:127–138, January 2014Copyright ª 2013 by The American Society for Pharmacology and Experimental Therapeutics
Role of the N-Terminal Region in G Protein–Coupled ReceptorFunctions: Negative Modulation Revealed by 5-HT2BReceptor Polymorphisms s
Arnauld Belmer, Stephane Doly, Vincent Setola,1 Sophie M. Banas, Imane Moutkine,Katia Boutourlinsky, Terry Kenakin, and Luc MaroteauxInstitut du Fer à Moulin, Université Pierre et Marie Curie, Institut National de la Santé et de la Recherche Médicale Unité Mixte deRecherche S-839, Paris, France (A.B., S.D., V.S., S.M.B., I.M., K.B., L.M.); and Department of Pharmacology, School ofMedicine, University of North Carolina, Chapel Hill, North Carolina (T.K.)
Received August 9, 2013; accepted October 30, 2013
ABSTRACTThe putative role of the N-terminal region of rhodopsin-like 7transmembrane biogenic amine receptors in agonist-inducedsignaling has not yet been clarified despite recent advances in7 transmembrane receptor structural biology. Given the exis-tence of N-terminal nonsynonymous polymorphisms (R6G;E42G) within the HTR2B gene in a drug-abusing population, weassessed whether these polymorphisms affect 5-hydroxytryp-tamine 2B (5-HT2B) receptor in vitro pharmacologic and couplingproperties in transfected COS-7 cells. Modification of the5-HT2B receptor N terminus by the R6G;E42G polymorphismsincreases such agonist signaling pathways as inositol phos-phate accumulation as assessed by either classic or operationalmodels. The N-terminal R6G;E42G mutations of the 5-HT2Breceptor also increase cell proliferation and slow its desensiti-zation kinetics compared with the wild-type receptor, further
supporting a role for the N terminus in transduction efficacy.Furthermore, by coexpressing a tethered wild-type 5-HT2B re-ceptor N terminus with a 5-HT2B receptor bearing a N-terminaldeletion, we were able to restore original coupling. This rever-sion to normal activity of a truncated 5-HT2B receptor by coex-pression of the membrane-tethered wild-type 5-HT2B receptorN terminus was not observed using a membrane-tethered 5-HT2Breceptor R6G;E42G N terminus. These data suggest that theN terminus exerts a negative control over basal as well as agonist-stimulated receptor activity that is lost in the R6G;E42G mutant.Our findings reveal a new and unanticipated role of the 5-HT2Breceptor N terminus as a negative modulator, affecting bothconstitutive and agonist-stimulated activity. Moreover, our datacaution against excluding the N terminus and extracellular loopsin structural studies of this 7 transmembrane receptor family.
IntroductionThe roles of the N-terminal region of 7 transmembrane
receptors (7TMRs) in receptor functions, such as ligandbinding, surface expression, signaling, and desensitization,vary depending on the size and structure of the N terminus.
It is well established that in some 7TMRs, the N terminusparticipates in cognate agonist binding and activation.These 7TMRs include secretin-like (family 2), metabotropicglutamate-like (family 3), and the rhodopsin-like (family 1)7TMRs that bind peptides. Within this latter family, theN-terminal domain interaction with the core of the receptorhas been associated with different modes of activation. Forprotease-activated receptors, cleavage of an N-terminal in-hibitor initiates receptor activation (Ramachandran andHollenberg, 2008). For glycoprotein hormone receptors,activation is dependent on the release of constitutive in-hibition upon binding of a ligand to the N terminus of thereceptor (Vassart et al., 2004). For the melanocortin 4 re-ceptor, a diffusible inverse agonist inhibits N-terminal acti-vation (Srinivasan et al., 2004; Ersoy et al., 2012). However,the N-terminal regions of the rhodopsin-like 7TMR familyinteracting with such small ligands as monoamines havebeen the least studied among the superfamily, and their
This work was supported by the Centre National de la RechercheScientifique; the Institut National de la Santé et de la Recherche Médicale;the Université Pierre et Marie Curie; by grants from the Fondation de France;the French Ministry of Research Agence Nationale pour la Recherche [ANR-12-BSV1-0015-01]; a Lefoulon-Lalande fellowship (to S.D.); a Société Française dePharmacologie Thérapeutique fellowship (to A.B.); and a Marie Curie EUfellowship (to V.S.). L.M.’s team is part of the École des Neurosciences de ParisIle-de-France network and of the Bio-Psy Labex.
A.B. and S.D. contributed equally to this work.1Current affiliation: Department of Physiology and Pharmacology, Robert C.
Byrd Health Sciences Center, School of Medicine, West Virginia University,Morgantown, West Virginia.
dx.doi.org/10.1124/mol.113.089086.s This article has supplemental material available at molpharm.
aspetjournals.org.
ABBREVIATIONS: BRET, bioluminescence resonance energy transfer; BSA, bovine serum albumin; BW 723C86, a-methyl-5-(2-thienylmethoxy)-1H-indole-3-ethanamine hydrochloride; DF, (1)-fenfluramine; DOI, (6)-2,5-dimethoxy-4-iodoamphetamine hydrochloride; ECL, extracellular loop;5-HT, serotonin or 5-hydroxytryptamine; IP, inositol phosphate; MDMA, (6)-3,4-methylenedioxymethamphetamine; NDF, nor-(1)-fenfluramine;DNter, N-terminal deletion of the first 32 amino acids; PBS, phosphate-buffered saline; PIP2, phosphatidyl inositol-bisphosphate; RA, relativeactivity ratio; SNP, single-nucleotide polymorphism; TM, transmembrane; 7TMR, 7 transmembrane receptor; Rluc, Renilla luciferase; WT, wild-type;YFP, yellow fluorescent protein.
127
http://molpharm.aspetjournals.org/content/suppl/2013/10/30/mol.113.089086.DC1Supplemental material to this article can be found at:
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

roles in agonist-induced signaling and desensitization havenot been reported.In some 7TMRs, such as the cannabinoid CB1 receptor, a1-
adrenergic receptor, and GPR37, published data show thattruncation of the N-terminal region increases plasma mem-brane expression of these receptors (Andersson et al., 2003;Hague et al., 2004; Dunham et al., 2009). The extracellularsurface of biogenic amine 7TMRs affects ligand recognition,ligand “escorting” into the binding pocket, and ligand bind-ing kinetics (Wittmann et al., 2011). We previously observedthat deletion of the long N terminus of the Drosophila 5-hydroxytryptamine (5-HT)2Dro receptor (which most closelyresembles the mammalian 5-HT2B receptor) affects its phar-macology, with some compounds exhibiting a 10-fold changein affinity (Colas et al., 1997). This early finding suggestedthat the N terminus affects the conformation of the receptortransmembrane (TM) regions involved in ligand interactions,which may be relevant for receptor structure, because mod-elers usually exclude (often for technical reasons) 7TMR Ntermini, including the recently published crystal structuresfor 5-HT2B receptors (Wacker et al., 2013). These observationssuggested that the N-terminal regions of 7TMRsmodulate theactivity of biogenic amine 7TMRs by as-yet-uncharacterizedmechanisms.Drug abuse vulnerability is a complex trait with strong
genetic influences. Understanding the genetic bases of drugabuse vulnerability and particular allelic variants that con-tribute to this vulnerability can strongly improve our under-standing of human addictions. We have recently extended therole of serotonin (5-HT) in the behavioral responses to psycho-stimulants by revealing that the genetic ablation of Htr2B inmice, the gene encoding 5-HT2B receptors, eliminates locomotorresponse and place preference induced by MDMA [(6)-3,4-methylenedioxymethamphetamine] (Doly et al., 2008, 2009).Independently, three novel single nucleotide polymorphisms,two of which result in a point mutation, were described ina drug-abusing population (Lin et al., 2004). Arg6, a conservedbasic residue, and the conserved acidic Glu42 are mutatedsimultaneously into Gly, termed R6G;E42G.Starting from these observations, we established whether
these polymorphisms within the 5-HT2B receptor N terminusaffect receptor pharmacology. We discovered a new and un-anticipated function of this extracellular region, and that itacts as a negative structural modulator of receptor basal andstimulated activity.
Materials and MethodsPlasmid Constructs. Human 5-HT2B receptor cDNA was subcl-
oned into the p513 vector, a derivative of the pSG5 mammalianexpression vector (Green et al., 1988), which replicates in SV40T-antigen–transformed cells and drives 5-HT2B receptor expressionunder the control of the SV40 early promoter. N-terminal variant5-HT2B receptorswere generated using the Quikchange II (Stratagene,La Jolla, CA) site-directed mutagenesis kit according to themanufacturer’s protocol. The N-terminal truncated 5-HT2B receptormutant was generated by polymerase chain reaction deletionmutagenesis. TM1 constructs carrying different N-terminal sequen-ces were obtained from each full construct by removing a PstI-PstIfragment (from AA85 to the C-terminal end of the 5-HT2B receptorsequence).
TM1-tethered (wild-type [WT], R6G;E42G, or N-terminal deletionof the first 32 amino acids [DNter]) and DNter 5-HT2B receptor coding
regions were amplified from their respective cDNAs using appropri-ate sense and antisense primers. The fragments were then subclonedin frame in either a plasmid encoding C terminus yellow fluorescentprotein (YFP) (Clontech/BD Biosciences, Mountain View, CA) orRenilla luciferase (Rluc). The coding regions of all constructs wereentirely sequenced. The Rluc construct was a kind gift from StefanoMarullo (U1016; Institut Cochin, Paris, France).
Cell Culture. COS-7 cells were cultured asmonolayers inDulbecco’smodified Eagle’s medium (GIBCO/Invitrogen, Carlsbad, CA) supple-mented with 10% fetal calf serum (Biowest SAS, Nuaillé, France)and 1% penicillin/streptomycin (Sigma-Aldrich, St. Louis, MO), in9-cm dishes (Falcon/Corning Life Sciences, Tewksbury, MA). Cellswere incubated at 37°C in a 5% CO2 atmosphere. Cells were 80%confluent when transfected with 10 mg of DNA using Nanofectin(PAA Laboratories GmBH, Pasching, Austria), according to themanufacturer’s protocol, in an antibiotic-free medium. Four hourslater, the medium was replaced with fresh medium. Twenty-fourhours after transfection, cells were incubated in serum-free mediumfor membrane radioligand binding or trypsinized (Trypsin 1X 0.05%EDTA; Invitrogen) and plated onto 24-well plates for inositolphosphate (IP) accumulation and whole-cell radioligand binding.
[3H]Thymidine Incorporation Assay. Proliferation as mea-sured by incorporation of radioactive desoxythymidine incorporationwas performed, as previously described elsewhere (Deraet et al.,2005).
[3H]Radioligands and Drugs. [3H]Myo-inositol (51.0 Ci/mmol),[3H]mesulergine (99 Ci/mmol), [3H]5-HT (80.0 Ci/mmol), 125I-DOI[125I-(6)-2,5-dimethoxy-4-iodoamphetamine hydrochloride] were ob-tained from PerkinElmer Life and Analytical Sciences (Waltham,MA). The 5-HT, BW 723C86 [a-methyl-5-(2-thienylmethoxy)-1H-indole-3-ethanamine hydrochloride], DOI, and nor-(1)-fenfluramine(NDF) were obtained from Sigma-Aldrich. The (1)-fenfluramine (DF)came from Tocris Bioscience (Bristol, UK). The mesulergine wasobtained from RBI (Natick, MA), and MDMA from Lipomed AG(Arlesheim, Switzerland).
[3H]Inositol Phosphate Accumulation Assay. Twenty-fourhours before the experiment, cells were incubated in 24-well platesovernight with 20 nM of [3H]myo-inositol diluted in an inositol-freemedium (BME, Lonza, Basel, Switzerland). Just before receptorstimulation, medium was replaced by Krebs-Ringer-Hepes buffer(130 mMNaCl, 1.3 mMKCl, 2.2 mMCaCl2, 1.2 mMNaH2PO4, 1.2 mMMgSO4, 10 mM Hepes, 10 mM glucose, pH 7.4) supplemented with20mMLiCl to prevent IP1 degradation. Cellswere stimulated in duplicatein a final volume of 500 ml for 2 hours. The experiment was stopped byreplacing the stimulation medium with 1023 M formic acid at roomtemperature for 20 minutes, and at 4°C overnight. Thus, IP1 accumu-lated from IP3 and IP2 hydrolysis was released from lysed and fixedcells. The accumulated IP1 was eluted on anion exchange columns(Bio-Rad AG-1X8; Bio-Rad Laboratories, Hercules, CA) with 0.2 Mammonium formate in 0.1 M formic acid. Scintillation cocktail(Ultima Gold XR; PerkinElmer) was added to the eluted [3H]IPsample, and radioactivity was counted in a Beckman Coulter (Brea,CA) scintillation counter. At least three independent experimentswere performed in duplicate, or in triplicate for tethered N terminusexperiments.
Desensitization Assay. To evaluate receptor desensitization, IPaccumulation assays were performed as described earlier, except thata first agonist stimulation, in the absence of LiCl (to avoid IPaccumulation), occurred at different time points (15, 30, 45, 60, 90,and 120 minutes) before a second stimulation with 100 nM of DOI.After the first stimulation, the medium was removed, the cells werewashed 3 times with fresh medium, and fresh medium containingLiCl 1 DOI (100 nM) was added. The end of the experiment wasperformed as previously described, and data were also normalized byeach control condition, that is, without the first stimulation (time:0 minutes). At least three independent experiments were performedin duplicate.
128 Belmer et al.
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

Membrane Radioligand Binding Assay. Membrane bindingassays were performed on transfected cells plated in 9-cm dishes.Cells were first washed with phosphate-buffered saline (PBS) andscraped into 10 ml of PBS on ice, then centrifuged for 5 minutes at1,000 g. Cell pellets were dissociated and lysed in 2 ml of bindingbuffer (50 mM Tris HCl, 10 mM MgCl2, 0.1 mM EDTA, pH 7.4) andcentrifuged for 30 minutes at 10,000g. Membrane preparations werethen resuspended in binding buffer to obtain a final concentration of0.2–0.4 mg of protein/well. Aliquots of membrane suspension (200 ml/well) were incubated with 25 ml/well of [3H]radioligand at a finalconcentration between half theKD and theKD for the 5-HT2B receptor,diluted in binding buffer, and 25 ml/well of increasing concen-trations of homologous compound (from 10211 to 1026 M, diluted inbinding buffer) in 96-well plates for 90 minutes at room temperature.Membranes were harvested by rapid filtration onto Whatman GF/Bglass fiber filters (Brandel, Gaitherburg, MD) presoaked with coldsaline solution and washed 3 times with cold saline solution to reducenonspecific binding. Filters were placed in 6-ml scintillation vials andallowed to dry overnight. The next day, 4 ml of scintillation cocktailwere added to the samples, which were counted as before. Data indisintegrations per minute were converted to fmoles and normalizedto protein content (ranging from 0.1 to 0.5 mg/well). Specific bindingrepresented 61.7% 6 6.2% of maximal binding for heterologous and52.0% 6 3.8% for homologous competition of the maximal binding, asdetermined in the presence of an excess of cold competitor. At leastthree independent experiments were performed in duplicate (seeSupplemental Fig. 1).
Nonpermeabilized Whole-Cell Radioligand Binding Assay.Cells were plated in 24-well clusters. Twenty-four hours before theexperiment, the cells were incubated in serum-free medium over-night. The next day, the medium was replaced by 400 ml/well ofKrebs-Ringer-Hepes buffer (130 mM NaCl, 1.3 mM KCl, 2.2 mMCaCl2, 1.2 mM NaH2PO4, 1.2 mM MgSO4, 10 mM Hepes, 10 mMglucose, pH 7.4). Then, 50 ml of tritium-labeled compounds were di-luted in Krebs-Ringer-Hepes buffer at a final concentration betweenhalf the KD and the KD for the 5-HT2B receptor. The tritiated radio-ligand was competed with 50 ml of increasing concentrations of non-radioactive ligand, also diluted in Krebs-Ringer-Hepes buffer. Cellswere then incubated for 30 minutes (5-HT2B receptor) at room temper-ature and then washed twice on ice with cold PBS. Washed cells weresolubilized by the addition of 500 ml of SDS 1%. The next day, 4 ml ofscintillation cocktail were added to the samples, and the radioactivitywas counted using a scintillation counter (Beckman Coulter). Data indisintegrations per minute were converted to femtomoles and normal-ized to protein content (0.2–0.4 mg of protein/well). Specific bindingrepresented 44.2% 6 2.1% for heterologous and 44.5% 6 5.3% for ho-mologous competition of the maximal binding, as determined in thepresence of an excess of cold competitor. At least three independentexperiments were performed in duplicate (see Supplemental Fig. 1).
Immunocytochemistry. COS-7 cells plated on coverslips aftertransfection with either the WT TM1-N terminus (TM1-WT), TM1-R6G;E42G, or TM1-DNter constructs fused to YFP together with5-HT2B receptor DNter were incubated overnight in serum-freeDulbecco’sModified Eagle’sMedium before the immunocytochemistryexperiments. Then, cells were washed with 3� cold PBS and fixed for15 minutes with 4% paraformaldehyde. After three washes with PBS,cells were permeabilized with PBS 0.25% triton and then incubated atroom temperature for 20 minutes in PBS 1 bovine serum albumin(BSA) 3%, cells were incubated at room temperature for 4 hours withan antibody against the C terminus of the 5-HT2B receptor (1:1000).After three washes with PBS 1 BSA (3%), the primary antibody wasrevealed by a 1-hour incubation with a donkey anti-mouse antibodycoupled to CY3 (1:1000). After three washes, coverslips were mountedwith Mowiol.
Similar experiments were performed after transfection with TM1-N terminus (TM1-WT) alone. After three washes with PBS1 BSA 3%without permeabilization, cells were incubated at room temperaturefor 4 hour with a rabbit antibody against the N terminus of the 5-HT2B
receptor (1:1000) (Pharmingen, San Diego, CA). After three washeswith PBS1 BSA (3%), the primary antibody was revealed by a 1-hourincubation with a goat-anti-rabbit antibody coupled to AlexaFluor 488(1:5000). After three washes, coverslips were mounted with Mowiol.
For coimmunoprecipitation experiments, FLAG-tagged WT 5-HT2Band YFP-tagged TM1 constructs were transfected in COS-7 cells. After48 hours, the cells were washed in PBS, sonicated, and solubilized inlysis buffer (75 mMTris, 2 mMEDTA, 12 mMMgCl2, 10 mMCHAPS,protease inhibitor cocktail EDTA free, pH 7.4) during 5 hours at 4°C.Lysates were centrifuged at 12,000g during 30 minutes at 4°C. Immu-noprecipitations were performed using EZview Red FLAG M2 AffinityGel (Sigma-Aldrich) according to themanufacturer’s recommendations.Immunoprecipitated proteins and 50 mg of total proteins were com-bined with Laemmli buffer, heated at 70°C for 10 minutes, and run onNuPAGE 4–12% Bis-Tris gel (Life Technologies). Immunoblots wereprobed with anti-gfp (Roche, Basel, Switzerland) or anti-flag (Sigma-Aldrich) antibodies diluted 1/2000 and immunoreactivity was revealedusing secondary antibody coupled to 680 nm fluorophores using theOdyssey LI-COR infrared fluorescent scanner (Li-Cor Biosciences,Lincoln, NE).
Bioluminescence Resonance Energy Transfer Assays. Bio-luminescence resonance energy transfer (BRET) assays were per-formed according to published methods (Achour et al., 2011). Briefly,COS-7 cells (5 � 105 per well of a 6-well plates) were transfected with30-ng plasmid DNA coding for the BRET donor (DNter 5-HT2B-Luc)and increasing amounts of BRET acceptor plasmids (TM1-tetheredWT, R6G;E42G or DN-ter-YFP; 100–4000 ng per well) or YFP. Twenty-four hours after transfection, cells were washed in PBS, detached using10 mM EDTA in PBS, centrifuged (1400g for 5 minutes), resuspendedin Hanks’ balanced salt solution, and distributed in 96-well plates(PerkinElmer plates; 105 cells per well). After addition of the luciferasesubstrate, coelenterazine h (5 mM final concentration), luminescence,and fluorescence were measured simultaneously (at 485 and 530 nm,respectively) in a Mithras LB940 plate reader (Berthold Technologies,Bad Wildbad, Germany).
The BRET ratio was calculated as: ([Emission at 530 nm/Emissionat 485 nm] 2 [Background at 530 nm/Background at 485 nm]), wherebackground corresponds to signals in cells expressing the Rluc fusionprotein alone under the same experimental conditions. For betterreadability, results were expressed in milli-BRET units (mBRET),where 1 mBRET corresponded to the BRET ratio multiplied by 1000.BRET ratios were plotted as a function of ([YFP 2 YFP0]/YFP0)/(Rluc/Rluc0), where YFP is the fluorescence signal at 530 nm afterexcitation at 485 nm, and Rluc is the signal at 485 nm after addition ofcoelenterazine h. YFP0 and Rluc0 correspond to the same values incells expressing the Rluc fusion protein alone.
Data Analysis. Binding data were analyzed using the iterativenonlinear regression model (GraphPad Prism 6.0; GraphPad Soft-ware, Inc., San Diego, CA). This allowed the calculation of inhibitionconstants (Ki) and the maximal number of sites (Bmax). All valuesrepresent the average of independent experiments 6 S.E.M. (n 5number of experiments as indicated in the text). For IP couplingexperiments, data obtained in disintegrations per minute wereanalyzed using GraphPad Prism, converted to femtomoles, andnormalized to protein content and receptor Bmax (obtained in whole-cell radioligand binding experiments of the same transfected cells). Asingle dose-response experiment may not accurately determineaffinity and efficacy because a drug that binds with high affinity buthas low efficacy will produce the same dose-response curve as a drugwith low affinity and high efficacy.
To untangle affinity from efficacy, we used the operational modelinitially described by (Black and Leff, 1983), which defines responseas:
Response A5 ½A�ntnEm=½A�ntn 1 ð½A�1KAÞn (1)
where Em represents the maximal response capability of the system,KA is the equilibrium dissociation constant of the agonist–receptor
N Terminus Structural Modulation of 5-HT2B Receptor 129
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

complex, n is the slope of the dose–response curve, and t is the ef-ficacy. This final term represents the receptor density and molecularinteraction of the agonist– receptor complex interaction with thestimulus–response system of the cell, which itself has ligand-specificelements (efficacy of the ligand) and system-specific elements (ef-ficiency of coupling of receptors to signaling pathway). A theoreticallycomplete term to describe the power of a ligand to active a cellularpathway is t/KA, which incorporates both elements of efficacy andaffinity. Considering that the most common difference between sys-tems is receptor density, ratios of t/KA account for these and should besystem-independent measures of the relative power of ligands toactivate pathways.
Another useful measure of agonist activity is Ehlert’s activity ratio(denoted RA, or relative activity ratio). Both affinity and efficacy fora single number of agonism for dose–response curves of unit slopehave been solved with the application of RA ratios. Proposed by Ehlertand colleagues (Ehlert, 2005), the RA value is themaximal response ofthe agonist divided by the EC50. This furnishes a single numberreflecting the power of a molecule to produce agonism in any system.In terms of the maximal response and EC50 for dose–response curvesin the Black Leff operational model (Black and Leff, 1983), it can beshown that RA represents the relation:
RA5 tn�ð21 tnÞ1=n 21
�Em=KAð11 tnÞ (2)
For dose–response curves of unit slope (n 5 1), it can be seen that RAreduces to the term Em(t/KA). Ratios of these terms for particularagonists cancel the Em term and are therefore system independent.
Values of t/KA have the added advantage of circumventing anotherissue with full agonists, namely, that an infinite variety of combina-tions of efficacy and affinity can describe any given curve to a fullagonist (there is no unique value of efficacy that can be identifiedunless the curve indicates partial agonism). The Δlog(RA) values were
calculated as: Δlog(RA)agonist/reference 5 log(RA)agonist 2 log(RA)reference to compare various agonists within a signaling pathway.The Δlog(RA) values were calculated as: Δlog(RA)R6G;E42G/wildtype5 Δlog(RA)R6G;E42G2 Δlog(RA)wild type. The ΔΔlog(RA) valuesprovided a scale to compare partial agonist (DOI) and reference fullagonist (5-HT) between the WT and mutant R6G;E42G receptors. Thebias (10ΔΔlog(RA)) represents the signaling difference between the wild-type and mutant receptor for the specific coupling pathway, here the IPcoupling, for DOI relative to 5-HT. The calculations of 95% confidenceintervals were made with the program Mathematica 5.0 (Wolfram,Champaign, IL) (Kenakin et al., 2012).
Statistics. Comparisons between groups were performed withStudent’s unpaired t test or one-way or two-way analysis of variancewith Bonferroni’s post hoc test, depending on the experiment. P ,0.05 was considered statistically significant.
ResultsThe N Terminus of the 5-HT2B Receptor Affects
Agonist Affinity. Starting from the report by Lin et al.(2004), we explored the pharmacologic properties of the R6G;E42G variant 5-HT2B receptor, which contains two amino acidchanges in the N-terminal region (Fig. 1). To study the effectof the N-terminal mutations, we compared WT and variant5-HT2B receptor–transfected COS-7 cells using radioligandbinding assays. Homologous competition experiments showeda left shift in the displacement curve for 5-HT binding (Fig.2A). The 5-HT2B variant receptor had a 5-fold increase inaffinity for 5-HT—that is, a 5-fold decrease in the Ki for theR6G;E42G (P , 0.05) variant relative to the WT receptor(Table 1). Using the 5-HT2B/2C receptor–selective agonist
Fig. 1. Molecular description of the 5-HT2B receptor. Amino acid sequence and localization of the variant R6G;E42G in the N terminus of 5-HT2Breceptor. The known 5-HT binding site residues are presented in dark gray circles with black letters. The R6G;E42G variant substitution position ispresented in red circles with white letters, and the R6E;E42R inverted variant is shown in light blue with black letters on the N-terminal sequence. Adouble arrowhead locates the DNter.
130 Belmer et al.
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

BW723C86 as a competitor, a 4-fold increase in affinity wasfound compared with the WT receptor (Table 1).A similar effect was observed using the 5-HT2 agonist DOI
as a competitor, which displayed a 2.4-fold increase in affinityfor the N-terminal variant receptor compared with the WT.Because none of the tested antagonists were affected by thesingle-nucleotide polymorphisms (SNPs) (Fig. 2B; Table 1),it was likely that the mutations affect receptor coupling.However, the fact that only some of the tested agonists areaffected by this variant may be related to a probe-dependenceissue, agonism being a combined interaction of the modulator(agonist), receptor, and signaling protein, known to be probedependent. Nevertheless, this gain of affinity seems indepen-dent of the type of agonism, full agonists being not differ-entially affected compared with partial agonists (Table 1)(Cussac et al., 2008).In light of the finding that N-terminal deletion of the
5-HT2Dro affected the affinity of some agonists (Colas et al.,1997), we tested the effect of DNter (Fig. 1). Interestingly, thisN-terminal truncation generated an apparent shift in agonistaffinity, with a 30-fold increase in affinity for 5-HT (Fig. 2A).Because the R6G;E42G variant results in neutralization oftwo oppositely charged amino acid side chains, we hypothe-sized that these residues interacted electrostatically. Basedon this, we tested whether exchanging the position of thecharged residues would alter 5-HT2B receptor ligand affinity
vis-à-vis the WT receptor. Thus, we generated the “chargeswap” mutant R6E;E42R. This inversion also increased itsapparent affinity for 5-HT (Fig. 2A), indicating that a missingelectrostatic interaction in the R6G;E42G variant likelyaffects its pharmacologic properties.The N Terminus of the 5-HT2B Receptor CanModulate
Receptor-Gaq Activation. We then explored the impact ofN-terminal modifications on an immediate effector of 5-HT2B
receptors, the Gaq/11–sensitive membrane phospholipase C.Upon agonist binding, the 5-HT2B receptor stimulates hydro-lysis of membrane phosphatidyl inositol-bisphosphate (PIP2)into cytoplasmic inositol phosphates (IP3, which promotesintracellular calcium release, and the degradation products IP2
and IP1) and diacylglycerol (an activator of protein kinase C)(Deraet et al., 2005). Because the N-terminal mutations mayaffect cell surface expression, we evaluated the cell surface ex-pression as receptor Bmax using tritiated mesulergine compe-tition comparing purified membrane to nonpermeabilizedwhole-cell binding (Supplemental Fig. 1). Only a fraction ofthe total membrane binding capacity was found on whole cells,supporting the rationale to used whole cell Bmax as an estimateof surface receptor to normalize coupling efficiency.Integrating these Bmax values to normalize for surface
receptor expression, increasing concentrations of a partial5-HT2 agonist, DOI, activated the R6G;E42G variant moreefficaciously than theWT receptor, leading to an increase of both
Fig. 2. Pharmacologic characterization of the R6G;E42G variant of human 5-HT2B receptor. (A) Representative example of [3H]5-HT radioligandbinding homologous competition experiments performed on membrane preparation of COS-7 cells transfected with plasmids encoding the WT, the R6G;E42G, the R6E;E42R, and the DNter receptors showing a left shift in the DNter, the R6E;E42R, and the R6G;E42G variant receptors curve (Ki of 3.3460.76, 1.08 6 0.60, 2.34 6 0.79, 0.107 6 0.016 nM, Bmax of 560 6 30, 240 6 70, 420 6 75, 120 6 25 fmoles/mg prot for WT, R6G;E42G, R6E;E42R, andDNter, respectively). Representative result of five independent experiments each performed in triplicate (see Table 1 for compete results). (B)Homologous antagonist competition binding (mesulergine) shows no difference between cells transfected with WT, R6G;E42G, or DNter 5-HT2Breceptors. (C) Dose-response of agonist-induced IP accumulation of COS-7 cells transfected with plasmids encoding theWT, DNter, R6G;E42G, and R6E;E42R receptors stimulated for 2 hours with increasing concentrations of DOI (from 1029 to 1025M). The calculated EC50 revealed very little impact of thedifferent mutations on apparent coupling efficiency (31 6 11, 18 6 12, 37 6 10, 34 6 7 nM for WT, DNter, R6E;E42R, and R6G;E42G, respectively).Representative results of at least three independent experiments each performed in duplicate, normalized in fmoles of IP per fmoles of surface receptors;that is, corrected by whole cells Bmax and statistically analyzed by two-way analysis of variance repeated measures with mutation and drugconcentration as main factors; effect of mutations on IP accumulation, F(3,24) = 148.35 P , 0.0001. A Bonferroni post tests was also applied on eachgraph: **P , 0.01; ****P , 0.0001 versus WT receptor.
N Terminus Structural Modulation of 5-HT2B Receptor 131
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from
minimal (Emin 1 48%) and maximal (Emax 1 56%) cytoplasmicIP accumulation as well as activation efficiency (Emax-Emin) withlittle effect on apparent agonist potency EC50 (Fig. 2C). TheDNter also generated an increase in IP basal activity (DNter68%). DOI also induced a stronger 5-HT2B receptor–mediated IPaccumulation, as did basal 5-HT2B receptor–mediated PIP2
hydrolysis, with a noticeable increase for the DNter mutant(DNter 97%), while the inverse mutant displayed a moderateincrease in Emax (15%) (Fig. 2C). This basal activity was testedagainst ritanserin, a known inverse agonist for the receptor,which was able to reverse this basal activity to similar extent(Supplemental Fig. 2A).Other tested agonists followed the same trend as DOI,
activating the R6G;E42G variant receptor more efficaciouslythan the WT receptor (125% for 5-HT, P , 0.05; 117% for DF,P, 0.01;120% forNDF,P, 0.05;121% for DOI,P, 0.001; and130% for MDMA, P , 0.05) (Supplemental Fig. 2, B–E). Takentogether, these results indicate that the human R6G;E42Gpolymorphism leads to increased coupling of the 5-HT2B receptorto phospholipase C activity. Notably, the IP responses for theR6G;E42G variant were significantly increased for DOI as
well as for other agonists including MDMA, DF, and itsmetabolite, NDF.Validation Using the Operational Model of the Mod-
ulation by the N Terminus of the 5-HT2B Receptor-Gaq
Activation. To take into account putative alteration of theefficiency of receptor coupling, which may vary among themutants, we used the Black-Leff operational model to deriveEhlert’s activity ratio (denoted RA) (Ehlert, 2005), the max-imal response of the agonist divided by the EC50, and tocalculate the putative difference in signaling character-istics for IP production for each mutant as ΔΔLog(RA)(Black and Leff, 1983; Ehlert, 2005; Kenakin et al., 2012).Using ΔΔLog(RA) values and comparing two agonist probes(5-HT as full agonist and DOI as partial agonist) for eachreceptor species cancels all effects of the measurement—expression level, assay coupling, and so on—simply revealingdifferences in the way the two probes (agonists) interact withthe receptor conformation. Differences in Log(RA) values areinformative about the intrinsic differences in the receptorconformation (see “Data Analysis”; Fig. 3, A–C; SupplementalFig. 3).
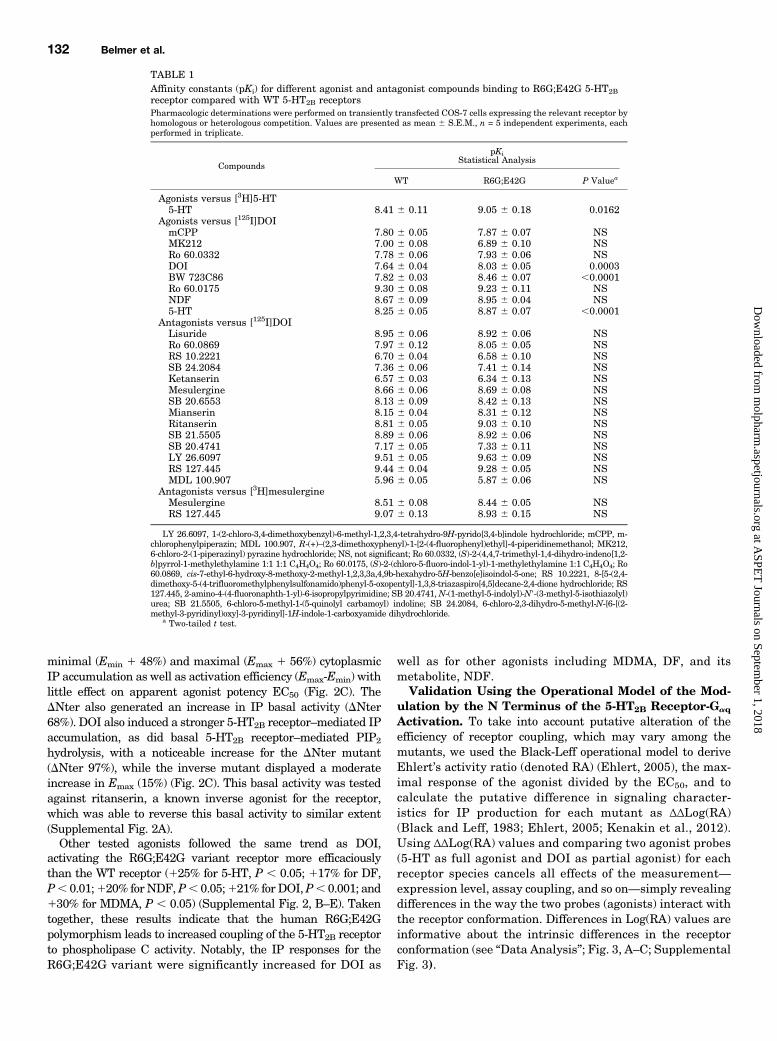
TABLE 1Affinity constants (pKi) for different agonist and antagonist compounds binding to R6G;E42G 5-HT2Breceptor compared with WT 5-HT2B receptorsPharmacologic determinations were performed on transiently transfected COS-7 cells expressing the relevant receptor byhomologous or heterologous competition. Values are presented as mean 6 S.E.M., n = 5 independent experiments, eachperformed in triplicate.
Compounds
pKiStatistical Analysis
WT R6G;E42G P Valuea
Agonists versus [3H]5-HT5-HT 8.41 6 0.11 9.05 6 0.18 0.0162
Agonists versus [125I]DOImCPP 7.80 6 0.05 7.87 6 0.07 NSMK212 7.00 6 0.08 6.89 6 0.10 NSRo 60.0332 7.78 6 0.06 7.93 6 0.06 NSDOI 7.64 6 0.04 8.03 6 0.05 0.0003BW 723C86 7.82 6 0.03 8.46 6 0.07 ,0.0001Ro 60.0175 9.30 6 0.08 9.23 6 0.11 NSNDF 8.67 6 0.09 8.95 6 0.04 NS5-HT 8.25 6 0.05 8.87 6 0.07 ,0.0001
Antagonists versus [125I]DOILisuride 8.95 6 0.06 8.92 6 0.06 NSRo 60.0869 7.97 6 0.12 8.05 6 0.05 NSRS 10.2221 6.70 6 0.04 6.58 6 0.10 NSSB 24.2084 7.36 6 0.06 7.41 6 0.14 NSKetanserin 6.57 6 0.03 6.34 6 0.13 NSMesulergine 8.66 6 0.06 8.69 6 0.08 NSSB 20.6553 8.13 6 0.09 8.42 6 0.13 NSMianserin 8.15 6 0.04 8.31 6 0.12 NSRitanserin 8.81 6 0.05 9.03 6 0.10 NSSB 21.5505 8.89 6 0.06 8.92 6 0.06 NSSB 20.4741 7.17 6 0.05 7.33 6 0.11 NSLY 26.6097 9.51 6 0.05 9.63 6 0.09 NSRS 127.445 9.44 6 0.04 9.28 6 0.05 NSMDL 100.907 5.96 6 0.05 5.87 6 0.06 NS
Antagonists versus [3H]mesulergineMesulergine 8.51 6 0.08 8.44 6 0.05 NSRS 127.445 9.07 6 0.13 8.93 6 0.15 NS
LY 26.6097, 1-(2-chloro-3,4-dimethoxybenzyl)-6-methyl-1,2,3,4-tetrahydro-9H-pyrido[3,4-b]indole hydrochloride; mCPP, m-chlorophenylpiperazin; MDL 100.907, R-(+)–(2,3-dimethoxyphenyl)-1-[2-(4-fluorophenyl)ethyl]-4-piperidinemethanol; MK212,6-chloro-2-(1-piperazinyl) pyrazine hydrochloride; NS, not significant; Ro 60.0332, (S)-2-(4,4,7-trimethyl-1,4-dihydro-indeno[1,2-b]pyrrol-1-methylethylamine 1:1 1:1 C4H4O4; Ro 60.0175, (S)-2-(chloro-5-fluoro-indol-1-yl)-1-methylethylamine 1:1 C4H4O4; Ro60.0869, cis-7-ethyl-6-hydroxy-8-methoxy-2-methyl-1,2,3,3a,4,9b-hexahydro-5H-benzo[e]isoindol-5-one; RS 10.2221, 8-[5-(2,4-dimethoxy-5-(4-trifluoromethylphenylsulfonamido)phenyl-5-oxopentyl]-1,3,8-triazaspiro[4,5]decane-2,4-dione hydrochloride; RS127.445, 2-amino-4-(4-fluoronaphth-1-yl)-6-isopropylpyrimidine; SB 20.4741,N-(1-methyl-5-indolyl)-N9-(3-methyl-5-isothiazolyl)urea; SB 21.5505, 6-chloro-5-methyl-1-(5-quinolyl carbamoyl) indoline; SB 24.2084, 6-chloro-2,3-dihydro-5-methyl-N-[6-[(2-methyl-3-pyridinyl)oxy]-3-pyridinyl]-1H-indole-1-carboxyamide dihydrochloride.
a Two-tailed t test.
132 Belmer et al.
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

The data show that the R6G;E42G and R6E;E42Rmutationsproduced significant differences in the relative way 5-HT andDOI bind to and active the receptor. These show statisticallysignificant differences between the relative activity of 5-HT andDOI produced by the R6G;E42G mutation (i.e., DOI was moreactive when compared with 5-HT) (Fig. 3C). The R6E;E42Rmutation (DOI was relatively less active when compared with5-HT) and the ΔNtermutation produced little effect in the way5-HT and DOI interact (or at the least, the changes induced bythe mutation are the same for both probes) (SupplementalFig. 3).So far, the results obtained by two independent approaches
support a role for the N terminus variant R6G;E42G incontrolling the efficacy and affinity of DOI-stimulated 5-HT2B
receptor-mediated PIP2 hydrolysis, suggesting that the twoamino acid substitutions encoded by this polymorphism aresufficient to modify N-terminal function over the receptor.Validation of N-Terminal Domain Interactions. Fur-
ther evidence for the involvement of the 5-HT2B N terminus inreceptor function was obtained using dual genetic constructs:the first TM domain of the 5-HT2B receptor (TM1) was used asa membrane tether for various N-terminal sequences and wascotransfected with N-terminally truncated receptors (Fig. 4,A–I) to explore putative intermolecular (in trans) effects. Theunderlying hypothesis was that expression of the TM1-tethered WT N terminus could interact and thus suppressthe effect of N-terminal truncation on receptors by directinteraction.To first validate putative interactions between TM1-
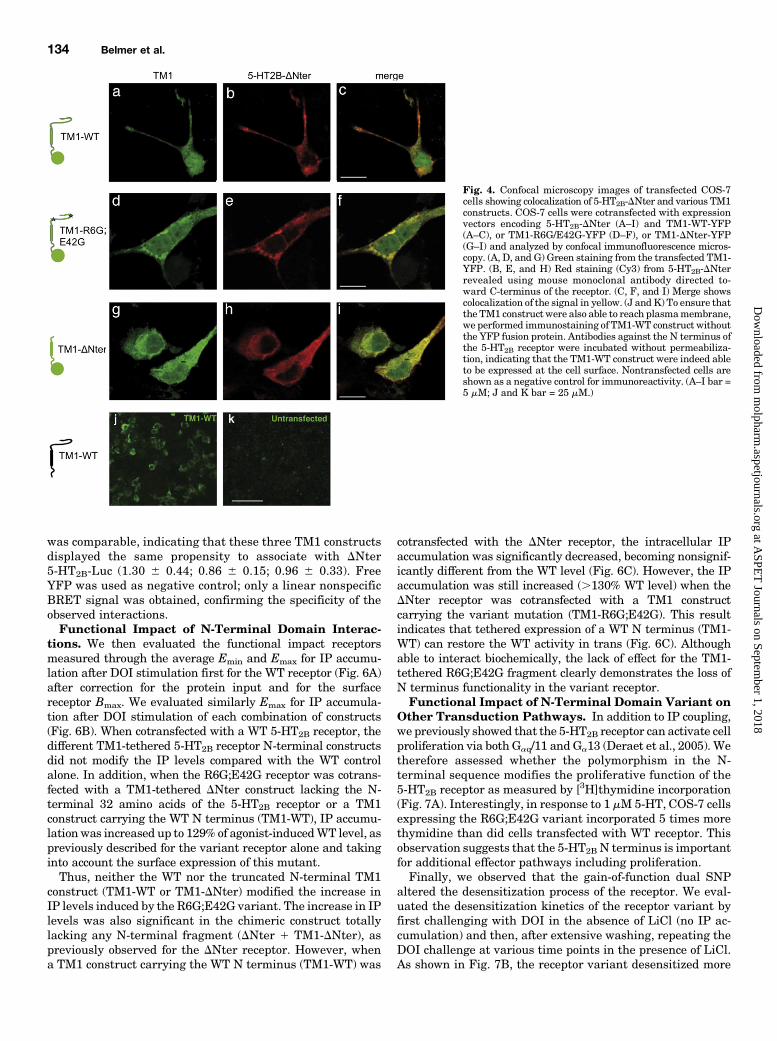
tethered construct and 7TMR, COS-7 cells were cotransfectedwith plasmids coding for ΔNter 5-HT2B receptor and TM1-tethered (WT, R6G;E42G or DNter)-YFP. Confocal analysisusing a 5-HT2B receptor antibody revealed colocalization with
the YFP for all tested TM1-tethered (WT, R6G;E42G orDNter-YFP) construct (Fig. 4, A–I). This observation supportsthe putative close localization between these different re-ceptor fragments upon coexpression. Proper membrane ex-pression of the TM1-tethered N terminus was also verified byimmunofluorescence and confocal microscopy on nonpermea-bilized cells using an antibody raised against the N terminusof the human 5-HT2B receptor (Fig. 4, J and K).To validate putative interactions between the TM1-tethered
construct and N-terminally truncated 5-HT2B receptor, COS-7cells were then cotransfected with plasmids encoding ΔNter5-HT2B receptor-FLAG and TM1-tethered (WT, R6G;E42Gor DNter)-YFP or free YFP (negative controls). Western blotanalysis of proteins immunoprecipitated with anti-FLAGantibody and probed with green fluorescent protein antise-rum indicated that all tested TM1-tethered (WT, R6G;E42Gor DNter)-YFP constructs coimmunoprecipitated with 5-HT2B
receptor-FLAG but not with YFP (Fig. 5, A–D). This resultfurther supports protein–protein interactions between thevarious receptor constructs.Interactions between the TM1-tethered (WT, R6G;E42G, or
DNter) constructs and DNter 5-HT2B receptor were furtherexamined in living COS-7 cells using BRET analysis. TheDNter 5-HT2B receptor fused to Rluc (DNter 5-HT2B-Luc) wasexpressed in the presence of increasing concentrations ofBRET acceptors, consisting of TM1-tethered WT, R6G;E42G,or DNter linked to cytoplasmic YFP. In all cases, hyperbolicsaturation curves were obtained, further supporting protein–protein interactions (Fig. 5, E–H). The BRET signals mark-edly depended on the relative distance of donor and acceptor,explaining why the maximal BRET values were slightlydifferent (57.206 5.33; 67.446 2.93; 61.96 7.3). The value ofthe YFP:Luc ratio at which half-maximal BRET was obtained
Fig. 3. Black-Leff operational model to derive putative biasin coupling of variant receptor. To take into accountefficiency of receptor coupling, which may vary among themutants, we used the Black-Leff operational model tocalculate putative difference in signaling characteristics forIP production for each mutant as ΔΔLog(RA). We used twoagonist probes (5-HT as full agonist and DOI as partialagonist) to cancel putative impact of receptor densitydifferences. DOI- and 5-HT–stimulated IP dose responsecurves are shown for COS-7 cells transfected with plasmidsencoding the WT receptor (A) and the R6G;E42G (B). Using5-HT and DOI as probes of the two receptor types, there wasa difference produced in the relative activity of 5-HT andDOI with the mutation from WT to R6G;E42G (C). This isdemonstrated by the ΔΔLog(RA) value 95% confidence limits.There was a significant increase in the relative activity ofDOI over 5-HT produced by the mutation (Bias of 1.23).Results for 5-HT (full reference agonist and DOI as partialagonist) are expressed in femtomoles of IP accumulation (n =3 independent experiments performed in duplicate).
N Terminus Structural Modulation of 5-HT2B Receptor 133
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from
was comparable, indicating that these three TM1 constructsdisplayed the same propensity to associate with DNter5-HT2B-Luc (1.30 6 0.44; 0.86 6 0.15; 0.96 6 0.33). FreeYFP was used as negative control; only a linear nonspecificBRET signal was obtained, confirming the specificity of theobserved interactions.Functional Impact of N-Terminal Domain Interac-
tions. We then evaluated the functional impact receptorsmeasured through the average Emin and Emax for IP accumu-lation after DOI stimulation first for the WT receptor (Fig. 6A)after correction for the protein input and for the surfacereceptor Bmax. We evaluated similarly Emax for IP accumula-tion after DOI stimulation of each combination of constructs(Fig. 6B). When cotransfected with a WT 5-HT2B receptor, thedifferent TM1-tethered 5-HT2B receptor N-terminal constructsdid not modify the IP levels compared with the WT controlalone. In addition, when the R6G;E42G receptor was cotrans-fected with a TM1-tethered DNter construct lacking the N-terminal 32 amino acids of the 5-HT2B receptor or a TM1construct carrying the WT N terminus (TM1-WT), IP accumu-lationwas increased up to 129% of agonist-inducedWT level, aspreviously described for the variant receptor alone and takinginto account the surface expression of this mutant.Thus, neither the WT nor the truncated N-terminal TM1
construct (TM1-WT or TM1-DNter) modified the increase inIP levels induced by the R6G;E42G variant. The increase in IPlevels was also significant in the chimeric construct totallylacking any N-terminal fragment (DNter 1 TM1-DNter), aspreviously observed for the DNter receptor. However, whena TM1 construct carrying the WT N terminus (TM1-WT) was
cotransfected with the DNter receptor, the intracellular IPaccumulation was significantly decreased, becoming nonsignif-icantly different from the WT level (Fig. 6C). However, the IPaccumulation was still increased (.130% WT level) when theDNter receptor was cotransfected with a TM1 constructcarrying the variant mutation (TM1-R6G;E42G). This resultindicates that tethered expression of a WT N terminus (TM1-WT) can restore the WT activity in trans (Fig. 6C). Althoughable to interact biochemically, the lack of effect for the TM1-tethered R6G;E42G fragment clearly demonstrates the loss ofN terminus functionality in the variant receptor.Functional Impact of N-Terminal Domain Variant on
Other Transduction Pathways. In addition to IP coupling,we previously showed that the 5-HT2B receptor can activate cellproliferation via both Gaq/11 and Ga13 (Deraet et al., 2005). Wetherefore assessed whether the polymorphism in the N-terminal sequence modifies the proliferative function of the5-HT2B receptor as measured by [3H]thymidine incorporation(Fig. 7A). Interestingly, in response to 1 mM 5-HT, COS-7 cellsexpressing the R6G;E42G variant incorporated 5 times morethymidine than did cells transfected with WT receptor. Thisobservation suggests that the 5-HT2B N terminus is importantfor additional effector pathways including proliferation.Finally, we observed that the gain-of-function dual SNP
altered the desensitization process of the receptor. We eval-uated the desensitization kinetics of the receptor variant byfirst challenging with DOI in the absence of LiCl (no IP ac-cumulation) and then, after extensive washing, repeating theDOI challenge at various time points in the presence of LiCl.As shown in Fig. 7B, the receptor variant desensitized more
Fig. 4. Confocal microscopy images of transfected COS-7cells showing colocalization of 5-HT2B-DNter and various TM1constructs. COS-7 cells were cotransfected with expressionvectors encoding 5-HT2B-DNter (A–I) and TM1-WT-YFP(A–C), or TM1-R6G/E42G-YFP (D–F), or TM1-DNter-YFP(G–I) and analyzed by confocal immunofluorescence micros-copy. (A, D, and G) Green staining from the transfected TM1-YFP. (B, E, and H) Red staining (Cy3) from 5-HT2B-DNterrevealed using mouse monoclonal antibody directed to-ward C-terminus of the receptor. (C, F, and I) Merge showscolocalization of the signal in yellow. (J and K) To ensure thatthe TM1 construct were also able to reach plasmamembrane,we performed immunostaining of TM1-WT construct withoutthe YFP fusion protein. Antibodies against the N terminus ofthe 5-HT2B receptor were incubated without permeabiliza-tion, indicating that the TM1-WT construct were indeed ableto be expressed at the cell surface. Nontransfected cells areshown as a negative control for immunoreactivity. (A–I bar =5 mM; J and K bar = 25 mM.)
134 Belmer et al.
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

slowly than did the WT receptor, consistent with a modifiedsignal transduction capacity of the polymorphic 5-HT2B
receptor.
DiscussionWe report a functional study of two HTR2B SNPs that
modify two amino acids (R6G;E42G) within the N terminus ofthe receptor. The polymorphisms were previously identified indrug-abusing patients. We show that the R6G;E42G variant
of the 5-HT2B receptor displays increased agonist affinity andefficacy, as highlighted by an increased basal activity Emin
and increased Emax of agonist-induced IP production afternormalization by protein content and cell surface receptorexpression Bmax or in the Black–Leff operational model.Furthermore, this variant also increases the ability of thereceptor to trigger cell proliferation and leads to decreaseddesensitization kinetics.Historically, rhodopsin-like 7TMRs have been identified
and studied on the basis of their ability to be activated by
Fig. 5. TM1-tethered (WT, R6G;E42G, orDNter) interaction with DNter 5-HT2Breceptor. (A–D) Immunoprecipitation ofTM1-tethered (WT, R6G;E42G or DNter)with DNter 5-HT2B receptor. COS-7 cellswere cotransfected with plasmids codingfor 5-HT2B receptor-FLAG (2B-FLAG) andTM1-tethered (WT, R6G;E42G, or DNter-YFP) or free YFP (negative controls). Rep-resentative Western blots are shown ofthree independent immunoprecipitationexperiments. (A) Direct blot of input ofproteins revealed using flag antibody. (B)Direct blot of input of proteins revealedusing green fluorescent protein (GFP)antibody. (C) Blot of immunoprecipitatedproteins using the FLAG antibody andrevealed with the FLAG antiserum. (D)Blot of immunoprecipitated proteins us-ing the FLAG antibody and revealed withthe GFP antiserum. (E–H) BRET analysisof TM1-tethered (WT, R6G;E42G orDNter) interaction with DNter 5-HT2Breceptor. COS-7 cells were cotransfectedwith plasmids coding for DNter 5-HT2Breceptor-Rluc (the BRET donor) and in-creasing concentrations of TM1-tethered(WT, R6G;E42G, or DNter-YFP, the BRETacceptors) or free YFP (negative controls).Energy transfer was measured after addi-tion of the membrane permeable luciferasesubstrate coelenterazine h. The BRET sig-nal was determined by calculating the ratioof light emitted at 530 nm over the lightemitted at 485 nm. The value of the YFP:Luc ratio, for which half-maximal BRETobtained, was comparable, indicating thatthese three forms of TM1 display the samepropensity to associate with DNter 5-HT2Breceptor. Representative BRET saturationcurves are shown of three independent ex-periments. Error bars indicate S.E.M. ofspecific BRET-ratio values obtained fromtriplicate.
N Terminus Structural Modulation of 5-HT2B Receptor 135
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

high-affinity diffusible pharmacologic ligands interactingdirectly with the core transmembrane region of the receptor.Interestingly, we found in this study that R6G;E42G 5-HT2B
receptors are more efficiently coupled to G proteins than areWT receptors, as shown by agonist binding and functional
assays. Using various TM1-tethered N termini coexpressedwith the DNter receptor, we demonstrate that these TM1-tethered N termini can interact with the 7TM domains usingcomplementary approaches: colocalization in fixed cells, co-immunoprecipitation in solubilized lysates, and BRET exper-iments on living cells. Furthermore, when the DNter receptorwas cotransfected with a TM1 construct carrying the WT Nterminus, coupling was reversed to WT levels, indicating thattethered expression of a WT N terminus was able to reversethe gain of coupling due to the deletion of the N terminusfrom the 5-HT2B receptor. This was not observed when theDNter receptor was cotransfected with a TM1 constructcarrying the variant mutation (TM1-R6G;E42G). Thus, the Nterminus can alter the coupling efficiency of the receptor intrans.The structural basis of 7TMR oligomerization has been
recently obtained in b1-adrenergic receptors (Huang et al.,2013). In an inactive conformation, two dimer interfaces wereidentified, one involving TM1 and the other engaging residuesfrom TM4 and TM5 (Huang et al., 2013). Previous findingsusing biochemical approaches have identified similar inter-actions in 5-HT2C receptors (Mancia et al., 2008). It is thus nottotally surprising that, using TM1-tethered N-terminal con-structs, we found direct interactions with the 7TM receptor.However, the fact that these interactions were able to mod-ulate basal and agonist-stimulated receptor activity was notanticipated.Recently solved crystal structures have directly confirmed
previous findings that biogenic amine 7TMRs including 5-HT2B
receptors bind endogenous ligands within the TM helicalbundle of the receptor (Manivet et al., 2002; Setola et al.,2005; Wacker et al., 2013). Conventional agonists target thesame binding site on a receptor as the endogenous ligand,termed the orthosteric site (Smith et al., 2011). Our resultssupport the view that the 5-HT2B receptor N terminus, byinteracting with TM and/or extracellular loops, acts nega-tively on coupling efficiency in both basal and stimulatedconditions without directly interfering with the orthostericsite. Thus, we propose that the 5-HT2B receptor N terminusbehaves as a negative structural modulator of the receptoractivation. The fact that the R6G;E42G variant also increasesthe ability of the receptor to trigger cell proliferation and leadsto a decreased desensitization kinetic indicates that thismodulation acts generally on the receptor activation processrather than specifically affecting certain intracellular signal-ing pathways.These findings extend those of previous studies that demon-
strated a role for the N terminus in the function of rhodopsin-like 7TMRs. An interaction between the N terminus and theextracellular loop (ECL)3 of CXCR4 and C5aR has also beenimplicated in an activation microswitch region: whereas theN terminus/ECL3 interaction stabilizes the active state ofCXCR4, it acts as an inverse agonist in C5aR, possibly bymaking multiple contacts with the TM domains to stabilizethe inactive state (Rana and Baranski, 2010). Furthermore,the N terminus of histamine H1 receptors interacts withECL2 to influence receptor pharmacology by contributing toligand binding, receptor activation, and agonist selectivity(Strasser et al., 2008).Since the publication of the b2-adrenergic receptor crystal
structure, a number of new 7TMR homology models based onthis template have been reported. Accurate prediction of the
Fig. 6. Functional study of different variants of 5-HT2B receptors. (A)Basal IP accumulation (Emin) of COS-7 cells transfected with plasmidsencoding the WT, R6G;E42G, R6E;E42R, and DNter receptors. Bmax forWT, R6G;E42G, R6E;E42R, and DNter receptors are 9826 84, 7846 102,860 6 74, 508 6 100 fmoles/mg of proteins, respectively. (B) DOI-stimulated IP maximum (Emax) accumulation of COS-7 cells transfectedwith plasmids encoding the WT, R6G;E42G, R6E;E42R, and DNterreceptors stimulated with DOI (1 mM). Results are presented as thepercentage of WT level from three independent experiments, each per-formed in duplicate, which were normalized in femtomoles of IP perfemtomoles of surface receptors per mg of protein (i.e., corrected by proteinand cell surface Bmax). A typical full-dose response curve is shown on Fig.2C. Statistical analysis was performed by Welch corrected t test; n = 3–5independent experiments performed at least in duplicate: ****P , 0.0001versus WT receptor. (C) Functional study of different variants of 5-HT2Breceptors coexpressed with a TM1-tethered N terminus. TM1-WT andTM1-R6G;E42G constructs are 93 amino-acids long and the TM1-DNterconstruct is deleted of the 32 first amino acids (i.e., 61 amino-acids long).The IP accumulation was measured after DOI (1 mM) stimulation of COS-7 cells transfected with plasmids encoding the WT, variant (R6G;E42G),and DNter receptors. Results were normalized in fmoles of IP per fmoles ofsurface receptors per mg of protein (i.e., corrected by protein and cellsurface Bmax) and then to WT level and were statistically analyzed by one-way analysis of variance and Bonferroni’s multiple comparison post test.F(7, 64) = 6.83, P, 0.0001, n = 3 independent experiments, each performedin triplicate. (Statistical significance: *P , 0.05; **P , 0.01 versus WT +TM1-DNter; #P , 0.05; ##P , 0.01 versus WT + TM1-WT; xP , 0.05; xxP ,0.01 versus WT + TM1-R6G;E42G; $P , 0.05 versus DNter + TM1-DNter;£P , 0.05 versus DNter + TM1-R6G;E42G). Bmax for the WT + TM1ΔNter,WT + TM1WT,WT + TM1R6G;E42G, R6G;E42G + TM1ΔNter, R6G;E42G+TM1WT, and DNter + TM1ΔNter, DNter + TM1WT, DNter + TM1R6G;E42G receptors are 7696 306, 8006 345, 7676 309, 6866 278, 6676 224,554 6 256, 643 6 321, 585 6 266 fmoles/mg of proteins, respectively.
136 Belmer et al.
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

loops, particularly those surrounding the orthosteric bindingsite, remains one of the more difficult aspects in 7TMR ho-mology modeling. In a previous report of homology models,5-HT2B receptors were identified as presenting difficulty inmodeling their ECL2 (McRobb et al., 2010). This has beensolved by the recently published crystal structure for the5-HT2B receptor (Wacker et al., 2013). The E212-R213-F214residues of the 5-HT2B receptor form an additional helicalturn stabilized by a structured water molecule at the extra-cellular tip of helix V (Wacker et al., 2013). As a result, thesegment of ECL2 that connect helices III and V via the con-served disulfide bond is shortened in the 5-HT2B receptor.However, the N terminus being absent in this crystal struc-ture, further investigations are necessary to fully understandthe interactions of the N terminus with the ECLs/TMs of5-HT2B receptors.The N-terminal regions of 7TMRs have been shown to
control the surface expression of receptor proteins, but theireffects vary depending on the structural features of eachreceptor. Polymorphisms in the b2-adrenergic receptor (R16G;Q27E) display normal agonist binding and functional cou-pling to Gs, but markedly alter the degree of agonist-promoteddownregulation of receptor expression (Green et al., 1994).In the human b1-adrenergic receptor, the N-terminal G49variant was found to display amore profound agonist-promoteddownregulation and both basal and agonist-stimulatedadenylyl cyclase activity than the S49 variant (Levin et al.,2002). Here, we show that the increased basal and agonist-stimulated IP activity, gain of function in R6G;E42G receptorcoupling is associated with a reduced rate of receptor de-sensitization, which may participate to this gain of function.Our study further revealed that not only signal trans-
duction pathways but also desensitization kinetic could beaffected by modifying the N terminus of 5-HT2B receptors.Although in vivo validation of these findings must beperformed, it is clear that our findings support the need formodeling 7TMRwith their N terminus and extracellular loopsto fully understand their activation mechanisms. In conclu-sion, by using deletion or mutation that leads to a gain ofcoupling efficiency both in basal and stimulated conditions,
our results support that the human 5-HT2B receptor Nterminus behaves as a negative modulator of the receptoractivity.
Acknowledgments
The authors thank Jean-Marie Launay for helpful discussions andfor the critical reading of this manuscript, and Stefano Marullo forthe Rluc plasmid and advice on BRET experiments.
Authorship Contributions
Participated in research design: Belmer, Doly, Setola, Maroteaux.Conducted experiments: Belmer, Doly, Banas, Setola, Moutkine,
Boutourlinsky.Performed data analysis: Belmer, Doly, Banas, Setola, Maroteaux.Wrote or contributed to the writing of the manuscript: Belmer, Doly,
Setola, Kenakin, Maroteaux.
References
Achour L, Kamal M, Jockers R, and Marullo S (2011) Using quantitative BRET toassess G protein-coupled receptor homo- and heterodimerization.Methods Mol Biol756:183–200.
Andersson H, D’Antona AM, Kendall DA, Von Heijne G, and Chin CN (2003) Mem-brane assembly of the cannabinoid receptor 1: impact of a long N-terminal tail.MolPharmacol 64:570–577.
Black JW and Leff P (1983) Operational models of pharmacological agonism. Proc RSoc Lond B Biol Sci 220:141–162.
Colas J-F, Choi D-S, Launay J-M, and Maroteaux L (1997) Evolutionary conservationof the 5-HT2B receptors. Ann N Y Acad Sci 812:149–153.
Cussac D, Boutet-Robinet E, Ailhaud MC, Newman-Tancredi A, Martel JC, Danty N,and Rauly-Lestienne I (2008) Agonist-directed trafficking of signalling at serotonin5-HT2A, 5-HT2B and 5-HT2C-VSV receptors mediated Gq/11 activation and cal-cium mobilisation in CHO cells. Eur J Pharmacol 594:32–38.
Deraet M, Manivet P, Janoshazi A, Callebert J, Guenther S, Drouet L, Launay JM,and Maroteaux L (2005) The natural mutation encoding a C terminus-truncated 5-hydroxytryptamine 2B receptor is a gain of proliferative functions. Mol Pharmacol67:983–991.
Doly S, Bertran-Gonzalez J, Callebert J, Bruneau A, Banas SM, Belmer A,Boutourlinsky K, Hervé D, Launay JM, and Maroteaux L (2009) Role of sero-tonin via 5-HT2B receptors in the reinforcing effects of MDMA in mice. PLoS ONE 4:e7952.
Doly S, Valjent E, Setola V, Callebert J, Hervé D, Launay JM, and Maroteaux L (2008)Serotonin 5-HT2B receptors are required for 3,4-methylenedioxymethamphetamine-induced hyperlocomotion and 5-HT release in vivo and in vitro. J Neurosci 28:2933–2940.
Dunham JH, Meyer RC, Garcia EL, and Hall RA (2009) GPR37 surface expressionenhancement via N-terminal truncation or protein-protein interactions. Bio-chemistry 48:10286–10297.
Ehlert FJ (2005) Analysis of allosterism in functional assays. J Pharmacol Exp Ther315:740–754.
Ersoy BA, Pardo L, Zhang S, Thompson DA, Millhauser G, Govaerts C, and Vaisse C(2012) Mechanism of N-terminal modulation of activity at the melanocortin-4 re-ceptor GPCR. Nat Chem Biol 8:725–730.
Fig. 7. Proliferation rate of R6G;E42G 5-HT2B receptor compared with WT 5-HT2B receptors. (A) Measurement of proliferation rate by [3H]thymidineincorporation performed for COS-7 cells transfected with plasmids encoding the WT receptor and for the variant R6G;E42G. Thymidine levels arenormalized to WT. Results of four independent experiments each performed in triplicate were statistically analyzed by unpaired t test with Welch’scorrection (****P , 0.0001). (B) Desensitization kinetics were evaluated after a first stimulation with 100 nM of DOI replaced by fresh mediumcontaining LiCl + DOI (100 nM). A delay of desensitization of the R6G;E42G variant receptor was observed during the first 30 minutes, compared withWT. Results of experiments performed in duplicate were statistically analyzed by two-way analysis of variance with mutation and time as main factors;effect of mutations on desensitization, F(1,2) = 19.35, P = 0.048. Bonferroni post test was also applied on each graph: *P , 0.05; **P , 0.01 versus WTreceptor.
N Terminus Structural Modulation of 5-HT2B Receptor 137
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from

Green S, Issemann I, and Sheer E (1988) A versatile in vivo and in vitroeukaryotic expression vector for protein engineering. Nucleic Acids Res 16:369.
Green SA, Turki J, Innis M, and Liggett SB (1994) Amino-terminal polymorphisms ofthe human beta 2-adrenergic receptor impart distinct agonist-promoted regulatoryproperties. Biochemistry 33:9414–9419.
Hague C, Chen Z, Pupo AS, Schulte NA, Toews ML, and Minneman KP (2004) The Nterminus of the human alpha1D-adrenergic receptor prevents cell surface expres-sion. J Pharmacol Exp Ther 309:388–397.
Huang J, Chen S, Zhang JJ, and Huang X-Y (2013) Crystal structure of oligomericb1-adrenergic G protein-coupled receptors in ligand-free basal state. Nat StructMol Biol 20:419–425.
Kenakin T, Watson C, Muniz-Medina V, Christopoulos A, and Novick S (2012) Asimple method for quantifying functional selectivity and agonist bias. ACS ChemNeurosci 3:193–203.
Levin MC, Marullo S, Muntaner O, Andersson B, and Magnusson Y (2002) Themyocardium-protective Gly-49 variant of the beta 1-adrenergic receptor exhibitsconstitutive activity and increased desensitization and down-regulation. J BiolChem 277:30429–30435.
Lin Z, Walther D, Yu X-Y, Drgon T, and Uhl GR (2004) The human serotonin receptor2B: coding region polymorphisms and association with vulnerability to illegal drugabuse. Pharmacogenetics 14:805–811.
Mancia F, Assur Z, Herman AG, Siegel R, and Hendrickson WA (2008) Ligand sen-sitivity in dimeric associations of the serotonin 5HT2c receptor. EMBO Rep 9:363–369.
Manivet P, Schneider B, Smith JC, Choi DS, Maroteaux L, Kellermann O,and Launay JM (2002) The serotonin binding site of human and murine 5-HT2Breceptors: molecular modeling and site-directed mutagenesis. J Biol Chem 277:17170–17178.
McRobb FM, Capuano B, Crosby IT, Chalmers DK, and Yuriev E (2010) Homologymodeling and docking evaluation of aminergic G protein-coupled receptors. J ChemInf Model 50:626–637.
Ramachandran R and Hollenberg MD (2008) Proteinases and signalling: patho-physiological and therapeutic implications via PARs and more. Br J Pharmacol153 (Suppl 1):S263–S282.
Rana S and Baranski TJ (2010) Third extracellular loop (EC3)-N terminus in-teraction is important for seven-transmembrane domain receptor function: impli-cations for an activation microswitch region. J Biol Chem 285:31472–31483.
Setola V, Dukat M, Glennon RA, and Roth BL (2005) Molecular determinants for theinteraction of the valvulopathic anorexigen norfenfluramine with the 5-HT2B re-ceptor. Mol Pharmacol 68:20–33.
Smith NJ, Bennett KA, and Milligan G (2011) When simple agonism is not enough:emerging modalities of GPCR ligands. Mol Cell Endocrinol 331:241–247.
Srinivasan S, Lubrano-Berthelier C, Govaerts C, Picard F, Santiago P, Conklin BR,and Vaisse C (2004) Constitutive activity of the melanocortin-4 receptor is main-tained by its N-terminal domain and plays a role in energy homeostasis in humans.J Clin Invest 114:1158–1164.
Strasser A, Wittmann HJ, and Seifert R (2008) Ligand-specific contribution of the Nterminus and E2-loop to pharmacological properties of the histamine H1-receptor.J Pharmacol Exp Ther 326:783–791.
Vassart G, Pardo L, and Costagliola S (2004) A molecular dissection of the glyco-protein hormone receptors. Trends Biochem Sci 29:119–126.
Wacker D, Wang C, Katritch V, Han GW, Huang XP, Vardy E, McCorvy JD, Jiang Y,Chu M, and Siu FY et al. (2013) Structural features for functional selectivity atserotonin receptors. Science 340:615–619.
Wittmann H-J, Seifert R, and Strasser A (2011) Influence of the N-terminus and theE2-loop onto the binding kinetics of the antagonist mepyramine and the partialagonist phenoprodifen to H(1)R. Biochem Pharmacol 82:1910–1918.
Address correspondence to: Luc Maroteaux, INSERM UMR S-839,Université Pierre et Marie Curie, Institut du Fer à Moulin, 17 rue du Fer àMoulin 75005 Paris, France. E-mail: [email protected]
138 Belmer et al.
at ASPE
T Journals on Septem
ber 1, 2018m
olpharm.aspetjournals.org
Dow
nloaded from