roles of small regulatory rnas in determining neuronal identity
TRANSCRIPT

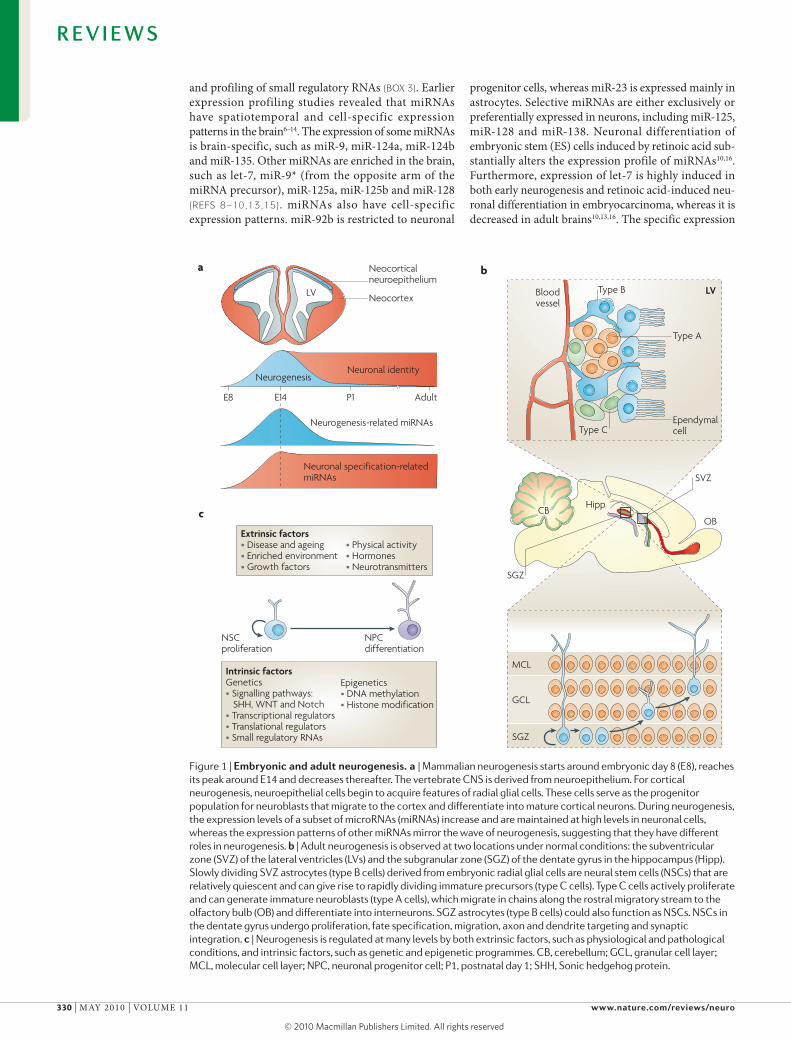
Neurogenesis is the process by which functionally integrated neurons are generated from neural stem cells (NSCs). It involves the proliferation and neuronal fate specification of NSCs, as well as the maturation and functional integration of the neuronal progeny into neu-ronal circuits1,2. NSCs exhibit the two essential proper-ties of stem cells: self-renewal and multipotency. During embryogenesis, NSCs are located in the ventricular zone of the neural tube. Embryonic NSCs can give rise to all the cell types required for the formation of the CNS. Contrary to the earlier dogma that neurogenesis occurs only during development, it is now generally accepted that neurogenesis occurs throughout life in mammalian brains. Adult neurogenesis has been observed at two locations under normal conditions: the subventricular zone (SVZ) of the lateral ventricles and the subgranu-lar zone (SGZ) of the dentate gyrus in the hippocampus (FIG. 1). Neurogenesis is regulated at many levels by both extrinsic factors, such as physiological and pathological conditions (BOX 1; FIG. 1c), and intrinsic factors, such as genetic and epigenetic programmes (FIG. 1c). The main-tenance and differentiation of NSCs are tightly control-led by intricate molecular networks. Uncovering these regulatory mechanisms is crucial for understanding the functions and plasticity of the CNS.
Genome projects have shown that at least 93% of nucleotides in the human genome are transcribed in dif-ferent cells, with similar findings in the mouse and other eukaryotes. However, only around 1.2% of transcribed RNAs encode proteins, indicating that there may be a vast reservoir of biologically meaningful non-coding RNAs, often referred to as the ‘dark matter’ of the cell3. Non-coding RNAs may regulate gene expression at many
levels and through a range of mechanisms. Uncovering the functions of these non-coding RNAs could substan-tially improve our understanding and the treatment of human diseases.
The small regulatory RNAs identified so far include microRNAs (miRNAs), endogenous small interfering RNAs (esiRNAs), Piwi-interacting RNAs (piRNAs) and promoter-associated short RNAs (PASRs; also known as transcription start site RNAs (TSS RNAs) and transcrip-tion initiation RNAs (tiRNAs)). These small RNAs range from 18 to 30 nucleotides in length and can shape diverse cellular pathways (BOX 2). Recent studies have shown that several classes of small regulatory RNAs have impor-tant roles in stem cell biology; indeed, the stem cell and miRNA fields have converged with the identification of stem cell-specific and lineage-specific miRNAs4,5. It has become clear that miRNAs provide a new dimension to the regulation of stem cell functions. For example, owing to their functions in the regulation of translation, miRNAs can regulate stem cell fate and behaviour by fine-tuning the protein levels of various factors required for stem cell proliferation and multipotency. Furthermore, piRNAs are also important for stem cell self-renewal, as their partner Piwi proteins are required for stem cell main-tenance. In this Review, we focus on recent progress in understanding the roles of small regulatory RNAs in neurogenesis.
Expression of small RNAs in neurogenesisDetecting nucleic acids that are only 18–30 nucle-otides in length is inherently difficult, simply because of their small size. The past several years have seen the emergence of new techniques for the identification
Department of Human Genetics, Emory University School of Medicine, 615 Michael Street, Suite 301, Atlanta, Georgia 30322, USA.Correspondence to P.J. e‑mail: [email protected]:10.1038/nrn2739Published online 31 March 2010; corrected online 30 April 2010
Self-renewalThe capacity of a cell to proliferate and produce identical cells.
MultipotencyThe potential of a cell to give rise to multiple lineage cells. Neural stem cells, for example can generate neurons, astrocytes and oligodendrocytes.
Roles of small regulatory RNAs in determining neuronal identityXuekun Li and Peng Jin
Abstract | Neurogenesis, the process of generating functional neurons from neural stem cells, is tightly controlled by many intrinsic and extrinsic mechanisms. Uncovering these regulatory mechanisms is crucial for understanding the functions and plasticity of the human brain. Recent studies in both invertebrates and vertebrates point to the importance of small regulatory RNAs in regulating lineage-specific gene expression and determining neuronal identity during neurogenesis. These new observations suggest that small regulatory RNAs could function at many levels to regulate self-renewal of neural stem cells and neuronal fate specification, implicating small regulatory RNAs in the complexity of neurogenesis.
C N S p R E C u R S o R S
R E V I E W S
NATURE REVIEwS | NeuroscieNce VolUmE 11 | mAy 2010 | 329
© 20 Macmillan Publishers Limited. All rights reserved10
Nature Reviews | Neuroscience
Neurogenesis
Neocorticalneuroepithelium
Neuronal identity
Neurogenesis-related miRNAs
Neuronal specification-related miRNAs
E8 E14 P1 Adult
NeocortexLV Bloodvessel
Type B
Type C
Type A
LV
Ependymalcell
MCL
GCL
SGZ
a b
cHipp
SVZ
OB
SGZ
CB
NSCproliferation
NPCdifferentiation
Extrinsic factors• Disease and ageing• Enriched environment• Growth factors
• Physical activity• Hormones• Neurotransmitters
Intrinsic factorsGenetics• Signalling pathways: SHH, WNT and Notch• Transcriptional regulators• Translational regulators• Small regulatory RNAs
Epigenetics• DNA methylation• Histone modification
and profiling of small regulatory RNAs (BOX 3). Earlier expression profiling studies revealed that miRNAs have spatiotemporal and cell-specific expression patterns in the brain6–14. The expression of some miRNAs is brain-specific, such as miR-9, miR-124a, miR-124b and miR-135. other miRNAs are enriched in the brain, such as let-7, miR-9* (from the opposite arm of the miRNA precursor), miR-125a, miR-125b and miR-128 (ReFs 8–10,13,15). miRNAs also have cell-specific expression patterns. miR-92b is restricted to neuronal
progenitor cells, whereas miR-23 is expressed mainly in astrocytes. Selective miRNAs are either exclusively or preferentially expressed in neurons, including miR-125, miR-128 and miR-138. Neuronal differentiation of embryonic stem (ES) cells induced by retinoic acid sub-stantially alters the expression profile of miRNAs10,16. Furthermore, expression of let-7 is highly induced in both early neurogenesis and retinoic acid-induced neu-ronal differentiation in embryocarcinoma, whereas it is decreased in adult brains10,13,16. The specific expression
Figure 1 | embryonic and adult neurogenesis. a | Mammalian neurogenesis starts around embryonic day 8 (E8), reaches its peak around E14 and decreases thereafter. The vertebrate CNS is derived from neuroepithelium. For cortical neurogenesis, neuroepithelial cells begin to acquire features of radial glial cells. These cells serve as the progenitor population for neuroblasts that migrate to the cortex and differentiate into mature cortical neurons. During neurogenesis, the expression levels of a subset of microRNAs (miRNAs) increase and are maintained at high levels in neuronal cells, whereas the expression patterns of other miRNAs mirror the wave of neurogenesis, suggesting that they have different roles in neurogenesis. b | Adult neurogenesis is observed at two locations under normal conditions: the subventricular zone (SVZ) of the lateral ventricles (LVs) and the subgranular zone (SGZ) of the dentate gyrus in the hippocampus (Hipp). Slowly dividing SVZ astrocytes (type B cells) derived from embryonic radial glial cells are neural stem cells (NSCs) that are relatively quiescent and can give rise to rapidly dividing immature precursors (type C cells). Type C cells actively proliferate and can generate immature neuroblasts (type A cells), which migrate in chains along the rostral migratory stream to the olfactory bulb (OB) and differentiate into interneurons. SGZ astrocytes (type B cells) could also function as NSCs. NSCs in the dentate gyrus undergo proliferation, fate specification, migration, axon and dendrite targeting and synaptic integration. c | Neurogenesis is regulated at many levels by both extrinsic factors, such as physiological and pathological conditions, and intrinsic factors, such as genetic and epigenetic programmes. CB, cerebellum; GCL, granular cell layer; MCL, molecular cell layer; NPC, neuronal progenitor cell; P1, postnatal day 1; SHH, Sonic hedgehog protein.
R E V I E W S
330 | mAy 2010 | VolUmE 11 www.nature.com/reviews/neuro
© 20 Macmillan Publishers Limited. All rights reserved10

Deep sequencingAn approach enabled by next-generation sequencing technology that is particularly useful for identifying low- abundance RNAs or low-frequency mutations.
Cre–loxP systemA site-specific recombination system derived from Escherichia coli bacteriophage P1. Two short DNA sequences (loxP sites) are engineered to flank the target DNA. Activation of the Cre-recombinase enzyme catalyses recombination between the loxP sites, leading to excision of the intervening DNA sequence.
Environmental enrichmentProviding animals under managed care with environmental stimuli to improve the quality of life by increasing physical activity, stimulating natural behaviours and preventing or reducing neural disorders including stereotypical behaviours.
of miRNA in the nervous system strongly suggests that miRNAs could play key parts in brain development and neuronal fate specification (FIG. 2a).
Studies in human and mouse ES cells have shown that the complete repertoire of miRNAs in stem cells can only be uncovered through deep sequencing, as some miRNAs are expressed solely in ES cells. Thus, it is important to use this technology to profile the expres-sion of small regulatory RNAs in both proliferating and differentiated NSCs — an approach that has not been taken so far.
NSC fate determination by miRNAsTo determine whether miRNAs play a crucial part in neurogenesis and the development of the nervous system, early studies focused on key components of the miRNA pathway. loss of Dicer (an RNase III enzyme that cleaves the miRNA precursor (pre-miRNA) to pro-duce mature miRNA) in zebrafish was found to lead to defective neural tubes, reduced ventricle size and loss of the midbrain–hindbrain boundary17. Surprisingly, these defects could be partially rescued by a single miRNA, miR-430 (ReF. 17). Similarly, the disruption of miRNA-induced silencing complex (miRISC) activity due to a mutation in Argonaute 2 (AGo2), an essential component of the complex, prevented neural-tube clo-sure in mice18. These two studies provide strong in vivo evidence that the miRNA pathway is important for brain development and morphogenesis. Further advances were provided by studies in which Dicer activity was condi-tionally removed in specific neuronal cell types using the Cre–loxP system. This led to various defects in neural development, such as increased cortical apoptosis, smaller cortex size, enlarged lateral ventricles, profound astrogliosis, altered dendritic maturation, smaller neural cells and abnormal neurogenesis19–22. These findings indicate an essential role for the miRNA pathway in neural development.
During neurogenesis, neuronal diversity can be achieved through the asymmetrical cell division of NSCs, which produces two daughter cells with different developmental potentials. whereas one differenti-ates into a neuron, the other maintains a stem cell fate. Although the mechanism is more complex in verte-brates than invertebrates, the unequal inheritance of cell fate determinants (DNA or protein) ensures that each daughter cell acquires a different fate23–25. The TRIm-NHl (tripartite motif and NCl1, HT2A and lIN41 domain-containing) protein TRIm32, a previously identified E3 ubiquitin ligase, was recently found to enhance the activity of selected miRNAs by interacting with AGo1, a member of the Argonaute protein family involved in miRNA-mediated gene regulation26. During asymmetrical cell division in NSCs and neural progeni-tor cells (NPCs), TRIm32 is enriched in one of the two daughter cells. TRIm32 suppresses cell proliferation and induces neuronal differentiation by degrading the tran-scription factor myC and enhancing let-7a activity26. Because TRIm32 and let-7a are also present in NSCs and NPCs, it is possible that another pathway might exist to antagonize the effect of TRIm32–let7a and maintain their multipotency27. These findings suggest that the unequal distribution of a ribonucleoprotein complex-containing TRIm32 and its target miRNAs could regu-late the balance between differentiation and proliferation in neural stem cell lineages, and thus that miRNAs could participate in cell fate determination and the regulation of asymmetrical cell division.
Recent studies on individual miRNAs have begun to shed light on their specific functions in neurogenesis and neuronal development (see below)4,28–32. Further research in this area could reveal novel mechanisms regulating neurogenesis and targets to manipulate this process.
Interplay between miRNAs and transcription factors. The interplay between miRNAs and transcription factors was first uncovered in Caenorhabditis elegans. In C. elegans, two taste neurons, ASE right (ASER) and ASE left (ASEl), display left–right asymmetrical expres-sion patterns of putative chemoreceptor genes, which are important for functional lateralization of taste neurons. ASEl and ASER were found to express distinct miRNAs, lsy-6 and miR-273, respectively33,34. lsy-6 and miR-273 regulate the establishment of neuronal left–right asym-metry by suppressing the translation of their targets, cog‑1 and die‑1. CoG-1 and DIE-1 can also inhibit the expression of lsy-6 and miR-273 to form a double- negative feedback loop33–35. These studies provide an excellent example of how miRNAs can specify distinct neuronal cell fates from a multipotent precursor state.
miR-124 is highly conserved in both invertebrates and vertebrates and is the most abundant miRNA in both the embryonic and adult CNS8. Its temporal expres-sion pattern suggests it may have a function in neuronal development7,16. Ectopic expression of miR-124 in cul-tured non-neuronal cells increased the expression of neuronal genes and inhibited non-neuronal genes16,36,37. This is required during early embryonic neuro genesis to ensure that differentiation occurs at the correct
Box 1 | Multiple extrinsic factors regulate neurogenesis
Diverse intrinsic and extrinsic mechanisms are known to regulate neurogenesis1,2. The following extrinsic factors have been found to be involved: hormones and neural transmitters, including dopamine, serotonin, acetylcholine and glutamate; growth factors, such as fibroblast growth factor 2, epidermal growth factor and vascular endothelial growth factor; exercise and learning; environmental enrichment and pathological conditions, such as stress, seizure, ischaemia and ageing.
Mounting evidence indicates the essential role of the stem cell niche in the maintenance and regulation of neurogenesis. In the subventricular zone (SVZ) of adult mice, the cilia of ependymal cells transform the flow of cerebrospinal fluid into a chemical signal that directs the establishment of the chemorepulsive gradients, which guides the migration of newly born neurons88. Recent studies revealed that the vascular system is also a key component of the stem cell niche and is involved in regulating neurogenesis in the SVZ89,90. Proliferating neural stem cells (NSCs) are tightly apposed to the blood vessels through the laminin receptor α6β1 integrin, and neuroblast chains run parallel to blood vessels in the direction of migration towards the rostral migratory stream89,90. Vasculature-derived signals can access the SVZ, and most of the proliferating cells that were recovered from treatment with the anti-mitotic reagent cytosine–arabinofurano-side were directly coupled to the vasculature90, suggesting that the vasculature provides signalling cues to NSCs and regulates their function. Given the roles of small regulatory RNAs in other types of stem cell niche5, it is likely that they also have important roles in the NSC niche, although this remains to be explored.
R E V I E W S
NATURE REVIEwS | NeuroscieNce VolUmE 11 | mAy 2010 | 331
© 20 Macmillan Publishers Limited. All rights reserved10

Transit-amplifying cellsCells that arise from adult stem cells and divide a finite number of times until they become differentiated. They are committed progenitor cells.
time and that the appropriate number of neurons is generated. RE1-silencing transcription factor (REST; also known as NRSF) suppresses neuronal genes in non-neuronal cells by binding to a conserved repressor element (RE1) in neuronal gene loci and recruiting the co-repressor complex containing histone deacetylases and the methyl-CpG-binding protein mECP2 (ReF. 38). Downregulation of REST during the transition from pro-genitors to post-mitotic neurons allows neuronal gene expression. Small carboxy-terminal domain phosphatase 1 (SCP1; also known as CTDSP1) is another anti-neural factor expressed in non-neuronal tissues39. SCP1 can be recruited to RE1-containing neuronal genes by REST in non-neuronal cells. The timely downregulation of the anti-neural REST–SCP1 pathway is thought to be important in embryonic neurogenesis. REST binds to RE1 sites within the promoter regions of three miR-124 genomic loci and represses their expression through the recruitment of two co-repressor complexes, SIN3 and CoREST40. miR-124 has been shown to directly target the SCP1 3′-untranslated region (UTR), which sup-presses SCP1 expression39. In addition to SCP1, compu-tational approaches have also uncovered miR-124 target sites in the 3′-UTR of mECP2 and CoREST. These findings suggest the existence of a negative-feedback loop between REST–SCP1 and miR-124 that mediates the rapid transition between neural progenitors and post-mitotic neurons during embryonic neurogenesis39
(FIG. 2b).
In the SVZ of the adult mouse, miR-124 shows cell type-specific expression. It is expressed in neuroblasts as well as mature granule and periglomerular neu-rons, but not in dividing doublecortin-negative transit-amplifying cells or in oligodendrocytes or astrocytes41. During the transition from amplifying type C cells to neuroblasts, the expression of miR-124 increases and is much higher in mature neurons, suggesting miR-124 might be important for cell lineage progression. Further gain- and loss-of-function studies suggest that miR-124 regulates the progression of neuronal differentiation by targeting SoX9, a transcription factor that can maintain the pluripotent state of NSCs and inhibit neuronal dif-ferentiation41 (FIG. 2b). whether SCP1 is regulated by miR-124 in adult NSCs remains to be determined. As well as the interplay between miR-124 and transcrip-tion factors, it has also been shown that miR-124 could promote neuronal differentiation and neuronal line-age commitment by inducing neuron-specific alterna-tive splicing of polypyrimidine tract-binding protein 1 (PTBP1; also known as PTB and hnRNP I), a global repressor of neuron-specific splicing37. These findings suggest that miR-124 fine-tunes neurogenesis by target-ing multiple mRNAs that are involved in different gene regulatory mechanisms.
miR-9 is also highly conserved and is expressed in most of the NPCs of zebrafish, chickens and mice6,15,42. In Drosophila melanogaster, sensory organs develop from the divisions of single sensory organ precursor cells (SoPs). Ectopic miR-9a expression led to a signifi-cant reduction in the number of SoPs, whereas the loss of miR-9a resulted in extra production of SoPs. This indicates that miR-9a ensures that the precise number of neuronal precursor cells are generated by controlling cell fate specification43. Appropriate expression of the tran-scription factor Senseless (SENS) is required to control the formation of the precise number of SoPs: high levels of SENS in SoPs promote pro-neural gene transcrip-tion, whereas low levels of SENS in non-SoPs repress the expression of pro-neural genes. miR-9a inhibits neuronal fate in non-SoP cells by repressing SENS expression in the embryonic and adult stages43.
During vertebrate brain development, the midbrain–hindbrain boundary (mHB) functions as an organizing centre in the neural tube that is both necessary and suf-ficient for the ordered development of the hindbrain. The mHB contains a pool of NPCs that contributes neurons to the entire mHB. Both gain- and loss-of-function studies in zebrafish have shown that miR-9 is required for the normal generation of new neurons in the mHB by repressing the anti-neurogenic genes her5 and her9 (ReF. 44). moreover, ectopic expression of miR-9 in embryonic mouse cortex through in vivo electroporation led to premature neuronal differentiation in the cor-tex, produced more neurons in the ventricular zone by repressing the expression of forkhead box protein G1 (which has been shown to suppress early cortical cell fate) and disrupted the migration of new neurons in the cortex45. In adult neurogenesis, miR-9 inhibits NSC pro-liferation and accelerates neural differentiation by tar-geting the nuclear receptor TlX, an essential regulator
Box 2 | Types of small regulatory RNAs and their biogenesis
With the advance of high-throughput sequencing technology, the field of small regulatory RNAs is rapidly expanding. Currently, the small regulatory RNAs identified in mammals include microRNAs (miRNAs), endogenous small interfering RNAs (esiRNAs), Piwi-interacting RNAs (piRNAs) and promoter-associated short RNAs (PASRs; also known as transcription start site RNAs (TSS RNAs) and transcription initiation RNAs (tiRNAs))75,83,91,92.
miRNAs are 18–25-nucleotide non-coding RNAs that can regulate the translation of target mRNA molecules in a sequence-specific manner. Most endogenous miRNA genes are transcribed initially as primary transcripts that contain one or more extended hairpin structures. The nuclear RNase III enzyme Drosha, working with DGCR8, cleaves both strands near the base of the primary stem loop and yields the precursor miRNA (pre-miRNA). After being exported to the cytoplasm by exportin 5 (also known as RANGTP), pre-miRNAs are further cleaved by the RNase III enzyme Dicer into mature miRNAs, which will be incorporated into the RNA-induced silencing complex. It has been shown that a single miRNA can simultaneously regulate the expression of many mRNA targets and thereby fine-tune protein expression. Many miRNAs are expressed in the nervous system, where they affect numerous neuronal genes.
High-throughput sequencing has recently identified three new classes of small regulatory RNAs: esiRNAs, piRNAs and PASRs. The biogenesis of esiRNAs starts with either bidirectional transcription or the transcription of an inverted repeat. The resulting double-stranded RNA or the hairpin RNA precursor is then processed by the components of the miRNA pathway. Although miRNA and esiRNA biogenesis is generally Dicer-dependent, piRNAs are generated from a long single-stranded precursor independently of Dicer. piRNAs also differ from miRNAs and siRNAs in that they are usually 26–31 nucleotides long and associated with Piwi subfamily proteins. Most piRNAs map to the repetitive regions of the genome and are thought to control the activity of transposons. PASRs map to the 5′ end or promoter region of the protein-coding genes and are proposed to have a role in the transcription of splice variants. Whether these small regulatory RNAs are expressed during neurogenesis remains to be determined.
R E V I E W S
332 | mAy 2010 | VolUmE 11 www.nature.com/reviews/neuro
© 20 Macmillan Publishers Limited. All rights reserved10

Locked nucleic acidA modified RNA nucleotide with high stability, which can be used as a highly sensitive detection probe.
of NSC proliferation and self-renewal, by activating wNT–β-catenin signalling and repressing the cyclin-dependent kinase inhibitor 1A (CDKN1; also known as wAF1 or CIP1)46. miR-9 suppresses TlX expres-sion through the 3′-UTR of TlX mRNA47. Introducing a TlX expression vector lacking the endogenous TlX 3′-UTR rescued the proliferation deficiency induced by miR-9 overexpression and reversed the miR-9-induced premature differentiation47. Furthermore, TlX binds to the 3′ genomic sequences of miR-9 and inhibits miR-9 expression. Thus, miR-9 and TlX form a feedback loop to regulate the switch between NSC proliferation and differentiation (FIG. 2b).
In summary, these studies show that transcription factors can directly regulate the expression of specific miRNAs, and specific miRNAs can then target other transcription factors and regulate their expression post-transcriptionally. The result is a transcription factor to miRNA to another transcription factor (sometimes even of the miRNA itself ) paradigm for regulating neurogenesis. This represents an interesting, evolution-arily conserved strategy to keep the balance between miRNAs and their transcriptional regulatory pro-grammes. However, it has been well documented that many transcription factors can coordinate the regulation of a given gene and that one miRNA can regulate many mRNA targets. Thus, although the current studies focus on one miRNA and one transcription factor at a time, it is likely that there is a more complex interplay between the miRNA network and the transcriptional regulation network in regulating neurogenesis.
miRNA-mediated regulation of signalling cascades. miR-9 not only controls neurogenesis at the mHB, but also regulates the formation of the mHB in embry-onic zebrafish despite not being expressed at this site. overexpression of miR-9 in the mHB leads to loss of the mHB by inhibiting the fibroblast growth factor (Fgf)
signalling pathway components Fgf8, Fgf receptor 1 and canopy 1 (ReF. 44). These results indicate that miR-9 inhibits the Fgf signalling pathway and promotes neuro-genesis by targeting different mRNAs, which act con-vergently to negatively delimit the mHB, and suggest a mechanism whereby organizer activity and neurogenesis are spatially coordinated in the mHB. It also illustrates a common theme in miRNA-mediated gene regulation; namely, that a given miRNA often regulates several com-ponents within the same biological pathway to fine-tune the output of this pathway.
The Sonic hedgehog (SHH) signalling pathway acts through the Patched 1–Smoothened (Smo) receptor complex to activate downstream effectors and has a piv-otal function in cell differentiation, survival and prolif-eration, and in tissue patterning48. For example, during Xenopus laevis retinal development, blocking the SHH pathway delayed cell cycle exit and induced the release of the translational inhibition of two homeobox genes, otx2 and vsx1, which act as key positive regulators of bipolar neuron generation49. It was shown that four miRNAs (miR-129, miR-155, miR-214 and miR-222) concertedly bind to the 3′-UTR of otx2 and vsx1 mRNAs, inhibiting their translation and thus the generation of bipolar neu-rons49. Inhibition of these four miRNAs promotes pro-genitor cell commitment to the bipolar cell fate49. The SHH pathway also controls the proliferation of cerebellar granule cell progenitors (GCPs), which respond to SHH produced by Purkinje cells. Excessive SHH signalling in GCPs induces neoplastic transformation into medullob-lastoma. Some miRNAs, including miR-125b, miR-324-5p and miR-326, are abnormally expressed in medullo-blastoma, and these miRNAs inhibit medullo blastoma cell proliferation by repressing components of the SHH pathway, such as GlI1 and Smo50. Conversely, loss of these miRNAs has been shown to promote GCP differ-entiation into granule cells50. A recent study showed that the miR-17/92 cluster is upregulated by SHH signalling
Box 3 | Expression profiling of small regulatory RNAs
Diverse methodologies have been used to determine the expression of small regulatory RNAs, including northern blot, in situ hybridization, microarray hybridization-based detection (using locked nucleic acid (LNA) and DNA), reverse transcription PCR-based detection (TaqMan and SYBR Green), direct sequencing of adaptor-ligated cDNA that is complementary to the small RNA and solution hybridization to oligonucleotide (LNA and DNA)-conjugated beads.
In recent years, next-generation sequencing technology has been widely used to profile small regulatory RNAs. Currently, three high-throughput sequencing platforms are commercially available: the Illumina Solexa Genome Analyser, the Roche 454 GS FLX Genome Analyser and Applied Biosystems’s SOLiD system93–95. All three systems require similar preparation of a small RNA library, including small RNA isolation, reverse transcription and ligation with system-specific adaptors. With the Roche 454 system, cDNAs are mixed with agarose beads and clonally amplified by emulsion PCR. The products are randomly deposited to a picotitre plate, and each well holds only one bead, which is pyrosequenced. With the Illumina Solexa platform, single cDNA strands bind to the surface flow cell channels and are amplified in situ by bridge PCR. Using a sequencing-by-synthesis approach, each sequencing cycle adds a mixture of four modified deoxynucleotides that are labelled with four different fluorescence colours. By imaging four channels with laser excitation, the identity of the first base in each cluster is recorded. After removing the fluorescent label, the next cycle is initiated, and the process is repeated until the full oligomer is sequenced. With the SOLiD platform, cDNA is clonally amplified by emulsion PCR on the surface of magnetic beads and subsequently deposited onto the surface of a glass slide. Rather than using polymerase, ligase-mediated sequencing is performed by introducing a primer to the adaptor sequence of cDNA and then ligating a fluorescently labelled eight-base oligonucleotide to the sequencing primer. After the fluorescence signal is recorded with a laser scanner, the octamer is cleaved between the fifth and sixth bases (removing the fluorescent group), and the next round is repeated until the sequence is completed.
R E V I E W S
NATURE REVIEwS | NeuroscieNce VolUmE 11 | mAy 2010 | 333
© 20 Macmillan Publishers Limited. All rights reserved10

SOX9
NSC NPC Neuron
TLX
miR-9
miR-124
a
c
b
Nature Reviews | Neuroscience
RESTSCP1
let-7miR-125miR-9
GFAP+
Nestin+
s100β–
Nestin+
DCX+
PSA–NCAM+
NeuN+
miR-92b miR-124amiR-124miR-128miR-138
YAN mRNA
miR-7 Target genes
Target genes
EGFR
YAN
PNT-P1?
miR-7 gene
ORF AAAA
NSC NPC Mature neuron
in medulloblastoma and promotes GCP proliferation51, suggesting that a feedback loop might exist here as well. This feedback loop has been shown more clearly in D. melanogaster. In the eye of the fruitfly, expression of miR-7 is activated in retinal progenitor cells as they begin to differentiate into photoreceptors52. This process is dependent on epidermal growth factor receptor (EGFR) signalling, which triggers extracellular signal-regulated kinase-mediated degradation of the transcription factor yAN. In non-stimulated cells, yAN represses miR-7 tran-scription, whereas miR-7 RNA represses yAN protein expression in photoreceptors by binding to sequences within its mRNA 3′-UTR52. This reciprocal negative feed-back between yAN and miR-7 ensures mutually exclusive expression, with yAN in progenitor cells and miR-7 in
photoreceptor cells. Expression is switched when EGFR signalling transiently triggers yAN degradation (FIG. 2c). A major question in signal transduction research is how weak or transient activation of signalling pathways can lead to robust and long-term switches in gene expres-sion. These studies suggest that, during neurogenesis, miRNAs may act through feedback loops to reinforce and stabilize changes in gene expression in response to signalling input.
Crosstalk between miRNAs and epigenetic regulation. Epigenetic regulation, including DNA methylation and histone modification, is known to be involved in the modulation of stem cell proliferation and differentia-tion, including the proliferation and differentiation of NSCs53,54. Recent genome-wide analyses have shown a role for DNA methylation and chromatin remodelling, particularly by the polycomb group proteins, in defining the properties and regulating the functions of stem cells55. Epigenetic regulation and chromatin remodel-ling are also thought to have important roles in neuro-genesis56–58; however, the identification of downstream targets has proved elusive59–61.
Functional interactions between small regulatory RNAs and epigenetic mechanisms in gene silencing were initially demonstrated in plants and yeast62, and recently in mammals63. on one hand, epigenetic regu-lation can modulate small RNA expression and, on the other hand, small regulatory RNAs can induce epigenetic changes that lead to mitotically heritable transcriptional silencing62,64. The crosstalk between epigenetic regulation and miRNAs in brain develop-ment and function is now beginning to be investigated. A neuronal activity-dependent miRNA, miR-132, was shown to regulate mECP2 expression in neurons65, and the level of mECP2-regulated miR-184 was increased in neurons following depolarization66. mECP2 belongs to a family of DNA methyl-CpG-binding proteins that interpret DNA methylation as a signal to sup-press gene expression67. De novo mutations in MECP2 cause neurodevelopmental disorders, including Rett’s syndrome67–69. How the loss of functional mECP2 protein leads to neurodevelopmental deficits is still under investigation.
Some studies are now revealing a link between miRNAs and epigenetic regulation in neurogenesis. For example, gliomas retain many features of NPCs, including the ability to grow as neurospheres in culture, self-renew and migrate in the brain. moreover, profiling studies have shown that gliomas express many genes that are expressed by NPCs. miRNA expression profiling of human glioblastoma specimens revealed a marked reduction of miR-128 in tumour samples. miR-128 could reduce glioma cell proliferation by suppressing BmI1, a member of the polycomb repressive complex70. It will be interesting to further examine the role of miR-128 in neurogenesis.
NSC differentiation into neurons is accompanied by a switch of neuronal progenitor-specific BRG1-associated factor (npBAF) to neuron-specific BAF (nBAF) through ATP-dependent chromatin-remodelling mechanisms.
Figure 2 | role of mirNAs in fine-tuning neurogenesis. a | Distinct microRNA (miRNA) expression profiles characterize the transition from neural stem cells (NSCs) to neural progenitor cells (NPCs) and neurons. Marker genes for each of these cell types are listed below the miRNA profiles. b | Schematic illustration of a transcription factor to miRNA to another transcription factor (sometimes even of the miRNA itself) paradigm for regulating neurogenesis. This negative-feedback loop keeps the balance between miRNAs and their transcriptional regulatory programmes during neurogenesis (the blue triangles indicate the expression levels of transcription factors and miRNAs). c | Schematic illustration of the feedback loop between miR-7 and its target, YAN. During eye development in Drosophila melanogaster, progenitor cells express YAN, which directly inhibits miR-7 transcription. When progenitor cells differentiate into photoreceptor neurons, epidermal growth factor receptor (EGFR) signalling is activated, which results in the degradation of YAN. Thus, miR-7 expression is initiated and promotes photoreceptor neuron differentiation by directly repressing YAN. Transcription activator pointed P1 (PNT-P1) can also activate miR-7 expression and contribute to YAN repression. This reciprocal feedback ensures mutually exclusive expression of YAN in progenitor cells and miR-7 in photoreceptor cells. DCX, doublecortin; GFAP, glial fibrillary acidic protein; NeuN, neuronal nuclei; ORF, open reading frame; PSA–NCAM, polysialylated neural cell adhesion molecule; REST, RE1-silencing transcription factor; SCP1, small carboxy-terminal domain phosphatase 1.
R E V I E W S
334 | mAy 2010 | VolUmE 11 www.nature.com/reviews/neuro
© 20 Macmillan Publishers Limited. All rights reserved10

Nature Reviews | Neuroscience
L1 gene
L1 gene Neuronal gene
Altered neuronalgene expression
Small regulatory RNAs
L1 RNA
Neuronaldiversity
??
?
Retrotransposition
A2B5A cell surface ganglioside epitope expressed in developing thymic epithelial cells, oligodendrocyte progenitors and neuroendocrine cells.
Long interspersed nuclear (L1) elementsFull-length active L1 elements are ~6 kb long, consisting of a 5′-untranslated region that has promoter activity, two open reading frames (encoding a nucleic acid-binding protein and an endonuclease), a reverse transcriptase protein and a poly(A) tail.
RetrotransposonGenetic elements that can amplify themselves in a genome through an RNA intermediate. They are ubiquitous components of the DNA of many eukaryotic organisms.
The BAF complex in mammals is functionally related to the SwI–SNF complex in Saccharomyces cerevisiae and D. melanogaster, which is thought to facilitate tran-scriptional activation of specific genes by antagonizing chromatin-mediated transcriptional repression. npBAF has two subunits, BAF53A and BAF45A, which are essential for NSC proliferation; BAF53B and BAF45B, by contrast, are subunits of nBAF that are crucial for dendritic development71. The repression of BAF53A by miR-9* and miR-124 was found to mediate the switch between npBAF and nBAF, suggesting that miRNAs could target chromatin remodelling factors during neuro genesis71. However, whether there is an epige-netic circuitry with a feedback regulatory mechanism mediated by miRNAs in the regulation of neurogenesis remains to be determined.
miRNAs in the specification of gliaProfiling studies have also identified a range of miRNAs that help to establish the lineage specificity for both astrocytes and oligodendrocytes. The phosphorylation of signal transducer and activator of transcription 3 (STAT3) can inhibit the differentiation of NPCs into neurons and promote differentiation into glia. Knockdown of miR-124a and miR-9 increased the level of phosphorylated STAT3, whereas overexpression of miR-124a and miR-9 decreased STAT3 expression, suggesting that these miRNAs could have a role in modulating glial cell fate determination by regulating the levels of STAT3 (ReF. 16). In addition, the expression of miR-23, a miRNA previously implicated in neural specification, was found to be restricted in astrocytes10; however, whether miR-23 is involved in the specification of astrocytes requires further study.
To delineate the regulation of oligodendrocyte differ-entiation by miRNAs, oligodendrocyte progenitor cells (oPCs) and oligodendrocyte lineage cells (olCs) were isolated based on their differential expression of cell sur-face markers by fluorescence-activated cell sorting72. There are dynamic changes in the expression of 43 miRNAs during the transition from A2B5-positive oPCs to pre-myelinating olCs. Combined expression profiling of miRNA and mRNA revealed that miR-9 is downregulated during oligodendrocyte differentiation, which inversely correlates with the expression of its predicted targets, particularly peripheral myelin protein 22 (ReF. 72). Recent studies have identified specific miRNAs involved in the differentiation of oPCs into oligodendrocytes (see Note added in proof).
other small regulatory RNAs in neurogenesisBesides miRNAs, numerous piRNAs and esiRNAs have been identified in genomes73–75. piRNAs in particular have been implicated in controlling the mobilization of transposable elements in mice and D. melanogaster 76. Although they were initially discovered only in repro-ductive systems, mounting evidence suggests that piRNAs and esiRNAs are present in both germline and somatic tissues73–75. In mammals, nearly 15% of piRNAs have been mapped to long interspersed nuclear (L1) elements (also known as lINE-1 elements). l1 elements
are abundant retrotransposons that make up ~17% of the human genome. most l1 elements are retrotrans position defective; nonetheless, there are approximately 100 full-length l1 elements that are potentially capable of retrotransposition in the diploid genome. Intriguingly, recent data indicate that l1 element retrotransposition occurs in rodent and human NPCs, suggesting that l1 elements can have an effect on the expression of neuronal genes and therefore influence neuronal cell fate and contribute to neuronal diversity77–80 (FIG. 3). In addition to the piRNAs that have been mapped to l1 elements, the bidirectional transcripts derived from the l1 5′-UTR can also be processed to siRNAs that sup-press retrotransposition81. It is unclear whether these piRNAs or l1-derived siRNAs can modulate l1 element retrotransposition in adult NSCs and thereby modulate neurogenesis.
PASRs are thought to be involved in the regulation of gene expression82,83. Although the exact biological roles of these small RNAs remain to be determined, the involvement of small RNA-mediated transcriptional regulation in neurogenesis has been established. In this case, a nuclear double-stranded small RNA contain-ing the RE1 sequence that can be recognized by REST was found to be expressed in NSCs. This small RNA induced the neuronal differentiation of NSCs by acti-vating the expression of RE1-containing genes84. It can switch the action of REST from a repressor to an acti-vator, possibly through chromatin modification, and its absence blocks neuronal differentiation, suggesting that it is necessary and sufficient to direct NSCs to the neuronal lineage84.
Figure 3 | A model of neuronal diversity generated by L1 retrotransposition. During neuronal differentiation, long interspersed nuclear (L1) element transcription is activated. This allows subsequent retrotransposition of L1 elements into neuronal genes, which could alter the expression of adjacent neuronal genes and contribute to neuronal diversity. L1 RNAs could be processed into small regulatory RNAs, which could potentially regulate L1 transcription and retrotransposition.
R E V I E W S
NATURE REVIEwS | NeuroscieNce VolUmE 11 | mAy 2010 | 335
© 20 Macmillan Publishers Limited. All rights reserved10

Summary and perspectivesThe discovery of different small regulatory RNAs reveals a new layer of complexity in the regulation of gene expres-sion28,85,86, and it is becoming increasingly clear that small regulatory RNAs are bona fide regulators of neurogenesis. Several principles have emerged from these studies. First, small regulatory RNAs usually function as a fine-tuning mechanism, not as an on–off switch, to ensure the proper progression through each stage of neurogenesis (from NSC self-renewal, to lineage specification following differentia-tion, to the integration of new neurons into existing neural circuits) and to coordinate their action with other biologi-cal regulators such as transcription factors, signalling mol-ecules and epigenetic effectors. Second, several miRNAs can regulate one mRNA, and a single miRNA is capable of regulating many mRNA targets, which could be different components within the same pathway (TABLe 1). Finally, sev-eral feedback regulatory loops exist between miRNAs and their targets, which could ensure the precision of miRNA-mediated gene regulation. This regulatory circuit involves not only the transcription factors, signalling cascades and epigenetic factors discussed above, but also components or regulators of the miRNA pathway itself, such as the feedback loop between let-7 and lIN28 (ReF. 87).
with the emergence of powerful deep-sequencing technologies, there is little doubt that discoveries to date represent the ‘tip of the iceberg’, with many more small regulatory RNAs involved in regulating neuro-genesis awaiting identification. Eventually, the interplay between these small RNAs and different gene regula-tory mechanisms will be systematically unravelled. These discoveries should paint a more complete pic-ture of the intricate and complex web of regulatory net-works through which small regulatory RNAs regulate neurogenesis.
Note added in proofTwo recent studies have shown that the miRNA path-way acts as a ‘micro-brake’ on proliferating oPCs and promotes the switch of oPCs into post-mitotic oligo-dendrocytes102,103. Dicer-knockout mice display severe defects in CNS myelination. miR-219 and miR-338 were found to promote differentiation into oligodendrocytes by repressing several mRNA targets, including those encoding platelet-derived growth factor, SoX6 and HES5. These studies provide compelling evidence that miRNAs are essential for regulating oligodendrocyte differentiation and myelination.
Table 1 | MicroRNAs (miRNAs) involved in neurogenesis and their mRNA targets
mirNA Function in neurogenesis cells and tissues Targets refs
let-7 Promotes neuronal lineage commitment Promotes neural tube closure
NSC LIN28 (mouse) LIN41 (mouse)
87 96
miR-7 Promotes neuronal lineage commitment Photoreceptor cell YAN (Drosophila melanogaster) 52
miR-9 Inhibits proliferation Promotes neuronal lineage commitment Inhibits glial lineage commitment
NSC and NPC ES cell Developing telencephalon
Her5 and Her9 (zebrafish) STAT3 (mouse) FOXG1 (mouse) TLX (mouse) BAF53A (mouse) FGF8 and FGFR1 (mouse) PMP22 (rat)
44 16 45 47 71 44 72
miR-9a Inhibits neuronal fate Epithelial cell Senseless (D. melanogaster) 43
miR-17 Inhibits neural differentiation SH-SY5Y cell Unknown 97
miR-23 Promotes glial lineage commitment Promotes neuronal differentiation
ES cell and P19 cell NT2 cell
Unknown HES1 (human)
10 98
miR-124 Inhibits NSC and NPC proliferation Promotes neuronal lineage commitment
Chicken neural tube P19 cell NSC and NPC
REST–SCP1 (chicken and mouse) PTBP1 (mouse) SOX9 (mouse) BAF53A (mouse)
40 39 37 41 71
miR-125(b) Inhibits NSC and NPC proliferation Promotes neuronal lineage commitment
Medulloblastoma cell and GCP SH-SY5Y cell
GLI1, SMO and LIN28 (mouse and human) Multiple targets (human)
50 99
miR-128 Inhibits NSC and NPC proliferation Glioblastoma BMI1 (human) 70
miR-132 Regulates homeostasis Neuron MECP2 (rat) 65
miR-200 Promotes neuronal differentiation Maintains olfactory progenitor cells
Olfactory progenitor cells Notch and TGFβ signalling pathways and FOXG1 (zebrafish and mouse)
100
miR-279 Promotes neuronal differentiation Sensory organ precursor Nerfin 1 (D. melanogaster) 101
miR-324-5p Inhibits NSC and NPC proliferation Medulloblastoma cell and GCP GLI1 and SMO (mouse and human) 50
miR-326 Inhibits NSC and NPC proliferation Medulloblastoma cell and GCP GLI1 and SMO (mouse and human) 50
ES cell, embryonic stem cell; FGF8, fibroblast growth factor 8; FGFR1, fibroblast growth factor receptor 1; FOXG1, forkhead box protein G1; GCP, granule cell progenitor; MECP2, methyl-CpG-binding protein 2; NPC, neural progenitor cell; NSC, neural stem cell; PMP22, peripheral myelin protein 22; PTBP1, polypyrimidine tract-binding protein 1; REST, RE1-silencing transcription factor; SCP1, small carboxy-terminal domain phosphatase 1; SMO, Smoothened; STAT3, signal transducer and activator of transcription 3; TGFβ, transforming growth factor-β.
R E V I E W S
336 | mAy 2010 | VolUmE 11 www.nature.com/reviews/neuro
© 20 Macmillan Publishers Limited. All rights reserved10

1. Ming, G. L. & Song, H. Adult neurogenesis in the mammalian central nervous system. Annu. Rev. Neurosci. 28, 223–250 (2005).
2. Zhao, C., Deng, W. & Gage, F. H. Mechanisms and functional implications of adult neurogenesis. Cell 132, 645–660 (2008).
3. Mattick, J. S. The functional genomics of noncoding RNA. Science 309, 1527–1528 (2005).
4. Gangaraju, V. K. & Lin, H. MicroRNAs: key regulators of stem cells. Nature Rev. Mol. Cell Biol. 10, 116–125 (2009).
5. Stadler, B. M. & Ruohola-Baker, H. Small RNAs: keeping stem cells in line. Cell 132, 563–566 (2008).
6. Kapsimali, M. et al. MicroRNAs show a wide diversity of expression profiles in the developing and mature central nervous system. Genome Biol. 8, R173 (2007).
7. Krichevsky, A. M., King, K. S., Donahue, C. P., Khrapko, K. & Kosik, K. S. A microRNA array reveals extensive regulation of microRNAs during brain development. RNA 9, 1274–1281 (2003).
8. Lagos-Quintana, M. et al. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735–739 (2002).This is the first paper to report the tissue-specific expression of miRNAs in mammals.
9. Sempere, L. F. et al. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 5, R13 (2004).
10. Smirnova, L. et al. Regulation of miRNA expression during neural cell specification. Eur. J. Neurosci. 21, 1469–1477 (2005).
11. Lagos-Quintana, M., Rauhut, R., Lendeckel, W. & Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 294, 853–858 (2001).
12. Landgraf, P. et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 129, 1401–1414 (2007).
13. Wulczyn, F. G. et al. Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB J. 21, 415–426 (2007).
14. Wienholds, E. & Plasterk, R. H. MicroRNA function in animal development. FEBS Lett. 579, 5911–5922 (2005).
15. Wienholds, E. et al. MicroRNA expression in zebrafish embryonic development. Science 309, 310–311 (2005).
16. Krichevsky, A. M., Sonntag, K. C., Isacson, O. & Kosik, K. S. Specific microRNAs modulate embryonic stem cell-derived neurogenesis. Stem Cells 24, 857–864 (2006).
17. Giraldez, A. J. et al. MicroRNAs regulate brain morphogenesis in zebrafish. Science 308, 833–838 (2005).This study provides the first in vivo evidence that miRNAs are crucial for brain development.
18. Liu, J. et al. Argonaute2 is the catalytic engine of mammalian RNAi. Science 305, 1437–1441 (2004).
19. Cuellar, T. L. et al. Dicer loss in striatal neurons produces behavioral and neuroanatomical phenotypes in the absence of neurodegeneration. Proc. Natl Acad. Sci. USA 105, 5614–5619 (2008).
20. Davis, T. H. et al. Conditional loss of Dicer disrupts cellular and tissue morphogenesis in the cortex and hippocampus. J. Neurosci. 28, 4322–4330 (2008).
21. De Pietri Tonelli, D. et al. miRNAs are essential for survival and differentiation of newborn neurons but not for expansion of neural progenitors during early neurogenesis in the mouse embryonic neocortex. Development 135, 3911–3921 (2008).
22. Kawase-Koga, Y., Otaegi, G. & Sun, T. Different timings of Dicer deletion affect neurogenesis and gliogenesis in the developing mouse central nervous system. Dev. Dyn. 238, 2800–2812 (2009).
23. Gotz, M. & Huttner, W. B. The cell biology of neurogenesis. Nature Rev. Mol. Cell Biol. 6, 777–788 (2005).
24. Neumuller, R. A. & Knoblich, J. A. Dividing cellular asymmetry: asymmetric cell division and its implications for stem cells and cancer. Genes Dev. 23, 2675–2699 (2009).
25. Zhong, W. & Chia, W. Neurogenesis and asymmetric cell division. Curr. Opin. Neurobiol. 18, 4–11 (2008).
26. Schwamborn, J. C., Berezikov, E. & Knoblich, J. A. The TRIM-NHL protein TRIM32 activates microRNAs and prevents self-renewal in mouse neural progenitors. Cell 136, 913–925 (2009).This paper was the first to show that a miRNA-containing ribonucleoprotein complex could be unequally distributed and contribute to asymmetrical cell division.
27. Loedige, I. & Filipowicz, W. TRIM-NHL proteins take on miRNA regulation. Cell 136, 818–820 (2009).
28. Cao, X., Yeo, G., Muotri, A. R., Kuwabara, T. & Gage, F. H. Noncoding RNAs in the mammalian central nervous system. Annu. Rev. Neurosci. 29, 77–103 (2006).
29. Fineberg, S. K., Kosik, K. S. & Davidson, B. L. MicroRNAs potentiate neural development. Neuron 64, 303–309 (2009).
30. Kosik, K. S. The neuronal microRNA system. Nature Rev. Neurosci. 7, 911–920 (2006).
31. Schratt, G. microRNAs at the synapse. Nature Rev. Neurosci. 10, 842–849 (2009).
32. Coolen, M. & Bally-Cuif, L. MicroRNAs in brain development and physiology. Curr. Opin. Neurobiol. 19, 461–470 (2009).
33. Chang, S., Johnston, R. J. Jr, Frokjaer-Jensen, C., Lockery, S. & Hobert, O. MicroRNAs act sequentially and asymmetrically to control chemosensory laterality in the nematode. Nature 430, 785–789 (2004).
34. Johnston, R. J. & Hobert, O. A microRNA controlling left/right neuronal asymmetry in Caenorhabditis elegans. Nature 426, 845–849 (2003).References 33 and 34 provide the first evidence that miRNAs regulate neuronal fate determination.
35. Johnston, R. J. Jr, Chang, S., Etchberger, J. F., Ortiz, C. O. & Hobert, O. MicroRNAs acting in a double-negative feedback loop to control a neuronal cell fate decision. Proc. Natl Acad. Sci. USA 102, 12449–12454 (2005).
36. Lim, L. P. et al. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 433, 769–773 (2005).
37. Makeyev, E. V., Zhang, J., Carrasco, M. A. & Maniatis, T. The microRNA miR-124 promotes neuronal differentiation by triggering brain-specific alternative pre-mRNA splicing. Mol. Cell 27, 435–448 (2007).
38. Ballas, N., Grunseich, C., Lu, D. D., Speh, J. C. & Mandel, G. REST and its corepressors mediate plasticity of neuronal gene chromatin throughout neurogenesis. Cell 121, 645–657 (2005).
39. Visvanathan, J., Lee, S., Lee, B., Lee, J. W. & Lee, S. K. The microRNA miR-124 antagonizes the anti-neural REST/SCP1 pathway during embryonic CNS development. Genes Dev. 21, 744–749 (2007).
40. Conaco, C., Otto, S., Han, J. J. & Mandel, G. Reciprocal actions of REST and a microRNA promote neuronal identity. Proc. Natl Acad. Sci. USA 103, 2422–2427 (2006).
41. Cheng, L. C., Pastrana, E., Tavazoie, M. & Doetsch, F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nature Neurosci. 12, 399–408 (2009).
42. Kloosterman, W. P. et al. Cloning and expression of new microRNAs from zebrafish. Nucleic Acids Res. 34, 2558–2569 (2006).
43. Li, Y., Wang, F., Lee, J. A. & Gao, F. B. MicroRNA-9a ensures the precise specification of sensory organ precursors in Drosophila. Genes Dev. 20, 2793–2805 (2006).
44. Leucht, C. et al. MicroRNA-9 directs late organizer activity of the midbrain–hindbrain boundary. Nature Neurosci. 11, 641–648 (2008).
45. Shibata, M., Kurokawa, D., Nakao, H., Ohmura, T. & Aizawa, S. MicroRNA-9 modulates Cajal-Retzius cell differentiation by suppressing Foxg1 expression in mouse medial pallium. J. Neurosci. 28, 10415–10421 (2008).
46. Sun, G., Yu, R. T., Evans, R. M. & Shi, Y. Orphan nuclear receptor TLX recruits histone deacetylases to repress transcription and regulate neural stem cell proliferation. Proc. Natl Acad. Sci. USA 104, 15282–15287 (2007).
47. Zhao, C., Sun, G., Li, S. & Shi, Y. A feedback regulatory loop involving microRNA-9 and nuclear receptor TLX in neural stem cell fate determination. Nature Struct. Mol. Biol. 16, 365–371 (2009).
48. Ruiz i Altaba, A., Palma, V. & Dahmane, N. Hedgehog–Gli signalling and the growth of the brain. Nature Rev. Neurosci. 3, 24–33 (2002).
49. Decembrini, S. et al. MicroRNAs couple cell fate and developmental timing in retina. Proc. Natl Acad. Sci. USA 106, 21179–21184 (2009).
50. Ferretti, E. et al. Concerted microRNA control of Hedgehog signalling in cerebellar neuronal progenitor and tumour cells. EMBO J. 27, 2616–2627 (2008).
51. Northcott, P. A. et al. The miR-17/92 polycistron is up-regulated in sonic hedgehog-driven medulloblastomas and induced by N-myc in sonic hedgehog-treated cerebellar neural precursors. Cancer Res. 69, 3249–3255 (2009).
52. Li, X. & Carthew, R. W. A microRNA mediates EGF receptor signaling and promotes photoreceptor differentiation in the Drosophila eye. Cell 123, 1267–1277 (2005).This paper was the first to show that miRNAs regulate neuronal fate through EGF receptor signalling.
53. Li, X. & Zhao, X. Epigenetic regulation of mammalian stem cells. Stem Cells Dev. 17, 1043–1052 (2008).
54. Mehler, M. F. Epigenetics and the nervous system. Ann. Neurol. 64, 602–617 (2008).
55. Bernstein, B. E., Meissner, A. & Lander, E. S. The mammalian epigenome. Cell 128, 669–681 (2007).
56. Lim, D. A. et al. Chromatin remodelling factor Mll1 is essential for neurogenesis from postnatal neural stem cells. Nature 458, 529–533 (2009).
57. Ma, D. K. et al. Neuronal activity-induced Gadd45b promotes epigenetic DNA demethylation and adult neurogenesis. Science 323, 1074–1077 (2009).
58. Zhao, X. et al. Mice lacking methyl-CpG binding protein 1 have deficits in adult neurogenesis and hippocampal function. Proc. Natl Acad. Sci. USA 100, 6777–6782 (2003).References 56–58 show the key role of epigenetic regulation in neurogenesis.
59. Cheng, L. C., Tavazoie, M. & Doetsch, F. Stem cells: from epigenetics to microRNAs. Neuron 46, 363–367 (2005).
60. Hsieh, J. & Gage, F. H. Epigenetic control of neural stem cell fate. Curr. Opin. Genet. Dev. 14, 461–469 (2004).
61. Abel, T. & Zukin, R. S. Epigenetic targets of HDAC inhibition in neurodegenerative and psychiatric disorders. Curr. Opin. Pharmacol. 8, 57–64 (2008).
62. Tijsterman, M., Ketting, R. F. & Plasterk, R. H. A. The genetics of RNA silencing. Annu. Rev. Genet. 36, 489–519 (2002).
63. Morris, K. V., Chan, S. W., Jacobsen, S. E. & Looney, D. J. Small interfering RNA-induced transcriptional gene silencing in human cells. Science 305, 1289–1292 (2004).
64. Kawasaki, H. & Taira, K. Induction of DNA methylation and gene silencing by short interfering RNAs in human cells. Nature 431, 211–217 (2004).
65. Klein, M. E. et al. Homeostatic regulation of MeCP2 expression by a CREB-induced microRNA. Nature Neurosci. 10, 1513–1514 (2007).
66. Nomura, T. et al. MeCP2-dependent repression of an imprinted miR-184 released by depolarization. Hum. Mol. Genet. 17, 1192–1199 (2008).
67. Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 16, 6–21 (2002).
68. Amir, R. E. et al. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nature Genet. 23, 185–188 (1999).
69. Chahrour, M. & Zoghbi, H. Y. The story of Rett syndrome: from clinic to neurobiology. Neuron 56, 422–437 (2007).
70. Godlewski, J. et al. Targeting of the Bmi-1 oncogene/stem cell renewal factor by microRNA-128 inhibits glioma proliferation and self-renewal. Cancer Res. 68, 9125–9130 (2008).
71. Yoo, A. S., Staahl, B. T., Chen, L. & Crabtree, G. R. MicroRNA-mediated switching of chromatin-remodelling complexes in neural development. Nature 460, 642–646 (2009).
72. Lau, P. et al. Identification of dynamically regulated microRNA and mRNA networks in developing oligodendrocytes. J. Neurosci. 28, 11720–11730 (2008).The first paper to report the potential involvement of miRNAs in the development of oligodendrocytes.
73. Kawamura, Y. et al. Drosophila endogenous small RNAs bind to Argonaute 2 in somatic cells. Nature 453, 793–797 (2008).
74. Czech, B. et al. An endogenous small interfering RNA pathway in Drosophila. Nature 453, 798–802 (2008).
75. Okamura, K. & Lai, E. C. Endogenous small interfering RNAs in animals. Nature Rev. Mol. Cell Biol. 9, 673–678 (2008).
76. Aravin, A. A., Hannon, G. J. & Brennecke, J. The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science 318, 761–764 (2007).
77. Coufal, N. G. et al. L1 retrotransposition in human neural progenitor cells. Nature 460, 1127–1131 (2009).
R E V I E W S
NATURE REVIEwS | NeuroscieNce VolUmE 11 | mAy 2010 | 337
© 20 Macmillan Publishers Limited. All rights reserved10

78. Muotri, A. R. et al. Somatic mosaicism in neuronal precursor cells mediated by L1 retrotransposition. Nature 435, 903–910 (2005).References 77 and 78 show that retrotransposition of the L1 element occurs in human and rodent NPCs, which could contribute to neuronal diversity.
79. Kuwabara, T. et al. Wnt-mediated activation of NeuroD1 and retro-elements during adult neurogenesis. Nature Neurosci. 12, 1097–1105 (2009).
80. Muotri, A. R. & Gage, F. H. Generation of neuronal variability and complexity. Nature 441, 1087–1093 (2006).
81. Yang, N. & Kazazian, H. H. Jr. L1 retrotransposition is suppressed by endogenously encoded small interfering RNAs in human cultured cells. Nature Struct. Mol. Biol. 13, 763–771 (2006).
82. Kapranov, P. et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 316, 1484–1488 (2007).
83. Taft, R. J. et al. Tiny RNAs associated with transcription start sites in animals. Nature Genet. 41, 572–578 (2009).
84. Kuwabara, T., Hsieh, J., Nakashima, K., Taira, K. & Gage, F. H. A small modulatory dsRNA specifies the fate of adult neural stem cells. Cell 116, 779–793 (2004).
85. Bushati, N. & Cohen, S. M. microRNA functions. Annu. Rev. Cell Dev. Biol. 23, 175–205 (2007).
86. Bartel, D. P. MicroRNAs: target recognition and regulatory functions. Cell 136, 215–233 (2009).
87. Rybak, A. et al. A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nature Cell Biol. 10, 987–993 (2008).
88. Sawamoto, K. et al. New neurons follow the flow of cerebrospinal fluid in the adult brain. Science 311, 629–632 (2006).
89. Shen, Q. et al. Adult SVZ stem cells lie in a vascular niche: a quantitative analysis of niche cell-cell interactions. Cell Stem Cell 3, 289–300 (2008).
90. Tavazoie, M. et al. A specialized vascular niche for adult neural stem cells. Cell Stem Cell 3, 279–288 (2008).References 89 and 90 provide evidence that the vasculature is an important component of neurogenic niches in the adult mammalian brain.
91. Ghildiyal, M. & Zamore, P. D. Small silencing RNAs: an expanding universe. Nature Rev. Genet. 10, 94–108 (2009).
92. Kim, V. N., Han, J. & Siomi, M. C. Biogenesis of small RNAs in animals. Nature Rev. Mol. Cell Biol. 10, 126–139 (2009).
93. Mardis, E. R. Next-generation DNA sequencing methods. Annu. Rev. Genomics Hum. Genet. 9, 387–402 (2008).
94. Metzker, M. L. Sequencing technologies — the next generation. Nature Rev. Genet. 11, 31–46 (2010).
95. Rusk, N. & Kiermer, V. Primer: Sequencing — the next generation. Nature Methods 5, 15 (2008).
96. Maller Schulman, B. R. et al. The let-7 microRNA target gene, Mlin41/Trim71 is required for mouse embryonic survival and neural tube closure. Cell Cycle 7, 3935–3942 (2008).
97. Beveridge, N. J., Tooney, P. A., Carroll, A. P., Tran, N. & Cairns, M. J. Down-regulation of miR-17 family expression in response to retinoic acid induced neuronal differentiation. Cell Signal. 21, 1837–1845 (2009).
98. Kawasaki, H. & Taira, K. Hes1 is a target of microRNA-23 during retinoic-acid-induced neuronal differentiation of NT2 cells. Nature 423, 838–842 (2003).
99. Le, M. T. et al. MicroRNA-125b promotes neuronal differentiation in human cells by repressing multiple targets. Mol. Cell. Biol. 29, 5290–5305 (2009).
100. Choi, P. S. et al. Members of the miRNA-200 family regulate olfactory neurogenesis. Neuron 57, 41–55 (2008).
101. Cayirlioglu, P. et al. Hybrid neurons in a microRNA mutant are putative evolutionary intermediates in insect CO2 sensory systems. Science 319, 1256–1260 (2008).
102. Dugas, J. C. et al. Dicer1 and miR-219 are required for normal oligodendrocyte differentiation and myelination. Neuron 65, 597–611 (2010).
103. Zhao, X. et al. MicroRNA-mediated control of oligodendrocyte differentiation. Neuron 65, 612–626 (2010).
AcknowledgementsWe thank C. Strauss for critical reading of the manuscript. We apologize to those whose works are not cited here owing space limitations. The work in our laboratory was supported in part by the grants from the National Institutes of Health and International Rett Syndrome Foundation. P.J. is a recipi-ent of the Beckman Young Investigator Award and the Basil O’Connor Scholar Research Award, as well as an Alfred P. Sloan Research Fellowship in Neuroscience. X.L. is supported by a FRAXA Fellowship.
Competing interests statementThe authors declare no competing financial interests.
DATABASESOMIM: http://www.ncbi.nlm.nih.gov/omimRett’s syndromeUniProtKB: http://www.uniprot.orgMECP2 | PTBP1 | REST | SCP1 | STAT3 | TLX | TRIM32
FuRTHER INFoRMATIoNPeng Jin’s homepage: http://genetics.emory.edu/labs/jinlab
ALL LiNks Are AcTive iN The oNLiNe pdF
R E V I E W S
338 | mAy 2010 | VolUmE 11 www.nature.com/reviews/neuro
© 20 Macmillan Publishers Limited. All rights reserved10