seed bank and vegetation of a constructed reservoir
TRANSCRIPT

WETLANDS. Vol. 15, No. 4, December 1995, pp. 374-385 © 1995, The Society of Wetland Scientists
S E E D B A N K A N D V E G E T A T I O N OF A C O N S T R U C T E D R E S E R V O I R
Bever ly Col l ins and G a r y Wein Savannah River Ecology Laboratoo,
P.O. Drawer E Aiken, SC 29802
Abstract: Vegetation development in constructed wetlands may be accelerated by planting aquatic and emergent species, especially when wetland seed banks are not initially present_ Environmental patchiness of convoluted shorelines may enhance or retard establishment of planted or natural vegetation. Development of the seed bank may be influenced both by planting patterns and by shoreline convolutions. To determine if seed bank composition differed among locations, sediment cores were removed from coves, points, and straights, and from planted and unplanted regions of the shoreline of L-Lake, on the Savannah River Site near Aiken, South Carolina. This reservoir was built in 1985; portions of the shoreline were planted in 1987; and samples were taken in 1992. Sets of cores were taken from areas with water depths up to 1 m, the permanent waterline, and 1 m above water on the shore to determine the effects of hydrology on seed bank composition. Of the 136 seed bank taxa, only 33% were well represented and 35~-16% occurred only once. Water depth w~s the primary influence on seed bank composition. Points, coves, and straights differed in species richness over water depths but did not differ in number of germinable seeds over depths, nor did locations differ in species richness or number of plants at the waterline. Planting vegetation in L-Lake did not significantly influence the seed bank; only 10% of laxa in the seed bank were planted in 1987. Of all taxa recorded, 56% were only in the seed bank, 22% were only in the vegetation, and 13% were in both the seed bank and the vegetation.
Key Words: constructed wetland, hydroperiod, seed bank, water depths, wetland management
I N T R O D U C T I O N
The seed bank o f a site harbors a potent ia l c o m - muni ty (Leck 1989). The cont r ibu t ion o f this com- muni ty to ex tan t vege ta t ion m a y di f fer wi th the local phys ica l or biotic env i ronment . In wet lands , hydroper - iod, phys ica l charac ter i s t ics of the shorel ine, age, and m a n a g e m e n t may all inf luence seed bank and vegela- l ion compos i t i on and s imi la r i ty (van der Valk and Da- vis 1976, 1978, Haag 1983, N icho l son and K e d d y 1983, Ger r i t sen and G r e e n i n g 1989, Haukos and Smi th 1992, 1994).
H y d r o p e r i o d may pr imar i ly de te rmine seed bank and vege ta t ion c o m p o s i t i o n of wet lands . S imi la r i ty o f seed bank and vege ta t ion c o m m u n i t i e s differs both among wet lands and among vegeta t ion types or zones wi thin wet lands (van der Valk and Davis 1976, 1979, Leck and Grave l ine 1979, K e d d y and Rezn icek 1982, K e d d y 1983, Pa rker and Leck 1985, Smi th and Kad lec 1985a, Schne ide r and Shar i tz 1986. Ger r i t sen and Green ing t989 , Hauko~ and Smith 1992, Ungar and Woode l l 1993, Wi l son et al. 1993, Leck and S impson 1993, 1994, K i r k m a n and Shar i tz 1994). Herbaceous wet lands with s table wate r levels may be domina t ed by clonal pe renn ia l s and have litt le recru i tment f rom
the seed bank. Often, the seed bank is depaupera te because the perennia l s p roduce few seeds and seeds o f annuals and nonclona l perennia l s do not persist . Al - ternat ively , the seed bank o f s table wet lands may be rich but c o m p r i s e d o f spec ies that cannot es tab l i sh wi thin the extant vege ta t ion (van tier Valk and Davis 1978, 1979, Keddy and Rezn icek 1982, Smi th and Kad lec 1985b, Wel l ing et al. 1988 a,b, Gerr i t sen and Green ing 1989, Unga r and Woode l l 1993, K i r k m a n and Shari tz , 1994, ter Heerd t and Drost 1994). F luc- tuat ing hydropei- iod or d r a w d o w n s in wet lands can p romote d o m i n a n c e by annuals and nonc lona l peren- nials , some t imes in zones or pa tches that ref lect ger- mina t ion windows , with regenera t ion from a va r ious ly s imi la r and va r ious ly zoned seed bank (van der Valk and Davis 1978, 1979, K e d d y and Rezn icek 1982, Wel l ing et al. 1988 a,b, Gerr i t sen and Green ing 1989, Haukos and Smi th 1992, 1994, K i r k m a n and Shar i tz 1994, L e e k and S impson 1994, ter Hecrd t and Drost 1994),
Phys ica l charac te r i s t i cs o f the shore l ine can influ- ence d ivers i ty o f the seed bank and vege ta t ion in wet- lands. In lakes, for example , water depth , s ed imen t texture, and organic mat ter affect seed bank compo- si t ion, s eed l ing number, and vege ta t ion c o m p o s i t i o n
374

Collins & Wein, R E S E R V O I R SEED B A N K 375
(Haag 1983, Wilson et al. 1993). Gently sloping shore- lines may promote vegetation and seed bank zone ex- pansion, especially if propagule diversity and plant survival decrease with water depth (van tier Valk and Davis 1979, Haag 1983, Nicholson and Keddy 1983). Shorel ine convolu t ions could p romote patchiness . Compared to straight shoreline, coves may have little scouring and serve as deposition sites for seeds and plant propagules. In contrast, points may be scoured and exposed to wind and water movement that re- moves seeds, propagules, and plants (Keddy 1982, 1983).
Young or constructed wetlands that have not been seeded may have a depauperate seed bank (Smith and Kadlcc 1985a, Reinartz and Warne 1993). Planting wetland species to facilitate restoration or vegetation development on a constructed site may contribute to seed bank development if seeds are transported on plants or accumulate from planted vegetation (Reinartz and Warne 1993). Over time, wetland management , including planting regimes and water-level manipula- tions, may interact with environmental variation to en- hance or diminish complexity in the established plant community and seed bank. Augmenting seed bank and vegetation diversity through planting and drawdown management may be desirable to provide stability or resistance to disturbance. In addition, managing for similar species composit ion and distribution between planted and naturally vegetated sites may enhance their functional similarity (van der Valk and Pederson 1989) as required by mitigation action plans.
The broad objective of our research was to deter- mine if either planting vegetation or shoreline shape influence seed bank development in a created wetland. Specific objectives were to
(1) compare the seed banks of different types of lo- cations, points, coves, and straights, along the shoreline of a created reservoir:
(2) compare the seed bank composition of planted and unplanted, naturally vegetated sites along the res- ervoir shoreline;
(3) determine the effect of water depth on the seed bank: and
(4) compare species composition of the seed bank with extant vegetation among locations, between planted and nonplanted areas, and over water depths.
M E T H O D S
The study site is L-Lake, a 400-ha constructed cool- ing reservoir on the Savannah River Site (SRS) near Aiken, South Carolina. During construction, the shore- line was scoured by soil graders; this removed all veg-
etation and much soil. L-Lake was filled in 1985 by pumping water from the Savannah River. Wetland veg- etation was planted along portions of the southern shoreline in 1987 (Wein et al. 1987, Kroeger 1990).
To examine the effects of shoreline shape and plant- ing on seed bank composit ion at the waterline, a set of cores was taken as described below from the wa- terline on 8 planted and 7 unplanted coves, 5 planted and 7 unplanted points, and 7 planted and 6 unplanted straights. To more intensively determine the effect of water depth on seed bank composition within shoreline locations, a second set of cores was collected from three planted and three unplanted coves, points, and straights. At each location, five soil cores (8.25-em diameter × 15-cm deep) were collected in December, 1992 from each of several depths (1 m, 66 cm, 33 cm), at the waterline, and 1 m above the waterline. These five sampling points were at 25-cm intervals along a 1-m transect. Each sampling transect was centered on a line that was perpendicular to the waterline and at the midpoint of each cove, point, or straight. Vegeta- tion was sampled at each seed bank core location by estimating percent cover of all species in t .0 × 1.0 m paired plots in July.
Cores were returned to the laboratory where an equal volume of each thoroughly mixed core (365 cm ~) was spread on potting soil in dishpans (33.7 cm × 28 cm). The remaining portion of each core was given a cold treatment (stratified at 4°C) for 90 days, after which an equal volume (365 cm ~) was spread on pot- ting soil. Dishpans were arrayed in a randomized block design on greenhouse benches. Seedlings were count- ed and removed as soon as they could be identified or transplanted for later identification.
Data Analysis
Data from the immediately-planted and cold-treated cores were combined for analysis because the seed bank was assumed to be composed of both fall and spring-germinating species that could be favored in the unstratified and stratified cores, respectively. For both the depth and waterline cores, seedling counts and spe- cies richness in each dishpan were compared among planting regimes (planted, unplanted); locations (point. cove, or straight); and, for depth cores, water depths (1 m, 66 cm, 33 cm, 0, or I m on shore) using analysis of variance (ANOVA). Water depth was nested within location and planting regime. Count and species rich- ness data were transformed (logLo) (Madansky 1988). Normality was assessed by analysis of residuals. An additional ANOVA was used on the subset of species that were planted to determine if their seed bank dif- fered among locations or water depths in planted and unplanted regions.

376 W E T L A N D S , Volume 15, No. 4, 1995
Similarity of the seed bank and vegetation at each sampling plot was calculated with presence absence data using Jaccard's coefficient (Mueller-Dombois and Ellenberg 1974) and compared among shorcline loca- tions, between planting regimes, and among depths us- ing nonparametric tests.
All analyses were conducted with Statistical Anal- ysis Software (SAS). Throughout the text, d.f. = de- grees of freedom; m.s. - mean square.
RESULTS
Seed Bank Composition
Depth-stratified cores. In all, 33 taxa were well rep- resented (> 2% of total abundance) in L-Lake (Table 1). Individual cores averaged 3-28 taxa and 4-389 plants.
Across water depths, taxa that comprised > 2% of the seed bank were distributed over points, coves, and straights (Table 1, Figure 1). However, species richness differed among locations (Table 2L Coves were richer than points or straights (Figure 1); the total number of taxa in the seed bank ranged among locations f rom 54 in planted straights to 101 in planted coves. In addi- tion, 30% of taxa, including Cyperus ovularis, Digi- taria ischaemum, and Habenaria repens, were fre- quent (> 10%) in only one location. More than one- third of taxa occurred only once in the seed bank, but this proportion ranged narrowly from 35% in unplant- ed coves to 46% in planted straights.
Neither species richness nor number of germinated plants differed between the depth-stratified cores from planted and unplanted regions (Table 2). Only 14 of the 131 taxa that germinated from the seed bank cores werc plantcd in L-Lake (Table 1). There was no effect of water depth, planting regime, or shoreline location on number of plants of these taxa that germinated from the seed bank (Table 3). Across depths, seven planted taxa, Hydrocotyle umbellata, Juncus diffusissimus, J. effusus, Panicum hemitomon, Polygonum hydropipe- roides, Sacciolepis striata, and Typha latifotia, ger- minated f rom planted and unplanted coves, points, and straights (Table 1). Only one species, Sagittaria lati- folia, germinated from planted, but not unplanted, points and coves (Table 1).
Water depth affected species richness and number of plants in the seed bank of cove, point, and straight locations but not of planted and unplanted regions (Ta- ble 2). Cores f rom underwater tended to be species- and number-poor compared to those from the waterline and from the shore (Table !, Figure 1 ). However, spe- cies showed individualistic responses to water depth (Table 1). Of well-represented taxa (> 2% of total abundance), some, such as Habenaria repens and Mi-
kania scandens, were restricted to the waterline and above, whereas others, such as Linaria canadensis, oc- curred over all depths. However, no taxa germinated exclusively from underwater cores (Table 1). Waterline cores. A total of 106 taxa germinated from cores taken from the waterline on planted and unplant- ed coves, points, and straights of L-Lake. Thirty-four taxa were well represented ( > 2% of total richness).
Neither species richness nor number of plants from waterline cores differed among shoreline locations (Ta- ble 4). Of the well represented species, 5 (15%; Eu- patorium capiff~f~Jtium. Gnaphatium spp., Juncus ef fusus, Juncus spp., and Panicum hemitomon) germi- nated from coves, points, and straights, whereas 7 (21%; Aneilema keisak, Cyperus spp., Eleocharis quadrangulata, Erechtites hieracifolia, Juncus diffus- issimus, Myrica cerifera, and Ptilimnium capillaceum) germinated f rom only one location.
Neither species richness nor number of plants dif- fered between planted and unplanted regions of lake- shore (Table 4). Only 10 of the 34 emergent species planted in L-Lake germinated from waterline cores. These comprised only 9% of total species richness from waterline cores.
Seed Bank and Vegetation Similarity: Depth-stratified Samples
There were many fewer species in the vegetation than in the seed bank (Table 1). Over all depths, the seed bank contained 131 taxa, the vegetation had only 87. The vegetation consistently held few species in common with its underlying seed bank (Table 5; Fig- ure 1, 2). Of the taxa distributed over depths, 56% were only in the seed bank, 22% were only in the vegetation, and 13% were in both communities (Figure 2). Similarity of the seed bank and vegetation did not differ among shoreline locations, nor did similarity dif- fer between planted and unplanted areas of lakeshore (Table 5), although two species showed effects of planting. Eleocharis quadrangulata seeds and plants were more frequent and more abundant where planted. Vallisneria americana plants appeared in both planted and unplanted areas but were more abundant where planted (Table 1 ). Similarity did differ among depths (Table 5). Mean similarity dropped sharply at water depths > 33 cm (Table 5). The taxa common to both vegetation and seed bank communit ies were absent from the deepest water (Figure 2~.
Taxa showed individualistic responses to depth (Ta- t3le 1). Thirty-tlaree or those taxa only in me seed 0ank, including Arnmannia coccinea, Ambrosia artemisiifol- ia, Gratiola virginiana, Lycopus rubellus, and Verbena spp., were lound only at or above the waterline; no taxa were collected exclusively from underwater (Ta-

Collins & Wein, RESERVOIR SEED BANK 377
Table 1, qhxa present (>2% abundance) in the vegetation and seed bank (seed bank taxa in parentheses) at each depth (1 m above waterline (A). at the waterline (W), and at 33 cm, 66 cm, and 1 m below waterline) on planted (p) and unplanted (u) coves (C), points (P), and straights (S) of L-Lake.
Species Depths Locations
Acer rubrum L. Acer negundo L. Alternantha philoxeroides Grisebach
Ambrosia artemisiifolia L. Ammannia coccinea Rottboell Andropogon spp.
Aneilema keisak Hasskarl.
Aster pilosus Willd Azotfa spp.
Baccharis hafimiJblia L.
Bacopa caroliniana Robinson* Bidens spp. Boehmeria cvlindrica Swmlz
Cephalanthus occidentalis L. Carex spp.**
Carex albolutescens Schweinitz Cenchrus longispinus Fernald Cyperus spp. Cyperus ovtdaris Torrey Cyperus strigosus L. Dig#aria spp. Dig#aria ischaemum Schreber Echinochloa crusgalli Beauvois*
Eclipta alba Hasskarl Eleocharis accicularis Roemer, Schultes Eleocharis quadrangulata Schultes*
Erechtites hieracifi~lia Raf.
Erigeron spp.
Eupatorium capillifolium Small
Fuirena squarrosa Michaux Galium spp.
Gelsemium sempervirens W. T. Aiton Gnaphalium spp. Gratiola virginiana L. Habenaria repens Nuttal]
l I 'vdroc~Jtyle u n r b c l l a t a L . ~
Hypericum spp.
Juncus spp.
A , W A A, W, 33
(A, W, 33, 66, 1 (W (W A,W
(A, W A , W
(A, W, 33, 66, 1 (.W W
(W A
(A, W, 33, 1 A , W A A , W
(A, W, 33, 66 A A , W
(A (A. W, 33, 66, 1 A
(A. W, 33, 66, 1 (A, W, 33, 66, 1 ¢A. W, 33, 66, 1 (A, W, 33, I (A, 66 A
(A, W, 33, 66 (A, W, 33, I W A, W, 33
(A, W, 33 A , W
tA, W, 66, 1 A
(A, W, 33, 66, 1 A
(A, W, 33, 66, 1 (A,W, 1 A
fA W
(A, W, 33, 1 (A A
(A A, W, 33, 66, l
(A, W. 33, 66, 1 A
(A, l A
(A, W, 33, 66, 1
Sp Sp
Cp, Cu, Pp, Pu, Sp, Su Cp, Cu, Pp, Pu, Sp. Su)
Cu, Pu, Sp, Su) Cu, Pu)
Cu, Pu, Sp, Su Cp, Cu, Pu, Sp. Su)
Cp, Cu, Sp, Su Cp, Cu, Pp, Pu)
Su) Cp
Pu, Su) Su
Cp, Pp, Pu, Sp, Su) Cn, Pp
Su Pn, Sp, Su
Cp, Cu, Pp, Pu, Sp, Su) Pu
Cu, Sp, Su Cp)
Cp, Cu, Pp, Pu, Sp, Su) Su
Cp, Cu, Pp, Pu, Sp, Su) Cp, Cu, Pp, Pu, Sp, Su) Cp, Cu, Pp, Pu, Sp, Su)
Cp, Pu, Su) Pp, Pu, Sp, Su)
Cp Cp, Cu, Pu, Sp, Su)
Cp, Cu, Pp, Pu, Sp, Su) Cu
Cp, Cu, Pp, Sp Cp, Cu, Pp, Pu)
Sp, Su Cp, Cu, Pp, Pu, Sp, Su)
Sp, Su Cp, Cu, Pp, Pu~ Sp, Su)
Sp, Su Cp, Cu, Pp, Pu. Sp, Su) Cp, Cu, Pp, Pu, Sp, Su)
Sp Cp, Cu, Pp, Pu)
Pu Cp, Cu, Pp, Pu, Sp, Su)
Cp, Pu) Su
Cp, Cu, Pp, Su) Cp, Cu, Pp, Pu, Sp. Su Cp, Ctt, Pp, Pu, Sp, Su)
Su Cp. Su)
Cu Cp, Cu, Pp, Pu, Sp, Su)

378 WETLANDS, Volume 15, No. 4, 1995
Table 1. Continued.
Species Depths Locations
Juncus debilis Gray Juncus dichotomus Ell. Juncus diffusissimus Buckley*
Juncus effusus L.*
Juncus tenuous Willd. Leersia spp. Leersia hexandra Swat-tz*
Lernna spp. Lespedeza cuneata G. Don Linaria canadensis Dumont Ludwigia spp. Ludwigia atternifolia L. Ludwigia decurrens Walter Ludwigia leptocarpa Hara
Ludwigia palustris Ell. Lycopus spp. Lycopus americanus Muhl.
Lycopus rubetlus Moench* Mikania scandens Willd.*
MoUugo verticillata L. Myrica cerifera L.
Myriophyllum spp.
Nelumbo lutea Pcrsoon* Panicum spp. Panicum anceps Michaux Panicum hernitomon Schultes*
Parthenocissus quinquefofia Planchon Paspalum spp. Pa.~7)alum notalum Parodi
Paspalum u~'iltei Steudel
Phytolacca americana L.
Pluchea foetida de Candotle Polygonum spp.
Palygonum densiflorum Meissner** Polygonum hydropiperoides Michaux**
Polygonum sagittatum L. **
Polypremum procumbens L. Pontederia cordata L. Patomogeton diversifolius Raf. Ptilimnium capillaceum Raf.
(A, W, 33, 66 Cp, Cu. Pp, Pu, Sp, Su) (A, W, 33, 66 Cp, Cu. Pp, Pu, Sp, Su) A Pp, Su
(A, W, 33, 66, 1 Cp, Cu. Pp, Pu, Sp, Su) A, W Pu, Su
(A. W, 1 Cp. Cu, Pp, Pu, Sp, Su) A Su A, W Cu A, W Cp, Pu, Su
(A, W Cp, Cu, Pu, Su) W Cp
(A Pu, Su) (A, W, 33, 66, 1 Cp, Cu, Pp, Pu, Sp, Su) A, W Cp, Sp
(A, W, 33 Cp, Cu. Pp, Pu, Sp, Su) (A, W, 33, 1 Cp, Cu, Pu, Sp, Su) A Pu, Su
(A, W, 33, 66, 1 Cp, Cu, Pp, Pu, Sp, Su) [A, W, 33, 1 Cp, Cu, Pp, Pu, Sp, Su) (W Cp, Pu, Su A Sp, Su
(A Cu (A Pp. Su) A, W Sp
(A, W Cp, Cu, Pu, Sp, Su) (A, 1 Pp, Pu) A , W Pu, Sp
(A Cp, Cu, Pp, Pu, Sp, Su) W Cu
(A Su) 33, 66, 1 Cu, Pu, Sp
(A, W. 33, 66, 1 Cp, Cu, Pu, Sp, Su) (A, W. 66, 1 Cp, Cu, Pu, Su) A, W Cp, Cu, Pp, Pu, Sp, Su
(A. W. 33. 66 Cp. Cu. Pp. Pu. Sp, Su) W Pu
( 1 Cp) A. W Cp, Cu, Su
(w Cp) A Su
(A Cp) W Su
(W Cu) (W, 1 Sp, Su) W Sp
(W, 33, 1 Cp, Cu, Sp) (A, W Cp, Pu, Sp, Su} A, W, 1 Cp, Cu, Pa, Sp
(A, W, 33, 66 Cp, Cu, Pp, Pu, Sp, Su) A Sp
(A, W, 1 Cp, Pp) (A, W. 33, 66, I Cp, Pp, Pu, Sp, Su) A Cu A, W, 33, 66, 1 Cp, Cu, Pp, Pu, Sp, Su A, W Cp, Cu, Sp, Su
(A, W Cp, Cu, Pp, Pu, Sp)

Col l in s & Wein , R E S E R V O I R S E E D B A N K 379
Table 1. Continued.
Species Depths Locations
Raphanus raphanistrum L.
Rorippa islandica Borbas Rubus spp.
Rumex acetosetla L_ Sacciolepis striata Nash*
Sagittaria latiJbtia L.*
Salix nigra Marshall Scirpus cyperinus Kunth
Setaria geniculata Beauvois Solidago spp. Sofidago rugosa Miller
Specularia perfoliata de Candolle Sphenopholis obtusata Scribner Typha latifolia L.*
Ulmus spp. Vallisneria americana Michaux*
A Su (A Pu) ( 1 Pp) A, W Pu, Sp, Su
(A, W, 66, I Cp, Cu, Pp, Pu, Sp, Su) A Su A, W Cp, Pu, Sp
(A, W, 33, 66, I Cp, Cu, lap, Pu. Sp, Su) A, W Cp, Cu
(A, W Cp, Pp) A, W Cu, Pp, Pu, Sp A, W Sp
(A, W, 33, 66, 1 Cp, Cu, P'p, Pu, Sp, Su) (W Cu. Pu Su) (A. W. 33, 66 Cp, Cu, Pp, Pu, Sp, Su) A Su
(A, W Cu, Pu, Sp) (A, W Cp, Pu, Su) (A, W, 33 Cp, Cu, Pp, Pu, Sp, Su) A, W, 33, 66 Cu, Pp, Pu, Sp
(A, W, 33, 66, 1 Cp, Cu, Pp, Pu, Sp, Su) W Sp A, W, 33, 66, 1 Cp, Cu, Pp, Pu, Sp
* Planted species, ** multiple or mixed species planted.
ble 1). F ive seed bank taxa, Carex albolutescens, Cy- perus spp., Gnaphalium spp., Ludwigia alternifolia, and L. patustris, b e c a m e less f requent wi th depth. In contras t , Ludwigia decurrens, Baccharis halimifotia, Digitaria spp., Eclipta alba, and all Juncus spp. except J. effusus were even ly d is t r ibu ted a m o n g the depths at which they occur red (Table 1).
S ix teen o f the taxa only in the vege ta t ion , inc luding Acer rubrum, A. negundo, Bacopa caroliniana, Leer- sia spp., Ludwigia spp., and Satiric nigra were found at or a b o v e the water l ine . Vallisneria americana and Po- tomogeton diversifolius were represen ted at all depths . On ly Nelumbo lutea was found exc lus ive ly be low the water l ine (Table 1 ).
Table 2. ANOVAs of species richness and number of seedlings per seedbank core (stratified and unstratified cores combined) from several depths on points, coves, and straights in planted and unplanted regions of L-Lake. Data were log,,, transformed for analysis.
Degrees of Effect Freedom Mean Square F Significance I,evel
Species richness Location 2 4.1001 4.89 0.01 Planting regime 1 0.273f1 0.33 0.57 Depth (location) 12 3.4889 4.16 0.0001 Deplh (planting) 4 0.8314 0.99 0.42 Location × Planting 2 0.1676 0.20 0.82 Error 81 0.8392
Number of pfants Location 2 5.7160 2,24 0.11 Planting regimc I 0.0132 0.Ot 0.94 Depth (location) 12 9.6746 3.79 0.0002 Depth (planting) 4 2.(1417 0.80 0.53 Location × Planting 2 0.5799 0.23 0.80 Error 77 2.5556
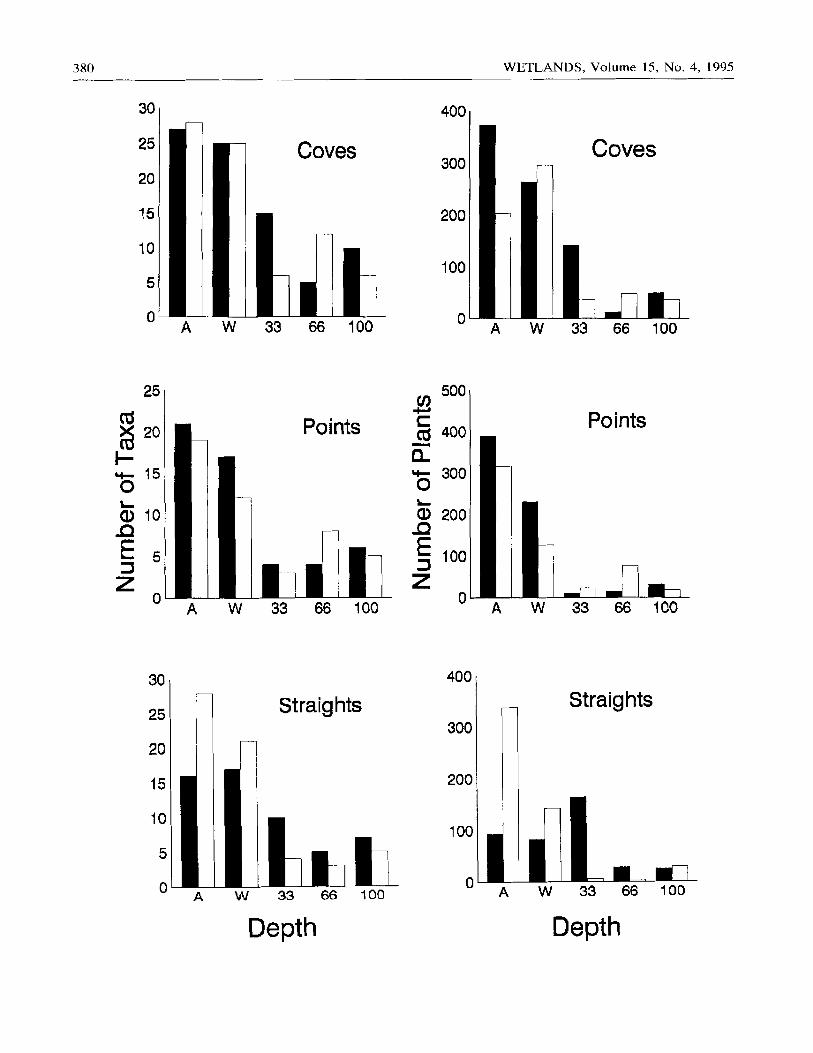
380 WETLANDS, Volume 15, No. 4, 1995
30
25
20
15
10
5
0 A W 33 66 100
400
300
200
100
A W 33 66 100
2~p ~o
~-- 15 O t _
10
Points .~ 5O0 I
i ~ - 300 O
200 ~3
I ' ,oo 0 ~ 0 A W 33 66 10O A W
Points
33 66 100
30
25
20
Straights 400
300
Straights
~o ~5 ~ 2oo ~ OOo 0 A W 33 66 1 O0 A W 33 66 100
Depth Depth

Collins & Wein, RESERVOIR SEED BANK 381
Table 3. ANOVA of location (cove, point, straight), planting regime, and water depth effects en number of plant,; of planted species in seedcores from L-Lake.
Degrees of Effect Freedom Mean Square F Significant Level
Location 2 0.7270 1.98 0.14 Planting regime 1 0.0738 0.20 0.65 Depth (location) 12 0.2925 0.80 0.65 Depth (planting) 4 0.0697 t).19 0.94 Location x Planting 2 0.0614 0.17 0.85 Error 250 0.3667
Ten taxa in both seed bank and vegetation com- munities, including Habenaria repens, Ptilimnium capillaceum, Mikania scandens, Leersia hexandra, Myrica cerifera, Andropogon spp., Paspalum notatum, and Sagittaria lat(~lia, were restricted to the waterline or above (Table 1). An additional 6 taxa had plants restricted to the upland, with seeds found over all depths. These taxa included Rubus spp., Scirpus cv- perinus, Aneilema keisak, and Boehmeria cylindrica (Table 1). Seven taxa, including Panicum hemitomon, Potygonnm hydropiperoides, Sacciolepis striata, Hy- droco~le umbellata, and Juncus effusus, were evenly represented across depths with a seed bank that mir- rored plant distribution.
The individualistic taxa regeneration patterns and responses to depth contributed to community-level similarity patterns across depths. At water depths < 66 cm, the vegetation of L-Lake was a mix of annual and perennial emergents, including Alternantha philoxer- oides, Panicum hemitomon, and Typha latifotia, and was a subset of the seed bank community (Tables 1, 5: Figure 2). Below this depth, species richness of veg- etation declined and aquatic, submersed or floating clonal perennials Potomogeton divers~folius, Vallisner- ia americana, and Nelumbo lutea, which did not ger- minate from the seed bank, dominated (Tables 1, 5: Figure 2).
DISCUSSION
Seed Bank Patterns
Along shorelines and in wetlands with stable water level, seed banks often become depauperate in species and abundance with depth (Haag 1983, Nicholson and Keddy 1983, Wilson et al. 1993). In L-Lake, water depth significantly influenced the species composition
and number of germinable seeds in the seed bank. Cores from 33-cm and deeper water had as few as four individuals and three taxa per 851 cm ~ core. In con- trast, cores from the waterline and 1 m above water had at least 83 germinable seeds and 16 taxa per core. Two factors may contribute to these patterns. First, dominant emergent and floating-leaved species may have predominantly vegetative reproduction and ac- cumulate a sparse seed bank. Second, nonelonal an- nuals and perennials may not persist in the seed bank under stable water levels (Pederson 1981, Haag 1983, Smith and Kadlec 1983, Schneider and Sharitz 1986, Welling et al. 19gSa,b, Poiani and Johnson 1989, Un- gar and Woodell 1993).
Three clonal perennials that dominated vegetation below the waterline in L-Lake, Vallisneria americana, Potomogeton diverxifolius, and Netumbo futea, did not germinate from the seed bank, either because germi- nation requirements were not met or seeds were not present in the sample cores (Poiani and Johnson 1988). Potomogeton is absent from the seed bank of other wetlands as well (Smith and Kadlec 1985b, Poiani and Johnson 1988). In L-Lake, water depth filtered pres- ence of taxa and abundance of individuals in the seed bank cores, Taxa such as Mikania scandens and Phv tolacca americana were restricted to the waterline or above-water and may have seeds that do not sink or do not survive inundation. Some of the taxa that oc- curred to i m depth, such as Eupatorium capillifolium and Ludwigia alternifolia, became less frequent or abundant with depth. These taxa were not in the veg- etation, and their abundance in the seed bank may re- flect historic plant distributions or long-term seed vi- ability. However, other taxa, including Juncus spp., Ludwigia decurrens, and Baccharis hatimifolia, did not change frequency or abundance with depth. These
Figure 1. Species richness and number of plants per seed bank core (stratified and unstratified cores combined) from several depths on points, coves, and straights in planted (shaded) and unplanted (unshaded) regions of L-Lake. A = I m above water; W = at the waterline.

382 W E T L A N D S , V o l u m e 15, No. 4, 1995
Table 4. ANOVA of species richness and number of plants from seedhank cores removed from the waterline in planted and unplanted regions of cove, point, and straight locations on L-Lake. Data were log,, transformed for analysis.
Degrees of Effect Freedom Mean Square F Significance Level
Species richness Location 2 O. 191 1.93 O. 1 6 Planting regime I 0.096 0.97 0.33 Location × Planting 2 tt.024 0.24 (1.79 Error 39 0.099
Number of plants Location 2 0.3009 0.84 0.44 Planting regime 1 0.0034 0.01 0.92 Location × Planting 2 0.1455 0.41 0.67 Error 39 0.3580
taxa were absent or infrequent in the vege ta t ion , and may es tab l i sh on ly when inf requent d i s tu rbances or d r a w d o w n s r e m o v e s tanding vege ta t ion and p rov ide f avo rab le ge rmina t ion cond i t ions (van der Valk and Davis 1979, Keddy and Rezn icek 1982, Wel l ing et al. 1988b, Gerr i t sen and Green ing 1989, Haukos and Smith 1992, 1994, K i r k m a n and Shar i tz 1994, ter Heerd t and Drost 19941.
The seed bank of coves , points , and s t ra ights o f L- L a k e d i f fe red s igni f icant ly in spec ies r ichness across wa te r depths , wi th coves showing grea tes t r ichness , as m a y be expec ted i f they are she l te red or depos i t ion si tes ( K e d d y 1982, 19831. However , loca t ion did not af fec t number of ge rminab l e seeds o r spec ies r ichness o f water l ine cores. Taxa showed ind iv idua l i s t i c dis t r i - bu t ions ove r shore l ine convo lu t ions ; some spec ies (15 % o f those wi th > 2 % f requency) occu r red ove r all loca t ions , whereas others were wel l r ep resen ted in on ly one locat ion. Our research d id not reveal the causes o f seed d is t r ibut ions . Poss ib ly , seed bank den-
sity reflects p lant dens i ty and fecundi ty , which, more than p lan t spec ies compos i t i on , di f fers a m o n g shore- l ine locat ions .
Re la t ive ly few (9-1 I%) o f the spec ies thai occur red in the seed bank were p lan ted in L-Lake . Further, p lant ing r eg ime d id not g rea t ly inf luence spec ies com- posi t ion, r ichness , or number o f ind iv idua ls . On ly Eleocharis quadrangulata plants and seeds occur red p r imar i ly where planted. The only o ther species to show effects o f p lant ing , Vallisneria ~mericana, did not p roduce a seed bank. Thus, a l though p lan t ing may have a f fec ted vege ta t ion e s t ab l i shment on the L - L a k e shorel ine, it d id not have a last ing ef fect on spec ies c o m p o s i t i o n o f the seed bank.
Seed Bank and Vegeta t ion S imi la r i ty
In L -L a ke , s imi la r i ty o f the seed bank and vegeta- tion c o m m u n i t i e s s ign i l ican t ly d i f fe red over depths . In deeper wa te r (66 cm, 1 m), the seed bank showed li t t le
Table 5. Means and nonparametric tests of Jaccard's coefficient of similarity (%/ of seed bank and standing vegetation from coves, points, and straights and from planted and unplanted areas in L-Lake.
Mean Dcgrees of Sum of Similarity N Freedom Mean Score Scores X 2 P
2 1.73 0.42 Location Cove 9.5 35 Point 7.5 37 Straight l 1.3 33
Planting Planted 10.5 48 Unplanted 8.4 57
Depth @1 m l l . I 20 waterline 11.8 24
33 cm 12.2 20 - 6 6 cm 4.1 19 - 1 m 7.0 22
56.2 1967.5 47.9 1774.0 55.2 ] 823.5
I 1 . 7 0 0 . 1 9
57.1 2738.5 49.6 2826.5
4 12.5 0.01 62.7 1254.0 59.8 1436.0 58.2 l 164.0 37.1 706.0 45.7 1005.0

Collins & Wein, RESERVOIR SEED BANK 383
V
t-
80
60
40
20
0
100 80 60 4(3 20
(3
100 80 60 40 20
0-
Planted ~ ~c-~ Above c~ Unplanted ~ ~ Waterline
C P S C P S C P S
C P S
C P S
'1 Waterline
C P S C P S
C P S
33 cm
C P S 10(3 8(3 60 40 20
0 C P S
F ~ 66 cm
C P S C P S
60 40 2O
0 C P S
Vegetation
lm
C P S C P S Seedbank Both
Figure 2. Percent of taxa only in the vegetation, only in the seed bank, or in both over depths in planted (shaded) and unplanted areas on points (P), coves (C), and straights (S) in L-Lake.
similarity with the vegetation, which was dominated by aquatic, submersed or floating clonal perennials that did not germinate in the greenhouse. In shallow water, e m e r g e n t c l o n a l p c [ ~ n n i a l x s u c h a s Typh~ tarifolia, Panicum hemitomon, and Hydroco~te umbetlata pro- duced a seed bank, and there was greater similarity between the communities. However, even in shallow water, as at the waterline and on shore (1 m above
water), the seed bank and vegetation showed low sim- ilarity (< 12 %). The seed bank was richer than the vegetation and contained upland and wetland species t h a t w c r c no t f o u n d in t h e v e g e t a t i o n .
The patterns of similarity between seed bank and vegetation communities of L-Lake agree with those found in other wetlands. In general, wetland seed banks are more species-rich than the vegetation (Get-

384 WETLANDS, Volume 15, No. 4, 1995
ritsen and Greening 1989, Haukos and Smith 1992, 1994, Wilson et al. 1993). Where clonal perennials comprise the vegetation, similarity with the underlying seed bank may depend upon the extent to which the plants produce seed and the contribution of annuals or nonclonal perennials that typically germinate with drawdown or disturbance (van der Valk and Davis 1979. Gerritsen and Greening 1989, ter Heerdt and Drost 1994). For example, Ungar and Woodell (1993) found low correlation between the vegetation and seed bank of perennial-dominated zones in a salt marsh. Most perennial species had no seed bank, and the per- centage of annual plant cover positively correlated with vegetation and seed bank similarity. However, where vegetation is composed of annuals and non- clonal perennials, the extant community may mirror the seed bank or may be a subset of it. For example, in Carolina bays, seed bank and vegetation composi- tion match where the vegetation is not dominated by clonal perennials (Kirkman and Sharitz 1994). In freshwater tidal marsh dominated by annual vegeta- tion. the seed bank and vegetation tend to coincide (Leck and Graveline 1979, Parker and Leek 1985, Leck and Simpson 1994).
Seed Bank Composition and Wetland Management
Five years after planting, the seed bank of L-Lake did not reflect planted and unplanted regions. Thus, results of our research do not indicate that planting wetland vegetation in a created reservoir enhances seed bank development or creates a seed bank that differs from natural revegetation. Further, shoreline convolutions, which might be constructed, apparent~.y have little influence on the number of seeds, although coves were tound to accumulate species. This would suggest that the inclusion of variable shoreline in cre- ated wetlands design would not enhance the develop- ment of the seed bank in systems with a stable water level. In contrast, the common management practice of a periodic drawdown may enhance seed bank and vegetation development in a reservoir such as I,-Lake by redistributing seeds with the waterline and by al- lowing input of seeds of facultative wetland species.
ACKNOWLEDGMENTS
This research was partially funded by the U.S. Army Corps of Engineers Waterways Experiment Station. Additional facilities, supplies, and technical support were provided by ~avannall River Ecology Laboratory ISREL). Manuscript preparation was supported by contract DE-AC09 76SROO-819 between the Depart- ment of Energy and SREL. We thank Philip Dixon for statistical advice. The Wetlands discussion group, Kar-
en Poiani, and an anonymous reviewer provided help- ful comments.
LITERATURE CITED
Gerritsen, J. and H. S_ Greening. 1989. Marsh seed banks of the Okefenokee Swamp: Effects of hydrologic regime and nutrients. Ecology 70:750-763.
Haag, R.W. 1983. Emergenee o f seedlings of aquatic macrophytcs from lake sediments. Canadian Journal of Botany 61 : 148-156.
Haukos, D. A. and L, M. Smith. 1992. Seed bank richness of playas in relation to disturbance, p. 562-568. In M. C. l,andin (ed.) Wet- lands: Prnceedings of the 13th Annual Conference, Society o f Wetland Scientists, New" Orleans, LA, USA.
Haukos, D. A. and L. M. Smith. 1994. Composit ion of seed banks along an elevational gradient in playa wetlands. Wetlands 14:301 307.
Keddy, R A. 1982. Quantifying within-lake gradients of wave en- ergy: interrelationships of wave energy, substrate particle size and shoreline plants in Axe Lake, Ontario. Aquatic Botany 14:41-58.
Keddy, P A. 1983. Shoreline vegetation in Axe Lake, Ontario: e f fects of exposure on zonation patterns. Ecology 64:331-344.
Keddy, P. A, and A. A. Reznicek. 1982. "the role of seed banks in the persistence of Ontario's coastal plain flora. American Journal of Botany 69:13-22,
Keddy, R A., I. C. Wisheu, B. Shippley, and C. Gaudet. 1989. Seed banks and vegetation management fbr conservation: Toward pre dictive communi ty ecology, p_ 347-363. In M . A l,eck, V.T. Par- ker, and R.L. Simpson (eds.) Ecology of Seed Banks. Academic Press, Ncw York, NY, USA.
Kirkman, L. K. and R. R. Sharitz. 1994. Vegetation disturbance and maintenance of diversity in internlittently flooded Carolina bays in South Carolina. Ecological Applications 4:177-188.
Kroeger, S, R. 1990. Wetland vegetation establishment in L-Lake. Unpublished report, Savannah River Site, United States Depart- ment o f Energy, Aiken, SC, USA,
Leek, M, A, 1989. Wetland ,~eed banks, p. 283-305, In M. A. Leek, V. T. Parker, and R. L. Simpson (eds.L Ecology of Seed Banks. Academic Press, New York, NY, LISA.
Leek, M. A. and K. J. Graveline. 1979. The seed bank of a fresh- water ~i,I' ,1 marsh. American Journal of Botany 66: f 006-1015.
Leek, M. A. and R. L, Simpson. 1993. Seeds and ,seedlings of theHamilton Marshes, a Delaware River tidal freshwater wetland. Proceedings of The Academy of Natural Sciences of Philadelphia !. 44:267-281.
Leek, M. A. and R. L Simpson. 1994. Tidal freshwater wetland zonation: seed and seedling dynamics. Aquatic Botany 47:61-75.
Madansky, A. 1988. Prescriptions for Working Statisticians. Spring- e~Verlag, New York, NY, USA.
Mueller-Dombois, D. and H, Ellenberg. 1974. Aims and Methods of Vegetation Ecology. John Wiley and Sons, New York, NY. USA,
Nicholson, A. and P. A. Keddy. 1983. The depth profile of a shore- line seed bank in Matchedash I.ake, Ontario. Canadian Journal of Botany 61:3293-3296.
Parker, V. T. and M. A. Leek. 1985. Relationships of seed banks to plant distribution patterns in a freshwater tidal wetland. American Journal of Botany 72:161-174,
Pedersom R. L. 1981. Seexl bank characteristics of the Delta Marsh, Manitoba: Applications for wetland management , p. 61 69. In B. Richardson (ed.) Selected Proceedings of the Midwest Conference on Wetland Values and Malmgement. Freshwater Society, Navar- re, MN, USA.
Poiani, K. A. and Vq. C. Johnson. 1988. Evaluation of the e m e r gence method in estimating seed bank conrposition of prairie wet lands. Aquatic Botany 32:91-97.
Poiani, K, A. and W. C. Johnson. 1989. Effect of hydroperk~ on seed-bank composit ion in semipermanent prairie wetlands- Ca- nadian Journal of Botany 67:856--864.
Reinartz, J. A. and E. L. Warne. 1993. Development of vegetation

C o l l i n s & W e i n , R E S E R V O I R S E E D B A N K 3 8 5
in small created wetlands in southeastern Wisconsin. Wetlands 13: 153-164.
Schtleider, R, L. and R. R. Sharitz, 1986, Seed bank dynamics in a southeastern riverine swamp. American Journal of Botany 73: 1022-1030.
Smith, L. M. and J. A, Kadlec, 1983. Se~:d hanks and their role during drawdown of a North American marsh. Journal of Applied Ecology 20:673-684.
Smith, L. M. and J. A. Kadlec. 1985a. The cffccts of disturbance on marsh seed hanks. Canadian Journal of Botany 63:2133-2137.
Smith, 1,. M. and J. A. Kadlec. 1985b. Predictions of vegetation change following fire in a Great Salt Lake marsh. Aquatic Botany 21:43-51.
ter Heerdt, G. N. J. and H. J. Drool. 1994. Potential for the devel- opment of marsh vegetation from the seed bank aftcr a drawdown. Biological Conservat ion 67:1-1 I.
Ungar, I. A. and S. R. J. Woodell. 1993, The relationship between the seed bank and species composit ion of plant communi t ies in two British salt marshes. Journal of Vegetation Science 4 :531- 536.
van der Valk, A. G. and C. B. Davis. 1976. The seed banks o f prairie glacial marshes. Canadian Journal of Botany 54:1832- 1838.
van der Valk, A. G. and C. B. Davis. 1978. The role of secd banks in vegetation dynamics of prairie glacial marshes. Ecology 59: 322-335.
van der Valk, A. G. and C. lg. Davis. 1979. A reconstruction of the recent vegetational history of a prairie marsh, EagleLake, Iowa, from its seed bank. Aquatic Botany 6:29-51.
van der Valk, A. G. and R. L. Pederson. 1989. Seed banks and thc management and restoration of natural vegetation, p. 329-346. In M. A. Leck, V. T_ Parker, and R. L Simpson (eds.) Ecology of Seed Banks, Academic Press, New York, NY, USA.
Wein, G. R., S. R. Kroeger, and G. J. Picrcc. 1987. Lacustrine vegetation establishment within a cooling reservoir, p. 206-218. In E J. Webb (ed.) Proceedings of the 14th Annual Conference on Wetlands Restoration and Creation. Hillsborough Communi ty College, Plant City. FL, USA.
Welling, C. H., R. L. Pederson, and A, G. van der Valk. 1988a. Temporal patterns in recruitment from the seed hank during draw- downs in a prairie wetland, Journal of Applied Ecology 25:999- 1007.
Welling, C. H., R_ L. Pederson, and A. G. van der Valk. 1988b. Recruitment from the seed bank and the development of emer- gence zonation during a drawdown in a prairie wetland. Journal of Ecology 76:483--496.
Wilson, S_ D, D. R. J, Moore, and E A. Keddy. 1993. Relationships of marsh seed banks to vegetation patterns along environmental gradients. Freshwater Biology 29:361-370.
Manuscript received 23 March ]995; revisions received 26 June 1995 and 11 Augus t 1995; accepted 31 August 1995.