seed dormancy in camellia sinensis l. (theaceae): … seed dormancy in camellia sinensis l....
TRANSCRIPT

Draft
Seed dormancy in Camellia sinensis L. (Theaceae): Effects
of cold-stratification and exogenous gibberellic acid application on germination
Journal: Botany
Manuscript ID cjb-2016-0149.R1
Manuscript Type: Article
Date Submitted by the Author: 18-Aug-2016
Complete List of Authors: Song, Danping; University of Shanghai for Science and Technology
Jaganathan, Ganesh; University of Shanghai for Science and Technology, Institute of Biothermal Engineering Han, Yingying; University of Shanghai for Science and Technology Liu, Baolin; University of Shanghai for Science and Technology
Keyword: Camellia sinensis, Cold-stratification, Germination, Physiological dormancy, Seedling emergence
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Seed dormancy in Camellia sinensis L. (Theaceae): Effects of cold-stratification and
exogenous gibberellic acid application on germination
Danping Song¶, Ganesh K. Jaganathan¶*, Yingying Han, Baolin Liu*
Institute of Biothermal Science and Technology, University of Shanghai for Science and technology, China.
¶ Equal first author
Running title: Germination ecology of C. sinensis
*Corresponding author
Tel: 08621-55277768; Fax: 08621-55270695
Email: [email protected]; [email protected]
Page 1 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Abstract
There are several different opinions about the presence of dormancy in tea (Camellia sinensis),
but no strong evidence is available to conclude whether or not seeds are dormant. Freshly
matured tea seeds collected from Hangzhou, China, at the natural dispersal time did not
germinate in light at daily alternative temperature regimes of 10/15, 15/20, 20/25, 25/35º C or at
a constant temperature of 25º C. Seeds were permeable to water and the embryos did not grow
prior to radicle emergence, thus, the seeds have no physical, morphological or
morphophysiological dormancy. When cold-stratified at 4º C for 1, 2 and 3 months, 64, 88 and
93% of the seeds germinated respectively. Intact fresh seeds failed to germinate after treating
with 0, 10, 500 and 1000 ppm GA3, whereas 3, 4, 61 and 86% of cracked seeds germinated
respectively. Thus, the seeds have non-deep and intermediate physiological dormancy. Two-
months cold-stratified seeds buried at 0, 1 and 5 cm soil depth in pots showed that seeds at 1 cm
depth established significantly higher number of seedlings (P <0.05) than at other two depths.
Because tea seeds are susceptible to summer temperature drying, these seeds do not establish a
persistent seed bank.
Keywords: Camellia sinensis; Cold-stratification; Germination; Physiological dormancy;
Seedling emergence.
Page 2 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Introduction
The successful establishment of newly germinated seedlings into young plants largely depends
on synchronizing the timing of germination with the growing season, where adequate moisture
and favorable temperatures are available (Fenner and Thompson 2005). Given the inherent risk
of seedling death during conditions unfavorable for growth at times other than the growing
season, many species produce seeds with some form of dormancy and specific requirement for
light, so that the germination is restricted to the growing season (Baskin and Baskin 2014; Long
et al. 2015). Consequently, the presence of dormancy (or lack thereof) is an important selective
mechanism influencing the distribution of plant species across various ecosystems (Willis et al.
2014). Thus, understanding seed dormancy and germination is critical to further our knowledge
on ecosystem complexities and species distribution.
Camellia is an important genus with more than 200 species in the family Theaceae. The vast
majority of species belonging to this genus are either endemic or adapted to China (Chen et al.
2007). However, many are being threatened in the wild due to deforestation and other
anthropogenic pressures imposed on their natural habitat. This genus also includes Camellia
sinensis L., commonly known as tea, which is an important commercial crop cultivated in over
40 countries for its leaves (Chen et al. 2007). Tea is the second most commonly consumed
beverage after water (Nair 2010). The natural vegetation of tea has been undergoing significant
alteration in recent years with the introduction of novel cultivars. As a result, studying the
storage and germination of seeds has attracted some interest, but whether or not tea seeds have
dormancy has not been explicitly quantified. This lack of understanding poses severe problems
in providing viable seed materials to the next sowing season and during long-term storage of
Page 3 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
seeds for conservation purposes because there is a risk of judging viable (but dormant) seeds as
dead. This is crucial because tea seeds are considered to be recalcitrant, i.e. drying to low
moisture content is lethal, but excised embryos can survive drying to very low moisture content
(Chandel et al. 1995; Kim et al. 2002). Thus, it is not known whether the whole seeds have some
dormancy which can be alleviated when the embryos are excised and cultured.
According to Baskin and Baskin (2004), a dormant seed is the one that does not germinate within
30 days when incubated under favorable conditions (including optimal temperature, moisture,
light etc.) that stimulat germination in non-dormant seeds. From the studies available on tea
seeds, several inferences can be made about the presence or absence of dormancy. Tubbs (1932)
showed that germination of intact seeds buried under 1 cm of soil was slow during the first
month, but 100% of seeds germinated when seeds were retrieved and tested after three months.
Likewise, Visser and Tillekeratne (1958) reported that intact seeds required 49 days to reach 75%
germination. The results presented by Sebastiampillai and Anandappa (1979) also show that
germination of intact seeds was slow and continued for up to 9 weeks. However, these results
contrast with Chandel et al. (1990) and Bhattacharya et al. (2002), who concluded that the tea
seeds are non-dormant, as germination occurred within a week. Therefore, to gain more insights
into this knowledge gap, we addressed the following questions: (1) Are tea seeds dormant? (2) If
dormant, what class of dormancy do they have and how can it be lifted? and (3) What are the
germination requirements for the seeds in the field?
Materials and methods
Seed collection
Page 4 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Mature fruits of C. sinensis were collected from Hangzhou, China (30.2741° N, 120.1551° E)
from a minimum of 30 plants growing at an altitude of 50-80m m.a.s.l. (meter above sea level).
The average temperature and rainfall of the study site recorded in the past 30 years is 17±1º C
and 119.84±0.6 mm. The warmest month is July with an average temperature of 29º C and the
coldest month is January with an average temperature of 5º C. Fruits collected during the natural
shedding period before the onset of winter in late November 2015 were transported to University
of Shanghai for Science and Technology, Shanghai, China, on the same day and upon arrival,
seeds were removed from the fruits by ripping them open manually or with a scalpel. Each
capsule contained approximately 3 to 4 brown colored seeds. Seeds suspected of being affected
by fungal or bacterial infection or immature seeds (judged by green color and small size) were
discarded and the healthy seeds were immediately placed in plastic bags and stored at 4º C until
used in the experiments. All the experiments began within 4 days of seed collection.
Seed weight and moisture content
The average weight of 100 seeds was determined by weighing eight replicates of 100 randomly
selected seeds using a standard balance (0.001 g). The moisture content of the seeds was
determined gravimetrically using four replicates of 15 seeds dried at 103º C for 17 h. The
difference in fresh and dry weight was expressed as a percentage of wet weight basis.
Germination tests
Seeds used in the germination experiments were thoroughly surface sterilized with 1% sodium
hypochloride for 15 min, then rinsed thrice in distilled water. Germination tests were conducted
at four alternating temperatures (10/15, 15/20, 20/25, 25/35º C) and at a constant temperature of
Page 5 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
25º C. For all the temperatures tested, light was provided for 14 h during the warm phase or at
25º C in the case of constant temperature, at an intensity of c. 25 µ mol m−2
s−1
. The criterion for
judging germination was radical emergence by at least 3 mm. Germination was scored daily or
every alternative day until no seeds had germinated for 1 week. Final germination was expressed
as a percentage of germinated seeds.
Exogenous application of GA3
Intact and cracked seeds were treated with 0, 10, 500 and 1000 ppm GA3 solution for 24 hours
under laboratory conditions (20-25º C; 50-65% RH). These seeds were then surface-disinfected
and germinated at 20/25º C, as described above.
Effect of cold-stratification
Intact seeds were cold-stratified at 4º C under white fluorescent light at an irradiance of 10 µ mol
m−2
s−1
for 1, 2 and 3 months. The moisture content of the seeds at the end of each month was
also determined in three replicates of 10 seeds as above. In addition, cracked seeds were also
cold-stratified at 4º C for 1 month. All the stratification experiments were conducted in four
replicates of 15 seeds taken in Petri dishes. Following the stratification period, seeds were
examined for germination at 20/25º C with the same photoperiod used in the germination
experiments.
Photographs of seed germination
In order to determine the changes in the external surface of the seeds during germination and
identify the location where the radicle emerges on the seed coat, intact seeds cold-stratified for
Page 6 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
one month and germinated at 20/25º C were photographed every 4 hours using a CanonTM
camera.
Germination of seeds at different soil depths
To understand the germination ability of seeds in the natural environment, we sowed four
replicates of 50 seeds previously cold-stratified for two months in pots filled with natural soil.
Three depths were chosen: (1) soil surface; (2) 1 cm and (3) 5 cm. The pots were incubated in a
growth chamber set at 20/25º C with the same photoperiod used in the germination experiments.
The pots were watered regularly and the establishment of seedlings was monitored for 2 months.
Statistical analysis
The germination percentage of cold-stratified seeds and seedling establishment data were
analyzed using one-way analysis of variance (ANOVA) followed by Tukey post-hoc test for
statistical significance. For all the statistical procedures, data were arcsine transformed to
improve the normality before performing ANOVA. All the data were subjected to statistical
analysis in SPSS software (version 21.0).
Results
Seed weight and moisture content
The average weight of 100 seeds was 117.7±0.3g. Thus, on average, each seed weighed about 1
g. The average moisture content at the time of collection was 43.41.6%.
Germination of fresh seeds
Page 7 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
No seed germinated at any temperature regimes tested, except at 15/20º C, where 3 seeds showed
cracking of seed coat but no visible emergence of radicle. The germination experiment was
terminated after 65 days.
Effect of GA3 on seed germination
No fresh intact seeds treated with GA3 germinated even after 67 days (Table 1). However, fresh
seeds cracked and treated with GA3 improved germination significantly only at higher
concentration (Table 1).
Cold-stratification
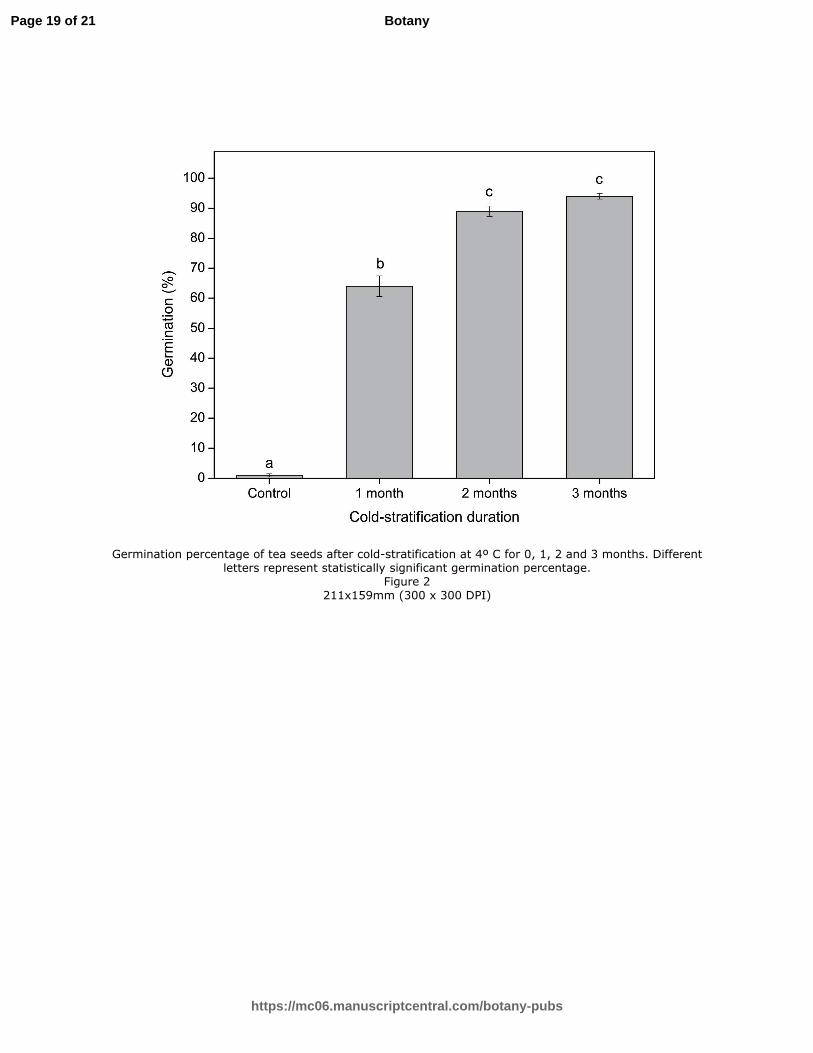
Cold-stratification at 4º C for one month significantly improved germination compared with
control seeds (P <0.05; Figure 2). However, seeds that were subjected to two months cold-
stratification germinated to 89%, which is significantly higher than the germination percentage
of seeds that received one-month stratification (P <0.05). Although three-month cold-stratified
seeds germinated to 94%, this was not significantly different from the two-month stratification
treatment (P >0.05; Figure 2), suggesting two-months of stratification could alleviate dormancy
in most of the seeds. Ninety-two percent of cracked seeds cold-stratified for one-month
germinated.
Photomicrographs
Photographs taken during germination are shown in Figure 3. The cracking of the seed coat
occurred within 4 hours of imbibition and this was followed by the radicle elongation on the
micropyle side. The radicle continued to grow and the shoot emerges from the elongated radicle.
Page 8 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Seedling establishment from various depths
Seedling establishment differed with the position of seeds in the soil. Overall, seedling
establishment was poor in buried seeds compared to those sown on the soil surface. Seeds
subjected to a two-month cold stratification treatment established a significantly higher
percentage of seedlings at 1 cm depth (P <0.05; Figure 4) compared with seeds buried at 5 cm
depth or at the soil surface. This suggests that there is an optimal burial depth at which seedlings
emerge successfully.
Discussion
Freshly collected tea seeds at the natural dispersal time did not germinate even after 60 days at
any of the temperatures tested, indicating that the seeds were dormant. Since seeds were
permeable to water and their weight increased to 97% within three days, dormancy was not due
to an impermeable seed coat (data not shown). In general, however, cracked seeds increased in
mass more quickly than the intact seeds (data not shown), which is in agreement with Tubbs
(1932). Intact seeds failed to germinate after treating them with GA3. Nonetheless, germination
percentage of cracked seeds increased with increasing concentration of GA3 (Table 1).
Germination of cracked seeds treated with GA3 occurred within a week confirming that the
embryos were fully developed and the seeds have no morphological dormancy, as observed by
Bhattacharya et al. (2002). Cold-stratification also improved germination significantly (Figure
2). Collectively, these results suggest that the tea seeds have physiological dormancy.
From our data, it appears that about 64% of the seeds collected had non-deep physiological
Page 9 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
dormant seeds which germinated after 1-month cold-stratification and nearly 24% had
intermediate physiological dormant seeds that required 2 months cold-stratification for
germination (Figure 2). The climatic data recorded in the study site between 1981 and 2010 show
that winter begins at the end of November and lasts up to early February. The average
temperature during winter was 5 ºC (Figure 1). The seeds matured and were dispersed at the end
of autumn, i.e. late October to early November, persist in soil during winter and germinate during
spring. The requirement of cold-stratification to break dormancy is satisfied by low winter
temperatures. Further, the results that GA3 treated intact seeds did not germinate suggest that the
hard seed coat acts as a mechanical barrier at the time of seed dispersal as noted previously
(Sebastiampillai and Anandappa 1979). It is possible that cold-stratification received during
winter months weakens the seed coat allowing the embryo to elongate. This was reported in
some species with a hard endocarp, e.g. Prunus armeniaca (Jaganathan et al. 2015), but, to the
best of our knowledge, no such reports are available for Theaceae. Our results might explain the
ecological significance of physiological dormancy in tea seeds preventing germination during
harsh winter conditions.
The difference in opinion on whether or not tea seeds have dormancy can be attributed to several
factors including the time of seed collection and location at which the seeds mature or both.
Katsuo et al. (1970) convincingly demonstrated that ripe tea seeds of Japanese and Assam
varieties germinated rapidly, but unripe seeds required 3 weeks of cold-stratification to germinate
to a higher percentage. However, seeds collected in our study are mature; even some of the seeds
had already dispersed to the ground at the time of collection. In a separate experiment, we
germinated the seeds fallen to the ground at 25º C in Petri dishes with 1% agar-water and found
Page 10 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
that the seeds can germinate only after one-month cold-stratification (data not shown). Given the
fact that tea is indigenous to subtropical climate, seeds maturing in locations where winter
conditions are warmer might not benefit from cold-stratification. However, we also rule out this
possibility because, in most of the countries, including those in tropics, tea is commonly
cultivated or grown at altitudes ranging between 1000 and 1500 masl (Chen et al. 2012;
Sebastiampillai and Anandappa 1979), where the winter temperatures are cold. But the question
still remains, why are tea seeds reported as non-dormant? The answer may lie in the post-harvest
storage conditions employed before submitting the seeds to germination test. Chandel et al.
(1990) stored seeds for 5 weeks at 10-15º C and subsequently germinated them, which possibly
served as a cold-stratification treatment and resulted in immediate germination. Interestingly,
Bhattacharya et al. (2002) also reported tea seeds were non-dormant, but whether the seeds were
stored before germination is not clear. In spite of this, they collected seeds during January, i.e. at
the end of winter, when the seeds would have been cold-stratified in the field. Alternatively, there
might be some variation at the population-level driven by the environmental conditions selecting
dormant or non-dormant seeds in a particular location. This is particularly true for many other
physiological dormant species (Baskin and Baskin 2014). Thus, more studies on tea species from
different locations are required in the future to answer this question clearly.
Light is an important environmental factor regulating the germination of seeds in soil (Pons and
Fenner 2000). Previous studies conducted on Camellia and other Theaceae genera have shown
that light is not an important requirement in some species for germination, while in others
darkness inhibited germination (Shen et al. 2010; Shen et al. 2015). The ability of C. sinensis to
germinate both in light and darkness at a wide-range of temperatures implies that this mechanism
Page 11 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
is important in allowing more seeds to germinate. It seems that the only chance for autumn
dispersed tea seeds to germinate is the following spring. Seeds of C. sinensis that remain non-
germinated after spring cannot persist in soil for a long-time because the high summer
temperatures likely dry the seeds below a critical moisture level required for maintaining
viability. In addition, tea seeds are also susceptible to soil microbes and fungi leading to viability
loss. The moisture content of the seeds during the cold-stratification period remained above 30%
compared with 43% at the time of seed collection. The three month cold-stratification period
used in the current study reflects the conditions prevailing in the natural environment and the
seeds do not lose viability at sub-zero temperatures. A previous study from seeds collected at the
same site showed that partial drying is not harmful to seeds (Hu et al. 1994), but we assume the
natural conditions during summer may dry seeds below the safe level. It is of interest to note that
our earlier field visits showed that no seeds found on the soil surface during late summer were
viable and most of them were completely empty (Jaganathan et al. unpub).
Based on the photographs taken and visual observations made during germination experiments,
we conclude that germination in C. sinensis is hypogeal (Figure 3; see also Chandel et al. 1990).
The visible sign of germination is the breaking of the seed coat, followed by radicle elongation
through this crack on the side of micropyle. The radicle has to grow enough in length before the
shoot starts growing. The successful germination of tea seeds, as indicated by radicel emergence,
in the present and previous studies, cannot be used as a useful indicator to conclude that seeds
would develop into seedlings and establish plants in soil. Thus, we undertook seed burial
experiments using cold-stratified seeds. The depth at which seeds are present in the soil
significantly affects the establishment of seedlings. Seeds buried at 1 cm depth showed higher
Page 12 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
seedling establishment compared with seeds buried at 5 cm depth or on the surface (Figure 4).
Examination of seeds buried at 5 cm depth revealed that the radicle grows deep into the soil but
the shoot failed to reach the soil surface. However, the conditions at 1 cm depth are optimal for
the shoot to emerge and reach the soil surface. Similar results have also been reported in other
Camellia species including C. nitidissima (Yang et al. 2008). It can be surmised that the tea seeds
shed in autumn do not become buried deep in the soil before spring due to their large size and a
slight burial during winter is therefore advantageous for seedling establishment.
Acknowledgements
We thank Xiao Qun for her help throughout the project. Thanks are also to Sheng Rui, Sha Lu
and Xiaoyan Song for their help in seed collection.
References
Baskin, C.C., and Baskin, J.M. 2014. Seeds: ecology, biogeography, and evolution of dormancy
and germination. Second ed. Elsevier, San Diego, USA.
Baskin, J.M., and Baskin, C.C. 2004. A classification system for seed dormancy. Seed science
research 14(1): 1-16.
Bhattacharya, A., Nagar, P., and Ahuja, P.S. 2002. Seed development in Camellia sinensis (L.) O.
Kuntze. Seed Science Research 12(01): 39-46.
Chandel, K., Chaudhury, R., Radhamani, J., and Malik, S. 1995. Desiccation and freezing
sensitivity in recalcitrant seeds of tea, cocoa and jackfruit. Annals of Botany 76(5): 443-450.
Chandel, K., Lakhanpaul, S., Singh, I., and Chaudhury, R. 1990. Germination and desiccation
tolerance of tea (Camellia sinensis (L) O. Kuntze) seeds and feasibility of cryopresevation.
Page 13 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Srilankan Journal of Tea Science 59(2): 89-94.
Chen, H., Pritchard, H.W., Seal, C.E., Nadarajan, J., Li, W., Yang, S., and Kranner, I. 2012. Post
desiccation germination of mature seeds of tea (Camellia sinensis L.) can be enhanced by pro-
oxidant treatment, but partial desiccation tolerance does not ensure survival at− 20° C. Plant
science 184: 36-44.
Chen, L., Zhou, Z.-X., and Yang, Y.-J. 2007. Genetic improvement and breeding of tea plant
(Camellia sinensis) in China: from individual selection to hybridization and molecular breeding.
Euphytica 154(1-2): 239-248.
Fenner, M., and Thompson, K. 2005. The ecology of seeds. Cambridge University Press.
Hu, J., Guo, C., and Shi, S. 1994. Partial drying and post-thaw preconditioning improve the
survival and germination of cryopreserved seeds of tea (Camellia sinensis). Plant Genetic
Resources Newsletter (IPGRI/FAO) 98: 25-28.
Jaganathan, G., Wu, G., Song, X., and Liu, B. 2015. Cryopreservation of dormant Prunus
armeniaca L. seeds. Seed Science and Technology 43(3): 456-466.
Katsuo, K., Toyao, T., and Kayumi, S. 1970. The germination of tea seed. Part 1. Relations
between picking period, storage conditions and seed germination. Study of Tea 39: 14-19.
Kim, H.-H., Cha, Y.-S., Baek, H.-J., Cho, E.-G., Chae, Y.-A., and Engelmann, F. 2002.
Cryopreservation of tea (Camellia sinensis L.) seeds and embryonic axes. CryoLetters 23(4):
209-216.
Long, R.L., Gorecki, M.J., Renton, M., Scott, J.K., Colville, L., Goggin, D.E., Commander, L.E.,
Westcott, D.A., Cherry, H., and Finch‐Savage, W.E. 2015. The ecophysiology of seed
persistence: a mechanistic view of the journey to germination or demise. Biological Reviews
90(1): 31-59.
Page 14 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Nair, K.P. 2010. The agronomy and economy of important tree crops of the developing world.
Elsevier, London, U.K. .
Pons, T.L., and Fenner, M. 2000. Seed responses to light. In Seeds: the ecology of regeneration
in plant communities. Edited by M. Fenner. CABI. pp. 237-260.
Sebastiampillai, A., and Anandappa, T. 1979. The influence of moisture and temperature in the
germinability and longevity of tea (Camellia sinensis L.) seeds. Tea Quarterly 48: 8-20.
Shen, S.K., Wang, Y.H., and Ma, H.Y. 2010. Seed germination requirements and responses to
desiccation and storage of Apterosperma oblata (Theaceae), an endangered tree from south‐
eastern China: implications for restoration. Plant Species Biology 25(2): 158-163.
Shen, S.K., Wu, F.Q., Yang, G.S., Wang, Y.H., and He, S.Z. 2015. Seed germination and seedling
emergence of Euryodendron excelsum HT Chang: implications for species conservation and
restoration. Plant Species Biology: doi: 10.1111/1442-1984.12105.
Tubbs, F. 1932. The germination of tea seed. Tea Quarterly 5: 66-69.
Visser, T., and Tillekeratne, L. 1958. Observations on the germination and storage of tea pollen
and seed. Tea Quarterly 29: 30-35.
Willis, C.G., Baskin, C.C., Baskin, J.M., Auld, J.R., Venable, D.L., Cavender‐Bares, J.,
Donohue, K., and Rubio de Casas, R. 2014. The evolution of seed dormancy: environmental
cues, evolutionary hubs, and diversification of the seed plants. New Phytologist 203(1): 300-309.
Yang, Q.-H., Wei, X., Zeng, X.-L., Ye, W.-H., Yin, X.-J., Zhang-Ming, W., and Jiang, Y.-S. 2008.
Seed biology and germination ecophysiology of Camellia nitidissima. Forest Ecology and
Management 255(1): 113-118.
Page 15 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Table 1. Germination percentage of intact and cracked tea seeds at the time of collection after
treating with different concentration of GA3. Significant difference in germination percentages
are specified with different alphabets.
GA3 concentration
(ppm) Intact seeds Cracked seeds
0 0 3±0.5a
10 0 4±0.8a
500 0 61±2.6b
1000 2±0.6 86±2.0c
Page 16 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Figure 1. The monthly average temperature and rainfall of the seed collection site recorded in the
past 30 years.
Figure 2. Germination percentage of tea seeds after cold-stratification at 4º C for 0, 1, 2 and 3
months. Different letters represent statistically significant germination percentage.
Figure 3. Photomicrographs of a germinating seed that had been cold-stratified for two-months at
4º C and germinated at 20/25º C. Fresh seeds before placing in 1% agar-water (a); after 4h (b);
8h (c); 12 h (d); 16h (e); 20 h (f); 24h (g); 28h (h); 32h (i); 40 h (j); cross section of fresh seeds
(k); backside of 8h germinating seed (l). C, cotyledons; E, embryo; EA, embryonic axes; M,
micropyle; L, leaves; R, radicle; S, shoot; and SC, seed coat. Scale – 5 mm.
Figure 4. Percentage of seedlings developed from two-months cold-stratified seeds when buried
at 0, 1 and 5 cm depths. Different letters indicate significant difference between groups.
Page 17 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
195x136mm (300 x 300 DPI)
Page 18 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany
Draft
Germination percentage of tea seeds after cold-stratification at 4º C for 0, 1, 2 and 3 months. Different letters represent statistically significant germination percentage.
Figure 2 211x159mm (300 x 300 DPI)
Page 19 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Photomicrographs of a germinating seed that had been cold-stratified for two-months at 4º C and germinated at 20/25º C. Fresh seeds before placing in 1% agar-water (a); after 4h (b); 8h (c); 12 h (d); 16h (e); 20 h (f); 24h (g); 28h (h); 32h (i); 40 h (j); cross section of fresh seeds (k); backside of 8h
germinating seed (l). C, cotyledons; E, embryo; EA, embryonic axes; M, micropyle; L, leaves; R, radicle; S, shoot; and SC, seed coat. Scale – 5 mm.
Figure 3 62x83mm (300 x 300 DPI)
Page 20 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Percentage of seedlings developed from two-months cold-stratified seeds when buried at 0, 1 and 5 cm depths. Different letters indicate significant difference between groups.
Figure 4
240x207mm (300 x 300 DPI)
Page 21 of 21
https://mc06.manuscriptcentral.com/botany-pubs
Botany