s equoia maguanensis m the s china: i 1 · abundance of more recent apparent records from the...
TRANSCRIPT

103
American Journal of Botany 102 ( 1 ): 103 – 118 , 2015 .
American Journal of Botany 102 ( 1 ): 103 – 118 , 2015 ; http://www.amjbot.org/ © 2015 Botanical Society of America
Sequoia sempervirens (D. Don) Endlicher, the coast red-wood, is an evergreen conifer and the only relict species of the genus Sequoia Endlicher (Cupressaceae sensu lato). Presently, it grows along a narrow strip of land along the Pacifi c coast of northern California and southern Oregon in the United States ( Watson and Eckenwalder, 1993 ). This strip of land is approxi-mately 724 km long and 8–56 km wide, characterized by mod-erate to heavy winter rain and summer fog, which are vital to this species ( Olson et al., 1990 ; Humboldt County Visitors Bureau, 2014 ). However, the paleogeographical origin of S. sem-pervirens has remained controversial. Fossil records show that
the origin of the foliage characteristics in this species is no older than the Neogene ( Fields, 1993 ; Ahuja and Neale, 2002 ). Al-though several fossils from the Miocene of eastern Asia have been considered to have affi nities with the modern S. sempervi-rens , their leafy shoots have no dimorphic leaves and lack scale leaves ( Endo, 1933 ; Ma et al., 2005 ), or the leaves have no epi-dermal structures ( Endo, 1933 ). The fertile shoots with cones in attachment are missing ( Endo, 1933 ; Ma et al., 2005 ), and thus the structure and shape of the cones are unknown.
Fossils that have been purported to be Sequoia have been reported from nearly every continent in the world. The oldest representatives were from the Middle Mesozoic, e.g., the im-pression of a leafy twig from the Middle to Late Jurassic of Northeast China ( Endo, 1951 ) and seed cones from the Jurassic of Boulogne, France ( Fliche and Zeiller, 1904 ). There is an abundance of more recent apparent records from the Creta-ceous to Neogene of North America, Eurasia, and Australia ( Velenovsky, 1885 ; Endo, 1928 , 1933 ; Florin, 1931 , 1963 ; Miki, 1941 ; Chaney, 1951 ; Miki and Hikita, 1951 ; Sveshnikova, 1967 ; Kilpper, 1968 ; Knobloch, 1969 ; Sveshnikova and Budantsev, 1969 ; Dorofeev, 1975 ; Miller, 1977 ; Mai and Walther, 1978 ; Peters and Christophel, 1978 ; WGCPC, 1978 ; Chen et al., 1988 ; Samylina, 1988 ; Fields, 1993 ; Hill et al., 1993 ; Hably, 1994 ; Palamarev, 1994 ; Mai, 1995 ; Pole, 1995 ; Meyer and Manchester, 1997 ; Barrón and Diéguez, 2001 ; Hill, 2001 ;
1 Manuscript received 4 August 2014; revision accepted 8 December 2014. This work was supported by the National Natural Science Foundation of
China (41372035) and the Chinese Academy of Sciences 135 program (XTBG-F01). The authors thank Prof. S. R. Manchester, Florida Museum of Natural History, USA, and Dr. Q. W. Ma, Beijing Museum of Natural History, China, for improving the manuscript, Dr. N. Nosova, Komarov Botanical Institute of the Russian Academy of Sciences, and Dr. L. Wang from XTBG, for providing some references, and staff at the Laboratory Center of XTBG for preparing the scanning electron micrographs of cuticles. This work is a contribution to NECLIME (Neogene Climate Evolution in Eurasia).
6 Author for correspondence (e-mail: [email protected])
doi:10.3732/ajb.1400347
SEQUOIA MAGUANENSIS , A NEW MIOCENE RELATIVE OF THE COAST REDWOOD, SEQUOIA SEMPERVIRENS , FROM
CHINA: IMPLICATIONS FOR PALEOGEOGRAPHY AND PALEOCLIMATE 1
JIAN-WEI ZHANG 2 , ASHALATA D’ROZARIO 3 , JONATHAN M. ADAMS 4 , YA LI 5 , XIAO-QING LIANG 2 , FRÉDÉRIC M. JACQUES 2 , TAO SU 2 , AND ZHE-KUN ZHOU 2,6
2 Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, the Chinese Academy of Sciences, Mengla 666303, China; 3 Department of Botany, Narasinha Dutt College, 129 Bellilious Road, Howrah 711101, India; 4 The college of Natural Sciences, Seoul National University, 1 Gwanak-ro, Gwanak-gu, Seoul 151–742, Korea; and 5 Institute of
Geology, the Chinese Academy of Geological Sciences, 26, Baiwanzhuang Road, Beijing 100037, China
• Premise of the study: The paleogeographical origin of the relict North American Sequoia sempervirens is controversial. Fossil records indicate a Neogene origin for its foliage characteristics. Although several fossils from the Miocene sediments in eastern Asia have been considered to have close affi nities with the modern S. sempervirens , they lack the typical features of a leafy twig bearing linear as well as scale leaves, and the fertile shoots terminating by a cone. The taxonomic status of these fossils has remained unclear.
• Methods: New better-preserved fossils from the upper Miocene of China indicate a new species of Sequoia . This fi nding not only confi rms the former presence of this genus in eastern Asia, but it also confi rms the affi nity of this Asian form to the modern relict S. sempervirens .
• Key results: The principal foliage characteristics of S. sempervirens had already originated by the late Miocene. The eastern Asian records probably imply a Beringian biogeographic track of the ancestor of S. sempervirens in the early Neogene, at a time when the land bridge was not too cool for this thermophilic conifer to spread between Asia and North America.
• Conclusions: The climatic context of the new fossil Sequoia in Southeast Yunnan, based on other fl oristic elements of the fossil assemblage in which it is found, is presumed to be warm and humid. Following the uplift of the Qinghai-Tibet Plateau, this warm, humid climate was replaced by the present monsoonal climate with dry winter and spring. This change may have led to the disappearance of this hygrophilous conifer from eastern Asia.
Key words: China; comparative morphology; Cupressaceae; fossil; late Miocene; monsoon climate; phytogeography; Sequoia sempervirens ; North America.

104 AMERICAN JOURNAL OF BOTANY [Vol. 102
previously assigned to the Huazhige Formation ( Zhang, 1976 ; BGMRYP, 1990 ). Recently, they have been assigned to the Xiaolongtan Formation ( BGM-RYP, 1996 ; Zheng et al., 1999 ).
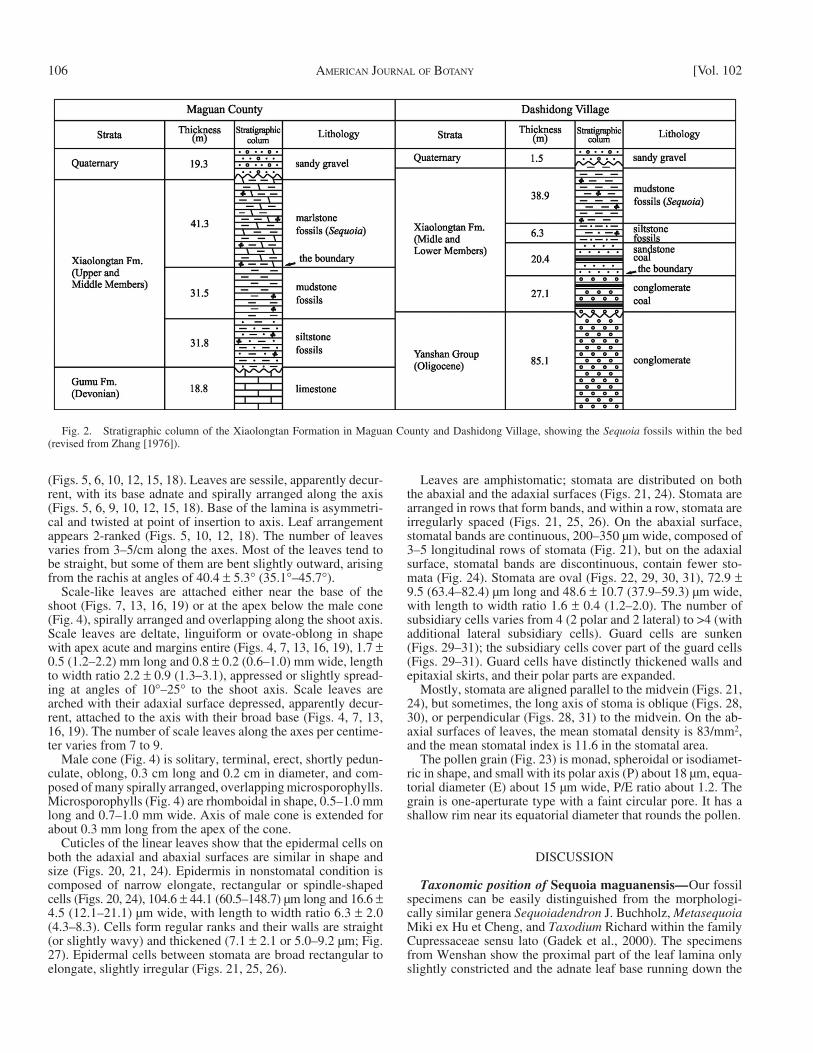
Exposure of the Xiaolongtan Formation around Wenshan State is about 200–300 m thick and generally divided into three members: the Upper Member (about 41.3 m thick) is composed of thin layers (<0.1 m) of gray to white-gray marlite; the Middle Member (63.3–65.6 m thick) consists of thin to medium layers (<0.5 m) of gray-green to gray-yellow mudstone or fi ne sandstone, with interlayers of coal; and the Lower Member (27.1 m thick) comprises medium layers (0.1–0.5 m) of gray to dark-gray conglomerate or sandstone. The Upper and Middle Members lie primarily around the Maguan County and the Middle and Lower Members around the Wenshan City ( Zhang, 1976 ; Zheng et al., 1999 ) ( Fig. 2 ). The present Sequoia fossils were collected from the Upper and Middle Members of the formation. The Xiaolongtan Formation is late Miocene in age based on lithology, biologic assemblage, and regional comparisons ( Dong, 1987 ; BGMRYP, 1996 ). It lies unconformably above the upper Oligo-cene of Yanshan Group and is overlain unconformably by the Quaternary strata ( Zhang, 1976 ).
The sedimentary sequences in Maguan County and Dashidong Village in Wenshan State ( Fig. 2 ) yield abundant plant fossils, such as Quercus L., Salix L., Dryophyllum Debey ex Saporta, Ulmus L., Zelkova Spach, Ampelopsis Mi-chaux, Glyptostrobus Endlicher ( Zhang, 1976 ), Ailanthus confucii Unger ( Su et al., 2013 ), Bauhinia Wenshanensis Meng et Zhou ( Meng et al., 2014 ), Pinus massoniana Lambert, and Sequoia , and fossil fi sh and insects (e.g., mosquitos, ants).
Fossil specimens — Fossil materials (DMS 0130–0134, 5 specimens) de-scribed in the present paper consist of compressions, represented by vegetative twigs as well as a foliaged shoot bearing a male cone. Fossil specimens are deposited in the fossil repository of the Paleoecology Research Group, Xish-uangbanna Tropical Botanical Garden, the Chinese Academy of Sciences.
Leaf morphology and cuticle — Photographs were obtained using a Nikon D700 digital camera (Nikon, Kanagawa, Japan) under tungsten light. Some specimens were immersed in kerosene to enhance details. Detailed structures of these specimens were observed and photographed with a Leica S8AP0 stereo-scope microscope (Leica, Wetzlar, Germany). Fossil materials were treated with Schulze’s solution (HNO 3 and KClO 3 , 3 : 1) and KOH, and washed with water to obtain clear cuticles ( Kerp, 1990 ). Epidermal characters were observed with a Leica DM750 stereomicroscope and photographed with a Leica DFC295 and Zeiss EVOLS10 scanning electron microscope (Carl Zeiss, Jena, Germany).
Pollen grains — Pollen grains were extracted from the male cone. The male cone was detached from the foliage shoot and put into HCl (10%) solution to remove the surrounding rock matrix. The clean cone was then soaked in Schul-ze’s solution for 12 h and crushed using a glass rod. The mixed liquor that suspended solids was fi ltrated using a fi ne sieve, and the residue was washed and transferred to a glass slide. Numerous pollen grains were recovered. They were observed with a Leica DM750 stereomicroscope and photographed with a Leica DFC295.
RESULTS
Systematics — Family— Cupressaceae sensu lato.
Subfamily— Sequoioideae (Luerss.) Quinn.
Genus— Sequoia Endlicher.
Type species— Sequoia sempervirens (D. Don) Endlicher.
S. maguanensis— J.W. Zhang et Z.K. Zhou, sp. nov. ( Figs. 3, 8, 11, 14, 17 ).
Specifi c diagnosis— Fertile and sterile shoots with dimorphic leaves, one with an expanded blade and another scale-like. The
Kovar-Eder et al., 2001 ; Spicer et al., 2002 ; Jungwirth, 2004 ; Ma et al., 2005 ; Palamarev et al., 2005 ; Grimsson et al., 2007 ; Kva č ek and Teodoridis, 2007 ; Worobiec et al., 2008 ; Sawangchote et al., 2009 ; Bozukov and Tsenov, 2012 ; Golovneva and Nosova, 2012 ; Holý et al., 2012 ). However, these supposed Sequoia fossils were often only preserved as isolated shoots or cones, and most of them have not yet been confi rmed on the basis of detailed anatomical characters ( Miller, 1977 ; Sawyer et al., 2000 ).
Additional Sequoia -like remains were assigned to the genera Sequoiites Brongniart ( Seward, 1919 ; Miller, 1977 ), Sequoiop-sis Velenovsky et Viniklar ( Miller, 1977 ), Parasequoia Krasilov ( Blazer, 1975 ), Austrosequoia Peters et Christophel ( Peters and Christophel, 1978 ), Quasisequoia Srinivasan et Friis emend. Kunzmann ( Srinivasan and Friis, 1989 ; Kunzmann, 1999 ), and Sequoiaoxylon Andrews ( Andrews, 1936 ). All these have uncertain affi nities with the modern-day Sequoia .
Most fi rmly identifi ed Sequoia fossils have been recognized from the Paleogene and early Neogene sediments of mid-north-ern latitudes ( Miller, 1977 ; Sawyer et al., 2000 ). Fossils of Se-quoia after the late Miocene are scarce and absent from Asia and Europe in the late Pliocene. Since at least that time, Sequoia has existed at its present location in western North America ( Sakai, 1971 ).
On the basis of the most well-established fossils among these records, the modern species S. sempervirens is thought to have had a Neogene origin because its foliage characteris-tics are more similar to Neogene forms than to Paleogene ones ( Ahuja and Neale, 2002 ). The extant species of Sequoia dif-fers quite clearly from Sequoia fossils of Eocene and Oligocene, but only slightly from Miocene fossils ( Fields, 1993 ). For ex-ample, the fossils from the Eocene–Oligocene bear narrow, appressed leaves and small cones, while the Miocene materi-als have comparatively longer leaves and larger cones and ap-pear much more similar to the extant species ( Ahuja and Neale, 2002 ). The early Pliocene materials from Japan show no signifi cant differences with the extant species in the mor-phological characters of cones, foliated shoots, epidermal structures, and even in the size of guard cells ( Miki, 1941 ; Miki and Hikita, 1951 ).
Female cones and foliage compressions of Sequoia cf. sem-pervirens have been reported from the Miocene of Chuxiong City, Central Yunnan ( Ma et al., 2005 ). In this paper, new fossil materials of Sequoia have been found from the upper Miocene of Southeast Yunnan, Southwest China. These remains are rep-resented by foliated shoots with linear and scale leaves, which have well-preserved cuticles. Additionally, one foliated shoot bearing a male cone at the apex was also found. In the present study, we describe a new species, S. maguanensis sp. nov., based on comparisons with related fossil and extant species of Sequoia . We also explore its biogeographic and paleoclimatic implications as well as the probable reasons for the disappear-ance of Sequoia from eastern Asia.
MATERIALS AND METHODS
Geological settings and age — Fossils of Sequoia were collected from out-crops located in Maguan County (23 ° 00 ′ N, 104 ° 23 ′ E, 1320 m a.s.l.) and Dashi-dong Village (23 ° 15 ′ N, 104 ° 15 ′ E, 1482 m a.s.l.), Wenshan State, SE Yunnan Province, SW China ( Fig. 1 ). The outcrops in both areas are characterized by light-gray or light-yellow pelitic laminated siltstone and mudstone. The lami-nated sedimentary sequences in Maguan County and Dashidong Village were

ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 105January 2015]
Paratypes— DMS 0131 ( Fig. 8 ), DMS 0132 ( Fig. 11 ), DMS 0133 ( Fig. 14 ), DMS 0134 ( Fig. 17 ) (designated here).
Type locality— Maguan County, Wenshan City, Yunnan Province, China.
Age and stratigraphy— Late Miocene, Xiaolongtan Formation.
Description— Foliated shoots are 4.7 ± 1.5 (3.2–6.2) cm long and 1.1 ± 0.2 (0.9–1.3) cm wide, length to width ratio 4.1 ± 0.8 (3.3–4.9). The axis of the shoots is 0.9–1.1 mm wide. Leaves are dimorphic, one with expanded lamina and another scale-like ( Figs. 3, 8, 11, 14, 17 ).
The expanded leaves are 2-ranked ( Figs. 5, 6, 9, 10, 12, 15, 18 ), attached obliquely to the axis of the shoot, linear or linear-lanceolate in shape, 7.9 ± 1.3 (6.6–9.2) mm long, 1.0 ± 0.1 (0.9–1.1) mm wide, length to width ratio 8.3 ± 1.3 (7.0 –9.6). Laminas are bifacially fl attened with smooth surface, entire margins and acute apex. Abaxial surface of leaf has a strong median vein appearing as a low ridge ( Fig. 10 ) and is clearly visible in the proximal part, less in the distal apical part ( Figs. 5, 6, 12, 15, 18 ). Abaxial surface of the leaf is planar or slightly concave; adaxial surface planar or slightly convex, not keeled,
expanded leaves linear or linear-lanceolate, 7.9 ± 1.3 (6.6–9.2) mm long, 1.0 ± 0.1 (0.9–1.1) mm wide, length to width ratio 8.3 ± 1.3 (7.0–9.6), lamina bifacially fl attened, margins entire, apex acute, base adnate, abaxial surface with a strong median vein appearing as a low ridge, spirally arranged along the axis, twisted, appearing 2-ranked, attached obliquely to the axis of the shoot. Scale-like leaves attached either near the base of the shoot or at the apex, below the male cone, spirally arranged, overlapping, deltate, linguiform or ovate-oblong in shape, 1.7 ± 0.5 (1.2–2.2) mm long, 0.8 ± 0.2 (0.6–1.0) mm wide, length to width ratio 2.2 ± 0.9 (1.3–3.1), apex acute, margins entire. Male cone solitary, oblong, 0.3 cm long and 0.2 cm in diameter. Micro-sporophylls rhomboidal, bearing small, monad-type pollen. Cells of epidermis often form the upper ranks, rectangular or spindle-shaped, walls straight or slightly wavy. Stomata distributed on both abaxial and adaxial surfaces, ovoid, subsidiary cells vary-ing from 4 to 6. Guard cells sunken, with distinctly thickened wall and epitaxial skirts, polar parts expanded. Long axis of stoma parallel, oblique, or perpendicular to the midvein.
Derivation of specifi c epithet— The specifi c name, magua-nensis , is derived from the fossil locality, China.
Holotype— DMS 0130 ( Fig. 3 ) (designated here).
Fig. 1. Maps of localities for Sequoia maguanensis sp. nov. in Southeast Yunnan, Southwest China.
106 AMERICAN JOURNAL OF BOTANY [Vol. 102
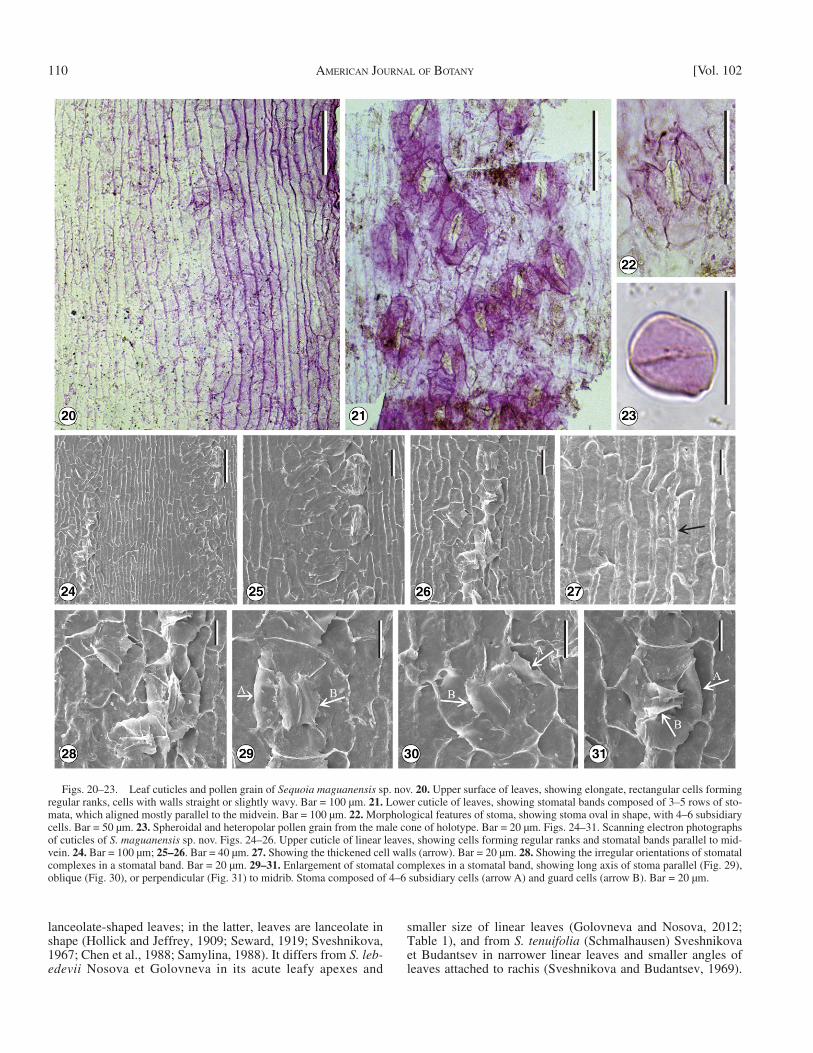
Leaves are amphistomatic; stomata are distributed on both the abaxial and the adaxial surfaces ( Figs. 21, 24 ). Stomata are arranged in rows that form bands, and within a row, stomata are irregularly spaced ( Figs. 21, 25, 26 ). On the abaxial surface, stomatal bands are continuous, 200–350 µm wide, composed of 3–5 longitudinal rows of stomata ( Fig. 21 ), but on the adaxial surface, stomatal bands are discontinuous, contain fewer sto-mata ( Fig. 24 ). Stomata are oval ( Figs. 22, 29, 30, 31 ), 72.9 ± 9.5 (63.4–82.4) µm long and 48.6 ± 10.7 (37.9–59.3) µm wide, with length to width ratio 1.6 ± 0.4 (1.2–2.0). The number of subsidiary cells varies from 4 (2 polar and 2 lateral) to >4 (with additional lateral subsidiary cells). Guard cells are sunken ( Figs. 29–31 ); the subsidiary cells cover part of the guard cells ( Figs. 29–31 ). Guard cells have distinctly thickened walls and epitaxial skirts, and their polar parts are expanded.
Mostly, stomata are aligned parallel to the midvein ( Figs. 21, 24 ), but sometimes, the long axis of stoma is oblique ( Figs. 28, 30 ), or perpendicular ( Figs. 28, 31 ) to the midvein. On the ab-axial surfaces of leaves, the mean stomatal density is 83/mm 2 , and the mean stomatal index is 11.6 in the stomatal area.
The pollen grain ( Fig. 23 ) is monad, spheroidal or isodiamet-ric in shape, and small with its polar axis (P) about 18 µm, equa-torial diameter (E) about 15 µm wide, P/E ratio about 1.2. The grain is one-aperturate type with a faint circular pore. It has a shallow rim near its equatorial diameter that rounds the pollen.
DISCUSSION
Taxonomic position of Sequoia maguanensis — Our fossil specimens can be easily distinguished from the morphologi-cally similar genera Sequoiadendron J. Buchholz, Metasequoia Miki ex Hu et Cheng, and Taxodium Richard within the family Cupressaceae sensu lato ( Gadek et al., 2000 ). The specimens from Wenshan show the proximal part of the leaf lamina only slightly constricted and the adnate leaf base running down the
( Figs. 5, 6, 10, 12, 15, 18 ). Leaves are sessile, apparently decur-rent, with its base adnate and spirally arranged along the axis ( Figs. 5, 6, 9, 10, 12, 15, 18 ). Base of the lamina is asymmetri-cal and twisted at point of insertion to axis. Leaf arrangement appears 2-ranked ( Figs. 5, 10, 12, 18 ). The number of leaves varies from 3–5/cm along the axes. Most of the leaves tend to be straight, but some of them are bent slightly outward, arising from the rachis at angles of 40.4 ± 5.3 ° (35.1 ° –45.7 ° ).
Scale-like leaves are attached either near the base of the shoot ( Figs. 7, 13, 16, 19 ) or at the apex below the male cone ( Fig. 4 ), spirally arranged and overlapping along the shoot axis. Scale leaves are deltate, linguiform or ovate-oblong in shape with apex acute and margins entire ( Figs. 4, 7, 13, 16, 19 ), 1.7 ± 0.5 (1.2–2.2) mm long and 0.8 ± 0.2 (0.6–1.0) mm wide, length to width ratio 2.2 ± 0.9 (1.3–3.1), appressed or slightly spread-ing at angles of 10 ° –25 ° to the shoot axis. Scale leaves are arched with their adaxial surface depressed, apparently decur-rent, attached to the axis with their broad base ( Figs. 4, 7, 13, 16, 19 ). The number of scale leaves along the axes per centime-ter varies from 7 to 9.
Male cone ( Fig. 4 ) is solitary, terminal, erect, shortly pedun-culate, oblong, 0.3 cm long and 0.2 cm in diameter, and com-posed of many spirally arranged, overlapping microsporophylls. Microsporophylls ( Fig. 4 ) are rhomboidal in shape, 0.5–1.0 mm long and 0.7–1.0 mm wide. Axis of male cone is extended for about 0.3 mm long from the apex of the cone.
Cuticles of the linear leaves show that the epidermal cells on both the adaxial and abaxial surfaces are similar in shape and size ( Figs. 20, 21, 24 ). Epidermis in nonstomatal condition is composed of narrow elongate, rectangular or spindle-shaped cells ( Figs. 20, 24 ), 104.6 ± 44.1 (60.5–148.7) µm long and 16.6 ± 4.5 (12.1–21.1) µm wide, with length to width ratio 6.3 ± 2.0 (4.3–8.3). Cells form regular ranks and their walls are straight (or slightly wavy) and thickened (7.1 ± 2.1 or 5.0–9.2 µm; Fig. 27 ). Epidermal cells between stomata are broad rectangular to elongate, slightly irregular ( Figs. 21, 25, 26 ).
Fig. 2. Stratigraphic column of the Xiaolongtan Formation in Maguan County and Dashidong Village, showing the Sequoia fossils within the bed (revised from Zhang [1976] ).

ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 107January 2015]
Figs. 3–7. Sequoia maguanensis sp. nov. 3. Holotype from Maguan County; foliated shoot with a male cone attached at the apex. Specimen DMS 0130. Bar = 1 cm. 4. Enlargement of oblong male cone in Fig. 3 (arrow A), 0.3 cm long and 0.2 cm in diameter, composed of many spirally arranged, overlapping microsporophylls, with axis (arrow A) extended at the tip and scale leaves (arrow B) attached below. Bar = 0.25 cm. 5. Enlargement of linear leaves in Fig. 3 (arrow B), showing linear leaves spirally arranged along the axis, bifacially fl attened blade, entire margins, acute apex, and sessile bases, decurrent and adnate to the axis (arrow). Bar = 0.5 cm. 6. Enlargement of the foliated shoot in Fig. 3 (arrow C), showing the linear leaves (arrow A) and scale leaves (arrow B) attached on the same axis. Bar = 0.5 cm. 7. Enlargement of scale leaves in Fig. 3 (arrow D), showing scale leaves spirally arranged and overlapping along the axis, deltate in shape with apex acute and margins entire (arrow). Bar = 0.25 cm.

108 AMERICAN JOURNAL OF BOTANY [Vol. 102
oblong; (5) pollen spherical, not winged ( Watson and Eckenwalder, 1993 ).
Leaf epidermal features also confi rm the generic identity of these twigs. In the Wenshan materials, epidermal cells with straight walls, long axis of stoma mostly parallel to the mid-vein, are consistent with those of Sequoia ( Florin, 1931 ; Ma and Li, 2002 ). While Taxodium differs from the present speci-men in the transverse arrangement of stomata in respect to the midvein, Metasequoia in having conspicuous undulated epider-mal cells, which are papillate ( Sveshnikova, 1963 ).
Comparisons with fossil species of Sequoia — Currently, about 19 putative fossil species of Sequoia have been docu-mented from Eurasia, North America and Australia ( Fliche and Zeiller, 1904 ; Seward, 1919 ; Torrey, 1923 ; Chaney, 1951 ; Endo, 1951 ; Knobloch, 1964 ; Sveshnikova, 1967 ; Sveshnikova and Budantsev, 1969 ; Dorofeev, 1975 ; Peters and Christophel, 1978 ; WGCPC, 1978 ; Ammons et al., 1987 ; Samylina, 1988 ; Hill et al., 1993 ; Palamarev, 1994 ; Ma et al., 2005 ; Grímsson
axis as a wide ridge, these features are different from Taxo-dium . In Taxodium , the proximal part of the leaf is markedly constricted, forming a false petiole, and its adnate leaf base is decurrent as a narrow ridge along the axis ( Florin, 1931 ; De Laubenfels, 1953 ). Sequoia differs from Sequoiadendron in that the former contains both linear and scale-like leaves, and the linear leaves are twisted, appearing 2-ranked, whereas the latter contains uniform radiating, needle-like leaves ( Chaney, 1951 ; Watson and Eckenwalder, 1993 ). The specimens from Wenshan State show linear leaves alternately arranged, leaf base decurrent, lacking a petiole, and are distinct from Metase-quoia . In the latter, the linear leaves are oppositely arranged and petiolate ( Miki, 1941 ).
Our fossil specimens are assignable to the genus Sequoia on the basis of the following morphological characters: (1) leafy twigs bearing densely placed, dimorphic leaves, viz., deltate scale-like and linear leaves; (2) linear leaves sessile, base decurrent, spirally arranged, twisted, appearing 2-ranked; (3) scale leaves appressed; (4) male cone solitary, terminal,
Figs. 8–13. Sequoia maguanensis sp. nov. 8. Leafy twig from Dashidong Village. Specimen DMS 0131. Bar = 1 cm. 9. Enlargement of the leafy twig in Fig. 8 (arrow A), showing a linear leaf with decurrent base (arrow). Bar = 0.5 cm. 10. Enlargement of the leafy twig in Fig. 8 (arrow B), showing linear leaves spirally arranged, midrib on abaxial surface of leaves (arrow). Bar = 0.5 cm. 11. Leafy twig from Dashidong Village. Specimen DMS 0132. Bar = 0.5 cm. 12. Enlargement of leafy twig in Fig. 11 (arrow A), showing linear to lanceolate leaves spirally arranged along the axis, with decurrent bases (ar-row). Bar = 0.5 cm. 13. Enlargement of the leafy twig in Fig. 11 (arrow B), showing scale leaves spirally arranged in base of twig, midrib on abaxial surface (arrow). Bar = 0.25 cm.

ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 109January 2015]
et al., 1993 ; Hill, 2001 ). Despite being assigned to different genera, the differences between Austrosequoia Peters et Chris-tophel and Sequoia appear to be at the specifi c rather than ge-neric level ( Peters and Christophel, 1978 ; Hill et al., 1993 ; Hill, 2001 ; Pole, 1995 ). The new species differs from these two species in leaf shape. The former bears linear to lanceolate leaves, while the latter contains only scaly leaves ( Peters and Christophel, 1978 ; Hill et al., 1993 ; Hill, 2001 ).
Sequoia heterophylla Velenovsky has been newly removed from Sequoia to genus Elatocladus an account of epidermal structure ( Kva č ek, 2014 ).
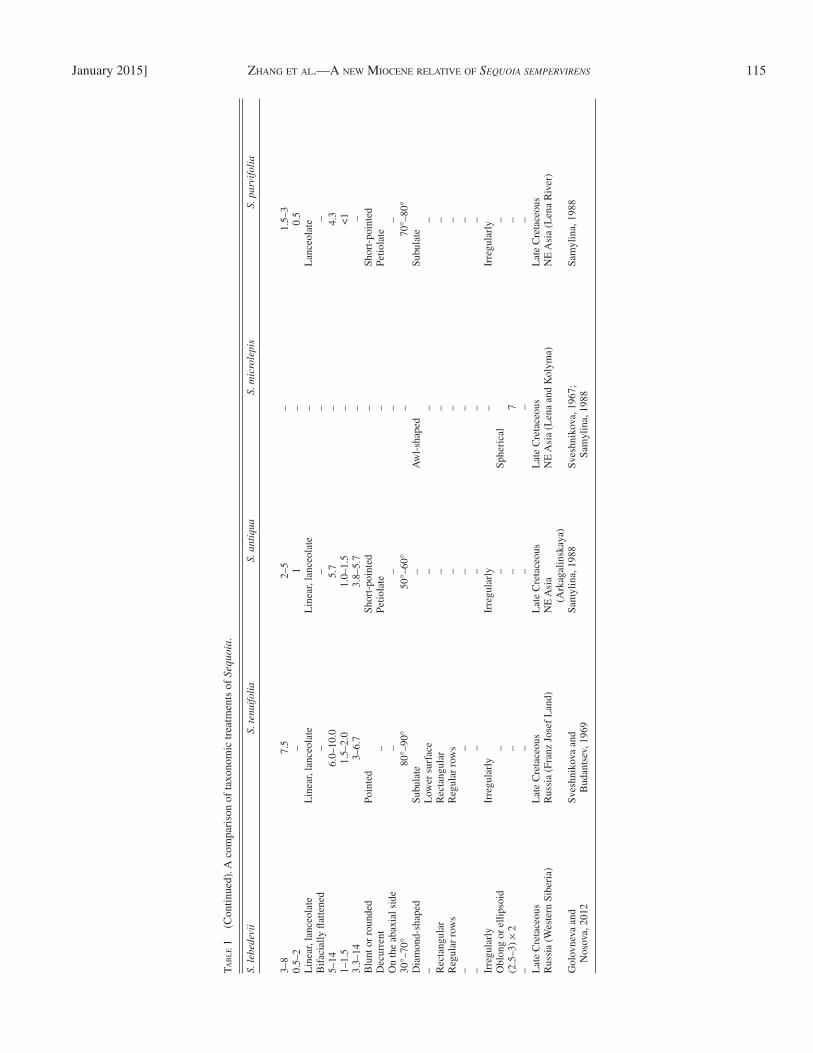
The new species differs from Sequoia jeholensis Endo ( Table 1 ) mainly in its bifacially fl attened linear leaves and larger angles of linear leaves to axis; in the latter, its linear leaves are furrowed and with smaller angles to axis ( Endo, 1951 ). It differs from S. minuta Sveshnikova in its linear to
et al., 2007 ; Golovneva and Nosova, 2012 ; Kva č ek, 2014 ). Among these records, leafy twigs with the linear and scale leaves and a male cone in attachment are rarely seen. In the present discussion, six putative species of Sequoia are excluded because either the leaves or male cones are lacking. Sequoia portlandica Fliche et Zeiller ( S. problematica Fliche et Zeiller) and S. dakotensis Brown were recognized on the basis of el-liptical or spherical seed cones ( Fliche and Zeiller, 1904 ; Chaney, 1951 ), S. burgessii (Penhallow) Torrey and S . mag-nifi ca Knowlton on the basis of petrifi ed woods and stumps ( Torrey, 1923 ; Ammons et al., 1987 ), and S. lithuanica Doro-feev and S. rossica Dorofeev on the basis of seeds ( Dorofeev, 1975 ; Palamarev, 1994 ).
Austrosequoia wintonensis Peters et Christophel and A. tas-manica Hill, Jordan et Carpenter are two seed cones with foli-age attached from Australia ( Peters and Christophel, 1978 ; Hill
Figs. 14–19. Sequoia maguanensis sp. nov. 14. Leafy twig from Maguan County. Specimen DMS 0133. Bar = 1 cm. 15. Enlargement of the leafy twig in Fig. 14 (arrow A), showing linear leaves spirally arranged along the axis, decurrent bases (arrow). Bar = 1 cm. 16. Enlargement of the leafy twig in Fig. 14 (arrow B), showing scale leaves spirally arranged in the base of the twig, with midrib on the abaxial surface (arrow). Bar = 0.5 cm. 17. Leafy twig from Maguan County. Specimen DMS 0134. Bar = 1 cm. 18. Enlargement of the leafy twig in Fig. 17 (arrow A), showing linear leaves twisted and spirally ar-ranged along the axis, with decurrent bases (arrow). Bar = 0.5 cm. 19. Enlargement of the leafy twig in Fig. 17 (arrow B), showing scale leaves spirally arranged at base of twig (arrow). Bar = 0.5 cm.
110 AMERICAN JOURNAL OF BOTANY [Vol. 102
smaller size of linear leaves ( Golovneva and Nosova, 2012 ; Table 1 ), and from S. tenuifolia (Schmalhausen) Sveshnikova et Budantsev in narrower linear leaves and smaller angles of leaves attached to rachis ( Sveshnikova and Budantsev, 1969 ).
lanceolate-shaped leaves; in the latter, leaves are lanceolate in shape ( Hollick and Jeffrey, 1909 ; Seward, 1919 ; Sveshnikova, 1967 ; Chen et al., 1988 ; Samylina, 1988 ). It differs from S. leb-edevii Nosova et Golovneva in its acute leafy apexes and
Figs. 20–23. Leaf cuticles and pollen grain of Sequoia maguanensis sp. nov. 20. Upper surface of leaves, showing elongate, rectangular cells forming regular ranks, cells with walls straight or slightly wavy. Bar = 100 µm. 21. Lower cuticle of leaves, showing stomatal bands composed of 3–5 rows of sto-mata, which aligned mostly parallel to the midvein. Bar = 100 µm. 22. Morphological features of stoma, showing stoma oval in shape, with 4–6 subsidiary cells. Bar = 50 µm. 23. Spheroidal and heteropolar pollen grain from the male cone of holotype. Bar = 20 µm. Figs. 24–31. Scanning electron photographs of cuticles of S. maguanensis sp. nov. Figs. 24–26. Upper cuticle of linear leaves, showing cells forming regular ranks and stomatal bands parallel to mid-vein. 24. Bar = 100 µm; 25–26 . Bar = 40 µm. 27. Showing the thickened cell walls (arrow). Bar = 20 µm. 28. Showing the irregular orientations of stomatal complexes in a stomatal band. Bar = 20 µm. 29–31. Enlargement of stomatal complexes in a stomatal band, showing long axis of stoma parallel (Fig. 29), oblique (Fig. 30), or perpendicular (Fig. 31) to midrib. Stoma composed of 4–6 subsidiary cells (arrow A) and guard cells (arrow B). Bar = 20 µm.

ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 111January 2015]
Figs. 32–37. S. sempervirens (D. Don) Endlicher . 32. Foliated shoot with a male cone attached at the apex, showing the adaxial surface of linear leaves. Bar = 1 cm. 33. Enlargement of the male cone in Fig. 32 (arrow A), showing globose shape and composed of many spirally arranged, overlapping microspo-rophylls (arrow A), with scale leaves (arrow B) attached below. Bar = 0.5 cm. 34, 35. Enlargement of linear leaves in Fig. 32 (arrows B, C), showing linear leaves spirally arranged along the axis, sessile, with bifacially fl attened lamina, entire margins, decurrent base, and adnate to the axis (arrow). Bar = 0.5 cm. 36. Enlargement of scale leaves in Fig. 32 (arrow D), showing scale leaves spirally arranged and overlapping along the axis, deltate in shape with apex acute and margins entire (arrow). Bar = 0.5 cm. 37. The same foliated shoot, showing the midrib on the abaxial surface of linear leaves (arrow). Bar = 1 cm.
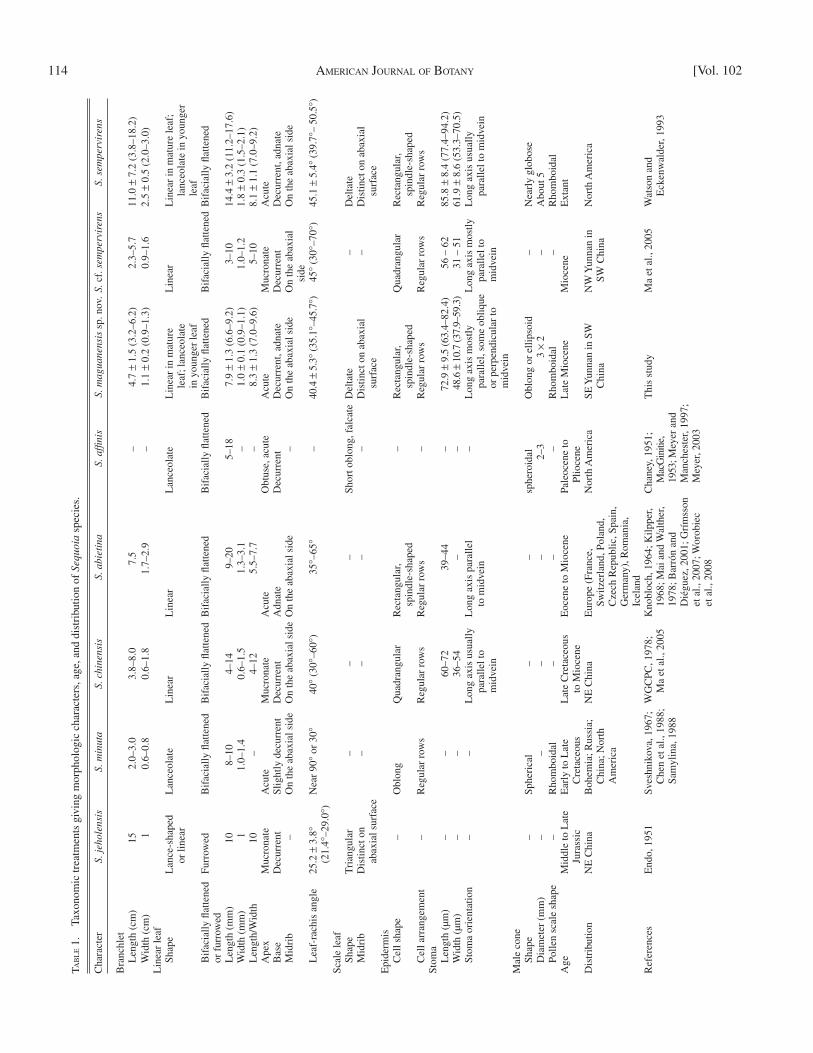
The new species differs from S. antiqua Samylina and S. parvifolia Samylina typically in the longer linear leaves and the lacks of petiole ( Samylina, 1988 ). The new species differs from S. micro-lepis Sveshnikova by its smaller and oblong or ellipsoid-shaped cone, the latter has larger and spherical cones ( Sveshnikova, 1967 ; Samylina, 1988 ; Table 1 ). It differs from S. chinensis Endo emend. Wang et Li in its smaller, linear to lanceolate leaves; in the latter, leaves are larger and linear; besides, the leaves of S. chinensis are hypostomatic, with the stomata only distributed on the abaxial surface of leaves ( WGCPC, 1978 ; Ma et al., 2005 ; Table 1 ). The new species differs from S. abietina (Brongniart) Knobloch ( S. langsdorfi i (Brongniart) Heer) in its smaller, linear to lanceolate leaves, smaller angles of linear leaves to axis and larger size of stomata; in the latter, leaves are larger and linear, with larger angles of linear leaves to axis and smaller size of stomata ( Table 1 ).
A variety of leaf morphologies has been included under the name Sequoia affi nis Lesquereux ( Table 1 ), from scaly and nondistichous, to needle-like and distichous ( Chaney, 1951 ). In the Florissant collections, there is as much distichous, needle-like as scaly foliage ( Chaney, 1951 ). The new species differs from S. affi nis generally in its typical dimorphic leaves attached on the same leafy twig, expanded leaves linear or linear to lan-ceolate in shape, scale leaves deltate, linguiform or ovate-oblong in shape and both with acute apex; while in the latter, its spreading leaves are usually thinner and more delicate, scale
leaves are short oblong with obtuse apex, and lanceolate leaves with acute apex ( Chaney, 1951 ; MacGinitie, 1953 ; Meyer, 2003 ).
Among the fossils of S. affi nis , several show similar morpho-logical features to the foliar shoots of S. maguanensis . In par-ticular, a specimen from the Oligocene Bridge Creek fl ora of North America ( Meyer and Manchester, 1997 ) shows branchlet with dimorphic foliage. The outspread foliar needles are alter-nately arranged (distichous) and possess pointed apexes. Scale-like, spirally arranged needles are appressed at the base. The fossils could be related to S. maguanensis ; however, several factors lead us to not make this assignment. (1) As discussed, the name “ S. affi nis ” includes a variety of leaf morphologies, and many identifi cations have not been confi rmed by their epi-dermal structures; (2) the Bridge Creek fossil lacks the detailed epidermal structure; (3) generally, specimens assigned to S. affi -nis lack dimorphic leaves and are more delicate than those ob-served in the extant S. sempervirens ( Chaney, 1951 ; MacGinitie, 1953 ; Meyer, 2003 ); (4) the spheroidal clustered bud of the Bridge Creek specimen is different from the oblong or ellipsoid male cone in S. maguanensis .
Comparison with the extant species of Sequoia — Morpho-logically, S. maguanensis shows close affi nity with the extant S. sempervirens in the characters of its foliated shoots ( Figs. 3, 8, 32, 37 ), the morphological features of linear ( Figs. 5, 6, 12, 18,
112 AMERICAN JOURNAL OF BOTANY [Vol. 102
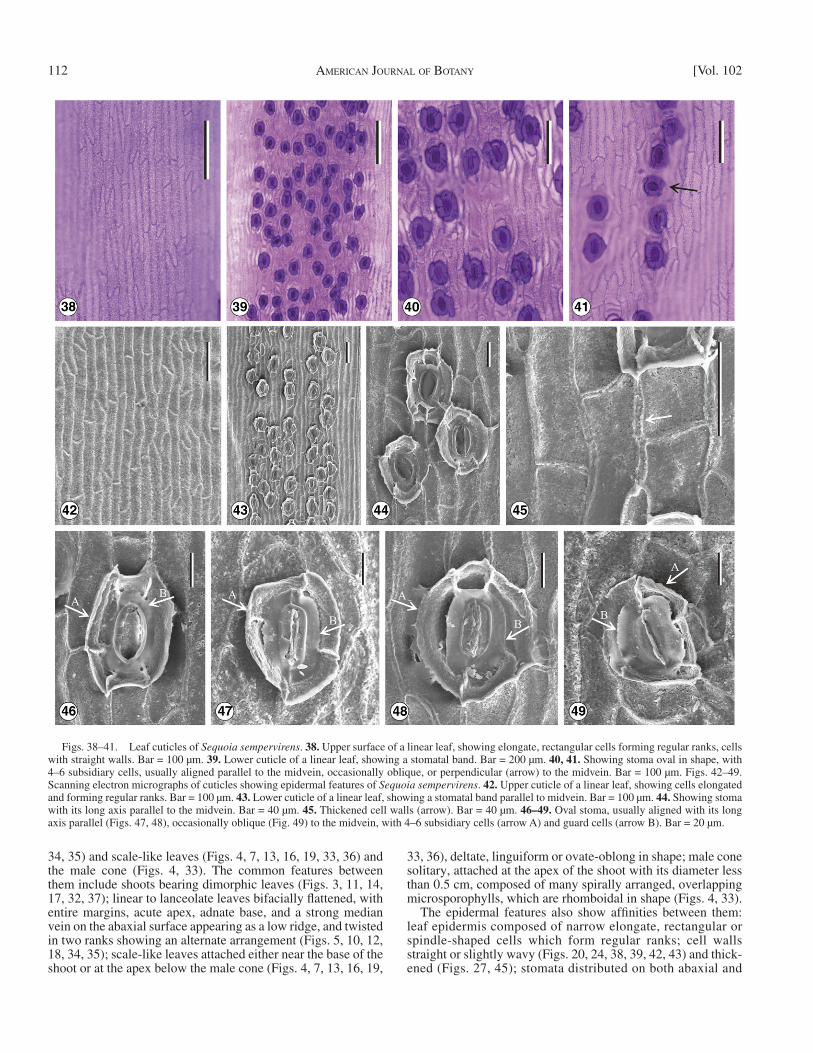
Figs. 38–41. Leaf cuticles of Sequoia sempervirens . 38. Upper surface of a linear leaf, showing elongate, rectangular cells forming regular ranks, cells with straight walls. Bar = 100 µm. 39. Lower cuticle of a linear leaf, showing a stomatal band. Bar = 200 µm. 40, 41. Showing stoma oval in shape, with 4–6 subsidiary cells, usually aligned parallel to the midvein, occasionally oblique, or perpendicular (arrow) to the midvein. Bar = 100 µm. Figs. 42–49. Scanning electron micrographs of cuticles showing epidermal features of Sequoia sempervirens . 42. Upper cuticle of a linear leaf, showing cells elongated and forming regular ranks. Bar = 100 µm. 43. Lower cuticle of a linear leaf, showing a stomatal band parallel to midvein. Bar = 100 µm. 44. Showing stoma with its long axis parallel to the midvein. Bar = 40 µm. 45. Thickened cell walls (arrow). Bar = 40 µm. 46–49. Oval stoma, usually aligned with its long axis parallel (Figs. 47, 48), occasionally oblique (Fig. 49) to the midvein, with 4–6 subsidiary cells (arrow A) and guard cells (arrow B). Bar = 20 µm.
34, 35 ) and scale-like leaves ( Figs. 4, 7, 13, 16, 19, 33, 36 ) and the male cone ( Figs. 4, 33 ). The common features between them include shoots bearing dimorphic leaves ( Figs. 3, 11, 14, 17, 32, 37 ); linear to lanceolate leaves bifacially fl attened, with entire margins, acute apex, adnate base, and a strong median vein on the abaxial surface appearing as a low ridge, and twisted in two ranks showing an alternate arrangement ( Figs. 5, 10, 12, 18, 34, 35 ); scale-like leaves attached either near the base of the shoot or at the apex below the male cone ( Figs. 4, 7, 13, 16, 19,
33, 36 ), deltate, linguiform or ovate-oblong in shape; male cone solitary, attached at the apex of the shoot with its diameter less than 0.5 cm, composed of many spirally arranged, overlapping microsporophylls, which are rhomboidal in shape ( Figs. 4, 33 ).
The epidermal features also show affi nities between them: leaf epidermis composed of narrow elongate, rectangular or spindle-shaped cells which form regular ranks; cell walls straight or slightly wavy ( Figs. 20, 24, 38, 39, 42, 43 ) and thick-ened ( Figs. 27, 45 ); stomata distributed on both abaxial and

ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 113January 2015]
2001 ; Lu et al., 2011 ). Thus, exchange between East Asia and North America via the Beringian land bridge was still possible in the early Neogene. With the global cooling of the Neogene in gen-eral, and in particular after the middle Miocene warm interval, the Beringian land bridge would have become too cool for many ther-mophilic plants to spread across this route ( Tanai, 1967 ; Wolfe, 1978 ; Tiffney and Manchester, 2001 ). Therefore, the exchange of the ancestor of S. sempervirens via the Beringian Strait has hap-pened at a time when the land bridge was not too cool for this thermophilic conifer to spread across between eastern Asia and North America, probably in the early middle Miocene. The present distribution of S. sempervirens is a relictual distribution with the extirpation of the Asian fossil representatives.
Paleoclimatic implications — The Wenshan fl ora is preserved in laminated beds of light-gray, gray-white or grayish yellow pelitic siltstone or carbonaceous mudstone. The sheet-like sedi-mentary sequence ( Fig. 2 ) contains abundant plant fossils, namely, Quercus sp., Salix sp., Dryophyllum sp., Ulmus sp., Z elkova sp., Ampelopsis sp., Glyptostrobus europaeus (Brongniart) Unger ( Zhang, 1976 ), Ailanthus confucii Unger ( Su et al., 2013 ), Bauhinia wenshanensis Meng et Zhou ( Meng et al., 2014 ), Pinus massoniana Lambert, and Sequoia maguanensis , as well as animal fossils, namely, Gyraulus sp., Galba sp., and Physa sp. ( Zhang, 1976 ). Based on the deciduous broadleaved spe-cies, the climate was considered to have been humid and tem-perate ( Zhang, 1976 ).
Ailanthus confucii Unger closely resembles the extant one, namely, A. altissima (Mill.) Swingle ( Su et al., 2013 ). The extant A. altissima is the only member of the genus found in temperate climates rather than in the tropics. Based on the distribution of this extant species (Northeast China, Central China, and Taiwan Province) and the assemblage of the fossil fl ora (Cupressaceae, Rosaceae, Saliaceae, and Ulmaceae), the authors reconstructed a subtropical to temperate humid climate during the late Miocene in Southeast Yunnan ( Su et al., 2013 ).
The paleoclimate reconstruction of Xiaolongtan flora (Xiaolongtan Formation, upper Miocene) in Kaiyuan (about 120 km NW of Dashidong Village, Fig. 1) shows a humid subtropical climate, being more humid with a higher mean annual tempera-ture (MAT) than today in Southeast Yunnan ( Xia et al., 2009 ).
The close morphological resemblance of Sequoia maguanensis to S. sempervirens implies similar environmental requirement be-tween them ( Thiel et al., 2012 ; Pittermann et al., 2012 ), which fur-ther suggests that the environment in the late Miocene of Southeast Yunnan and western North America must have been similar. Cur-rently, S. sempervirens lives in climates with MAT between 10.0 ° C and 15.3 ° C, January temperatures between 5.5 ° C and 12.2 ° C, July temperatures between 11.6 ° C and 21.9 ° C, and annual precipitation between 560 and 1810 mm, January precipitation be-tween 135 and 331 mm, and July precipitation between 1 and 9 mm ( Humboldt County Visitors Bureau, 2014 ; USGS, 2014 ), accompanied regions with summer fog. Thus, this conifer lives in a warm and humid environment throughout the year ( Barbour and Major, 1977 ; Snyder, 1992 ). The present day environmental limits of S. sempervirens by analogy indicate a warm climate with humid seasons in Southeast Yunnan during the late Miocene.
The disappearance of Sequoia from eastern Asia — Tempera-ture and precipitation are the climatic factors that usually confi ne the distribution of plants ( Woodward, 1987 ). Based on the dis-cussion above, S. maguanensis would probably have lived in
adaxial surface ( Figs. 21, 24, 40, 41 ), oval in shape, with guard cells having distinctly thickened walls and epitaxial skirts ( Figs. 29–31, 46–49 ); mostly, stomata being aligned parallel to the midvein ( Figs. 26, 44 ).
The new species differs from the extant one primarily in the shape and size of male cone ( Figs. 4, 33 ), size of linear leaves ( Figs. 3, 8, 11, 14, 17, 32, 37 ), angular divergence of linear leaves from rachis ( Figs. 5, 10, 12, 18, 34, 35 ), and the arrange-ment of stomata ( Figs. 21, 25, 26, 40, 41, 44 ; Table 1 ). In the new species, its male cone is oblong in shape ( Fig. 4 ) rather than spherical or globose in shape as in extant species ( Fig. 33 ); the male cone and the linear leaves in the new species are smaller than in the extant species. In addition in the new spe-cies, its linear leaves form a smaller angle with the rachis ( Figs. 5, 10, 12, 18 ) and its stomata are smaller ( Figs. 29–31 ) and ar-ranged in a different manner ( Fig. 28 ) than the extant species ( Figs. 34, 35, 46–49 ; Table 1 ).
Leaf shoots with associated female cones were described from the upper Miocene and lower Pliocene of Japan ( Endo, 1933 ; Miki, 1941 ) and Miocene of Northwest Yunnan, China ( Ma et al., 2005 ). These fossils were assigned to the modern species of S. sempervirens (or S. cf. sempervirens ) based on the similarities of gross morphological features in leaves, cones, and alternate arrangement of linear leaves. However, these leafy shoots have no dimorphic leaves and lack scale leaves at their bases. Because the female cones were usually fi lled by the surrounding rock matrix, their detailed structure is unknown. Fur-ther, the epidermal structures of the Japanese specimens are miss-ing. Therefore, the assignment of these fossils to the extant species is doubtful. The linear leaves on these fossil shoots also show simi-lar morphological features to the Wenshan materials, suggesting that these fossils can also be S. maguanensis or a relative.
Biogeographic implications — In eastern Asia, Sequoia cf. sempervirens and S. sempervirens were reported from the Mio-cene or early Pliocene ( Endo, 1933 ; Miki, 1941 ; Ma et al., 2005 ). They all bear spreading, bifacially fl attened, linear to lanceolate leaves, and show similar foliage characteristics to the modern S. sempervirens . But their leafy shoots have no di-morphic leaves and lack scale leaves on the bases, the leaves have no epidermal structures, and no fertile structures are pres-ent. Thus, as discussed already, their taxonomic status is un-clear, and affi nity with the extant species is controversial. The present fi nding of S. maguanensis in the upper Miocene of Southeast Yunnan, China, which mostly resembles the extant species, confi rms the affi nity of the Asian forms to the modern relict S. sempervirens .
In Europe, S. abietina has been recognized frequently from Paleogene and Neogene sediments ( Knobloch, 1969 ; Grímsson et al., 2007 ). Its leaf morphology as well as epidermal structure show clearer difference to the extant species than those of S. maguanensis ( Table 1 ). In North America, S. affi nis is mostly found from the Paleogene and contains a variety of leaf mor-phologies. It differs from the extant species in its spreading leaves as well as scale leaves ( Table 1 ). Therefore, according to leaf morphology and anatomy, Asian S. maguanensis is more closely related to the extant S. sempervirens than European fos-sils and North American S. affi nis.
The presence of S. maguanensis in the upper Miocene of eastern Asia, which closely resembles the extant North Ameri-can S. sempervirens , probably implies a Beringian biogeo-graphic track of the ancestor of S. sempervirens in the early Neogene ( Hopkins, 1967 ; Tiffney, 1985 ; Donoghue et al.,
114 AMERICAN JOURNAL OF BOTANY [Vol. 102 TA
BL
E 1
. Ta
xono
mic
trea
tmen
ts g
ivin
g m
orph
olog
ic c
hara
cter
s, a
ge, a
nd d
istr
ibut
ion
of S
equo
ia s
peci
es.
Cha
ract
er S.
jeho
lens
is
S. m
inut
a S.
chi
nens
is
S. a
biet
ina
S. a
ffi ni
s S.
mag
uane
nsis
sp.
nov
. S.
cf.
sem
perv
iren
s S.
sem
perv
iren
s
Bra
nchl
et
Len
gth
(cm
)15
2.0–
3.0
3.8–
8.0
7.5
–4.
7 ±
1.5
(3.2
–6.2
)2.
3–5.
711
.0 ±
7.2
(3.
8–18
.2)
W
idth
(cm
)1
0.6–
0.8
0.6–
1.8
1.7–
2.9
–1.
1 ±
0.2
(0.9
–1.3
)0.
9–1.
62.
5 ±
0.5
(2.0
–3.0
)L
inea
r le
af
Shap
eL
ance
-sha
ped
or li
near
Lan
ceol
ate
Lin
ear
Lin
ear
Lan
ceol
ate
Lin
ear
in m
atur
e le
af; l
ance
olat
e in
you
nger
leaf
Lin
ear
Lin
ear
in m
atur
e le
af;
lanc
eola
te in
you
nger
le
af
Bif
acia
lly fl
atte
ned
or f
urro
wed
Furr
owed
Bif
acia
lly fl
atte
ned
Bif
acia
lly fl
atte
ned
Bif
acia
lly fl
atte
ned
Bif
acia
lly fl
atte
ned
Bif
acia
lly fl
atte
ned
Bif
acia
lly fl
atte
ned
Bif
acia
lly fl
atte
ned
L
engt
h (m
m)
108–
104–
149–
205–
187.
9 ±
1.3
(6.6
–9.2
)3–
1014
.4 ±
3.2
(11
.2–1
7.6)
W
idth
(m
m)
11.
0–1.
40.
6–1.
51.
3–3.
1–
1.0
± 0.
1 (0
.9–1
.1)
1.0–
1.2
1.8
± 0.
3 (1
.5–2
.1)
L
engt
h/W
idth
10–
4–12
5.5–
7.7
–8.
3 ±
1.3
(7.0
–9.6
)5–
108.
1 ±
1.1
(7.0
–9.2
)
Ape
xM
ucro
nate
Acu
teM
ucro
nate
Acu
teO
btus
e, a
cute
Acu
teM
ucro
nate
Acu
te
Bas
eD
ecur
rent
Slig
htly
dec
urre
ntD
ecur
rent
Adn
ate
Dec
urre
ntD
ecur
rent
, adn
ate
Dec
urre
ntD
ecur
rent
, adn
ate
M
idri
b–
On
the
abax
ial s
ide
On
the
abax
ial s
ide
On
the
abax
ial s
ide
–O
n th
e ab
axia
l sid
eO
n th
e ab
axia
l si
deO
n th
e ab
axia
l sid
e
L
eaf-
rach
is a
ngle
25.2
± 3
.8 °
(21.
4 ° –2
9.0 °
)N
ear
90 ° o
r 30
° 40
° (30
° –60
° )35
° –65
° –
40.4
± 5
.3 ° (
35.1
° –45
.7 ° )
45 ° (
30 ° –
70 ° )
45.1
± 5
.4 ° (
39.7
° – 5
0.5 °
)
Scal
e le
af
Shap
eT
rian
gula
r–
––
Shor
t obl
ong,
fal
cate
Del
tate
–D
elta
te
Mid
rib
Dis
tinct
on
abax
ial s
urfa
ce–
––
–D
istin
ct o
n ab
axia
l su
rfac
e–
Dis
tinct
on
abax
ial
surf
ace
Epi
derm
is
Cel
l sha
pe–
Obl
ong
Qua
dran
gula
rR
ecta
ngul
ar,
spin
dle-
shap
ed–
Rec
tang
ular
, sp
indl
e-sh
aped
Qua
dran
gula
rR
ecta
ngul
ar,
spin
dle-
shap
ed
Cel
l arr
ange
men
t–
Reg
ular
row
sR
egul
ar r
ows
Reg
ular
row
s–
Reg
ular
row
sR
egul
ar r
ows
Reg
ular
row
sSt
oma
L
engt
h (µ
m)
––
60–7
239
–44
–72
.9 ±
9.5
(63
.4–8
2.4)
56 –
62
85.8
± 8
.4 (
77.4
–94.
2)
Wid
th (
µm)
––
36–5
4–
–48
.6 ±
10.
7 (3
7.9–
59.3
)31
– 5
161
.9 ±
8.6
(53
.3–7
0.5)
St
oma
orie
ntat
ion
––
Lon
g ax
is u
sual
ly
para
llel t
o m
idve
in
Lon
g ax
is p
aral
lel
to m
idve
in–
Lon
g ax
is m
ostly
pa
ralle
l, so
me
obliq
ue
or p
erpe
ndic
ular
to
mid
vein
Lon
g ax
is m
ostly
pa
ralle
l to
mid
vein
Lon
g ax
is u
sual
ly
para
llel t
o m
idve
in
Mal
e co
ne
Shap
e–
Sphe
rica
l–
–sp
hero
idal
Obl
ong
or e
llips
oid
–N
earl
y gl
obos
e
Dia
met
er (
mm
)–
––
–2–
33
× 2
–A
bout
5
Polle
n sc
ale
shap
e–
Rho
mbo
idal
––
–R
hom
boid
al–
Rho
mbo
idal
Age
Mid
dle
to L
ate
Jura
ssic
Ear
ly to
Lat
e C
reta
ceou
sL
ate
Cre
tace
ous
to M
ioce
neE
ocen
e to
Mio
cene
Pale
ocen
e to
Pl
ioce
neL
ate
Mio
cene
Mio
cene
Ext
ant
Dis
trib
utio
nN
E C
hina
Boh
emia
; Rus
sia;
C
hina
; Nor
th
Am
eric
a
NE
Chi
naE
urop
e (F
ranc
e,
Switz
erla
nd, P
olan
d,
Cze
ch R
epub
lic, S
pain
, G
erm
any)
, Rom
ania
, Ic
elan
d
Nor
th A
mer
ica
SE Y
unna
n in
SW
C
hina
NW
Yun
nan
in
SW C
hina
Nor
th A
mer
ica
Ref
eren
ces
End
o, 1
951
Sves
hnik
ova,
196
7 ;
Che
n et
al.,
198
8 ;
Sam
ylin
a, 1
988
WG
CPC
, 197
8 ;
Ma
et a
l., 2
005
Kno
bloc
h, 1
964 ;
Kilp
per,
1968
; Mai
and
Wal
ther
, 19
78 ; B
arró
n an
d D
iégu
ez, 2
001 ;
Grí
mss
on
et a
l., 2
007 ;
Wor
obie
c et
al.,
200
8
Cha
ney,
195
1 ;
Mac
Gin
itie,
19
53 ; M
eyer
and
M
anch
este
r, 19
97 ;
Mey
er, 2
003
Thi
s st
udy
Ma
et a
l., 2
005
Wat
son
and
Eck
enw
alde
r, 19
93
ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 115January 2015]
TAB
LE 1
(C
ontin
ued)
. A c
ompa
riso
n of
taxo
nom
ic tr
eatm
ents
of
Sequ
oia .
S. le
bede
vii
S. te
nuif
olia
S.
ant
iqua
S.
mic
role
pis
S. p
arvi
foli
a
3–8
7.5
2–5
–1.
5–3
0.5–
2–
1–
0.5
Lin
ear,
lanc
eola
teL
inea
r, la
nceo
late
Lin
ear,
lanc
eola
te–
Lan
ceol
ate
Bif
acia
lly fl
atte
ned
––
––
5–14
6.0–
10.0
5.7
–4.
31–
1.5
1.5–
2.0
1.0–
1.5
–<
13.
3–14
3–6.
73.
8–5.
7–
–B
lunt
or
roun
ded
Poin
ted
Shor
t-po
inte
d–
Shor
t-po
inte
dD
ecur
rent
–Pe
tiola
te–
Petio
late
On
the
abax
ial s
ide
––
––
30 ° –
70 °
80 ° –
90 °
50 ° –
60 °
–70
° –80
° D
iam
ond-
shap
edSu
bula
te–
Aw
l-sh
aped
Subu
late
–L
ower
sur
face
––
–R
ecta
ngul
arR
ecta
ngul
ar–
––
Reg
ular
row
sR
egul
ar r
ows
––
––
––
––
––
––
–Ir
regu
larl
yIr
regu
larl
yIr
regu
larl
y–
Irre
gula
rly
Obl
ong
or e
llips
oid
––
Sphe
rica
l–
(2.5
–3)
× 2
––
7–
––
––
–L
ate
Cre
tace
ous
Lat
e C
reta
ceou
sL
ate
Cre
tace
ous
Lat
e C
reta
ceou
sL
ate
Cre
tace
ous
Rus
sia
(Wes
tern
Sib
eria
)R
ussi
a (F
ranz
Jos
ef L
and)
NE
Asi
a (A
rkag
alin
skay
a)N
E A
sia
(Len
a an
d K
olym
a)N
E A
sia
(Len
a R
iver
)
Gol
ovne
va a
nd
Nos
ova,
201
2 Sv
eshn
ikov
a an
d B
udan
tsev
, 196
9 Sa
myl
ina,
198
8 Sv
eshn
ikov
a, 1
967 ;
Sa
myl
ina,
198
8 Sa
myl
ina,
198
8
116 AMERICAN JOURNAL OF BOTANY [Vol. 102
coast redwood to other genera of Taxodiaceae. Silvae Genetica 51 : 93 – 100 .
AMMONS , R. , W. J. FRITZ , R. B. AMMONS , AND A. AMMONS . 1987 . Cross-identifi cation of ring signatures in Eocene trees ( Sequoia magnifi ca ) from the Specimen Ridge locality of the Yellowstone Fossil Forests. Palaeogeography, Palaeoclimatology, Palaeoecology 60 : 97 – 108 .
AN , Z. S. , J. E. KUTZBACH , W. L. PRELL , AND S. C. PORTER . 2001 . Evolution of Asian monsoons and phased uplift of the Himalaya–Tibetan Plateau since late Miocene times. Nature 411 : 62 – 66 .
ANDREWS , H. N. 1936 . A new Sequoioxylon from Florissant, Colorado. Annals of the Missouri Botanical Garden 23 : 439 – 446 .
BARBOUR , M. G. , AND J. MAJOR . 1977 . Terrestrial vegetation of California . John Wiley, New York, New York, USA.
BARRÓN , E. , AND C. DIÉGUEZ . 2001 . Estudio macrofl orístico del Mioceno Inferior lacustre de la Cuenca de Rubielos de Mora (Teruel, España). Boletín Geológico y Minero 112 : 13 – 56 [in Spanish] .
BGMRYP [ BUREAU OF GEOLOGY AND MINERAL RESOURCES OF YUNNAN PROVINCE ] . 1990 . Regional geology of Yunnan Province . Geological Publishing House, Beijing, Beijing, China [in Chinese with English introduction].
BGMRYP [BUREAU OF GEOLOGY AND MINERAL RESOURCES OF YUNNAN PROVINCE] . 1996 . Stratigraphy (lithostratigraphy) of Yunnan Province . China University of Geosciences Press, Wuhan, Hubei, China [in Chinese with English introduction].
BLAZER , A. M. 1975 . Index of generic names of fossil plants, 1966–1973, based on the Compendium Index of Paleobotany of the U.S. Geological Survey . U.S. Geological Survey 1396, U.S. Government Printing, Washington D.C., USA.
BOZUKOV , V. S. , AND B. V. TSENOV . 2012 . Catalogue of the Cenozoic plants of Bulgaria (Eocene to Pliocene). Addendum and corrigendum. Phytologia Balcanica 18 : 237 – 261 .
CHANEY , R. W. 1951 . A revision of fossil Sequoia and Taxodium in western North America based on the recent discovery of Metasequoia. Trans-actions of the American Philosophical Society, New Series 40 : 171 – 263 .
CHEN , F. , X. Y. MENG , S. Q. REN , AND C. L. WU . 1988 . The Early Cretaceous fl ora of Fuxin Basin and Tiefa Basin, Liaoning Province . Geological Publishing House, Beijing, Beijing, China [in Chinese].
DE LAUBENFELS , D. J. 1953 . The external morphology of coniferous leaves. Phytomorphology 3 : 1 – 20 .
DONG , W. 1987 . Further investigations upon the age and characteristics of the Xiaolongtan Fauna, Kaiyuan Co., Yunnan Province. Vertebrata Palasiatica 25 : 116 – 123 .
DONOGHUE , M. J. , C. D. BELL , AND J. LI . 2001 . Phylogenetic patterns in northern hemisphere plant geography. International Journal of Plant Sciences 162 : S41 – S52 .
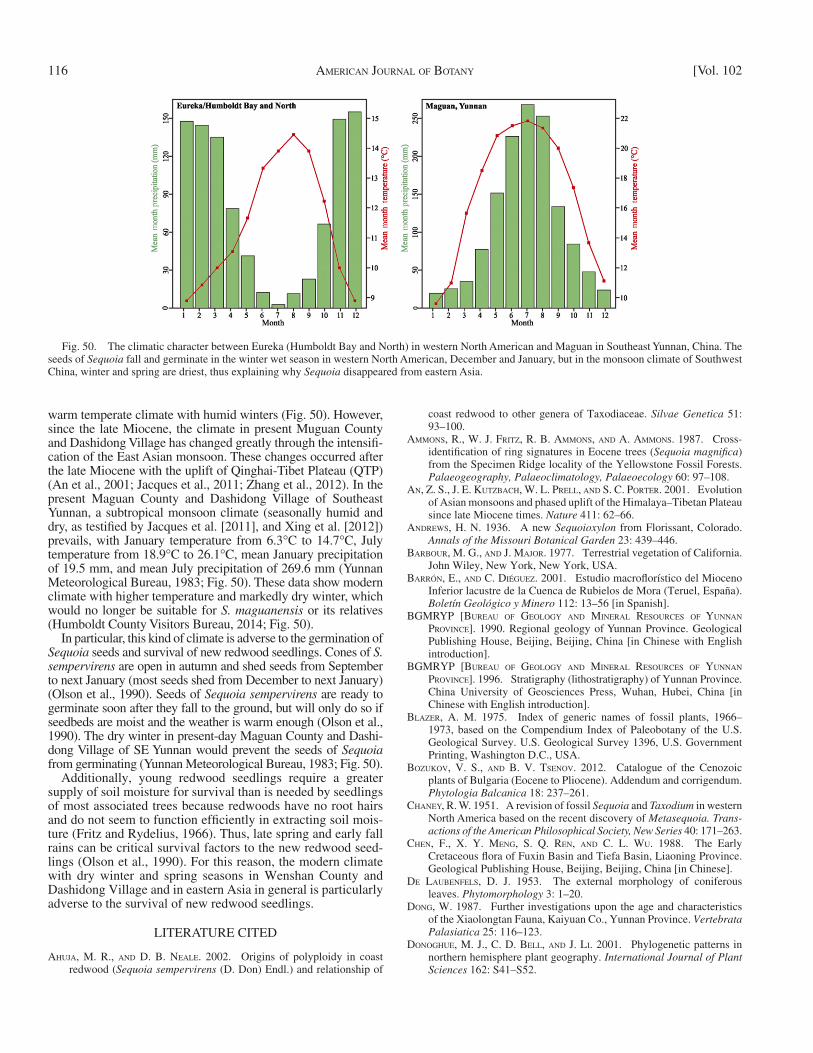
warm temperate climate with humid winters ( Fig. 50 ). However, since the late Miocene, the climate in present Muguan County and Dashidong Village has changed greatly through the intensifi -cation of the East Asian monsoon. These changes occurred after the late Miocene with the uplift of Qinghai-Tibet Plateau (QTP) ( An et al., 2001 ; Jacques et al., 2011 ; Zhang et al., 2012 ). In the present Maguan County and Dashidong Village of Southeast Yunnan, a subtropical monsoon climate (seasonally humid and dry, as testifi ed by Jacques et al. [2011] , and Xing et al. [2012] ) prevails, with January temperature from 6.3 ° C to 14.7 ° C, July temperature from 18.9 ° C to 26.1 ° C, mean January precipitation of 19.5 mm, and mean July precipitation of 269.6 mm ( Yunnan Meteorological Bureau, 1983 ; Fig. 50 ). These data show modern climate with higher temperature and markedly dry winter, which would no longer be suitable for S. maguanensis or its relatives ( Humboldt County Visitors Bureau, 2014 ; Fig. 50 ).
In particular, this kind of climate is adverse to the germination of Sequoia seeds and survival of new redwood seedlings. Cones of S. sempervirens are open in autumn and shed seeds from September to next January (most seeds shed from December to next January) ( Olson et al., 1990 ). Seeds of Sequoia sempervirens are ready to germinate soon after they fall to the ground, but will only do so if seedbeds are moist and the weather is warm enough ( Olson et al., 1990 ). The dry winter in present-day Maguan County and Dashi-dong Village of SE Yunnan would prevent the seeds of Sequoia from germinating ( Yunnan Meteorological Bureau, 1983 ; Fig. 50 ).
Additionally, young redwood seedlings require a greater supply of soil moisture for survival than is needed by seedlings of most associated trees because redwoods have no root hairs and do not seem to function effi ciently in extracting soil mois-ture ( Fritz and Rydelius, 1966 ). Thus, late spring and early fall rains can be critical survival factors to the new redwood seed-lings ( Olson et al., 1990 ). For this reason, the modern climate with dry winter and spring seasons in Wenshan County and Dashidong Village and in eastern Asia in general is particularly adverse to the survival of new redwood seedlings.
LITERATURE CITED
AHUJA , M. R. , AND D. B. NEALE . 2002 . Origins of polyploidy in coast redwood ( Sequoia sempervirens (D. Don) Endl.) and relationship of
Fig. 50. The climatic character between Eureka (Humboldt Bay and North) in western North American and Maguan in Southeast Yunnan, China. The seeds of Sequoia fall and germinate in the winter wet season in western North American, December and January, but in the monsoon climate of Southwest China, winter and spring are driest, thus explaining why Sequoia disappeared from eastern Asia.

ZHANG ET AL.—A NEW MIOCENE RELATIVE OF SEQUOIA SEMPERVIRENS 117January 2015]
DOROFEEV , P. I. 1975 . Zur Systematik einigen Taxodiaceen. Paleontologicheski Zhurnal 1 : 105 – 116 [in Russian] .
ENDO , S. 1928 . A new Paleocene species of Sequoia. Japanese Journal of Geology and Geography 6 : 27 – 29 .
ENDO , S. 1933 . A Neogene species of Sequoia from Japan. Botanical Gazette 94 : 605 – 610 .
ENDO , S. 1951 . A record of Sequoia from the Jurassic of Manchuria. Botanical Gazette 113 : 228 – 230 .
FIELDS , P. F. 1993 . A newly recognized Neogene Sequoia in the Pacifi c Northwest of North America. American Journal of Botany 80 ( supple-ment ): 89 .
FLICHE , P. , AND R. ZEILLER . 1904 . Note sur une fl orule portlandienne des environs de Boulogne-sur-Mer. Bulletin de la Société Géologique de France 4 : 787 – 812 .
FLORIN , R. 1931 . Untersuchungen zur Stammesgeschichte der Coniferales und Cordaitales. Kungliga Svenska Vetenskaps Akademiens Handlingar Series 10 : 1 – 588 .
FLORIN , R. 1963 . The distribution of conifer and taxad genera in time and space. Acta Horti Bergiani 20 : 121 – 311 .
FRITZ , E. , AND J. A. RYDELIUS . 1966 . Redwood reforestation problems: An experimental approach to their solution . Foundation for American Resource Management, San Francisco, California, USA.
GADEK , P. A. , D. L. ALPERS , M. M. HESLEWOOD , AND C. J. QUINN . 2000 . Relationships within Cupressaceae sensu lato: A combined morphologi-cal and molecular approach. American Journal of Botany 87 : 1044 – 1057 .
GOLOVNEVA , L. B. , AND N. B. NOSOVA . 2012 . Albian-Cenomanian fl ora of Western Siberia . Marathon Publishers, St. Petersburg, Russia [in Russian with English introduction].
GRÍMSSON , F. , T. DENK , AND L. A. SÍMONARSON . 2007 . Middle Miocene fl oras of Iceland—The early colonization of an island? Review of Palaeobotany and Palynology 144 : 181 – 219 .
HABLY , L. 1994 . Egerian plant fossils from Pom á�z, Hungary. Fragmenta Mineralogica et Palaeontologica 17 : 5 – 70 .
HILL , R. S. 2001 . The Cenozoic macrofossil record of the Cupressaceae in the Southern Hemisphere. Acta Palaeobotanica 41 : 123 – 132 .
HILL , R. S. , G. J. JORDAN , AND R. J. CARPENTER . 1993 . Taxodiaceous macrofossils from Tertiary and Quaternary sediments in Tasmania. Australian Systematic Botany 6 : 237 – 249 .
HOLLICK , A. , AND E. C. JEFFREY . 1909 . Studies of Cretaceous coniferous remains from Kreischerville, New York. Memoirs of the New York Botanical Garden 3 : 1 – 76 .
HOLÝ , F. , Z. KVA EK , AND V. TEODORIDIS . 2012 . A review of the early Miocene mastixioid fl ora of the Kristina Mine at Hr á�dek and Nisou in North Bohemia (The Czech Republic). Acta Musei Nationalis Pragae, B 68 : 53 – 118 .
HOPKINS , D. M. 1967 . The Bering land bridge . Stanford University Press, Palo Alto, California, USA.
HUMBOLDT COUNTY VISITORS BUREAU . 2014 . California’s Redwood Coast, Humboldt County [online]. Website http://redwoods.info/showrecord.asp?id=2465&source=Search [accessed 03 November 2014].
JACQUES , F. M. B. , S. X. GUO , T. SU , Y. W. XING , Y. J. HUANG , Y. S. LIU , D. K. FERGUSON , AND Z. K. ZHOU . 2011 . Quantitative reconstruction of the late Miocene monsoon climates of southwest China: A case study of the Lincang fl ora from Yunnan Province. Palaeogeography, Palaeoclimatology, Palaeoecology 304 : 318 – 327 .
JUNGWIRTH , E. 2004 . The determination and taxonomic problems of Paleogene fossil macrofl ora from Slovenia and Croatia. Natura Croatica 13 : 187 – 196 .
KERP , H. 1990 . The study of fossil gymnosperms by means of cuticular analysis. Palaios 5 : 548 – 569 .
KILPPER , K. 1968 . Koniferen aus den tertiären Deckschichten des Niederrheinischen Hauptfl özes, 3. Taxodiaceae und Cupressaceae. Palaeontographica Abt B 124 : 102 – 111 .
KNOBLOCH , E. 1964 . Haben Cinnamomum scheuchzeri Heer und Cinnamomum polymorphum (A. Braun) Heer nomenklatorisch richtige Namen? Neues Jahrbuch für Geologie und Paläontologie 10 : 597 – 603 .
KNOBLOCH , E. 1969 . Tertiäre Floren von Mähren . Moravské Museum Brno and Musejní Spolek Brno, Brno, Southern Moravia, Czechoslovakia.
KOVAR-EDER , J. , Z. KVA Č EK , AND B. MELLER . 2001 . Comparing early to middle Miocene fl oras and probable vegetation of Oberdorf N
Voitsberg (Austria), Bohemia (Czech Republic), and Wackersdorf (Germany). Review of Palaeobotany and Palynology 114 : 83 – 125 .
KUNZMANN , L. 1999 . Koniferen der Oberkreide und ihre Relikte im Tertiär Europas. Ein Beitrag zur Kenntnis ausgestorbener Taxodiaceae und Geinitziaceae fam. nov. Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden 45 : 1 – 191 .
KVA Č EK , J. 2014 . The conifer Elatocladus velenovskyi from the Bohemian Cenomanian. Palaeontographica B291 : 1 – 3 .
KVA Č EK , Z. , AND V. TEODORIDIS . 2007 . Tertiary Macrofloras of the Bohemian Massif: A review with correlations within Boreal and Central Europe. Bulletin of Geosciences 82 : 383 – 408 .
LU , J. M. , D. Z. LI , S. LUTZ , A. SOEJIMA , T. S. YI , AND J. WEN . 2011 . Biogeographic disjunction between eastern Asia and North America in the Adiantum pedatum complex (Pteridaceae). American Journal of Botany 98 : 1680 – 1693 .
MA , Q. W. , AND C. S. LI . 2002 . Epidermal structures of Sequoia sempervi-rens (D. Don) Endl. (Taxodiaceae). Taiwania 47 : 194 – 202 .
MA , Q. W. , F. L. LI , AND C. S. LI . 2005 . The coast redwoods ( Sequoia , Taxodiaceae) from the Eocene Heilongjiang and the Miocene of Yunnan, China. Review of Palaeobotany and Palynology 135 : 117 – 129 .
MACGINITIE , H. D. 1953 . Fossil plants of the Florissant beds, Colorado. Carnegie Institution of Washington Publication 599 : 86 – 87 .
MAI , D. H. 1995 . Tertiäre Vegetationsgeschichte Europas Methoden und Ergebnisse . Gustav Fischer Verlag, Jena, Thuringia Land, Germany [in German].
MAI , D. H. , AND H. WALTHER . 1978 . Die fl oren der Haselbacher Serie im Weisselster-Becken (Bezirk Leipzig, DDR). Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden 28 : 1 – 200 [in German] .
MENG , H. H. , F. B. JACQUES , T. SU , Y. J. HUANG , S. T. ZHANG , H. J. MA , AND Z. K. ZHOU . 2014 . New biogeographic insight into Bauhinia s.l. (Leguminosae) integration from fossil records and molecular analy-ses. BMC Evolutionary Biology 14 : 181 – 195 .
MEYER , H. W. 2003 . The fossils of Florissant . Smithsonian Books, Washington D.C., USA.
MEYER , H. W. , AND S. R. MANCHESTER . 1997 . The Oligocene Bridge Creek fl ora of the John Day Formation, Oregon. University of California Publications in Geological Sciences 141 : 1 – 195 .
MIKI , S. 1941 . On the change of fl ora in eastern Asian since Tertiary Period (I), the clay or lignite beds fl ora in Japan with special refer-ence to the Pinus trifolia beds in Central Hondo. Journal of Japanese Botany 11 : 237 – 303 .
MIKI , S. , AND S. HIKITA . 1951 . Probable chromosome number of fossil Sequoia and Metasequoia found in Japan. Science 113 : 3 – 4 .
MILLER , C. N. 1977 . Mesozoic conifers. Botanical Review 43 : 217 – 280 .
OLSON , D. F. , D. F. ROY , AND G. A. WALTERS . 1990 . Sequoia sempervirens (D. Don) Endl. Redwood . In R. M. Burns and B. H. Honkala [eds.], Silvics of North America, vol. 1, Conifers. Agriculture Handbook 654, 541–551. USDA Forest Service, Washington D.C., USA.
PALAMAREV , E. 1994 . Palaokarpologische Untersuchungen des Braunkohlenjungtertiars in Bulgarien. Palaeontographica 232 : 129 – 154 [in German] .
PALAMAREV , E. , V. BOZUKOV , K. UZUNOVA , A. PETKOVA , AND G. KITANOV . 2005 . Catalogue of the Cenozoic plants of Bulgaria (Eocene to Pliocene). Phytologia Balcanica 11 : 228 – 236 .
PETERS , M. D. , AND D. C. CHRISTOPHEL . 1978 . Austrosequoia wintonen-sis , a new taxodiaceous cone from Queensland, Australia. Canadian Journal of Botany 56 : 3119 – 3128 .
PITTERMANN , J. , S. A. STUART , T. E. DAWSON , AND A. MOREAU . 2012 . Cenozoic climate change shaped the evolutionary ecophysiology of the Cupressaceae conifers. Proceedings of the National Academy of Sciences, USA 109 : 9647 – 9652 .
POLE , M. 1995 . Late Cretaceous macrofloras of Eastern Otago, New Zealand: Gymnosperms. Australian Systematic Botany 8 : 1067 – 1106 .
SAKAI , A. 1971 . Freezing resistance of relicts from the Arcto-Tertiary fl ora. New Phytologist 70 : 1199 – 1205 .

118 AMERICAN JOURNAL OF BOTANY
SAMYLINA , V. A. 1988 . Arkagalinskaya stratofl ora Severo-Vostoka Azii (Arkagalinskaya stratofl ora of North-East Asia) . Nauka Publishers, Leningrad, Leningrad Region, Russia [in Russian].
SAWANGCHOTE , P. , P. J. GROTE , AND D. L. DILCHER . 2009 . Tertiary leaf fossils of Mangifera (Anacardiaceae) from Li Basin, Thailand as ex-amples of the utility of leaf marginal venation characters. American Journal of Botany 96 : 2048 – 2061 .
SAWYER , J. O. , S. C. SILLETT , J. H. POPENOE , A. LABANCE , T. SHOLARS , D. L. LARGENT , F. EUPHRAT , ET AL . 2000 . Characteristics of redwood for-ests. In R. F. Noss [ed.], The redwood forest: History, ecology, and conservation of the coast redwoods, 39–79. Island Press, Washington D.C., USA.
SEWARD , A. C. 1919 . Fossil plants , Ginkgoales, Coniferales, Gnetales, vol. 4. Cambridge University Press, Cambridge, UK.
SNYDER , J. A. 1992 . The ecology of Sequoia sempervirens : An adden-dum to “On the Edge: Nature’s Last Stand for Coast Redwoods”. D.M.A. dissertation, San Jose State University, San Jose, California, USA.
SPICER , R. A. , A. AHLBERG , A. B. HERMAN , S. P. KELLEY , M. I. RAIKEVICH , AND P. M. REES . 2002 . Palaeoenvironment and ecology of the middle Cretaceous Grebenka fl ora of northeastern Asia. Palaeogeography, Palaeoclimatology, Palaeoecology 184 : 65 – 105 .
SRINIVASAN , V. , AND E. M. FRIIS . 1989 . Taxodiaceous conifers from the Upper Cretaceous of Sweden . Biologiske Skrifter—Kongelige Danske Videnskabernes Selskab 35 : 5 – 57 .
SU , T. , F. B. JACQUES , H. J. MA , AND Z. K. ZHOU . 2013 . Fossil fruits of Ailanthus confucii from the upper Miocene of Wenshan, Yunnan Province, SW China. Palaeoworld 22 : 153 – 158 .
SVESHNIKOVA , I. N. 1963 . Atlas and a key for the identifi cation of the liv-ing and fossil Sciadopityaceae and Taxodiaceae based on the structure of the leaf epidermis. Acta Science USSR Paleobotany 4 : 207 – 229 [in Russian with an English abstract] .
SVESHNIKOVA , I. N. 1967 . Late Cretaceous conifers of Soviet Union I, fos-sil conifers of the Viliuyian Depression . Paleobotanika 6 : 177 – 203 [in Russian].
SVESHNIKOVA , I. N. , AND L. Y. BUDANTSEV . 1969 . Iskopaemye Flory Arktiki I . Nauka Publishers, Leningrad, Russia [in Russian].
TANAI , T. 1967 . Miocene fl oras and climate in East Asia. Abhandlungen des Zentralen Geologischen Instituts, Berlin 10 : 195 – 205 .
THIEL , C. , S. KLOTZ , AND D. UHL . 2012 . Palaeoclimate estimates for se-lected leaf fl oras from the late Pliocene (Reuverian) of Central Europe based on different palaeobotanical techniques. Turkish Journal of Earth Sciences 21 : 263 – 287 .
TIFFNEY , B. H. 1985 . Perspectives on the origin of the fl oristic similar-ity between eastern Asia and eastern North America. Journal of the Arnold Arboretum 66 : 73 – 94 .
TIFFNEY , B. H. , AND S. R. MANCHESTER . 2001 . The use of geological and paleontological evidence in evaluating plant phylogeographic
hypotheses in the northern hemisphere Tertiary. International Journal of Plant Sciences 162 ( 6 supplement ): S3 – S17 .
TORREY , R. E. 1923 . The comparative anatomy and phylogeny of the Coniferales, part 3. Mesozoic and Tertiary coniferous woods. Memoirs of the Boston Society of Natural History 6 : 39 – 106 .
USGS [U. S. GEOLOGICAL SURVEY] . 2014 . Climate–vegetation atlas of North America [online]. Website http://pubs.usgs.gov/pp/p1650-a/datatables/ and http://pubs.usgs.gov/pp/p1650-a/pages/conifers.html/ [accessed 23 May, 2014].
VELENOVSKY , J. 1885 . Die Gymnospermen der böhmischen Kreide Formation . Selbstverlag in Commission bei Fr. Ř�ivn á� č , Prague, Prague, Czechoslovakia.
WATSON , F. D. , AND J. E. ECKENWALDER . 1993 . Cupressaceae . In Flora of North America Editorial Committee [ed.], Flora of North America, north of Mexico, vol. 2, 399 – 422. Oxford University Press, Oxford, UK.
WGCPC [WRITING GROUP OF CENOZOIC PLANTS OF CHINA] . 1978 . Cenozoic plants from China, fossil plants of China, vol. 3 . Science Press, Beijing, China [in Chinese].
WOLFE , J. A. 1978 . A paleobotanical interpretation of Tertiary climates in the Northern Hemisphere. American Scientist 66 : 694 – 703 .
WOODWARD , F. I. 1987 . Climate and plant distribution . Cambridge University Press, Cambridge, UK.
WOROBIEC , G. , E. WOROBIEC , AND J. KASINSKI . 2008 . Plant assemblages of the drill cores from the Neogene Ruja lignite deposit near Legnica (Lower Silesia, Poland). Acta Palaeobotanica 48 : 191 – 275 .
XIA , K. , T. SU , Y. S. LIU , Y. W. XING , F. M. B. JACQUES , AND Z. K. ZHOU . 2009 . Quantitative climate reconstructions of the late Miocene Xiaolongtan megafl ora from Yunnan, southwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 276 : 80 – 86 .
XING , Y. W. , T. UTESCHER , F. JACQUES , T. SU , Y. CHRISTOPHER , Y. HUANG , AND Z. K. ZHOU . 2012 . Paleoclimatic estimation reveals a weak win-ter monsoon in southwestern China during the late Miocene: Evidence from plant macrofossils. Palaeogeography, Palaeoclimatology, Palaeoecology 358–360 : 19 – 26 .
YUNNAN METEOROLOGICAL BUREAU . 1983 . Climatic data of Yunnan agri-culture . Yunnan People’s Press, Kunming, China [in Chinese].
ZHANG , C. H. 1976 . The report to the regional geological survey (1/200,000) of Wenshan/Maguan Scope (F-48-3, F-48-9) . Geological Bureau of Yunnan Province, Yunnan, China [online book, http://search.ngac.cn/CGML/DZ000659/z01_0107.pdf and http://search.ngac.cn/CGML/DZ000659/z01_0190.pdf] [in Chinese].
ZHANG , Q. Q. , D. K. FERGUSON , V. MOSBRUGGER , Y. F. WANG , AND C. S. LI . 2012 . Vegetation and climatic changes of SW China in response to the uplift of Tibetan Plateau. Palaeogeography, Palaeoclimatology, Palaeoecology 363–364 : 23 – 36 .
ZHENG , J. J. , X. X. HE , S. W. LIU , Z. J. LI , X. S. HUANG , G. F. CHEN , AND Z. D. QIU . 1999 . Chinese stratigraphical thesaurus, Tertiary . Geological Publishing House, Beijing, China [in Chinese].