saltatory search in free-living callithrix jacchus: environmental and age influences
TRANSCRIPT

Saltatory Search in Free-Living Callithrixjacchus: Environmental and Age Influences
Antonio Souto & Bruna M. Bezerra & Nicola Schiel &Ludwig Huber
Received: 19 October 2004 /Revised: 30 January 2006 /Accepted: 10 July 2006 /Published online: 15 August 2007# Springer Science + Business Media, LLC 2007
Abstract Researchers generally categorize motile foraging behavior into 3strategies: ambush, cruise, and saltatory searching. During saltatory searches,predators move from one location to the next, frequently pausing to scan for preythat are hard to detect and widely distributed. We investigated whether 1) theforaging strategy of free-living common marmosets conforms to the strategy; 2)scanning occurs solely when the individual is stationary; 3) the environment (denseand sparse vegetation) influences foraging behavior; and 4) the age of the marmosetsis related to their foraging behavior. Bezerra carried out the observations in a 32-hafragment of Atlantic Forest in the Northeast of Brazil and in an adjacentcondominium. Using the focal sampling method, we observed 31 commonmarmosets, including adults, juveniles, and infants, Bezerra recorded the followingbehaviors (in dense and sparse vegetation): locomotion (subdivided into minorlocomotion—moving distances ≤1 m; major locomotion—moving distances >1 m),scan, pause, and capture. The frequency of scanning behavior was significantlygreater when individuals were stationary. Adults and juveniles exhibited the clearestdifferentiation in terms of locomotion, both adjusting the behavior in accordancewith the environment; periods of major locomotion were more frequent in sparsevegetation, whereas minor locomotion was more frequent in dense vegetation. Incontrast, infants exhibited major locomotion more frequently in dense vegetation.We conclude that common marmosets use a saltatory strategy when foraging, andthat their foraging behavior is plastic, changing both with the age of the individualand with the density of the vegetation.
Int J Primatol (2007) 28:881–893DOI 10.1007/s10764-007-9165-1
A. Souto (*) : B. M. BezerraDepartment of Zoology, Federal University of Pernambuco, 50.670-420 Recife, Brazile-mail: [email protected]
N. Schiel : L. HuberDepartment of Developmental and Theoretical Biology, Institute of Zoology, University of Vienna,Vienna, Austria

Keywords commonmarmoset . environmental influences . foraging behavior .
ontogeny . saltatory search
Introduction
Researchers broadly classify behaviors used in the search for prey into 3 foragingstrategies based on the degree of motility employed: ambush, cruise, and saltatorysearching (O’Brien et al. 1990).
Ambush searches, or sit-and-wait, involve the predator remaining in a stationaryposition for long periods of time waiting for prey to approach (Huey and Pianka1981). The foraging strategy is defined by <1 movement per min (MPM) (Butler2005; Cooper 2005) and by <10% of the total time spent moving, the latter criterionformally known as percent time moving (PTM; Cooper 2005; Reaney and Whiting2002). Species that seek out prey items that are densely distributed and highlymobile employ the ambush strategy (Bell 1991), i.e., arthropods, reptiles, birds, andmammals, including tarsiers (Barbosa and Moreno 1999; Cooper 2005; Hopcraftet al. 2005; Leyhe and Ritchison 2004; Miyashita 2005; Robakiewicz and Daigle2004; Roberts and Kohn 1993; Secor 2005; Shine and Li-Xing 2002; Tye 1989). Incontrast, the cruise search, or widely foraging, strategy involves moving through theenvironment in an active search for prey (Bell 1991). A cruise searcher has an MPM<1, like the ambush searchers, but a PTM ≥10 (Cooper 2005; Reaney and Whiting2002). They are specialist predators of sedentary or widely distributed prey (Bell1991). Some species of arthropods, fish, reptiles, and birds employ the cruise search(Barbosa and Moreno 1999; Bell 1991; Cooper 2005; Day et al. 1999; O’Brien et al.1990; Reaney and Whiting 2002). However, if individuals cannot easily detect thewidely distributed prey because of crypticness or small size or both, it may be amore effective strategy to stop and scan for them (Anderson et al. 1997; Bell 1991;O’Brien et al. 1990). Researchers first called this stop and go pattern of foragingpause-travel search (Anderson 1981) and later, saltatory search (Bell 1991; O’Brienet al. 1989). The saltatory search strategy is therefore a frequent stop and go patternof foraging, in which the forager scans for available resources only while stationary(Bell 1991; O’Brien et al. 1990). Butler (2005) and Cooper (2005) developed theconcept by specifying that such foragers should exhibit an MPM ≥1, thus higherthan that of ambush searchers and cruise searchers. While the frequent stop and gopattern is a fundamental characteristic of saltatory searching, some researchersbelieve that scanning for prey items is possible during locomotion as well as duringstationary periods. However, Getty (1993) conjectured that if scanning occurs whilethe forager is moving, then detection cannot be as efficient as when it is stationary.More recently Anderson et al. (1997) adopted a model that assumes that individualsare able to scan while moving, even if acuity decreases with locomotion. Whethersaltatory foragers scan for prey items while moving is still open to debate.
Unlike cruise and ambush searching, saltatory search varies according toenvironmental conditions (O’Brien et al. 1990). For example, in starlings andEuropean thrushes, the distance of relocating moves within a foraging patch varies
882 A. Souto, et al.

with vegetation density (Brownsmith 1977; Smith 1974). In such birds, movingshort distances in open areas was the modal pattern, whereas moving longerdistances was more common in dense vegetation.
Studies have revealed the use of the saltatory search strategy in different speciesof birds, reptiles, fish, and insects (Anderson et al. 1997; Andersson 1981;Brownsmith 1977; Butler 2005; Cody 1968; Eifler and Eifler 1999; Getty andPullian 1991; O’Brien et al. 1989, 1990; Smith 1974; Sonerud 1992; Tye 1989),though conclusive evidence of the strategy in mammals is still lacking. Kenagy(1973) suggested the presence of saltatory searching in some species of rodents.
Common marmosets (Callithrix jacchus) are small Neotropical primates,measuring (head and body length) about 21 cm and 16 cm (adults and younganimals, respectively; Hershkovitz 1977). They are endemic to Brazil, inhabitingmainly second growth or scrub forest (caatinga) as well as the Atlantic Forest innortheast Brazil (Auricchio 1995; Hershkovitz 1977; Rylands and De Faria 1993;Stevenson and Rylands 1988). Insects are an essential part of their diet (Ferrari andLopes Ferrari 1989; Stevenson and Rylands 1988; Sussman and Kinzey 1984), andthey spend 24–30% of their daily activities searching for animal prey (Rylandsand De Faria 1993; Stevenson and Rylands 1988). The main insects commonmarmosets prey upon are orthopterans (grasshoppers, crickets), hymenopterans(ants), isopterans (termites), homopterans (cicadas), and lepidopterans (usuallylarvae from butterflies and moths: Alonso and Langguth 1989; Maier et al. 1982;Rylands and De Faria 1993; Schiel 2000). The common marmoset foraging strategycomprises a slow creeping movement, showing permanent attentiveness andconcentration to the surroundings (Maier et al. 1982; Stevenson and Rylands1988). The strategy is presumably advantageous to them given that their prey areoften cryptic (Stevenson and Rylands 1988). Among their insect prey, crypticness isusually achieved via coloration (green, brown, or olive-green; Schiel 2000), thoughbehavior also plays an important role because the insects must select a suitablesubstrate on which to stay (Atkins 1978). Moreover, their relatively small sizeaffords a form of protection against predators (Buzzi 2005; Gullan and Cranston2000; Maranhão 1976). The prey size of common marmosets generally ranges from0.5 cm, e.g., termites and some ants, to ca. 11 cm, e.g., grasshoppers (Buzzi 2005).To capture insects, common marmosets quickly seize them when they are stationarywith 1 or 2 hands, or directly by mouth, and then chew and ingest them (Maier et al.1982; Schiel 2000). There is no record of marmosets pursuing prey. Though thereare few studies on the distribution of insects in tropical forests, the herbivorousspecies (the most numerous types) are widely distributed (Barbosa et al. 2005;Novotny and Basset 2005; Novotny et al. 2002). If the majority of insect speciespreyed upon by common marmosets are widely distributed and hard to detect, thenthey should forage via a stop and go pattern of locomotion—the saltatory searchstrategy—as O’Brien et al. (1990) and Anderson et al. (1997) suggested.
Accordingly, our objectives were to investigate whether 1) the foraging strategyof free-living common marmosets conforms to the strategy; 2) scanning occurssolely when the individual is stationary; 3) the environment (either dense or sparsevegetation) affects foraging behavior; and 4) the age of the marmosets influencestheir foraging behavior.
Saltatory Search in Free-Living Callithrix jacchus 883

Methods
Study Site
The study site is ca. 40 km from Recife, the capital of the state of Pernambuco innortheast Brazil. The climate has a wet and a dry season. In the rainy season, June–August, the temperature ranges between 17 and 29°C and the humidity between 90%and 100%. During the dry season, the temperature can vary between 24 and 32°Cand humidity between 70% and 80%.
We conducted observations in an Atlantic Forest fragment at 7°56′97″ S, 35°01′23′ W), and also in its surroundings where there is a small condominium, occupiedmostly during the summer months. We carried out all observations during periodswhen few of the occupants of the condominium were present, to limit humaninterference.
The native primary and secondary Atlantic Forest vegetation in situ is representedmostly by 15–20-m-high mature trees, which have other, younger trees growingaround them, producing a dense arrangement. We measured the average vegetationdensity using the nearest neighbor method (Beasom and Haucke 1975) by countingthe number of trees in 5 m2 with a trunk circumference of 5 cm-1.50 m (Curtis andMcIntosh 1950). We measured the distance between the trunks 1 m above theground. The average vegetation density is 3.2 trees/m2 and the average distance tothe nearest neighbor is 61 cm.
The condominium consists of 70 houses with gardens connecting each house tothe surrounding forest. The gardens usually include trees, which also commonlygrow alongside the unpaved, narrow roadways of the condominium. The vegetationin the areas is relatively sparse (average density was 0.4 tree/m2), while the averagedistance to the nearest neighbor is 5.85 m. Though consisting of fewer trees thandense vegetation, the canopy in sparse vegetation provided good support forquadrupedal locomotion inside the canopy and between the trees. In both areas(Atlantic Forest and gardens), the locations where Bezerra conducted observationshad a continuous canopy, and some trees in the gardens were directly connected withthe forest via the canopy. The home range of the common marmosets included thetrees in the gardens, which they visited daily during periods of foraging.
Subjects
Bezerra observed a total of 31 individuals of varying ages, in their naturalenvironment. We classified the observed individuals into 3 age classes (Ingram1977; Stevenson and Rylands 1988): infants (9 wk–4 mo), juveniles (5–10 mo), andadults (≥15 mo). The number of individuals within each age group was 10 infants,11 juveniles, and 10 adults. There was no subadult (10–15 mo) in the focal groups.
Bezerra identified individuals by their natural marks: size and color of the ear-tuffs, scars, color of the fur, and any physical defect. In 6 infants she cut, withscissors, a small portion of their tail fur to aid in distinguishing them. To ensure thatthe infant that was to be marked was distracted sufficiently to enable the tail fur to becut, one of us (Schiel) placed a slice of banana on a tree trunk about 1.5 m above theground, attracting the target individual to eat it. Bezerra would then carefully
884 A. Souto, et al.

approach from behind and properly mark it. As the groups were used to bothresearchers, the adults showed no perceivable aversive reaction, i.e., alarm calls orevasion. We offered a slice of banana only for marking purposes, on average onceevery 45 d of the observational period, such that the marmosets did not make anassociation between the observer and food. The identification avoided unnecessarystress and suffering, which otherwise might have occurred with capturing andmarking techniques. Further, no associations were created between observer and anegative stimulus, and thus the individuals allowed the observer to approach closely,facilitating observations. We generated an identification table, which included thename, group, classification code, estimated age, gender, main natural marks, and adrawing spotting the main natural marks of each individual.
Forms of Behavior
Bezerra’s observations were based on the ethogram Caine (1996) provided, whichspecifically describes the behavior of captive Geoffroyi marmosets (Callithrixgeoffroyi) foraging for animal prey.
In investigating the presence and use of saltatory searching by foraging commonmarmosets in the wild, Bezerra recorded specific behaviors: locomotion, scan, pause,and capture. Locomotion is movement from place to place as the result of changingthe position of all 4 limbs in relation to the substrate. Locomotion comprised allmovements, including those that preceded prey capture, and we subdivided it intominor locomotion and major locomotion. Minor locomotion is moving from onelocation to another ≤1 m away, and major locomotion is moving from one locationto another >1 m away. We chose 1 m to categorize locomotion subsequent topreliminary observations of marmoset foraging behavior, when we noticed thatlocomotion distance varied considerably. Sometimes individuals undertook move-ments over quite short distances—as little as 20 cm—and sometimes they moved >1 m.The 2 types of locomotion occurred in areas of continuous vegetation. We interpretedthe act of leaping from one branch to another as part of the locomotion behavior, and assuch included it in minor and major locomotion. A focal individual could engage inboth major and minor locomotion during a single observational session. We used thebehavior scan—slow arcing movement of the head— to indicate that foraging activitywas occurring (Caine 1996). Bezerra did not record scans clearly associated withvigilance, defined as looking around quickly into the surrounding vegetation (Halseyet al. 2006). We defined capture as the act of capturing a prey item, per Maier et al.(1982) and Schiel (2000).
General Methods
The study had 3 phases of observations: 1) During 6 wk, totaling ca. 90 h, Bezerraand Schiel followed the marmosets so that they would get used to their presence; bythe end of this period Bezerra and Schiel could easily approach to ≤1 m withoutdisturbing them and also identified the individuals. 2) With 30 h of observations viathe focal sampling method (Lehner 1996), they assessed and confirmed thesuitability of using the proposed definitions to categorize behaviors. They deemedthis step necessary in light of the fact that the descriptions Caine (1996) provided
Saltatory Search in Free-Living Callithrix jacchus 885

were for captive Callithrix geoffroyi. 3) Bezerra recorded the foraging behavior ofthe identified individuals, employing the proposed behavioral categories, via thefocal sampling method: 155 sessions of 10 min duration, with continuousobservations of 1 individual before Bezerra selected a new focal individual. Bezerraafforded 5 observational sessions to each individual in total, for a total of 50 min ofdata per individual. She recorded 80 sessions in dense vegetation and 75 in sparsevegetation during April–June 2002, February–March 2003, and October andDecember 2003. Bezerra observed each individual in only 1 of the 2 environmentalconditions. She observed 16 individuals —5 adults, 6 juveniles, and 5 infants—only in dense vegetation, and she observed 15 others —5 adults, 5 juveniles, and 5infants— only in sparse vegetation.
The marmosets used vegetation with a continuous canopy, which enabledindividuals to explore both habitats in the study site without constraint. Wediscarded the data of a focal individual that eventually used both areas during thesame observational session. We also discarded observational sessions in which anysort of disturbance due to the approach of unfamiliar humans or domestic animalsoccurred. Finally, we also excluded sessions containing >3 min of nonforagingbehaviors, e.g., playing, resting, grooming etc. That is, all individuals foraged for≥70% of each session.
We considered behavior to be a discrete unit for recording and analysis when ithad a clear start and end point. For example, in a given 10-min session an individualcould pause, start scanning, and then stop scanning, e.g., engaging in a groomingactivity, and then return to scanning again a few moments later, in which case wescored 2 acts of scanning. Although the categories major locomotion, minorlocomotion, and pause are mutually exclusive, it is possible for scan to occur duringany of the other behaviors.
We were especially attentive to forms of behavior that denoted 1 of the 3 foragingstrategies —saltatory, ambush, and cruise search— as described by Anderson et al.(1997), Bell (1991), Getty (1993), Huey and Pianka (1981), and O’Brien et al.(1990). We adopted the criteria that Cooper (2005) and Butler (2005) used todistinguish ambush and cruise search from saltatory search. Thus, we consideredindividuals exhibiting ≥1 MPM to be saltatory foragers (Butler 2005; Cooper 2005).We also measured the PTM value because it provides more details of the foragingbehavior of common marmosets and might be useful in future comparisons withother primate species when more data are available. The MPM value of anindividual is the sum of its movements, minor and major locomotion together,divided by the total observational time for that individual. We obtained the PTM valueof each individual by multiplying the total time it spent moving by 100 and thendividing by the total observation time.
Bezerra recorded all forms of foraging behavior via a Psion Organizer II(Observer 2.0, Noldus Corporation) and microcassette recorders (Sony-M-529V;Aiwa NRF TP-M 330).
Statistical Analyses
We used Friedman 2-way of analyses of variance by ranks to calculate possibledifferences in the frequency of >2 paired samples. We used Kruskal-Wallis 1-way
886 A. Souto, et al.

analyses of variance by ranks to test for >2 independent samples. For thecomparisons of 2 independent samples we used the Mann-Whitney U test. Weconsidered p≤0.05 (2-tailed) to indicate significant effects (Siegel 1975). Values forMPM and PTM are reported as means ± SE.
Results
Strategy Employed During Foraging Activity, Scan Behavior, and Locomotion Patterns
The common marmosets showed only intermittent locomotion during the observa-tional sessions. The average number of MPM is 2.49±0.09 (N=31), and PTM is11.78±0.50 (N=31).
Scanning for prey occurred both when individuals were moving and when theywere stationary. The lowest frequency of scan behavior occurred when individualsdisplayed major locomotion, while the highest frequency for the behavior occurredwhen they were stationary (pause), irrespective of their age group (adults: N=10, Fr=15.8, d.f.=2 p≤0.001; juveniles: N=11, Fr=18.7, d.f.=2, p<0.001; infants: N=10,Fr=18.2, df=2, p≤0.001; Friedman 2-way analyses of variance by ranks; Fig. 1).
Prey Captured and Common Marmosets’ Age Class
Common marmosets captured the following animals: ants and termites (15 events),larvae (9 events), cicadas (2 events), 1 spider, 1 mollusc, 1 grasshopper, 1 smallbutterfly, and unidentified small invertebrates (≤2 cm, 26 events). The largest preythat the common marmosets captured was a grasshopper, ca. 10 cm long, and thesmallest prey were ants and termites (ca. 0.5 cm long). The captured prey wasusually brown, green, or olive-green, similar to the predominant colors of theenvironment in which they lived.
There was a total of 56 capturing events, from which 40 occurred in the denseenvironment (15 by adults, 22 by juveniles, and 3 by infants) and 16 in the sparse (9
distribution of scan
adults juveniles infants0
10
20
30
40
50
60
maj
or l.
maj
or l.
maj
or l.m
inor
l.
paus
e
paus
e
paus
e
*** *** ***
min
or l.
min
or l.
%
Fig. 1 Distribution of scanbehavior within the behaviors’major locomotion (major l.),minor locomotion (minor l.), andpause for each age group. Sta-tistics: Friedman 2-way analysisof variance by ranks (2-tailed);***p≤0.001.
Saltatory Search in Free-Living Callithrix jacchus 887
by adults, 4 by juveniles, and 3 by infants). Common marmosets captured more preyin dense than in sparse vegetation (Mann-Whitney U test: n1=16, n2=15, U=69.5,p<0.05). Adults and juveniles captured more prey than infants did in densevegetation (n1, 2=5, U=2, p<0.05 and n1=5, n2=6, U=1.5, p<0.05, respectively),whereas there is no difference between adults and juveniles there (n1=5, n2=6, U=14.5, ns). Regardless of age classes, in sparse vegetation there is no statisticaldifference in the frequency of captures (adults and juveniles: n1, 2=5, U=6.5, ns;adults and infants: n1, 2=5, U=5.5, ns; juveniles and infants: n1, 2=5, U=10.5, ns).In 3 events, all in sparse vegetation, infants actively obtained the rest of the uneatenprey older individuals captured.
Age Group in Common Marmosets and the Ability for Locomotion While Foraging
Overall locomotion —values of minor and major categories summed— does notdiffer between age classes, either in dense or in sparse vegetation (dense vegetation:N=16, X 2=3.054, d.f.=2, ns; sparse vegetation: N=15, X 2=1.954, d.f.=2, ns;Kruskal-Wallis 1-way analysis of variance by ranks).
Age Group, Environmental Differences, and Locomotion Pattern
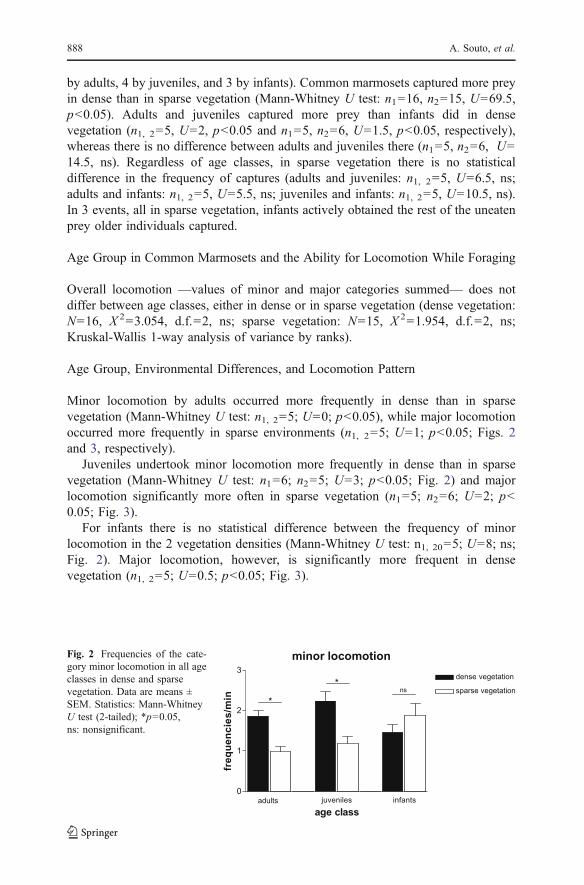
Minor locomotion by adults occurred more frequently in dense than in sparsevegetation (Mann-Whitney U test: n1, 2=5; U=0; p<0.05), while major locomotionoccurred more frequently in sparse environments (n1, 2=5; U=1; p<0.05; Figs. 2and 3, respectively).
Juveniles undertook minor locomotion more frequently in dense than in sparsevegetation (Mann-Whitney U test: n1=6; n2=5; U=3; p<0.05; Fig. 2) and majorlocomotion significantly more often in sparse vegetation (n1=5; n2=6; U=2; p<0.05; Fig. 3).
For infants there is no statistical difference between the frequency of minorlocomotion in the 2 vegetation densities (Mann-Whitney U test: n1, 20=5; U=8; ns;Fig. 2). Major locomotion, however, is significantly more frequent in densevegetation (n1, 2=5; U=0.5; p<0.05; Fig. 3).
minor locomotion
0
1
2
3
adults juveniles infants
dense vegetation
sparse vegetation
*
*ns
age class
freq
uen
cies
/min
Fig. 2 Frequencies of the cate-gory minor locomotion in all ageclasses in dense and sparsevegetation. Data are means ±SEM. Statistics: Mann-WhitneyU test (2-tailed); *p=0.05,ns: nonsignificant.
888 A. Souto, et al.

Discussion
The foraging behavior of common marmosets showed a clear sequence of stop andgo movements (MPM ≥1), which supports the prediction of O’Brien et al. (1989)and Anderson et al. (1997) for predators searching for species that are difficult todetect and widely distributed. But scanning also occurred when marmosetsundertook locomotion over relatively short distances. Thus, scanning did not occurexclusively when the individual was sessile, as O’Brien et al. (1989) and Bell (1991)assumed. However, scanning decreased significantly as locomotion increased. In thissense, the PTM was slightly >10, suggesting an advantage in being stationary whensearching for prey.
Our data support the model of Anderson et al. (1997) in which movement andscanning for prey are possible at the same time. It is important to note that theproposition is still compatible with O’Brien’s hypothesis but includes the possibilityof scanning during movement. We believe that common marmosets tend to optimizetheir foraging by scanning while moving, even if in a limited way. The difference inthe frequency of scanning events when the individuals performed major locomotion,minor locomotion, and pause might be related to the need for vision to maintainbalance while moving (Eibl-Eibesfeldt 1999). One shortfall of our research was thelack of measurements of the speed of locomotion of the marmosets while foraging.However, if one assumes that the speed of locomotion is similar for periods of minorand major locomotion then it might be reasonable to suppose that it is easier to scanand maintain balance while moving short distances. An alternative possibility is thatminor locomotion is associated with lower speeds, in which case individuals thatmove slowly from one location to another are probably more able to scan whilemoving than otherwise. More studies are required to clarify this point. We suggestcarrying out such measurements with individuals in captivity because measuringspeed in free-living marmosets is particularly difficult for several methodologicalreasons.
Adults, juveniles, and infants were active foragers, though on 3 occasions infantshad access to animal protein by getting the remains of prey captured by olderindividuals. However, adults never freely shared the captured prey with the infants.Adults and juveniles tried to consume the captured item rapidly and actively avoidedthem. In fact, older individuals did not share any captured prey with the infantsduring the observations, despite their begging vocalizations. Similarly, Price and
major locomotion
0
1
2dense vegetation
sparse vegetation
age classfr
equ
enci
es/m
in
*
*
*
adults juveniles infants
Fig. 3 Frequencies of majorlocomotion in all age classes indense and sparse vegetation.Data are means ± SEM. Statis-tics: Mann-Whitney U test (2-tailed); *p≤0.05.
Saltatory Search in Free-Living Callithrix jacchus 889

Feistner (1993) found that food sharing in captive lion tamarins occurs mostly wheninfants have a restricted access to food. It is then possible that in locations wherefree-living marmosets regularly obtain food, older individuals are also reluctant toshare it with the infants, at least ones >9 wk old. We believe that behavior by olderindividuals may stimulate infants to start acquiring the skills required to capture theirown food.
The prey common marmosets captured were similar to the ones in previous studies(Alonso and Langguth 1989; Stevenson and Rylands 1988), with the exception of amollusc. Common marmosets have a flexible diet; thus one eating of a mollusc is notsurprising. To access the flesh of the mollusc, the adult marmoset broke the distalpart of the shell with its teeth and then sucked out the flesh.
Our results indicate that the saltatory search behavior in common marmosets isdependent on the density of the vegetation in which they are foraging. The factorsupports a claim of O’Brien et al. (1990), in which such a foraging strategy couldvary according to environmental conditions. Saltatory search also varied inaccordance with the age class, which previously received little attention in termsof the saltatory foraging strategy.
Adults and juveniles tended to display periods of minor locomotion morefrequently in dense vegetation. A similar behavioral adjustment occurs in adultthrushes and starlings, which also move shorter distances when searching for ahidden prey in a dense environment (Brownsmith 1977; Smith 1974), because thereis more area to be visually scanned in dense vegetation. In dense vegetation, shorterlocomotion distances are likely to be a better strategy for prey detection, being lessenergetically costly than moving further and leaving a closer and unsearched areathat is potentially inhabited by prey. In contrast to dense areas, sparse environmentscan be more easily scanned and relatively distant vegetation can be observed.Individuals can therefore afford to move further each time they locomote duringperiods of foraging.
Though in dense and sparse vegetation, adults and juveniles frequently appliedthe foraging strategies, infants demonstrated a different behavioral trend in that theyundertook major locomotion more frequently in dense vegetation than in sparsevegetation. Given that the amount of locomotion infants, juveniles, and adults usedwas similar during periods of foraging, the difference in foraging strategy by theinfants seems to explain why they captured fewer prey than adults and juveniles didin dense areas. Thus, it is possible that the observed infants are yet to develop themost appropriate foraging skills for all types of environment. Long-term studies onforaging groups of marmosets, focusing on individuals from birth to adulthood,should provide a considerable amount of information on the development offoraging skills in common marmosets.
However, when marmosets were confronted with sparse vegetation, the numberof prey captures did not differ statistically with age class, despite juveniles and adultsusing major locomotion more frequently, whereas infants employed minorlocomotion more frequently. A possible reason for this similarity may be a higherperception of predation risk, affecting their foraging behavior. The main predators ofthe Callitrichidae are aerial raptors (Ferrari and Lopes Ferrari 1990), to whichmarmosets are most vulnerable when in sparse vegetation because they are morevisible then. Such a variable could confound data obtained on foraging activity.
890 A. Souto, et al.

Unfortunately, there has been no field study on how the perception of predation riskaffects the behavior of marmosets. Future studies should specifically be carried outto address this important and complex issue (Caine 1996; Dunbar 1994).
We here introduce the concept of saltatory searching as a foraging strategy intothe primate literature, providing a clear example of a mammalian species thatemploys the strategy. Common marmosets possess the basic requirements for using asaltatory search strategy and they clearly forage following a stop and go pattern.Their foraging behavior is plastic, changing both with the age of the individual andwith the density of the vegetation.
The traditional definition of saltatory search states that the forager exhibits afrequent stop and go pattern, scanning for available recourses only while stationary(Bell 1991; O’Brien et al. 1990). Based on the claim of Anderson et al. (1997), onthe studies of Butler (2005) and Cooper (2005), and on our results, we propose anadjustment to the current definition of saltatory searching, as follows: Saltatorysearch describes foraging behavior of species that exhibit ≥1 MPM, scanning foravailable resources mostly or exclusively while stationary. We believe the definitioncharacterizes the saltatory search strategy more precisely.
Finally, it is quite likely that some other primate species forage for prey items thatare hard to detect and are widely distributed, and thus will use a stop-and-go pattern.To test the supposition we suggest that further researchers at least investigate theforaging behavior of other members of the Callitrichidae, because they share manyecological similarities with common marmosets (Dietz et al. 1997; Kierulff et al.2002).
Acknowledgments We thank 2 anonymous referees for their very useful suggestions and Dr. LewisHalsey for his advice on grammar. A Ph.D. grant (BX 171399-0) from the Brazilian Higher EducationAuthority—CAPES funded the study.
References
Alonso, C., & Langguth, A. (1989). Ecologia e comportamento de Callithrix jacchus (Primates:Callitrichidae) numa ilha de Floresta Atlântica. Revista Nordestina de Biologica, 6, 105–137.
Anderson, J. P., Stephens D. W., & Dumbar S. R. (1997). Saltatory search: A theoretical analysis.Behavioral Ecology, 8, 307–317.
Andersson, M. (1981). On optimal predator search. Theoretical Population Biology, 19, 58–86.Atkins, M. D. (1978). Insects in Perspective. New York: Collier Macmillan.Auricchio, P. (1995). Primatas do Brasil. São Paulo: Editora Terra Brasilis.Barbosa, A., & Moreno, E. (1999). Evolution of foraging strategies in shorebirds: An ecomorphological
approach. The Auk, 116, 712–725.Barbosa, V. S., Leal, I. R., Iannuzzi, L., & Almeida-Cortez, J. (2005). Distribution pattern of herbivorous
insects in a remnant of Brazilian Atlantic Forest. Neotropical Entomology, 34, 701–711.Beasom, S. L., & Haucke, H. H. (1975). A comparison of four distance sampling techniques in South
Texas live oak mottes. Journal of Range Management, 28, 142–144.Bell, W. J. (1991). Searching Behaviour: The Behavioral Ecology of Finding Resources. London:
Chapman and Hall.Brownsmith, C. B. (1977). Foraging rates of starling in two habitats. Condor, 79, 386–387.Butler, M. A. (2005). Foraging mode of the chameleon, Bradypodion pumilum: A challenge to the sit-
and-wait versus active forager paradigm? Biological Journal of the Linnean Society, 84, 797–808.Buzzi, J. Z. (2005). Entomologia. Editora da Universidade Federal do Paraná: Curitiba.
Saltatory Search in Free-Living Callithrix jacchus 891

Caine, N. G. (1996). Foraging for animal prey by outdoor groups of Geoffroy’s marmosets (Callithrixgeoffroyi). International Journal of Primatology, 17, 933–945.
Cody, M. L. (1968). On the methods of resource division in grassland bird communities. AmericanNaturalist, 102, 107–147.
Cooper, W. E. Jr. (2005). The foraging mode controversy: Both continuous variation and clustering offoraging movements occur. Journal of Zoology, London, 267, 179–190.
Curtis, J. T., & McIntosh, R. P. (1950). The interrelation of certain analytic and syntheticphytosociological characters. Ecology, 31, 434–455.
Day, L. B., Crews, D., & Wilczynski, W. (1999). Spatial and reversal learning in congeneric lizards withdifferent foraging strategies. Animal Behavior, 57, 393–407.
Dietz, J. M., Peres, C. A., & Pinder, L. (1997). Foraging ecology and use of space in wild golden lionstamarins (Leontopithecus rosalia). American Journal of Primatology, 41, 289–305.
Dunbar, R. I. M. (1994). Primate Social Systems. London: Chapman & Hall.Eibl-Eibesfeldt, I. (1999). Grundriss der vergleichenden Verhaltensforshung. München: Piper Verlag.Eifler, D. A., & Eifler, M. A. (1999). Foraging behavior and spacing patterns of the lizard Oligosoma
grande. Journal of Herpetology, 33, 632–639.Ferrari, S. F., & Lopes Ferrari, M. A. (1989). A re-evaluation of the social organization of the
Callitrichidae, with reference to the ecological differences between genera. Folia Primatologica, 52,132–147.
Ferrari, S. F., & Lopes Ferrari, M. A. (1990). Predator avoidance behaviour in the buffy-headed marmoset,Callithrix flaviceps. Primates, 31, 323–338.
Getty, T. (1993). Search tactics and frequency-dependent prey detection. American Naturalist, 141, 804–811.Getty, T., & Pullian, H. R. (1991). Random prey detection with pause-travel search. American Naturalist,
138, 1459–1477.Gullan, P. J., & Cranston, P. S. (2000). The Insects: An outline of Entomology. Oxford: Blackwell Science.Halsey, L. G., Bezerra, B. M., & Souto, A. S. (2006). Can wild common marmosets (Callithrix jacchus)
solve the parallel strings task? Animal Cognition. Online First. Published online: March 15, 2006.Hershkovitz, P. (1977). Living New World Monkeys (Platyrrini): With an Introduction to Primates, Vol. 1.
Chicago: University of Chicago Press.Hopcraft, J. G. C., Sinclair, A. R. E., & Packer, C. (2005). Planning for success: Serengeti lions seek prey
accessibility rather than abundance. Journal of Animal Ecology, 74, 559–566.Huey, R. B., & Pianka, E. R. (1981). Ecological consequences of foraging mode. Ecology, 62, 991–999.Ingram, J. C. (1977). Interaction between parents and infants and the development of independence in the
common marmoset (Callithrix jacchus). Animal Behavior, 25, 811–827.Kenagy, G. J. (1973). Daily and seasonal patterns of activity and energetic in a heteromyid rodent
community. Ecology, 54, 1201–1219.Kierulff, M. C. M., Raboy, B. E., Oliveira, P. P., Miller, K., Passos, F., & Prado, F. (2002). Behavioral
ecology of lion tamarins. In D. Kleiman, & A. B. Rylands, (eds.), Lion Tamarins: Biology andConservation (pp. 157–187). Washington, D.C.: Smithsonian Institution Press
Lehner, R. A. (1996). Handbook of Ethological Methods. London: Cambridge University Press.Leyhe, J. E., & Ritchison, G. (2004). Perch sites and hunting behavior of red-tailed hawks (Buteo
jamaicensis). Journal of Raptor Research, 38, 19–25.Maier, W., Alonso, C., & Langguth, A. (1982). Field observations on Callithrix jacchus jacchus L.
Zeitshrift Säugertierkunde 47:334–346.Maranhão, Z. C. (1976). Entomologia geral. São Paulo: Livraria Nobel S.A.Miyashita, T. (2005). Contrasting patch residence strategy in two species of sit-and-wait foragers under the
same environment: A constraint by life history? Ethology, 111, 159–167.Novotny, V., & Basset, Y. (2005). Host specificity of insect herbivores in tropical forests. Proceedings of
the Royal Society B—Biological Sciences, 272, 1083–1090.Novotny, V., Basset, Y, Miller, S. E., Weiblen, G. D., Bremer, B., Cizek, L., & Drozd, P. (2002). Low host
specificity of herbivorous insects in a tropical forest. Nature, 416, 841–844.O’Brien W. J., Browman, H. I., & Evans B. I. (1990). Search strategies of foraging animals. American
Scientist, 78, 152–160.O’Brien, W. J., Evans, B. I., & Browman, H. I. (1989). Flexible search tactics and efficient foraging in
saltatory searching animals. Oecologia, 80, 100–110.Price E. C., & Feistner A. T. C. (1993). Food sharing in lion tamarins: Tests of three hypotheses. American
Journal of Primatology, 31, 211–221.Reaney, L. T., & Whiting, M. J. (2002). Life on a limb: Ecology of the tree agama (Acanthocercus a.
atricollis) in southern Africa. Journal of Zoology, London, 257, 439–448.
892 A. Souto, et al.

Robakiewicz, P., & Daigle, W. (2004). Path quality and foraging time in the crab spider Misumenopsasperatus Hentz (Aranea: Thomisidae). Northeastern Naturalist, 11, 23–32.
Roberts, M., & Kohn, F. (1993). Habitat use, foraging behavior, and activity patterns in reproducingwestern tarsiers, Tarsius bancanus, in captivity—A management synthesis. Zoo Biology, 12, 217–232.
Rylands, A. B., & De Faria, S. (1993). Habitats, feeding ecology, and home range size in the genusCallithrix. In A. B. Rylands, (ed.), Marmosets and Tamarins: Systematics, Behaviour, and Ecology(pp. 262–272). Oxford: Oxford University Press.
Schiel, N. (2000). Das estratégias de captura do Callithrix jacchus (Primates, Callitrichidae) àcapacidade de fuga da presa. Master’s thesis, Brazil: Universidade Federal de Pernambuco.
Secor, S. M. (2005). Physiological responses to feeding, fasting and estivation for anurans. The Journal ofExperimental Biology, 208, 2595–2608.
Shine, R., & Li-Xing, S. (2002). Arboreal ambush site selection by pit-vipers Gloydius shedaoensis.Animal Behavior, 63, 565–576.
Siegel, S. (1975). Estatística não-paramétrica para ciências do comportamento. São Paulo: EditoraMc-Graw-Hill.
Smith, J. N. M. (1974). The food searching behaviour of two European thrushes. II: The adaptativeness ofthe search patterns. Behaviour, 59, 1–61.
Sonerud, G. A. (1992). Search tactics of pause and travel predators: Adaptative adjustments of perchingtimes and move distances by hawk owls (Surnia ulula). Behavioral Ecology and Sociobiology, 30,207–217.
Stevenson, M. F., & Rylands, A. B. (1988). The marmosets, genus Callithrix. In A. R. Mittermeier, B. A.Rylands, A. Coimbra-Filho, & B. A. G. Fonseca, (eds.), Ecology and Behaviour of NeotropicalPrimates, Vol. 2 (pp. 131–222). Washington, D.C.: World Wildlife Fund
Sussman, R. W., & Kinzey, W. G. (1984). The ecological role of the Callitrichidae: A review. AmericanJournal of Physical Anthropology, 64, 419–449.
Tye, A. (1989). A model of search behaviour for the northern wheatear Oenanthe oenanthe (Aves,Turbidae) and other pause-travel predators. Ethology, 83, 1–18.
Saltatory Search in Free-Living Callithrix jacchus 893