saturable transport of insulin from plasma central nervous
TRANSCRIPT

Saturable Transport of Insulin from Plasmainto the Central Nervous System of Dogs In VivoA Mechanism for Regulated Insulin Delivery to the Brain
Gail D. Baura, David M. Foster, Daniel Porte, Jr., Steven E. Kahn, Richard N. Bergman,*Claudio Cobelli,* and Michael W. SchwartzDepartments of Bioengineering and Medicine, University of Washington and Seattle Veterans Affairs Medical Center, Seattle,Washington 98195, *Department of Physiology and Biophysics, University of Southern California, Los Angeles, California 90033; andtDepartment of Electronics and Informatics, Universita di Padova, 35131 Padova, Italy
Abstract
By acting in the central nervous system, circulating insulin mayregulate food intake and body weight. We have previouslyshown that the kinetics of insulin uptake from plasma into cere-brospinal fluid (CSF) can best be explained by passagethrough an intermediate compartment. To determine if trans-port kinetics into this compartment were consistent with aninsulin receptor-mediated transport process, we subjected over-night fasted, anesthetized dogs to euglycemic intravenous insu-lin infusions for 90 min over a wide range of plasma insulinlevels (69-5,064 ,U/ml) (n = 10). Plasma and CSFsampleswere collected over 8 h for determination of immunoreactiveinsulin levels, and the kinetics of insulin uptake from plasmainto CSF were analyzed using a compartmental model withthree components (plasma -- intermediate compartment -CSF). By sampling frequently during rapid changes of plasmaand CSFinsulin levels, we were able to precisely estimate threeparameters (average standard deviation 14%) characterizingthe uptake of insulin from plasma, through the intermediatecompartment and into CSF (klk2); insulin entry into CSFandinsulin clearance from the intermediate compartment (k2 + k3);and insulin clearance from CSF (k4). At physiologic plasmainsulin levels (80±7.4,gU/ml), klk2 was determined to be 10.7X 10'±+1.3 X 10-6 min2. With increasing plasma levels, how-ever, k1k2 decreased progressively, being reduced sevenfold atsupraphysiologic levels (5,064 ,U/ml). The apparent KMofthis saturation curve was 742 uUU/ml ( - 5 nM). In contrast,the rate constants for insulin removal from the intermediatecompartment and from CSF did not vary with plasma insulin(k2 + k3 = 0.011±0.0019 min' and k4 = 0.046±0.021 min').Weconclude that delivery of plasma insulin into the centralnervous system is saturable, and is likely facilitated by an insu-lin-receptor mediated transport process. (J. Clin. Invest. 1993.92:1824-1830.) Key words: central nervous system * compart-mental model * insulin action * SAAMmodeling program
Introduction
Several lines of investigation suggest that circulating insulinlevels within the physiologic range exert biologic effects onfunctioning of the central nervous system (CNS). For exam-
Address reprint requests to Dr. Michael W. Schwartz, Metabolism(151), Va Medical Center, 1660 South Columbian Way, Seattle, WA98108.
Receivedfor publication 10 September 1992 and in revisedform 9May 1993.
ple, experimental elevations of plasma insulin to postprandialvalues suppress feeding behavior (1, 2), an effect also observedduring direct insulin infusion into the brain (3). That this oc-curs despite the extensive network of endothelial tight junc-tions in cerebral vasculature comprising the blood-brainbarrier, which limits diffusion of peptides of this size, suggeststhat circulating insulin may enter the brain via a facilitatedtransport process (4). In vitro studies (5,6) demonstrating thatinsulin transport across vascular endothelium is dependent onits binding to endothelial insulin receptors have led to the hy-pothesis that receptor-mediated insulin transport may facilitatethe delivery of circulating insulin to target tissues in vivo. More-over, brain microvascular endothelium expresses insulin re-ceptors (7), and the uptake of plasma insulin into cerebrospi-nal fluid (CSF)' demonstrates specificity when compared toanalogues with reduced affinity for the insulin receptor, such asproinsulin (8). These and other studies (9) provide support forthe hypothesis that the entry of circulating insulin into thebrain is facilitated by an insulin receptor-mediated transportprocess. However, quantitative measurements of insulin trans-port kinetics into the CNShave yet to address the possibility ofsaturable insulin transport.
In our previous work ( I0), we provided evidence that insu-lin uptake from plasma into CSF requires passage through anintermediate compartment, and reported on the use of a com-partmental model to facilitate the analysis of this uptake pro-cess. Wenow report the quantitation of the dose-response re-lationship between the plasma insulin level and kinetics ofinsulin uptake from plasma, through an intermediate compart-ment, and into CSF. Wehypothesized that if insulin receptors,localized to either the blood-brain or blood-CSF barrier, facili-tate this uptake process, then the rate constant characterizingCSFinsulin uptake should be saturable, decreasing in magni-tude with increasing plasma insulin levels. Moreover, theplasma insulin concentration at which half-maximal transportoccurs (KM) should be comparable to the dissociation constant(KD) of the insulin receptor.
Methods
Study animals and conditions. Six normal adult male mongrel dogsweighing 20-30 kg were studied after an overnight fast. Their care wassupervised by a licensed veterinarian. The dogs were housed in individ-ual cages that included a 5 X 20 ft area for exercise and were fed 0.3kg/d of dry standard dog laboratory diet (Wayne Pro-Mix, Allied
The Journal of Clinical Investigation, Inc.Volume 92, October 1993, 1824-1830
1. Abbreviations used in this paper: CSF, cerebrospinal fluid; IRI, im-munoreactive insulin.
1824 Baura et al.

Mills, Inc., Memphis, TN) with unlimited access to water. For eachstudy, each dog was anesthetized with thiamylol (Surital, Parke-Davis,Morris Plains, NJ), 20 mg/kg i.v., and was placed on mechanical ven-tilation, with 1-2% halothane and 40% 02- Intravenous catheters wereplaced for both sampling and infusion, and the cisternum magnumwascannulated with a no. 22 gauge spinal needle for sampling of CSF, asreported previously (10). Body weight was measured immediately be-fore each study. All studies were approved by the Animal Care Com-mittee of the Seattle VA Medical Center.
Insulin infusion and sampling. The study protocol consisted of a90-mm i.v. insulin infusion period with sampling of plasma and CSFfor a total of 490 min. Intravenous insulin was administered as a 3-minprimed infusion (8.5-50 mU/kg-min) commencing at t = 0 min, fol-lowed by a continuous infusion for 87 min at 20% of the primed infu-sion rate. Blood samples ( 1.8 ml) were obtained at t = - 10, -5, 1,2, 3,4, 6, 8, 10, 13, 16, 20, 25, 30, 35, 40, 65, and 90 min, at 5-min intervalsfor 90 < t < 150 min, and at 20-min intervals for 150 < t < 490 min.CSFsamples (0.4 ml) were obtained at t = - 10, -5, 20, 40, 65, 90, 95,100, 110, 120, 135, and 150 min and thereafter at 20-min intervalsuntil 490 min. In three studies (P81, P520, and P1195), additionalblood and CSF samples were taken at 20-min intervals for 510 < t< 590 min. Euglycemia was maintained in all studies by a variable-ratei.v. infusion of 50%dextrose with on-line monitoring of blood glucoselevels using a hand-held, computerized glucose meter (Glucoscan, Life-scan, American Medical Systems, Cincinnati, OH). Blood and CSFsamples were placed on ice before processing. After blood separation,both plasma and CSFwere frozen at -20'C until assay. Plasma levelsof immunoreactive insulin (IRI) were measured in duplicate by radio-immunoassay using a modification of the double-antibody method(11). CSF IRI was measured in triplicate using a further modificationof this method which enhances its sensitivity (8, 12). Plasma glucosewas determined by the glucose oxidase method with a glucose autoana-lyzer (Beckman Instruments, Inc., Brea, CA). The average plasma in-sulin concentration during infusion, Pave, was determined by taking theaverage of plasma insulin values during the interval 1 . t < 90 min.Two dogs were studied once at either a low or high insulin dose, andfour were studied twice, at both a low and high insulin dose. Thus, 10independent data sets were obtained.
Multicompartmental model. A schematic illustration of the multi-compartment model used to interpret the data is shown in Fig. 1; thismodel was previously described in Schwartz et al. (10). Because of thedelay in the appearance of insulin in the CSFafter intravenous insulininfusion, insulin entry from plasma into CSFcan best be explained bypassage through an intermediate compartment, INS,(t). Insulin inplasma, INSp(t), enters CSF, INSc(t), after passing through INSI(t),and irreversible loss can occur from both INSc(t) and INSI(t). INSp(t)provides a functional description of the plasma data providing input tothe model. By assuming that flux of insulin from the CNSback intoplasma has a negligible influence on the plasma concentration, theneed to postulate a structure for the kinetics of insulin outside of thebrain is eliminated.
In this model, k, represents the uptake of plasma insulin into theintermediate compartment, and k2 represents the movement of insulinfrom the intermediate compartment into CSF. k3 represents insulinclearance from the intermediate compartment, and k4 represents insu-lin clearance from CSF. All rate constants are expressed in min-'.
Using this model, the rates of change of exogenously administeredinsulin in the intermediate compartment, INS,(t), and of CSFinsulin,INSc(t), are described by the following differential equations:
d-INS(t) = kINSp(t) - (k2 + k3)INSI(t) (1)dt
dINSc(t) = k2INSI(t) - k4INSc(t) (2)
INSp(0) = INSJ(0) = INSc(0) = 0 (3)All insulin variables, INSX(t), are expressed in concentration units ofmicrounits per milliliter. To obtain the initial conditions of Eq. 3, basal
plasma and CSFlevels were subtracted from all values of each data setbefore modeling, such that for modeling purposes the insulin concen-tration in each compartment was set equal to zero at t = 0 min. There-fore, only the kinetics of infused insulin were analyzed, and it was notnecessary to assume that at the basal state all CSF IRI must be derivedfrom plasma. Because the intermediate compartment was not directlysampled, only the rate constant characterizing insulin clearance fromCSF, k4, can be identified independently, whereas ki, k2, and k3 are notindependently identifiable. Instead, the product klk2, which representsthe throughput of insulin from plasma, through the intermediate com-partment, and subsequently into CSF, and sum (k2 + k3), which repre-sents insulin flux from the intermediate compartment into CSFas wellas insulin clearance from the intermediate compartment, were esti-mated. A detailed derivation of unique parameter identification is pro-vided in the Appendix.
Parameter identification. By using the mathematical modeling pro-gram SAAM( 13) on a model 486 computer (Gateway 2000, Gateway,Sioux City, SD), the rate constants kk2 (min2), (k2 + k3) (min-'),and k4 (min -') were estimated from the plasma and CSFinsulin data.To reduce the number of samples and therefore total fluid volumesrequired per study, only CSF insulin data points taken during t 2 90min were used for modeling. It can be shown that as long as enoughplasma and CSFsamples are taken and steady state is reached, the sameparameters will be obtained from CSFsamples taken at 0 < t < 3, 3 < t< 90, or t . 90 min ( 14). The intraassay errors for the plasma and CSFinsulin radioimmunoassays were assessed by measuring eight samplesof various insulin concentrations (data not shown). These error esti-mates (plasma insulin assay 4% for concentrations < 80 AU/ml, 12%otherwise; CSF insulin assay 1 1%) were used by SAAMto determinethe optimal curve fit and to identify the rate constant parameters. Foreach parameter, the corresponding coefficient of variation (CV), alsoknown as the fractional standard deviation, was determined from thecovariance matrix generated by SAAM.
Independent estimation of the CSFinsulin clearance parameter, ki4.To obtain an independent estimate of k4, the CSF insulin clearanceparameter, insulin (750 /U in 750 AI of saline) was infused over 1 mininto the cistemum magnumof three dogs which had been fasted over-night, anesthetized, and maintained on mechanical ventilation. Imme-diately before the infusion was administered, 750 Ml of CSF was re-moved from the cisternum magnum, in order to preserve the originalvolume of CSF during the infusion. CSF samples (0.4 ml) were ob-tained at t = -10, -5, 2, 5, 10, 15, 25, and 40 min, and thereafter at20-min intervals till 240 min. The resulting CSFlevels decayed biex-ponentially, and were fitted to a two-compartment model. This two-compartment analysis has previously been used to model transportkinetics of substances administered intracerebroventricularly, with onecompartment representing CSFand the other representing brain inter-stitium ( 15, 16). The CSFclearance rate was calculated by dividing theestimated bolus concentration administered by the area under thecurve of the two exponentials ( 17 ).
Estimation of KM, the plasma insulin concentration at which half-maximal transport occurs. To estimate KM, the plasma insulin concen-tration at which half-maximal transport occurs, the transport velocityfor various plasma insulin concentrations was required. Velocity isdetermined by multiplying ki, the rate constant representing uptake ofplasma insulin into the intermediate compartment, by the correspond-ing plasma insulin concentration during infusion, Pave. As kI could notbe uniquely identified, the product klk2, which estimated ki, was usedfor this calculation. Therefore, velocity* k2, which is directly propor-tional to the rate of insulin uptake from plasma into CSF, was insteadcalculated. Velocity * k2 due to nonspecific uptake is linear, such that athigh plasma insulin levels, velocity * k2 does not plateau. As this linearcomponent is readily apparent at high plasma insulin levels, ve-
locity *2 due to nonspecific uptake was estimated through graphicalanalysis as 2 X 10-7*Pave. This estimate was subtracted from veloc-ity * k2 to isolate velocity * k2 due to saturable uptake ( 1 8 ). Velocity* k2was then plotted as a function of Pave. KMwas calculated by using a
variation of the Michaelis-Menten equation,
Saturable Transport of Plasma Insulin into the Central Nervous System 1825

Velocity*k2= VMAxk2PavePaVe + KM
to fit the resulting curve. In this equation, VMAX= maximum velocity.
Results
The 90-min intravenous insulin infusion protocol resulted in acharacteristic time course of change in CSF IRI levels, illus-trated in Fig. 2. During i.v. insulin infusion, CSFinsulin levelsrose gradually, reaching peak values at 105 min ( 15 min afterdiscontinuation of infusion), and fell exponentially thereafter.The estimated parameters and corresponding coefficients ofvariation for each study are given in Table I. The mean CVforall estimated parameters was 14%.
The data in Table I demonstrate that studies conducted atlow plasma insulin levels are associated with relatively highvalues of k1k2. This was evident when examining all dogs as agroup, as well as when comparing results obtained in separatestudies at two different plasma levels. Examining data obtainedfrom two studies in individual dogs, the value of k1k2 washigher in studies conducted at lower plasma insulin levels in allcases, whereas no consistent relationship between either (k2+ k3) or k4 and the plasma level was evident.
To further characterize these relationships, klk2, (k2 + k3),and k4 were plotted as a function of the average plasma insulinconcentration during infusion, Pave, to determine dose-re-sponse relationships. As shown in Fig. 3, the parameter charac-terizing insulin uptake from plasma into CSFvaried inverselywith plasma insulin levels, being reduced sevenfold from physi-ologic to supraphysiologic levels. The apparent KM for thistransport process was determined from a Michaelis-Mentendose-response curve based on the data in Table I. As shown inFig. 4, the apparent KMwas approximated to be 742 ,uU/ml( - 5 nM). In contrast, the mean rate constant values for insu-lin removal from the intermediate compartment (k2 + k3) andinsulin clearance from CSF (k4) did not appear to vary withplasma insulin (k2 + k3 = 0.011±0.0019 min-' and k4= 0.046±0.021 min-') (Fig. 5). Therefore, delivery of circu-lating insulin into the intermediate compartment and subse-quently into CSFdemonstrates saturable kinetics, whereas in-sulin clearance from the intermediate compartment does not.Data are insufficient to assess the possibility that a saturableprocess also contributes to CSF insulin clearance.
To validate the model-derived parameter representing CSFinsulin clearance, the mean CSF clearance rate constant was
INS(t)k 1 (t)kk
Figure 1. Schematic illustration of the pharmacokinetic model used toanalyze the kinetics of insulin uptake from plasma into CSFin thedog. In this model, k1 represents the uptake of plasma insulin into theintermediate compartment, hypothesized to occur across the blood-brain barrier, and k2 represents the transport of insulin from the in-termediate compartment into CSF. k3 represents insulin clearancefrom the intermediate compartment, and k4 represents insulin clear-ance from CSF. All rate constants are expressed in min-1. Modifiedfrom Schwartz et al. (10).
Table I. Identified Parameters from Optimal Curve Fitsof Data from Nine studies in Dogs of Various Insulin Doses
Study P.". k,k2 k2 + k3 k4
UU/mI min-2 min-'
P69 69 10.2e-6 0.0086 0.060(14%) (11%) (16%)
P81 81 9.1e-6 0.014 0.043(14%) (28%) (16%)
P83 83 11.5e-6 0.011 0.10(16%) (8%) (19%)
P86 86 12.0e-6 0.012 0.047(10%) (19%) (23%)
P520 520 6.7e-6 0.011 0.026(4%) (5%) (8%)
P989 989 7.6e-6 0.0095 0.035(12%) (18%) (22%)
P1195 1195 3.8e-6 0.011 0.034(16%) (24%) (22%)
P1203 1203 3.1e-6 0.012 0.032(10%) (11%) (19%)
P2076 2076 3.4e-6 0.011 0.042(13%) (12%) (18%)
P5064 5064 1.6e-6 0.0073 0.042(8%) (6%) (12%)
In studies P81 and P1194, plasma insulin samples were not taken at t= 1, 2, 3, 4, 6, 8, 10, 13, and 16 min, which resulted in higher coeffi-cients of variation than in studies in which these samples were ob-tained. Studies conducted in the same dog were P69 and P989, P81and P1203, P520 and P1194, and P86 and P5064.
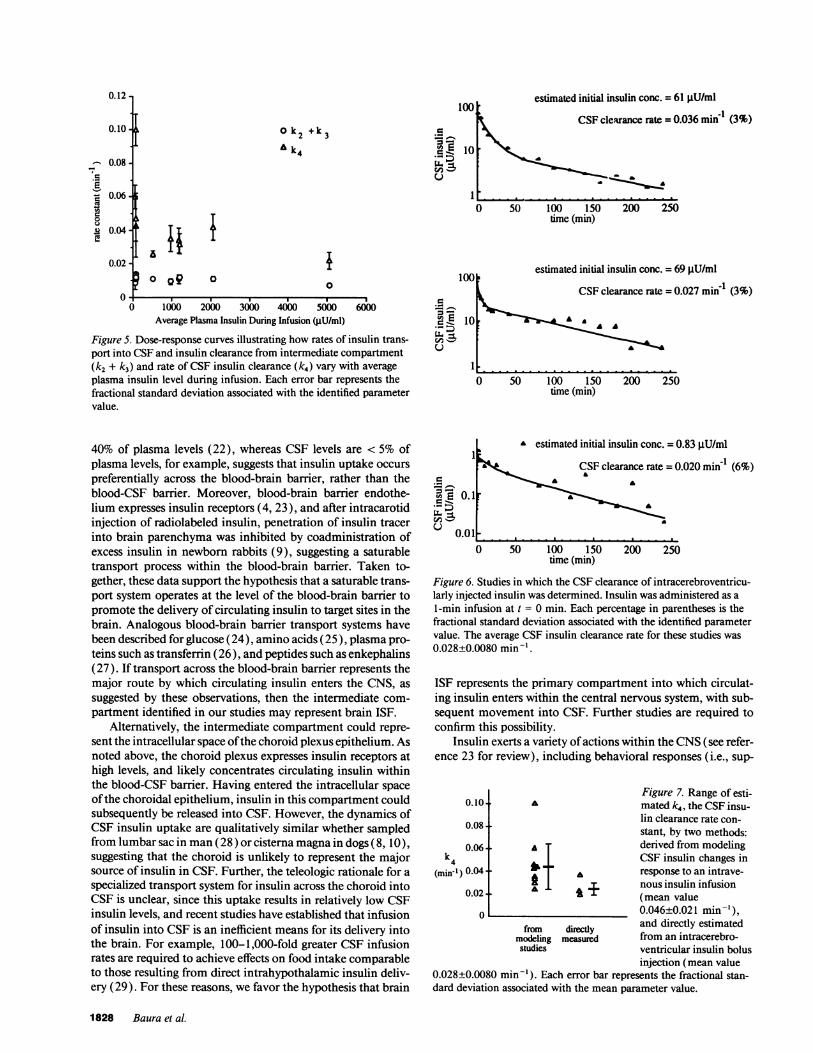
also determined independently by modeling data from intra-cisternal insulin injection (n = 3). The intracisternal insulininjection protocol resulted in a characteristic time course ofchange in CSF IRI levels, illustrated in Fig. 6. The mean CSFinsulin clearance rate determined from these studies was 0.028± 0.0080 min-', which is within the error of the mean value ofk4 determined during i.v. insulin infusion (Fig. 7). These re-sults validate the CSF insulin clearance parameter identifiedduring intravenous insulin infusion.
Discussion
In the current studies, we analyzed the kinetics of CSF insulinuptake from plasma using a previously presented multicom-partmental model ( 10). Our results indicate that the efficiencywith which circulating insulin enters CSFvia passage throughan intermediate compartment varies inversely with the plasmainsulin concentration. These data support the hypothesis thatinsulin entry into the CNSfrom plasma is facilitated by a satu-rable transport process. As the efficiency of uptake into theCNSdecreases sevenfold with increasing plasma insulin con-centration, we estimate that 85%ofthis uptake is mediated by asaturable process at physiologic plasma insulin levels (< 100,uU/ml). The apparent KMof this transport process was 742/gU/ml (- 5 nM), which is similar to the KD of the insulinreceptor identified in brain microvasculature (2 nM) (19).Therefore, data are consistent with the hypothesis that insulinreceptors facilitate transport of plasma insulin into the centralnervous system.
1826 Baura et al.

c
=v 83 PU/mi
E 3EXL >
Ic (' _ M _ W _ __
_ _0) 1()() 2(X() .300(t 4(X) 5(X)
time (min)
._ 0).
_-
().
c 8(1
B. c4_
,)
c
-._E EL
c _
._ _
V, _4c;
6.4.20
X]
O)
I(X) 200 (X) 400time (min)
V
.hd 4 75(X)
Pave = 520 ilU/ml
OF ^- - - _^^ --be _ ^_-
0 100 200 300 400 500 600time (min)
6
4 L2
0 1(X) 200 300 400 500 600time (min)
In
0
0 1000 2000 3000 4000 5000Average Plasma Insulin During Infusion (iU/ml)
6000
Figure 3. Dose-response curve illustrating the relationship betweenthe rate of uptake of plasma insulin into the intermediate compart-ment and subsequently into CSF. kjk2 decreases sevenfold with in-creasing plasma insulin levels from the physiologic to supraphysio-logic range during intravenous insulin infusion. Each error bar repre-sents the fractional standard deviation associated with the identifiedparameter value.
c
, _1.2(X)C. S* E 8(X)
:D-
c
r__C
P?==1195 pU/ml
t) I(X) 200 30) 4(X) (X)
time (min)8 [6[4 (mm)
() I(X) 2(X 3() 4X) S(X)time (min)
the choroid plexus. One hypothesis to account for this observa-tion is that insulin entering the choroid plexus is sequestered bychoroidal insulin receptors, which are expressed at high con-
centrations, preventing its diffusion into CSF, and are possibly6(X) responsible for degrading insulin following binding, as occurs
in the placenta (20, 21 ).The intermediate compartment through which insulin
passes en route to CSF likely represents either brain ISF or
choroid plexus epithelium. Although the identity of this com-
partment can not be established without undertaking the diffi-60( cult task of sampling the compartment itself, data are available
which indirectly address these two possibilities. The observa-Figure 2. Representative studies in which the kinetics of CSFuptakeof intravenously infused insulin were determined. Insulin was infusedduring 0 t 90 min. Studies shown are representative of low (A),medium (B), and high (C) insulin doses. Upper panels: time course
of plasma insulin. Lower panels: time course of CSFinsulin and op-timal curve fit, from which rate constant parameters were derived.
Two barrier systems exist in the central nervous systemwhich regulate solute movement between the blood and CNS.The first of these is the blood-brain barrier, which arises fromepithelial-like tight junctions that virtually cement adjoiningcapillary endothelium together in the brain microvasculature.Similarly, the blood-CSF barrier arises from tight junctions be-tween adjacent epithelial cells investing the choroid plexus, theprimary site of CSF formation. In general, most solutes gainaccess from blood to CSF via the blood-CSF barrier at thechoroid plexus. Transport of solutes such as Na' occurs di-rectly from blood to CSF, and does not necessitate postulatingtransport through an intermediate compartment ( 16). Thus,while insulin may gain access to CSF via either of the CNSbarrier systems, the observation that plasma insulin passes
through an intermediate compartment en route to CSFattestsnot only to the uniqueness with which insulin enters the CSF,but suggests that a barrier to diffusion of insulin exists across
tion that insulin concentrations in rat brain extracts are - 30-
0.008
ll.l.5EE
=L1-1
eq
19
>1
8lu
0.006
0.004-
0.002-
aa
0 1000 2000 3000 4000 5(000Average Plasma Insulin During Infusion (pU/ml)
Figure 4. Michaelis-Menten curve, based on calculated kk2, the rateof insulin uptake from plasma into intermediate compartment andsubsequently into CSF, for various insulin doses. The apparent KM,the concentration during which half maximal transport occurs, is742 ,U/ml (-. 5 nM).
Saturable Transport of Plasma Insulin into the Central Nervous System 1827
A.
.kw
By
a
Velocity * k2 = 0.0074 P
P +74ave
a
Pave
42
_m
estimated initial insulin conc. = 61 ptU/ml100o
o k2 + k3Ak
a i? o Q9 0
=._
cw
ii'
UA
CSFclearance rate = 0.036 min 1 (3%)
_ft. _
10o
1000
O 1000 2000 3000 4000 5000Average Plasma Insulin During Infusion (pU/ml)
600010
Figure 5. Dose-response curves illustrating how rates of insulin trans-port into CSFand insulin clearance from intermediate compartment(k2 + k3) and rate of CSF insulin clearance (k4) vary with averageplasma insulin level during infusion. Each error bar represents thefractional standard deviation associated with the identified parametervalue.
U.O -3
co
I .L . .
o 50 100 150time (min)
200 250
estimated initial insulin conc. = 69 gU/mlCSFclearance rate = 0.027 mind (3%)
A -
AA
0 50 100 150
time (min)200 250
40% of plasma levels (22), whereas CSF levels are < 5% ofplasma levels, for example, suggests that insulin uptake occurs
preferentially across the blood-brain barrier, rather than theblood-CSF barrier. Moreover, blood-brain barrier endothe-lium expresses insulin receptors (4, 23), and after intracarotidinjection of radiolabeled insulin, penetration of insulin tracerinto brain parenchyma was inhibited by coadministration ofexcess insulin in newborn rabbits (9), suggesting a saturabletransport process within the blood-brain barrier. Taken to-gether, these data support the hypothesis that a saturable trans-port system operates at the level of the blood-brain barrier topromote the delivery of circulating insulin to target sites in thebrain. Analogous blood-brain barrier transport systems havebeen described for glucose (24), amino acids (25), plasma pro-teins such as transferrin (26), and peptides such as enkephalins(27). If transport across the blood-brain barrier represents themajor route by which circulating insulin enters the CNS, as
suggested by these observations, then the intermediate com-
partment identified in our studies may represent brain ISF.Alternatively, the intermediate compartment could repre-
sent the intracellular space of the choroid plexus epithelium. Asnoted above, the choroid plexus expresses insulin receptors athigh levels, and likely concentrates circulating insulin withinthe blood-CSF barrier. Having entered the intracellular space
of the choroidal epithelium, insulin in this compartment couldsubsequently be released into CSF. However, the dynamics ofCSF insulin uptake are qualitatively similar whether sampledfrom lumbar sac in man(28) or cisterna magna in dogs (8, 10),suggesting that the choroid is unlikely to represent the majorsource of insulin in CSF. Further, the teleologic rationale for a
specialized transport system for insulin across the choroid intoCSF is unclear, since this uptake results in relatively low CSFinsulin levels, and recent studies have established that infusionof insulin into CSFis an inefficient means for its delivery intothe brain. For example, 100-1,000-fold greater CSF infusionrates are required to achieve effects on food intake comparableto those resulting from direct intrahypothalamic insulin deliv-ery (29). For these reasons, we favor the hypothesis that brain
0.1
0.011
a estimated initial insulin conc. = 0.83 uU/mI
~~~~CSFclearance rate:=
a
=0.020 min'I (6%)
0 50 100 150 200 250time (min)
Figure 6. Studies in which the CSFclearance of intracerebroventricu-larly injected insulin was determined. Insulin was administered as a1-min infusion at t = 0 min. Each percentage in parentheses is thefractional standard deviation associated with the identified parametervalue. The average CSF insulin clearance rate for these studies was0.028±0.0080 min-'.
ISF represents the primary compartment into which circulat-ing insulin enters within the central nervous system, with sub-sequent movement into CSF. Further studies are required toconfirm this possibility.
Insulin exerts a variety of actions within the CNS(see refer-ence 23 for review), including behavioral responses (i.e., sup-
0.10.
0.08-
0.06-k4
(mind ) 0.04-
0.02-
o
A
AT
A~L
a
i +
from directlymodeling measured
studies
Figure 7. Range of esti-mated k4, the CSFinsu-lin clearance rate con-
stant, by two methods:derived from modelingCSF insulin changes inresponse to an intrave-nous insulin infusion
(mean value0.046±0.021 min'),and directly estimatedfrom an intracerebro-ventricular insulin bolusinjection (mean value
0.028±0.0080 min-'). Each error bar represents the fractional stan-dard deviation associated with the mean parameter value.
1828 Baura et al.
S0.10.
0.08.
0.06.
0.04-
0.02-
fi
1-
.s
0u84IL
0.12-
vu
II
.-

pression of food intake), regulation of neuropeptide biosynthe-sis and catecholamine metabolism, and activation of the sym-pathetic nervous system. Collectively, these central actionspromote a state of negative energy balance, favoring weightloss. These observations have led to the hypothesis that circu-lating insulin acts in the brain as a negative feedback signalproportional to body adipose mass in the process of long-termbody weight regulation (30, 31 ). From the available data, webelieve that plasma insulin is delivered to brain interstitial fluidat least in part via an insulin receptor-mediated transcytoticprocess. As most insulin-receptor mediated actions are subjectto a variety of regulatory influences, it is possible that this insu-lin transport process may also be regulated, which would beunlikely were transport dependent solely on nonsaturablemechanisms such as diffusion. Theoretically, regulation ofCNS insulin transport could alter the efficiency with whichcirculating insulin gains access to target sites in the brain, andcould thus represent an important determinant of the levelabout which body weight is maintained.
Receptor-mediated insulin transport from vasculature totissue interstitium may regulate processes other than bodyweight. The movement of insulin from blood into the intersti-tium of tissues is an important determinant of the rate of insu-lin action on carbohydrate utilization in vivo. Yang et al. (32)have shown a strong correlation between the dynamics of tho-racic duct lymph insulin, which is derived from interstitialfluid, and insulin mediated glucose utilization. Further, lymphinsulin is proportional to glucose utilization within the physio-logic but not pharmacologic range of insulin, indicating thattranscapillary insulin transport is rate-limiting for insulin ac-tion in this range (33). Thus, the movement of insulin fromvasculature to interstitium is a critical step in determining tis-sue sensitivity to insulin, and it is possible that defects in trans-port may contribute to the pathogenesis of insulin resistance inconditions such as non-insulin-dependent diabetes mellitusand obesity. It should be stressed, however, that while we havedemonstrated saturable uptake of insulin into CSF, studieshave yet to establish a saturable mechanism of insulin trans-port into peripheral interstitial fluid. It is possible that mecha-nisms operating within brain vasculature differ from those ofother tissues where insulin uptake is concerned.
It should be noted that the estimated k1k2 values in thesupraphysiologic range of plasma insulin levels in the currentstudies are about half those calculated previously (10). Thesedifferences are probably attributable to modified plasma andCSFsampling schedules, which have improved parameter esti-mates. Specifically, the number of samples taken during rapidchanges of plasma and CSFinsulin levels was increased. Thischange of sampling times reduced the variability of estimatedparameters, compared to those generated using conventionalsampling procedures (34). In our previous work, estimatedparameters often had associated coefficients of variation> 100%. In contrast, the mean coefficient of variation for 30identified parameters in the current studies is 14%. In bothcases, the true parameter value was located within the coeffi-cient of variation of each estimate. However, because the coeffi-cients of variation have been reduced in the current studies, themean estimates are now much closer to the true parametersthan in the previous work.
In the current studies, we measured k4, the rate of insulinclearance from CSF, to be 0.046±0.021 min-'. This clearancerate was independently verified by intracerebroventricular ad-
ministration of insulin, and is much higher than the rate ofbulk flow CSFclearance in the dog (0.0025±0.0003 min '), asdetermined from previous inulin studies (10, 15, 16). Thislarge disparity between CSFclearance rates suggests that mech-anisms other than bulk flow contribute to insulin removalfrom CSF. One possibility is that insulin receptors on the cho-roid plexus bind and remove/degrade insulin in CSF, as sug-gested by Manin et al. (35). While this would imply a saturablemechanism for CSF insulin removal, saturability would not bedemonstrable at such low CSF insulin levels as occurred in ourstudies (< 10 ,U/ml). Thus, while CSF insulin removal mayinvolve a saturable mechanism, further study is required toaddress this possibility.
In summary, we have demonstrated that uptake of plasmainsulin into the central nervous system is saturable. This mecha-nism is consistent with insulin binding to blood-brain barrierand/or blood-CSF insulin receptors and subsequent transcyto-sis into the central nervous system.
Appendix
To determine if model parameters are uniquely identifiable,the system impulse response, that is, the cause-effect relation-ship between input and output, is analyzed. For this system, theCSF impulse response is:
INSc(t) = 1c- _c- kw [e-w- - e-(k2+k3)t]I(5)
For theoretical, or a priori, identifiability, all the rate constantsmust be uniquely identified from Eq. 4. For the functionD(e-' - end), D, a, and 3 can be determined from a semilogplot of the natural log of this function versus time. Therefore,k4 can be identified, as can the quantity (k2 + k3). These threeconstants make up the denominator of the terms multiplyingthe exponentials, so the numerator (klk2) can also be identi-fied. In order to uniquely identify k, k2, and k3, more infor-mation is needed (36). Therefore, this model is not theoreti-cally identifiable.
Eqs. 1 and 2 can be rearranged to simplify parameter identi-fication by dividing both sides of Eq. 1 by ki, subtracting andadding the same term to Eq. 1, and rewriting Eq. 2 in terms ofINSI(t)/kl:
d INS, (t) INS-(t)= INS~() (k2+ k3- k~k2-Fk1k2) (6)
dt k, k
d INSc(t) = k2l- k INSI(t) -_k4INSc(t)dt ' ,(7).
These equations are realized in the rearranged model of Fig. 8.
1 ki k2k4INS p(t) I (t) INS$ (t)
k2 +k3 - ki k2
Figure 8. Schematic illustration of the rearranged pharmacokineticmodel used to analyze the kinetics of insulin uptake from plasmainto CSF.
Saturable Transport of Plasma Insulin into the Central Nervous System 1829
(5)

The parameters identified with this model are k,k2, (k2 + k3- klk2), and k4. Once klk2 and (k2 + k3 - klk2) are determined,(k2 + k3) is also known.
Acknowledgments
Wegratefully acknowledge critical assistance with study design pro-vided by Dr. Alfredo Ruggieri. Expert technical assistance was pro-vided by Rix Kuester, Wendy Hamer, and Dave DeGroot in conduct-ing these studies. Insulin assays were performed by Richard Chan andHong Nguyen, and glucose assays by Ruth Hollingworth.
This work was supported by grants RRO2176, DK-12829, DK-17047, and DK-29867 from the National Institutes of Health, by anAssociate Investigator Award and Research Associate Award of theDept. of Veterans Affairs, and by an NIH Physician Scientist Award.
References
1. Woods, S. C., D. Porte, Jr., E. Bobbioni, E. Ionescu, J. Sauter, F. Rohner-Jeanrenaud, and B. Jenrenaud. 1985. Insulin: its relationship to the central ner-vous system and the control of food intake and body weight. Am. J. Clin. Nutr.42:1063-1071.
2. Vanderweele, D. A., E. Haraczkiewicz, and T. B. Van Itallie. 1982. Elevatedinsulin and satiety in obese and normal weight rats. Appetite. 3:99-109.
3. Woods, S. C., E. C. Lotter, L. D. McKay, and D. Porte, Jr. 1979. Chronicintracerebroventricular infusion of insulin reduces food intake and body weightof baboons. Nature (Lond.). 282:503-505.
4. Pardridge, W. M. 1987. Receptor-mediated transport through the blood-brain barrier. Endocr. Rev. 7:314-330.
5. Bar, R. S., M. Boes, and A. Sandra. 1985. Vascular transport of insulin torat cardiac muscle. J. Clin. Invest. 81:1225-1233.
6. King, G. L., and S. M. Johnson. 1985. Receptor-mediated transport ofinsulin across endothelial cells. Science (Wash. DC). 277:1583-1586.
7. Frank, H. J. L., and W. M. Pardridge. 1983. Insulin binding to brainmicrovessels. Adv. Metab. Disord. 10:291-302.
8. Schwartz, M. W., A. J. Sipols, S. E. Kahn, D. F. Lattemann, G. J. Taborsky,Jr., R. N. Bergman, and D. Porte, Jr. 1990. Kinetics and specificity of insulinuptake from plasma into cerebrospinal fluid. Am. J. Physiol. 259 (Endocr. Me-tab. 22):E378-E383.
9. Duffy, K. R., and W. M. Pardridge. 1987. Blood-brain barrier transcytosisof insulin in developing rabbits. Brain Res. 420:32-38.
10. Schwartz, M. W., R. N. Bergman, S. E. Kahn, G. J. Taborsky, Jr., L. D.Fisher, A. J. Sipols, S. C. Woods, G. M. Steil, and D. Porte, Jr. 1991. Evidence forentry of plasma insulin into cerebrospinal fluid through an intermediate com-partment in dogs. J. Clin. Invest. 88:1272-1281.
11. Morgan, C. R., and A. Lazarow. 1963. Immunoassay of insulin: twoantibody system. Diabetes. 12:115-126.
12. Baskin, D. G., S. C. Woods, D. B. West, M. van Houten, B. I. Posner,D. M. Dorsa, and D. Porte, Jr. 1983. Immunocytochemical detection of insulinin rat hypothalamus and its possible uptake from cerebrospinal fluid. Endocrinol-ogy. 113:1818-1825.
13. Berman, M., and M. F. Weiss. 1978. The SAAMManual. DHEWPubli-cation No. (NIH) 78-180. U.S. Government Printing Office, Washington, DC.
14. Gabel, R. A., and R. A. Roberts. 1980. Signals and Linear Systems. 2ndedition. John Wiley & Sons, Inc., NewYork.
15. Reed, D. J., and D. M. Woodbury. 1963. Kinetics of movement of iodide,
sucrose, inulin and radio-iodinated serum albumin in the central nervous systemand cerebrospinal fluid of the rat. J. Physiol. (Lond.). 169:816-850.
16. Davson, H. 1970. Physiology of the Cerebrospinal Fluid. Churchill, Lon-don.
17. Gilman, A. G., T. W. Rall, A. S. Nies, and P. Taylor, editors. 1990.Goodmanand Gilman's The Pharmacological Basis of Therapeutics. 8th edition.Pergamon Press, NewYork. 21.
18. Tallarida, R. J., and L. S. Jacob. 1979. The Dose-Response Relation inPharmacology. Springer-Verlag, Inc., NewYork. 50.
19. Frank, H. J. L., and W. M. Pardridge. 1981. A direct in vitro demonstra-tion of insulin binding to isolated brain microvessels. Diabetes. 30:757-761.
20. Haugaul, F., and V. Desmaizieres. 1986. The effect of insulin on glucoseuptake and metabolism in the human placenta. J. Clin. Endocrinol. Metab.62:803-807.
21. Keller, J. M., and J. S. Krohmer. 1968. Insulin transfer in the isolatedhuman placenta. Obstet. Gynecol. 32:77-80.
22. Yalow, R. S., and J. Eng. 1983. Insulin in the central nervous system. Adv.Metab. Disord. 10:341-354.
23. Schwartz, M. W., D. P. Figlewicz, D. G. Baskin, S. C. Woods, and D.Porte, Jr. 1992. Insulin in the brain: a hormonal regulator of energy balance.Endocr. Rev. 13:387-414.
24. Crone, C. 1965. Facilitated transfer of glucose from blood into braintissue. J. Physiol. (Lond.). 181:103-1 13.
25. Cutler, R. W. P. 1980. Neurochemical aspects of blood-brain cerebrospi-nal fluid barriers. In Neurobiology of Cerebrospinal Fluid. J. H. Wood, editor.Plenum Press, NewYork. p. 43.
26. Fishman, J. B., J. B. Rubin, J. V. Handrahan, J. R. Connor, and R. E.Fine. 1987. Receptor-mediated transcytosis of transferrin across the blood-brainbarrier. J. Neurosci. Res. 18:299-304.
27. Zlokovic, B. V., M. N. Lipovac, D. J. Begley, H. Davson, and L. Rakie.1987. Transport of leucine-enkephalin across the blood-brain barrier in the per-fused guinea pig brain. J. Neurochem. 49:310-315.
28. Wallum, B. J., G. J. Taborsky, Jr., D. Porte, Jr., D. P. Figlewicz, L.Jacobson, J. C. Beard, W. K. Ward, and D. Dorsa. 1987. Cerebrospinal fluidinsulin levels increase during intravenous insulin infusions in man. J. Clin. Endo-crinol. Metab. 64:190-194.
29. McGowan, M. K., K. M. Andrews, J. Kelly, and S. P. Grossman. 1990.Effects of chronic intrahypothalamic infusion of insulin on food intake and diur-nal meal patterning in the rat. Behav. Neurosci. 104:373-385.
30. Woods, S. C., and D. Porte, Jr. 1976. Insulin and the set-point regulationof body weight. In Hunger: Basic Mechanisms and Clinical Implications. D.Novin, G. A. Bray, and W. Wyrwichka, editors. Raven Press, New York. 273-280.
31. Woods, S. C., D. P. Figlewicz Lattemann, M. W. Schwartz, and D. Porte,Jr. 1990. A re-assessment of the regulation of adiposity and appetite by the braininsulin system. Int. J. Obesity. 14 (Suppl. 3):1063-1071.
32. Yang, Y. J., I. D. Hope, M. Ader, and R. N. Bergman. 1989. Insulintransport across capillaries is rate limiting for insulin action in dogs. J. Clin.Invest. 84:1620-28.
33. Ader, M., R. A. Poulin, Y. J. Yang, and R. N. Bergman. 1992. Dose-re-sponse relationship between lymph insulin and glucose uptake reveals enhancedinsulin sensitivity of peripheral tissues. Diabetes. 41:241-53.
34. D'Argenio, D. Z. 1981. Optimal sampling times for pharmacokineticexperiments. J. Pharmacokin. Biopharm. 9:739-756.
35. Manin, M., Y. Broer, M. Balage, W. Rostene, and J. Grizard. 1990. Meta-bolic clearance of insulin from the cerebrospinal fluid in the anesthetized rat.Peptides. 11:5-12.
36. Carson, E. R., C. Cobelli, and L. Finkelstein. 1983. The MathematicalModeling of Metabolic and Endocrine Systems. John Wiley & Sons, Inc., NewYork.
1830 Baura et aLe