seasonal variation in food web composition and structure...
TRANSCRIPT

Seasonal Variation in Food Web Composition and Structure in
a Temperate Tidal Estuary
S. AKIN{ and K. O. WINEMILLER*
Section of Ecology, Evolutionary Biology and Systematics, Department of Wildlife and Fisheries Sciences,Texas A&M University, College Station, Texas 77843-2258
ABSTRACT: Seasonal variation in aquatic food web structure at Mad Island Marsh, Matagorda Bay, Texas, was examinedusing dietary information obtained from the analysis of gut contents from large samples of fish and crustacean specimens.Unique aspects of this study include the use of large samples of consumer gut contents (n 5 6,452), long-term sampling(bimonthly surveys over 18 mo), and standard methods of data collection and analysis facilitating comparisons with otheraquatic food webs. Dietary data were partitioned for analysis into warm (summer) and cold (winter) seasons. Most consumersfed low in the food web, with trophic levels ranging from about 2 to 3.5 during both summer and winter. Vegetative detrituswas more important in macroconsumer diets than live algae and macrophytes. Low trophic levels of consumers reflected theimportant role of abundant detritivores (e.g., striped mullet Mugil cephalus, Gulf menhaden Brevortia patronum, andmacroinvertebrates) in linking detritus to top predators via short food chains, a finding consistent with many other estuarinefood web studies. Despite changes in community composition and population size structure of certain species, most food webproperties revealed comparatively little seasonal variation. The summer food web had more nodes (86), more links (562),a higher density of links as indicated by connectance (0.08), and a slightly higher predator:prey ratio (0.51) compared to thewinter food web (75 nodes, 394 links, connectance 5 0.07, predator:prey ratio 5 0.47). Proportions of top (0.06–0.07),intermediate (0.75–0.76), and basal (0.19) species did not vary significantly between seasons, but mean trophic level washigher during summer. Addition of feeding links based on information from the literature increased connectance to 0.13during both seasons; other web parameters had values similar to those obtained for our directly estimated food webs.Seasonal variation in food web structure was influenced by changes in community composition (e.g., influxes of postlarvalestuarine-dependent marine fishes during winter), availability of resources (e.g., more submerged macrophytes andamphipods during summer), and size structure and ontogenetic diet shifts of dominant consumer taxa. Our findings suggestthat some basic properties of estuarine food web are resilient to seasonal changes in population and community structuresand food web architecture.
Introduction
The concept of the food web, a summary ofresource-consumer interactions in a community,enhances understanding of ecosystem structure andpopulation dynamics (Pimm et al. 1991; Winemillerand Polis 1996; de Ruiter et al. 2005; Winemillerand Layman 2005). Motivated in part by May’stheoretical research (1973) on the relationshipbetween community diversity and stability (see alsoMcCann 2000), a main theme of food web studieshas been empirical descriptions of different ecosys-tems in pursuit of general patterns or rules(Schoener 1989; Cohen et al. 1993). Analysis ofcollections of published food webs (e.g., Cohen1978; Pimm 1982; Briand 1983; Sugihara et al. 1989;Cohen et al. 1990) indicated that structure isnonrandom, possibly due to biological processes,such as predator-prey dynamics or energetic andmorphological constraints. These food webs tend to
have short food chains (Pimm 1982; Cohen et al.1986; Briand and Cohen 1987), constant fractionsof top, intermediate, and basal species (Briand andCohen 1984; Sugihara et al. 1989; Cohen et al.1990), an inverse relationship between web con-nectance and number of species (Briand andCohen 1984; Lawton 1989), infrequent omnivory(Pimm 1982; Pimm and Kitching 1987), anda constant predator : prey ratio (Briand and Cohen1984; Jeffries and Lawton 1985).
These early food web studies stimulated new areasof research, but the data from which generalizationswere derived have been strongly criticized (Paine1988; Winemiller 1990; Polis 1991; Hall andRaffaelli 1993; Yodzis 1993). Many of these general-izations were artifacts due to taxonomic incomplete-ness, inappropriate or inconsistent scales of resolu-tion, and other sources of bias. Recent years haveseen greater efforts to obtain better food webdescriptions based on data with higher resolutionand encompassing temporal and spatial variations.Analyses of these higher quality food webs haverejected many of the generalizations derived fromearly food web compilations (Winemiller 1989; Hall
{Current address: Faculty of Agriculture, Department ofFisheries, Gaziosmanpasa University, 60240 Tokat, Turkey.
* Corresponding author; tele: 979/862-4020; fax 979/845-4096;e-mail: [email protected]
Estuaries and Coasts Vol. 29, No. 4, p. 552–567 August 2006
552� 2006 Estuarine Research Federation

and Raffaelli 1991; Martinez 1991; Closs et al. 1993;Thompson and Townsend 2005). Leaving asideother shortcomings (lack of standard methods toobtain abundance and diet data, aggregation ofspecies), the early food webs did not take intoaccount temporal and spatial variations in commu-nity structure and predator-prey interactions. Mostecological communities occur within fluctuatingenvironments where primary production sourcesand the abundance, size structure, and trophicinteractions of populations vary accordingly (Wine-miller 1990; Polis et al. 1996). Aquatic animalschange their diets in association with ontogeny andseasonal dynamics of prey availability (Werner andGilliam 1984). Many food web comparisons werebased on snapshots of the community and failed toreveal variation over time and space.
A recent challenge in food web studies is tounderstand the effects of seasonal environmentalchange on food web properties. In temperateestuarine ecosystems, the composition and diversityof biota undergo strong interannual and seasonalchanges due to behavioral and demographic re-sponses to changes in temperature, salinity, andother environmental factors (Akin et al. 2003;Garcia et al. 2003). Seasonal influxes of estuarine-dependent marine fishes and shrimp alter commu-nity structure and the potential number of feedinginteractions. Warmer temperatures during summershould increase metabolism and rates of feeding byecotherms. These factors might be expected toincrease the density of feeding interactions in thefood web. The large detrital pool normally associ-ated with estuarine systems could influence trophicdynamics and increase stability of food web compo-nents (DeAngelis 1992). To our knowledge, noestuarine study has directly investigated seasonalvariation on food web structure and properties. Ourstudy examines seasonal variation in feeding inter-actions estimated from gut content analyses of largesamples from an estuarine fish and macroinverte-brate community on the Texas Gulf coast. Seasonaleffects on the food web were determined byanalyzing differences in web structure based onmonthly data aggregated as two periods, winter andsummer. A second objective was to determine majorsources of primary production that support the foodweb during each season. A third objective was tocompare characteristics of the Mad Island Marshweb with those of other webs reported in theliterature.
Materials and Methods
STUDY AREA
The study was conducted on Mad Island MarshPreserve (MIM) located on the northwestern coast
of the Gulf of Mexico, in Matagorda Bay, Texas(Fig. 1). Samples for stomach contents analysis weretaken from six sites located along a longitudinalestuarine gradient. Aquatic habitats in MIM spanphysiochemical gradients from freshwater in MadIsland Slough and diked wetlands, to an oligoha-line-mesohaline lake and salt marsh, to a polyhalinetidal bayou. The area is managed by the NatureConservancy of Texas, which has a restorationprogram that conducts controlled burns and aug-ments freshwater inflow to restore and enhancemarsh habitat. Dominant vegetation of the marshincludes saltwater cordgrass (Spartina alterniflora)and gulf cordgrass (Spartina spartinae). Submergedaquatic vegetation, such as widgeon grass (Ruppiamaritima) and skunkweed (Chara sp.), occurs sea-sonally with widgeon grass especially dominant atupper estuarine sites. Percent coverage of aquaticvegetation ranged from 0% to 90% and is highest inupper estuarine reaches during late spring throughearly fall (Akin et al. 2003). Water depth in theestuary ranged from 0.14 to 0.74 m. In most of theestuary and fringing marsh, substrate consisted ofa dense mud-sand mixture, covered by an approx-imately 5-cm layer of decomposing vegetation.Substrate near the intracoastal waterway consistedof mud, sand, and crushed oyster shell. Watertemperature ranged annually from 16.5uC to37.2uC, range of salinities was 3.5–29.4% withhigher values generally recorded from locationscloser to the bay, and dissolved oxygen concentra-tion varied from 2.66 to 13.9 mg l21. The measure-ments were taken every other month for 18 mofrom multiple locations at the site. Recordedenvironmental values were intended to reflectconditions during the period of sampling andreveal seasonal rather than diel variation (more
Fig. 1. Map of estuary at Mad Island Marsh Preserve on theTexas Gulf of Mexico coast showing locations of the survey sites.
Seasonal Variation in an Estuarine Food Web 553

detailed descriptions of environmental conditionsthroughout the study period appear in Akin et al.2003). Diurnal tidal flux at the site is low (0.1–0.25 m, with 0.5 m infrequently), and the largestvariation in water depth is associated with localprecipitation (Gelwick et al. 2001).
FIELD SAMPLING
Every 2 mo for 18 mo, a metal drop sampler wasused to quantify the density of benthic macroinver-tebrates, as well as the density of macrophytesenclosed within an area of 0.09 m2. During eachsurvey, two samples were taken at each of five sitesspaced approximately evenly along the longitudinalgradient of the estuary. The sampler was 1.0 m high,had no bottom, and was dropped by hand over anundisturbed area. All material inside the sampler(organisms, vegetation, and sediment to a depth of10 cm) was collected by sweeping the water columnand surface of the substrate with a fine-mesh dipnet. Samples were then placed into 50-l plastic bagsand preserved with 10% formalin. Entire sedimentsamples were sieved through a 0.25-mm mesh in thelaboratory. Macrophyte fragments were identifiedand weighed, and invertebrates were sorted, identi-fied to higher taxonomic/functional groups (inmost cases a coarse resolution was employed tomatch that which was feasible for specimensobtained during stomach contents analysis), andcounted.
Samples of fish and macroinvertebrate were takenevery 2 mo between March 1998 and August 1999along the longitudinal estuarine gradient (Fig. 1).Fishes and macroinvertebrates were captured usinga bag seine (6 3 1.2 m, 4.5 mm mesh, bag 1.2 31.2 m) and experimental gill nets (38 m, 5 panelsconsisted of 25, 38, 50, 64, and 76 mm mesh).Number of hauls and distances seined wererecorded in order to estimate relative abundanceas catch per unit effort of fish and macroinverte-brate taxa (additional details of collecting methodsappear in Akin et al. 2003). The duration of eachgill net set was recorded, and relative abundancewas estimated as number of individuals captured perhour. Captured fish and invertebrates were anesthe-tized in MS-222 then fixed in a 10% formalinsolution in the field. For each sample, fish weresorted, identified to species, measured (standardlength to nearest 0.1 mm), weighed (to nearest0.1 g), and counted in the laboratory.
Prior to seining each site, water depth wasmeasured with a tape measure, and temperature,dissolved oxygen, and salinity were measured witha Hyrdolab Datasonde or YSI 85. Aquatic macro-phyte abundance was estimated visually at each siteas the approximate percentage of a 100-m2 area withmacrophyte coverage (widgeon grass stands extend-
ing near or to the surface) when viewed from abovethe water surface.
STOMACH CONTENTS ANALYSIS
Stomach contents analysis was performed todocument food web structure at MIM usingmethods described in Winemiller (1990). Wheneverpossible at least 30 specimens of each fish speciesand 3–5 specimens of each macroinvertebratespecies (e.g., shrimp and crabs) from each siteduring each month were dissected for analysis.When a sample contained .30 specimens, individ-uals were selected for dissection so that size classeswere represented in proportions approximatingthose in the field sample. All available specimensof piscivores were dissected for stomach contents.All food items were removed from the anterior halfof the gut and examined under a dissectingmicroscope or compound microscope dependingon the size of the prey.
Most food items recovered from stomachs wererecorded as fairly broad functional and taxonomiccategories (Akin and Winemiller unpublished data),and this was largely due to limited capabilities toidentify fragmented or partially digested items withfiner taxonomic resolution. Taxonomic aggregationof invertebrate prey and plants (algae) reducedestimates of species richness and feeding linkswithin lower levels of the food web. The level ofscale and resolution for these food web componentsare quite similar to those employed by previousstudies with which results are compared. Smallinvertebrate prey, such as microcrustacea andinsects, were classified to taxonomic order. Fishesand macrocrustacea, such as grass shrimp (Palaemo-netes pugio), brown shrimp (Farfantepenaeus aztecus),white shrimp (Litopenaeus setiferus), blue crab (Call-linectes sapidus) and mud crabs (Neopenope sayi andEurypanopeus debressus), were classified to species.Some categories (Akin and Winemiller unpublisheddata) (e.g., fish parts, unidentified fish) weredeleted from the food web analyses that involvedtaxonomic nodes. For larger prey items (.0.1 ml),the entire prey item was blotted dry and its volumewas measured by water displacement in a graduatedcylinder. For volumes ,0.1 ml, items were placedon a glass slide and visually estimated by comparingits size with a water droplet of known volumeextracted from a graduated pipette.
Detrivores and algivores were handled differently.The entire gut was removed and a small sample ofits contents from the foregut was placed on a glassslide for examination under a compound micro-scope. The volume of the entire gut and its contentswas measured by water displacement. The relativeproportion (by area) of each identifiable item onthe slide was estimated, then multiplied by the
554 S. Akin and K. O. Winemiller

volume of the guts plus contents to yield percent oftotal gut volume.
FOOD WEB MATRICES
Because some species were abundant only atsome of the collection sites, we were not able toexamine spatial variation in food web structure.Monthly consumer-resource matrices were con-structed by pooling dietary data of each speciesfrom all six sites. Seasonal variation in fish andmacroinvertebrate assemblage structure is muchgreater than spatial variation within our study reach(Akin et al. 2003). Based on the seasonal pattern ofwater temperatures at MIM, we divided the year intothe warmer summer (April–September; monthlymean water temperature .29uC) and colder winter(October–March; monthly mean temperature,29uC) periods. In addition to these two seasonalfood webs, an annual food web was created bypooling dietary data from both seasons intoa cumulative resource matrix. Only those specieswith sample sizes having at least 6 specimens withstomachs containing food were included in re-source matrices, so these food webs representinteractions among common food web elements.For each consumer species, the reliability of dietestimates was evaluated using yield-effort curvesplotted as the cumulative percentage of feedinglinks in relation to the cumulative number ofspecimens examined (Cohen et al. 1993). Yield-effort curves for all common species (n . 100) andmost of the uncommon consumers (5 , n , 100)attained or approached an asymptote, indicatingthat our samples had revealed most of the feedinglinks.
Four of the fish species included as consumers inour matrix were not documented as prey within anystomachs analyzed during this study, yet they havebeen reported by other studies to be prey of certainspecies included in the MIM food web. Instead ofconsidering these potential prey species as toppredators (species having no predators), we as-signed trophic links (having the minimum observedvolumetric proportion for the consumer taxon)based on literature accounts. Information on pre-dators of gizzard shad (Dorosoma cepedianum), gulfpipefish (Syngnathus scovelli), bluntnose jack (Hemi-caranx amblyrhynchus), and bay whiff (Citharichthysspilopterus) was obtained from Pattillo et al. (1997).The seasonal and annual food webs comprised offield-estimated trophic links leading to fish andmacroinvertebrate consumers are referred to hereas original food webs.
To facilitate comparisons of our results withrecently published food webs, we compiled a secondversion of each of the three food webs that includedour dietary data for fishes and macroinvertebrates
(original webs) plus feeding links for the commonsmall invertebrates, such as worms and amphipods,inferred from literature reports (Pennak 1978;Merrit and Cummins 1996). These webs containingcrude approximations for diets of small inverte-brates are referred to as composite webs. Theliterature-based feeding links contained withincomposite webs were kept consistent betweenseasons.
ANALYSIS OF TROPHIC DATA
Since our aim was to look at the relative amountsof various categories consumed by the entirecommunity each season and annually, we standard-ized dietary data relative to sampling effort. Fishesand macroinvertebrates were collected using seineand gill nets. Since seine samples yielded moreindividuals than gill net samples, each species’dietary data were standardized in relation tosampling effort using the following procedure.Since striped mullet (Mugil cephalus) were collectedeffectively by both sampling gears, this species’biomass data were used to derive a crude conversionfactor for seine and gill net efforts. These calcula-tions revealed that 100 m of seining effort approx-imated 0.2 h of gill netting for mullet. For eachspecies, we estimated its overall relative abundance(Aj) based on both seine and gill net captures.Although admittedly crude, this method permits thecombination abundance data for both capturemethods, each of which has its own bias in termsof vulnerability to capture of certain taxa. Theconversion factor only applied to species that werecaptured by both methods.
The average volume of each prey item consumedby a given predator species (Vij) was calculated bydividing its total volume taken from stomachs (VTij)by the number of specimens dissected (Nj) thenmultiplying this ratio by the predator’s standardizedabundance (Aj) as defined above. In order todetermine proportional volumetric consumptionof individual food items (pij), the total volume ofeach prey category consumed by a given species(VTij) was divided by the total volume of foodconsumed by that species (VTj).
Food resource diversity (Simpson 1949) withinthe entire food web was calculated using Simpson’sdiversity index formula:
D ~1Pp2
i
where pi is the proportion of prey item i consumedby the entire community of fishes and macrocrus-tacea in a season. Levin’s standardized index wasused to calculate diet breadth of individual con-sumer species (Krebs 1999).
Seasonal Variation in an Estuarine Food Web 555

Dietary overlap (similarity) was calculated usingPianka’s (1973) symmetrical measure of nicheoverlap:
hjk ~
Pni ~ 1
pijpikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPni ~ 1
pij2Pn
i ~ 1pik
2
s
where hjk measures niche overlap between species jand k, pij is the volumetric proportion of resource iused by species j, pik is the volumetric proportion ofresource i used by species k, and n is the totalnumber of resource states. Dietary overlap wascalculated based on seasonal matrices from whichunidentified prey items and terrestrial insects wereremoved. Calculations were performed for thosesamples that had at least 6 stomachs with food.Dietary overlap also was calculated for conspecificsduring two different seasons as means to assessseasonal diet shifts of individual species.
Trophic levels of fishes and macrocrustacea werecalculated for each season using the formulapresented in Adams et al. (1983):
Ti ~ 1:0 zXn
j ~ 1
Tj pij
� �
where Tj is the trophic level of prey species j, and pij
is the fraction of the consumed food (volume) ofspecies i consisting of prey species j. Becauseprimary producers were coded as Tj 5 1, primaryconsumers (feeding on only plant material) were Tj
5 2.0, and a consumer eating exactly half plant andhalf herbivore tissue was Tj 5 2.5. Tj values forherbivorous species range from 2 to 2.5, those foromnivores ranged from 2.5 to 3.0, those for primarycarnivores ranged from 3 to 3.5, and tertiarycarnivores had Tj . 3.5 (Winemiller 1990; Halland Raffaelli 1993).
In order to evaluate the importance of prey itemsand primary production sources for summer andwinter communities, two separate seasonal principalcomponents analyses (PCA) were performed ondietary data, from which sand and unidentifieditems were removed (independent variables), thatwere summarized for each species (observations).The matrices for PCA were manipulated accordingto the method reported by de Crespin de Billy et al.(2000).
Seasonal variation in diet breadth and trophiclevel were tested using analysis of variance. Prior toanalysis, all variables were tested for normality(Kolmogorov-Smirnov) and equality of variances(Cochran’s test). All variables satisfied both criteria.Statistical tests, including tests of bivariate relation-
ships (Pearson’s correlation) and PCA, were per-formed using MINITAB version 13.1.
FOOD WEB STATISTICS
The number of species in the web (S) was thenumber of taxonomic species (fishes and macro-invertebrates) plus aggregated trophospecies, suchas diatoms and calanoid copepods, that were prey ofconsumers in original webs. The number ofpredator-prey interactions (L) was the number ofnon-zero elements in the community resourcematrix. Connectance, a measure of system complex-ity or the degree to which species interact in theweb, was calculated in three ways. Trophic con-nectance (Warren 1989) is the number of linksdivided by the number of possible links, defined asCT 5 L/(S(S 2 1)). Lower connectance (Pimm etal. 1991) was defined as CL 5 2L/(S(S 2 1)). Thisformula takes into account the fact that S(S 2 1) isthe correct estimate of possible links only whencannibalism is excluded. Directed connectance(Martinez 1991) was calculated as CD 5 L/S2, andtakes into account both cannibalism and mutualpredation between species.
The number of basal species (autotrophs), in-termediate species (taxa that eat other species andare themselves preyed upon), and top species(consumer taxa not fed upon by any of the otherspecies included in the web) were determined andconverted to proportions of the total species in theweb. The ratio of the number of predator taxadivided by the number of prey taxa (Jefferies andLawton 1985) was calculated for each web. Theincidence of omnivory was estimated as the fractionspecies with trophic levels between 2.5 and 3.0.
Results
SPECIES ABUNDANCE AND COMPOSITION
A total of 36,773 fishes from 40 species and 19families was collected with the seine during summer(Akin and Winemiller unpublished data). The twomost abundant species, comprising 85% of the totalseine samples, were Gulf menhaden (Brevortiapatronus) (40.7%) and bay anchovy (Anchoa mitchilli)(44.2%). Other common species, in order ofabundance, were pinfish (Lagodon rhomboides), tide-water silverside (Menidia peninsulae), striped mullet,and inland silverside (Menidia beryllina). A markeddecline in the abundance of young Atlantic croaker(Micropogonias undulatus) and spot (Leiostomusxanthurus), species that recruit into the estuary aspostlarvae during late winter, was apparent over thesummer months. A total of 32,637 macroinverte-brates representing 8 species was collected with theseine during summer (Akin and Winemiller un-published data). Of these species, grass shrimp was
556 S. Akin and K. O. Winemiller
the most abundant (85.2%), followed by brownshrimp (11.2%). A total of 12,762 fishes represent-ing 27 species and 14 families was collected in theseine during winter (Akin and Winemiller unpub-lished data). Gulf menhaden, Atlantic croaker, bayanchovy, and spot comprised 91.4% of individualsin the seine. Seine samples yielded a total of 22,592macroinvertebrates from 8 species during winter.Grass shrimp and white shrimp comprised morethan 98.5% of individuals in seine samples.
A total of 581 individual fishes from 17 speciesand 10 families was captured in gill nets duringsummer (Akin and Winemiller unpublished data).An increase in the relative abundance of stripedmullet (38.2%) and hardhead catfish (Arius felis;11.2%) and a pronounced decline in the relativeabundance of gizzard shad (from 35.8% to 19.45%)were the major differences in summer gill netcapture data compared to winter. Winter gill netsamples yielded a total of 463 individual fishes from18 species and 10 families (Akin and Winemillerunpublished data). Striped mullet, gizzard shad,and red drum (Sciaenops ocellatus) were the mostabundant species during winter. Hardhead catfish,black drum (Pogonias cromis), and alligator gar(Atractosteus spatula) also were common in wintergill net samples.
A summary of benthic fauna and infauna insediment core samples appears in Table 1.
Total abundance of benthic organisms variedseasonally, with the summer total almost twice thatof winter. Annelid worms, gastropods, chironomidlarvae, and mysid shrimp made up more than 85%of the total benthic sample during winter, butduring summer amphipods and annelid wormsalone made up over 80% of the total sample.Abundances of chironomid larvae and mysid shrimpdeclined to just 1% and 4% during summer,respectively. Annually, the most abundant benthictaxa were amphipods, annelid worms, and chiron-omid larvae.
TROPHIC DATA
We dissected a total of 5,865 fish and 587macroinvertebrate specimens for stomach contentsanalysis (3,658 fish and 349 invertebrates forsummer; 2,207 fish and 238 macroinvertebratesfor winter). Based on diets standardized relative tosampling effort for these macroconsumers (seine,gill net), the total volume of food recovered fromconsumer stomachs during summer was 127 mlcompared to only 27 ml during winter. Resourcediversity was higher during summer (6.04) thanwinter (5.77). According to volumetric percentage,detritus was the most import food category (.50%by volume) for macroconsumers during bothseasons, with more detritus consumed during
summer (Fig. 2). Based on the estimated totalvolume of material consumed from standardizedcommunity samples, consumption of the detrituswas almost six times higher during summer thanwinter. Algae, collectively, were the second mostimportant food resource at MIM, with a greaterpercentage consumed by the community of macro-consumers during winter (Fig. 2) but a slightlyhigher absolute volume consumed during summer.The total volume of fishes consumed by thecommunity was almost three times higher duringsummer than winter. The high volume of con-sumed fish prey was largely due to fact that severalpiscivores, such as red drum, alligator gar, south-ern flounder (Paralichthys lethostigma), and spottedsea trout (Cynoscion nebulosus), that were moreabundant during summer (Akin and Winemillerunpublished data). Spotted sea trout fed heavilyon striped mullet during summer (81% of dietby volume) and consume mostly Gulf menha-den (35%) and grass shrimp (27.5%) duringwinter. Hardhead catfish fed mostly on mollusks
TABLE 1. Estimates of benthic organism densities fromcore samples.
SpeciesSummer(# m2)
Summer(%)
Winter(# m2)
Winter(%)
Spider 0.00 0.00 0.93 0.08Mayfly 0.00 0.00 0.93 0.08Amphipoda 1,051.85 49.89 38.89 3.37Mysid shrimp 28.70 1.36 150.00 13.02Bivalvia 21.30 1.01 0.93 0.08Chironomid larvae 77.78 3.69 201.39 17.48Other Diptera larvae 2.78 0.13 0.00 0.00Diptera adult 0.00 0.00 5.56 0.48Gastropoda 55.56 2.64 293.52 25.47Isopoda 0.00 0.00 1.39 0.12Other annelid worm 616.67 29.25 355.56 30.86Polychaete worm 90.74 4.30 42.59 3.70Myrophis punctatus 1.85 0.09 0.00 0.00Eurypanopeus debressus 0.93 0.04 9.26 0.80Palaemonetes pugio 123.15 5.84 35.19 3.05Farfantepenaeus aztecus 0.93 0.04 0.00 0.00Litopenaeus setiferus 0.00 0.00 1.39 0.12Callinectes sapidus 8.33 0.40 4.63 0.40Callinectes similis 1.85 0.09 0.00 0.00Anchoa mitchilli 0.00 0.00 0.93 0.08Citharichthys spilopterus 0.00 0.00 1.85 0.16Cyprinodon variegatus 8.33 0.40 0.00 0.00Gobionellus boleosoma 0.00 0.00 1.39 0.12Gobiosoma bosc 0.93 0.04 0.93 0.08Gobiosoma robustum 0.00 0.00 0.93 0.08Other Gobiidae 1.85 0.09 0.00 0.00Lagodon rhomboides 1.85 0.09 0.00 0.00Lucania parva 4.63 0.22 0.00 0.00Microgobius gulosus 0.00 0.00 0.93 0.08Paralichthys lethostigma 0.00 0.00 0.93 0.08Unidentified Sciaenidae 0.00 0.00 0.46 0.04Unidentified fish 8.33 0.40 1.85 0.16Total 2,108.33 100.00 1,152.32 100.00# m2 488.04 355.65
Seasonal Variation in an Estuarine Food Web 557

(42%) and blue crabs (37%) during summer, andpinfish (42.5%), white shrimp (41.5%), mud crabs(N. sayi, 17.5%), and blue crabs (16%) duringwinter.
Seasonal distributions of diet breadth values arepresented in Table 2. Mean diet breadth of bothsummer (0.12) and winter (0.15) consumers did notdiffer significantly (F1,31 5 0.91, p 5 0.35). Dietbreadths of some species revealed strong seasonalvariation. Bay anchovy had greater diet breadth insummer than winter. Mean trophic level was notsignificantly associated with diet breadth duringsummer (r 5 0.25, p 5 0.24) but was marginallysignificant for winter (r 5 0.38, p 5 0.049).Although the mean trophic level for summer(2.76) was higher than winter (2.64), this differencewas not statistically significant (F1,31 5 0.95, p 50.33). Associations between diet breadth andconsumer body length (standard length) duringboth seasons were negative; associations were notsignificantly different for summer (r 5 20.133, p 50.45) or winter (r 5 20.099, p 5 0.62). In general,diet breadths of juvenile fishes and invertebrateswere broader than those of large fishes. Dietbreadth of large predatory species (e.g., red drum
Fig. 2. Percentages of the total standardized volumes of the 8most important food resources used by the assemblage ofconsumer taxa at Mad Island Marsh during summer and winter.
TABLE 2. Total number of stomachs examined, diet breadth (B), proportional frequency of occurrence of detritus (Dfreq), and trophiclevel (TL) of Mad Island Marsh macroconsumers during summer and winter.
Summer Winter
N Stomachs B Dfreq TL N Stomachs B Dfreq TL
Adinia xenica 53 0.11 0.32 2.02 — — —Anchoa mitchilli 523 0.29 0.16 3.31 387 0.06 0.28 3.03Arius felis 62 0.20 0.03 3.35 16 0.47 0.00 3.20Atractosteus spatula 20 0.07 0.00 3.28 10 0.10 0.00 3.07Bairdiella chrysoura 189 0.13 0.00 3.21 9 0.16 0.00 3.10Brevoortia patronus 210 0.03 0.83 2.01 314 0.07 0.64 2.02Callinectes sapidus 124 0.15 0.08 2.75 79 0.26 0.36 2.41Callinectes similis 15 0.14 0.22 2.02 — — —Citharichthys spilopterus 49 0.07 0.00 3.25 — — —Cynoscion arenurius 8 0.39 0.00 3.14 — — —Cynoscion nebulosus 50 0.04 0.00 3.18 42 0.38 0.00 3.23Cyprinodon variegatus 123 0.04 0.33 2.01 17 0.06 0.69 2.00Dorosoma cepedianum 111 0.09 0.59 2.24 90 0.03 0.83 2.11Elops saurus 8 0.53 0.00 3.07 — — —Farfantepenaeus aztecus 82 0.15 0.42 2.36 35 0.34 0.36 2.37Fundulus confluentus 7 0.60 0.10 2.70 — — —Fundulus grandis 108 0.18 0.83 2.54 25 0.09 0.64 2.15Fundulus pulvereus 11 0.02 0.02 3.00 — — —Gobionellus boleosoma 43 0.42 0.31 2.48 32 0.17 0.52 2.23Gobiosoma bosc 36 0.06 0.01 3.01 129 0.10 0.16 2.85Gobisoma robustum — — — 10 0.52 0.25 2.75Hemicaranx amblyrhyncus 14 0.03 0.00 3.22 — — —Lagodon rhomboides 316 0.03 0.05 2.16 48 0.07 0.04 2.74Leiostomus xanthurus 265 0.17 0.00 3.21 273 0.13 0.00 2.99Litopenaeus setiferus 37 0.11 0.82 2.24 52 0.03 0.83 2.03Lucania parva 123 0.03 0.02 3.00 49 0.24 0.45 2.53Menidia beryllina 423 0.14 0.00 2.83 196 0.20 0.03 2.52Menidia peninsulae 70 0.46 0.06 2.89 — — —Microgobius gulosus 6 0.63 0.00 3.06 — — —Micropogonias undulatus 175 0.18 0.04 2.87 339 0.26 0.09 2.94Mugil cephalus 402 0.05 0.67 2.16 115 0.10 0.59 2.07Palaemonetes pugio 90 0.04 0.85 2.03 66 0.03 0.83 2.13Paralichthys lethostigma 35 0.21 0.00 3.14 10 0.06 0.00 3.40Poecilia latipinna 77 0.04 0.82 2.02 11 0.00 1.00 2.00Pogonias cromis 43 0.06 0.76 3.15 14 0.16 0.82 2.03Sciaenops ocellatus 62 0.04 0.00 3.53 33 0.11 0.00 3.27Syngnathus scovelli — — — 15 0.49 0.04 2.98
558 S. Akin and K. O. Winemiller
and alligator gar) ranged from 0.04 to 0.11seasonally, whereas the darter goby (Gobionellusboleosoma) ranged from 0.17 to 0.42, and the inlandsilverside ranged from 0.14 to 0.20. Sample size(number of stomachs examined) did not affect dietbreadth during summer (r 5 0.041, p 5 0.813) orwinter (r 5 0.247, p 5 0.215).
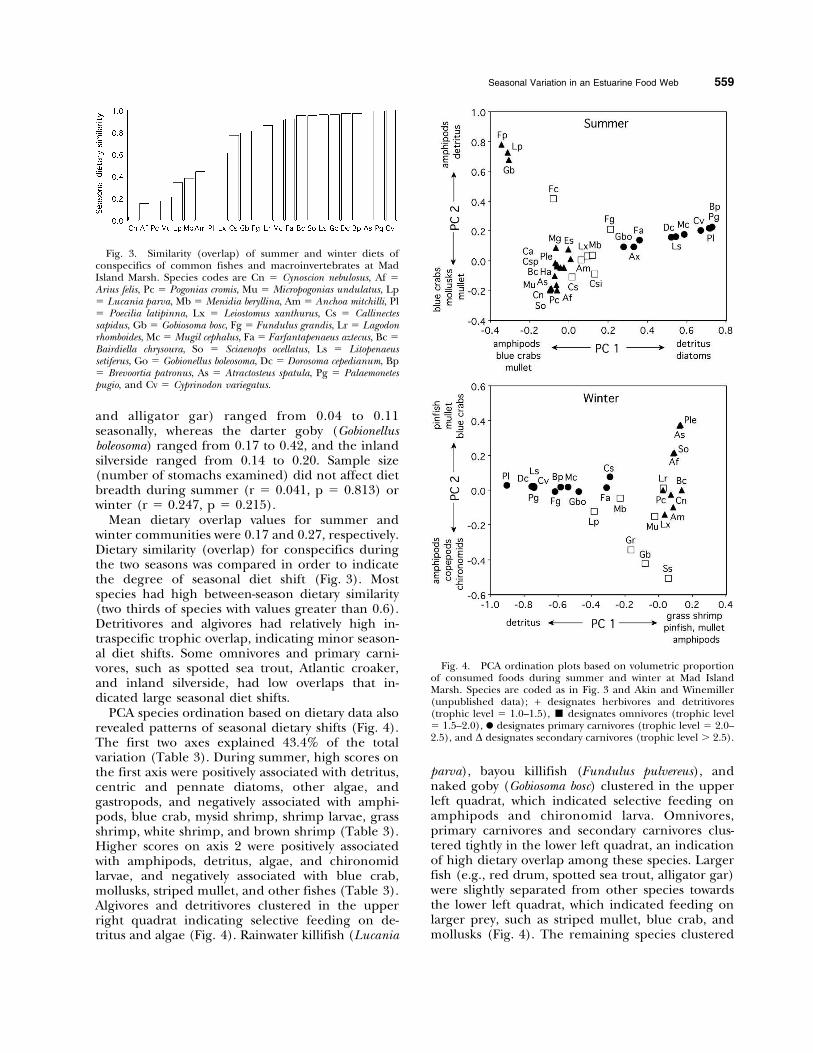
Mean dietary overlap values for summer andwinter communities were 0.17 and 0.27, respectively.Dietary similarity (overlap) for conspecifics duringthe two seasons was compared in order to indicatethe degree of seasonal diet shift (Fig. 3). Mostspecies had high between-season dietary similarity(two thirds of species with values greater than 0.6).Detritivores and algivores had relatively high in-traspecific trophic overlap, indicating minor season-al diet shifts. Some omnivores and primary carni-vores, such as spotted sea trout, Atlantic croaker,and inland silverside, had low overlaps that in-dicated large seasonal diet shifts.
PCA species ordination based on dietary data alsorevealed patterns of seasonal dietary shifts (Fig. 4).The first two axes explained 43.4% of the totalvariation (Table 3). During summer, high scores onthe first axis were positively associated with detritus,centric and pennate diatoms, other algae, andgastropods, and negatively associated with amphi-pods, blue crab, mysid shrimp, shrimp larvae, grassshrimp, white shrimp, and brown shrimp (Table 3).Higher scores on axis 2 were positively associatedwith amphipods, detritus, algae, and chironomidlarvae, and negatively associated with blue crab,mollusks, striped mullet, and other fishes (Table 3).Algivores and detritivores clustered in the upperright quadrat indicating selective feeding on de-tritus and algae (Fig. 4). Rainwater killifish (Lucania
parva), bayou killifish (Fundulus pulvereus), andnaked goby (Gobiosoma bosc) clustered in the upperleft quadrat, which indicated selective feeding onamphipods and chironomid larva. Omnivores,primary carnivores and secondary carnivores clus-tered tightly in the lower left quadrat, an indicationof high dietary overlap among these species. Largerfish (e.g., red drum, spotted sea trout, alligator gar)were slightly separated from other species towardsthe lower left quadrat, which indicated feeding onlarger prey, such as striped mullet, blue crab, andmollusks (Fig. 4). The remaining species clustered
Fig. 3. Similarity (overlap) of summer and winter diets ofconspecifics of common fishes and macroinvertebrates at MadIsland Marsh. Species codes are Cn 5 Cynoscion nebulosus, Af 5Arius felis, Pc 5 Pogonias cromis, Mu 5 Micropogonias undulatus, Lp5 Lucania parva, Mb 5 Menidia beryllina, Am 5 Anchoa mitchilli, Pl5 Poecilia latipinna, Lx 5 Leiostomus xanthurus, Cs 5 Callinectessapidus, Gb 5 Gobiosoma bosc, Fg 5 Fundulus grandis, Lr 5 Lagodonrhomboides, Mc 5 Mugil cephalus, Fa 5 Farfantapenaeus aztecus, Bc 5
Bairdiella chrysoura, So 5 Sciaenops ocellatus, Ls 5 Litopenaeussetiferus, Go 5 Gobionellus boleosoma, Dc 5 Dorosoma cepedianum, Bp5 Brevoortia patronus, As 5 Atractosteus spatula, Pg 5 Palaemonetespugio, and Cv 5 Cyprinodon variegatus.
Fig. 4. PCA ordination plots based on volumetric proportionof consumed foods during summer and winter at Mad IslandMarsh. Species are coded as in Fig. 3 and Akin and Winemiller(unpublished data); + designates herbivores and detritivores(trophic level 5 1.0–1.5), & designates omnivores (trophic level5 1.5–2.0), N designates primary carnivores (trophic level 5 2.0–2.5), and D designates secondary carnivores (trophic level . 2.5).
Seasonal Variation in an Estuarine Food Web 559
above these large fishes, which indicated consump-tion of mysid shrimp, juvenile menhaden, brownshrimp, white shrimp, and grass shrimp.
During winter, the first two axes explained 50.7%of the total variation (Table 3). High scores on thefirst PCA axis were associated with greater consump-tion of grass shrimp, mullet, pinfish, amphipods,blue crab, calanoid copepods, white shrimp, men-haden, and chironomid larvae, and less consump-
tion of detritus, gastropods, diatoms, and otheralgae (Fig. 4). The second PCA axis was associatedwith more pinfish, striped mullet, blue crab, whiteshrimp, and widgeon grass in the diet, and lessamphipods, calanoid copepods, chrinomoid larvae,and annelid worms (Table 3). Detrivores andalgivores (e.g., striped mullet, Gulf menhaden,gizzard shad, and shrimp) had negative scores onPCA1 and scores near 0 on PCA2. Omnivoresclustered on the lower right quadrat that wasinfluenced by consumption of amphipods andother benthic invertebrates (Fig. 4). Primary carni-vores that consumed shrimp larvae, grass shrimp,copepods, and other benthic invertebrates hadpositive scores on PCA1. Piscivores were located atthe upper right quadrat and consumed mostlystriped mullet, pinfish, and macroinvertebrates,such as blue crab and white and brown shrimp(Fig. 4).
FOOD WEB COMPOSITION
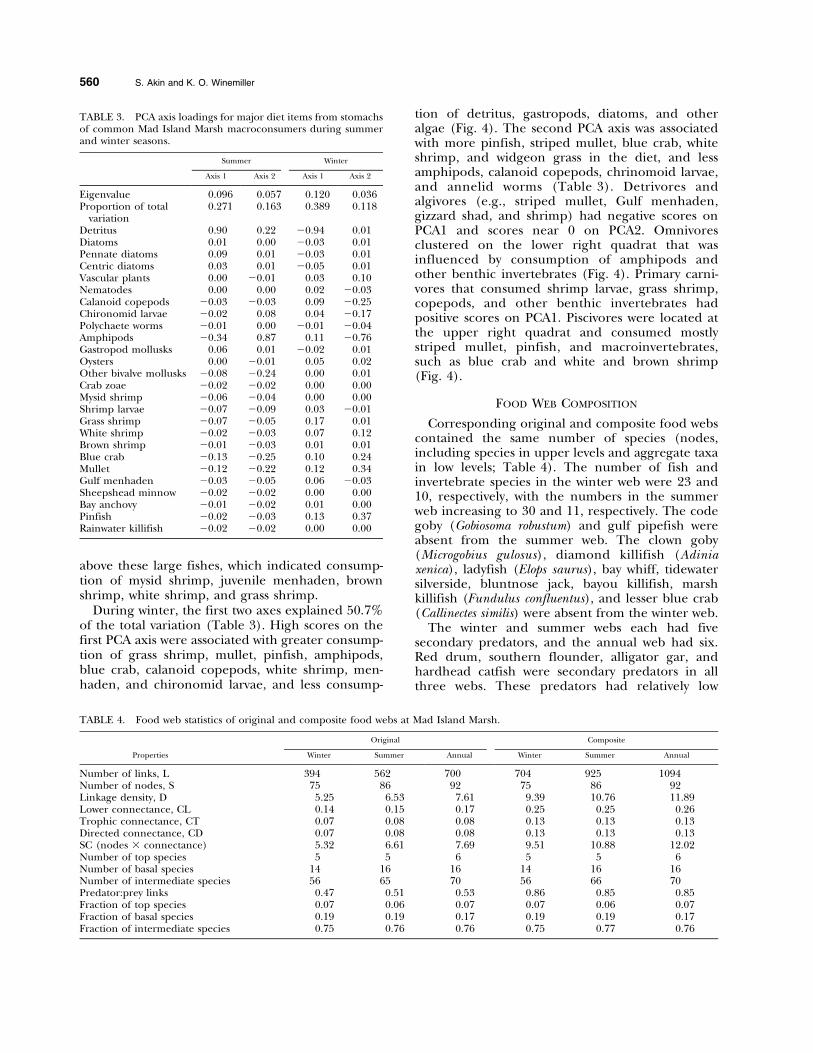
Corresponding original and composite food webscontained the same number of species (nodes,including species in upper levels and aggregate taxain low levels; Table 4). The number of fish andinvertebrate species in the winter web were 23 and10, respectively, with the numbers in the summerweb increasing to 30 and 11, respectively. The codegoby (Gobiosoma robustum) and gulf pipefish wereabsent from the summer web. The clown goby(Microgobius gulosus), diamond killifish (Adiniaxenica), ladyfish (Elops saurus), bay whiff, tidewatersilverside, bluntnose jack, bayou killifish, marshkillifish (Fundulus confluentus), and lesser blue crab(Callinectes similis) were absent from the winter web.
The winter and summer webs each had fivesecondary predators, and the annual web had six.Red drum, southern flounder, alligator gar, andhardhead catfish were secondary predators in allthree webs. These predators had relatively low
TABLE 3. PCA axis loadings for major diet items from stomachsof common Mad Island Marsh macroconsumers during summerand winter seasons.
Summer Winter
Axis 1 Axis 2 Axis 1 Axis 2
Eigenvalue 0.096 0.057 0.120 0.036Proportion of total
variation0.271 0.163 0.389 0.118
Detritus 0.90 0.22 20.94 0.01Diatoms 0.01 0.00 20.03 0.01Pennate diatoms 0.09 0.01 20.03 0.01Centric diatoms 0.03 0.01 20.05 0.01Vascular plants 0.00 20.01 0.03 0.10Nematodes 0.00 0.00 0.02 20.03Calanoid copepods 20.03 20.03 0.09 20.25Chironomid larvae 20.02 0.08 0.04 20.17Polychaete worms 20.01 0.00 20.01 20.04Amphipods 20.34 0.87 0.11 20.76Gastropod mollusks 0.06 0.01 20.02 0.01Oysters 0.00 20.01 0.05 0.02Other bivalve mollusks 20.08 20.24 0.00 0.01Crab zoae 20.02 20.02 0.00 0.00Mysid shrimp 20.06 20.04 0.00 0.00Shrimp larvae 20.07 20.09 0.03 20.01Grass shrimp 20.07 20.05 0.17 0.01White shrimp 20.02 20.03 0.07 0.12Brown shrimp 20.01 20.03 0.01 0.01Blue crab 20.13 20.25 0.10 0.24Mullet 20.12 20.22 0.12 0.34Gulf menhaden 20.03 20.05 0.06 20.03Sheepshead minnow 20.02 20.02 0.00 0.00Bay anchovy 20.01 20.02 0.01 0.00Pinfish 20.02 20.03 0.13 0.37Rainwater killifish 20.02 20.02 0.00 0.00
TABLE 4. Food web statistics of original and composite food webs at Mad Island Marsh.
Properties
Original Composite
Winter Summer Annual Winter Summer Annual
Number of links, L 394 562 700 704 925 1094Number of nodes, S 75 86 92 75 86 92Linkage density, D 5.25 6.53 7.61 9.39 10.76 11.89Lower connectance, CL 0.14 0.15 0.17 0.25 0.25 0.26Trophic connectance, CT 0.07 0.08 0.08 0.13 0.13 0.13Directed connectance, CD 0.07 0.08 0.08 0.13 0.13 0.13SC (nodes 3 connectance) 5.32 6.61 7.69 9.51 10.88 12.02Number of top species 5 5 6 5 5 6Number of basal species 14 16 16 14 16 16Number of intermediate species 56 65 70 56 66 70Predator:prey links 0.47 0.51 0.53 0.86 0.85 0.85Fraction of top species 0.07 0.06 0.07 0.07 0.06 0.07Fraction of basal species 0.19 0.19 0.17 0.19 0.19 0.17Fraction of intermediate species 0.75 0.76 0.76 0.75 0.77 0.76
560 S. Akin and K. O. Winemiller
trophic levels ranging from 3.1 to 3.5, whichreflected consumption of species low in the foodchain. The alligator gar’s diet consisted mostly ofstriped mullet that had a trophic level of 2.06 and2.16 for winter and summer, respectively. Red drumfed mainly on blue crab and striped mullet duringwinter and blue crab during summer.
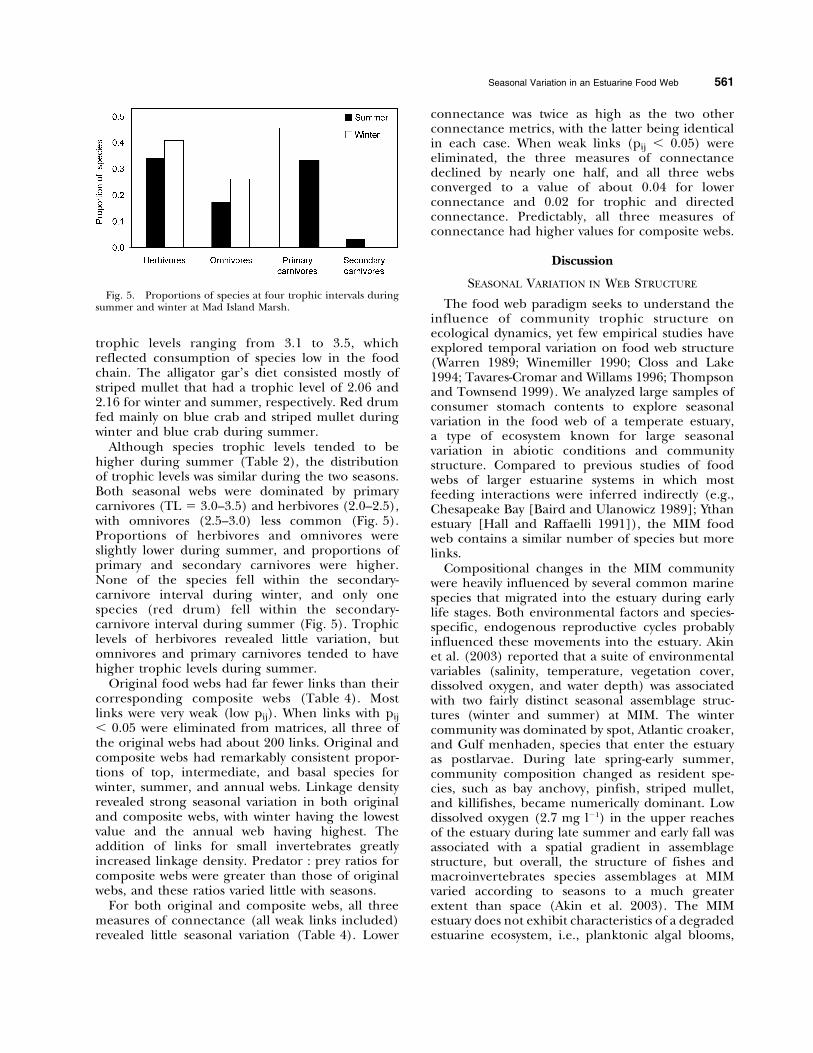
Although species trophic levels tended to behigher during summer (Table 2), the distributionof trophic levels was similar during the two seasons.Both seasonal webs were dominated by primarycarnivores (TL 5 3.0–3.5) and herbivores (2.0–2.5),with omnivores (2.5–3.0) less common (Fig. 5).Proportions of herbivores and omnivores wereslightly lower during summer, and proportions ofprimary and secondary carnivores were higher.None of the species fell within the secondary-carnivore interval during winter, and only onespecies (red drum) fell within the secondary-carnivore interval during summer (Fig. 5). Trophiclevels of herbivores revealed little variation, butomnivores and primary carnivores tended to havehigher trophic levels during summer.
Original food webs had far fewer links than theircorresponding composite webs (Table 4). Mostlinks were very weak (low pij). When links with pij
, 0.05 were eliminated from matrices, all three ofthe original webs had about 200 links. Original andcomposite webs had remarkably consistent propor-tions of top, intermediate, and basal species forwinter, summer, and annual webs. Linkage densityrevealed strong seasonal variation in both originaland composite webs, with winter having the lowestvalue and the annual web having highest. Theaddition of links for small invertebrates greatlyincreased linkage density. Predator : prey ratios forcomposite webs were greater than those of originalwebs, and these ratios varied little with seasons.
For both original and composite webs, all threemeasures of connectance (all weak links included)revealed little seasonal variation (Table 4). Lower
connectance was twice as high as the two otherconnectance metrics, with the latter being identicalin each case. When weak links (pij , 0.05) wereeliminated, the three measures of connectancedeclined by nearly one half, and all three websconverged to a value of about 0.04 for lowerconnectance and 0.02 for trophic and directedconnectance. Predictably, all three measures ofconnectance had higher values for composite webs.
Discussion
SEASONAL VARIATION IN WEB STRUCTURE
The food web paradigm seeks to understand theinfluence of community trophic structure onecological dynamics, yet few empirical studies haveexplored temporal variation on food web structure(Warren 1989; Winemiller 1990; Closs and Lake1994; Tavares-Cromar and Willams 1996; Thompsonand Townsend 1999). We analyzed large samples ofconsumer stomach contents to explore seasonalvariation in the food web of a temperate estuary,a type of ecosystem known for large seasonalvariation in abiotic conditions and communitystructure. Compared to previous studies of foodwebs of larger estuarine systems in which mostfeeding interactions were inferred indirectly (e.g.,Chesapeake Bay [Baird and Ulanowicz 1989]; Ythanestuary [Hall and Raffaelli 1991]), the MIM foodweb contains a similar number of species but morelinks.
Compositional changes in the MIM communitywere heavily influenced by several common marinespecies that migrated into the estuary during earlylife stages. Both environmental factors and species-specific, endogenous reproductive cycles probablyinfluenced these movements into the estuary. Akinet al. (2003) reported that a suite of environmentalvariables (salinity, temperature, vegetation cover,dissolved oxygen, and water depth) was associatedwith two fairly distinct seasonal assemblage struc-tures (winter and summer) at MIM. The wintercommunity was dominated by spot, Atlantic croaker,and Gulf menhaden, species that enter the estuaryas postlarvae. During late spring-early summer,community composition changed as resident spe-cies, such as bay anchovy, pinfish, striped mullet,and killifishes, became numerically dominant. Lowdissolved oxygen (2.7 mg l21) in the upper reachesof the estuary during late summer and early fall wasassociated with a spatial gradient in assemblagestructure, but overall, the structure of fishes andmacroinvertebrates species assemblages at MIMvaried according to seasons to a much greaterextent than space (Akin et al. 2003). The MIMestuary does not exhibit characteristics of a degradedestuarine ecosystem, i.e., planktonic algal blooms,
Fig. 5. Proportions of species at four trophic intervals duringsummer and winter at Mad Island Marsh.
Seasonal Variation in an Estuarine Food Web 561

low abundance of submerged aquatic plants andbenthic microalgae, depleted apex predators, anddepletion of oyster stocks (Baird et al. 2004). Evenduring late summer and early fall, apex predators,such as alligator gar and red drum, were capturedthroughout the system.
Web size increased from winter (75 nodes) tosummer (86 nodes), which was expected since thenumber of species in the estuary increased duringsummer when temperature and salinity were higher(Akin et al. 2003). The summer increase in speciesrichness was largely due to the addition of fishes atintermediate trophic levels, since numbers of lowerlevel consumers (e.g., zooplankton and benthicinvertebrates and top predators) varied little.Studies of lotic freshwater systems also have revealedstrong seasonal variation in food web structure inrelation to environmental conditions, such ashydrology and temperature, as well as life historycharacteristics (Winemiller 1990, 1996; Closs andLake 1994; Tavares-Cromar and Williams 1996;Peterson 1997). Shallow temperate estuaries arehighly dynamic ecosystems with hydrologic changesinfluenced by precipitation, wind, and tides. Hydro-logic variation was relatively low at MIM during ourstudy (Akin et al. 2003), and most of the seasonalvariation in community composition probably wascaused by combined effects of endogenous migra-tion cycles by estuarine-dependent species andassociated trophic dynamics (i.e., functional re-sponse, prey switching).
Seasonal dietary shifts were common in bothfishes and macroinvertebrates in MIM. In general,there was little indication of dietary shifts forherbivores, but seasonal changes in diet wereevident for omnivores and carnivores. The diet ofjuvenile spotted sea trout in winter consisted mostlyof zooplankton, amphipods, mysid shrimp, andsmall fishes, such as Gulf menhaden and bayanchovy; larger fish and invertebrates, includingstriped mullet, pinfish, white shrimp, and bluecrabs, dominated its diet during summer when itsmean standard length had increased to 102 mm.The same was true for Atlantic croaker with a dietshifting from copepods, amphipods, chironomidlarvae, and small fishes during winter when meanstandard length was 23.5 mm, to blue crabs andlarger polychaete worms during summer whenmean standard length was 43.7 mm.
In addition to ontogenetic dietary shifts, dietcomposition shifts seasonally in response to changesin food resource availability at MIM. More amphi-pods were consumed by rainwater killifish and othersmall fishes during summer when amphipodscomprised half of the total benthic fauna in coresamples. An increase in direct consumption ofvascular plants (mostly submerged widgeon grass)
also can be attributed to greater availability of thisresource during summer. The greater summerconsumption of mollusks was largely attributed toincreased abundance of black drum and hardheadcatfish, given that the abundance of mollusks incore samples was similar during winter and summer.An increase in piscivory during summer wasassociated with greater summertime abundances ofpiscivores, such as alligator gar.
An increase in trophic level of consumers and theproportion of primary carnivores during summerprovides additional evidence that diet shifts werecommon features of consumers in MIM. Duringwinter, many fishes were postlarvae of estuarine-dependent marine species that fed on zooplanktonand benthic invertebrates. By summer these fisheshad grown and became major food resources forpiscivores, which caused increases in both meantrophic level and the proportion of primarycarnivores in the web.
Seasonal variation in consumption of detritus wasapparent in the MIM web. Fishes and invertebratesin our samples were estimated to consume six timesmore detritus (volumetrically) during summer. Thisestimate even takes into account the observationthat the frequency of occurrence of detritus in thestomachs was slightly higher during winter (79%)than summer (72%). The increase in volume ofdetritus consumed during summer was influencedby a summertime increase in abundance of adultsize classes of striped mullet, a major detritivore incoastal ecosystems. Greater stomach fullness alsocould reflect greater energy demands of fishesduring warmer summer months (Baird et al. 2004).
SOURCES OF PRIMARY PRODUCTION AND DETRITUS
Benthic microalgae and macroalgae, phytoplank-ton, and vascular plants are sources of primaryproduction for estuarine food webs. Several studieshave concluded that vascular plants and plantdetritus are the principal basal resource supportingfish production in coastal systems (Darnell 1961;Canuel et al. 1995; Currin et al. 1995; Creach et al.1997). Other research has revealed greater assimi-lation of algal production by estuarine consumers(Peterson et al. 1985; Page 1997; Stribling andCornwell 1997; Riera et al. 1999; Currin et al. 2003).Studies using stable isotope techniques have re-vealed that carbon derived from C4 grasses, such assaltwater cordgrass, is assimilated in variable propor-tions into elements of marsh food webs (Haines1977; Currin et al. 1995; Kwak and Zedler 1997).Widgeon grass has been inferred to be a significantcarbon source for some consumers in coastal foodwebs (Harrigan et al. 1989). Evidence from stomachcontents analysis and sediment samples suggeststhat widgeon grass is a significant production source
562 S. Akin and K. O. Winemiller

supporting fishes and invertebrates in MIM. Inaddition to its role as a physical refuge for fishes,widgeon grass (both living and dead tissue) serves asfood and habitat for benthic invertebrates that inturn are food resources for almost all juvenile fishesin the system. Availability of these invertebratesincreased greatly during summer.
Consistent with other estuarine studies, detritus(originating from either algae or macrophytes) wasthe most important food source for the MIMmacrofauna. Microscopic examination of detritusrecovered from stomachs indicated mostly fineparticulate matter of unknown origin. It may beassumed that this material contains variable frac-tions of microbial and fungal biomass, althoughthese materials could not be quantified from ourmethod. Consumption of detritus was not limited todetritivores, but also consumed to a lesser extent byprimary carnivores and even secondary carnivores.Consumption of detritus by carnivores probably wasincidental with questionable nutritional contribu-tion. The most important foods of primary andsecondary consumers were benthic organisms thatfeed on detritus, either suspended or depositedparticles, and live aquatic vegetation. Herbivoresand detritivores comprised almost half of the totalfish and macroinvertebrate community during bothseasons. As reported by Odum and Heald (1975) foran Atlantic coast estuary, detritivorous fishes andinvertebrates link detritus to top predators via shortfood chains that presumably yield high ecologicalefficiency.
Based on dietary analysis, diatoms are anotherimportant production source supporting estuarineconsumers in MIM. Since stomach contents analysisonly quantifies ingested material, it is difficult toascertain whether or not algae, cordgrass, orwidgeon grass is the main source of carbon forconsumers. Phytoplankton are ubiquitous at MIM,and cordgrasses (Spartina alterniflora, S. patens, S.spartinae) fringe the entire study reach, with a thinlayer of benthic algae generally covering submergedstems and leaves. By providing estimates of assimi-lated (not just ingested) time-integrated diet of ananimal, stable isotope ratios of consumer tissues canbe used to infer primary production sources ofassimilated carbon (Peterson and Fry 1987). Arecent study comparing dietary and stable isotoperatios of MIM food web elements (Winemiller andAkin unpublished data) indicated that most con-sumer taxa probably are supported to some extentby algae, a finding consistent with other estuarinestudies (e.g., Currin et al. 1995; Moncreiff andSullivan 2001). Several species associated withshallow marginal areas of the estuary were estimatedto assimilate larger fractions of carbon from iso-topically heavier sources, such as cordgrass or
widgeon grass (Winemiller and Akin unpublisheddata).
COMPARISONS WITH OTHER FOOD WEBS
Due to inconsistent criteria that have been usedto compile food webs based on information in theliterature (Paine 1988) as well as the sensitivity ofweb properties to methods (Winemiller 1990; Polis1991; Hall and Raffaelli 1993), we consider compar-isons of webs from field studies using differentmethods and criteria to be tenuous. Food webs canbe compared if similar methods are used to samplehabitats, quantify feeding interactions, and desig-nate prey categories. We compared the MIMoriginal web with those from studies by Winemiller(1990), Peterson (1997), and Link (2002) that usedsimilar methods and followed most of the criteriaproposed by Cohen et al. (1993). For heuristicpurposes, we also compared several properties ofour composite webs with food webs created fromliterature data and that only characterize webtopology (e.g., Little Rock Lake [Martinez 1991]and topological webs compiled in ECOWEB [Cohenet al. 1993]).
The number of species in our original andcomposite annual food webs was greater than mostof the ECOWEB webs. Exceptions were Ythanestuary (S 5 134; Hall and Raffaelli 1991), LittleRock Lake (S 5 183), and El Verde rainforest,Puerto Rico (S 5 136; Regan et al. 1996). Criteriaused to compile these webs vary widely. Cohen et al.(1993, p. 254) suggested ‘‘a yield-effort curveshould be accompanied by an explicit report ofthe temporal and spatial sampling frame, thesampling plan, sampling gears, and methods forpreserving samples in the field.’’ Our MIM compos-ite annual web had more links (1,094) than theYthan estuary web (500) and all of the other websfrom ECOWEB (average 34, range 3–138), exceptfor the Little Rock Lake (2,366), El Verde (1,428),and marine coastal shelf webs (1,562; Link 2002).The Ythan estuary web included 1 mammal and 18bird species, and some data were gathered from theliterature. The greater number of links in the MIMweb seems to be derived from finer taxonomicresolution among primary producers and inverte-brates.
Winemiller’s (1990) tropical stream webs con-tained an average of 514 links when all interactionswere included, and 99 links when weak interactionswith pij , 0.05 were eliminated from the web. MIMoriginal webs contained an average of 552 links withall interactions included and 144 links when weaklinks (pij , 0.05) were excluded. A potentialexplanation is that these webs have similar rangesfor web size (number of nodes), and consumers inboth types of aquatic systems tend to have fairly
Seasonal Variation in an Estuarine Food Web 563

broad diets, with individual species consuming, onaverage, about 10–15% of available prey nodes.
Because it represents trophic complexity asa single value, connectance has received muchattention, particularly in the context of theoriespredicting effects of complexity on stability (e.g.,Pimm et al. 1991; Martinez 1992; Warren 1994).Early published food webs indicated that directedconnectance, CD, decreases hyperbolically withincreasing number of species, S (Fig. 6). Subse-quent analyses of published food webs rejected thispattern and showed that CD increases (Winemiller1989) or is constant (Warren 1989; Martinez 1992;Martinez et al. 1999) in relation to S. Montoya andSole (2002) concluded that CD is scale variant ina new collection of 12 food webs, with valuesranging from 0.02 to 0.24. CD of MIM webs onlyranged from 0.07 during winter to 0.08 for thecumulative annual web, even though the number ofspecies was 75 and 92 for these webs, respectively.Our findings agree with results obtained by Tavares-Cromar (1996) and Peterson (1997), but our CD
values were smaller than those obtained by Wine-miller (1990) for tropical streams. Differences in CD
could be due to more trophic links derived fromgreater sampling effort in the latter study.
Briand and Cohen’s (1984) link-species scalinglaw theorized that the average number of links perspecies in food webs (linkage density) is approxi-mately constant at 1.86, with this presumed asderived from dynamic constraints on system stabil-ity. Others have claimed that linkage densityincreases with increasing number of species in webs(Schoener 1989; Hall and Raffaelli 1991; Martinez1992; Havens 1993; Goldwasser and Roughgarden1993). Linkage density, D, was higher in the MIMannual web than the time-specific webs largelybecause both the numbers of species and links weregreater in the annual web. The average linkagedensity for three original MIM webs (6.4) was nearlythe same as the mean of 6.3 estimated for tropicalstream webs (Winemiller 1990), whereas linkagedensities of composite MIM webs (9.39–11.89) werenearly twice the values of the original MIM webs,and were much higher than the value reported forYthan estuary (4.1). Such patterns should beinterpreted with caution, especially given thatMartinez (1991) demonstrated the high sensitivityof linkage density to taxonomic aggregation.
The fractions of top species, basal, and interme-diate species are claimed to be constant at the ratioof 0.29, 0.19, and 0.52, respectively (Briand andCohen 1984; Cohen et al. 1990), but these ratioshave been criticized (Winemiller 1990; Polis 1991;Closs et al. 1993; Havens 1993). To define a toppredator is not a simple matter, especially when oneconsiders that even the fiercest predators suffersome level of predation (Winemiller 1990). In theMIM web, top, intermediate, and basal species werenearly constant between seasons. Because they sumto unity, these proportions are not independent(Closs et al. 1993), and the proportion of basalspecies has been shown to decrease with increasingnumber of species in the web (Winemiller 1990;Schoenly and Cohen 1991). Schoenly and Cohen(1991) suggested that smaller proportions of basalspecies in cumulative webs relative to time-specificwebs are due to the fact that turnover amongintermediate species and top species is much moreprevalent than among basal species. This appears tobe the case at MIM and estuaries in general. Thenumber of intermediate species was greater (70) inthe annual web compared to seasonal webs (56, 65),but the number of basal nodes changed little (14–16).
Average food chain length in the MIM web waslow, with top species, such as alligator gar and reddrum, having trophic level estimates between 3.1–3.5. These low values were strongly influenced byconsumption of common detritivores, such as Gulfmenhaden and striped mullet. Our small samplesize for the alligator gar (30, including 12 emptystomachs) could have influenced results. Alligator
Fig. 6. Directed connectance plotted in relation to thenumber of species of the Mad Island Marsh food web (symbolcircled) and 19 published webs: 1) Canadian Lake (Sprules andBowerman 1988), 2) Chesapeake Bay (Baird and Ulanowicz1989), 3) Coachella Valley-aggregated (Polis 1991), 4) Skipwithpond (Warren 1989), 5) Benguela ecosystem (Yodzis 1988), 6)northwest U.S. shelf ecosystem (Link 2002), 7) St. Martin Island(Goldwasser and Roughgarden 1993), 8) Duffin Creek (Tavares-Cromar and Williams 1996), 9) Australian stream (Closs and Lake1994), 10) Bridge Brook Lake (Havens 1993), 11) neotropicalwetlands and streams (Winemiller 1990), 12) UK grassland(Martinez et al. 1999), 13) UK grassland (Dawah et al. 1995),14) Ythan estuary-1 (Hall and Raffaell 1991), 15) Ythan estuary-2(Huxham et al. 1996), 16) Silwood park (Memmot et al. 2000),17) El Verde rainforest (Reagen and Waide 1996), and 18) LittleRock Lake (Martinez 1991).
564 S. Akin and K. O. Winemiller

gars have been reported to feed on gizzard shad(Garcıa et al. 2001), another common detritivore inMIM, but no gizzard shad were found in our sampleof alligator gar stomachs. For red drum, sample sizewas adequate (95, with 35 empty) to representsubadult and adult feeding habits accurately (stan-dard length range of specimens 200–665 mm). Ourestimate of the red drum diet is similar to thatobtained from an investigation of stomach contentsof red drum from Galveston Bay on the Texas Gulfcoast (Scharf and Schlicht 2000).
Omnivory, feeding on more than one trophiclevel, is reported to be common in food webs(Winemiller 1990; Hall and Raffaelli 1991; Polis1991; Havens 1993; Holyoak and Sachdev 1998).The percentage of omnivorous fish and macro-invertebrate species at MIM varied from 17% to26%, a range that overlaps with those reported forthe Ythan estuary (21–34%) and tropical streams(15–25%). A decline in omnivory in MIM duringsummer can be attributed to the fact that manyestuarine-dependent fishes and invertebrates mi-grate into the estuary as postlarvae during winterand early spring. Diets of most postlarvae andjuvenile estuarine fishes consisted of mixtures ofdetritus, diatoms, zooplankton, and other micro-invertebrates. As some of these species grow duringthe warmer months, they became more specializedfeeders. Recent theoretical research indicates thatthese kinds of flexible feeding strategies (adaptiveforaging) can increase food web stability (Kondoh2005).
ACKNOWLEDGMENTS
The Nature Conservancy of Texas and M. Dumesnil providedaccess to field sites and facilities at Mad Island Marsh Preserve. Wethank D. Arrington, J. Arrington, S. Aydin, Y. Bolek, T. Lantz, H.Lopez, M. Morgan, M. Robertson, S. Tarim, and J. Walther forassistance with field data and specimen collection. W. Neill and J.Calvin provided helpful statistical advice. Funding was provided bythe Nature Conservancy of Texas, International Sportfish Fund,and a fellowship from Gaziosmanpasa University to the firstauthor.
LITERATURE CITED
ADAMS, S. M., B. L. KIMMEL, AND G. R. PLOSKEY. 1983. Sources oforganic carbon for reservoir fish production: A trophic-dynamics analysis. Canadian Journal of Fisheries Aquatic Sciences40:1480–1495.
AKIN, S., K. O. WINEMILLER, AND F. P. GELWICK. 2003. Seasonal andspatial variation in fish and macrocrustacean assemblagestructure in Mad Island Marsh estuary, Texas. Estuarine Coastaland Shelf Science 56:1–14.
BAIRD, D., R. R. CHRISTIAN, C. H. PETERSON, AND G. A. JOHNSON.2004. Consequences of hypoxia on estuarine ecosystemfunction: Energy diversion from consumers to microbes.Ecological Applications 14:805–822.
BAIRD, D. AND R. E. ULANOWICZ. 1989. The seasonal dynamics ofthe Chesapeake Bay ecosystem. Ecological Monographs 59:329–364.
BRIAND, F. 1983. Environmental control of food web structure.Ecology 64:253–263.
BRIAND, F. AND J. E. COHEN. 1984. Community food webs havescale-invariant structure. Nature 307:264–266.
BRIAND, F. AND J. E. COHEN. 1987. Environmental correlates offood chain length. Science 238:956–960.
CANUEL, E. A., J. E. CLOERN, D. B. RINGELBERG, J. B. GUCKERT, AND
G. H. RAU. 1995. Molecular and isotopic tracers used toexamine sources of organic-matter and its incorporation intothe food webs of San Francisco Bay. Limnology and Oceanography40:67–81.
CLOSS, G. P. AND P. S. LAKE. 1994. Spatial and temporal variationin the structure of an intermittent-stream food web. EcologicalMonographs 64:1–21.
CLOSS, G., G. A. WATTERSON, AND P. J. DONNELLY. 1993. Constantpredator-prey ratios: An arithmetic artifact? Ecology 74:238–243.
COHEN, J. E. 1978. Food Webs and Niche Space. PrincetonUniversity Press, Princeton, New Jersey.
COHEN, J. E., R. A. BEAVER, S. H. COUSINS, D. L. ANGELIS, L.GOLDWASSER, K. L. HEONG, R. K. HOLT, A. J. KOHN, J. H.LAWTON, N. MARTINEZ, R. O’MALLEY, L. M. PAGE, B. C. PATTERN,S. L. PIMM, G. A. POLIS, M. REJMANEK, T. W. SCHOENER, K.SCHOENLY, W. G. SPRULES, J. M. TEAL, R. E. ULANOWICZ, P. H.WARREN, H. M. WILBUR, AND P. YODZIS. 1993. Improving foodwebs. Ecology 74:252–258.
COHEN, J. E., F. BRIAND, AND C. M. NEWMAN. 1986. A stochastictheory of community food webs. III. Predicted and observedlengths of food chains. Proceedings of the Royal Society of London B228:317–353.
COHEN, J. E., F. BRIAND, AND C. M. NEWMAN. 1990. CommunityFood Webs: Data and Theory. Springer-Verlag, New York.
CREACH, V., M. T. SCHRICKE, G. BERTRU, AND A. MARIOTTI. 1997.Stable isotopes and gut analyses to determine feeding relation-ships in saltmarsh macroconsumers. Estuarine Coastal and ShelfScience 44:599–611.
CURRIN, C. A., S. Y. NEWELL, AND H. W. PEARL. 1995. The role ofstanding dead Spartina alterniflora and benthic microalgae insalt-marsh food webs—Considerations based on multiple stable-isotope analysis. Marine Ecology Progress Series 121:99–116.
CURRIN, C. A., S. C. WAINWRIGHT, K. W. ABLE, M. P. WEINSTEIN, AND
C. M. FULLER. 2003. Determination of food web support andtrophic position of the mummichog, Fundulus heteroclitis, inNew Jersey smooth cordgrass (Spartina alterniflora), commonreed grass (Phragmites australis), and restored marshes. Estuaries26:495–510.
DARNELL, R. M. 1961. Trophic spectrum of an estuarinecommunity based on studies of Lake Pontchartrain, Louisiana.Ecology 42:553–568.
DAWAH, H. A., B. A. HAWKINS, AND M. F. CLARIDGE. 1995. Structureof the parasitoid communities of grass-feeding chalcid wasps.Journal of Animal Ecology 64:708–720.
DEANGELIS, D. L. 1975. Stability and connectance in food webmodels. Ecology 56:238–243.
DEANGELIS, D. L. 1992. Dynamics of Nutrient Cycling and FoodWebs. Chapman and Hall, London.
DE CRESPIN DE BILLY, V., S. DOLEDEC, AND D. CHESSEL. 2000. Biplotpresentation of diet composition data: An alternative for fishstomach contents analysis. Journal of Fish Biology 56:961–973.
DE RUITER, P. C., A.-M. NEUTEL, AND J. C. MOORE. 1995. Energetics,patterns of interaction strengths, and stability in real ecosys-tems. Science 269:1257–1260.
DE RUITER, P. C., V. WOLTERS, J. C. MOORE, AND K. O. WINEMILLER.2005. Food web ecology: Playing Jenga and beyond. Science 309:68–69.
GARCıA, F. J., L. GONZALES-GARCIA, J. M. HERRERA-CASTILLO, K. O.WINEMILLER, AND A. BANDA-VALDES. 2001. Ecology of thealligator gar, Atractosteus spatula, in the Vicente GuerreroReservoir, Tamaulipas Mexico. The Southwestern Naturalist 46:151–157.
Seasonal Variation in an Estuarine Food Web 565

GARCIA, A. M., J. P. VIEIRA, AND K. O. WINEMILLER. 2003. Influencesof El Nino on the dynamics of an estuarine fish assemblage.Estuarine Coastal and Shelf Science 57:489–500.
GELWICK, F. P., S. AKIN, D. A. ARRINGTON, AND K. O. WINEMILLER.2001. Seasonal dynamics in abiotic factors and distribution offishes in the Mad Island Marsh Preserve, Texas. Estuaries 24:285–296.
GOLDWASSER, L. AND J. ROUGHGARDEN. 1993. Construction andanalysis of a large Caribbean food web. Ecology 74:1216–1233.
HAINES, E. B. 1977. Origins of detritus in Georgia salt-marshestuaries. Oikos 29:254–260.
HALL, S. J. AND D. G. RAFFAELLI. 1991. Food web pattern: Lessonsfrom a species rich web. Journal of Animal Ecology 60:823–842.
HALL, S. J. AND D. G. RAFFAELLI. 1993. Food webs. Theory orreality? p. 187–239. In M. Begon and A. H. Fitter (eds.),Advances in Ecological Research. Academic Press, London.
HARRIGAN, P., J. C. ZIEMAN, AND S. A. MACkO. 1989. The base ofnutritional support for the gray snapper (Lutjanus griseus): Anevaluation based on a combined stomach content and stableisotope analysis. Bulletin of Marine Science 44:65–77.
HAVENS, K. E. 1993. Pelagic food web structure in AdirondackMountain, USA, lakes of varying acidity. Canadian Journal ofFisheries and Aquatic Sciences 50:149–155.
HOLYOAK, M. AND S. SAMBHAV. 1998. Omnivory and the stability ofsimple food webs. Oecologia 117:413–419.
HUXHAM, M., S. BEANEY, AND D. RAFFAELLI. 1996. Do parasitesreduce the chances of triangulation in a real food web? Oikos76:284–300.
JEFFRIES, M. J. AND J. H. LAWTON. 1985. Predator-prey ratios incommunities of freshwater invertebrates: The role of enemyand free space. Freshwater Biology 15:105–112.
KONDOH, M. 2005. Linking flexible food web structure topopulation viability: A theoretical consideration on adaptivefood webs, p. 101–113. In P. de Ruiter, V. Wolters, and J. C.Moore (eds.), Dynamic Food Webs: Multispecies Assemblages,Ecosystem Development, and Environmental Change. Elsevier,Amsterdam.
KREBS, C. J. 1999. Ecological Methodology, 2nd edition. AddisonWesley, Menlo Park, California.
KWAK, T. J. AND J. B. ZEDLER. 1997. Food web analysis of southernCalifornia coastal wetlands using multiple stable isotopes.Oecologia 110:262–277.
LAWTON, J. H. 1989. Food webs, p. 43–78. In J. M. Cherrett (ed.),Ecological Concepts. Blackwell Scientific, Oxford, U.K.
LINK, J. 2002. Does food web theory work for marine ecosystems.Marine Ecology Progress Series 230:1–9.
MARTINEZ, N. D. 1991. Artifacts or attributes? Effects of resolutionon the Little Rock Lake food web. Ecological Monographs 61:367–392.
MARTINEZ, N. D. 1992. Constant connectance in community foodwebs. American Naturalist 139:1208–1218.
MARTINEZ, N. D., B. A. HAWKINS, H. A. DAWAH, AND B. P. FEIFAREK.1999. Effects of sampling effort on characterization of food-webstructure. Ecology 80:1044–1055.
MAY, R. M. 1973. Stability and Complexity in Model Ecosystems.Princeton University Press, Princeton, New Jersey.
MCCANN, K. S. 2000. The diversity-stability debate. Nature 405:228–233.
MEMMOT, J., N. D. MARTINEZ, AND J. E. COHEN. 2000. Predators,parasitoids and pathogens: Species richness, trophic generalityand body sizes in a natural food web. Journal of Animal Ecology69:1–15.
MERRITT, R. W. AND K. W. CUMMINS (EDS.). 1996. An Introductionto the Aquatic Insects of North America, 3rd edition. Kendall/Hunt Publishing Company, Dubuque, Iowa.
MONCREIFF, C. A. AND M. J. SULLIVAN. 2001. Trophic importance ofepiphytic algae in subtropical seagrass beds: Evidence frommultiple stable isotope analyses. Marine Ecology Progress Series215:93–106.
MONTOYA, J. M. AND R. V. SOLE. 2002. Small world patterns in foodwebs. Journal of Theoretical Biology 214:405–421.
ODUM, W. E. AND E. J. HEALD. 1975. The detritus based food webof an estuarine mangrove community, p. 265–286. In L. E.Cronin (ed.), Estuarine Research, Volume 1. Academic Press,New York.
PAGE, H. M. 1997. Importance of vascular plant and algalproduction to macroinvertebrate consumers in a southernCalifornia salt marsh. Estuarine Coastal and Shelf Science 45:823–834.
PAINE, R. T. 1988. Food webs: Road maps of interactions or gristfor theoretical development? Ecology 69:1648–1654.
PATTILLO, M. E., T. E. CZAPLA, D. M. NELSON, AND M. E. MONACO.1997. Distribution and Abundance of Fishes and Invertebratesin Gulf of Mexico, Volume II: Species Life History Summaries.ELMR Rep. No. 11. NOAA/NOS Strategic EnvironmentalAssessments Division, Silver Spring, Maryland.
PENNAK, R. W. 1978. Freshwater-Invertebrates of the United States,2nd edition. John Wiley and Sons, New York.
PETERSON, C. C. 1997. Food webs of two Venezuelan clear-waterstreams with seasonal fluctuations in hydrology. M.S. Thesis,Texas A&M University, College Station, Texas.
PETERSON, B. J. AND B. FRY. 1987. Stable isotope in ecosystemsstudies. Annual Review of Ecology and Systematics 198:293–320.
PETERSON, B. J., R. W. HOWARTH, AND R. H. GARRITT. 1985.Multiple stable isotopes to trace the flow of organic matter inestuarine food webs. Science 227:1361–1363.
PIANKA, E. R. 1973. The structure of lizard communities. AnnualReview of Ecology and Systematics 4:53–74.
PIMM, S. L. 1982. Food Webs, 1st edition. Chapman and Hall.London, England.
PIMM, S. L. AND R. L. KITCHING. 1987. The determinants of foodchain length. Oikos 50:302–307.
PIMM, S. L., J. H. LAWTON, AND J. E. COHEN. 1991. Food webpattern and their consequences. Nature 350:669–674.
POLIS, G. A. 1991. Complex trophic interactions in deserts: Anempirical critique of food web theory. American Naturalist 138:123–155.
POLIS, G. A., R. D. HOLT, B. A. MENGE, AND K. O. WINEMILLER.1996. Temporal and spatial components of food webs, p. 435–460. In G. A. Polis and K. O. Winemiller (eds.), Food Webs:Integration of Patterns and Dynamics. Chapman and Hall, NewYork.
REAGAN, D. P. AND R. B. WAIDE (EDS.). 1996. The Food Web ofa Tropical Rain Forest. University of Chicago Press, Chicago,Illinois.
RIERA, P., L. J. STAL, AND J. NIEUWENHUIZE. 1999. Determination offood sources for benthic invertebrates in a salt marsh (AiguillonBay, France) by carbon and nitrogen stable isotopes: Impor-tance of locally produced sources. Marine Ecology Progress Series187:301–307.
SCHARF, F. S. AND K. K. SCHLICHT. 2000. Feeding habits of reddrum (Sciaenops ocellatus) in Galveston Bay, Texas: Seasonal dietvariation and predator-prey size relationships. Estuaries 23:128–139.
SCHOENER, T. W. 1989. Food webs from the small to the large:Probes and hypotheses. Ecology 70:1559–1589.
SCHOENLY, K. AND J. E. COHEN. 1991. Temporal variation in foodweb structure: Sixteen empirical cases. Ecological Monographs 61:267–298.
SIMPSON, E. H. 1949. Measurement of diversity. Nature 163:688.SPRULES, G. AND J. BOWERMAN. 1988. Omnivory and food chain
lengths in zooplankton food webs. Ecology 69:418–426.STRIBLING, J. M. AND J. C. CORNWELL. 1997. Identification of
important primary producers in a Chesapeake Bay tidal creeksystem using stable isotopes of carbon and sulfur. Estuaries 20:77–85.
SUGIHARA, G., K. SCHOENLY, AND A. TROMBLA. 1989. Scaleinvariance in food webs properties. Science 245:48–52.
566 S. Akin and K. O. Winemiller

TAVARES-CROMAR, A. F. AND D. D. WILLIAMS. 1996. The importanceof temporal resolution in food web analysis: Evidence froma detritus-based stream. Ecological Monographs 66:91–113.
THOMPSON, R. M. AND C. R. TOWNSEND. 1999. The effect ofseasonal variation on the community structure and food-webattributes of two streams: Implications for food-web science.Oikos 87:75–88.
THOMPSON, R. M. AND C. R. TOWNSEND. 2005. Food-web topologyvaries with spatial scale in a patchy environment. Ecology 86:1916–1925.
WARREN, P. H. 1989. Spatial and temporal variation in thestructure of a freshwater food web. Oikos 55:299–311.
WARREN, P. H. 1994. Making connections in food webs. Trends inEcology and Evolution 9:136–141.
WERNER, E. E. AND J. F. GILLIAM. 1984. The ontogenetic niche andspecies interactions in size structured populations. AnnualReview of Ecology and Systematics 15:393–425.
WINEMILLER, K. O. 1989. Must connectance decrease with speciesrichness? American Naturalist 134:960–968.
WINEMILLER, K. O. 1990. Spatial and temporal variation in tropicalfish trophic networks. Ecological Monographs 60:331–367.
WINEMILLER, K. O. AND C. A. LAYMAN. 2005. Food web science:Moving on the path from abstraction to prediction, p. 10–23. InP. C. de Ruiter, V. Wolters, and J. C. Moore (eds.), DynamicFood Webs: Multispecies Assemblages, Ecosystem Developmentand Environmental Change. Elsevier, Amsterdam.
WINEMILLER, K. O. AND G. A. POLIS. 1996. Food webs: What canthey tell us about world? p. 1–22. In G. A. Polis and K. O.Winemiller (eds.), Food Webs: Integration of Patterns andDynamics. Chapman and Hall, New York.
YODZIS, P. 1988. The indeterminancy of ecological interactions asperceived through perturbation experiments. Ecology 69:508–515.
YODZIS, P. 1993. Environmental and trophodiversity, p. 26–38. InR. E. Ricklefs and D. Schluter (eds.), Species Diversity inEcological Communities. University of Chicago Press, Chicago,Illinois.
Received, September 15, 2005Revised, February 27, 2006
Accepted, February 28, 2006
Seasonal Variation in an Estuarine Food Web 567