schwannoma gene therapy by adeno-associated virus delivery ... · of the pore-forming protein...
TRANSCRIPT

Cancer Gene Therapyhttps://doi.org/10.1038/s41417-018-0077-3
ARTICLE
Schwannoma gene therapy by adeno-associated virus deliveryof the pore-forming protein Gasdermin-D
Sherif G. Ahmed1● Abdelanabi Ahmed1
● Mohamed Doha1 ● Gary J. Brenner1
Received: 7 September 2018 / Revised: 28 November 2018 / Accepted: 9 December 2018© Springer Nature America, Inc. 2019
AbstractSchwannomas are peripheral nerve sheath tumors associated with three genetically distinct disease entities, namely sporadicschwannoma, neurofibromatosis type-2, and schwannomatosis. Schwannomas are associated with severe disability and incases lead to death. The primary treatment is operative resection that itself can cause neurologic damage and is at timescontra-indicated due to tumor location. Given their homogenous Schwann-lineage cellular composition, schwannomas areappealing targets for gene therapy. In the present study, we have generated an adeno-associated serotype 1 virus (AAV1)-based vector delivering N-terminal of the pyroptotic gene Gasdermin-D; (GSDMDNterm) under the control of the Schwann-cell specific promoter, P0. we have demonstrated that AAV1-P0-GSDMDNterm injection into intra-sciatic schwannomasreduces the growth of these tumors and resolves tumor-associated pain without causing neurologic damage. This AAV1-P0-GSDMDNterm vector holds promise for clinical treatment of schwannomas via direct intra-tumoral injection.
Introduction
Schwannomas are slow growing tumors of Schwann cellorigin that develop along peripheral nerves [1]. In ~60% ofthe cases these lesions arise in children and young adultsand new tumors continue to develop in multiple locationsthroughout the patient’s life [2]. Schwannomas are asso-ciated with a variety of disabling neurological symptoms,both positive (gain of function) and negative (loss offunction). Pain, deafness, motor dysfunction includingparalysis and even death are among these symptoms [3]. Inaddition, while they initially present as benign and non-invasive tumors, they can then develop into malignantperipheral nerve sheath tumors [4]. Conventional treatmentof these tumors includes surgical resection and radio-therapy. Operative resection is not always possible due to
the risk of neurologic damage and radiotherapy is limited byrisk of malignant transformation in NF2-deficient schwan-nomas [5]. Clinical trials testing cancer chemotherapeutics,such as anti-angiogenic compounds and small molecules,are ongoing. However, the efficacy of the tested drugs hasbeen limited and transient [6].
The typically slow growth and mitotic index of schwan-nomas makes these tumors an appropriate target for genetherapy. Further, gene therapy via direct tumor injection isminimally invasive compared with operative resection.There are very few reported preclinical schwannoma genetherapy studies. Intra-tumoral injection of a herpes simplexvirus-1 (HSV-1) amplicon vector expressing caspase-1(interleukin-β-converting enzyme; ICE) under the Schwanncell-specific P0 promoter led to tumor regression in axenograft model in which a human schwannoma cell linewas implanted subcutaneously [7]. In further development ofthis strategy, we utilized adeno-associated viral vectorserotype-1 (AAV1) expressing a pro-apoptotic caspase-1gene under the control of the Schwann-lineage cell-specificpromoter, P0 to target intra-sciatic schwannomas in anotherxenograft model [8]. These studies together demonstratedthat the P0 promoter, which is selectively expressed inSchwann and schwann-lineage cells (including schwannomatumor cells), prevented transgene expression in neurons, andtumor treatment was not associated with any neural toxicity.Thus, gene therapy utilizing tissue-specific delivery of
* Gary J. [email protected]
1 Department of Anesthesiology, Critical Care, and Pain Medicine,Massachusetts General Hospital, Harvard Medical School,Boston, MA 02114, USA
Supplementary information The online version of this article (https://doi.org/10.1038/s41417-018-0077-3) contains supplementarymaterial, which is available to authorized users.
1234
5678
90();,:
1234567890();,:

transgene capable of selectively killing schwannoma tumorrepresents a promising treatment.
Gasdermin-D (GSDMD), a member of the gasderminprotein family, is an essential mediator of inflammasome-mediated pyroptosis in human and mouse cells [9–11].Inflammasome activation and assembly with caspase-1,caspase-11, caspase-4, and caspase-5 and apoptosis-associated speck-like protein containing a CARD (ASC)results in GSDMD cleavage and induction of pyroptoticcell death. The N-terminal fragment of GSDMD(GSDMDNterm) is sufficient to induce pyroptotic cell death[10, 11]. The mechanism of GSDMDNterm induction ofcell death is still not fully elucidated. Pyroptosis had longbeen demonstrated to involve the formation of a plasmamembrane pore, and subsequent osmotic rupture of the hostcell [12]. It is likely that GSDMDNterm either promotes theformation of this pore or itself has pore-formingactivity [13].
Here we report on the synthesis of a GSDMDNtermtransgene and its AAV1-mediated delivery as a pro-pyroptotic protein by AAV-1 under the Schwann cell spe-cific promoter, P0. Our data show that exogenous expres-sion of GSDMDNterm in cultured cells of thehuman schwannoma cell line, HEI-193, induces cell death.Intra-tumoral injection of AAV1-rP0-GSDMDNterm intointra-sciatic schwannomas in both xenograft human andsyngeneic mouse schwannoma models led to increasedtumor cell death and decreased tumor cell proliferation,which in turn was associated with a significant reduction intumor burden. There was no evidence that this treatmentwas associated with toxicity. This work demonstrates thatGSDMDNterm is a promising transgene for schwannomagene therapy.
Materials and methods
Animals
All animal experimentation was approved by and conductedunder the oversight of the Massachusetts General Hospital(MGH, Boston, MA) Institutional Animal Care and UseCommittee. Animals, nu/nu mice and FVB/N, were kept ona 12:12 light-to-dark cycle with ad libitum access to foodand water. Animal health was evaluated daily.
Cell culture and transfection
The HEI-193 human schwannoma cell line (from D.J. Lim,House Ear Institute, Los Angeles, CA) was established froma schwannoma in a patient with NF2, immortalized withhuman papillomavirus E6/E7 genes and grown as described[14, 15]. Mouse 08031-9 schwannoma cells (from Dr.
Marco Giovannini, Univ. of California, Los Angeles, CA)were grown as described [16]. cells were infected withlentivirus encoding Fluc and mCherry for bioluminescenceimaging and IHC, respectively [7]. HEI-193 cells weretransfected with lipofectamine 2000 (Invitrogen, Carlsbad,CA) per the manufacturer’s instructions. Skin fibroblastswere kindly provided by (Dr. Christopher Bragg laboratory,Massachusetts General Hospital, MA, USA). Briefly, skinbiopsies were performed by standard procedures [17]. Tis-sue explants were transferred to culture dishes containingfibroblast growth medium [Dulbecco's Modified EagleMedium (DMEM) with 20% fetal bovine serum (FBS) and1% penicillin/streptomycin] and placed under sterile cov-erslips to promote cell attachment. Cultures were main-tained in a humidified incubator at 37 °C and 5% CO2 withmedium exchanges every 3–4 days as fibroblasts migratedfrom the explant. Cultures were expanded by trypsinization,collecting cells by centrifugation, and resuspending ingrowth medium. Skin fibroblasts were transfected usingHuman Dermal Fibroblast Nucleofector Kit (Amaxa Bio-systems) per the manufacturer’s instructions. All tissueculture media and supplements were obtained from LifeTechnologies (Grand Island, NY, USA). All cell lines wereconfirmed to be free of contamination, including myco-plasma, prior to experimental use.
Cell viability and toxicity assay
The Cell Titer-Glo luminescent cell viability assay (Pro-mega, Madison, WI) was used to determine the number ofviable cells in culture based on ATP level quantification, anindicator of metabolically active cells. Measurement of theluminescence was conducted with a luminometer (DynexTechnologies, Chantilly, VA) per to the manufacturer'sinstructions. Cell death was measured by the LDH assayusing CytoTox 96 Non-Radioactive Cytotoxicity Assay kit(Promega, Madison, WI) per the manufacturer'sinstructions.
Cell death detection by Flow Cytometry
HEI-193 cells were transfected with pAAV-CBA-GSDMDNterm, pAAV-CBA-GFP or exposed to lipofecta-mine alone then stained with annexin V-conjugated tofluorescein isothiocyanate (FITC) and propidium iodide (PI)by using the annexin-V-FITC staining kit (Invitrogen, USA)per manufacturer's instructions. The cells were analyzed onthe FACSAria II platform (BD Bioscience).
AAV vectors
The AAV vector plasmid self-complementary (sc)AAV-CBA-GSDMDNterm was derived from scAAV-CBA-BGHpA
S. G. Ahmed et al.

(Dr. Miguel Sena-Esteves, University of MassachusettsMedical Center, Worcester, MA). The rat P0 promoter hasbeen previously described [18]. scAAV-rP0-GSDMDNtermand scAAV-rP0-null plasmids were derived from the plasmidscAAV-rP0-ICE [8]. These plasmids carry two AAV invertedterminal repeat (ITR) elements, one wild type and one inwhich the terminal resolution site was deleted, as described[19], generating a vector that is packaged as a self-com-plementary, double-stranded-like molecule. scAAV-rP0-nullwas generated by digesting the ICE transgene out of scAAV-rP0-ICE using EcoRV and then re-ligating the blunt site.scAAV-rP0-GSDMDNterm plasmid was generated by insert-ing PCR-amplified mouse GSDMDNterm (828 bp) intoscAAV-rP0-null that was linearized with ECORI.GSDMDNterm (828 bp) sequence was described previously[11] and synthetized by Genscript, USA. Resulting plasmidclones were screened by restriction digest and sequencing forcorrect GSDMDNterm orientation in relation to the P0 pro-moter. The primers that were used for GSDMDNtermamplification contain an ECORI restriction site, the sequencesare: Forward GATCAGAATTCGCCACCATGCCATCGGand reverse CATGTGAATTCTCAATCTGACAGGAGAC.The scAAV-CBA- GSDMDNterm plasmid was producedthrough inserting PCR-amplified mouse GSDMDNterm(828 bp) into scAAV-CBA-BGHpA that was linearized withHindIII and SpeI. The primers that were used forGSDMDNterm amplification contain HindIII and SpeIrestriction sites, the sequences are: ForwardGTCAAGCTTGCCACCAtGCCATCGG and reverse CAACTAGTTCAATCTGACAGGAGAC. All AAV vectors carrythe bovine growth hormone polyadenylation signal. Theidentity of all the cloned transgenes was confirmed bysequencing. AAV1 serotype (AAV1) vectors were producedby transient co-transfection of 293T cells by calcium phos-phate precipitation of vector plasmid (scAAV1-rP0-GSDMDNterm), adenoviral helper plasmid pAdΔF6, and aplasmid encoding the AAV1 cap gene (pXR1), as previouslydescribed [20]. Viral vectors titer (genome copies [GC]/ml)was determined by real time TaqMan PCR amplification withprimers and probe specific for the bovine growth hormonepolyadenylation signal. AAV1 vectors were stored at −80 °Cuntil use.
Generation of tumors and vector injection
Sciatic nerve schwannomas were generated by directinjection of HEI-193FC human or 08031-8FC mouseschwannoma cells into the left sciatic nerve of isoflurane-anesthetized mice, as described [21]. HEI-193FC or 08031-9FC cells were trypsinized and rinsed with cold PBS, and30,000 (or 10,000 for 08031-9) cells in a volume of 0.5 µl ofPBS were injected into the sciatic nerve of athymic nudemice (nu/nu, 5–7-week-old males; National Cancer Institute
[NCI]), or syngeneic FVB/N mice (5–7-week-old males;Charles River Laboratories), respectively, using a glassmicropipette and a gas-powered microinjector (IM-300;Narishige, Tokyo, Japan). Tumors were injected withAAV1-rP0-GSDMDNterm, AAV1-rP0-null or PBS (n= 8/group) either 2-weeks’ post HEI-193 tumor-cell implanta-tion or 4-days post 08031-9 tumor cell implantation, in eachcase with 2 × 1010 vector GC in 2 µl PBS, targeting thelocation of the sciatic nerve where tumor cells wereimplanted. Tumor growth was monitored blindly by in vivobioluminescence imaging at weekly intervals for HEI-193and twice a week for 08031-9, as described [21]. Briefly,mice were injected intraperitoneally with the Fluc substrated-luciferin, and, 10 min later, signal was acquired with ahigh efficiency IVIS Spectrum (Caliper Life Sciences,Hopkinton, MA)
Behavioral analysis
Behavioral testing utilized the von Frey method for pain/mechanical sensitivity, the Hargreaves plantar test for Pain/Thermal sensitivity, and by rotarod for gross motor func-tion, all according to published methods [22, 23]. Nu/numice were used for the behavioral experiments (n= 8/group). All animals were habituated to the behavioralapparatus for 1 week before testing for baseline. Threebaseline measurements on three separate days preceded thefirst injection. Mice were then tested the day after eachinjection and twice per week for 8 weeks. Mechanicalsensitivity of the hind paw was measured by determiningwithdrawal thresholds assessed with von Frey filamentsemployed to determine mechanical sensitivity of the plantarsurface of the hind paw, as described [8]. The 50%threshold for each paw withdrawal was calculated, as pre-viously described [24]. Thermal sensitivity of the hind pawwas measured by determining withdrawal latency assessedwith Hargreaves plantar test, as described [25]. A rotatingrod apparatus (Columbus Instruments, Columbus, OH) wasused to assess motor performance. Mice were placed on theelevated accelerating rod beginning at 1 rpm/min for twotrials per day twice per week. Animals’ fall latency (inseconds) was scored, as described [8].
Histological and immunohistochemical analysis
Six-weeks post tumor implantation and 4-weeks post virusinjection, animals were terminally anesthetized with iso-flurane (3%) and killed by decapitation. Sciatic nerves wereremoved and snap frozen for hematoxylin and eosin (H&E)and immunohistochemical staining, as described [26]. Thesciatic nerves were kept in OCT blocks at −80 °C. Sectionswere stained with H&E in accordance with routine proto-cols. Proliferation marker staining was performed using
Schwannoma gene therapy by adeno-associated virus delivery of the pore-forming protein Gasdermin-D

antibody against Ki67 (Abcam, Cambridge, MA). Briefly,sections were dried at room temperature (RT) overnight.They were fixed in pre-chilled acetone at 4 °C for 10 min,allowed to dry, and then immediately stained. Sections werewashed in PBS, blocked with serum-free protein block(Dako, Carpinteria, CA) and quenched for peroxidases indual endogenous enzyme block (Dako). Sections werewashed in PBS then incubated with anti-Ki67 primaryantibody for 1 h at room temperature, then washed in PBSand incubated with horseradish peroxidase-conjugated sec-ondary antibody for 30 min at room temperature. Sectionswere washed in PBS and incubated with DAB solution(Dako). Counterstaining was accomplished by dippingsections in ethanol and xylene before mounting in Cytoseal(Richard Allan Scientific, San Diego, CA) and covered withcover slips for microscopic visualization. In vivo apoptosisstaining was assessed using the TACS® 2 TdT-DAB in Situapoptosis Detection Kit (Trevigen, Gaithersburg, MD).Fifteen micrometre sections (cryostat) of fresh-frozennerves fixed with 3.7% formaldehyde after mounting onslides was stained and visualized with diaminobenzidine(DAB) under light microscope according to the manu-facturer’s instructions.
Data analysis
All data are presented as group averages ± standard error ofthe mean (SEM). All baseline behavioral values represent
average of all measurements obtained before injection. Datawere analyzed with GraphPad Prism and Microsoft Excel.Two-tailed t-test and repeated-measure analysis of variance(ANOVA) were utilized, as described [27]. p < 0.05 wasaccepted as significant.
Results
Over-expression of GSDMDNterm leads to death ofschwannoma cells in vitro
To assess the effect of exogenous expression ofGSDMDNterm on the schwannoma cell lines, we havegenerated an AAV plasmid construct to express the poreforming GSDMDNterm, the end-product and effector ofinflammasome pathway activation (Fig. 1a). We expressedGSDMDNterm under either constitutive promoter CBA orthe Schwann cell specific promoter, P0 (Fig. 1b); pAAV-CBA-GSDMDNterm was used to over-expressGSDMDNterm in cultured schwannoma cell lines, whilepAAV-rP0-GSDMDNterm was used for AAV1 packagingand production for the in vivo testing.
Transfection of human the schwannoma cell line,HEI-193, with pAAV-CBA-GSDMDNterm led to celldeath as indicated by increased LDH release anddecreased ATP-based viability (Fig. 2a). Microscopicexamination of the transfected cells confirmed the
Fig. 1 a a schematic diagram of the inflammasome pathway and acti-vation of Gasdermin-D. Inflammasome activation leads to assembly withASC and caspase-1 which results in cleavage of pro-caspase-1 to itsactive form. The activated caspase-1 causes IL-1β and IL-18 maturationand cleavage of gasdermin-D to its active N-terminal form which gen-erates pyroptotic cell death. b Schematic diagram of the self-
complementary AAV vector encoding the mouse GSDMDNterm geneunder the control of the CBA or rat P0 promoter. GSDMDNterm clea-vage site is at arrow, inflammatory caspases cleave mouse gasdermin-Dat site 276 between aspartic acid (D) and glycine (G). AAV-rP0-GSDMDNterm was used for AAV viral vectors production for in-vivoapplication. ITR inverted terminal repeats, PA polyadenylation signal
S. G. Ahmed et al.

pAAV-CBA-GSDMDNterm mediated killing (Fig. 2b).We further confirmed the GSDMDNterm induced celldeath after transfection of mouse schwannoma cell line,08031-9, with pAAV-CBA-GSDMDNterm (Supplemen-tary Figure 1). To examine whether GSDMDNterm over-expression induces cell death in non-tumor tissue, wetransfected normal human dermal fibroblasts with pAAV-CBA-GSDMDNterm and measured LDH release and ATP-based viability. We did not find a difference in eitherparameter in comparison to cells transfected with thecontrol plasmid, pAAV-CBA-GFP or non-transfected cells
(Fig. 2c). Furthermore, there was no altered morphology inthe cells transfected with GSDMDNterm construct com-pared to control (Fig. 2d). There was no significant dif-ference in transfection efficiency of HEI-193 and culturedhuman dermal fibroblasts (Supplementary Figure 2). Flowcytometric analysis demonstrated significantly more (*p <0.05) double Annexin V and propidium iodide (PI) staining(indicative of cell death) of GSDMDNterm-transfectedHEI-193 cells compared with GFP-transfected or lipo-fectamine exposed cells (Fig. 2e). GSDMDNterm over-expression resulted in ~45% dead cells, compared with 6%
Fig. 2 a Lactate dehydrogenase (LDH) release cytotoxicity and ATP-based viability of human HEI-193 were measured 24 or 72 hours,respectively, post pAAV-CBA-GSDMDNterm transfection, pAAV-CBA-GFP transfection or lipofectamine exposure alone.b GSDMDNterm over-expression causes altered morphological pat-tern and noticeable detached cells compared with the GFP transfectedcells and lipofectamine control. Western blot of GSDMDNterm pro-tein after transfection of HEI-193 demonstrated GSDMDNtermexpression (31 KDa) in pAAV-CBA-GSDMDNterm transfected cells,but not in pAAV-CBA-GFP transfected or lipofectamine exposedcells. c Lactate dehydrogenase (LDH) release cytotoxicity and ATP-
based viability of normal human dermal fibroblasts 24 or 72 hours,respectively, after GSDMDNterm over-expression by transfection didnot show any difference from the control GFP or non-transfected cells.d GSDMDNterm did not induce any morphological alteration orresulted in detached cells upon transfection. e Flow cytometric analysisdemonstrated significantly greater double Annexin V and propidiumiodide (PI) staining of GSDMDNterm-transfected HEI-193 cells,indicative of cell death, compared with GFP-transfected or lipofecta-mine exposed cells. GSDMDNterm over-expression resulted in ~45%dead cells, compared with 6% for CBA-GFP transfected and lipo-fectamine exposed cells (graph on right)
Schwannoma gene therapy by adeno-associated virus delivery of the pore-forming protein Gasdermin-D
for both CBA-GFP transfected and lipofectamine exposedcells.
Intra tumoral injection of AAV1-rP0-GSDMDNtermcontrols schwannoma growth
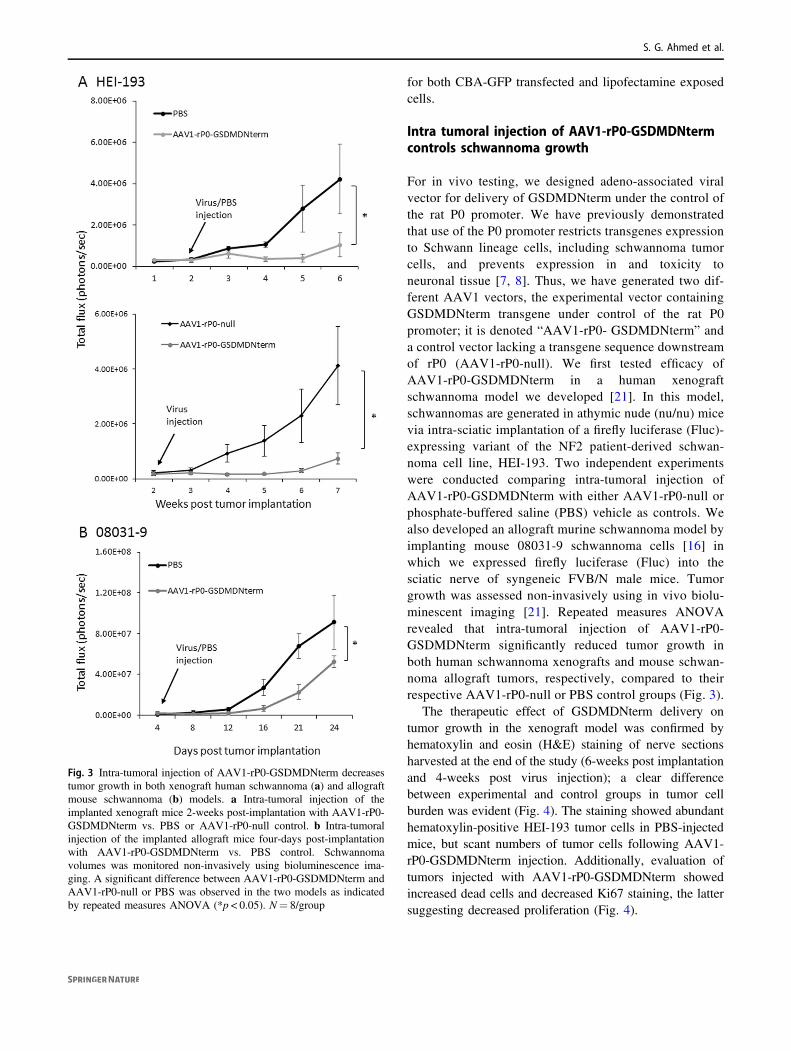
For in vivo testing, we designed adeno-associated viralvector for delivery of GSDMDNterm under the control ofthe rat P0 promoter. We have previously demonstratedthat use of the P0 promoter restricts transgenes expressionto Schwann lineage cells, including schwannoma tumorcells, and prevents expression in and toxicity toneuronal tissue [7, 8]. Thus, we have generated two dif-ferent AAV1 vectors, the experimental vector containingGSDMDNterm transgene under control of the rat P0promoter; it is denoted “AAV1-rP0- GSDMDNterm” anda control vector lacking a transgene sequence downstreamof rP0 (AAV1-rP0-null). We first tested efficacy ofAAV1-rP0-GSDMDNterm in a human xenograftschwannoma model we developed [21]. In this model,schwannomas are generated in athymic nude (nu/nu) micevia intra-sciatic implantation of a firefly luciferase (Fluc)-expressing variant of the NF2 patient-derived schwan-noma cell line, HEI-193. Two independent experimentswere conducted comparing intra-tumoral injection ofAAV1-rP0-GSDMDNterm with either AAV1-rP0-null orphosphate-buffered saline (PBS) vehicle as controls. Wealso developed an allograft murine schwannoma model byimplanting mouse 08031-9 schwannoma cells [16] inwhich we expressed firefly luciferase (Fluc) into thesciatic nerve of syngeneic FVB/N male mice. Tumorgrowth was assessed non-invasively using in vivo biolu-minescent imaging [21]. Repeated measures ANOVArevealed that intra-tumoral injection of AAV1-rP0-GSDMDNterm significantly reduced tumor growth inboth human schwannoma xenografts and mouse schwan-noma allograft tumors, respectively, compared to theirrespective AAV1-rP0-null or PBS control groups (Fig. 3).
The therapeutic effect of GSDMDNterm delivery ontumor growth in the xenograft model was confirmed byhematoxylin and eosin (H&E) staining of nerve sectionsharvested at the end of the study (6-weeks post implantationand 4-weeks post virus injection); a clear differencebetween experimental and control groups in tumor cellburden was evident (Fig. 4). The staining showed abundanthematoxylin-positive HEI-193 tumor cells in PBS-injectedmice, but scant numbers of tumor cells following AAV1-rP0-GSDMDNterm injection. Additionally, evaluation oftumors injected with AAV1-rP0-GSDMDNterm showedincreased dead cells and decreased Ki67 staining, the lattersuggesting decreased proliferation (Fig. 4).
Fig. 3 Intra-tumoral injection of AAV1-rP0-GSDMDNterm decreasestumor growth in both xenograft human schwannoma (a) and allograftmouse schwannoma (b) models. a Intra-tumoral injection of theimplanted xenograft mice 2-weeks post-implantation with AAV1-rP0-GSDMDNterm vs. PBS or AAV1-rP0-null control. b Intra-tumoralinjection of the implanted allograft mice four-days post-implantationwith AAV1-rP0-GSDMDNterm vs. PBS control. Schwannomavolumes was monitored non-invasively using bioluminescence ima-ging. A significant difference between AAV1-rP0-GSDMDNterm andAAV1-rP0-null or PBS was observed in the two models as indicatedby repeated measures ANOVA (*p < 0.05). N= 8/group
S. G. Ahmed et al.

Intra tumoral injection of AAV1-rP0-GSDMDNtermdoes not cause neuronal toxicity
Delivery of pore-forming proteins such as GSDMDNterminto schwannoma tumor may carry the risk of neurotoxicity.To investigate whether the GSDMDNterm gene therapy isassociated with toxicity to injected nerves, we have con-ducted behavioral tests. Three behavioral tests were per-formed. Thermal sensitivity was assessed using Hargreaves’Method in which an infrared heat source stimulates the hindpaw ipsilateral to tumor; and withdrawal latency is theprimary dependent measure (Fig. 5a) [25]. Mechanicalsensitivity – which can detect allodynia or decreased sen-sation – was tested using von Frey filaments to establishwithdrawal threshold of the hind paw ipsilateral to thetumor-containing sciatic nerve (Fig. 5b) [22]. Finally, grossmotor performance was assayed by the accelerating rotarodtest as a measure of neurotoxicity, (Fig. 5c) [23].
Intra-tumoral AAV1-rP0-GSDMDNterm inhibited tumorgrown (Fig. 3a). Tumor growth itself lead to sensitization to
thermal stimulation (Fig. 5a, PBS control group) which isinterpreted as development of pain sensitivity in the tumorbearing limb. Interestingly, as indicated by repeated mea-sure ANOVA [F (1,120)= 65.3, p < 0.01] AAV1-rP0-GSDMDNterm treatment was associated with normal-ization of heat sensitization (Fig. 5a). Von Frey withdrawalthreshold and gross motor performance did not changefollowing treatment with AAV-rP0-GSDMDNterm treat-ment compared with PBS controls (Fig. 5b, c, respectively).Taken together, these behavioral data indicate lack oftoxicity of AAV-rP0-GSDMDNterm treatment in the HEI-193 xenograft schwannoma model.
Discussion
We have previously shown that over-expression of pro-caspase-1 can control experimental schwannomas using anAAV1-based gene therapy vector [8]. In the experimentsdescribed here, we report on the use of a novel transgene,
Fig. 4 Intra-tumoral injection of AAV1-rP0-GSDMDNterm into HEI-193 human schwannoma increases pyroptosis and reduces tumor-cellproliferation. a Tumors injected with AAV1-rP0- GSDMDNtermshowed an increased intra-tumoral pyroptosis (brown stained, top left)while few scant dead bodies were observed in PBS injected tumors(top right). Immunohistochemical staining for the proliferation markerKi67 was lower in HEI-193 tumors injected with AAV1-rP0-GSDMDNterm (middle left) when compared to the PBS injected
tumors (middle right). Hematoxylin and eosin (H&E) staining showedmore HEI-193 tumor cells in PBS-injected mice (bottom right) than inthe AAV1-rP0-GSDMDNterm injected ones (bottom left). b Quanti-fication of brown stained dead bodies and Ki67-positive cells showedsignificantly greater cell death and less proliferation in the tumorsinjected with AAV1-rP0- GSDMDNterm in comparison to tumorsinjected with PBS (*p < 0.05, Student’s t-test)
Schwannoma gene therapy by adeno-associated virus delivery of the pore-forming protein Gasdermin-D

specifically the N-terminal of gasdermin D (GSDMDterm)under the control of the schwann-lineage specific promotor,P0, in an AAV1-based schwannoma gene therapy; the vectoris denoted “AAV1-rP0-GSDMDNterm.” GSDMD, like cas-pase-1, constitutes a primary component of the inflammasomepyroptotic cell death pathway. To our knowledge, this is thefirst study to propose GSDMDNterm as a transgene that couldbe used in a schwannoma gene therapy.
GSDMD was identified as a substrate for the inflam-matory caspases 1/4/5/11 [11]. Pyroptotic cell death wasshown to be dependent upon Cleavage of GSDMD and therelease of the GSDMDNterm which produces pores in bothartificial liposomes and the plasma membrane of cells [28].
Our in vitro data show that over-expression ofGSDMDNterm in the human NF2 schwannoma cell line,HEI-193, induces cell death as measured by ATP-based via-bility, LDH release and PI/annexin flowcytometric analysis.In contrast, GSDMDNterm did not cause cell death whenover-expressed in normal human dermal fibroblasts. How-ever, it is unclear why GSDMDNterm was not toxic to thenormal dermal fibroblasts. These observations suggest theselective killing of schwannoma tumor cells compared tohealthy cells following exogenous over-expression ofGSDMDNterm. Thus, we have generated pre-clinical datasupporting a potentially effective gene therapy strategy for thetreatment of schwannomas. Our therapeutic approach utilizesintra-tumoral injection of an AAV1 vector for delivery of aGSDMDNterm transgene. Tissue specificity and hence neu-ronal protection is generated using the Schwann-lineage cellspecific promoter, P0. Previous work from our lab demon-strated that there is essentially no expression of P0-controlledtransgenes in sensory neurons [8, 29]. Intra-tumoral injectionof AAV1-rP0-GSDMDNterm into mouse sciatic nervedemonstrated tumor cell death without signs of neuronalaxons toxicity. Behavioral testing, both sensory and motor,supported safety of AAV1-rP0-GSDMDNterm.
Although we have tested the therapeutic outcome of AAV1-rP0-GSDMDNterm using both xenograft and allograft mouseschwannoma models, preclinical assessment of this productrequires further studies in our immune-competent schwannomamouse model. This includes investigation of potential innateand adaptive immune responses associated with AAV1-rP0-GSDMDNterm treatment, and any effects of these responseson efficacy or toxicity.
In summary, we have identified GSDMDNterm as poten-tially therapeutic transgene with promise for schwannomagene therapy and generated a therapeutic strategy that utilizesa novel mechanism that does not interfere or overlap withcurrent schwannoma treatments. Translation of this AAV1-rP0-GSDMDNterm schwannoma gene therapy to clinicalapplication could reduce the need for surgical resection and itsassociated morbidity. Thus, AAV1-rP0-GSDMDNterm holds
Fig. 5 Effects of AAV1-rP0-GSDMDNterm or PBS injectionof implanted sciatic nerve HEI-193 schwannomas on painbehaviors, both thermal nociception (Hargreaves test, a) andmechanical sensitivity (Von Frey method, b), and on gross motorfunction (rotarod, c) were evaluated at 2- to 7-day intervalsfor ~7 weeks following tumor cell implantation. The Hargreaveswithdrawal latency (thermal nociception, a) return to values thatare not significantly different from baseline in AAV1-rP0-GSDMDNterm injected mice, while the nociceptive functionof PBS injected mice remains significantly lower than baselinevalues. Repeated measures ANOVA with post-hoc testing wasused to compare behaviors of the AAV1-rP0-GSDMDNterm andPBS injected mice to each other, as well as within each groupcomparing post virus injection at each time point to their respectivebaseline value. Results are represented as the mean ± SEM; n= 8mice per group. Asterisks indicate significant difference between theAAV1-rP0-GSDMDNterm injected and control group at the giventime point; *p < 0.05
S. G. Ahmed et al.

the promise of filling a major unmet clinical need for theschwannoma patients and their families.
Acknowledgements We thank Dr. Marco Giovannini, M.D., Ph.D.(UCLA Health) for the HEI-193 and 08031-9 cell lines, and TimothyHoule, Ph.D. (Department of Anesthesia, Critical Care and PainMedicine, MGH) for his assistance with statistical analysis.
Funding This work was supported by NIH/NINDS R01 NS081146,NIH/NINDS tR21 NS088013, and a charitable donation from Neu-rofibromatosis Northeast to the Rebecca Grasso Fund.
Compliance with ethical standards
Conflict of interest The authors declare that they have no conflict ofinterest.
Publisher’s note: Springer Nature remains neutral with regard tojurisdictional claims in published maps and institutional affiliations.
References
1. Evans DG. Neurofibromatosis type 2 (NF2): a clinical andmolecular review. Orphanet J Rare Dis. 2009;4:16.
2. Antinheimo J, Sankila R, Carpen O, Pukkala E, Sainio M, Jaas-kelainen J. Population-based analysis of sporadic and type 2neurofibromatosis-associated meningiomas and schwannomas.Neurology. 2000;54:71–6.
3. Lu-Emerson C, Plotkin SR. The neurofibromatoses. Part 2: NF2and schwannomatosis. Rev Neurol Dis. 2009;6:E81–6.
4. Carlson ML, Jacob JT, Habermann EB, Glasgow AE, Raghu-nathan A, Link MJ. Malignant peripheral nerve sheath tumors ofthe eighth cranial nerve arising without prior irradiation. J Neu-rosurg. 2016;125:1120–9.
5. Yamanaka R, Hayano A. Radiation-induced malignant peripheralnerve sheath tumors: a systematic review. World Neurosurg.2017;105:961–70 e8.
6. Wong HK, Lahdenranta J, Kamoun WS, Chan AW, McClatcheyAI, Plotkin SR, et al. Anti-vascular endothelial growth factortherapies as a novel therapeutic approach to treatingneurofibromatosis-related tumors. Cancer Res. 2010;70:3483–93.
7. Prabhakar S, Brenner GJ, Sung B, Messerli SM, Mao J, Sena-Esteves M, et al. Imaging and therapy of experimental schwannomasusing HSV amplicon vector-encoding apoptotic protein underSchwann cell promoter. Cancer Gene Ther. 2010;17:266–74.
8. Prabhakar S, Taherian M, Gianni D, Conlon TJ, Fulci G,Brockmann J, et al. Regression of schwannomas induced byadeno-associated virus-mediated delivery of caspase-1. Hum GeneTher. 2013;24:152–62.
9. He WT, Wan H, Hu L, Chen P, Wang X, Huang Z, et al. Gas-dermin D is an executor of pyroptosis and required for interleukin-1beta secretion. Cell Res. 2015;25:1285–98.
10. Kayagaki N, Stowe IB, Lee BL, O'Rourke K, Anderson K,Warming S, et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature. 2015;526:666–71.
11. Shi J, Zhao Y, Wang K, Shi X, Wang Y, Huang H, et al. Cleavageof GSDMD by inflammatory caspases determines pyroptotic celldeath. Nature. 2015;526:660–5.
12. Lamkanfi M. Emerging inflammasome effector mechanisms. NatRev Immunol. 2011;11:213–20.
13. Broz P. Immunology: caspase target drives pyroptosis. Nature.2015;526:642–3.
14. Hung G, Li X, Faudoa R, Xeu Z, Kluwe L, Rhim JS, et al.Establishment and characterization of a schwannoma cell linefrom a patient with neurofibromatosis 2. Int J Oncol.2002;20:475–82.
15. Lepont P, Stickney JT, Foster LA, Meng JJ, Hennigan RF, Ip W.Point mutation in the NF2 gene of HEI-193 human schwannomacells results in the expression of a merlin isoform with attenuatedgrowth suppressive activity. Mutat Res. 2008;637:142–51.
16. Tanaka K, Eskin A, Chareyre F, Jessen WJ, Manent J, Niwa-Kawakita M, et al. Therapeutic potential of HSP90 inhibition forneurofibromatosis type 2. Clin Cancer Res. 2013;19:3856–70.
17. Wray S, Self M, Consortium NPsDi, Consortium NPsDi, Con-sortium NAi, Lewis PA, et al. Creation of an open-access,mutation-defined fibroblast resource for neurological diseaseresearch. PLoS ONE 2012;7:e43099.
18. Brown AM, Lemke G. Multiple regulatory elements controltranscription of the peripheral myelin protein zero gene. J BiolChem. 1997;272:28939–47.
19. McCarty DM, Fu H, Monahan PE, Toulson CE, Naik P, SamulskiRJ. Adeno-associated virus terminal repeat (TR) mutant generatesself-complementary vectors to overcome the rate-limiting step totransduction in vivo. Gene Ther. 2003;10:2112–8.
20. Broekman ML, Comer LA, Hyman BT, Sena-Esteves M. Adeno-associated virus vectors serotyped with AAV8 capsid are moreefficient than AAV-1 or -2 serotypes for widespread gene deliveryto the neonatal mouse brain. Neuroscience. 2006;138:501–10.
21. Saydam O, Ozdener GB, Senol O, Mizrak A, Prabhakar S, Stemmer-Rachamimov AO, et al. A novel imaging-compatible sciatic nerveschwannoma model. J Neurosci Methods. 2011;195:75–7.
22. Agarwal N, Pacher P, Tegeder I, Amaya F, Constantin CE,Brenner GJ, et al. Cannabinoids mediate analgesia largely viaperipheral type 1 cannabinoid receptors in nociceptors. Nat Neu-rosci. 2007;10:870–9.
23. Kirschbaum KM, Hiemke C, Schmitt U. Rotarod impairment:catalepsy-like screening test for antipsychotic side effects. Int JNeurosci. 2009;119:1509–22.
24. Chen Q, Peto CA, Shelton GD, Mizisin A, Sawchenko PE,Schubert D. Loss of modifier of cell adhesion reveals a pathwayleading to axonal degeneration. J Neurosci. 2009;29:118–30.
25. Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new andsensitive method for measuring thermal nociception in cutaneoushyperalgesia. Pain. 1988;32:77–88.
26. Messerli SM, Tang Y, Giovannini M, Bronson R, Weissleder R,Breakefield XO. Detection of spontaneous schwannomas byMRI in a transgenic murine model of neurofibromatosis type 2.Neoplasia. 2002;4:501–9.
27. Harris JE, Sheean PM, Gleason PM, Bruemmer B, Boushey C.Publishing nutrition research: a review of multivariate techniques--part 2: analysis of variance. J Acad Nutr Diet. 2012;112:90–8.
28. Sborgi L, Ruhl S, Mulvihill E, Pipercevic J, Heilig R, Stahlberg H,et al. GSDMD membrane pore formation constitutes themechanism of pyroptotic cell death. EMBO J. 2016;35:1766–78.
29. Prabhakar S, Messerli SM, Stemmer-Rachamimov AO, Liu TC,Rabkin S, Martuza R, et al. Treatment of implantableNF2 schwannoma tumor models with oncolytic herpes simplexvirus G47Delta. Cancer Gene Ther. 2007;14:460–7.
Schwannoma gene therapy by adeno-associated virus delivery of the pore-forming protein Gasdermin-D